Змеиный яд

Snake Venom - очень токсичная слюна [ 1 ] содержащий зоотоксины , которые облегчает иммобилизацию и пищеварение добычи . Это также обеспечивает защиту от угроз. Snake Venom обычно вводит уникальные клыки во время укуса , хотя некоторые виды также способны плевать яд . [ 2 ]
Ядовитые железы , которые секретируют Zootoxins, представляют собой модификацию околоушных слюнных желез, найденных у других позвоночных , и обычно расположены на каждой стороне головы, внизу и за глазами, и заключены в мышечную оболочку. Яд хранится в больших железах, называемых альвеолами, а затем передается протоком в основание канала или трубчатых клыков, через которые его выброшен. [ 3 ] [ 4 ]
Venom содержит более 20 различных соединений, которые в основном являются белками и полипептидами . [ 3 ] [ 5 ] Сложная смесь белков, ферментов и различных других веществ обладает токсичными и летальными свойствами. [ 2 ] Яд служит для иммобилизации добычи. [ 6 ] Ферменты в яде играют важную роль в пищеварении добычи, [ 4 ] и различные другие вещества несут ответственность за важные, но несмертельные биологические эффекты. [ 2 ] Некоторые из белков в змеином яде оказывают очень специфическое влияние на различные биологические функции, включая коагуляцию крови, регуляцию артериального давления и передачу нервных или мышечных импульсов. Эти яды были изучены и разработаны для использования в качестве фармакологических или диагностических инструментов и даже лекарств. [ 2 ] [ 5 ]
Химия
[ редактировать ]Белки составляют 90-95% сухого веса Venom и отвечают практически за все его биологические эффекты. [ 5 ] [ 7 ] Сотни, даже тысячи белков, обнаруженных в ядах, включают токсины, в частности нейротоксины , а также нетоксичные белки (которые также обладают фармакологическими свойствами) и множество ферментов, особенно гидролитических . [ 2 ] Ферменты ( молекулярная масса составляют 80-90% виперидов и 25-70% элапидных ядов, включая пищеварительные гидролазы , L-аминокислотную оксидазу , фосфолипазы , тромбин про-коагулян 13-150 кДа ) - подобный сериновые протеазы и металлопротеиназы (геморрагины), которые повреждают эндотелий сосудов . Полипептидные токсины (молекулярная масса 5-10 кДа) включают цитотоксины , кардиотоксины и постсинаптические нейротоксины (такие как α-бунгунгаротоксин и α-корбратоксин ), которые связываются с рецепторами ацетилхолиновых в ней нервно-мышечных соединений. Соединения с низкой молекулярной массой (до 1,5 кДа) включают металлы, пептиды, липиды, нуклеозиды , углеводы, амины и олигопептиды , которые ингибируют ангиотензин-конвертирующий фермент (ACE) и потенциальный брадикинин (BPP). Изменение меж и внутривидового химического состава яда является географическим и онтогенным. [ 3 ] Фосфодиэстеразы мешают сердечной системе добычи, в основном для снижения артериального давления . Фосфолипаза А2 вызывает гемолиз лизируя фосфолипидные клеточные мембраны эритроцитов , . [ 8 ] Аминокислотные оксидазы и протеазы используются для пищеварения. Аминокислотная оксидаза также запускает некоторые другие ферменты и отвечает за желтый цвет яда некоторых видов. Гиалуронидаза увеличивает проницаемость ткани, чтобы ускорить поглощение других ферментов в ткани. Некоторые змеиные яды несут фаскулины , такие как мамба ( дендроаспис ), которые ингибируют холинэстеразу , чтобы добыча терять мышечную контроль. [ 9 ]
| Тип | Имя | Источник |
|---|---|---|
| Оксидоредуктазы | лактатдегидрогеназа | Elapidae |
| L-аминокислотная оксидаза | Все виды | |
| Каталаза | Все виды | |
| Трансферы | Аланиновая аминотрансфераза | |
| Гидролазы | Фосфолипаза A2 | Все виды |
| Лизофосфолипаза | Elapidae, Viperidae | |
| Ацетилхолинэстераза | Elapidae | |
| Щелочная фосфатаза | Bothrops Atrox | |
| Кислотная фосфатаза | Deinagkistrodon Sharp | |
| 5'-нуклеотидаза | Все виды | |
| Фосфодиэстераза | Все виды | |
| Дезоксирибонуклеаза | Все виды | |
| Рибонуклеаза 1 | Все виды | |
| Аденозин трихосфатаза | Все виды | |
| Амилаза | Все виды | |
| Гиалуронидаза | Все виды | |
| NAD-NUCLEOTIDASE | Все виды | |
| Кининогеназа | Viperidae | |
| Фактор X активатор | Viperidae, Crotalinae | |
| Гепариназа | Crotalinae | |
| α-фибриногеназа | Viperidae, Crotalinae | |
| β-фибриногеназа | Viperidae, Crotalinae | |
| Аб-фибриногеназа | Bity Gabonica | |
| Фибринолитический фермент | Crotalinae | |
| Активатор протромбина | Crotalinae | |
| Коллагеназа | Viperidae | |
| Эластаза | Viperidae | |
| Лиазы | Глюкозамита-аммиак-линейаза |
Змеиные токсины сильно различаются в своих функциях. Два широких класса токсинов, обнаруженных в ядобах змеи, - это нейротоксины (в основном обнаруженные в элапидах) и гемотоксины (в основном обнаруженные в Viperids). Тем не менее, исключения происходят-яд черного шепта кобры ( Naja nigricollis ), элаппа, состоит в основном из цитотоксинов , в то время как у гремучей змеи мохаве ( Crotalus scutulatus ), гадюка, является главным образом нейротоксичным. Как элапиды, так и гадюки могут нести множество других типов токсинов.
| α-нейротоксины | α-бунгаротоксин , α-токсин, эрабутоксин , кобритоксин |
|---|---|
| β-нейротоксины ( PLA2 ) | β-бонгаротоксин , нотксин, аммодитоксин , кротоксин , тайпоксин |
| κ-нейротоксины | Каппа Бунгаротоксин |
| Дендротоксины ( Куниц ) | Дендротоксин , токсины I и K; Возможно, β-бонгаротоксин цепь B |
| Кардиотоксины | Naja nigricollis y-toxin, кардиотоксин III (он же цитотоксины) |
| Миотоксины | Миотоксин -а, кротамин |
| Сарафотоксины | Сарафотоксины A, B и C |
| Геморрагины (металлопротеаза) | Мукролизин , атролизины , акутолизины и т. Д. [ 11 ] |
| Гемотоксины (сериновая протеаза) | Венобин а |
Токсины
[ редактировать ]В этом разделе отсутствует информация о том, как эволюционные/структурные классификации соответствуют функциональным классификациям. ( Август 2021 г. ) |
Нейротоксины
[ редактировать ]
Начало нового нейронного импульса заключается в следующем:
- Обмен ионами (заряженными атомами) через мембрану нервной клеток посылает деполяризующий ток к концу нервной клетки (клеточный концерн).
- Когда деполяризующий ток прибывает в конце нервного клеток, нейротрансмиттер ацетилхолин (ACH), который удерживается в везикулах , выделяется в пространство между двумя нервами (синапс). Он перемещается через синапс к постсинаптическим рецепторам.
- ACH связывается с рецепторами и передает сигнал в целевую ячейку, и через короткое время он разрушается ацетилхолинэстеразой.
Фасцикулины
[ редактировать ]- Эти токсины атакуют холинергические нейроны (те, которые используют ACH в качестве передатчика) путем разрушения ацетилхолинэстеразы (ACHE). Следовательно, ACH не может быть сломана и остается в рецепторе. Это вызывает тетанию (непроизвольное сокращение мышц), что может привести к смерти. Токсины были названы фаскулинами с момента инъекции в мышей , они вызывают тяжелые, генерализованные и долговечные (5-7 ч) фасцикуляции (быстрые сокращения мышц).
- Пример змеи: найдено в основном в яде мамбы ( дендроаспс spp.) И некоторые гремучие змеи ( Crotalus spp.)
Дендротоксины
[ редактировать ]- Дендротоксины ингибируют нейротрансмиссию, блокируя обмен положительными и отрицательными ионами через нейрональную мембрану, приводят к отсутствию нервного импульса, тем самым парализуя нервы.
- Пример змеи: мамба
α-нейротоксины
[ редактировать ]- Альфа-нейротоксины -большая группа; Более 100 постсинаптических нейротоксинов были идентифицированы и секвенированы. [ 12 ] α-нейротоксины атакуют никотиновые ацетилхолиновые рецепторы холинергических нейронов. Они имитируют форму молекулы ацетилхолина и таким образом вписываются в рецепторы, где они блокируют поток ACH, что приводит к ощущению онемения и паралича.
- Примеры змеи: король кобра ( Ophiophagus hannah ) (известный как ханнахтоксин, содержащий α-нейротоксины), [ 13 ] Морские змеи (гидрофийны) (известные как эрабутоксин), многополосный крайт ( мультикшинг Bungarus ) (известный как α-бунгунгаротоксин ) и кобры ( Naja spp.) (известный как кобритоксин )
Цитотоксины
[ редактировать ]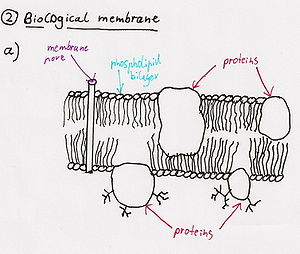

Фосфолипазы
[ редактировать ]- Фосфолипаза - это фермент, который трансформирует молекулу фосфолипидов в лизофосфолипид (мыло) → Новая молекула привлекает и связывает жиры и разрывает клеточные мембраны. Фосфолипаза A2 является одним из специфических типов фосфолипаз, обнаруженных в яде змеи.
- Пример змеи: Окинаван Хабу ( Trimeresurus flavoviridis )
Кардиотоксины / цитотоксины
[ редактировать ]- Кардиотоксины - это компоненты, которые специально токсичны для сердца. Они связываются с определенными сайтами на поверхности мышечных клеток и вызывают деполяризацию → токсин предотвращает сокращение мышц. Эти токсины могут привести к тому, что сердце нерегулярно преодолеть или перестать бить, вызывая смерть. Примером является с тремя пальцами кардиотоксин III из китайской кобры , пример короткой семьи с тремя пальцами ( InterPro : IPR003572 ).
- Пример змеи: мамба и некоторые Naja виды
Гемотоксины
[ редактировать ]- Гемотоксины вызывают гемолиз, разрушение эритроцитов (эритроциты) или индуцируют коагуляцию крови (свертывание, например, мукроцетин ). Общее семейство гемотоксинов включает в себя металлопротеиназы змеиного яда, такие как мукролизин . [ 11 ] [ 14 ]
- Примеры змеи: большинство гадюков и многие виды Cobra : тропическая гремучая змея Crotalus durissus производит коннульсин , коагулянт. [ 15 ]
Миотоксины
[ редактировать ]
Миотоксины - это маленькие, основные пептиды, обнаруженные в гремучей змее [ 16 ] [ 17 ] и ящерица (например, мексиканская ящерица из бисера ) [ 18 ] яды. Это включает в себя негментативный механизм, который приводит к тяжелому скелетных мышц некрозу . Эти пептиды действуют очень быстро, вызывая мгновенный паралич , чтобы предотвратить сбег добычи и в конечном итоге смерть из -за диафрагмального паралича.
Первым миотоксином, который должен быть идентифицирован и изолирован, был кротамин , обнаруженный в 1950 -х годах бразильским ученым Хосе Морой Гонсалвесом из яда тропического южноамериканского гремучих змеев Crotalus durissus Terrificus . Его биологические действия, молекулярная структура и ген, ответственный за его синтез, были выяснены за последние два десятилетия.
Определение токсичности яда (LD 50 )
[ редактировать ]В этом разделе отсутствует информация о том, почему развод альбумина работает лучше. ( Август 2021 г. ) |
Токсичность змеиного яда оценивается с помощью токсикологического теста, называемого медианной летальной дозой , летальной дозы 50% (сокращенно как LD 50 ), которая определяет концентрацию токсина, необходимую для убийства половины членов испытанной популяции. Потенция яда дикой змеи значительно варьируется из -за различных влияний, таких как биофизическая среда , физиологический статус, экологические переменные , генетические вариации ( адаптивные или случайные) и другие молекулярные и экологические эволюционные факторы. [ Цитация необходима ] Это верно даже для членов одного вида. Такие различия меньше в популяциях в неволе в лабораторных условиях, хотя его нельзя устранить. Тем не менее, исследования для определения потенции ядом змеи должны быть разработаны, чтобы минимизировать изменчивость.
Несколько методов были разработаны с этой целью. Одним из подходов является использование 0,1% бычьего сывороточного альбумина (также известного как «Фракция V» в процессе COHN ) в качестве разбавителя при определении значений LD 50 . Это приводит к более точным и последовательным определениям LD 50 , чем использование 0,1% физиологического раствора в качестве разбавителя. Например, фракция V производит около 95% очищенного альбумина (сушеный сырой яд). Солевой раствор в качестве разбавителя постоянно дает широко различные результаты LD 50 почти для всех ядовитых змей. Он производит непредсказуемые различия в чистоте осадков (35-60%). [ 19 ] Фракция V является структурно стабильной, потому что у нее семнадцать дисульфидных связей ; Он уникален тем, что имеет самую высокую растворимость и самую низкую изоэлектрическую точку основных белков плазмы. Это делает его последней частью, чтобы быть ускоренной из его решения. Бычий сывороточный альбумин расположен во фракции V. Осаждение альбумина осуществляется путем уменьшения pH до 4,8, вблизи pH белков и поддержания концентрации этанола при 40%с концентрацией белка 1%. Таким образом, только 1% от первоначальной плазмы остается в пятой фракции. [ 20 ]
Когда конечной целью обработки плазмы является очищенный плазменный компонент для инъекции или переливания , компонент плазмы должен быть очень чистым. Первый практическое крупномасштабное метод фракционирования плазмы крови было разработано Эдвином Дж. Коном во время Второй мировой войны . Это известно как процесс COHN (или метод COHN). Этот процесс также известен как фракционирование холодного этанола, поскольку он включает постепенное увеличение концентрации в растворе этанола при 5 ° C и 3 ° C. [ 21 ] Процесс COHN использует различия в свойствах плазменных белков, в частности, высокой растворимости и низкой PI альбумина. Поскольку концентрация этанола увеличивается поэтапно с 0 до 40%, pH снижается с нейтрального (pH ~ 7) до 4,8, что находится рядом с PI альбумина. [ 21 ] На каждом этапе белки осаждаются из раствора и удаляются. Последним осадком является очищенный альбумин. Существует несколько вариантов этого процесса, включая адаптированный метод Nitschmann и Kistler, который использует меньше шагов, и заменяет центрифугирование и объемное замораживание фильтрацией и диафильтрацией. [ 21 ] [ 22 ] Некоторые новые методы очистки альбумина добавляют дополнительные шаги очищения в процесс COHN и его вариации. Обработка хроматографического альбумина появилась в 1980-х годах, однако она не была широко принята до тех пор, пока позже из-за неравнолечительной хроматографической оборудования. Методы, включающие хроматографию, как правило, начинаются с криоазубельной плазмы плазмы, проходящей обмен буферными буферами с помощью диафильтрации или хроматографии буфера, для подготовки плазмы для выполнения этапов ионообменной хроматографии . После ионного обмена обычно происходят шаги очистки и обмен буферным обменом. [ 21 ]
Тем не менее, хроматографические методы стали приняты в 1980 -х годах. [ Цитация необходима ] Продолжались события между тем, когда началось фракционирование COHN в 1946 году, и когда в 1983 году появилась хроматография . В 1962 году процесс Kistler и Nistchmann был создан в качестве отдачи от процесса COHN. В 1990 -х годах были созданы процессы Zenalb и CSL Albumex, которые включали хроматографию с вариациями. Общий подход к использованию хроматографии для фракционирования плазмы для альбумина: восстановление супернатанта I, делипидация, анионная хроматография , катионная хроматография и гель -фильтрационная хроматография. Извлеченный очищенный материал составлен с комбинациями октаноата натрия и N-ацетил-триптофаната натрия, а затем подвергается процедурам вирусной инактивации, включая пастеризацию при 60 ° C. Это более эффективная альтернатива, чем процесс COHN, потому что:
- плавная автоматизация и относительно недорогая завод, необходимость,
- легче стерилизовать оборудование и поддерживать хорошую производственную среду
- Хроматографические процессы менее вредны для белка альбумина
- Более успешный результат альбумина может быть достигнут. [ Цитация необходима ]
По сравнению с процессом COHN чистота альбумина увеличилась с 95% до 98% с использованием хроматографии, а выход увеличился с 65% до 85%. Небольшое процентное увеличение имеет значение в отношении чувствительных измерений, таких как чистота. Большой недостаток связан с экономикой. Хотя метод предлагался эффективным, приобретение необходимого оборудования было трудным. Необходима большая техника, и в течение долгого времени отсутствие доступности оборудования ограничивало его широкое использование. [ Цитация необходима ]
Эволюция
[ редактировать ]Venom развивался только один раз среди всех Toxicofera около 170 миллионов лет назад, а затем диверсифицировался в огромное разнообразие яда, которое наблюдалось сегодня. [ 23 ] Оригинальный токсичный яд был очень простым набором белков, которые были собраны в паре желез. Впоследствии этот набор белков развивался независимо в различных линиях токсичныхферов, включая змеи , ангуморфы и игуании . [ 24 ] С тех пор несколько линий змеи утратили способность производить яд, часто из -за изменения диеты или изменения в хищной тактике. [ 23 ] В дополнение к этому, прочность и состав яда изменились из -за изменений в добыче определенных видов змей. Например, яд мраморной морской змеи ( aipysurus eydouxii ) стал значительно менее токсичным после того, как диета этого вида изменилась от рыбы на строго рыбные яйца. [ 23 ] Считается, что эволюция яда отвечает за огромное расширение змей по всему миру. [ 23 ] [ 25 ]
Механизм эволюции в большинстве случаев был дупликацией генов в тканях, не связанных с ядом. [ 24 ] Существующие белки слюны являются вероятными предками большинства генов токсинов яда. [ 26 ] Экспрессия нового белка в ядовитой железе последовала за дублированием. [ 24 ] Затем приступил к естественному отбору для адаптивных признаков после модели рождения и смерти, где за дублированием следует функциональная диверсификация, что приводит к созданию структурно связанных белков, которые имеют немного разные функции. [ 23 ] [ 24 ] [ 27 ] Изучение эволюции яда стало главным приоритетом для ученых с точки зрения научных исследований, из -за медицинской значимости змеиного яда с точки зрения проведения антиономических и раковых исследований. Знание больше о композиции яда и о том, как он потенциально может развиваться, очень полезно. Три основных фактора, которые влияют на эволюцию яда, были тщательно изучены: хищники змеи, которые устойчивы к яду змеи, добычу, которая находится в эволюционной гонке вооружений со змеями, и специфические диеты, которые влияют на внутривидовую эволюцию яда. [ 23 ] [ 28 ] Яды продолжают развиваться в виде специфических токсинов и модифицируются для нацеливания на определенную добычу, и токсины, как обнаружено, различаются в зависимости от диеты у некоторых видов. [ 29 ] [ 30 ]
Быстрая эволюция яда также может быть объяснена гонкой вооружений между молекулами, нацеленными на яд, у резистентных хищников, таких как опоссум , и ядом змеи, который нацелен на молекулы. Ученые провели эксперименты по опоссумам и обнаружили, что множественные испытания показали замену тихой замены в гене фон-фактора ( VWF ) фон Виллебранда, которые кодируют для белка гемостатического крови, нацеленного на яд. Считается, что эти замены ослабляют связь между VWF и токсичным лигандом ядом змеи (ботроцетин), который изменяет чистый заряд и гидрофобность. Эти результаты важны для эволюции яда, потому что это первое цитирование быстрой эволюции в молекуле, нацеленной на яд. Это показывает, что эволюционная гонка вооружений может происходить с точки зрения защитных целей. Альтернативные гипотезы предполагают, что эволюция яда обусловлена трофической адаптацией, тогда как эти ученые считают, что в этом случае отбор будет происходить на признаках, которые помогают при выживании добычи с точки зрения эволюции яда вместо успеха хищничества. Несколько других хищников ямы Viper (Mongooses и Hedgehogs) показывают такую же тип взаимосвязи между змеями, что помогает подтвердить гипотезу о том, что яд играет очень сильную оборонительную роль наряду с трофической ролью. Что, в свою очередь, подтверждает идею, что хищничество на змеях может быть гонкой вооружений, которая производит эволюцию ядом змеи. [ 31 ]
Некоторые из различных адаптаций, полученных в результате этого процесса, включают яд более токсичный к специфической добыче в нескольких линиях, [ 30 ] [ 32 ] [ 33 ] белки, которые добывают добычу, [ 34 ] а также метод отслеживания добычи после укуса. [ 35 ] Хотя функция яда развивалась как специфическая для класса добычи (например, особые коагуляторные эффекты), [ 36 ] Эволюция широких токсикологических эффектов (например, нейротоксичность или коагулотоксичность), по -видимому, широко влияет на тип добычи. [ 37 ] При наличии пищеварительных ферментов в ядом змеи когда -то считалось адаптацией для оказания помощи пищеварению. Тем не менее, исследования гремучей змеи Western Diamondback ( Crotalus atrox ), змеи с высокопротеолитическим ядом , показывают, что яд не оказывает влияния на время, необходимое для прохождения пищи . [ 38 ] Эти различные адаптации яда также привели к значительным дебатам об определении яд и ядовитых змей. [ 23 ]
Инъекция
[ редактировать ]Гадюки
[ редактировать ]В гадюках , которые имеют наиболее высокоразвитый аппарат для доставки яда, ядтная железа очень большая и окружена массовой или временной мышцей , которая состоит из двух полос, превосходных, возникающих из-за глаза, более низкого, простирающегося от железы к нижней челюсти. Поток несет яд от железы к клыку. В гадюках и элапидах эта канавка полностью закрыта, образуя игольчатую трубку для подкожного кодерки. У других видов канавки не покрыты или только частично покрыты. От передней конечности железы проток проходит под глазом и над верхней челюстной костью , до базального отверстия ядовитого клыка, который закреплен в толстой складке слизистой оболочки . С помощью подвижной верхнечелюстной кости шарнирной к префронтальной кости и соединенной с поперечной костью, которая продвигается вперед мышцами, установленными в действии путем отверстия рта, клык возведен и продюн сбрасывается через дистальное отверстие. Когда змея кусается, челюсти закрываются, и мышцы, окружающие сжимание железы, вызывая выброс яда через клыки.
Элапиды
[ редактировать ]В протероглифных элапидах клыки являются трубчатыми, но они короткие и не обладают подвижностью, наблюдаемой у гадюков.
Колубриды
[ редактировать ]Описхоглифные колубриды увеличили, рифмы, расположенные на задней конечности верхней челюсти , где небольшая задняя часть верхней губной или слюнной железы производит яд.
Механика укуса
[ редактировать ]
Несколько родов, включая азиатские коралловые змеи ( Calliophis ), Burrowing ASPS ( Atractaspis ) и ночные добавки ( причины ), замечательны тем, что имеют исключительно длинные ядовитые железы, простирающиеся вдоль каждой стороны тела, в некоторых случаях простирающиеся на протяжении всей сердце. Вместо мышц височной области, служащих, чтобы выдвинуть яд в проток, это действие выполняется по сторонам тела.
Знаки наблюдаются значительную изменчивость в поведении прикуса. При кусании змеи -гадюки часто ударяются быстро, сбрасывая яд, когда клыки проникают в кожу, а затем немедленно освобождаются. В качестве альтернативы, как и в случае с откликом на кормление, некоторые гадюки (например, Lachesis ) укусают и удерживают. Протероглиф опишлиф или может закрыть свои челюсти и укусить или прочно жевать в течение значительного времени.
Различия в длине клыка между различными ядовитыми змеями, вероятно, связаны с эволюцией различных поразительных стратегий. [ 39 ] Кроме того, было показано, что клыки разных видов ядовитых змей имеют разные размеры и формы в зависимости от биомеханических свойств добычи змеи. [ 40 ]
Механика плев
[ редактировать ]Прызгивая кобры родов Наджа и Хемачатуса , когда раздражаются или угрожают, могут выбросить потоки или распыление яда на расстоянии 1,2 метра (от 4 фута) до 2,4 метра (8 футов). Клыки этих змей были модифицированы для целей плев; Внутри клыков канал делает изгиб на 90 ° к нижнему передней части клыка. Спиттеры могут многократно плевать и при этом иметь возможность доставить смертельный укус.
Плютить - это только защитная реакция. Змеи, как правило, стремятся к глазам воспринимаемой угрозы. Прямой удар может привести к временному шоку и слепоте за счет воспаления роговицы тяжелого и конъюнктивы . Хотя обычно нет серьезных симптомов, если яд немедленно смыт с большим количеством воды, слепота может стать постоянной, если его не лечить. Краткий контакт с кожей не сразу опасен, но открытые раны могут быть векторами для эненомации.
Физиологические эффекты
[ редактировать ]Эта статья требует дополнительных цитат для проверки . ( август 2021 г. ) |
Четыре различных типа яда действуют по -разному:
- Протеолитический яд демонтирует молекулярное окружение, в том числе в месте укуса.
- Гемотоксичный яд действует на сердечно -сосудистую систему, включая сердце и кровь.
- Нейротоксический яд действует на нервную систему, включая мозг.
- Цитотоксический яд имеет локализованное действие на месте укуса.
Протероглифные змеи
[ редактировать ]Эффект яда протероглифных змей ( морские змеи , Крайты , Мамба , черные змеи , змеи тигров и добавки смерти ) в основном на нервной системе , а дыхательный паралич быстро вырабатывается путем введения яда в контакт с центральным нервным механизмом, который, как это контролирует дыхание; Боль и местный набухание, которые следуют за укусом, обычно не являются сильными. Укус всех протероглифных элапидов, даже самых маленьких и нежных, таких как коралловые змеи , насколько известно, смертоносен для людей. Однако остаются некоторые мягкие ядовитые элапиды, такие как змеи с капюшоном ( парасута ), бандитские полосы ( вермицелла ) и т. Д.
Гадюки
[ редактировать ]Viper Venom ( Viper's Russell , Saw-Scalled Vipers , кустарники и гремучие змеи ) больше действует на сосудистую систему, приводя к свертыванию крови и свертывания легочных артерий; Его действие на нервную систему не велик, ни одна отдельная группа нервных клеток, по-видимому, не выбран, и влияние на дыхание не так прямое; Влияние на циркуляцию объясняет Великую депрессию, которая является симптомом варневой энвеномации. Боль раны сильной и быстро сопровождается отеком и обесцвечиванием. Таким образом, симптомы, вызванные укусом европейских гадюков, описаны Мартином и Лэммом: [ 41 ]
Закуска сразу же сопровождается местной болью горящего персонажа; Вскоре конечность набухает и обесцвечивается, и через один -три часа великая прострация, сопровождаемая рвотой и часто диареей , наступает. Холодное, липкое пот является обычным. Пульс становится чрезвычайно слабым, и небольшую одышку можно увидеть и беспокойство. В тяжелых случаях, которые встречаются в основном у детей, пульс может стать незаметным, а конечности холодны; Пациент может пройти в кому . В течение двенадцати до двадцати четырех часов эти тяжелые конституционные симптомы обычно выходят из строя; Но тем временем отек и обесцвечивание широко распространились. Конечность становится флегмонозной и иногда грандиозной. В течение нескольких дней восстановление обычно происходит несколько внезапно, но смерть может быть результатом тяжелой депрессии или от вторичных эффектов наглучения . Эти случаи смерти, как у взрослых, так и у детей нечасты в некоторых частях континента, упоминаются в последней главе этого введения.
Viperidae сильно отличается среди себя в токсичности их ядов. Некоторые, такие как гадюка индийского Рассела ( Daboia Russelli ) и Saw-Scalled Viper ( E. carinatus ); Американские гремучие змеи ( Crotalus spp.), Bushmasters ( Lachesis spp.) и Lanceheads ( Bothrops spp.); и африканские добавки ( Bitis spp.), Ночные добавки ( Causeus spp.) и рогатые гадюки ( Cerastes spp.), вызывают смертельные результаты, если только лекарство не применяется. Укус более крупных европейских гадюков может быть очень опасным, а затем смертельные результаты, особенно у детей, по крайней мере, в более горячих частях континента; Пока маленький луговой гадюк ( Vipera ursinii ), который вряд ли когда -либо кусается, если не обрабатывается, не одержит очень вирулентному яду, и, хотя он очень распространен в некоторых частях Австрии и Венгрии , не вызывал никогда не вызвано серьезный случай.
Опишлифные колубриды
[ редактировать ]Биологи давно знали, что у некоторых змей были задние клыки, «низкие» механизмы инъекции ядом, которые могут обездвигать добычу; Хотя несколько смертельных случаев были зарегистрированы, до 1957 года, вероятность того, что такие змеи были смертельными для людей, казалась наиболее отдаленной. Смерть двух выдающихся герпетологов, Роберта Мертенса и Карла Шмидта , от африканских укусов Колубрида, изменили эту оценку, и недавние события показывают, что у нескольких других видов змей заднего фарнина есть ядовиты, которые потенциально смертельны для крупных позвоночных.
Boomslang ( Dispholidus typus ) и ветки змея ( TheLotornis spp.) Яды токсичны для клетки крови и тонкие кровь (гемотоксичны, геморрагические). Ранние симптомы включают головные боли, тошноту, диарею, летаргию, умственную дезориентацию, синяки и кровотечение на месте и все отверстия для тела. Обливание является основной причиной смерти от такого укуса.
Яд Boomslang-самый мощный из всех змей с задним панкой в мире, основанным на LD 50 . Хотя его яд может быть более сильным, чем некоторые гадюки и элапиды, он вызывает меньшее количество погибших из -за различных факторов (например, эффективность клыков не высока по сравнению со многими другими змеями, доставляемая доза яда низкая, а бумзланги, как правило, меньше меньше. агрессивно по сравнению с другими ядовитыми змеями, такими как кобры и мамба). Симптомы укуса от этих змей включают тошноту и внутреннее кровотечение, и его можно умереть от кровоизлияния в мозг и респираторного коллапса .
Агорные змеи
[ редактировать ]Эксперименты, проведенные с секрецией околоушной железы Rhabdophis показали и Zamenis, , что даже агорные змеи не полностью лишены яда, и указывают на вывод, что физиологическая разница между так называемыми безвредными и яд Поскольку различные шаги существуют в превращении обычной околоушной железы в ядовитую железу или сплошного зуба в трубчатый или рифленый клык.
Использование ядов змеи для лечения заболеваний
[ редактировать ]Учитывая, что змеиный яд содержит много биологически активных ингредиентов, некоторые могут быть полезны для лечения заболевания. [ 42 ]
Например, фосфолипазы типа A2 (PLA2S) из тунисских гадюков Cerastes и Macrovipera Lebetina обладают противоопухолевой активностью. [ 43 ] Целью активности также сообщалась для других соединений в змеином яде. [ 44 ] [ 45 ] Таким образом, гидролипиды PLA2S, таким образом, могут действовать на поверхности бактериальных клеток, обеспечивая новые антимикробные (антибиотики) активность. [ 46 ]
Анальгетическая (убивающая боль ) активность многих белков яда змеи давно известна. [ 47 ] [ 48 ] Основная проблема, однако, заключается в том, как доставить белок в нервные клетки: белки обычно не применимы как таблетки.
Иммунитет
[ редактировать ]Среди змей
[ редактировать ]Вопрос о том, невосприимчивы ли отдельные змеи к своему собственному яду, еще не был окончательно урегулирован, хотя пример известен о кобре, которая самооценка, что приводит к большому абсцессу , требующему хирургического вмешательства, но не показывая ни одного из других эффектов, которые будут иметь Доказано быстро смертельно у видов добычи или людей. [ 49 ] Кроме того, некоторые безвредные виды, такие как североамериканская общая цариская змея ( Lampropeltis getula ) и центральная и южноамериканская муссурана ( Clelia spp.), Являются доказательством яда кроталинов , которые часто встречаются те же районы, и которые они способны Чтобы одолеть и питаться. Куриная змея ( Spilotes pullatus ) является врагом Fer-De-Plance ( Bothrops Caribbaeus ) в Сент-Люсии, и в их встречах куриная змея неизменно является победителем. Повторные эксперименты показали, что европейская травяная змея ( Natrix Natrix ) не затронут укус европейского доклада ( Vipera Berus ) и европейской ASP ( Vipera Aspis ), что связано с присутствием, в крови безвредной змеи из токсичных принципов, секретируемых околоушными и губными железами, и аналогичными принципам яда этих гадюков. Несколько североамериканских видов змей крыс, а также короля змей оказались невосприимчивыми или очень устойчивыми к яду видов гремучей змеи. Говорят, что король кобры, которая добывает добычу на кобрах, является невосприимчивым к их яду.
Среди других животных
[ редактировать ]Ежик , как известно , (Erinaceidae), Mongoose (Herpestidae), медовый барсук ( Mellivora capensis ) и опоссум невосприимчивы к дозе змеиного яда. [ Цитация необходима ] Недавно было обнаружено, что у медового барсука и домашней свиньи были сходящиеся аминокислотные замены в их никотино-ацетилхолиновом рецепторе, которые, как известно, придают устойчивость к альфа-нейротоксинам в ежах. [ 50 ] Может ли свинья считаться невосприимчивой, все еще неясно, хотя ранние исследования показывают эндогенную устойчивость у свиней, протестированных против нейротоксинов. [ 51 ] слой жира свиньи Хотя подкожный может защитить его от яда змеи, большинство ядов легко проходят через слои сосудистого жира, что делает это маловероятным вклад в его способность противостоять ядам. Садовый Дормоз ( Eliomys Quercinus ) был недавно добавлен в список животных, рефрактерных к Viper Venom. Некоторые популяции калифорнийской наземной белки ( Otospermophilus beecheyi ), по крайней мере, частично невосприимчивы к яду гремучей змеи в качестве взрослых.
Среди людей
[ редактировать ]Приобретение человеческого иммунитета против змеиного яда является древним (около 60 г. н.э., племя Псилли ). Исследование разработки вакцин, которые приведут к иммунитету, продолжаются. Билл Хааст , владелец и директор Miami Serpentarium, ввел себя змеиным ядом в течение большей части своей взрослой жизни, пытаясь построить иммунитет широкому кругу ядовитых змей, в практике, известной как митридатизм . Хаст жил до 100 лет и пережил 172 укуса змей. Он пожертвовал свою кровь для лечения пациентов с укусом змеи, когда не было доступно подходящее противоядие. Более 20 человек, получавших себя, выздоровели. [ 52 ] [ 53 ] [ 54 ] Исследователь -любитель Тим Фриде также позволяет ядовитым змеям укусить его в надежде на развитие ядовита змеи на змею, и выжил более 160 укусов от разных видов по состоянию на январь 2016 года. [ 55 ]
Традиционные методы лечения
[ редактировать ]По оценкам Всемирной организации здравоохранения , 80% населения мира зависит от традиционной медицины для их потребностей в первичной медицинской помощи. [ 56 ] Методы традиционных методов лечения змеи, хотя и сомнительной эффективности и, возможно, даже вредных, тем не менее, являются актуальными.
Растения, используемые для обработки змеиных кубиков в Тринидаде и Тобаго, превращаются в настойки с спиртом или оливковым маслом и хранятся в роговых колбах, называемых змеиными бутылками, которые содержат несколько различных растений и/или насекомых. Используемые растения включают в себя лозу, называемую лестницей обезьян ( Bauhinia cumanensis или Bauhinia exisa , Fabaceae), которая забит и наносит укус. В качестве альтернативы, настойка сделана из куска лозы и хранится в бутылке змеи. Другие используемые растения включают корень Mat ( Aristolochia Rugosa ), кошачий коготь ( Pithecellobim unguis-cati ), табак ( Nicotiana tabacum ), куст змей ( Barleria lupulina ), семена Obie ( Cola nitida ) и дикий корень Gri Gri ( Acrocomia aculeata ). Некоторые бутылки змеи также содержат гусеницы ( Battus polydamas , Papilionidae), которые едят листья деревьев ( Aristolochia trilobata ). Аварийные змеиные лекарства получают путем пережевывания трехдюймового куска корня Bois Canôt ( Cecropia peltata ) и введения этого разжеванного корня для укусающего субъекта (обычно охотничьего собаки). Это обычное местное растение Латинской Америки и Карибского бассейна, которое делает его подходящим в качестве аварийного средства. Еще одним местным растением используется Марди Гра ( Ренеалмия Альпиния ) (ягоды), которые измельчаются вместе с соком дикого тростника ( костюма ) и дают укушеному. Быстрые исправления включали в себя применение жевательного табака из сигарет, сигар или труб. [ 57 ] В прошлом в прошлом понижение прокола или высасывание яда полезным, но этот курс лечения теперь сильно обескуражен из-за риска самосознания через порезы или порезы ножа во рту (всасывающие чашки из укуса змеи Наборы могут быть использованы, но всасывание редко обеспечивает какую -либо измеримую выгоду). [ 58 ] [ 59 ]
Серотерапия
[ редактировать ]Серотерапия с использованием антивинома является распространенным текущим лечением и описана еще в 1913 году. [ Примечание 1 ] Как адаптивный иммунитет, так и серотерапия специфичны для типа змеи; Яд с одинаковым физиологическим действием не перекрестно нейтрализуется. Boulenger 1913 описывает следующие случаи:
Европейский в Австралии, который стал невосприимчивым к яду смертельной австралийской тигровой змеи ( Notechis Scutatus ), безнаказанным манипулируя этими змеями и испытывал впечатление, что его иммунитет распространялся и на другие виды, низи укушен когда Superbus ), союзный элапин, умер на следующий день.
В Индии было обнаружено , что сыворотка, приготовленная с ядом моноселированной кобры Наджа Каутия, не влияет на яд двух видов Крайта ( бунгарус ), гадюка Рассела ( Дабоя Расселли ), шкалевой гадюки ( Эхис Каринатус ) Pome's Pit Viper ( Trimeresurus popeiorum ). Сыворотка Russell Viper без влияния на яды колубрина или эхис и тримерсурус .
В Бразилии сыворотка, приготовленная с ядом мрачных головок ( Bothrops spp.), Без действия на гремучую змею ( Crotalus spp.) Venom.
Обработка антивиномочного укубала должна соответствовать типу эневеномации, которая произошла. В Америке доступны поливалентные противоядие, которые эффективны против укусов большинства ямы. Crofab - это противоядия, разработанная для лечения укусов гадюков из Северной Америки. [ 60 ] Они не эффективны против эненомации коралловой змеи , которая требует специфического противоядия для их нейротоксического яда. Ситуация еще более сложна в таких странах, как Индия, с его богатым смесью гадюков (Viperidae) и высоко нейротоксических кобр и Kraits Elapidae.
Примечания
[ редактировать ]- ^ Этот раздел основан на книге 1913 года « Змеи Европы » Га -Бууленгер, которая в настоящее время находится в общественном доступе в Соединенных Штатах (и, возможно, в другом месте) из -за его возраста, текст в этой статье не обязательно должен рассматриваться как как Отражая текущие знания яда змеи
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Исследование яда рептилий» . Австралийский парк рептилий. Архивировано из оригинала 2 февраля 2010 года . Получено 21 декабря 2010 года .
- ^ Jump up to: а беременный в дюймовый и фон Баухот Р. (1994). Змеи: естественная история Нью -Йорк, Нью -Йорк, США: Sterling Publishing Co., Inc. Стр. 194–2 ISBN 978-1-4027-3181-5 .
- ^ Jump up to: а беременный в Хэллидей А., Крайг Т, ред. (2002). Энциклопедия Firefly от рептилий и амфибий . Торонто, Канада: Firefly Books Ltd. с. 202–203 . ISBN 978-1-55297-613-5 .
- ^ Jump up to: а беременный Bottrall JL, Madaras F, Biven CD, Venning MG, Mirtschin PJ (сентябрь 2010 г.). «Протеолитическая активность ядов элапидной и виперидной змеи и ее значение для пищеварения» . Журнал исследований яда . 1 (3): 18–28. PMC 3086185 . PMID 21544178 .
- ^ Jump up to: а беременный в дюймовый Оливейра, Ана Л.; Viegas, Matilde F.; Da Silva, Saulo L.; Соарс, Андреймар М.; Рамос, Мария Дж.; Фернандес, Педро А. (июль 2022 г.). «Химия змеиного яда и его лекарственный потенциал» . Природа обзоры химии . 6 (7): 451–469. Doi : 10.1038/s41570-022-00393-7 . ISSN 2397-3358 . PMC 9185726 . PMID 35702592 .
- ^ Mattison C (2007). Новая энциклопедия змей . Нью -Джерси, США (впервые опубликовано в Великобритании): издательство Принстонского университета (Принстон и Оксфорд) впервые опубликовано в Бландфорде. п. 117. ISBN 978-0-691-13295-2 .
- ^ Cardoso KC, Da Silva MJ, Costa GG, Torres TT, Del Bem Le, Vidal RO, et al. (Октябрь 2010). «Транскриптомный анализ экспрессии генов в ядовитой железе змеи Bothrops Alternatus (Urutu)» . BMC Genomics . 11 (1): 605. doi : 10.1186/1471-2164-11-605 . PMC 3017861 . PMID 20977763 .
- ^ Condrea E, Devries A, Mager J (февраль 1964 г.). «Гемолиз и расщепление фосфолипидов эритроцитов человека яду змеи». Biochimica et Biophysica Acta (BBA) - Специализированный раздел на липидах и связанных с ними субъектов . 84 (1): 60–73. doi : 10.1016/0926-6542 (64) 90101-5 . PMID 14124757 .

- ^ Rodríguez-ithurralde D, Silveira R, Barbeito L, Dajas F (1983). «Фаскулин, в мощном антихолинэстеразной полипептиде из яда Denroaspis angusticeps». Нейрохимия International . 5 (3): 267–74. Doi : 10.1016/0197-0186 (83) 90028-1 . PMID 20487949 . S2CID 8952817 .
- ^ Макси, Стивен П. (1 мая 2021 г.). Макси, Стивен П. (ред.). Справочник по ядам и токсинам рептилий (2 изд.). Второе издание. | Boca Raton: CRC Press, 2021.: CRC Press. doi : 10.1201/9780429054204 . ISBN 978-0-429-05420-4 .
{{cite book}}: CS1 Maint: местоположение ( ссылка ) - ^ Jump up to: а беременный «Ключевое слово: геморрагический токсин KW-1200» . Uniprot . Получено 1 июня 2019 года .
- ^ Hodgson WC, Wickramaratna JC (сентябрь 2002 г.). «Нервно -мышечная активность in vitro у змеиных ядов» . Клиническая и экспериментальная фармакология и физиология . 29 (9): 807–14. doi : 10.1046/j.1440-1681.2002.03740.x . PMID 12165047 . S2CID 20158638 .
- ^ Он Yy, Lee WH, Zhang Y (сентябрь 2004 г.). «Клонирование и очистка альфа-нейротоксинов из короля кобры (Ophiophagus hannah)». Токсикона . 44 (3): 295–303. doi : 10.1016/j.toxicon.2004.06.003 . PMID 15302536 .
- ^ Bernardoni JL, Sousa LF, Wermelinger LS, Lopes AS, Prezoto BC, Serrano SM, Zingali RB, Moura-Da-Silva AM (14 октября 2014 г.). «Функциональная изменчивость металлопротеиназ Snake Venom: адаптивные преимущества в нацеливании на различную добычу и последствия для эненомации человека» . Plos один . 9 (10): E109651. BIBCODE : 2014PLOSO ... 9J9651B . doi : 10.1371/journal.pone.0109651 . PMC 4196926 . PMID 25313513 .
- ^ Hermans C, Wittevrongel C, Thys C, Smethurst PA, Van Geet C, Freson K (август 2009 г.). «Составная гетерозиготная мутация в гликопротеине VI у пациента с кровоточащим расстройством» . Журнал тромбоза и гемостаза . 7 (8): 1356–63. doi : 10.1111/j.1538-7836.2009.03520.x . PMID 19552682 . S2CID 205728095 .

- ^ Griffin PR, AIRD SD (ноябрь 1990 г.). «Новый маленький миотоксин из яда прерии гремучей змеи (Crotalus viridis viridis)» . Письма Febs . 274 (1–2): 43–47. doi : 10.1016/0014-5793 (90) 81325-I . PMID 2253781 . S2CID 45019479 .
- ^ Somejima Y, Aoki Y, Mebs D (1991). «Аминокислотная последовательность миотоксина из яда Восточного Алмаза Гремучей змеи (Crotalus Adamanteus)». Токсикона . 29 (4–5): 461–468. doi : 10.1016/0041-0101 (91) 90020-r . PMID 1862521 .
- ^ Уиттингтон CM, Papenfuss AT, Bansal P, Torres AM, Wong ES, Deakin JE, et al. (Июнь 2008 г.). «Дефенсины и конвергентная эволюция генов яда и рептилий» . Исследование генома . 18 (6): 986–994. doi : 10.1101/gr.7149808 . PMC 2413166 . PMID 18463304 .
- ^ Широкий AJ, Сазерленд SK, Coulter AR (17 мая 1979 г.). «Смертность у мышей опасного австралийского и другого змею -яда» (PDF) . Токсикона . 17 (6): 661–4. doi : 10.1016/0041-0101 (79) 90245-9 . PMID 524395 .
- ^ Розен Ф.С. (31 июля 2003 г.). «Эдвин Дж. Кон и развитие химии белка». Новая Англия Журнал медицины . 349 (5): 511–512. doi : 10.1056/nejm200307313490522 .
- ^ Jump up to: а беременный в дюймовый Matejtschuk P, Dash CH, Gascoigne EW (декабрь 2000 г.). «Производство решения человеческого альбумина: постоянно развивающийся коллоид» . Британский журнал анестезии . 85 (6): 887–95. doi : 10.1093/bja/85.6.887 . PMID 11732525 .
- ^ Brodniewicz-Proba T (декабрь 1991 г.). «Фракционирование плазмы человека и влияние новых технологий на использование и качество продуктов, полученных из плазмы». Обзоры крови . 5 (4): 245–57. doi : 10.1016/0268-960x (91) 90016-6 . PMID 1782484 .
- ^ Jump up to: а беременный в дюймовый и фон глин Fry Bg, Casewell NR, Wüster W, Vidal N, Young B, Jackson TN (сентябрь 2012 г.). «Структурная и функциональная диверсификация системы яд рептилий Toxicofera». Токсикона . 60 (4): 434–48. doi : 10.1016/j.toxicon.2012.02.013 . PMID 22446061 .
- ^ Jump up to: а беременный в дюймовый Casewell NR, Wüster W, Vonk FJ, Harrison RA, Fry BG (апрель 2013 г.). «Сложные коктейли: эволюционная новинка ядов». Тенденции в экологии и эволюции . 28 (4): 219–29. doi : 10.1016/j.tree.2012.10.020 . PMID 23219381 .
- ^ Lomonte B, Fernández J, Sanz L, Angulo Y, Sasa M, Gutiérrez JM, Calvete JJ (июнь 2014 г.). «Ядовитые змеи Коста -Рики: биологические и медицинские последствия их протеомных профилей яда, проанализированные посредством стратегии ядных змеи». Журнал протеомики . 105 : 323–39. doi : 10.1016/j.jprot.2014.02.020 . PMID 24576642 .
- ^ Hargreaves, Adam D.; Суэйн, Мартин Т.; Хегарти, Мэтью Дж.; Логан, Даррен В.; Малли, Джон Ф. (август 2014 г.). «Ограничение и набор - генеральное дублирование и происхождение и эволюция токсинов яда змеи» . Биология и эволюция генома . 6 (8): 2088–2095. doi : 10.1093/gbe/evu166 . ISSN 1759-6653 . PMC 4231632 . PMID 25079342 .
- ^ Lynch VJ (январь 2007 г.). «Изобретение арсенала: адаптивная эволюция и неофункционализация генов фосфолипазы A2 змеи -яда» . BMC Эволюционная биология . 7 (2): 2. DOI : 10.1186/1471-2148-7-2 . PMC 1783844 . PMID 17233905 .
- ^ Barlow A, Pook CE, Harrison RA, Wüster W (июль 2009 г.). «Коаволюция диеты и специфичной для добычи активности яда подтверждают роль отбора в эволюции яда змеи» . Разбирательство. Биологические науки . 276 (1666): 2443–2449. doi : 10.1098/rspb.2009.0048 . PMC 2690460 . PMID 19364745 .
- ^ Пахари С., Бикфорд Д., Фрай Б.Г., Кини Р.М. (сентябрь 2007 г.). «Паттерн экспрессии генов токсина и фосфолипазы А2 фосфолипазы в ядных железах двух морских змей, лапемиса и акалиптофиса Peronii: Сравнение эволюции этих токсинов в сухопутных змеях, морских Kraits и морских змей» . BMC Эволюционная биология . 7 : 175. DOI : 10.1186/1471-2148-7-175 . PMC 2174459 . PMID 17900344 .
- ^ Jump up to: а беременный Barlow A, Pook CE, Harrison RA, Wüster W (июль 2009 г.). «Коаволюция диеты и специфичной для добычи активности яда подтверждают роль отбора в эволюции яда змеи» . Труды: биологические науки . 276 (1666): 2443–9. doi : 10.1098/rspb.2009.0048 . JSTOR 30244073 . PMC 2690460 . PMID 19364745 .
- ^ Янса С.А., Восс Р.С. (22 июня 2011 г.). «Адаптивная эволюция белка VWF, нацеленного на яд, в опоссумах, которые едят питвийперы» . Plos один . 6 (6): E20997. Bibcode : 2011ploso ... 620997j . doi : 10.1371/journal.pone.0020997 . PMC 3120824 . PMID 21731638 .
- ^ Calvete JJ, Ghezellou P, Paiva O, Matainaho T, Ghassempour A, Goudarzi H, Kraus F, Sanz L, Williams DJ (июль 2012 г.). «Снейк -ядом двух плохо известных гидрофийей: сравнительная протеомика ядов наземных токсикокаламусов Longissimus и морского гидрофиса Cyanocinctus». Журнал протеомики . 75 (13): 4091–101. doi : 10.1016/j.jprot.2012.05.026 . PMID 22643073 .
- ^ Li M, Fry Bg, Kini RM (январь 2005 г.). «Диета только для яиц: ее последствия для изменений профиля токсина и экологии мраморной морской змеи (Aipysurus eydouxii)». Журнал молекулярной эволюции . 60 (1): 81–9. Bibcode : 2005jmole..60 ... 81L . doi : 10.1007/s00239-004-0138-0 . PMID 15696370 . S2CID 17572816 .
- ^ Mackessy SP (июль 2010 г.). «Эволюционные тенденции в композиции яда в западных гремучих змеях (Crotalus viridis sensu lato): токсичность против тендеров». Токсикона . 55 (8): 1463–74. doi : 10.1016/j.toxicon.2010.02.028 . PMID 20227433 .
- ^ Saviola AJ, Chiszar D, Busch C, Mackessy SP (март 2013 г.). «Молекулярная основа для перемещения добычи у випер -змей» . BMC Biology . 11 (1): 20. doi : 10.1186/1741-7007-11-20 . PMC 3635877 . PMID 23452837 .
- ^ Янгман, Николас Дж; Llinas, Джошуа; Хаворт, Марк; Гиллетт, Эмбер; Джонс, Ли; Уокер, Эндрю А; Фрай, Брайан Дж (июнь 2022 г.). «Распутание взаимодействия между Bitis Vipers и их добычей с использованием коагулотоксичности против разнообразной плазмы позвоночных» . Токсикона . 216 : 37–44. doi : 10.1016/j.toxicon.2022.06.012 . ISSN 1879-3150 . PMID 35780972 . S2CID 250188352 .
- ^ Дэвис, Эмма-Луиза; Арбакл, Кевин (декабрь 2019 г.). «Коэволюция токсической активности и диеты с ядом змеи: доказательства того, что экологический общий общий способ предпочитает токсикологическое разнообразие» . Токсины . 11 (12): 711. doi : 10.3390/toxins11120711 . ISSN 2072-6651 . PMC 6950196 . PMID 31817769 .
- ^ McCue MD (октябрь 2007 г.). «Добыча эненомация не улучшает пищеварительные характеристики в западных бриллиантовых гремучих змеях (Crotalus atrox)». Журнал экспериментальной зоологии, часть а . 307 (10): 568–77. doi : 10.1002/jez.411 . PMID 17671964 .
- ^ Broeckhoven C, Du Plessis A (август 2017 г.). «Разве Snake Fang Evolution потеряла свой укус? Новые идеи с точки зрения структурной механики» . Биологические письма . 13 (8): 20170293. DOI : 10.1098/rsbl.2017.0293 . PMC 5582107 . PMID 28768797 .
- ^ Cleuren SG, Hocking DP, Evans AR (июнь 2021 г.). «Эволюция клыка у ядовитых змей: адаптация формы трехмерного зуба к биомеханическим свойствам их добычи» . Эволюция; Международный журнал органической эволюции . 75 (6): 1377–1394. doi : 10.1111/evo.14239 . PMID 33904594 . S2CID 233411378 .
- ^ Martin CJ, Lamb G (1907). "Змеиная и укусовая змея". В Allbutt TC, Rolleston ND (Eds.). Система медицины . Лондон: Макмиллан. С. 783–821.
- ^ МакКлири Р.Дж., Кини Р.М. (февраль 2013 г.). «Незенциатические белки из ядов змей: золотой рудник фармакологических инструментов и лекарств». Токсикона . 62 : 56–74. doi : 10.1016/j.toxicon.2012.09.008 . PMID 23058997 .
- ^ Zouari-Kessentinini R, Srairi-Abid N, Bazaa Ayeb M, Luis J, Merrakchi N (2013). «Противоотвержденный потенциал ядов тунисских змей секретируется фосфолией A2 » Biomed Research International 2013 : 1–9 Doi : 10.1155/ 2013/3 PMC 35812 PMID 23509718
- ^ Вьяс В.К., Брамбхатт К., Бхатт Х, Пармар У (февраль 2013 г.). «Терапевтический потенциал змеиного яда при терапии рака: текущие перспективы» . Азиатский тихоокеанский журнал тропической биомедицины . 3 (2): 156–62. doi : 10.1016/s2221-1691 (13) 60042-8 . PMC 3627178 . PMID 23593597 .
- ^ Jain D, Kumar S (2012). "Snake Venom: мощный противоопухолевый агент" . Азиатско -Тихоокеанский журнал профилактики рака . 13 (10): 4855–60. doi : 10.7314/apjcp.2012.13.10.4855 . PMID 23244070 .
- ^ De Oliveira Junior NG, E Silva Cardoso MH, Franco OL (декабрь 2013 г.). «Змеиные яды: привлекательные антимикробные белковые соединения для терапевтических целей» . Клеточные и молекулярные науки о жизни . 70 (24): 4645–58. doi : 10.1007/s00018-013-1345-x . PMC 11113393 . PMID 23657358 . S2CID 15127065 .
- ^ Вульф CJ (январь 2013 г.). «Боль: морфин, метаболиты, мамба и мутации». Lancet. Неврология . 12 (1): 18–20. doi : 10.1016/s1474-4422 (12) 70287-9 . PMID 23237896 . S2CID 8697382 .
- ^ Осипов А., Утки Ю (декабрь 2012 г.). «Влияние полипептидов яда змеи на центральную нервную систему». Центральная нервная система агентов в лекарственной химии . 12 (4): 315–28. doi : 10.2174/187152412803760618 . PMID 23270323 . S2CID 36274766 .
- ^ «Стерильный хвостовой абсцесс в Naja Annulifera - случай самооценка» . Архивировано из оригинала 27 октября 2004 года . Получено 2 апреля 2009 года .
- ^ Drabeck D, Jansa S (2015). «Почему медовый барсук не заботится: независимая эволюция устойчивости к трем токсинам пальца в никотиновом рецепторе ацетилхолина». Токсикона . 99 : 68–72. doi : 10.1016/j.toxicon.2015.03.007 . PMID 25796346 .
- ^ Grasset E, Zoutendykanda A, Schaafsma A (1935). «Исследования токсичных и антигенных свойств южноафриканских змеиных ядов со специальным отслеживанием к поливалентности южноафриканской антидионелен». Транс. R. Soc. Троп. Медик Гигиена . 28 (6): 601–612. doi : 10.1016/s0035-9203 (35) 90031-1 .
- ^ «Прощай с этими знаменитыми флоридцами» . Флоридская тенденция . 19 декабря 2011 года . Получено 2 апреля 2012 года .
- ^ Розенберг С (21 июня 2011 г.). «Билл Хааст умирает на 100; змей были очарованием знаменитости Южной Флориды» . Los Angeles Times . Получено 16 октября 2012 года .
- ^ Шудель М (18 июня 2011 г.). «Билл Хааст умирает в 100: Florida Snake Man предоставил яд для сыворотки Snakebite» . The Washington Post . Получено 16 октября 2012 года .
- ^ «Человек заставляет смертельных змей укусить его 160 раз в охоте на человеческое противоядие | Америка | Новости | Независимые» . Independent.co.uk . 21 января 2016 года . Получено 7 июля 2016 года .
- ^ Hiremath VT, Taranath TC (февраль 2010 г.). «Традиционная фитотерапия для укусов змей племенами района Читрадурга, штат Карнатака, Индия» . Этноботанические листочки . 14 (2): 120–125.
- ^ Zethelius M, Balick MJ (март 1982 г.). «Современная медицина и шаманский ритуал: случай позитивного синергетического ответа при лечении укуса змеи» (PDF) . Журнал этнофармакологии . 5 (2): 181–5. doi : 10.1016/0378-8741 (82) 90042-3 . PMID 7057657 .
- ^ «Лечение укусов змей» . Ces.ncsu.edu . Получено 16 октября 2012 года .
- ^ «CDC - Ядовитые змеи - тема безопасности и здоровья Niosh» . CDC.gov. 1 июля 2016 года . Получено 7 июля 2016 года .
- ^ http://www.savagelabs.com/products/crofab/home/crofab_frame.htm Архивировано 3 марта 2016 года по ссылке машины Wayback на PDF для полной информации, полученной 11/12/12
Дальнейшее чтение
[ редактировать ]- Jonassen I, Collins JF, Higgins DG (август 1995). «Поиск гибких паттернов в невыполненных белковых последовательностях» . Белковая наука . 4 (8): 1587–95. doi : 10.1002/pro.5560040817 . PMC 2143188 . PMID 8520485 .
- Шоу IC (2007). «Глава 19: Змеиные токсины». В Waring RH, Steventon GB, Mitchell SC (Eds.). Молекулы смерти (второе изд.). Ривер Эдж, Нью -Джерси: Имперская колледж Пресс. С. 329 –344. ISBN 978-1-86094-815-2 .
Внешние ссылки
[ редактировать ]- Обзор разнообразия и эволюции змеиных клыков.
- Змеиные яды Умич Ориентация белков в мембранах семейства/суперсемейства -55 - Расчетная ориентация фосфолипаз змеи А2 и миотоксинов в липидном бислое.
- LD50 для большинства токсичных ядов.
- Австралийский исследовательский подразделение - общий источник информации для ядовитых существ в Австралии.
- BiomedCentral.com - Лекарственные и этноветеринарные средства охотников в Тринидаде .
- reptilis.net - Как работает Venom.
- Snakevenom.net - сушка и хранение змеиного яда.