Рас ГТФаза
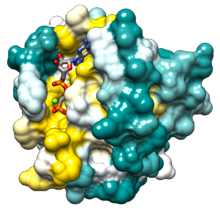 HRas Структура PDB 121p, поверхность окрашена в результате консервации в затравочном состоянии Pfam : золото, наиболее консервативное; темно-голубой, наименее сохранившийся. | |||
| Идентификаторы | |||
|---|---|---|---|
| Символ | Рас | ||
| Пфам | PF00071 | ||
| ИнтерПро | ИПР020849 | ||
| PROSITE | PDOC00017 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 5p21 / SCOPe / СУПФАМ | ||
| CDD | cd04138 | ||
| |||
Ras , от « вируса крысиной аркомы семейство », представляет собой родственных белков , которые экспрессируются во всех линиях и органах животных клеток. Все члены семейства белков Ras принадлежат к классу белков, называемых малой ГТФазой , и участвуют в передаче сигналов внутри клеток ( клеточного трансдукция сигнала ). Ras является прототипом члена суперсемейства белков Ras , которые связаны трехмерной структурой и регулируют разнообразное поведение клеток.
Когда Ras «включается» поступающими сигналами, он впоследствии включает другие белки, которые в конечном итоге активируют гены, участвующие в клеток росте , дифференцировке и выживании . Мутации в генах Ras могут привести к выработке постоянно активированных белков Ras, что может вызвать непреднамеренную и сверхактивную передачу сигналов внутри клетки даже при отсутствии входящих сигналов.
Поскольку эти сигналы приводят к росту и делению клеток, сверхактивная передача сигналов Ras может в конечном итоге привести к раку . [1] Три гена Ras у человека ( HRAS , KRAS и NRAS ) являются наиболее распространенными онкогенами при раке человека; мутации, которые постоянно активируют Ras, обнаруживаются в 20–25% всех опухолей человека и до 90% при определенных типах рака (например, раке поджелудочной железы ). [2] По этой причине ингибиторы Ras изучаются как средство лечения рака и других заболеваний со сверхэкспрессией Ras.
История
[ редактировать ]первые два гена Ras, HRAS и KRAS . Были идентифицированы [3] на основе исследований двух вызывающих рак вирусов, вируса саркомы Харви и вируса саркомы Кирстен, проведенных Эдвардом М. Скольником и его коллегами из Национальных институтов здравоохранения (NIH). [4] Эти вирусы были первоначально обнаружены у крыс в 1960-х годах Дженнифер Харви. [5] и Вернер Х. Кирстен , [6] название Rat соответственно, отсюда и s arcoma . [3] активированные и трансформирующие гены ras человека. обнаружил в раковых клетках человека В 1982 году Джеффри М. Купер из Гарварда [7] , и Стюарт А. Ааронсон из НИЗ [8] Роберт Вайнберг из Массачусетского технологического института, [9] и Майкл Виглер из лаборатории Колд-Спринг-Харбор. [10] Третий ген ras был впоследствии обнаружен исследователями из группы Робина Вайса в Институте исследования рака . [11] [12] и Майкл Виглер из лаборатории Колд-Спринг-Харбор, [13] назван NRAS в честь его первоначальной идентификации в клетках нейробластомы человека.
Три гена ras человека кодируют чрезвычайно похожие белки, состоящие из цепочек из 188–189 аминокислот. Их генные символы — HRAS , NRAS и KRAS , последний из которых производит изоформы K-Ras4A и K-Ras4B в результате альтернативного сплайсинга . [ нужна ссылка ]
Структура
[ редактировать ]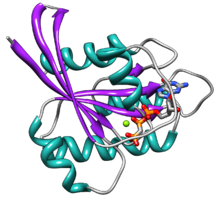
Ras содержит шесть бета-цепей и пять альфа-спиралей . [14] Он состоит из двух доменов: G-домена из 166 аминокислот (около 20 кДа), который связывает гуанозиновые нуклеотиды, и С-концевой области мембранного нацеливания (CAAX-COOH, также известной как CAAX-бокс ), которая модифицирована липидами фарнезилом . трансфераза , RCE1 и ICMT . [ нужна ссылка ]
Домен G содержит пять мотивов G, которые напрямую связывают GDP/GTP.Мотив G1, или P-петля, связывает бета-фосфат GDP и GTP.Мотив G2, также называемый Switch I или SW1, содержит треонин35, который связывает концевой фосфат (γ-фосфат) GTP и двухвалентный ион магния, связанный в активном центре. Мотив G3, также называемый Switch II или SW2, имеет мотив DXXGQ. D представляет собой аспартат57, который специфичен для связывания гуанина по сравнению с аденином, а Q представляет собой глютамин61, важнейший остаток, который активирует каталитическую молекулу воды для гидролиза GTP до GDP.Мотив G4 содержит мотив LVGNKxDL и обеспечивает специфическое взаимодействие с гуанином.Мотив G5 содержит консенсусную последовательность SAK. А — это аланин146, который обеспечивает специфичность к гуанину, а не к аденину.
Два мотива переключения, G2 (SW1) и G3 (SW2), являются основными частями белка, которые перемещаются, когда GTP гидролизуется с образованием GDP. Это конформационное изменение, вызванное двумя мотивами переключения, является тем, что обеспечивает базовую функциональность белка молекулярного переключения. Это связанное с GTP состояние Ras является состоянием «включено», а состояние, связанное с GDP, является состоянием «выключено». Два мотива переключения имеют ряд конформаций при связывании GTP или GDP или при отсутствии нуклеотида (при связывании с SOS1, который высвобождает нуклеотид). [15]
Ras также связывает ион магния , который помогает координировать связывание нуклеотидов.
Функция
[ редактировать ]Белки Ras функционируют как бинарные молекулярные переключатели, которые контролируют внутриклеточные сигнальные сети. Ras-регулируемые сигнальные пути контролируют такие процессы, как актинового цитоскелета целостность , пролиферацию клеток , дифференцировку клеток , адгезию клеток , апоптоз и миграцию клеток . Ras и Ras-родственные белки часто нарушаются при раке, что приводит к усилению инвазии и метастазированию , а также снижению апоптоза.
Ras активирует несколько путей, из которых каскад митоген-активируемых протеинкиназ (MAP) хорошо изучен . Этот каскад передает сигналы ниже по течению и приводит к транскрипции генов, участвующих в росте и делении клеток. [16] Другим Ras-активируемым сигнальным путем является путь PI3K/AKT/mTOR , который стимулирует синтез белка, клеточную миграцию и рост, а также ингибирует апоптоз. [17] [18]
Активация и деактивация
[ редактировать ]Ras представляет собой гуанозин - нуклеотидсвязывающий белок. В частности, это односубъединичная малая ГТФаза , которая по структуре родственна Gα субъединице гетеротримерных G-белков (больших ГТФаз). G-белки функционируют как бинарные сигнальные переключатели с состояниями «включено» и «выключено». В «выключенном» состоянии он связан с нуклеотидом гуанозиндифосфатом (GDP), а во «включенном» состоянии Ras связан с гуанозинтрифосфатом (GTP), который имеет дополнительную фосфатную группу по сравнению с GDP. Этот дополнительный фосфат удерживает две переключающие области в конфигурации «нагруженной пружины» (в частности, Thr-35 и Gly-60). При отпускании области переключения расслабляются, что вызывает конформационные изменения в неактивное состояние. Следовательно, активация и деактивация Ras и других малых G-белков контролируются путем циклического переключения между активной GTP-связанной и неактивной GDP-связанной формами.
Процессу обмена связанного нуклеотида способствуют факторы обмена гуаниновых нуклеотидов (GEF) и белки, активирующие ГТФазу (GAP). Согласно классификации, Ras обладает внутренней активностью ГТФазы , что означает, что белок сам по себе гидролизует связанную молекулу ГТФ с образованием ВВП. Однако этот процесс слишком медленный для эффективного функционирования, и, следовательно, GAP Ras, RasGAP, может связываться и стабилизировать каталитический механизм Ras, поставляя дополнительные каталитические остатки (« аргининовый палец »), так что молекула воды оптимально позиционируется для нуклеофильной реакции. атака на гамма-фосфат ГТФ. Высвобождается неорганический фосфат, и молекула Ras теперь связана с GDP. Поскольку форма, связанная с GDP, «выключена» или «неактивна» для передачи сигналов, белок, активирующий ГТФазу, инактивирует Ras, активируя его активность ГТФазы. Таким образом, GAP ускоряют инактивацию Ras .
ГЭФ катализируют реакцию «толкания и притяжения», которая высвобождает ВВП из Раса. Они встраиваются близко к P-петле и месту связывания катиона магния и ингибируют их взаимодействие с анионом гамма-фосфата . Кислотные (отрицательные) остатки в переключателе II «оттягивают» лизин в P-петле от GDP, что «отталкивает» переключатель I от гуанина. Контакты, удерживающие ВВП, разрываются, и он высвобождается в цитоплазму. Поскольку внутриклеточный ГТФ в избытке относительно ВВП (примерно в 10 раз больше) [16] GTP преимущественно повторно входит в карман связывания нуклеотидов Ras и перезагружает пружину. Таким образом, GEF способствуют активации Ras . [14] Хорошо известные GEF включают Son of Sevenless (Sos) и cdc25 , которые включают домен RasGEF .
Баланс между активностью GEF и GAP определяет статус гуаниновых нуклеотидов Ras, тем самым регулируя активность Ras.
В GTP-связанной конформации Ras обладает высоким сродством к многочисленным эффекторам , которые позволяют ему выполнять свои функции. К ним относится ПИ3К . Другие небольшие ГТФазы могут связывать адаптеры, такие как арфаптин , или системы вторичных мессенджеров, такие как аденилатциклаза . Связывающий домен Ras обнаружен во многих эффекторах и неизменно связывается с одной из областей переключения, поскольку они меняют конформацию между активной и неактивной формами. Однако они также могут связываться с остальной поверхностью белка.
Существуют и другие белки, которые могут изменить активность белков семейства Ras. Одним из примеров является GDI (ингибитор диссоциации ВВП). Они действуют, замедляя обмен ВВП на ГТП, тем самым продлевая неактивное состояние членов семьи Рас. Могут существовать и другие белки, дополняющие этот цикл.
Мембранное крепление
[ редактировать ]Ras прикрепляется к клеточной мембране за счет пренилирования и пальмитоилирования ( HRAS и NRAS ) или сочетания пренилирования и многоосновной последовательности, прилегающей к месту пренилирования ( KRAS ). С-концевой CaaX-бокс Ras сначала фарнезилируется по остатку Cys в цитозоле, что позволяет Ras свободно встраиваться в мембрану эндоплазматического ретикулума и другие клеточные мембраны. Затем трипептид (ааХ) отщепляется от С-конца специфической эндопротеазой, специфичной для пренил-белка, и новый С-конец метилируется метилтрансферазой . На этом обработка KRas завершена. Динамические электростатические взаимодействия между его положительно заряженной основной последовательностью и отрицательными зарядами на внутреннем листке плазматической мембраны объясняют его преобладающую локализацию на поверхности клетки в устойчивом состоянии. NRAS и HRAS подвергаются дальнейшему процессингу на поверхности аппарата Гольджи путем пальмитоилирования одного или двух остатков Cys, соответственно, соседних с CaaX-боксом . Таким образом, белки прочно закрепляются на мембране (липидные рафты) и транспортируются в Плазматическая везикул мембрана секреторного пути . Депальмитоилирование ацилпротеинтиоэстеразами в конечном итоге высвобождает белки из мембраны, позволяя им войти в новый цикл пальмитоилирования и депальмитоилирования. [19] Считается, что этот цикл предотвращает утечку NRAS и HRAS в другие мембраны с течением времени и поддерживает их стационарную локализацию вдоль аппарата Гольджи , секреторного пути , плазматической мембраны и взаимосвязанного пути эндоцитоза .
Члены
[ редактировать ]Клинически наиболее заметными представителями подсемейства Ras являются HRAS , KRAS и NRAS , в основном из-за их участия во многих типах рака. [20]
Однако есть и много других представителей этого подсемейства: [21] ДИРАС1 ; ДИРАС2 ; ДИРАС3 ; ЭРАС ; ДРАГОЦЕННЫЙ КАМЕНЬ ; МРАС ; НКИРАС1 ; НКИРАС2 ; РАЛА ; РАЛБ ; РАП1А ; РАП1Б ; РАП2А ; РАП2Б ; РАП2С ; РАСД1 ; РАСД2 ; РАСЛ10А ; РАСЛ10Б ; РАСЛ11А ; РАСЛ11Б ; РАСЛ12 ; РЕМ1 ; РЕМ2 ; РЕРГ ; РЕРГЛ ; РРАД ; РРАС ; RRAS2
Рас при раке
[ редактировать ]Мутации в семействе протоонкогенов Ras (включая H-Ras, N-Ras и K-Ras) очень распространены и обнаруживаются в 20–30% всех опухолей человека. [20] Разумно предположить, что фармакологический подход, ограничивающий активность Ras, может представлять собой возможный метод ингибирования определенных типов рака. Точечные мутации Ras являются единственной наиболее распространенной аномалией протоонкогенов человека. [22] Ингибитор Ras транс-фарнезилтиосалициловая кислота (ФТС, салирасиб ) проявляет глубокие антионкогенные эффекты во многих линиях раковых клеток. [23] [24]
Неподходящая активация
[ редактировать ]Было показано, что несоответствующая активация гена играет ключевую роль в неправильной передаче сигнала, пролиферации и злокачественной трансформации. [16]
Мутации в ряде различных генов, а также в самом РАС могут иметь такой эффект. Онкогены, такие как p210BCR-ABL или рецептор роста erbB, находятся выше Ras, поэтому, если они конститутивно активированы, их сигналы будут передаваться через Ras. [ нужна ссылка ]
Ген -супрессор опухоли NF1 кодирует Ras-GAP – его мутация при нейрофиброматозе будет означать, что Ras с меньшей вероятностью будет инактивирован. Ras также может усиливаться, хотя в опухолях это происходит лишь изредка.
Наконец, онкогены Ras могут быть активированы точечными мутациями, так что реакция GTPase больше не может стимулироваться GAP – это увеличивает период полураспада активных мутантов Ras-GTP. [25]
Конститутивно активный Ras
[ редактировать ]Конститутивно активный Ras ( Ras Д ) — тот, который содержит мутации, которые предотвращают гидролиз GTP, тем самым удерживая Ras в постоянно включенном состоянии.
Наиболее распространенные мутации обнаруживаются в остатке G12 в P-петле и каталитическом остатке Q61.
- Мутация глицина на валин в остатке 12 делает домен GTPase Ras нечувствительным к инактивации GAP и, таким образом, остается во «включенном состоянии». Ras требует GAP для инактивации, поскольку он сам по себе является относительно плохим катализатором, в отличие от других белков, содержащих G-домен, таких как альфа-субъединица гетеротримерных G-белков.
- Остаток 61 [26] отвечает за стабилизацию переходного состояния гидролиза ГТФ. Поскольку ферментативный катализ обычно достигается за счет снижения энергетического барьера между субстратом и продуктом, мутация Q61 в K (глутамин в лизин) обязательно снижает скорость внутреннего гидролиза Ras GTP до физиологически бессмысленного уровня.
См. также «доминантно-негативные» мутанты, такие как S17N и D119N.
Лечение рака, нацеленное на Ras
[ редактировать ]Реовирус был признан потенциальным средством лечения рака, когда исследования показали, что он хорошо размножается в определенных линиях раковых клеток. Он специфически реплицируется в клетках, которые имеют активированный путь Ras (клеточный сигнальный путь, который участвует в росте и дифференцировке клеток). [27] Реовирус реплицируется и в конечном итоге убивает Ras-активированные опухолевые клетки, а когда происходит гибель клеток, дочерние вирусные частицы могут свободно инфицировать окружающие раковые клетки. Считается, что этот цикл заражения, репликации и гибели клеток повторяется до тех пор, пока все опухолевые клетки, несущие активированный путь Ras, не будут уничтожены. [ нужна ссылка ]
Еще одним вирусом, лизисом опухоли, который специфически нацелен на опухолевые клетки с активированным путем Ras, является агент на основе вируса простого герпеса II типа (HSV-2), получивший обозначение FusOn-H2. [28] Активирующие мутации белка Ras и вышестоящих элементов белка Ras могут играть роль в более чем двух третях всех видов рака человека, включая большинство метастатических заболеваний. Реолизин , форма реовируса, и FusOn-H2 в настоящее время проходят клинические испытания или разрабатываются для лечения различных видов рака. [29] основанный на антимутированной миРНК K-RAS (G12D), называемый siG12D LODER (NCT01188785, NCT01676259). Кроме того, в настоящее время проходят клинические испытания метод лечения местно-распространенного рака поджелудочной железы, [30]
В моделях глиобластомы на мышах уровни SHP2 были повышены в раковых клетках головного мозга. Ингибирование SHP2 , в свою очередь, ингибирует дефосфорилирование Ras. Это уменьшило размеры опухоли и сопутствующее увеличение выживаемости. [31] [32]
Другие стратегии пытались манипулировать регуляцией вышеупомянутой локализации Ras. Ингибиторы фарнезилтрансферазы были разработаны для остановки фарнезилирования Ras и, следовательно, для ослабления его сродства к мембранам. [2] Другие ингибиторы воздействуют на цикл пальмитоилирования Ras путем ингибирования депальмитоилирования ацилпротеинтиоэстеразами , что потенциально приводит к дестабилизации цикла Ras. [33]
Новая стратегия поиска ингибиторов для мутированных молекул Ras была описана в . [34] Мутации Ras в 12-м положении остатка ингибируют связывание регуляторной молекулы GAP с мутированным Ras, вызывая неконтролируемый рост клеток. Новая стратегия предлагает найти небольшие молекулы клея, которые прикрепляют мутировавший Ras к GAP, запрещая неконтролируемый рост клеток и восстанавливая нормальную функцию. Для этой цели была разработана теоретическая конформация Ras-GAP с зазором в несколько Å между молекулами и проведен высокопроизводительный стыковочный процесс in silico для поиска клеящих агентов. В качестве доказательства концепции были описаны две новые молекулы с удовлетворительной биологической активностью.
У других видов
[ редактировать ]В большинстве типов клеток большинства видов большая часть Ras относится к типу GDP. Это справедливо для Xenopus ооцитов и мыши фибробластов . [35]
Ксенопус левис
[ редактировать ]Как упоминалось выше, большая часть X. ооцитов Ras является конъюгатом GDP. млекопитающих Ras индуцирует мейоз в ооцитах X. laevis почти наверняка путем усиления мейоза, индуцированного инсулином , но не индуцируемого прогестероном . Синтез белка , похоже, не является частью этого этапа. Инъекция увеличивает синтез диацилглицерина из фосфатидилхолина . Некоторым эффектам мейоза противодействует rap1 , (и Ras модифицированный для неправильной стыковки). И rap1 , и модифицированный Ras являются соантагонистами GAP p120Ras на этом пути. [35]
Дрозофила меланогастер
[ редактировать ]Экспрессируется во всех тканях Drosophila melanogaster , но преимущественно в нервных клетках. Сверхэкспрессия в некоторой степени смертельна и во время развития приводит к аномалиям глаз и крыльев. (Это соответствует - и может быть причиной - подобных аномалий, вызванных мутированными рецепторными тирозинкиназами .) D. Гены , вызывающие ras у млекопитающих, вызывают аномалии. [35]
Аплизия
[ редактировать ]Наибольшая экспрессия у Aplysia spp. находится в нервных клетках. [35]
Ценорабдитис элегантный
[ редактировать ]Ген у C. elegans — let 60 . В этой модели, по-видимому, также играют роль в образовании рецепторной тирозинкиназы. Сверхэкспрессия приводит к развитию нескольких влагалищ из-за ее участия в нормальном развитии этого региона; сверхэкспрессия в эффекторных сайтах приводит к летальному исходу. [35]
Диктиостелиум дискоидеум
[ редактировать ]Незаменим для Dictyostelium discoideum . Об этом свидетельствуют тяжелые нарушения развития при недостаточной экспрессии ras и значительные нарушения различных видов жизнедеятельности при их искусственной экспрессии, такие как: повышенная концентрация инозитолфосфатов ; вероятное снижение связывания цАМФ с хемотаксиса рецепторами ; и это, вероятно, причина цГМФ нарушения синтеза . На активность аденилатциклазы не влияет ras . [35]
Ссылки
[ редактировать ]- ^ Гудселл Д.С. (1999). «Молекулярная перспектива: онкоген ras» . Онколог . 4 (3): 263–4. doi : 10.1634/теонколог.4-3-263 . ПМИД 10394594 .
- ^ Jump up to: а б Вниз J (январь 2003 г.). «Нацеливание на сигнальные пути RAS в терапии рака». Обзоры природы. Рак . 3 (1): 11–22. дои : 10.1038/nrc969 . ПМИД 12509763 . S2CID 43074411 .
- ^ Jump up to: а б Малумбрес М., Барбацид М. (июнь 2003 г.). «Онкогены РАН: первые 30 лет». Обзоры природы. Рак . 3 (6): 459–65. дои : 10.1038/nrc1097 . ПМИД 12778136 . S2CID 27928171 .
- ^ Чанг Э.Х., Гонда М.А., Эллис Р.В., Сколник Э.М., Лоуи Д.Р. (август 1982 г.). «Геном человека содержит четыре гена, гомологичных трансформирующим генам вирусов мышиной саркомы Харви и Кирстен» . Труды Национальной академии наук Соединенных Штатов Америки . 79 (16): 4848–52. Бибкод : 1982PNAS...79.4848C . дои : 10.1073/pnas.79.16.4848 . ПМЦ 346782 . ПМИД 6289320 .
- ^ Харви Джей-Джей (декабрь 1964 г.). «Неопознанный вирус, вызывающий быстрое образование опухолей у мышей». Природа . 204 (4963): 1104–5. Бибкод : 1964Natur.204.1104H . дои : 10.1038/2041104b0 . ПМИД 14243400 . S2CID 4144311 .
- ^ Кирстен В.Х., Шауф В., Маккой Дж. (1970). «Свойства вируса мышиной саркомы». Гематологическая библиотека . Международный симпозиум по сравнительным исследованиям лейкемии. 36 (36): 246–9. дои : 10.1159/000391714 . ISBN 978-3-8055-1160-5 . ПМИД 5538357 .
- ^ Купер GM (август 1982 г.). «Клеточные трансформирующие гены». Наука . 217 (4562): 801–6. Бибкод : 1982Sci...217..801C . дои : 10.1126/science.6285471 . ПМИД 6285471 . S2CID 5807661 .
- ^ Сантос Э., Троник С.Р., Ааронсон С.А., Пульчиани С., Барбацид М. (июль 1982 г.). «Онкоген карциномы мочевого пузыря человека T24 представляет собой активированную форму нормального человеческого гомолога генов, трансформирующих BALB и Harvey-MSV». Природа . 298 (5872): 343–7. Бибкод : 1982Natur.298..343S . дои : 10.1038/298343a0 . ПМИД 6283384 . S2CID 37033023 .
- ^ Парада Л.Ф., Табин С.Дж., Ши С., Вайнберг Р.А. (июнь 1982 г.). «Онкоген карциномы мочевого пузыря EJ человека является гомологом гена ras вируса саркомы Харви». Природа . 297 (5866): 474–8. Бибкод : 1982Natur.297..474P . дои : 10.1038/297474a0 . ПМИД 6283357 . S2CID 4338225 .
- ^ Тапаровски Э., Суард Ю., Фазано О., Симидзу К., Гольдфарб М., Виглер М. (декабрь 1982 г.). «Активация гена, трансформирующего карциному Т24, связана с изменением одной аминокислоты». Природа . 300 (5894): 762–5. Бибкод : 1982Natur.300..762T . дои : 10.1038/300762a0 . ПМИД 7177195 . S2CID 34179063 .
- ^ Маршалл CJ, зал А, Вайс РА (сентябрь 1982 г.). «Трансформирующий ген, присутствующий в клеточных линиях саркомы человека». Природа . 299 (5879): 171–3. Бибкод : 1982Natur.299..171M . дои : 10.1038/299171a0 . ПМИД 6287287 . S2CID 4342747 .
- ^ Зал А, Маршалл С.Дж., Сперр Н.К., Вайс Р.А. (1983). «Идентификация трансформирующего гена в двух клеточных линиях саркомы человека как нового члена семейства генов ras, расположенного на хромосоме 1». Природа . 303 (5916): 396–400. Бибкод : 1983Natur.303..396H . дои : 10.1038/303396a0 . ПМИД 6304521 . S2CID 4372475 .
- ^ Симидзу К., Гольдфарб М., Перучо М., Виглер М. (январь 1983 г.). «Выделение и предварительная характеристика трансформирующего гена клеточной линии нейробластомы человека» . Труды Национальной академии наук Соединенных Штатов Америки . 80 (2): 383–7. Бибкод : 1983PNAS...80..383S . дои : 10.1073/pnas.80.2.383 . ПМЦ 393381 . ПМИД 6300838 .
- ^ Jump up to: а б Феттер И.Р., Виттингхофер А. (ноябрь 2001 г.). «Переключатель связывания гуаниновых нуклеотидов в трех измерениях». Наука . 294 (5545): 1299–304. Бибкод : 2001Sci...294.1299V . дои : 10.1126/science.1062023 . ПМИД 11701921 . S2CID 6636339 .
- ^ Паркер М.И., Мейер Дж.Э., Големис Э.А., Данбрек Р.Л. (5 июля 2022 г.). «Очерчивание конформационного ландшафта РАН» . Исследования рака . 82 (13): 2485–2498. дои : 10.1158/0008-5472.CAN-22-0804 . ПМЦ 9256797 . ПМИД 35536216 . Проверено 11 октября 2022 г.
- ^ Jump up to: а б с Лодиш Х., Берк А., Зипурски С.Л., Мацудайра П., Балтимор Д., Дарнелл Дж. (2000). «Глава 25, Рак» . Молекулярно-клеточная биология (4-е изд.). Сан-Франциско: WH Freeman. ISBN 0-7167-3706-Х .
- ^ Пал Д.С., Банерджи Т., Лин Ю., де Трогофф Ф., Борлейс Дж., Иглесиас П.А., Девреотес П.Н. (июль 2023 г.). «Активация отдельных нижестоящих узлов в сети факторов роста управляет миграцией иммунных клеток» . Развивающая клетка . 58 (13): 1170–1188.e7. дои : 10.1016/j.devcel.2023.04.019 . ISSN 1534-5807 . ПМЦ 10524337 . ПМИД 37220748 .
- ^ Линь Ю, Пал Д.С., Банерджи П., Банерджи Т., Цинь Дж., Дэн Ю., Борлейс Дж., Иглесиас П.А., Девреотес П.Н. (01.07.2024). «Подавление Ras усиливает поляризацию и миграцию клеток, обусловленную сократимостью заднего актомиозина» . Природная клеточная биология : 1–15. дои : 10.1038/s41556-024-01453-4 . ISSN 1476-4679 .
- ^ Рокс О, Пейкер А, Бастиенс П.И. (август 2006 г.). «Пространственно-временная сегрегация сигналов Раса: один корабль, три якоря, много гаваней». Современное мнение в области клеточной биологии . 18 (4): 351–7. дои : 10.1016/j.ceb.2006.06.007 . ПМИД 16781855 .
- ^ Jump up to: а б Бос Дж.Л. (сентябрь 1989 г.). «Онкогены ras при раке человека: обзор». Исследования рака . 49 (17): 4682–9. ПМИД 2547513 .
- ^ Веннерберг К., Россман К.Л., Der CJ (март 2005 г.). «Краткий обзор суперсемьи Рас». Журнал клеточной науки . 118 (Часть 5): 843–6. дои : 10.1242/jcs.01660 . ПМИД 15731001 . S2CID 40171018 .
- ^ Роббинс и Котран (2010). Патологические основы болезней 8-е изд . п. 282.
- ^ Ротблат Б., Эрлих М., Хаклай Р., Клоог Ю. (2008). «Ингибитор Ras фарнезилтиосалициловая кислота (Салирасиб) нарушает пространственно-временную локализацию активного Ras: потенциальное лечение рака». Малые ГТФазы в болезнях, Часть Б. Методы энзимологии. Том. 439. стр. 467–89. дои : 10.1016/S0076-6879(07)00432-6 . ISBN 978-0-12-374311-4 . ПМИД 18374183 .
- ^ Блюм Р., Джейкоб-Хирш Дж., Амариглио Н., Речави Г., Клоог Ю. (февраль 2005 г.). «Ингибирование Ras при глиобластоме подавляет индуцируемый гипоксией фактор-1альфа, вызывая остановку гликолиза и гибель клеток» . Исследования рака . 65 (3): 999–1006. дои : 10.1158/0008-5472.999.65.3 . ПМИД 15705901 . S2CID 21694752 .
- ^ Reuter CW, Морган М.А., Бергманн Л. (сентябрь 2000 г.). «Нацеливание на сигнальный путь Ras: рациональное, основанное на механизмах лечение гематологических злокачественных новообразований?». Кровь . 96 (5): 1655–69. дои : 10.1182/blood.V96.5.1655 . ПМИД 10961860 .
- ^ «Омим - гомолог вирусного онкогена нейробластомы Рас; Нрас» . Архивировано из оригинала 06 марта 2019 г. Проверено 10 сентября 2017 г.
- ^ Лал Р., Харрис Д., Постель-Винай С., де Боно Дж. (октябрь 2009 г.). «Реовирус: обоснование и обновленная информация о клинических испытаниях». Современное мнение о молекулярной терапии . 11 (5): 532–9. ПМИД 19806501 .
- ^ Фу Икс, Тао Л., Цай Р., Пригге Дж., Чжан Икс (май 2006 г.). «Мутантный вирус простого герпеса 2 типа с удаленным протеинкиназным доменом гена ICP10 является мощным онколитическим вирусом» . Молекулярная терапия . 13 (5): 882–90. дои : 10.1016/j.ymthe.2006.02.007 . ПМИД 16569513 .
- ^ Тируккумаран С., Моррис Д.Г. (2009). «Онколитическая вирусная терапия с использованием реовируса». Генная терапия рака . Методы молекулярной биологии. Том. 542. стр. 607–34. дои : 10.1007/978-1-59745-561-9_31 . ISBN 978-1-934115-85-5 . ПМИД 19565924 .
- ^ «Клинические испытания.gov» .
- ^ Бунда С., Баррелл К., Хейр П., Цзэн Л., Аламсахебпур А., Кано Ю., Раф Б., Чжан З.И., Заде Г., Ох М. (ноябрь 2015 г.). «Ингибирование SHP2-опосредованного дефосфорилирования Ras подавляет онкогенез» . Природные коммуникации . 6 : 8859. Бибкод : 2015NatCo...6.8859B . дои : 10.1038/ncomms9859 . ПМЦ 4674766 . ПМИД 26617336 .
- ^ Тауб Б. (3 декабря 2015 г.). «Ученые нашли способ дезактивировать наиболее распространенный белок, вызывающий рак» . IFLНаука . Проверено 20 февраля 2016 г.
- ^ Чавда Б., Арнотт Дж.А., Плани С.Л. (сентябрь 2014 г.). «Нацеленное пальмитоилирование белка: селективные ингибиторы и последствия при заболеваниях». Мнение экспертов об открытии лекарств . 9 (9): 1005–19. дои : 10.1517/17460441.2014.933802 . ПМИД 24967607 . S2CID 207494086 .
- ^ Ранджелович И., Ньири К., Коппани Г., Бараньи М., Товари Дж., Кидьос А., Тимар Дж., Вертесси Б.Г., Гролмуш В. (февраль 2024 г.). «Приклеивание GAP к мутантам RAS: новый подход к старой проблеме разработки лекарств от рака» . Международный журнал молекулярных наук . 25 (5): 2572. arXiv : 2312.05791 . дои : 10.3390/ijms25052572 . ПМЦ 10932042 . ПМИД 38473821 .
- ^ Jump up to: а б с д и ж Боллаг Дж., Маккормик Ф. (1991). «Регуляторы и эффекторы белков ras». Ежегодный обзор клеточной биологии . 7 (1). Годовые обзоры : 601–32. дои : 10.1146/annurev.cb.07.110191.003125 . ПМИД 1667084 .
Дальнейшее чтение
[ редактировать ]- Агравал АГ, Сомани Р.Р. (июнь 2009 г.). «Ингибитор фарнезилтрансферазы как противораковое средство». Мини-обзоры по медицинской химии . 9 (6): 638–52. дои : 10.2174/138955709788452702 . ПМИД 19519490 .
Внешние ссылки
[ редактировать ]- «Результаты опухолей головного мозга дают надежду на новую стратегию, говорит Канадское онкологическое общество» на ncic.cancer.ca
- «Новое лечение рака получает поддержку NCI» на arstechnica.com
- ras+Proteins Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
- ras+Genes в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- дрозофилы Онкоген Ras на 85D - The Interactive Fly
- «Анимация активации ras EGFR»
- «Rascore: инструмент для анализа белковых структур RAS»