Генетика
| Часть серии на |
| Генетика |
|---|
 |
|
|
Генетика - это изучение генов , генетических изменений и наследственности в организмах . [ 1 ] [ 2 ] [ 3 ] Это важная ветвь в биологии организмов , потому что наследственность жизненно важна для эволюции . Грегор Мендель , моравский августинский монах, работающий в 19 веке в Брно , был первым, кто научно изучал генетику. Мендель изучал «наследство черты», закономерности в том, как признаки передаются от родителей со временем. Он заметил, что организмы (растения гороха) наследуют признаки посредством дискретных «единиц наследования». Этот термин, все еще используемый сегодня, является несколько неоднозначным определением того, что называется геном.
Черта наследования и механизмы молекулярного наследования генов по -прежнему являются основными принципами генетики в 21 -м веке, но современная генетика расширилась для изучения функции и поведения генов. Структура и функция генов, изменение и распределение изучаются в контексте клетки , организма (например, доминирования ) и в контексте популяции. Генетика вызвала ряд подполи, включая молекулярную генетику , эпигенетику и генетику популяции . Организмы, изученные в широком поле, охватывают домены жизни ( археи , бактерии и эукарья ).
Генетические процессы работают в сочетании с окружающей средой и опытом организма, чтобы влиять на развитие и поведение , часто называемые природой и воспитанием . Внутриклеточная . или внеклеточная среда живой клетки или организма может увеличить или уменьшить транскрипцию генов Классическим примером являются два семяна генетически идентичной кукурузы, один из которых находится в умеренном климате и одно в засушливом климате (отсутствует достаточный водопад или дождь). В то время как средний рост, до того, как две кукурузные стебли могут вырасти, генетически определяется, тот в засушливом климате возрастает лишь на половину высоты в умеренном климате из -за отсутствия воды и питательных веществ в окружающей среде.
Этимология
[ редактировать ]Слово генетика проистекает из древнегреческого γενετικός Genetikos, означающего «родительный падеж»/«генеративный», который, в свою очередь, происходит от γένεσις генезис , означающего «происхождение». [ 4 ] [ 5 ] [ 6 ]
История
[ редактировать ]Наблюдение за тем, что живые существа наследуют черты от их родителей, использовались с доисторических времен для улучшения сельскохозяйственных растений и животных посредством селективного размножения . [ 7 ] [ 8 ] Современная наука генетики, стремящаяся понять этот процесс, началась с работы августинского монаха Грегора Менделя в середине 19-го века. [ 9 ]

До того, как Мендель, Имре Фетертика , венгерский дворянин, который жил в Кесшеге перед Менделем, был первым, кто использовал слово «генетический» в наследственном контексте и считается первым генетиком. Он описал несколько правил биологического наследования в своей работе генетическими законами природы (Die Genetischen Gesetze der Natur, 1819). [ 10 ] Его второй закон такой же, как и тот, который опубликовал Мендель. [ 11 ] В своем третьем законе он разработал основные принципы мутации (его можно считать предшественником Хьюго де Вриса ). [ 12 ] Феститики утверждали, что изменения, наблюдаемые в генерации сельскохозяйственных животных, растений и людей, являются результатом научных законов. [ 13 ] Festetics эмпирически вывели, что организмы наследуют их характеристики, а не приобретают их. Он признал рецессивные черты и присущие вариации, постулируя, что признаки прошлых поколений могут появиться позже, а организмы могут привести к потомству с различными атрибутами. [ 14 ] Эти наблюдения представляют собой важную прелюдию к теории наследования частиц Менделя, поскольку в ней есть переход наследственности от его статуса как мифа к научной дисциплине, предоставляя фундаментальную теоретическую основу для генетики в двадцатом веке. [ 10 ] [ 15 ]

Другие теории наследования предшествовали работе Менделя. Популярная теория в 19 -м веке, и подразумеваемая Чарльзом Дарвином 1859 г. о происхождении видов , была смешиванием наследства : идея, что люди наследуют плавную смесь черт от своих родителей. [ 16 ] Работа Менделя привела примеры, когда черты определенно не были смешаны после гибридизации, показывая, что признаки производятся комбинациями отдельных генов, а не непрерывной смесью. Смешивание признаков в потомстве теперь объясняется действием нескольких генов с количественным эффектом . Другая теория, которая получила некоторую поддержку в то время, - это наследство приобретенных характеристик : вера в то, что люди наследуют черты, усиленные их родителями. эта теория (обычно связанная с Жан-Батист Ламарк )-опыт людей не влияет на гены, которые они передают своим детям. В настоящее время известно, что [ 17 ] Дарвина Другие теории включали пангенезис (который был приобретен и унаследованные аспекты) и Фрэнсиса Гальтона как твердых и унаследованных. переформулировка пангенеза [ 18 ]
Мендельская генетика
[ редактировать ]
Современная генетика началась с исследований Менделя о природе наследования у растений. В своей статье « Версхэ -über pflanzenhybriden » (« Эксперименты по гибридизации растений »), представленные в 1865 году на Naturforschender Verein (Общество исследований в природе) в Brno , Мендель проследил схемы наследования некоторых признаков в растениях гороха и описал их математически математически. Хотя эта схема наследования могла наблюдаться только для нескольких черт, работа Менделя предполагала, что наследственность была частицами, а не приобретенной, и что модели наследования многих признаков могут быть объяснены с помощью простых правил и соотношений. [ 19 ]
Важность работы Менделя не получила широкого понимания до 1900 года, после его смерти, когда Хьюго де Врис и другие ученые вновь открыли его исследования. Уильям Бейтсон , сторонник работы Менделя, придумал слово генетику в 1905 году. [ 20 ] [ 21 ] Прилагательное генетика , полученное из греческого слова Genesis - γένεσις, «Происхождение», предшествует существительному и впервые использовалось в биологическом смысле в 1860 году. [ 22 ] Бейтсон оба действовали в качестве наставника, и ей значительно помогали работа других ученых из колледжа Ньюнхэма в Кембридже, в частности, работы Бекки Сондерс , Норы Дарвин Барлоу и Мюриэля Уилдейла Онслоу . [ 23 ] Бейтсон популяризировал использование слова генетики для описания изучения наследования в своем первом обращении к третьей международной конференции по гибридизации растений в Лондоне в 1906 году. [ 24 ]
После повторного открытия работы Менделя ученые попытались определить, какие молекулы в клетке были ответственны за наследование. В 1900 году Нетти Стивенс начала изучать еду. [ 25 ] В течение следующих 11 лет она обнаружила, что у женщин только X -хромосома и у мужчин были как X, так и y -хромосомы. [ 25 ] Она смогла сделать вывод, что секс является хромосомным фактором и определяется мужчинами. [ 25 ] В 1911 году Томас Хант Морган утверждал, что гены находятся на хромосомах связанной с полом, , основанные на наблюдениях за мутацией белого глаза, у плодовых мух . [ 26 ] В 1913 году его ученик Альфред Стертевант использовал феномен генетической связи, чтобы показать, что гены расположены линейно на хромосоме. [ 27 ]
Молекулярная генетика
[ редактировать ]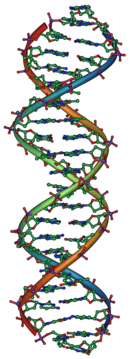
Хотя было известно, что гены существуют на хромосомах, хромосомы состоят из белка и ДНК, и ученые не знали, какие из них ответственны за наследование. В 1928 году Фредерик Гриффит обнаружил феномен трансформации : мертвые бактерии могут перенести генетический материал , чтобы «трансформировать» другие все ещеживые бактерии. Шестнадцать лет спустя, в 1944 году, эксперимент Avery -Macleod -McCarty идентифицировал ДНК как молекулу, ответственную за трансформацию. [ 28 ] Роль ядра в качестве хранилища генетической информации у эукариот была установлена Хеммерлингом в 1943 году в его работе над одноклеточной водоросшей . [ 29 ] Эксперимент Hershey -Chase в 1952 году подтвердил, что ДНК (а не белок) является генетическим материалом вирусов, которые заражают бактерии, предоставляя дополнительные доказательства того, что ДНК является молекулой, ответственной за наследование. [ 30 ]
Джеймс Уотсон и Фрэнсис Крик определили структуру ДНК в 1953 году, используя рентгеновские кристаллографические работы Розалинды Франклина и Мориса Уилкинса , которые указывают на то, что ДНК имеет спиральную структуру (то есть в форме, как штопора). [ 31 ] [ 32 ] Их модель с двойным спиралью имела две пряди ДНК, а нуклеотиды указывают внутрь, каждая из которых соответствует комплементарному нуклеотиду на другой цепи, чтобы сформировать то, что выглядит как ступеньки на витой лестнице. [ 33 ] Эта структура показала, что генетическая информация существует в последовательности нуклеотидов на каждой цепи ДНК. Структура также предложила простой метод репликации : если пряди разделены, новые партнерские нити могут быть реконструированы для каждой на основе последовательности старой нити. Это свойство-это то, что дает ДНК свою полуконсервативную природу, где одна цепь новой ДНК из оригинальной родительской пряди. [ 34 ]
Хотя структура ДНК показала, как работает наследование, все еще не было известно, как ДНК влияет на поведение клеток. В последующие годы ученые пытались понять, как ДНК контролирует процесс производства белка . [ 35 ] Было обнаружено, что клетка использует ДНК в качестве шаблона для создания соответствующей РНК мессенджера , молекулы с нуклеотидами, очень похожими на ДНК. Нуклеотидная последовательность РНК мессенджера используется для создания аминокислотной последовательности в белке; Эта трансляция между нуклеотидными последовательностями и аминокислотными последовательностями известна как генетический код . [ 36 ]
С новым молекулярным пониманием наследования произошло взрыв исследований. [ 37 ] Примечательная теория возникла из Томоко Охты в 1973 году с ее поправкой к нейтральной теории молекулярной эволюции посредством публикации почти нейтральной теории молекулярной эволюции . В этой теории OHTA подчеркнула важность естественного отбора и окружающей среды до скорости, с которой происходит генетическая эволюция . [ 38 ] с концевой цепью Одним из важных разработок было секвенирование ДНК в 1977 году Фредериком Сэнгером . Эта технология позволяет ученым читать нуклеотидную последовательность молекулы ДНК. [ 39 ] В 1983 году Кари Бэнкс Муллис разработал полимеразную цепную реакцию , обеспечивая быстрый способ изолировать и усилить специфический участок ДНК из смеси. [ 40 ] Усилия проекта генома человека , Министерство энергетики, NIH и параллельные частные усилия Celera Genomics привели к секвенированию генома человека в 2003 году. [ 41 ] [ 42 ]
Особенности наследования
[ редактировать ]Дискретное наследство и законы Менделя
[ редактировать ]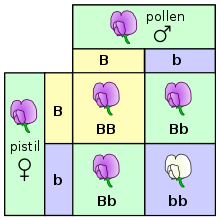
На самом фундаментальном уровне наследование в организмах происходит путем прохождения дискретных наследственных единиц, называемых генами , от родителей до потомства. [ 43 ] Это свойство впервые было замечено Грегором Менделем, который изучал сегрегацию наследственных признаков в растениях гороха , показывая, например, цветы на одном растении были либо фиолетовыми, либо белыми, но никогда не промежуточным между двумя цветами. Дискретные версии того же гена, контролирующего унаследованный внешний вид (фенотипы), называются аллелями . [ 19 ] [ 44 ]
В случае с горохом, который является диплоидным видом, каждое отдельное растение имеет две копии каждого гена, одна копия, унаследованная от каждого родителя. [ 45 ] Многие виды, в том числе люди, имеют эту схему наследования. Диплоидные организмы с двумя копиями одного и того же аллеля данного гена называются гомозиготными в этом геном локусе , в то время как организмы с двумя разными аллелями данного гена называются гетерозиготными . Набор аллелей для данного организма называется его генотипом , в то время как наблюдаемые признаки организма называются его фенотипом . Когда организмы гетерозиготные в ген, часто один аллель называется доминирующим, поскольку его качества доминируют в фенотипе организма, в то время как другой аллель называется рецессивным, поскольку его качества отступают и не наблюдаются. Некоторые аллели не имеют полного доминирования и вместо этого имеют неполное доминирование , экспрессируя промежуточный фенотип или кодоминантность , экспрессируя оба аллеля одновременно. [ 46 ]
Когда пара организмов размножается сексуально , их потомство случайно наследуют один из двух аллелей от каждого родителя. Эти наблюдения за дискретным наследством и сегрегацией аллелей вместе известны как первый закон Менделя или закон сегрегации. Тем не менее, вероятность получения одного гена на другое может измениться из -за доминирующих, рецессивных, гомозиготных или гетерозиготных генов. Например, Мендель обнаружил, что если вы пересекаете гетерозиготные организмы, ваши шансы получить доминирующую черту 3: 1. Реальное генетическое исследование и рассчитывать вероятности с использованием теоретических вероятностей, эмпирических вероятностей, правила продукта, правила суммы и многого другого. [ 47 ]
Обозначения и диаграммы
[ редактировать ]
Генетики используют диаграммы и символы для описания наследования. Ген представлен одним или несколькими буквами. Часто символ «+» используется для обозначения обычного, не мутантного аллеля для гена. [ 48 ]
В экспериментах по оплодотворениям и размножению (и особенно при обсуждении законов Менделя) родители называются генерацией «P» и потомством как «F1» (первое сыновное) поколение. Когда потомство F1 потомки друг с другом, потомство называется «F2» (второе сыновное) генерацию. Одной из общих диаграмм, используемых для прогнозирования результата перекрестного размножения, является площадь Паннетта . [ 49 ]
При изучении генетических заболеваний человека генетики часто используют родословные диаграммы для представления наследования признаков. [ 50 ] Эти диаграммы отображают наследство черты в семейном древе.
Многочисленные генные взаимодействия
[ редактировать ]
Организмы имеют тысячи генов, и в сексуально воспроизводящих организмах эти гены обычно ассорти независимо друг от друга. Это означает, что наследование аллеля на желтый или зеленый горох не связан с наследством аллелей для белых или фиолетовых цветов. Это явление, известное как « Второй закон Менделя » или «Закон независимого ассортимента», означает, что аллели разных генов перетасовывают между родителями для формирования потомства со многими различными комбинациями. Различные гены часто взаимодействуют, чтобы влиять на одну и ту же черту. Например, в голубоглазой Мэри ( Omphalodes Verna ) существует ген с аллелями, которые определяют цвет цветов: синий или пурпурный. Другой ген, однако, контролирует, имеют ли цветы вообще цвет или белые. Когда растение имеет две копии этого белого аллеля, его цветы белые - независимо от того, имеет ли первый ген аллели синего или пурпурного. Это взаимодействие между генами называется эпистазом , со вторым геном эпистатичным к первым. [ 51 ]
Многие черты являются не дискретными чертами (например, фиолетовые или белые цветы), а вместо этого являются непрерывными признаками (например, высота человека и цвет кожи ). Эти сложные черты являются продуктами многих генов. [ 52 ] Влияние этих генов в различной степени опосредовано окружающей средой, которую испытывал организм. Степень, в которой гены организма способствуют сложной черте, называется наследственностью . [ 53 ] Измерение наследуемости признака является относительным - в более переменной среде окружающая среда оказывает большее влияние на общее изменение признака. Например, человеческая высота является чертой с сложными причинами. В Соединенных Штатах он наследуемость составляет 89%. Однако в Нигерии, когда люди испытывают более важный доступ к хорошему питанию и медицинскому обслуживанию , рост имеет наследуемость всего на 62%. [ 54 ]
Молекулярная основа для наследования
[ редактировать ]ДНК и хромосомы
[ редактировать ]
Молекулярной ( ДНК основой для генов является дезоксирибонуклеиновая кислота ). ДНК состоит из дезоксирибозы (молекула сахара), фосфатной группы и основания (группа амина). Существует четыре типа оснований: аденин (A), цитозин (C), гуанин (G) и тимин (T). Фосфаты создают фосфодиэфирные связи с сахарами, чтобы сделать длинные фосфат-сахарные кости. Основы специально сочетаются вместе (T & A, C & G) между двумя магистралью и делают как ступеньки на лестнице. Основы, фосфаты и сахара вместе создают нуклеотид , который соединяется, чтобы сделать длинные цепи ДНК. [ 55 ] Генетическая информация существует в последовательности этих нуклеотидов, и гены существуют в виде участков последовательности вдоль цепи ДНК. [ 56 ] Эти цепи катутся в структуру двойной а-спирали и обертывают белки, называемые гистонами , которые обеспечивают структурную поддержку. ДНК, обернутая вокруг этих гистонов, называются хромосомами. [ 57 ] Вирусы иногда используют сходную молекулу РНК вместо ДНК в качестве их генетического материала. [ 58 ]
ДНК обычно существует в виде двухцепочечной молекулы, нарастанной в форме двойной спирали . Каждый нуклеотид в ДНК преимущественно сочетается со своим партнером нуклеотидом на противоположной цепи: пары с T и C пары C с G. Таким образом, в своей двухцепочечной форме каждая цепь эффективно содержит всю необходимую информацию, избыточную со своей партнерской нити. Эта структура ДНК является физической основой для наследования: репликация ДНК дублирует генетическую информацию, разделяя нити и используя каждую цепь в качестве шаблона для синтеза новой нити партнеров. [ 59 ]

Гены расположены линейно вдоль длинных цепей последовательностей основания ДНК. У бактерий каждая клетка обычно содержит один круглый генофор , в то время как эукариотические организмы (такие как растения и животные) имеют свою ДНК, расположенную в множественных линейных хромосомах. Эти пряди ДНК часто чрезвычайно длинные; Например, крупнейшая хромосома человека составляет около 247 миллионов пар оснований . [ 60 ] ДНК хромосомы связана со структурными белками, которые организуют, компактно и контролируют доступ к ДНК, образуя материал, называемый хроматином ; У эукариот хроматин обычно состоит из нуклеосом , сегментов раны ДНК вокруг ядер гистоновых белков. [ 61 ] Полный набор наследственных материалов в организме (обычно комбинированные последовательности ДНК всех хромосом) называется геномом .
ДНК чаще всего встречается в ядре клеток, но Рут Сагер помогала в открытии нехромосомных генов, обнаруженных за пределами ядра. [ 62 ] У растений они часто встречаются в хлоропластах и в других организмах, в митохондриях. [ 62 ] Эти некромосомные гены все еще могут быть переданы любым партнером в сексуальном размножении, и они контролируют различные наследственные характеристики, которые воспроизводят и остаются активными в течение поколений. [ 62 ]
В то время как гаплоидные организмы имеют только одну копию каждой хромосомы, большинство животных и многих растений являются диплоидными , содержащими две из каждой хромосомы и, следовательно, две копии каждого гена. Два аллеля для гена расположены на идентичных локусах двух гомологичных хромосом , каждый из которых унаследовал от другого родителя. [ 45 ]
Многие виды имеют так называемые половые хромосомы , которые определяют пол каждого организма. [ 63 ] У людей и многих других животных Y -хромосома содержит ген, который запускает развитие специально мужских характеристик. В эволюции эта хромосома потеряла большую часть своего содержания, а также большую часть ее генов, в то время как X -хромосома похожа на другие хромосомы и содержит много генов. При этом Мэри Фрэнсис Лион обнаружила, что во время размножения существует инактивация X-Chromosome, чтобы избежать передачи вдвое больше генов к потомству. [ 64 ] Открытие Лиона привело к открытию заболеваний, связанных с X. [ 64 ]
Размножение
[ редактировать ]
Когда клетки делятся, их полный геном копируется, а каждая дочерняя клетка наследует одну копию. Этот процесс, называемый митоз , является самой простой формой воспроизведения и является основой для бесполого воспроизведения. Асексуальное воспроизведение также может происходить в многоклеточных организмах, производящих потомство, которое наследует их геном от одного родителя. Потомки, которые генетически идентичны их родителям, называются клонами . [ 65 ]
Эукариотические организмы часто используют сексуальное размножение, чтобы генерировать потомство, которое содержит смесь генетического материала, унаследованного от двух разных родителей. Процесс сексуального размножения чередуются между формами, которые содержат отдельные копии генома ( гаплоид ) и двойных копий ( диплоид ). [ 45 ] Гаплоидные клетки сливаются и объединяют генетический материал, чтобы создать диплоидную клетку с парными хромосомами. Диплоидные организмы образуют гаплоиды, делясь без воспроизведения своей ДНК, чтобы создать дочерние клетки, которые случайно наследуют одну из каждой пары хромосом. Большинство животных и многие растения являются диплоидными на протяжении большей части своей жизни, а гаплоидная форма уменьшается до одноклеточных гаметов, таких как сперматозоиды или яйца . [ 66 ]
Несмотря на то, что они не используют гаоплаидный/диплоидный метод сексуального размножения, бактерии имеют много методов получения новой генетической информации. Некоторые бактерии могут подвергаться конъюгации , передавая небольшой круглый кусок ДНК в другую бактерию. [ 67 ] Бактерии также могут принимать необработанные фрагменты ДНК, обнаруженные в окружающей среде и интегрировать их в свои геномы, явление, известное как трансформация . [ 68 ] Эти процессы приводят к горизонтальному переносу генов , передачи фрагментов генетической информации между организмами, которые в противном случае были бы не связаны. Природная бактериальная трансформация происходит у многих видов бактерий и может рассматриваться как половой процесс для передачи ДНК из одной клетки в другую клетку (обычно из одного и того же вида). [ 69 ] Трансформация требует действия многочисленных продуктов бактериального гена , и ее первичной адаптивной функцией, по -видимому, является восстановление в повреждений ДНК клетке реципиента. [ 69 ]
Рекомбинация и генетическая связь
[ редактировать ]
Диплоидная природа хромосом позволяет генам на различных хромосомах независимо от ассорти или отделена от их гомологичной пары во время сексуального размножения, в котором образуются гаплоидные гаметы. Таким образом, новые комбинации генов могут происходить в потомке спаривающейся пары. Гены на той же хромосоме теоретически никогда не будут рекомбинировать. Однако они делают через клеточный процесс хромосомного кроссовера . Во время кроссовера хромосомы обмениваются участками ДНК, эффективно перетасовывая генные аллели между хромосомами. [ 70 ] Этот процесс хромосомного кроссовера обычно происходит во время мейоза , серии клеточных разделений, которые создают гаплоидные клетки. Мейотическая рекомбинация , особенно у микробных эукариот , выполняет адаптивную функцию восстановления повреждений ДНК. [ 69 ]
Первая цитологическая демонстрация пересечения была выполнена Гарриет Крейттон и Барбара МакКлинток в 1931 году. Их исследования и эксперименты на кукурузе предоставили цитологические доказательства генетической теории, которая связывала гены на парных хромосомах, на самом деле обменяют места от одного гомолога к другому. [ 71 ]
Вероятность хромосомного кроссовера, происходящего между двумя данными точками на хромосоме, связана с расстоянием между точками. Для произвольно большего расстояния вероятность кроссовера достаточно высока, чтобы наследование генов эффективно некоррелировано. [ 72 ] Однако для генов, которые ближе друг к другу, более низкая вероятность кроссовера означает, что гены демонстрируют генетическую связь; Аллели для двух генов, как правило, унаследованы вместе. Количество связей между серией генов можно объединить, чтобы сформировать линейную карту связи , которая примерно описывает расположение генов вдоль хромосомы. [ 73 ]
Экспрессия гена
[ редактировать ]Генетический код
[ редактировать ]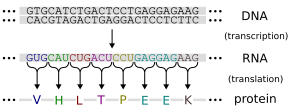
Гены экспрессируют свой функциональный эффект посредством производства белков, которые являются молекулами, ответственными за большинство функций в клетке. Белки состоит из одной или нескольких полипептидных цепей, каждая из которых состоит из последовательности аминокислот . Последовательность ДНК гена используется для получения специфической аминокислотной последовательности . Этот процесс начинается с производства молекулы РНК с последовательности, соответствующей последовательности ДНК гена, процессом, называемой транскрипцией .
Эта молекула РНК мессенджера затем служит для получения соответствующей аминокислотной последовательности посредством процесса, называемого трансляцией . Каждая группа из трех нуклеотидов в последовательности, называемой кодоном , соответствует либо одной из двадцати возможных аминокислот в белке, либо инструкции по прекращению аминокислотной последовательности ; Эта переписка называется генетическим кодом . [ 74 ] Поток информации является однонаправленным: информация переносится из нуклеотидных последовательностей в аминокислотную последовательность белков, но она никогда не переносится из белка обратно в последовательность ДНК - явление Фрэнсис Крик, называемый центральной догмой молекулярной биологии . [ 75 ]
Специфическая последовательность аминокислот приводит к уникальной трехмерной структуре для этого белка, и трехмерные структуры белков связаны с их функциями. [ 76 ] [ 77 ] Некоторые из них являются простыми структурными молекулами, как волокна, образованные протеиновым коллагеном . Белки могут связываться с другими белками и простыми молекулами, иногда действующими как ферменты , облегчая химические реакции в связанных молекулах (без изменения структуры самого белка). Структура белка динамична; Белок гемоглобин изгибается в несколько разных форм, поскольку он облегчает захват, транспортировку и высвобождение молекул кислорода в крови млекопитающих. [ Цитация необходима ]
Единственная разница в нуклеотидах в ДНК может вызвать изменение аминокислотной последовательности белка. Поскольку белковые структуры являются результатом их аминокислотных последовательностей, некоторые изменения могут значительно изменить свойства белка, дестабилизируя структуру или изменяя поверхность белка таким образом, чтобы изменить его взаимодействие с другими белками и молекулами. Например, серповидно-клеточная анемия человека является генетическим заболеванием , которое является результатом единого основания в кодирующей области для сечения β-глобина гемоглобина, что вызывает одно аминокислотное изменение, которое изменяет физические свойства гемоглобина. [ 78 ] Серповые версии гемоглобина придерживаются самих себя, укладывая с образованием волокон, которые искажают форму эритроцитов, несущих белок. Эти серповые клетки больше не протекают плавно через кровеносные сосуды , имея тенденцию засорять или деградировать, вызывая проблемы со здоровьем, связанные с этим заболеванием. [ Цитация необходима ]
Некоторые последовательности ДНК транскрибируются в РНК, но не переводятся в белковые продукты-такие молекулы РНК называются некодирующей РНК . В некоторых случаях эти продукты складываются в структуры, которые участвуют в критических клеточных функциях (например, рибосомальная РНК и передача РНК ). РНК также может иметь регуляторные эффекты посредством гибридизационных взаимодействий с другими молекулами РНК (такими как микроРНК ). [ Цитация необходима ]
Природа и воспитание
[ редактировать ]
Хотя гены содержат всю информацию, которую организм использует для функционирования, окружающая среда играет важную роль в определении конечных фенотипов, которые демонстрирует организм. Фраза « природа и воспитание » относится к этим дополнительным отношениям. Фенотип организма зависит от взаимодействия генов и окружающей среды. Интересным примером является окраска пальто сиамского кота . В этом случае температура тела кошки играет роль окружающей среды. Гены кошки кодируют темные волосы, таким образом, клетки, продуцирующие волосы в кошке, заставляют клеточные белки, что приводит к темным волосам. Но эти темные белки, продуцирующие волосы, чувствительны к температуре (т. Е. Имеют мутация, вызывающую чувствительность температуры) и денатуру в более высоких температурных средах, не образуя пигмента с темноволосой в областях, где у кошки более высокая температура тела. Однако в низкотемпературной среде структура белка стабильна и обычно производит пигмент с темным HAIR. Белок остается функциональным в областях кожи, которые холоднее - такие как его ноги, уши, хвост и лицо - так что у кошки на конечностях темные волосы. [ 79 ]
Окружающая среда играет важную роль в влиянии генетического заболевания человека фенилкетонурия . Мутация, которая вызывает фенилкетонурию, нарушает способность организма разрушать аминокислотный фенилаланин , вызывая токсическое наращивание промежуточной молекулы, которая, в свою очередь, вызывает серьезные симптомы прогрессирующей интеллектуальной инвалидности и приступов. Однако, если кто -то с мутацией фенилкетонурии следует строгой диете, которая избегает этой аминокислоты, он остается нормальным и здоровым. [ 80 ]
Общий метод для определения того, как гены и окружающая среда («природа и воспитание») способствуют фенотипу, включает в себя изучение идентичных и братских близнецов или других братьев и сестер с несколькими родами . [ 81 ] Идентичные братья и сестры генетически одинаковы, так как они происходят из той же зиготы. Между тем, братские близнецы так же генетически отличаются друг от друга, как и нормальные братья и сестры. Сравнивая, как часто определенное расстройство происходит в паре идентичных близнецов с тем, как часто оно происходит в паре братских близнецов, ученые могут определить, вызвано ли это расстройство генетическими или постнатальными факторами окружающей среды. Один известный пример включал в себя изучение четырехлетных четырехлетных квадруплетов , которые были идентичными четырехлетними в четыре раза, диагностированные с шизофренией . [ 82 ]
Генная регуляция
[ редактировать ]Геном данного организма содержит тысячи генов, но не все эти гены должны быть активными в любой момент. Ген экспрессируется, когда он транскрибируется в мРНК, и существует много клеточных методов контроля экспрессии генов, таких что белки продуцируются только при необходимости клетки. Факторы транскрипции - это регуляторные белки, которые связываются с ДНК, либо способствуют или ингибируют транскрипцию гена. [ 83 ] Например, в геноме бактерий Escherichia coli существует ряд генов, необходимых для синтеза аминокислотного триптофана . Однако, когда триптофан уже доступен для клетки, эти гены для синтеза триптофана больше не нужны. Наличие триптофана напрямую влияет на активность генов - молекулы для триптофана связываются с репрессором триптофана (фактор транскрипции), изменяя структуру репрессора, так что репрессор связывается с генами. Репрессор триптофана блокирует транскрипцию и экспрессию генов, тем самым создавая отрицательную регуляцию обратной связи процесса синтеза триптофана. [ 84 ]
Различия в экспрессии генов особенно ясны в многоклеточных организмах , где все клетки содержат один и тот же геном, но имеют очень разные структуры и поведение из -за экспрессии различных наборов генов. Все клетки в многоклеточном организме происходят из одной клетки, дифференцируя в вариантные типы клеток в ответ на внешние и межклеточные сигналы и постепенно устанавливая различные паттерны экспрессии генов для создания различных поведений. Поскольку ни один ген не несет ответственности за развитие структур в многоклеточных организмах, эти паттерны возникают из -за сложных взаимодействий между многими клетками. [ Цитация необходима ]
В эукариотах существуют структурные особенности хроматина , которые влияют на транскрипцию генов, часто в форме модификаций в ДНК и хроматина, которые стабильно унаследованы дочерними клетками. [ 85 ] Эти особенности называются « эпигенетическими », потому что они существуют »сверху» последовательности ДНК и сохраняют наследование от одной генерации клеток к другому. Из -за эпигенетических особенностей различные типы клеток, выращенные в одной и той же среде, могут сохранять очень разные свойства. Хотя эпигенетические особенности, как правило, динамичны в ходе разработки, некоторые, как и явление параметизации , имеют многопокол наследование и существуют как редкие исключения из общего правила ДНК в качестве основы для наследования. [ 86 ]
Генетическое изменение
[ редактировать ]Мутации
[ редактировать ]
Во время процесса репликации ДНК ошибки иногда возникают в полимеризации второй цепи. Эти ошибки, называемые мутациями, могут влиять на фенотип организма, особенно если они встречаются в кодирующей последовательности белка гена. Показатели ошибок обычно очень низки - 1 ошибка в каждые 10–100 миллионов баз - до «корректурирования» способности ДНК -полимеразы . [ 87 ] [ 88 ] Процессы, которые увеличивают скорость изменений в ДНК, называются мутагенными : мутагенные химические вещества способствуют ошибкам в репликации ДНК, часто вмешиваясь в структуру основания, в то время как ультрафиолетовое излучение индуцирует мутации, вызывая повреждение структуры ДНК. [ 89 ] Химическое повреждение ДНК также происходит естественным путем, и клетки используют механизмы репарации ДНК для восстановления несоответствий и разрывов. Ремонт, однако, не всегда восстанавливает исходную последовательность. Особенно важным источником повреждений ДНК, по -видимому, является реактивная форма кислорода [ 90 ] Производится клеточным аэробным дыханием , и они могут привести к мутациям. [ 91 ]
У организмов, которые используют хромосомный кроссовер для обмена ДНК и рекомбинированных генов, ошибки при выравнивании во время мейоза также могут вызывать мутации. Ошибки в кроссовере особенно вероятны, когда подобные последовательности заставляют партнерских хромосомов принять ошибочное выравнивание; Это делает некоторые области в геномах более склонными к мутированию таким образом. Эти ошибки создают большие структурные изменения в последовательности ДНК - дупликации , инверсии , делеции целых областей - или случайного обмена целыми частями последовательностей между различными хромосом, хромосомная транслокация . [ 92 ]

Естественный отбор и эволюция
[ редактировать ]Мутации изменяют генотип организма, и иногда это вызывает различные фенотипы. Большинство мутаций мало влияют на фенотип организма, здоровье или репродуктивную подготовку . [ 93 ] Мутации, которые оказывают эффект, обычно вредны, но иногда некоторые могут быть полезными. [ 94 ] Исследования на мух Drosophila melanogaster позволяют предположить, что если мутация изменяет белок, продуцируемый геном, около 70 процентов этих мутаций вредны, поскольку остальные являются либо нейтральными, либо слабо полезными. [ 95 ]

Генетика популяции изучает распределение генетических различий в популяциях и то, как эти распределения меняются с течением времени. [ 96 ] На изменения частоты аллеля в популяции в основном влияют естественный отбор , где данное аллель дает избирательное или репродуктивное преимущество для организма, [ 97 ] а также другие факторы, такие как мутация , генетический дрейф , генетический автостоп [ 98 ] Искусственный отбор и миграция . [ 99 ]
В течение многих поколений геномы организмов могут значительно измениться, что приводит к эволюции. В процессе, называемом адаптацией , отбор для полезных мутаций может привести к тому, что вид может развиваться в формы, способные выжить в окружающей среде. [ 100 ] Новые виды формируются в процессе видообразования , часто вызванных географическим разделением, которые предотвращают обменять гены населения друг с другом. [ 101 ]
Сравнивая гомологию между геномами разных видов, можно рассчитать эволюционное расстояние между ними и когда они могли разочароваться . Генетические сравнения, как правило, считаются более точным методом характеристики родства между видами, чем сравнение фенотипических характеристик. Эволюционные расстояния между видами могут быть использованы для формирования эволюционных деревьев ; Эти деревья представляют собой общий спуск и дивергенцию видов с течением времени, хотя они не показывают перенос генетического материала между неродственными видами (известными как горизонтальный перенос генов и наиболее распространенную у бактерий). [ 102 ]
Исследования и технологии
[ редактировать ]Модель организмов
[ редактировать ]
Хотя генетики первоначально изучали наследование в самых разных организмах, диапазон изученных видов сузился. Одна из причин заключается в том, что когда существует значительное исследование для данного организма, новые исследователи с большей вероятностью выбирают его для дальнейшего изучения, и поэтому в конечном итоге несколько модельных организмов стали основой для большинства исследований в области генетики. Общие темы исследования в генетике модели организма включают изучение регуляции генов и участие генов в развитие и рак . Организмы были выбраны, отчасти, для удобства - время поколения и легкие генетические манипуляции сделали некоторые организмы популярных инструментов исследования генетики. Widely used model organisms include the gut bacterium Escherichia coli , the plant Arabidopsis thaliana , baker's yeast ( Saccharomyces cerevisiae ), the nematode Caenorhabditis elegans , the common fruit fly ( Drosophila melanogaster ), the zebrafish ( Danio rerio ), and the common house mouse ( Mus Musculus ). [ 103 ]
Лекарство
[ редактировать ]
Медицинская генетика стремится понять, как генетические вариации связаны с здоровьем и заболеванием человека. [ 104 ] При поиске неизвестного гена, который может быть вовлечен в болезнь, исследователи обычно используют генетическую связь и генетические родословные , чтобы найти местоположение на геноме, связанном с заболеванием. На уровне популяции исследователи пользуются преимуществами рандомизации Менделя, чтобы искать места в геноме, которые связаны с заболеваниями, метод, особенно полезный для многогенных признаков, не четко определяемых одним геном. [ 105 ] После того, как ген кандидата обнаруживается, часто проводится дальнейшие исследования на соответствующих (или гомологичных ) генах модельных организмов. В дополнение к изучению генетических заболеваний, повышенная доступность методов генотипирования привела к области фармакогенетики : изучение того, как генотип может влиять на лекарственные реакции. [ 106 ]
Люди различаются по своей наследственной тенденции к развитию рака , а рак является генетическим заболеванием. Процесс развития рака в организме представляет собой комбинацию событий. Мутации иногда встречаются в клетках организма, когда они делятся. Хотя эти мутации не будут унаследованы какими -либо потомками, они могут влиять на поведение клеток, иногда вызывая их расти и разделиться чаще. Существуют биологические механизмы, которые пытаются остановить этот процесс; Сигналы даются неуместно делящим клеткам, которые должны вызывать гибель клеток , но иногда встречаются дополнительные мутации, которые заставляют клетки игнорировать эти сообщения. Внутренний процесс естественного отбора происходит в организме, и в конечном итоге мутации накапливаются в клетках, чтобы способствовать их собственному росту, создавая раковую опухоль , которая растет и вторгается в различные ткани тела. Обычно клетка делится только в ответ на сигналы, называемые факторами роста , и перестает расти один раз в контакте с окружающими клетками и в ответ на сигналы ингибирования роста. Обычно он делит ограниченное количество раз и умирает, оставаясь в пределах Эпителий , где он не может мигрировать в другие органы. Чтобы стать раковой клеткой, клетка должна накапливать мутации в ряде генов (от трех до семи). Раковая клетка может делиться без фактора роста и игнорировать ингибирующие сигналы. Кроме того, он бессмертный и может расти на неопределенный срок, даже после того, как он вступает в контакт с соседними клетками. Он может выйти из эпителия и, в конечном итоге, от первичной опухоли . Затем сбежанная клетка может пересечь эндотелий кровеносного сосуда и транспортироваться кровотоком, чтобы колонизировать новый орган, образуя смертоносные метастазии . Хотя в небольшой доли рака существуют некоторые генетические предрасположенности, основная фракция обусловлена набором новых генетических мутаций, которые первоначально появляются и накапливаются в одном или небольшом количестве клеток, которые будут делиться, образуя опухоль и не передаются на Потомство ( соматические мутации ). Наиболее частыми мутациями являются потеря функции белка p53 , супрессора опухоли или в пути p53 и усиление функциональных мутаций в Белки Ras , или в других онкогенах . [ 107 ] [ 108 ]
Методы исследования
[ редактировать ]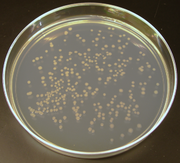
ДНК можно манипулировать в лаборатории. Ограничительные ферменты обычно используются ферментами, которые разрезают ДНК в специфических последовательностях, продуцируя предсказуемые фрагменты ДНК. [ 109 ] Фрагменты ДНК могут быть визуализированы путем использования гелевого электрофореза , который отделяет фрагменты в соответствии с их длиной. [ 110 ]
Использование лигирования ферментов позволяет подключать фрагменты ДНК. Связывая («лигирование») фрагменты ДНК вместе из разных источников, исследователи могут создавать рекомбинантную ДНК , ДНК часто ассоциируется с генетически модифицированными организмами . Рекомбинантная ДНК обычно используется в контексте плазмид : короткие кольцевые молекулы ДНК с несколькими генами на них. В процессе, известном как молекулярное клонирование , исследователи могут амплифицировать фрагменты ДНК, вставляя плазмиды в бактерии, а затем культивируя их на пластинах агара (для изоляции клонов бактерий клеток ). «Клонирование» также может относиться к различным средствам создания клонированных («клональных») организмов. [ 111 ]
ДНК также может быть амплифицирована с использованием процедуры, называемой полимеразной цепной реакцией (ПЦР). [ 112 ] Используя специфические короткие последовательности ДНК, ПЦР может изолировать и экспоненциально усилить целевую область ДНК. Поскольку он может амплифицироваться из чрезвычайно небольших количеств ДНК, ПЦР также часто используется для обнаружения присутствия специфических последовательностей ДНК. [ 113 ] [ 114 ]
Секвенирование ДНК и геномика
[ редактировать ]Секвенирование ДНК, одна из наиболее фундаментальных технологий, разработанных для изучения генетики, позволяет исследователям определять последовательность нуклеотидов в фрагментах ДНК. Техника секвенирования сети , разработанная в 1977 году командой, возглавляемой Фредериком Сэнгером , все еще обычно используется для последовательности фрагментов ДНК. Используя эту технологию, исследователи смогли изучить молекулярные последовательности, связанные со многими заболеваниями человека. [ 115 ]
Поскольку секвенирование стало дешевле, исследователи секвенировали геномы многих организмов, используя процесс, называемый сборкой генома , который использует вычислительные инструменты для сшивания последовательностей из многих различных фрагментов. [ 116 ] Эти технологии были использованы для последовательности генома человека в проекте генома человека, завершенного в 2003 году. [ 41 ] Новые высокопроизводительные технологии секвенирования значительно снижают стоимость секвенирования ДНК, и многие исследователи надеются снизить стоимость переоценки генома человека до тысячи долларов. [ 117 ]
Секвенирование следующего поколения (или высокопроизводительное секвенирование) произошло из-за постоянно растущей спроса на недорогое секвенирование. Эти технологии секвенирования позволяют одновременно производство потенциально миллионов последовательностей. [ 118 ] [ 119 ] Большое количество доступных данных о последовательности создало под поле геномики , исследования, которые используют вычислительные инструменты для поиска и анализа закономерностей в полных геномах организмов. Геномика также может считаться подполом биоинформатики , в которой используются вычислительные подходы для анализа больших наборов биологических данных . Распространенная проблема для этих областей исследований заключается в том, как управлять и делиться данными, которые касаются человеческого субъекта и личной информации . [ Цитация необходима ]
Общество и культура
[ редактировать ]19 марта 2015 года группа ведущих биологов призвала всемирный запрет на клиническое использование методов, в частности, использование CRISPR и цинкового пальца , чтобы редактировать геном человека таким образом, который может быть унаследован. [ 120 ] [ 121 ] [ 122 ] [ 123 ] В апреле 2015 года китайские исследователи сообщили о результатах фундаментальных исследований для редактирования ДНК нежизнеспособных человеческих эмбрионов с использованием CRISPR. [ 124 ] [ 125 ]
Смотрите также
[ редактировать ]- Бактериальный размер генома
- Криоконсервация генетических ресурсов животных
- Евгеника
- Эмбриология
- Генетическое расстройство
- Генетическое разнообразие
- Генетическая инженерия
- Генетическое улучшение
- Глоссарий генетики (M - z)
- Индекс статей генетики
- Медицинская генетика
- Молекулярные инструменты для изучения генов
- Нейроэпигенетика
- Схема генетики
- Временная шкала истории генетики
- Генетические ресурсы растений
Ссылки
[ редактировать ]- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Генетика и организм: введение» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Hartl D, Jones E (2005)
- ^ «Определение генетики» . www.dictionary.com . Получено 25 октября 2018 года .
- ^ "Genetikos (γενετ-ικός)" . Генри Джордж Лидделл, Роберт Скотт, греко-английский лексикон . Цифровая библиотека Персея, Университет Тафтса. Архивировано из оригинала 15 июня 2010 года . Получено 20 февраля 2012 года .
- ^ «Бытие (γένεσις)» . Генри Джордж Лидделл, Роберт Скотт, греко-английский лексикон . Цифровая библиотека Персея, Университет Тафтса. Архивировано из оригинала 15 июня 2010 года . Получено 20 февраля 2012 года .
- ^ «Генетический» . Онлайн этимологический словарь . Архивировано из оригинала 23 августа 2011 года . Получено 20 февраля 2012 года .
- ^ Наука: окончательное визуальное руководство . Пингвин. 2009. с. 362. ISBN 978-0-7566-6490-9 .
- ^ Poczai P, Santiago-Blay JA (июль 2022 г.). «Темы биологического наследства в размножении овец в начале девятнадцатого века, как выявили JM Ehrenfels» . Гены . 13 (8): 1311. doi : 10.3390/genes13081311 . PMC 9332421 . PMID 35893050 .
- ^ Weiling F (июль 1991 г.). «Историческое исследование: Иоганн Грегор Мендель 1822-1884». Американский журнал медицинской генетики . 40 (1): 1–25, обсуждение 26. doi : 10.1002/ajmg.1320400103 . PMID 1887835 .
- ^ Jump up to: а беременный Poczai P, Santiago-Blay JA (октябрь 2021 г.). «Принципы и биологические концепции наследственности перед Менделем» . Биология прямой . 16 (1): 19. DOI : 10.1186/S13062-021-00308-4 . PMC 8532317 . PMID 34674746 .
 Текст был скопирован из этого источника, который доступен в рамках Attribution Creative Commons 4.0 Международная лицензия, архивная 16 октября 2017 года на машине Wayback .
Текст был скопирован из этого источника, который доступен в рамках Attribution Creative Commons 4.0 Международная лицензия, архивная 16 октября 2017 года на машине Wayback .
- ^ Szabó AT, Poczai P (июнь 2019 г.). «Появление генетики от овец Festetics через горох Менделя до цыплят Бейтсона». Журнал генетики . 98 (2): 63. doi : 10.1007/s12041-019-1108-z . HDL : 10138/324962 . PMID 31204695 . S2CID 174803150 .
- ^ Poczai P, Bell N, Hyvönen J (январь 2014 г.). «Imre Festetics и Общество заводчиков овец Моравии: забытая исследовательская сеть Менделя » . PLOS Биология . 12 (1): E1001772. doi : 10.1371/journal.pbio.1001772 . PMC 3897355 . PMID 24465180 .
- ^ Poczai P (2022). Наследственность перед Менделем: праздники и вопрос о овечьей шерсти в центральной Европе . Бока Ратон, Флорида: CRC Press. п. 113. ISBN 978-1-032-02743-2 Полем Получено 30 августа 2022 года .
- ^ Poczai P, Santiago-Blay JA, Sekerák J, Bariska I, Szabó AT (октябрь 2022 г.). «Mimush Sheep и призрак инбридинга: исторический фон для органических и генетических законов Festetics за четыре десятилетия до экспериментов Менделя в городе» . Журнал истории биологии . 55 (3): 495–536. doi : 10.1007/s10739-022-09678-5 . PMC 9668798 . PMID 35670984 . S2CID 249433049 .
- ^ Poczai P, Santiago-Blay JA (2022). «Чисти старый блок: поколение, развитие и наследственные концепции наследственности» . Границы в генетике . 13 : 814436. DOI : 10.3389/fgene.2022.814436 . PMC 8959437 . PMID 35356423 .
- ^ Гамильтон Х (2011). Генетика популяции . Джорджтаунский университет. п. 26. ISBN 978-1-4443-6245-9 .
- ^ Lamarck, JB (2008). В Encyclopædia Britannica . Получено из Encyclopædia Britannica Online Archived 14 апреля 2020 года на машине Wayback 16 марта 2008 года.
- ^ Петр Дж. Боулер , Мендельская революция: чрезвычайная поступка наследственных концепций в современной науке и обществе (Балтимор: издательство Джона Хопкинса Университета, 1989): главы 2 и 3.
- ^ Jump up to: а беременный Blumberg RB. «Бумага Менделя на английском языке» . Архивировано с оригинала 13 января 2016 года.
- ^ Генетика, н. Оксфордский английский словарь , 3 -е изд.
- ^ Бейтсон В. "Письмо Уильяма Бейтсона Алану Седжвику в 1905 году" . Центр Джона Иннеса. Архивировано из оригинала 13 октября 2007 года . Получено 15 марта 2008 года . Письмо было Адам Седжвик, зоолог и «Читатель морфологии животных» в Тринити -колледже, Кембридж
- ^ Генетический, прил. Оксфордский английский словарь, 3 -е изд.
- ^ Ричмонд М.Л. (ноябрь 2007 г.). «Возможности для женщин в ранней генетике» . Природные обзоры. Генетика . 8 (11): 897–902. doi : 10.1038/nrg2200 . PMID 17893692 . S2CID 21992183 . Архивировано из оригинала 16 мая 2008 года.
- ^ Бейтсон В. (1907). «Прогресс генетических исследований». В Уилксе, W (ред.). Отчет Третьей Международной конференции по генетике 1906 года: гибридизация (скрещивание родов или видов), скрещивание разновидностей и общее размножение растений . Лондон: Королевское садоводческое общество. : Первоначально назвал «Международную конференцию по гибридизации и разведению растений», название было изменено в результате речи Бейтсона. Видеть: Cock AG, Forsdyke DR (2008). Создайте свои исключения: наука и жизнь Уильяма Бейтсона . Спрингер. п. 248 ISBN 978-0-387-75687-5 .
- ^ Jump up to: а беременный в «Нетти Стивенс: открыватель сексуальных хромосом» . Scaute . Природное образование . Получено 8 июня 2020 года .
- ^ Мур JA (1983). «Томас Хант Морган - генетик». Интегративная и сравнительная биология . 23 (4): 855–865. doi : 10.1093/icb/23.4.855 .
- ^ Sturtevant AH (1913). «Линейное расположение шести связанных с полом факторов у дрозофилы, как показано в их режиме ассоциации» (PDF) . Журнал экспериментальной биологии . 14 (1): 43–59. Bibcode : 1913jez .... 14 ... 43 с . Citeseerx 10.1.1.37.9595 . doi : 10.1002/jez.1400140104 . S2CID 82583173 . Архивировано (PDF) из оригинала 27 февраля 2008 года.
- ^ Avery OT, Macleod CM, McCarty M (февраль 1944 г.). «Исследования химической природы вещества, индуцирующей трансформацию пневмококковых типов: индукция трансформации с помощью дроссельной фракции дезоксирибонуклеиновой кислоты, выделенной из пневмококка типа III» . Журнал экспериментальной медицины . 79 (2): 137–158. doi : 10.1084/jem.79.2.137 . PMC 2135445 . PMID 19871359 . Перепечатка: Avery OT, Macleod CM, McCarty M (февраль 1979 г.). «Исследования химической природы вещества, индуцирующей трансформацию пневмококковых типов. Индукции трансформации с помощью дроссерибонуклеиновой кислоты, выделенной из пневмококка типа III» . Журнал экспериментальной медицины . 149 (2): 297–326. doi : 10.1084/jem.149.2.297 . PMC 2184805 . PMID 33226 .
- ^ Ханна П (2008). Клеточная и молекулярная биология . IK International Pvt Ltd. p. 221. ISBN 978-81-89866-59-4 .
- ^ Херши ad, Чейз М (май 1952). «Независимые функции вирусного белка и нуклеиновой кислоты в росте бактериофагов» . Журнал общей физиологии . 36 (1): 39–56. doi : 10.1085/jgp.36.1.39 . PMC 2147348 . PMID 12981234 .
- ^ Джадсон Х (1979). Восьмой день творения: производители революции в биологии . Cold Spring Harbor Laboratory Press. С. 51–169. ISBN 978-0-87969-477-7 .
- ^ Уотсон Д.Д., Крик Ф.Х. (апрель 1953 г.). «Молекулярная структура нуклеиновых кислот; структура дезоксирибозной нуклеиновой кислоты» (PDF) . Природа . 171 (4356): 737–738. Bibcode : 1953natur.171..737W . doi : 10.1038/171737a0 . PMID 13054692 . S2CID 4253007 . Архивировано (PDF) из оригинала 4 февраля 2007 года.
- ^ Watson JD, Crick FH (май 1953). «Генетические последствия структуры дезоксирибонуклеиновой кислоты» (PDF) . Природа . 171 (4361): 964–967. Bibcode : 1953natur.171..964W . doi : 10.1038/171964b0 . PMID 13063483 . S2CID 4256010 . Архивировано (PDF) из оригинала 21 июня 2003 года.
- ^ Stratmann SA, Van Oijen Am (февраль 2014 г.). «Репликация ДНК на уровне одной молекулы» (PDF) . Обзоры химического общества . 43 (4): 1201–1220. doi : 10.1039/c3cs60391a . PMID 24395040 . S2CID 205856075 . Архивировано (PDF) из оригинала 6 июля 2017 года.
- ^ Фредерик Б. (2010). Управление наукой: методология и организация исследований . Спрингер. п. 76. ISBN 978-1-4419-7488-4 .
- ^ Райс С.А. (2009). Энциклопедия эволюции . Infobase Publishing. п. 134. ISBN 978-1-4381-1005-9 .
- ^ Саркар С. (1998). Генетика и редукционизм . Издательство Кембриджского университета. п. 140. ISBN 978-0-521-63713-8 .
- ^ OHTA T (ноябрь 1973 г.). «Слегка вредные мутанты замены в эволюции». Природа . 246 (5428): 96–98. Bibcode : 1973natur.246 ... 96o . doi : 10.1038/246096a0 . PMID 4585855 . S2CID 4226804 .
- ^ Сангер Ф., Никлен С., Коулсон А.Р. (декабрь 1977 г.). «Секвенирование ДНК с цепными ингибиторами» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (12): 5463–5467. Bibcode : 1977pnas ... 74.5463S . doi : 10.1073/pnas.74.12.5463 . PMC 431765 . PMID 271968 .
- ^ Saiki RK, Scharf S, Faloona F, Mullis KB, Horn GT, Erlich HA, et al. (Декабрь 1985). «Ферментативная амплификация геномных последовательностей бета-глобина и анализа сайтов ограничения для диагностики серповидноклеточной анемии». Наука . 230 (4732): 1350–1354. Bibcode : 1985sci ... 230.1350S . doi : 10.1126/science.2999980 . PMID 2999980 .
- ^ Jump up to: а беременный «Информация о проекте человека генома человека» . Проект генома человека. Архивировано из оригинала 15 марта 2008 года . Получено 15 марта 2008 года .
- ^ «Последовательность человеческого генома». Наука . 291 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Модели наследования: введение» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Эксперименты Менделя» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Jump up to: а беременный в Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Мендельская генетика в эукариотических жизненных циклах» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Взаимодействие между аллелями одного гена» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ «Вероятности в генетике (статья)» . Ханская академия . Получено 28 сентября 2022 года .
- ^ Чейни Р.В. «Генетическая нотация» . Университет Кристофер Ньюпорт. Архивировано из оригинала 3 января 2008 года . Получено 18 марта 2008 года .
- ^ Müller-Wille S, Parolini G (9 декабря 2020 г.). «Паннетт квадраты и гибридные кресты: как Мендельанцы изучили свою торговлю по книге». Обучение книгой: Руководства и справочники в истории науки . Темы BJHS. Тол. 5. Британское общество по истории науки / издательство Кембриджского университета . С. 149–165. doi : 10.1017/bjt.2020.12 . S2CID 2293444415 . Архивировано из оригинала 29 марта 2021 года . Получено 29 марта 2021 года .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Человеческая генетика» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Взаимодействие генов и модифицированные дигибридные соотношения» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ MAYEUX R (июнь 2005 г.). «Картирование новой границы: сложные генетические расстройства» . Журнал клинических исследований . 115 (6): 1404–1407. doi : 10.1172/jci25421 . PMC 1137013 . PMID 15931374 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Количественная наследуемость» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Люк А., Го Х., Адеймо А.А., Уилкс Р., Форрестер Т., Лоу В. и др. (Июль 2001 г.). «Наследственность черт, связанных с ожирением, среди нигерийцев, ямайцев и нас, чернокожих людей» . Международный журнал ожирения и связанных с ними метаболических расстройств . 25 (7): 1034–1041. doi : 10.1038/sj.ijo.0801650 . PMID 11443503 .
- ^ Урри Л., Каин М., Вассерман С., Минорский П., Рис Дж., Кэмпбелл Н. "Биология Кэмпбелла" . plus.pearson.com . Получено 28 сентября 2022 года .
- ^ Пирсон H (май 2006 г.). "Генетика: что такое ген?" Полем Природа . 441 (7092): 398–401. Bibcode : 2006natur.441..398p . doi : 10.1038/441398a . PMID 16724031 . S2CID 4420674 .
- ^ "Гистон" . Genome.gov . Получено 28 сентября 2022 года .
- ^ Прескотт Л.М., Харли Дж.П., Кляйн Д.А. (1996). Микробиология (3 -е изд.). Wm. C. Браун. п. 343. ISBN 0-697-21865-1 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Механизм репликации ДНК» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Грегори С.Г., Барлоу К.Ф., Маклай К.Е., Каул Р., Сварбрек Д., Данхэм А. и др. (Май 2006 г.). «Последовательность ДНК и биологическая аннотация хромосомы человека 1» . Природа . 441 (7091): 315–321. Bibcode : 2006natur.441..315G . doi : 10.1038/nature04727 . PMID 16710414 .
- ^ Alberts et al. (2002), II.4. упаковка в хроматиновом волокно ее ДНК и хромосомы: хромосомная ДНК и
- ^ Jump up to: а беременный в "Рут Сагер" . Энциклопедия Британская . Получено 8 июня 2020 года .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Секс-хромосомы и связанные с полом наследство» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Jump up to: а беременный Rastan S (февраль 2015 г.). «Мэри Ф. Лион (1925-2014)» . Природа . 518 (7537). Springer Nature Limited: 36. Bibcode : 2015natur.518 ... 36r . doi : 10.1038/518036a . PMID 25652989 . S2CID 4405984 .
- ^ "Клон" . Мерриам-Уэбстер Словарь . Получено 13 ноября 2023 года .
- ^ "Гаплоид" . www.genome.gov . Получено 10 февраля 2024 года .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Бактериальное сопряжение» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Бактериальная трансформация» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Jump up to: а беременный в Бернштейн Х, Бернштейн С., Мичод Р.Е. (январь 2018 г.). «Секс в микробных патогенах» . Инфекция, генетика и эволюция . 57 : 8–25. Bibcode : 2018infge..57 .... 8b . doi : 10.1016/j.meegid.2017.10.024 . PMID 29111273 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbar, Eds. (2000). «Природа пересечения» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Creighton HB, McClintock B (август 1931 г.). «Корреляция цитологического и генетического пересечения в Зеа Мэйс» . Труды Национальной академии наук Соединенных Штатов Америки . 17 (8): 492–497. Bibcode : 1931pnas ... 17..492c . doi : 10.1073/pnas.17.8.492 . PMC 1076098 . PMID 16587654 .
- ^ Staub JE (1994). Кроссовер: концепции и приложения в генетике, эволюции и размножении . Университет Висконсин Пресс. п. 55. ISBN 978-0-299-13564-5 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbar, Eds. (2000). "Карты связи" . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Berg JM, Tymoczko JL, Stryer L, Clarke ND (2002). «I. 5. ДНК, РНК и поток генетической информации: аминокислоты кодируются группами из трех оснований, начиная с фиксированной точки» . Биохимия (5 -е изд.). Нью -Йорк: WH Freeman and Company. Архивировано из оригинала 11 апреля 2006 года.
- ^ Крик Ф. (август 1970 г.). «Центральная догма молекулярной биологии» (PDF) . Природа . 227 (5258): 561–563. Bibcode : 1970natur.227..561c . doi : 10.1038/227561A0 . PMID 4913914 . S2CID 4164029 . Архивировано (PDF) из оригинала 15 февраля 2006 года.
- ^ Alberts et al. (2002), I.3. Белки: форма и структура белков архивировали 1 января 2023 года на машине Wayback
- ^ Alberts et al. (2002), I.3. Белки: белковая функция архивировала 25 апреля 2006 года на машине Wayback
- ^ "Как серп -клетка вызывает болезнь?" Полем Бригам и женская больница: информационный центр серповых клеток и талассемических расстройств. 11 апреля 2002 года. Архивировано с оригинала 23 сентября 2010 года . Получено 23 июля 2007 года .
- ^ Imes DL, Geary LA, Grahn RA, Lyons LA (апрель 2006 г.). «Альбинизм у домашней кошки (Felis Catus) связан с мутацией тирозиназы (TYR)» . Животная генетика . 37 (2): 175–178. doi : 10.1111/j.1365-2052.2005.01409.x . PMC 1464423 . PMID 16573534 .
- ^ "MedlinePlus: фенилкетонурия" . NIH: Национальная библиотека медицины. Архивировано из оригинала 25 июля 2008 года . Получено 15 марта 2008 года .
- ^ Например, Ридли М. (2003). Природа через воспитание: гены, опыт и то, что делает нас людьми . Четвертое поместье. п. 73. ISBN 978-1-84115-745-0 .
- ^ Розенталь Д. (1964). «Четырехлеты Genain: тематическое исследование и теоретический анализ наследственности и окружающей среды при шизофрении». Поведенческая наука . 9 (4): 371. doi : 10.1002/bs.3830090407 .
- ^ Бриванлу А.Х., Дарнелл Дж. (Февраль 2002 г.). «Сигнальная трансдукция и контроль экспрессии генов». Наука . 295 (5556): 813–818. Bibcode : 2002sci ... 295..813b . Citeseerx 10.1.1.485.6042 . doi : 10.1126/science.1066355 . PMID 11823631 . S2CID 14954195 .
- ^ Alberts et al. (2002), II.3. Контроль экспрессии генов - репрессор триптофана - это простой переключатель, который включает и выключает гены в бактериях, архивировав 29 июня 2007 года на машине Wayback
- ^ Jaenisch R, Bird A (март 2003 г.). «Эпигенетическая регуляция экспрессии генов: как геном интегрирует внутренние и экологические сигналы». Природа генетика . 33 (Suppl): 245–254. doi : 10.1038/ng1089 . PMID 12610534 . S2CID 17270515 .
- ^ Chandler VL (февраль 2007 г.). «Paramutation: от кукурузы к мышам» . Клетка . 128 (4): 641–645. doi : 10.1016/j.cell.2007.02.007 . PMID 17320501 . S2CID 6928707 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Спонтанные мутации» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Freisinger E, Grollman AP, Miller H, Kisker C (апрель 2004 г.). «Повреждение (в) толерантность раскрывает понимание верности репликации ДНК» . Embo Journal . 23 (7): 1494–1505. doi : 10.1038/sj.emboj.7600158 . PMC 391067 . PMID 15057282 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Индуцированные мутации» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Кадет J, Вагнер -младший (февраль 2013 г.). «Повреждение основания ДНК реактивными формами кислорода, окислителями и ультрафиолетовым излучением» . Перспективы Cold Spring Harbor в биологии . 5 (2): A012559. doi : 10.1101/cshperspect.a012559 . PMC 3552502 . PMID 23378590 .
- ^ Йена Н.Р. (июль 2012 г.). «Повреждение ДНК реактивными видами: механизмы, мутация и восстановление». Журнал биологических наук . 37 (3): 503–517. doi : 10.1007/s12038-012-9218-2 . PMID 22750987 . S2CID 14837181 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Мутация хромосом I: изменения в структуре хромосом: введение» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Schaechter M (2009). Энциклопедия микробиологии . Академическая пресса. п. 551. ISBN 978-0-12-373944-5 .
- ^ Calver M, Lymbery A, McComb J, Bamford M (2009). Экологическая биология . Издательство Кембриджского университета. п. 118. ISBN 978-0-521-67982-4 .
- ^ Sawyer SA, Parsch J, Zhang Z, Hartl DL (апрель 2007 г.). «Распространенность положительного отбора среди почти нейтральных замен аминокислот у дрозофилы» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (16): 6504–6510. Bibcode : 2007pnas..104.6504S . doi : 10.1073/pnas.0701572104 . PMC 1871816 . PMID 17409186 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Изменение и ее модуляция» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). «Выбор» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Gillespie JH (ноябрь 2001 г.). "Является ли численность вида, имеет отношение к его эволюции?" Полем Эволюция; Международный журнал органической эволюции . 55 (11): 2161–2169. doi : 10.1111/j.0014-3820.2001.tb00732.x . PMID 11794777 . S2CID 221735887 .
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). "Случайные события" . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Дарвин С (1859). На происхождение видов . Лондон: Джон Мюррей. п. 1. ISBN 978-0-8014-1319-3 Полем Архивировано из оригинала 12 декабря 2006 года.
Ранее связанные идеи были признаны в Дарвин С (1861). О происхождении видов (3 -е изд.). Лондон: Джон Мюррей. xiii. ISBN 978-0-8014-1319-3 Полем Архивировано из оригинала 23 февраля 2011 года. - ^ Gavrilets S (октябрь 2003 г.). «Перспектива: модели видообразования: что мы узнали за 40 лет?». Эволюция; Международный журнал органической эволюции . 57 (10): 2197–2215. doi : 10.1554/02-727 . PMID 14628909 . S2CID 198158082 .
- ^ Вольф Йи, Рогозин И.Б., Грисин Н.В., Кунин Э.В. (сентябрь 2002 г.). «Деревья генома и дерево жизни». Тенденции в генетике . 18 (9): 472–479. doi : 10.1016/s0168-9525 (02) 02744-0 . PMID 12175808 .
- ^ «Использование модельных организмов в обучении» . Университет Висконсина: Висконсинские исследования по работе с общественностью. Архивировано из оригинала 13 марта 2008 года . Получено 15 марта 2008 года .
- ^ «NCBI: гены и болезнь» . NIH: Национальный центр биотехнологической информации. Архивировано из оригинала 20 февраля 2007 года . Получено 15 марта 2008 года .
- ^ Смит Г.Д. , Эбрагим С (февраль 2003 г.). « Менделяйская рандомизация»: может ли генетическая эпидемиология способствовать пониманию экологических детерминантов заболевания? ». Международный журнал эпидемиологии . 32 (1): 1–22. doi : 10.1093/ije/dyg070 . PMID 12689998 .
- ^ «Фармакогенетическая информация» . NIH: Национальный институт общих медицинских наук. Архивировано из оригинала 12 мая 2008 года . Получено 15 марта 2008 года .
- ^ Фрэнк С.А. (октябрь 2004 г.). «Генетическая предрасположенность к раку - понимание генетики популяции». Природные обзоры. Генетика . 5 (10): 764–772. doi : 10.1038/nrg1450 . PMID 15510167 . S2CID 6049662 .
- ^ Strachan T, Read AP (1999). Молекулярная генетика человека 2 (второе изд.). John Wiley & Sons Inc. Глава 18: Генетика рака архивировал 26 сентября 2005 года на машине Wayback
- ^ Lodish et al. (2000), Глава 7: 7.1. Клонирование ДНК с помощью плазмидных векторов заархивировано 27 мая 2009 г. на машине Wayback
- ^ Timms JF, Cramer R (декабрь 2008 г.). «Разница гель электрофорез» . Протеомика . 8 (23–24): 4886–4897. doi : 10.1002/pmic.200800298 . ISSN 1615-9853 . PMID 19003860 .
- ^ Кифер К.Л. (июль 2015 г.). «Искусственное клонирование домашних животных» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (29): 8874–8878. Bibcode : 2015pnas..112.8874k . doi : 10.1073/pnas.1501718112 . PMC 4517265 . PMID 26195770 .
- ^ Lodish et al. (2000), Глава 7: 7.7. Полимеразная цепная реакция: альтернатива клонированию
- ^ Chang D, Tram K, Li B, Feng Q, Shen Z, Lee CH, et al. (8 июня 2017 г.). «Обнаружение ДНК -ампликонов полимеразной цепной реакции с использованием лакмусового теста» . Научные отчеты . 7 (3110): 3110. Bibcode : 2017natsr ... 7.3110c . doi : 10.1038/s41598-017-03009-z . PMC 5465217 . PMID 28596600 .
- ^ Garibyan L, Nidhi (март 2013 г.). «Полимеразная цепная реакция» . Журнал расследования дерматологии . 133 (3): 1–4. doi : 10.1038/jid.2013.1 . PMC 4102308 . PMID 23399825 . Получено 27 февраля 2024 года .
- ^ Браун Т.А. (2002). «Раздел 2, глава 6: 6.1. Методология для секвенирования ДНК» . Геномы 2 (2 -е изд.). Оксфорд: Биос. ISBN 978-1-85996-228-2 .
- ^ Браун (2002), Раздел 2, Глава 6: 6.2. Сборка смежной последовательности ДНК архивирована 8 февраля 2007 года на машине Wayback
- ^ Сервис RF (март 2006 г.). «Секвенирование генов. Гонка за геном за 1000 долларов». Наука . 311 (5767): 1544–1546. doi : 10.1126/science.311.5767.1544 . PMID 16543431 . S2CID 23411598 .
- ^ Зал N (май 2007 г.). «Усовершенствованные технологии секвенирования и их более широкое влияние на микробиологию» . Журнал экспериментальной биологии . 210 (Pt 9): 1518–1525. doi : 10.1242/jeb.001370 . PMID 17449817 .
- ^ Церковь GM (январь 2006 г.). «Геномы для всех». Scientific American . 294 (1): 46–54. Bibcode : 2006sciam.294a..46c . doi : 10.1038/Scientificamerican0106-46 . PMID 16468433 . S2CID 28769137 . (требуется подписка)
- ^ Уэйд N (19 марта 2015 г.). «Ученые ищут запрет на метод редактирования человеческого генома» . New York Times . Архивировано из оригинала 19 марта 2015 года . Получено 20 марта 2015 года .
- ^ Поллак A (3 марта 2015 г.). «Мощный новый способ редактирования ДНК» . New York Times . Архивировано с оригинала 26 марта 2015 года . Получено 20 марта 2015 года .
- ^ Балтимор Д., Берг П., Ботчан М., Кэрролл Д., Чаро Р.А., Черч Г. и др. (Апрель 2015). «Биотехнология. Представленный путь вперед для геномной инженерии и модификации генов зародышевой линии» . Наука . 348 (6230): 36–38. Bibcode : 2015sci ... 348 ... 36b . doi : 10.1126/science.aab1028 . PMC 4394183 . PMID 25791083 .
- ^ Lanphier E, Urnov F, Haecker SE, Werner M, Smolenski J (март 2015 г.). «Не редактируйте человеческую линию зародышей» . Природа . 519 (7544): 410–411. Bibcode : 2015natur.519..410L . doi : 10.1038/519410a . PMID 25810189 .
- ^ Kolata G (23 апреля 2015 г.). «Китайские ученые редактируют гены человеческих эмбрионов, выявляя проблемы» . New York Times . Архивировано с оригинала 24 апреля 2015 года . Получено 24 апреля 2015 года .
- ^ Liang P, Xu Y, Zhang X, Ding C, Huang R, Zhang Z, et al. (Май 2015). «Редактирование генов CRISPR/CAS9 у человеческого трипноядерного зигот» . Белок и клетка . 6 (5): 363–372. doi : 10.1007/s13238-015-0153-5 . PMC 4417674 . PMID 25894090 .
Дальнейшее чтение
[ редактировать ]- Альбертс Б., Брей Д., Хопкин К., Джонсон А., Льюис Дж., Рафф М. и др. (2013). Основная клеточная биология, 4 -е издание . Гарлендская наука. ISBN 978-1-317-80627-1 .
- Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart, Eds. (2000). Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- Hartl D, Jones E (2005). Генетика: анализ генов и геномов (6 -е изд.). Джонс и Бартлетт. ISBN 978-0-7637-1511-3 .
- King RC, Mulligan PK, Stansfield WD (2013). Словарь генетики (8 -е изд.). Нью -Йорк: издательство Оксфордского университета. ISBN 978-0-19-976644-4 .
- Lodish H, Berk A, Zipursky LS, Matsudaira P, Baltimore D, Darnell J (2000). Молекулярная клеточная биология (4 -е изд.). Нью -Йорк: научные американские книги. ISBN 978-0-7167-3136-8 .
Внешние ссылки
[ редактировать ] Цитаты, связанные с генетикой в Wikiquote
Цитаты, связанные с генетикой в Wikiquote  Генетика в Wikibooks
Генетика в Wikibooks - Библиотечные ресурсы в вашей библиотеке и в других библиотеках о генетике
- Генетика в наше время в BBC
