Митохондриальная Ева
| Гаплогруппа L | |
|---|---|
 | |
| Возможное время происхождения | в. 100–230 тысяч лет назад [ примечание 1 ] [ примечание 2 ] |
| Возможное место происхождения | Восточная Африка |
| Предок | н/д |
| Потомки | Митохондриальные макрогаплогруппы L0 , L1 и L5 |
| Определение мутаций | Никто |
В генетике человека Митохондриальная Ева (более известная как Митохондриальный Самый Недавний Общий Предок , сокращенно до mt-Eve или mt-MRCA ) является по материнской линии самым недавним общим предком (MRCA) всех ныне живущих людей. Другими словами, она определяется как самая последняя женщина, от которой все живые люди происходят по непрерывной линии исключительно через своих матерей и через матерей этих матерей, до тех пор, пока все линии не сойдутся в одной женщине.
С точки зрения митохондриальных гаплогрупп , mt-MRCA расположен в месте расхождения макрогаплогруппы L на L0 и L1–6 . По состоянию на 2013 год оценки возраста этого раскола колебались примерно в 155 000 лет назад. [ примечание 3 ] соответствует дате, более поздней, чем видообразование но Homo sapiens, более ранней, чем недавнее расселение за пределы Африки . [ 4 ] [ 1 ] [ 5 ]
Мужским аналогом «Митохондриальной Евы» является « Y-хромосомный Адам » (или Y-MRCA), индивидуум, от которого по отцовской линии произошли все ныне живущие люди. Поскольку идентичность MRCA как по материнской, так и по отцовской линии зависит от генеалогической истории ( коллапс родословной ), они не обязательно жили в одно и то же время. По состоянию на 2015 год оценки возраста Y-MRCA варьируются от 200 000 до 300 000 лет назад, что примерно соответствует появлению анатомически современного человека. [ 6 ]
Название «Митохондриальная Ева» отсылает к библейской Еве , что привело к неоднократным искажениям или заблуждениям в журналистских сообщениях по этой теме. Научно-популярные презентации этой темы обычно указывают на такие возможные заблуждения, подчеркивая тот факт, что положение mt-MRCA не является фиксированным во времени (поскольку положение mt-MRCA движется вперед во времени по мере митохондриальной ДНК вымирания линий (мтДНК)). и это не относится ни к «первой женщине», ни к единственной живой женщине своего времени, ни к первому представителю «нового вида». [ примечание 4 ]
История
[ редактировать ]−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( О. praegens ) ( О. тугененсис ) ( Ар. кадабба ) ( Ар. ramidus ) |
| ||||||||||||||||||||||||||||
Ранние исследования
[ редактировать ]Первые исследования с использованием методов молекулярных часов проводились в конце 1970-х — начале 1980-х годов. Аллан Уилсон , Марк Стоукинг , Ребекка Л. Канн и Уэсли Браун обнаружили, что мутация в мтДНК человека происходит неожиданно быстро, со скоростью 0,02 замены на основание (1%) за миллион лет, что в 5–10 раз быстрее, чем в ядерной ДНК . [ 8 ] Связанная с этим работа позволила проанализировать эволюционные взаимоотношения между гориллами, шимпанзе ( обыкновенный шимпанзе и бонобо ) и людьми. [ 9 ] Опираясь на данные 21 человека, Браун опубликовал первую оценку возраста mt-MRCA — 180 000 лет назад, в 1980 году. [ 10 ] Статистический анализ, опубликованный в 1982 году, был принят в качестве доказательства недавнего африканского происхождения (гипотеза, которая в то время конкурировала с азиатским происхождением H. sapiens ). [ 11 ] [ 12 ] [ 13 ]
публикация 1987 г.
[ редактировать ]К 1985 году были доступны данные мтДНК 145 женщин из разных популяций и двух клеточных линий , HeLa и GM 3043, полученных от афроамериканца и ƃKung соответственно. После более чем 40 изменений черновика рукопись была отправлена в журнал Nature в конце 1985 или начале 1986 года. [ 13 ] и опубликовано 1 января 1987 года. Опубликованный вывод заключался в том, что вся нынешняя человеческая мтДНК произошла от одной популяции из Африки, датированной периодом от 140 000 до 200 000 лет назад. [ 14 ]
Датировка «Евы» стала ударом по мультирегиональной гипотезе , которая обсуждалась в то время, и послужила толчком для теории недавней модели происхождения . [ 15 ]
Канн, Стоункинг и Уилсон не использовали термин «Митохондриальная Ева» или даже имя «Ева» в своей оригинальной статье. Однако Канн использовал его в статье под названием «В поисках Евы» в сентябрьском – октябре 1987 года номере журнала The Sciences . [ 16 ] Оно также появляется в статье журнале Science в Роджера Левина за октябрь 1987 года , озаглавленной «Разоблачение митохондриальной Евы». [ 17 ] Библейский подтекст был очень ясен с самого начала. Сопутствующая исследовательская новость в журнале Nature называлась «Из Эдемского сада». [ 18 ]
Сам Уилсон предпочитал термин «Счастливая Мать». [ 19 ] и считал использование имени Ева «достойным сожаления». [ 17 ] [ 20 ] Но концепция Евы завоевала популярность среди публики и была повторена в статье на обложке Newsweek (в номере от 11 января 1988 года на обложке было изображение Адама и Евы под названием «В поисках Адама и Евы»), [ 21 ] и статья на обложке журнала Time от 26 января 1987 года. [ 22 ]
Критика и более поздние исследования
[ редактировать ]Вскоре после публикации 1987 года была опубликована критика его методологии и вторичных выводов. [ 23 ] И датировка Евы, и значимость возраста чисто матрилинейного происхождения для замещения населения были предметом споров в 1990-е годы; [ 24 ] [ 25 ] [ 26 ] [ 27 ] Алан Темплтон (1997) утверждал, что исследование «не подтвердило гипотезу недавнего африканского происхождения всего человечества после раскола между африканцами и неафриканцами, произошедшего 100 000 лет назад», а также «не подтвердило гипотезу недавнего глобального замещения людей, вышедших из Африки». [ 28 ]
Размещение Канном, Стоункингом и Уилсоном (1987) относительно небольшой популяции людей в Африке к югу от Сахары соответствовало гипотезе Канна (1982) и обеспечило значительную поддержку сценарию «недавнего выхода из Африки».
В 1999 году Крингс и др. устранил проблемы молекулярной синхронизации, постулированные Неем (1992). [ 29 ] когда было обнаружено, что последовательность мтДНК для того же региона существенно отличалась от MRCA по сравнению с любой последовательностью человека. [ 30 ] [ 31 ]
В 1997 году Парсонс и др. (1997) опубликовали исследование частоты мутаций мтДНК в одной хорошо задокументированной семье ( семья Романовых , принадлежащая к российской королевской семье). В этом исследовании они подсчитали, что частота мутаций в двадцать раз превышает предыдущие результаты. [ 32 ]
Хотя первоначальное исследование имело аналитические ограничения, оценка возраста mt-MRCA оказалась надежной. [ 33 ] [ 34 ] Более поздние оценки возраста остались в соответствии с оценкой в 140–200 тысяч лет назад , опубликованной в 1987 году: по оценке 2013 года, Митохондриальная Ева датируется примерно 160 тысячами лет назад (в пределах сдержанной оценки оригинального исследования), а Out of Africa II - примерно 95 тысяч лет назад. [ 3 ] Другое исследование 2013 года (основанное на секвенировании генома 69 человек из 9 различных популяций) показало, что возраст Митохондриальной Евы составляет от 99 до 148 тысяч лет назад, а возраст Y-MRCA - от 120 до 156 тысяч лет назад. [ 2 ]
Женское и митохондриальное происхождение
[ редактировать ]
Без образца ДНК невозможно восстановить полный генетический состав ( геном ) любого человека, который умер очень давно. Однако, анализируя ДНК потомков, ученые оценивают части геномов предков. Митохондриальная ДНК (мтДНК, ДНК, расположенная в митохондриях и отличающаяся от ДНК в ядре клетки) и ДНК Y-хромосомы обычно используются для отслеживания происхождения таким способом. мтДНК обычно передается от матери к детям обоих полов в несмешанном виде по материнской линии или по матрилинейной линии . [ 35 ] [ 36 ] Матрилинейное происхождение происходит от матерей к их матерям, пока все женские линии не сойдутся.
Ветви идентифицируются одним или несколькими уникальными маркерами, которые дают митохондриальную «подпись ДНК» или « гаплотип » (например, CRS является гаплотипом). Каждый маркер представляет собой пару оснований ДНК, возникшую в результате SNP мутации . Ученые сортируют результаты митохондриальной ДНК на более или менее родственные группы с более или менее недавними общими предками. Это приводит к построению генеалогического древа ДНК , где ветви в биологическом смысле представляют собой клады , а общие предки, такие как Митохондриальная Ева, находятся в точках ветвления этого дерева. Говорят, что основные ветви определяют гаплогруппу (например, CRS принадлежит гаплогруппе H ), а большие ветви, содержащие несколько гаплогрупп, называются «макрогаплогруппами».

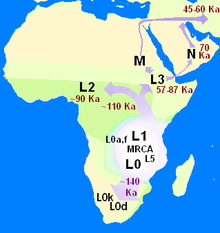
Митохондриальная клада, которую определяет Митохондриальная Ева, — это сам вид Homo sapiens sapiens или, по крайней мере, нынешняя популяция или « хроновид », существующий сегодня. В принципе, более ранние Евы также могут быть определены, выходящие за рамки вида, например, те, кто являются предками как современного человечества, так и неандертальцев , или, еще раньше, «Евы», предков всех представителей рода Homo и шимпанзе рода Pan . Согласно текущей номенклатуре, митохондриальная гаплогруппа Евы находилась в митохондриальной гаплогруппе L, поскольку эта макрогаплогруппа содержит все сохранившиеся сегодня человеческие митохондриальные линии, и она должна была предшествовать появлению L0 .
Вариации митохондриальной ДНК у разных людей можно использовать для оценки времени, прошедшего до общего предка, такого как Митохондриальная Ева. Это работает, потому что по любой конкретной линии происхождения митохондриальная ДНК накапливает мутации со скоростью примерно одна мутация каждые 3500 лет на нуклеотид. [ 1 ] [ 37 ] [ примечание 5 ] Определенное количество этих новых вариантов доживет до наших дней и будет идентифицировано как отдельные линии. В то же время некоторые ветви, даже очень старые, прекращают свое существование, когда в последней семье отдельной ветви не остается дочерей.
Митохондриальная Ева — самый недавний общий предок всех современных людей по материнской линии. Всякий раз, когда одна из двух самых древних ветвей вымирает (производя в это время только нематрилинейных потомков), MRCA переходит к более позднему предку женского пола, всегда самой последней матери, у которой было более одной дочери с живой материнской линией. потомки живы и сегодня. Число мутаций, которые можно обнаружить, отличающие современных людей, определяется двумя критериями: во-первых, и это наиболее очевидно, временем, обратным к ней, а во-вторых, менее очевидно, различной скоростью, с которой возникают новые ветви и становятся старыми. вымерший. Глядя на количество мутаций, накопленных в различных ветвях этого генеалогического древа, и глядя на то, в каких географических регионах имеется самый широкий спектр наименее связанных ветвей, можно предположить регион, в котором жила Ева.
Популярный прием и заблуждения
[ редактировать ]Newsweek сообщил о Митохондриальной Еве на основе данных Cann et al. исследование в январе 1988 года под заголовком «Ученые исследуют противоречивую теорию происхождения человека». Издание было продано рекордным тиражом. [ 38 ]
Популярное название «Митохондриальная Ева», отчеканенное в 1980-х годах. [ 17 ] способствовал появлению ряда популярных заблуждений. Поначалу заявление о «митохондриальной Еве» было даже встречено с одобрением со стороны молодых земных креационистов , которые рассматривали эту теорию как подтверждение библейской истории творения . [ 39 ] [ 40 ] [ 41 ] [ нужен неосновной источник ]
Из-за таких недопониманий авторы научно-популярных публикаций с 1990-х годов настойчиво подчеркивали, что это имя - всего лишь популярное условное обозначение и что mt-MRCA ни в коем случае не была «первой женщиной». [ 42 ] Ее позиция является чисто результатом генеалогической истории человеческих популяций позже, и по мере того, как матрилинейные линии вымирают, позиция mt-MRCA со временем продолжает смещаться к более молодым людям.
В книге «Река из рая» (1995) Ричард Докинз обсуждал человеческое происхождение в контексте «реки генов», включая объяснение концепции митохондриальной Евы. [ 43 ] «Семь дочерей Евы» (2002) представили широкой аудитории тему митохондриальной генетики человека. [ 44 ] Настоящая канун: путешествие современного человека из Африки Стивена Оппенгеймера (2003) [ 38 ] был адаптирован в документальный фильм Discovery Channel 2002 года . [ 45 ]
Не единственная женщина
[ редактировать ]Одно распространенное заблуждение относительно Митохондриальной Евы заключается в том, что, поскольку все живущие сегодня женщины произошли от нее по прямой непрерывной женской линии , она, должно быть, была единственной женщиной, жившей в то время. [ 42 ] Однако исследования ядерной ДНК показывают, что эффективная численность населения древних людей никогда не опускалась ниже десятков тысяч. [ 46 ] У других женщин, живших во времена Евы, могут быть потомки, живущие сегодня, но не по прямой женской линии. [ 47 ]
Не фиксированная личность с течением времени
[ редактировать ]Определение Митохондриальной Евы фиксировано, но женщина в доисторические времена, подходящая под это определение, может измениться. То есть не только наши знания о том, когда и где жила Митохондриальная Ева, могут измениться благодаря новым открытиям, но и сама Митохондриальная Ева может измениться. Митохондриальная Ева может измениться, когда линия матери-дочери подходит к концу. Из определения Митохондриальной Евы следует, что у нее было как минимум две дочери, обе из которых имеют непрерывную женскую линию, сохранившуюся до наших дней. В каждом поколении митохондриальные линии заканчиваются – когда женщина с уникальной мтДНК умирает, не оставив дочерей. Когда митохондриальные линии дочерей Митохондриальной Евы вымирают, титул «Митохондриальная Ева» переходит вперед от оставшейся дочери через ее потомков по материнской линии, пока не будет достигнут первый потомок, у которого было две или более дочерей, которые вместе имеют всех живых людей как их потомки по материнской линии. Как только линия вымирает, она безвозвратно утрачивается, и этот механизм может, таким образом, лишь сдвинуть название «Митохондриальная Ева» вперед во времени. [ 48 ]
Поскольку картирование мтДНК человека очень неполное, открытие живых линий мтДНК, которые предшествуют нашей нынешней концепции «Митохондриальной Евы», может привести к тому, что титул перейдет к более ранней женщине. Это случилось с ее коллегой-мужчиной, «Y-хромосомным Адамом», когда была обнаружена более старая линия Y, гаплогруппа A-00 . [ 49 ]
Не обязательно современник «Y-хромосомного Адама».
[ редактировать ]Иногда предполагается, что Митохондриальная Ева жила в то же время, что и Адам с Y-хромосомой (от которого все ныне живущие мужчины произошли по отцовской линии), и, возможно, даже встречалась с ним и спаривалась с ним. Даже если бы это было правдой, что в настоящее время считается маловероятным, это было бы всего лишь совпадением. Подобно митохондриальной «Еве», Y-хромосомный «Адам», вероятно, жил в Африке. Однако недавнее исследование (март 2013 г.) пришло к выводу, что «Ева» жила намного позже «Адама» — примерно на 140 000 лет позже. [ 50 ] (Более ранние исследования, наоборот, считали, что «Ева» жила раньше «Адама».) [ 51 ] Более поздние исследования показывают, что митохондриальная Ева и Адам с Y-хромосомой действительно могли жить примерно в одно и то же время. [ 52 ]
Не самый недавний предок, общий для всех людей.
[ редактировать ]Митохондриальная Ева — самый поздний общий предок по материнской линии , а не самый поздний общий предок . Поскольку мтДНК наследуется по материнской линии, а рекомбинация либо редка, либо отсутствует, относительно легко проследить происхождение линий до MRCA; однако этот MRCA действителен только при обсуждении митохондриальной ДНК. Приблизительная последовательность от самых новых к самым старым может перечислить различные важные моменты в происхождении современных человеческих популяций:
- Человеческий MRCA . Период времени, в течение которого жил человек MRCA, неизвестен. Роде и др. выдвинули «грубое предположение», что MRCA мог существовать 5000 лет назад; однако авторы заявляют, что эта оценка является «крайне предварительной, и модель содержит несколько очевидных источников ошибок, поскольку она была мотивирована больше соображениями теоретической проницательности и управляемости, чем реализмом». [ 53 ] Всего за несколько тысяч лет до того, как самый последний единый предок, общий для всех ныне живущих людей, был временем, когда все люди, которые тогда были живы, либо не оставили потомков в живых сегодня, либо были общими предками всех людей, живущих сегодня. Однако столь позднюю дату трудно совместить с географическим распространением нашего вида и последующей изоляцией разных групп друг от друга. Например, общепринято, что коренное население Тасмании было изолировано от всех остальных людей в период между подъемом уровня моря после последнего ледникового периода около 8000 лет назад и приходом европейцев. Оценки MRCA даже близкородственных человеческих популяций были сделаны гораздо больше, чем 5000 лет назад. [ 54 ]
- Тождественные предки указывают . Другими словами, «каждый современный человек имеет точно такой же набор генеалогических предков», живущих в «точку идентичных предков» во времени. Это произошло гораздо позже, чем тогда, когда предполагалось, что Митохондриальная Ева жила. [ 53 ]
- Митохондриальная Ева, последний общий предок всех ныне живущих людей по женской линии.
- «Y-хромосомный Адам», последний общий предок всех ныне живущих людей по мужской линии.
См. также
[ редактировать ]- Археогенетика
- Коалесцентная теория
- Евразийский Адам
- Генеалогический ДНК-тест
- Генетическая генеалогия
- Гаплогруппа L0
- Эволюция человека
- Гаплогруппы митохондриальной ДНК человека
- Последний вселенский предок
- Макрогаплогруппа L (мтДНК)
- Митохондриальный геном
- Моногенизм
- Нейтральная теория молекулярной эволюции
- Доисторическая демография
- Гипотеза единого происхождения
- Хронология эволюции
- Хронология эволюции человека
- Y-хромосома Аарон
- Y-хромосомный Адам
- Женщины в предыстории
|
Филогенетическое древо гаплогрупп митохондриальной ДНК (мтДНК) человека | |||||||||||||||||||||||||||||||||||||||
| Митохондриальная Ева ( L ) | |||||||||||||||||||||||||||||||||||||||
| Л0 | Л1–6 | ||||||||||||||||||||||||||||||||||||||
| Л1 | Л2 | Л3 | Л4 | Л5 | Л6 | ||||||||||||||||||||||||||||||||||
| М | Н | ||||||||||||||||||||||||||||||||||||||
| Чехия | Д | И | Г | вопрос | ТО | А | С | Р | я | В | Х | И | |||||||||||||||||||||||||||
| С | С | Б | Ф | Р0 | до JT | П | В | ||||||||||||||||||||||||||||||||
| Х.В. | Джей Ти | К | |||||||||||||||||||||||||||||||||||||
| ЧАС | V | Дж | Т | ||||||||||||||||||||||||||||||||||||
Примечания
[ редактировать ]- ^ «Скорость синонимических мутаций Кивисилда и др. [...] оценивает время слияния дерева мтДНК в целом в ~ 160 000 тысяч лет назад [...] Мы представляем пересмотренную хронологию с использованием полной скорости генома мтДНК и подхода ML для дерево мтДНК на рисунке 6, с полной информацией об оценках возраста и связанных с ними 95% доверительных областях в таблице S5». [ 1 ] См.: Дополнительные данные.
- ^ «Мы оцениваем время до появления самого недавнего общего предка (TMRCA) Y-хромосомы от 120 до 156 тысяч лет, а митохондриального генома TMRCA от 99 до 148 тысяч лет. Наши результаты показывают, что, вопреки предыдущим утверждениям, мужчины линии не сливаются значительно позже, чем женские линии». [ 2 ]
- ^ Два исследования, опубликованные в 2013 году, имели 95% доверительные интервалы, едва перекрывающиеся в районе 15 тыс. лет назад, в третьем исследовании 95% доверительный интервал был промежуточным между двумя другими: «от 99 до 148 тыс. лет назад», согласно Познику, 2013. [ 2 ] (оценка возраста цельной мтДНК ML: 178,8 [155,6; 202,2], оценка возраста цельной мтДНК ρ: 185,2 [153,8; 216,9], оценка синонимического возраста ρ: 174,8 [153,8; 216,9]), «от 134 до 188 тыс. лет назад», согласно Фу, 2013, [ 3 ] и от 150 до 234 тыс. лет назад (95% ДИ) из Соареса, 2009. [ 1 ]
- ^ «Внимание: это не делает Митохондриальную Еву первой женщиной, первым человеком или первым представителем нового вида. Дальнейшее предупреждение: это не означает, что другие женщины, жившие во времена Евы, не имеют потомков сегодня; они просто не имеют живых потомков, произошедших только по женским связям. Еще одно предостережение: если будет обнаружен человек, мтДНК которого покажет образец мутаций большей временной глубины, то это будет статус Митохондриальной Евы. будет переназначен самому последнему предку женского пола, общему как для этого человека, так и для человека, которого мы теперь называем Митохондриальной Евой». [ 7 ]
- ^ В мтДНК есть участки (такие как: 16129, 16223, 16311, 16362), которые развиваются быстрее и, как было отмечено, изменяются в пределах временных рамок внутри поколения - Excoffier & Yang (1999) .
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Соарес П., Эрмини Л., Томсон Н., Мормина М., Рито Т., Рёль А. и др. (июнь 2009 г.). «Поправка на очищающий отбор: улучшенные митохондриальные молекулярные часы человека» . Американский журнал генетики человека . 84 (6): 740–759. дои : 10.1016/j.ajhg.2009.05.001 . ПМЦ 2694979 . ПМИД 19500773 .
- ^ Jump up to: а б с Позник Г.Д., Хенн Б.М., Йи М.К., Сливерска Э., Ойскирхен Г.М., Лин А.А. и др. (август 2013 г.). «Секвенирование Y-хромосомы устраняет несоответствие во времени общему предку мужчин и женщин» . Наука . 341 (6145): 562–565. Бибкод : 2013Sci...341..562P . дои : 10.1126/science.1237619 . ПМК 4032117 . ПМИД 23908239 .
- ^ Jump up to: а б Фу К., Миттник А., Джонсон П.Л., Бос К., Лари М., Боллонгино Р. и др. (апрель 2013 г.). «Пересмотренная временная шкала эволюции человека, основанная на древних митохондриальных геномах» . Современная биология . 23 (7): 553–559. дои : 10.1016/j.cub.2013.02.044 . ПМК 5036973 . ПМИД 23523248 .
- ^ Эндикотт П., Хо С.Ю., Метспалу М., Стрингер С. (сентябрь 2009 г.). «Оценка митохондриальной шкалы эволюции человека». Тенденции в экологии и эволюции . 24 (9): 515–521. дои : 10.1016/j.tree.2009.04.006 . ПМИД 19682765 .
- ^ «Новые «молекулярные часы» помогают датировать историю миграции человечества» . Университет Лидса. 3 июня 2009 года . Проверено 23 декабря 2019 г.
- ^ Кармин; и др. (2015). «Недавнее ограничение разнообразия Y-хромосом совпало с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–66. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 . «Мы датируем самого последнего общего предка Y-хромосомы (MRCA) в Африке 254 (95% ДИ 192–307) тысячелетий назад и обнаруживаем кластер основных неафриканских гаплогрупп-основателей в узком временном интервале в 47–52 тысячелетий, что соответствует с моделью быстрой первоначальной колонизации Евразии и Океании после «узкого места» за пределами Африки. В отличие от демографических реконструкций, основанных на мтДНК, мы. Мы предполагаем, что это узкое место вызвано культурными изменениями, влияющими на различия в репродуктивном успехе среди самцов».
- ^ «Джордан: «Митохондриальная Ева» » . weber.ucsd.edu . 2011 . Проверено 7 января 2012 г.
- ^ Браун В.М., Джордж М., Уилсон AC (апрель 1979 г.). «Быстрая эволюция митохондриальной ДНК животных» . Труды Национальной академии наук Соединенных Штатов Америки . 76 (4): 1967–1971. Бибкод : 1979ПНАС...76.1967Б . дои : 10.1073/pnas.76.4.1967 . ПМЦ 383514 . ПМИД 109836 .
- ^ Феррис С.Д., Уилсон AC, Браун WM (апрель 1981 г.). «Эволюционное древо обезьян и человека, основанное на картах расщепления митохондриальной ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 78 (4): 2432–2436. Бибкод : 1981PNAS...78.2432F . дои : 10.1073/pnas.78.4.2432 . ПМК 319360 . ПМИД 6264476 .
- ^ Браун В.М. (июнь 1980 г.). «Полиморфизм в митохондриальной ДНК человека, выявленный с помощью эндонуклеазного анализа рестрикции» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (6): 3605–3609. Бибкод : 1980PNAS...77.3605B . дои : 10.1073/pnas.77.6.3605 . ПМК 349666 . ПМИД 6251473 .
- ^ Канн Р.Л., Браун В.М., Уилсон AC (1982). «Эволюция митохондриальной ДНК человека: предварительный отчет». Прогресс клинических и биологических исследований . 103 Пт А (Пт А): 157–165. ПМИД 6298804 .
- ^ Канн Р.Л., Уилсон AC (август 1983 г.). «Мутации длины в митохондриальной ДНК человека» . Генетика . 104 (4): 699–711. дои : 10.1093/генетика/104.4.699 . ПМК 1202135 . ПМИД 6311667 .
- ^ Jump up to: а б Канн Р. (май 2010 г.). «Все о митохондриальной кануне: интервью с Ребеккой Канн. Интервью Джейн Гитшир» . ПЛОС Генетика . 6 (5): e1000959. дои : 10.1371/journal.pgen.1000959 . ПМЦ 2877732 . ПМИД 20523888 .
- ^ Канн Р.Л., Стоункинг М., Уилсон AC (1987). «Митохондриальная ДНК и эволюция человека». Природа . 325 (6099): 31–36. Бибкод : 1987Natur.325...31C . дои : 10.1038/325031a0 . ПМИД 3025745 . S2CID 4285418 .
- ^ Vigilant L, Stoneking M, Harpending H, Hawkes K, Wilson AC (сентябрь 1991 г.). «Африканские популяции и эволюция митохондриальной ДНК человека». Наука . 253 (5027): 1503–1507. Бибкод : 1991Sci...253.1503V . дои : 10.1126/science.1840702 . ПМИД 1840702 .
- ^ Канн Р.Л. (1987). «В поисках Евы» . Науки . 27 (5): 30–37. дои : 10.1002/j.2326-1951.1987.tb02967.x . ISSN 2326-1951 .
- ^ Jump up to: а б с Левин Р. (октябрь 1987 г.). «Разоблачение митохондриальной Евы». Наука . 238 (4823): 24–26. Бибкод : 1987Sci...238...24L . дои : 10.1126/science.3116666 . ПМИД 3116666 .
- ^ Жилет J (1987). «Эволюция человека. Выход из Эдемского сада» . Природа . 325 (6099): 13. Бибкод : 1987Natur.325...13W . дои : 10.1038/325013a0 . ПМИД 3796736 . S2CID 13187170 .
- ^ Уилкинс А. (27 января 2012 г.). «Ученые, создавшие «Митохондриальную Еву», рассказывают нам о «счастливой матери», навсегда изменившей эволюцию человека» . Гизмодо . Проверено 23 декабря 2019 г.
- ^ Канн Р.Л. (1997). «Глава 4: Матери, ярлыки и женоненавистничество». В Хагере Л.Д. (ред.). Женщины в эволюции человека . Лондон: Рутледж. стр. 75–89. ISBN 9780415108331 .
- ^ Тирни Дж. (1992). «В поисках Адама и Евы» . Newsweek . Институт афроамериканских и африканских исследований Картера Г. Вудсона. Архивировано из оригинала 20 марта 2015 года . Проверено 21 июля 2019 г. - из Интернет-архива.
- ^ Лемоник, доктор медицины (26 января 1987 г.). «Всеобщая генеалогическая мать» . Время . Архивировано из оригинала 23 сентября 2008 года . Проверено 13 мая 2013 г.
- ^ Дарлу П., Тасси П. (1987). «Спорное африканское происхождение человеческой популяции» . Природа . 329 (6135): 111–112. Бибкод : 1987Natur.329..111D . дои : 10.1038/329111b0 . ПМИД 3114640 . S2CID 4313392 .
- ^ Мэддисон Д.Р. (1991). «Пересмотр африканского происхождения митохондриальной ДНК человека». Систематическая зоология . 40 (3): 355–63. дои : 10.2307/2992327 . JSTOR 2992327 .
- ^ Ней М (ноябрь 1992 г.). «Возраст общего предка митохондриальной ДНК человека» . Молекулярная биология и эволюция . 9 (6): 1176–1178. doi : 10.1093/oxfordjournals.molbev.a040785 . ПМИД 1435241 .
- ^ Аяла FJ (декабрь 1995 г.). «Миф о Еве: молекулярная биология и происхождение человека» . Наука . 270 (5244): 1930–1936. Бибкод : 1995Sci...270.1930A . дои : 10.1126/science.270.5244.1930 . ПМИД 8533083 . S2CID 42801341 .
- ^ Темплтон А (март 2002 г.). «Из Африки снова и снова». Природа . 416 (6876): 45–51. Бибкод : 2002Natur.416...45T . дои : 10.1038/416045а . ПМИД 11882887 . S2CID 4397398 .
- ^ Темплтон, Арканзас (декабрь 1997 г.). «Из Африки? Что говорят нам гены?». Текущее мнение в области генетики и развития . 7 (6): 841–847. дои : 10.1016/S0959-437X(97)80049-4 . ПМИД 9468796 .
- ^ Ней М (1992). «Возраст общего предка митохондриальной ДНК человека» . Молекулярная биология и эволюция . 9 (6): 1176–1178. doi : 10.1093/oxfordjournals.molbev.a040785 . ПМИД 1435241 .
- ^ Крингс М., Салем А.Е., Бауэр К., Гейзерт Х., Малек А.К., Шаикс Л. и др. (апрель 1999 г.). «Анализ мтДНК популяций долины реки Нил: генетический коридор или барьер для миграции?» . Американский журнал генетики человека . 64 (4): 1166–1176. дои : 10.1086/302314 . ПМЦ 1377841 . ПМИД 10090902 .
- ^ Крингс М., Стоун А., Шмитц Р.В., Крайницки Х., Стоункинг М., Паабо С. (июль 1997 г.). «Последовательности ДНК неандертальцев и происхождение современного человека» . Клетка . 90 (1): 19–30. дои : 10.1016/S0092-8674(00)80310-4 . hdl : 11858/00-001M-0000-0025-0960-8 . ПМИД 9230299 . S2CID 13581775 .
- ^ Парсонс Т.Дж., Мунец Д.С., Салливан К., Вудятт Н., Аллистон-Грейнер Р., Уилсон М.Р. и др. (апрель 1997 г.). «Высокая наблюдаемая скорость замен в контрольной области митохондриальной ДНК человека». Природная генетика . 15 (4): 363–368. дои : 10.1038/ng0497-363 . ПМИД 9090380 . S2CID 32812244 .
- ^ Холсингер К. (29 сентября 2012 г.). «(Математика слияния) Пример: Митохондриальная Ева» . Лаборатория Холсингера . Проверено 16 мая 2013 г.
- ^ Сайран К.А., Киммел М. (ноябрь 2010 г.). «Альтернативы модели Райта-Фишера: надежность митохондриального датирования Евы». Теоретическая популяционная биология . 78 (3): 165–172. дои : 10.1016/j.tpb.2010.06.001 . ПМИД 20600209 .
- ^ Джайлз Р.Э., Блан Х., Канн Х.М., Уоллес округ Колумбия (ноябрь 1980 г.). «Материнское наследование митохондриальной ДНК человека» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (11): 6715–6719. Бибкод : 1980PNAS...77.6715G . дои : 10.1073/pnas.77.11.6715 . ПМК 350359 . ПМИД 6256757 .
- ^ Бирки CW (август 2008 г.). «Однородительское наследование генов органелл» . Современная биология . 18 (16): Р692–Р695. дои : 10.1016/j.cub.2008.06.049 . ПМИД 18727899 . S2CID 9866662 .
- ^ Гиббонс А. (январь 1998 г.). «Калибровка митохондриальных часов». Наука . 279 (5347): 28–29. Бибкод : 1998Sci...279...28G . дои : 10.1126/science.279.5347.28 . ПМИД 9441404 . S2CID 29855766 .
- ^ Jump up to: а б Оппенгеймер С (2003). Настоящая канун: путешествие современного человека из Африки . Нью-Йорк: Основные книги. ISBN 978-0-7867-1192-5 .
- ^ Виланд С. (2005). «Митохондриальная Ева и библейская Ева выглядят хорошо: критика юного возраста преждевременна» . Журнал творения . 19 (1): 57–59.
- ^ Нельсон CW (1 апреля 2003 г.). «Генетика и библейские демографические события» . Ответы в Бытии . Проверено 16 мая 2013 г.
- ^ Оукс Дж. (25 января 2007 г.). «Могут ли человеческие аргументы о митохондриальной Еве и Y-хромосомном Адаме быть распространены на животный мир, чтобы проверить реальность Ноева потопа?» . Доказательства христианства . Ответы в Бытии . Проверено 16 мая 2013 г.
- ^ Jump up to: а б Докинз Р. (2004). Рассказ предка: паломничество к заре эволюции . Бостон: Хоутон Миффлин. ISBN 978-0-618-00583-3 .
- ^ Докинз Р. (1995). Река из рая: дарвиновский взгляд на жизнь . Основные книги. ISBN 978-0-465-06990-3 . Проверено 5 декабря 2011 г.
- ^ Сайкс Б. (2002). Семь дочерей Евы: наука, раскрывающая наше генетическое происхождение . WW Нортон и компания. ISBN 978-0-393-32314-6 .
- ^ Оппенгеймер, Стивен (2003). Настоящая канун: путешествие современного человека из Африки . Кэрролл и Граф. ISBN 978-0-7867-1192-5 .
- ^ Такахата Н. (январь 1993 г.). «Аллельная генеалогия и эволюция человека» . Молекулярная биология и эволюция . 10 (1): 2–2 doi : 10.1093/oxfordjournals.molbev.a039995 . ПМИД 8450756 .
- ^ «Неужели мы все произошли от общего предка женского пола?» . Как все работает . 14 января 2008 года . Проверено 17 мая 2022 г.
- ^ Изучите JP. «Нет, митохондриальная «Ева» — не первая самка вида» . Смитсоновский журнал . Проверено 23 февраля 2021 г.
- ^ Мендес Ф.Л., Кран Т., Шрак Б., Кран А.М., Вирама К.Р., Вернер А.Е. и др. (март 2013 г.). «Афроамериканская линия по отцовской линии добавляет чрезвычайно древний корень к филогенетическому дереву Y-хромосомы человека» . Американский журнал генетики человека . 92 (3): 454–459. дои : 10.1016/j.ajhg.2013.02.002 . ПМК 3591855 . ПМИД 23453668 .
- ^ Баррас С. (6 марта 2013 г.). «Отцу всех людей 340 000 лет» . Новый учёный . Проверено 13 марта 2013 г.
- ^ Кручиани Ф, Тромбетта Б, Массайя А, Дестро-Бисол Г, Селитто Д, Скоццари Р (июнь 2011 г.). «Пересмотренный корень филогенетического дерева Y-хромосомы человека: происхождение отцовского разнообразия в Африке» . Американский журнал генетики человека . 88 (6): 814–818. дои : 10.1016/j.ajhg.2011.05.002 . ПМК 3113241 . ПМИД 21601174 .
- ^ Каллауэй Э (6 августа 2013 г.). «Генетические Адам и Ева жили не слишком далеко друг от друга во времени» . Природа . дои : 10.1038/nature.2013.13478 . S2CID 170608686 .
- ^ Jump up to: а б Роде Д.Л., Олсон С., Чанг Дж.Т. (сентябрь 2004 г.). «Моделирование недавнего общего происхождения всех живых людей». Природа . 431 (7008): 562–566. Бибкод : 2004Natur.431..562R . CiteSeerX 10.1.1.78.8467 . дои : 10.1038/nature02842 . ПМИД 15457259 . S2CID 3563900 .
- ^ Чжоу, Цзинь; Тео, Йик-Ин (август 2016 г.). «Оценка времени до самого последнего общего предка (TMRCA): сравнение и применение восьми методов» . Европейский журнал генетики человека . 24 (8): 1195–1201. дои : 10.1038/ejhg.2015.258 . ISSN 1476-5438 . ПМЦ 4970674 . ПМИД 26669663 . S2CID 965600 .
Дальнейшее чтение
[ редактировать ]- Аткинсон QD, Грей RD, Драммонд AJ (январь 2009 г.). «Байесовский слиятельный вывод об основных расширениях гаплогрупп митохондриальной ДНК человека в Африке» . Слушания. Биологические науки . 276 (1655): 367–373. дои : 10.1098/rspb.2008.0785 . ПМЦ 2674340 . ПМИД 18826938 .
- Аяла FJ (декабрь 1995 г.). «Миф о Еве: молекулярная биология и происхождение человека» . Наука . 270 (5244): 1930–1936. Бибкод : 1995Sci...270.1930A . дои : 10.1126/science.270.5244.1930 . ПМИД 8533083 .
- Баллу Ф., Хэндли Л.Дж., Жомбар Т., Лю Х., Маника А. (октябрь 2009 г.). «Климат сформировал всемирное распространение вариаций последовательностей митохондриальной ДНК человека» . Слушания. Биологические науки . 276 (1672): 3447–3455. дои : 10.1098/rspb.2009.0752 . ПМК 2817182 . ПМИД 19586946 .
- Бехар Д.М., Виллемс Р., Судьялл Х., Блю-Смит Дж., Перейра Л., Метспалу Э. и др. (май 2008 г.). «Рассвет человеческого матрилинейного разнообразия» . Американский журнал генетики человека . 82 (5): 1130–1140. дои : 10.1016/j.ajhg.2008.04.002 . ПМК 2427203 . ПМИД 18439549 .
- Браун В.М. (июнь 1980 г.). «Полиморфизм в митохондриальной ДНК человека, выявленный с помощью эндонуклеазного анализа рестрикции» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (6): 3605–3609. Бибкод : 1980PNAS...77.3605B . дои : 10.1073/pnas.77.6.3605 . ПМК 349666 . ПМИД 6251473 .
- Член парламента Кокса (август 2008 г.). «Точность молекулярного датирования с помощью статистики ро: отклонения от объединенных ожиданий в рамках ряда демографических моделей». Биология человека . 80 (4): 335–357. дои : 10.3378/1534-6617-80.4.335 . ПМИД 19317593 . S2CID 207701422 .
- Докинз Р. (2004). Рассказ предка: паломничество к заре эволюции . Бостон: Хоутон Миффлин. ISBN 978-0-618-00583-3 .
- Эндикотт П., Хо С.Ю. (апрель 2008 г.). «Байесовская оценка скорости митохондриального замещения человека» . Американский журнал генетики человека . 82 (4): 895–902. дои : 10.1016/j.ajhg.2008.01.019 . ПМЦ 2427281 . ПМИД 18371929 .
- Эндикотт П., Хо С.Ю., Метспалу М., Стрингер С. (сентябрь 2009 г.). «Оценка митохондриальной шкалы эволюции человека». Тенденции в экологии и эволюции . 24 (9): 515–521. дои : 10.1016/j.tree.2009.04.006 . ПМИД 19682765 .
- Экскофье Л., Ян З. (октябрь 1999 г.). «Вариация скорости замещения между сайтами в митохондриальной гипервариабельной области I человека и шимпанзе» . Молекулярная биология и эволюция . 16 (10): 1357–1368. doi : 10.1093/oxfordjournals.molbev.a026046 . ПМИД 10563016 .
- Фельзенштейн Дж. (апрель 1992 г.). «Оценка эффективного размера популяции по образцам последовательностей: неэффективность парных и разделяющих сайтов по сравнению с филогенетическими оценками» . Генетические исследования . 59 (2): 139–147. дои : 10.1017/S0016672300030354 . ПМИД 1628818 .
- Феррис С.Д., Браун В.М., Дэвидсон В.С., Уилсон AC (октябрь 1981 г.). «Обширный полиморфизм в митохондриальной ДНК обезьян» . Труды Национальной академии наук Соединенных Штатов Америки . 78 (10): 6319–6323. Бибкод : 1981PNAS...78.6319F . дои : 10.1073/pnas.78.10.6319 . ПМК 349030 . ПМИД 6273863 .
- Гиббонс А. (январь 1998 г.). «Калибровка митохондриальных часов». Наука . 279 (5347): 28–29. Бибкод : 1998Sci...279...28G . дои : 10.1126/science.279.5347.28 . ПМИД 9441404 . S2CID 29855766 .
- Гондер М.К., Мортенсен Х.М., Рид Ф.А., де Соуза А., Тишкофф С.А. (март 2007 г.). «Анализ последовательности всего генома мтДНК древних африканских линий» . Молекулярная биология и эволюция . 24 (3): 757–768. дои : 10.1093/molbev/msl209 . ПМИД 17194802 .
- Хейзелвуд Л., Стил Дж. (2004). «Пространственная динамика расселения людей: ограничения на моделирование и археологическую проверку». Журнал археологической науки . 31 (6): 669–79. дои : 10.1016/j.jas.2003.11.009 .
- Хо С.Ю., Ларсон Дж. (февраль 2006 г.). «Молекулярные часы: когда времена меняются» . Тенденции в генетике . 22 (2): 79–83. дои : 10.1016/j.tig.2005.11.006 . ПМИД 16356585 .
- Ингман М., Кассманн Х., Паабо С., Гилленстен У. (декабрь 2000 г.). «Вариации митохондриального генома и происхождение современного человека». Природа . 408 (6813): 708–713. Бибкод : 2000Natur.408..708I . дои : 10.1038/35047064 . ПМИД 11130070 . S2CID 52850476 .
- Каессманн Х., Паабо С. (январь 2002 г.). «Генетическая история человека и человекообразных обезьян». Журнал внутренней медицины . 251 (1): 1–18. дои : 10.1046/j.1365-2796.2002.00907.x . ПМИД 11851860 . S2CID 7453672 .
- Кимура М., Ота Т. (2001). «Теоретические аспекты популяционной генетики». Монографии по популяционной биологии . 4 . Издательство Принстонского университета: 1–219. ISBN 978-0-691-08098-7 . ПМИД 5162676 .
- Лоу Л., Шерер С. (ноябрь 1997 г.). «Митохондриальная Ева: сюжет усложняется». Тенденции в экологии и эволюции . 12 (11): 422–423. дои : 10.1016/S0169-5347(97)01204-4 . ПМИД 21238138 .
- Лоогвяли Э.Л., Кивисилд Т., Маргус Т., Виллемс Р. (декабрь 2009 г.). О'Рурк Д. (ред.). «Объяснение несовершенства молекулярных часов митохондрий человекообразных» . ПЛОС ОДИН . 4 (12): е8260. Бибкод : 2009PLoSO...4.8260L . дои : 10.1371/journal.pone.0008260 . ПМЦ 2794369 . ПМИД 20041137 .
- Мака-Мейер Н., Гонсалес А.М., Ларруга Х.М., Флорес С., Кабрера В.М. (2001). «Основные геномные митохондриальные линии определяют раннюю экспансию человека» . БМК Генетика . 2 (1): 13. дои : 10.1186/1471-2156-2-13 . ПМК 55343 . ПМИД 11553319 .
- Мишмар Д., Руис-Песини Е., Голик П., Маколей В., Кларк А.Г., Хоссейни С. и др. (январь 2003 г.). «Естественный отбор сформировал региональные вариации мтДНК у людей» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (1): 171–176. Бибкод : 2003PNAS..100..171M . дои : 10.1073/pnas.0136972100 . ПМК 140917 . ПМИД 12509511 .
- Ней М (ноябрь 1992 г.). «Возраст общего предка митохондриальной ДНК человека» . Молекулярная биология и эволюция . 9 (6): 1176–1178. doi : 10.1093/oxfordjournals.molbev.a040785 . ПМИД 1435241 .
- Нильсен Р., Бомонт М.А. (март 2009 г.). «Статистические выводы в филогеографии» . Молекулярная экология . 18 (6): 1034–1047. дои : 10.1111/j.1365-294X.2008.04059.x . ПМИД 19207258 . S2CID 13613087 .
- Оппенгеймер С (2004). Настоящая канун: путешествие современного человека из Африки . Нью-Йорк: Кэрролл и Граф. ISBN 978-0-7867-1334-9 .
- Причард Дж.К. , Зейлстад М.Т., Перес-Лезон А., Фельдман М.В. (декабрь 1999 г.). «Рост популяции Y-хромосом человека: исследование микросателлитов Y-хромосомы» . Молекулярная биология и эволюция . 16 (12): 1791–1798. doi : 10.1093/oxfordjournals.molbev.a026091 . ПМИД 10605120 .
- Рид Ф.А., Тишкофф С.А. (декабрь 2006 г.). «Африканское человеческое разнообразие, происхождение и миграция». Текущее мнение в области генетики и развития . 16 (6): 597–605. дои : 10.1016/j.где.2006.10.008 . ПМИД 17056248 .
- Роде Д.Л., Олсон С., Чанг Дж.Т. (сентябрь 2004 г.). «Моделирование недавнего общего происхождения всех живых людей». Природа . 431 (7008): 562–566. Бибкод : 2004Natur.431..562R . CiteSeerX 10.1.1.78.8467 . дои : 10.1038/nature02842 . ПМИД 15457259 . S2CID 3563900 .
- Сантос С, Сьерра Б, Альварес Л, Рамос А, Фернандес Э, Ногес Р, Алуха М.П. (август 2008 г.). «Частота и характер гетероплазмии в контрольной области митохондриальной ДНК человека». Журнал молекулярной эволюции . 67 (2): 191–200. Бибкод : 2008JMolE..67..191S . дои : 10.1007/s00239-008-9138-9 . ПМИД 18618067 . S2CID 1143395 .
- Шаффнер С.Ф. (январь 2004 г.). «Х-хромосома в популяционной генетике». Обзоры природы. Генетика . 5 (1): 43–51. дои : 10.1038/nrg1247 . ПМИД 14708015 . S2CID 15772901 .
- Соарес П., Эрмини Л., Томсон Н., Мормина М., Рито Т., Рёль А. и др. (июнь 2009 г.). «Поправка на очищающий отбор: улучшенные митохондриальные молекулярные часы человека» . Американский журнал генетики человека . 84 (6): 740–759. дои : 10.1016/j.ajhg.2009.05.001 . ПМЦ 2694979 . ПМИД 19500773 .
- Суисса С., Ван З., Пул Дж., Витткопп С., Федер Дж., Шатт Т.Э. и др. (май 2009 г.). Десаль Р. (ред.). «Древние генетические варианты мтДНК модулируют транскрипцию и репликацию мтДНК» . ПЛОС Генетика . 5 (5): e1000474. дои : 10.1371/journal.pgen.1000474 . ПМК 2673036 . ПМИД 19424428 .
- Сайкс Б. (2001). Семь дочерей Евы . Нью-Йорк: Нортон. ISBN 978-0-393-02018-2 .
- Сайкс Б.Д. (2003). Проклятие Адама: будущее без мужчин . Лондон: Бантам. ISBN 978-0-593-05005-7 .
- Такахата Н. (январь 1993 г.). «Аллельная генеалогия и эволюция человека» . Молекулярная биология и эволюция . 10 (1): 2–2 doi : 10.1093/oxfordjournals.molbev.a039995 . ПМИД 8450756 .
- Такахата Н., Ли Ш., Сатта Ю. (февраль 2001 г.). «Тестирование мультирегиональности происхождения современного человека» . Молекулярная биология и эволюция . 18 (2): 172–183. doi : 10.1093/oxfordjournals.molbev.a003791 . ПМИД 11158376 .
- Тамура К., Ней М. (май 1993 г.). «Оценка количества нуклеотидных замен в контрольной области митохондриальной ДНК человека и шимпанзе» . Молекулярная биология и эволюция . 10 (3): 512–526. doi : 10.1093/oxfordjournals.molbev.a040023 . ПМИД 8336541 .
- Тан Х, Зигмунд Д.О., Шен П., Офнер П.Дж., Фельдман М.В. (май 2002 г.). «Частая оценка времени слияния на основе данных нуклеотидной последовательности с использованием древовидного разделения» . Генетика . 161 (1): 447–459. дои : 10.1093/генетика/161.1.447 . ПМК 1462078 . ПМИД 12019257 .
- Тишкофф С.А., Гондер М.К., Хенн Б.М., Мортенсен Х., Найт А., Жиньу С. и др. (октябрь 2007 г.). «История говорящих по щелчку популяций Африки, выведенная на основе генетических вариаций мтДНК и Y-хромосомы» . Молекулярная биология и эволюция . 24 (10): 2180–2195. дои : 10.1093/molbev/msm155 . ПМИД 17656633 .
- Тишкофф С.А., Рид Ф.А., Фридлендер Ф.Р., Эрет С., Ранчиаро А., Фромент А. и др. (май 2009 г.). «Генетическая структура и история африканцев и афроамериканцев» . Наука . 324 (5930): 1035–1044. Бибкод : 2009Sci...324.1035T . дои : 10.1126/science.1172257 . ПМЦ 2947357 . ПМИД 19407144 .
- Виджилант Л., Пеннингтон Р., Харпендинг Х., Кохер Т.Д., Уилсон А.С. (декабрь 1989 г.). «Последовательности митохондриальной ДНК в отдельных волосах населения Южной Африки» . Труды Национальной академии наук Соединенных Штатов Америки . 86 (23): 9350–9354. Бибкод : 1989PNAS...86.9350V . дои : 10.1073/pnas.86.23.9350 . ПМК 298493 . ПМИД 2594772 .
- Vigilant L, Stoneking M, Harpending H, Hawkes K, Wilson AC (сентябрь 1991 г.). «Африканские популяции и эволюция митохондриальной ДНК человека». Наука . 253 (5027): 1503–1507. Бибкод : 1991Sci...253.1503V . дои : 10.1126/science.1840702 . ПМИД 1840702 .
- Уотсон Э., Форстер П., Ричардс М., Бандельт Х.Дж. (сентябрь 1997 г.). «Митохондриальные следы человеческой экспансии в Африке» . Американский журнал генетики человека . 61 (3): 691–704. дои : 10.1086/515503 . ПМЦ 1715955 . ПМИД 9326335 .
- Уайт Т.Д., Асфау Б., Бейен И., Хайле-Селассие Ю., Лавджой К.О., Сува Г., ВольдеГабриэль Дж. (октябрь 2009 г.). «Ardipithecus ramidus и палеобиология ранних гоминид» Наука 326 (5949): 75–86. Бибкод : 2009Sci...326... 75W дои : 10.1126/science.1175802 . ПМИД 19810190 . S2CID 20189444 .
- Уайлдер Дж.А., Мобашер З., Хаммер М.Ф. (ноябрь 2004 г.). «Генетические доказательства неравной эффективной численности популяций женщин и мужчин» . Молекулярная биология и эволюция . 21 (11): 2047–2057. дои : 10.1093/molbev/msh214 . ПМИД 15317874 .
- Уилсон А.С., Канн Р.Л., Карр С.М., Джордж М., Гилленстен У.Б., Хелм-Быховски К.М. и др. (1985). «Митохондриальная ДНК и два взгляда на эволюционную генетику». Биологический журнал Линнеевского общества . 26 (4): 375–400. дои : 10.1111/j.1095-8312.1985.tb02048.x .