Клеточная биология
| Часть серии о |
| Биология |
|---|
Клеточная биология (также клеточная биология или цитология ) — раздел биологии , изучающий структуру , функции и поведение клеток . [1] [2] Все живые организмы состоят из клеток. Клетка – это основная единица жизни, которая отвечает за жизнь и функционирование организмов. [3] Клеточная биология — это изучение структурных и функциональных единиц клеток. Клеточная биология охватывает как прокариотические , так и эукариотические клетки и имеет множество подтем, которые могут включать изучение клеточного метаболизма , клеточной коммуникации , клеточного цикла , биохимии и клеточного состава . Исследование клеток проводится с использованием нескольких микроскопии методов , культуры клеток и фракционирования клеток . Они позволили и в настоящее время используются для открытий и исследований, касающихся функционирования клеток, что в конечном итоге дает понимание более крупных организмов. Знание компонентов клеток и того, как они работают, имеет фундаментальное значение для всех биологических наук, а также важно для исследований в биомедицинских областях, таких как рак и другие заболевания. Исследования в области клеточной биологии взаимосвязаны с другими областями, такими как генетика , молекулярная генетика , молекулярная биология , медицинская микробиология , иммунология и цитохимия .
История [ править ]
в 17 веке Клетки впервые были обнаружены в Европе с изобретением сложного микроскопа . В 1665 году Роберт Гук назвал строительные блоки всех живых организмов «клетками» (опубликовано в Micrographia ) после того, как посмотрел на кусок пробки и увидел клеточную структуру; [4] [5] однако клетки были мертвы. Они не дали никаких указаний на фактические общие компоненты клетки. Несколько лет спустя, в 1674 году, Антон Ван Левенгук первым проанализировал живые клетки при исследовании водорослей . Много лет спустя, в 1831 году, Роберт Браун открыл ядро . Все это предшествовало клеточной теории , которая утверждает, что все живые существа состоят из клеток и что клетки являются функциональными и структурными единицами организмов. В конечном итоге к такому выводу пришел ученый-растениевод Маттиас Шлейден. [5] и ученый-зоотехник Теодор Шванн в 1838 году, который изучал живые клетки в тканях растений и животных соответственно. [3] 19 лет спустя Рудольф Вирхов внес дальнейший вклад в клеточную теорию, добавив, что все клетки возникают в результате деления ранее существовавших клеток. [3] Вирусы не рассматриваются в клеточной биологии — им не хватает характеристик живой клетки, и вместо этого они изучаются в микробиологическом подклассе вирусологии . [6]
Техники [ править ]
Исследования клеточной биологии рассматривают различные способы культивирования и манипулирования клетками вне живого организма для дальнейших исследований в области человека анатомии и физиологии , а также для получения лекарств. Методы изучения клеток развивались. Благодаря достижениям в области микроскопии, методов и технологий учёным удалось лучше понять структуру и функции клеток. Ниже перечислены многие методы, обычно используемые для изучения клеточной биологии: [7]
- Культура клеток : использует быстро растущие клетки на среде, что позволяет получить большое количество клеток определенного типа и является эффективным способом изучения клеток. [8] Культура клеток является одним из основных инструментов, используемых в клеточной и молекулярной биологии, предоставляя превосходные модельные системы для изучения нормальной физиологии и биохимии клеток (например, метаболические исследования, старение), воздействия лекарств и токсичных соединений на клетки, а также мутагенеза. и канцерогенез. Он также используется при проверке и разработке лекарств, а также при крупномасштабном производстве биологических соединений (например, вакцин, терапевтических белков).
- Флуоресцентная микроскопия : флуоресцентные маркеры, такие как GFP , используются для маркировки определенного компонента клетки. После этого свет определенной длины волны используется для возбуждения флуоресцентного маркера, который затем можно визуализировать. [8]
- Фазово-контрастная микроскопия : использует оптический аспект света для представления изменений твердой, жидкой и газовой фаз в виде различий в яркости. [8]
- Конфокальная микроскопия : сочетает флуоресцентную микроскопию с визуализацией путем фокусировки света и моментальной съемки для формирования трехмерного изображения. [8]
- Просвечивающая электронная микроскопия : включает окрашивание металлов и прохождение электронов через клетки, которые отклоняются при взаимодействии с металлом. В конечном итоге это формирует изображение изучаемых компонентов. [8]
- Цитометрия : клетки помещаются в машину, которая использует луч для рассеивания клеток по различным аспектам и, следовательно, может разделять их по размеру и содержимому. Клетки также можно пометить флуоресценцией GFP и разделить таким же образом. [9]
- Фракционирование клеток . Этот процесс требует разрушения клеток с использованием высокой температуры или обработки ультразвуком с последующим центрифугированием для разделения частей клетки, что позволяет изучать их отдельно. [8]
Типы ячеек [ править ]

Существует две фундаментальные классификации клеток: прокариотические и эукариотические . Прокариотические клетки отличаются от эукариотических клеток отсутствием клеточного ядра или других мембраносвязанных органелл . [10] Прокариотические клетки намного меньше эукариотических клеток, что делает их самой маленькой формой жизни. [11] Прокариотические клетки включают бактерии и археи и не имеют закрытого клеточного ядра. Эукариотические клетки встречаются у растений, животных, грибов и простейших. Их диаметр составляет от 10 до 100 мкм, а их ДНК содержится в мембраносвязанном ядре. Эукариоты – это организмы, содержащие эукариотические клетки. Четыре эукариотических царства — это Animalia, Plantae, Fungi и Protista. [12]
Они оба размножаются посредством бинарного деления . Бактерии, наиболее распространенный тип, имеют несколько различных форм , хотя большинство из них имеют сферическую или палочковидную форму . Бактерии можно разделить на грамположительные и грамотрицательные в зависимости от состава клеточной стенки . Грамположительные бактерии имеют более толстый слой пептидогликана, чем грамотрицательные бактерии. Структурные особенности бактерий включают жгутик , который помогает клетке двигаться, [13] рибосомы для перевода РНК в белок, [13] и нуклеоид , который удерживает весь генетический материал в кольцевой структуре. [13] В прокариотических клетках происходит множество процессов, которые позволяют им выживать. У прокариот синтез мРНК инициируется с промоторной последовательности на матрице ДНК, содержащей две консенсусные последовательности, которые рекрутируют РНК-полимеразу. Прокариотическая полимераза состоит из основного фермента, состоящего из четырех белковых субъединиц, и σ-белка, который способствует только инициации. Например, в процессе, называемом конъюгацией , фактор фертильности позволяет бактериям обладать пилусом, который позволяет им передавать ДНК другим бактериям, у которых отсутствует фактор F, обеспечивая передачу устойчивости, позволяющую им выживать в определенных средах. [14]
Структура и функции [ править ]
Строение эукариотических клеток [ править ]
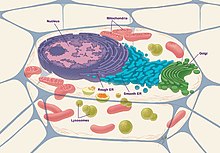
Эукариотические клетки состоят из следующих органелл:
- Ядро : Ядро клетки функционирует как геном и хранилище генетической информации клетки, содержащее всю ДНК, организованную в виде хромосом . Он окружен ядерной оболочкой , которая включает в себя ядерные поры, позволяющие транспортировать белки между внутренней и внешней частью ядра. [15] Это также сайт репликации ДНК и транскрипции ДНК в РНК. После этого РНК модифицируется и транспортируется в цитозоль для трансляции в белок. [16]
- Ядрышко : Эта структура находится внутри ядра, обычно плотная и сферическая. Это место синтеза рибосомальной РНК (рРНК), которая необходима для сборки рибосом.
- Эндоплазматическая сеть (ЭР) : функция синтеза, хранения и секреции белков для аппарата Гольджи. [17] Структурно эндоплазматическая сеть представляет собой сеть мембран, расположенных по всей клетке и связанных с ядром. Мембраны немного различаются от клетки к клетке, и функция клетки определяет размер и структуру ЭР. [18]
- Митохондрии : широко известные как электростанция клетки, представляют собой клеточные органеллы, связанные с двойной мембраной. [19] Это функция для производства энергии или АТФ внутри клетки. В частности, это место, где происходит цикл Кребса или цикл ТСА для производства НАДН и ФАДН. Впоследствии эти продукты используются в цепи переноса электронов (ETC) и окислительном фосфорилировании для окончательного производства АТФ. [20]
- Аппарат Гольджи : он предназначен для дальнейшей обработки, упаковки и секреции белков по назначению. Белки содержат сигнальную последовательность, которая позволяет аппарату Гольджи распознать и направить ее в нужное место. Аппарат Гольджи также производит гликопротеины и гликолипиды . [21]
- Лизосома : функция лизосомы заключается в разложении материала, поступающего извне клетки или старых органелл. Он содержит множество кислых гидролаз, протеаз, нуклеаз и липаз, которые расщепляют различные молекулы. Аутофагия — это процесс деградации через лизосомы, который происходит, когда везикула отпочковывается от ЭР и поглощает материал, а затем прикрепляется и сливается с лизосомой, позволяя материалу разлагаться. [22]
- Рибосомы : функции перевода РНК в белок. он служит местом синтеза белка. [23]
- Цитоскелет : Цитоскелет — это структура, которая помогает поддерживать форму и общую организацию цитоплазмы. Он закрепляет органеллы внутри клеток и обеспечивает структуру и стабильность клетки. Цитоскелет состоит из трех основных типов белковых нитей: актиновых нитей, промежуточных филаментов и микротрубочек, которые удерживаются вместе и связаны с субклеточными органеллами и плазматической мембраной с помощью множества вспомогательных белков. [24]
- Клеточная мембрана : Клеточная мембрана может быть описана как фосфолипидный бислой и также состоит из липидов и белков. [13] Поскольку внутренняя часть бислоя гидрофобна, и для того, чтобы молекулы могли участвовать в реакциях внутри клетки, они должны иметь возможность пересекать этот мембранный слой и проникать в клетку через осмотическое давление , диффузию , градиенты концентрации и мембранные каналы. [25]
- Центриоли : Функция производства веретенообразных волокон, которые используются для разделения хромосом во время деления клеток.
Эукариотические клетки также могут состоять из следующих молекулярных компонентов:
- Хроматин : Он составляет хромосомы и представляет собой смесь ДНК с различными белками.
- Реснички : они помогают продвигать вещества, а также могут использоваться для сенсорных целей. [26]
Клеточный метаболизм [ править ]
Клеточный метаболизм необходим для производства энергии для клетки и, следовательно, ее выживания, и включает в себя множество путей, а также поддержание основных клеточных органелл, таких как ядро, митохондрии, клеточная мембрана и т. д. Для клеточного дыхания , как только глюкоза становится доступной, происходит гликолиз. происходит в цитозоле клетки с образованием пирувата. Пируват подвергается декарбоксилированию с использованием мультиферментного комплекса с образованием ацетил-КоА, который можно легко использовать в цикле ТСА для производства НАДН и ФАДН 2 . Эти продукты участвуют в цепи переноса электронов , образуя в конечном итоге протонный градиент через внутреннюю мембрану митохондрий. Этот градиент может затем стимулировать выработку АТФ и H 2 O при окислительном фосфорилировании . [27] Метаболизм в растительных клетках включает фотосинтез , который является полной противоположностью дыхания, поскольку в конечном итоге производит молекулы глюкозы.
Передача сигналов сотовой связи [ править ]
Передача сигналов или клеточная коммуникация важна для регуляции клеток, а также для того, чтобы клетки обрабатывали информацию из окружающей среды и реагировали соответствующим образом. Передача сигналов может происходить посредством прямого контакта с клетками или посредством эндокринной , паракринной и аутокринной передачи сигналов . Прямой межклеточный контакт – это когда рецептор на клетке связывает молекулу, которая прикрепляется к мембране другой клетки. Эндокринная передача сигналов происходит через молекулы, секретируемые в кровоток. Паракринная передача сигналов использует для общения молекулы, диффундирующие между двумя клетками. Аутокрин — это клетка, посылающая самой себе сигнал, секретируя молекулу, которая связывается с рецептором на ее поверхности. Формами общения могут быть:
- Ионные каналы : могут быть разных типов, например, потенциал- или лиганд-управляемые ионные каналы. Они обеспечивают отток и приток молекул и ионов.
- Рецептор, связанный с G-белком (GPCR): широко признано, что он содержит семь трансмембранных доменов. Лиганд связывается с внеклеточным доменом, и как только лиганд связывается, это сигнализирует фактору обмена гуанина, который преобразует GDP в GTP и активирует субъединицу G-α. G-α может нацеливаться на другие белки, такие как аденилциклаза или фосфолипаза C, которые в конечном итоге продуцируют вторичные мессенджеры, такие как цАМФ, Ip3, DAG и кальций. Эти вторичные мессенджеры усиливают сигналы и могут воздействовать на ионные каналы или другие ферменты. Одним из примеров усиления сигнала является связывание цАМФ и активация ПКА путем удаления регуляторных субъединиц и высвобождения каталитической субъединицы. Каталитическая субъединица имеет последовательность ядерной локализации, которая побуждает ее проникать в ядро и фосфорилировать другие белки, чтобы либо подавить, либо активировать активность гена. [27]
- Рецепторные тирозинкиназы : связывают факторы роста, дополнительно стимулируя перекрестное фосфорилирование тирозина во внутриклеточной части белка. Фосфорилированный тирозин становится посадочной площадкой для белков, содержащих домен SH2, что позволяет активировать Ras и вовлечь путь MAP-киназы . [28]
Рост развитие и
Эукариотический клеточный цикл [ править ]

Клетки являются основой всех организмов и являются фундаментальными единицами жизни. Рост и развитие клеток необходимы для поддержания хозяина и выживания организма. В ходе этого процесса клетка проходит этапы клеточного цикла и развития, которые включают рост клетки, репликацию ДНК , деление клетки , регенерацию и смерть клетки .
Клеточный цикл делится на четыре отдельные фазы : G1, S, G2 и M. Фаза G, которая является фазой роста клеток, составляет примерно 95% цикла. Пролиферация клеток индуцируется предшественниками. Все клетки изначально имеют одинаковую форму и могут стать клетками любого типа. Передача сигналов в клетках, такая как индукция, может влиять на близлежащие клетки, чтобы определить тип клетки, которой они станут. Более того, это позволяет однотипным клеткам агрегироваться и образовывать ткани, затем органы и, наконец, системы. Фазы G1, G2 и S (репликация, повреждение и восстановление ДНК) считаются интерфазной частью цикла, а фаза М ( митоз ) — деления клеток частью цикла . Митоз состоит из многих стадий, которые включают профазу, метафазу, анафазу, телофазу и цитокинез соответственно. Конечным результатом митоза является образование двух одинаковых дочерних клеток.
Клеточный цикл регулируется в контрольных точках клеточного цикла с помощью ряда сигнальных факторов и комплексов, таких как циклины, циклин-зависимая киназа и р53 . Когда клетка завершает процесс роста и обнаруживается, что она повреждена или изменена, она подвергается клеточной гибели либо путем апоптоза , либо некроза , чтобы устранить угрозу, которую она может создать для выживания организма. [29]
Клеточная смертность, линий бессмертие клеточных
Предки каждой современной клетки, по-видимому, ведут непрерывную линию на протяжении более 3 миллиардов лет к зарождению жизни . являются не клетки, На самом деле бессмертными а клеточные линии, состоящие из нескольких поколений. [30] Бессмертие клеточной линии зависит от сохранения потенциала клеточного деления . Этот потенциал может быть утерян в любой конкретной линии из-за повреждения клеток, терминальной дифференцировки , как это происходит в нервных клетках, или запрограммированной гибели клеток ( апоптоза ) во время развития. Поддержание потенциала клеточного деления на протяжении последующих поколений зависит от предотвращения и точного восстановления клеточных повреждений, особенно повреждений ДНК . В половых организмах непрерывность зародышевой линии зависит от эффективности процессов предотвращения повреждений ДНК и восстановления тех повреждений ДНК , которые действительно происходят. Половые процессы у эукариот , как и у прокариот , открывают возможность эффективной репарации повреждений ДНК в зародышевой линии путем гомологичной рекомбинации . [30] [31]
Фазы клеточного цикла [ править ]
Клеточный цикл — это четырехэтапный процесс, который проходит клетка по мере своего развития и деления. Он включает Gap 1 (G1), синтез (S), Gap 2 (G2) и митоз (M). Клетка либо перезапускает цикл с G1, либо выходит из цикла через G0 после завершения цикла. Клетка может прогрессировать от G0 через терминальную дифференцировку.
Интерфаза относится к фазам клеточного цикла, которые происходят между одним митозом и следующим, и включает G1, S и G2.
Фаза G1 [ править ]
Размер клетки увеличивается.
Содержимое клеток реплицируется.
S-фаза [ править ]
Репликация ДНК
В клетке реплицируется каждая из 46 хромосом (23 пары).
Фаза G2 [ править ]
Клетка размножается.
При подготовке к делению клеток образуются органеллы и белки.
Фаза М [ править ]
После митоза происходит цитокинез (разделение клеток).
Образование двух дочерних клеток, идентичных
Фаза G0 [ править ]
Эти клетки покидают G1 и переходят в G0, стадию покоя. Клетка в G0 выполняет свою работу, не готовясь к делению. [32]
Патология [ править ]
Научная отрасль, изучающая и диагностирующая заболевания на клеточном уровне, называется цитопатологией . Цитопатология обычно используется на образцах свободных клеток или фрагментов тканей, в отличие от патологического раздела гистопатологии , который изучает целые ткани. Цитопатология обычно используется для исследования заболеваний, поражающих широкий спектр участков тела, часто для диагностики рака, а также для диагностики некоторых инфекционных заболеваний и других воспалительных состояний. Например, распространенным применением цитопатологии является мазок Папаниколау , скрининговый тест, используемый для выявления рака шейки матки , а также предраковых поражений шейки матки , которые могут привести к раку шейки матки. [33]
точки клеточного цикла и система восстановления ДНК повреждений Контрольные
Клеточный цикл состоит из ряда четко упорядоченных последовательных стадий, которые приводят к клеточному делению. Тот факт, что клетки не переходят к следующему этапу, пока не завершится последний, является важным элементом регуляции клеточного цикла. Контрольные точки клеточного цикла — это характеристики, которые представляют собой отличную стратегию мониторинга для точного клеточного цикла и делений. Cdks, связанные аналоги циклина, протеинкиназы и фосфатазы регулируют рост и деление клеток от одной стадии к другой. [34] Клеточный цикл контролируется временной активацией Cdks, которая регулируется партнерским взаимодействием циклинов, фосфорилированием определенными протеинкиназами и дефосфорилированием фосфатазами семейства Cdc25. В ответ на повреждение ДНК реакция восстановления ДНК клетки представляет собой каскад сигнальных путей, который приводит к задействованию контрольных точек, регулирует механизм восстановления ДНК, изменения клеточного цикла и апоптоз. Многочисленные биохимические структуры, а также процессы, обнаруживающие повреждения ДНК, представляют собой ATM и ATR, которые вызывают контрольные точки восстановления ДНК. [35]
Клеточный цикл — это последовательность действий, в ходе которой клеточные органеллы дублируются и впоследствии с точностью разделяются на дочерние клетки. Есть важные события, которые происходят во время клеточного цикла. Процессы, происходящие в клеточном цикле, включают развитие клеток, репликацию и сегрегацию хромосом. Контрольно-пропускные пункты клеточного цикла — это системы наблюдения, которые отслеживают целостность, точность и хронологию клеточного цикла. Каждая контрольная точка служит альтернативной конечной точкой клеточного цикла, в которой проверяются параметры ячейки, и только при достижении желаемых характеристик клеточный цикл продвигается через отдельные этапы. Цель клеточного цикла — точно скопировать ДНК каждого организма, а затем поровну разделить клетку и ее компоненты между двумя новыми клетками. У эукариот происходят четыре основные стадии. В G1 клетка обычно активна и продолжает быстро расти, тогда как в G2 рост клетки продолжается, пока молекулы белка не готовы к разделению. Сейчас не времена покоя; это когда клетки набирают массу, интегрируют рецепторы факторов роста, устанавливают реплицированный геном и готовятся к сегрегации хромосом. Репликация ДНК у эукариот ограничена отдельным синтезом, который также известен как S-фаза. Во время митоза, который также известен как М-фаза, происходит разделение хромосом. [36] ДНК, как и любая другая молекула, способна вступать в широкий спектр химических реакций. С другой стороны, модификации последовательности ДНК оказывают значительно большее влияние, чем модификации других клеточных компонентов, таких как РНК или белки, поскольку ДНК действует как постоянная копия клеточного генома. Когда ошибочные нуклеотиды включаются во время репликации ДНК, могут возникнуть мутации. Большинство повреждений ДНК устраняется путем удаления дефектных оснований и последующего повторного синтеза вырезанной области. С другой стороны, некоторые повреждения ДНК можно вылечить, обратив вспять повреждение, что может быть более эффективным методом борьбы с распространенными типами повреждений ДНК. Таким способом можно исправить только несколько форм повреждений ДНК, в том числе пиримидиновые димеры, вызванные ультрафиолетовым (УФ) светом, измененные в результате внедрения метильных или этильных групп в положение O6 пуринового кольца. [37]
мембраны митохондриальной Динамика
Митохондрии обычно называют «электростанциями» клеток из-за их способности эффективно производить АТФ, которая необходима для поддержания клеточного гомеостаза и метаболизма. Более того, исследователи получили лучшее представление о значении митохондрий в клеточной биологии благодаря открытию клеточных сигнальных путей с помощью митохондрий, которые являются решающими платформами для регуляции клеточных функций, таких как апоптоз. Его физиологическая адаптивность тесно связана с продолжающейся реконфигурацией клеточных митохондриальных каналов посредством ряда механизмов, известных как динамика митохондриальных мембран, которые включают слияние и фрагментацию (разделение) эндомембран, а также ультраструктурное ремоделирование мембраны. В результате динамика митохондрий регулирует и часто управляет не только метаболическими, но и сложными клеточными сигнальными процессами, такими как клеточные плюрипотентные стволовые клетки, пролиферация, созревание, старение и смертность. Взаимно, посттрансляционные изменения митохондриального аппарата и развитие сайтов трансмембранных контактов между митохондриями и другими структурами, которые потенциально могут связывать сигналы из разных путей, которые существенно влияют на динамику митохондриальных мембран. [36] Митохондрии окружены двумя мембранами: внутренней митохондриальной мембраной (IMM) и внешней митохондриальной мембраной (OMM), каждая из которых имеет особую функцию и структуру, что соответствует их двойной роли как клеточных электростанций и сигнальных органелл. Внутренняя митохондриальная мембрана делит просвет митохондрий на две части: внутреннюю пограничную мембрану, которая проходит параллельно ОММ, и кристы, которые представляют собой глубоко скрученные многоядерные впячивания, которые дают место для увеличения площади поверхности и вмещают митохондриальный дыхательный аппарат. Внешняя митохондриальная мембрана, напротив, мягкая и проницаемая. Таким образом, он действует как основа для клеточных сигнальных путей, которые собираются, расшифровываются и транспортируются в митохондрии. Кроме того, ОММ соединяется с другими клеточными органеллами, такими как эндоплазматический ретикулум (ЭР), лизосомы, эндосомы и плазматическая мембрана. Митохондрии играют широкий спектр ролей в клеточной биологии, что отражается в их морфологическом разнообразии. С самого начала изучения митохондрий было документально подтверждено, что митохондрии могут иметь множество форм, причем как их общая, так и ультраструктурная морфология сильно различается между клетками, в течение клеточного цикла и в ответ на метаболические или клеточные сигналы. . Митохондрии могут существовать как независимые органеллы или как часть более крупных систем; они также могут быть неравномерно распределены в цитозоле посредством регулируемого митохондриального транспорта и размещения для удовлетворения локализованных энергетических потребностей клетки. Митохондриальная динамика относится к адаптивному и изменчивому аспекту митохондрий, включая их форму и внутриклеточное распределение. [36]
Аутофагия [ править ]
Аутофагия — это механизм самодеградации, который регулирует источники энергии во время роста и реакции на пищевой стресс. Аутофагия также убирает за собой, очищая агрегированные белки, очищая поврежденные структуры, включая митохондрии и эндоплазматический ретикулум, и уничтожая внутриклеточные инфекции. Кроме того, аутофагия выполняет противовирусную и антибактериальную роль внутри клетки и участвует в начале характерных и адаптивных иммунных ответов на вирусное и бактериальное заражение. Некоторые вирусы содержат белки вирулентности, которые предотвращают аутофагию, тогда как другие используют элементы аутофагии для внутриклеточного развития или клеточного расщепления. [38] Макроаутофагия, микроаутофагия и шаперон-опосредованная аутофагия являются тремя основными типами аутофагии. Когда запускается макроаутофагия, мембрана исключения включает в себя часть цитоплазмы, образуя аутофагосому, характерную двухмембранную органеллу. Затем аутофагосома присоединяется к лизосоме, образуя аутолизосому, при этом лизосомальные ферменты разрушают компоненты. При микроаутофагии лизосома или вакуоль поглощают часть цитоплазмы, инвагинируя или выпячивая лизосомальную мембрану, закрывая цитозоль или органеллы. Обеспечение качества белка шаперон -опосредованной аутофагии (CMA) путем переваривания окисленных и измененных белков в стрессовых условиях и доставки аминокислот посредством денатурации белка. [39] Аутофагия — это основная внутренняя система деградации пептидов, жиров, углеводов и других клеточных структур. Как в физиологических, так и в стрессовых ситуациях такое клеточное развитие жизненно важно для поддержания правильного клеточного баланса. Нестабильность аутофагии приводит к различным симптомам заболеваний, включая воспаление, биохимические нарушения, старение и нейродегенерацию, из-за ее участия в контроле целостности клеток. Модификация аутофагически-лизосомальных сетей является типичным признаком многих неврологических и мышечных заболеваний. В результате аутофагия была определена как потенциальная стратегия профилактики и лечения различных заболеваний. Многие из этих расстройств можно предотвратить или улучшить, употребляя полифенолы с едой. В результате природные соединения, способные модифицировать механизм аутофагии, рассматриваются как потенциальный терапевтический вариант. [40] Создание двойной мембраны (фагофора), которое будет известно как нуклеация, является первым шагом в макроаутофагии. Фагофорный подход указывает на дисрегуляцию полипептидов или дефектных органелл, которые происходят из клеточной мембраны, аппарата Гольджи, эндоплазматического ретикулума и митохондрий. С заключением аутофагоцита расширение фагофора заканчивается. Аутофагосома соединяется с лизосомальными везикулами, образуя аутолизосому, которая расщепляет инкапсулированные вещества, что называется фагоцитозом. [41]
клеточные Известные биологи
- Жан Батист Карнуа
- Питер Агре
- Гюнтер Блобель
- Роберт Браун
- Джеффри М. Купер
- Кристиан де Дюв
- Анри Дютроше
- Роберт Гук
- Х. Роберт Хорвиц
- Марк Киршнер
- Антон ван Левенгук
- Ира Меллман
- Марта Минчиньска [42]
- Питер Д. Митчелл
- Рудольф Вирхов
- Пол Нерс
- Джордж Эмиль Паладе
- Кейт Р. Портер
- Рэй Раппапорт
- Майкл Суонн
- Роджер Тен
- Эдмунд Бичер Уилсон
- Кеннет Р. Миллер
- Маттиас Якоб Шлейден
- Теодор Шванн
- Ёсинори Осуми
- Ян Евангелиста Пуркине
 Чешский анатом Ян Евангелиста Пуркине наиболее известен своим открытием в 1837 году клеток Пуркинье .
Чешский анатом Ян Евангелиста Пуркине наиболее известен своим открытием в 1837 году клеток Пуркинье . Теодор Шванн , первооткрыватель шванновской клетки
Теодор Шванн , первооткрыватель шванновской клетки Ёсинори Осуми , лауреат Нобелевской премии за работы по аутофагии
Ёсинори Осуми , лауреат Нобелевской премии за работы по аутофагии



См. также [ править ]
- Американское общество клеточной биологии
- Клеточная биофизика
- Разрушение клеток
- Клеточная физиология
- Клеточная адаптация
- Клеточная микробиология
- Институт молекулярной и клеточной биологии (значения)
- Мейомитоз
- Органоид
- Очерк клеточной биологии
Примечания [ править ]
- ^ Альбертс, Брюс; Джонсон, Александр Д.; Морган, Дэвид; Рафф, Мартин; Робертс, Кейт; Уолтер, Питер (2015). «Клетки и геномы». Молекулярная биология клетки (6-е изд.). Нью-Йорк, штат Нью-Йорк: Garland Science. стр. 1–42. ISBN 978-0815344322 .
- ^ Бишелья, Ник. «Клеточная биология» . Возбудимый . www.nature.com.
- ^ Jump up to: Перейти обратно: а б с Гупта, П. (1 декабря 2005 г.). Клеточная и молекулярная биология . Публикации Растоги. п. 11. ISBN 978-8171338177 .
- ^ Гук, Роберт (сентябрь 1665 г.). Микрография .
- ^ Jump up to: Перейти обратно: а б Чабб, Гилберт Чарльз (1911). . В Чисхолме, Хью (ред.). Британская энциклопедия . Том. 7 (11-е изд.). Издательство Кембриджского университета. п. 710.
- ^ Паес-Эспино Д., Элое-Фадрош Е.А., Павлопулос Г.А., Томас А.Д., Хантеманн М., Михайлова Н., Рубин Е., Иванова Н.Н., Кирпидес Н.К. (август 2016 г.). «Открытие земного вирома » Природа 536 (7617): 425–30. Бибкод : 2016Nature.536..425P . дои : 10.1038/nature19094 . ПМИД 27533034 . S2CID 4466854 .
- ^ Лаванья, П. (1 декабря 2005 г.). Клеточная и молекулярная биология . Публикации Растоги. п. 11. ISBN 978-8171338177 .
- ^ Jump up to: Перейти обратно: а б с д и ж Купер, Джеффри М. (2000). «Инструменты клеточной биологии» . Клетка: молекулярный подход. 2-е издание .
- ^ Маккиннон, Кэтрин М. (21 февраля 2018 г.). «Проточная цитометрия: обзор» . Современные протоколы в иммунологии . 120 (1): 5.1.1–5.1.11. дои : 10.1002/cpim.40 . ISSN 1934-3671 . ПМЦ 5939936 . ПМИД 29512141 .
- ^ Добль, Мукеш; Гуммади, Сатьянараяна Н. (5 августа 2010 г.). Биохимическая инженерия . Нью-Дели: Prentice-Hall of India Pvt Ltd. ISBN 978-8120330528 .
- ^ Канеширо, Эдна (2 мая 2001 г.). Справочник по клеточной физиологии: молекулярный подход (3-е изд.). Академическая пресса. ISBN 978-0123877383 .
- ^ Леветин, Эстель; МакМахон, Карен (16 октября 2014 г.). Электронная книга: Растения и общество . МакГроу Хилл. п. 135. ИСБН 978-0-07-717206-0 .
- ^ Jump up to: Перейти обратно: а б с д Нельсон, Дэниел (22 июня 2018 г.). «Разница между эукариотическими и прокариотическими клетками». Тенденции науки . doi : 10.31988/scitrends.20655 . S2CID 91382191 .
- ^ Гриффитс, Энтони Дж. Ф.; Миллер, Джеффри Х.; Сузуки, Дэвид Т.; Левонтин, Ричард С.; Гелбарт, Уильям М. (2000). «Бактериальная конъюгация» . Введение в генетический анализ. 7-е издание .
- ^ Элосеги-Артола А., Андреу И., Бидл А.Е., Лезамиз А., Уроз М., Космальска А.Дж. и др. (ноябрь 2017 г.). «Сила вызывает проникновение YAP в ядерную зону, регулируя транспорт через ядерные поры» . Клетка . 171 (6): 1397–1410.e14. дои : 10.1016/j.cell.2017.10.008 . ПМИД 29107331 .
- ^ «Ядро» . Genome.gov . Проверено 27 сентября 2021 г.
- ^ «Эндоплазматический ретикулум (шероховатый и гладкий) | Британское общество клеточной биологии» . Проверено 6 октября 2019 г.
- ^ Studios, Эндрю Рейдер. «Biology4Kids.com: Структура клетки: эндоплазматическая сеть» . www.biology4kids.com . Проверено 27 сентября 2021 г.
- ^ «Энергостанция клетки имеет механизм самосохранения» . ЭврекАлерт! . Проверено 27 сентября 2021 г.
- ^ Пелли, Джон В. (2007), «Цикл лимонной кислоты, цепь переноса электронов и окислительное фосфорилирование», Интегрированная биохимия Elsevier , Elsevier, стр. 55–63, doi : 10.1016/b978-0-323-03410-4.50013-4 , ISBN 9780323034104
- ^ Купер, Джеффри М. (2000). «Аппарат Гольджи» . Клетка: молекулярный подход. 2-е издание .
- ^ Верити, Массачусетс. Лизосомы: некоторые патологические последствия . OCLC 679070471 .
- ^ «Рибосома | цитология» . Британская энциклопедия . Проверено 27 сентября 2021 г.
- ^ Купер, Джеффри М. (2000). Клетка: молекулярный подход . АСМ Пресс. ISBN 9780878931064 .
- ^ Купер, Джеффри М. (2000). «Транспорт малых молекул» . Клетка: молекулярный подход. 2-е издание .
- ^ «Каковы основные функции ресничек и жгутиков?» . Наука . Проверено 23 ноября 2020 г.
- ^ Jump up to: Перейти обратно: а б Ахмад, Мария; Кахваджи, Чади И. (2019), «Биохимия, цепь переноса электронов» , StatPearls , StatPearls Publishing, PMID 30252361 , получено 20 октября 2019 г.
- ^ Шлезингер, Джозеф (октябрь 2000 г.). «Передача сигналов клетками рецепторными тирозинкиназами» . Клетка . 103 (2): 211–225. дои : 10.1016/s0092-8674(00)00114-8 . ISSN 0092-8674 . ПМИД 11057895 . S2CID 11465988 .
- ^ Шекелфорд, RE; Кауфманн, ВК; Паулес, Р.С. (февраль 1999 г.). «Контроль клеточного цикла, механизмы контрольных точек и генотоксический стресс» . Перспективы гигиены окружающей среды . 107 (приложение 1): 5–24. дои : 10.1289/ehp.99107s15 . ISSN 0091-6765 . ПМК 1566366 . ПМИД 10229703 .
- ^ Jump up to: Перейти обратно: а б Бернштейн С., Бернштейн Х., Пейн С. Бессмертие клеток: поддержание потенциала клеточного деления. Прог Мол Субклеточная Биол. 2000;24:23-50. дои : 10.1007/978-3-662-06227-2_2 . ПМИД 10547857.
- ^ Авис JC. Перспектива: Эволюционная биология старения, полового размножения и восстановления ДНК. Эволюция. Октябрь 1993 г.; 47 (5): 1293–1301. дои : 10.1111/j.1558-5646.1993.tb02155.x . ПМИД 28564887.
- ^ «Клеточный цикл – Фазы – Митоз – Регуляция» . Научите меня физиологии . Проверено 7 октября 2021 г.
- ^ «Что такое патология?» . Новости-Medical.net . 13 мая 2010 года . Проверено 21 сентября 2021 г.
- ^ Медсестра Пол (7 января 2000 г.). «Долгий двадцатый век клеточного цикла и далее» . Клетка . 100 (1): 71–78. дои : 10.1016/S0092-8674(00)81684-0 . ISSN 0092-8674 . ПМИД 10647932 . S2CID 16366539 .
- ^ Цимприч, Карлин А.; Кортес, Дэвид (август 2008 г.). «ATR: важный регулятор целостности генома» . Nature Reviews Молекулярно-клеточная биология . 9 (8): 616–627. дои : 10.1038/nrm2450 . ISSN 1471-0080 . ПМЦ 2663384 . ПМИД 18594563 .
- ^ Jump up to: Перейти обратно: а б с Джакомелло, Марта; Пьякурел, Асвин; Глицова, Кристина; Скоррано, Лука (18 февраля 2020 г.). «Клеточная биология динамики митохондриальных мембран» . Nature Reviews Молекулярно-клеточная биология . 21 (4): 204–224. дои : 10.1038/s41580-020-0210-7 . ISSN 1471-0072 . ПМИД 32071438 . S2CID 211170966 .
- ^ Ты, Чжуншэн; Бейлис, Джули М. (июль 2010 г.). «Повреждение ДНК и решения: CtIP координирует восстановление ДНК и контрольные точки клеточного цикла» . Тенденции в клеточной биологии . 20 (7): 402–409. дои : 10.1016/j.tcb.2010.04.002 . ISSN 0962-8924 . ПМК 5640159 . ПМИД 20444606 .
- ^ Глик, Даниэль; Барт, Сандра; Маклауд, Кей Ф. (3 февраля 2010 г.). «Аутофагия: клеточные и молекулярные механизмы» . Журнал патологии . 221 (1): 3–12. дои : 10.1002/путь.2697 . ISSN 0022-3417 . ПМК 2990190 . ПМИД 20225336 .
- ^ Ёши, Саори Р.; Мидзусима, Нобору (28 августа 2017 г.). «Мониторинг и измерение аутофагии» . Международный журнал молекулярных наук . 18 (9): 1865. doi : 10.3390/ijms18091865 . ISSN 1422-0067 . ПМК 5618514 . ПМИД 28846632 .
- ^ Перроне, Лорена; Скилларо, Тициана; Наполитано, Филомена; Терраччано, Кьяра; Сампаоло, Симона; Мелоне, Мариароса Анна Беатрис (13 августа 2019 г.). «Сигнальный путь аутофагии: потенциальная многофункциональная терапевтическая мишень куркумина при неврологических и нервно-мышечных заболеваниях» . Питательные вещества . 11 (8): 1881. doi : 10.3390/nu11081881 . ISSN 2072-6643 . ПМК 6723827 . ПМИД 31412596 .
- ^ Левин, Бет; Кремер, Гвидо (11 января 2008 г.). «Аутофагия в патогенезе заболеваний» . Клетка . 132 (1): 27–42. дои : 10.1016/j.cell.2007.12.018 . ISSN 0092-8674 . ПМК 2696814 . ПМИД 18191218 .
- ^ «Исследования – ИИМКБ» . www.iimcb.gov.pl .
Ссылки [ править ]
- Пеннер-Хан, Джеймс Э. (2013). «Глава 2. Технологии обнаружения металлов в одиночных клетках. Раздел 4. Собственная рентгеновская флуоресценция». В Бани, Люсия (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. стр. 15–40. дои : 10.1007/978-94-007-5561-1_2 . ISBN 978-94-007-5560-4 . ПМИД 23595669 . электронная книга ISBN 978-94-007-5561-1 ISSN 1559-0836 электронный- ISSN 1868-0402
- Клеточная и молекулярная биология Карпа, 5-е изд., ISBN 0-471-46580-1
 В этой статье использованы общедоступные материалы из Научный букварь . НКБИ . Архивировано из оригинала 8 декабря 2009 года.
В этой статье использованы общедоступные материалы из Научный букварь . НКБИ . Архивировано из оригинала 8 декабря 2009 года.
Внешние ссылки [ править ]
 СМИ, связанные с клеточной биологией, на Викискладе?
СМИ, связанные с клеточной биологией, на Викискладе? - Клеточная биология в Керли
- Стареющая клетка
- «Фрэнсис Гарри Комптон Крик (1916–2004)» А. Андрея в Энциклопедии Embryo Project.
- «Биологический ресурс профессора Линя».

Отрасли химии |
|---|
| Базы данных органов управления : Национальные |
|---|