Липополисахарид

Липополисахарид , теперь более широко известный как эндотоксин , [ 1 ] является коллективным термином для компонентов самой внешней мембраны клеточной оболочки грамотрицательных бактерий, таких как кишечная палочка и сальмонелла [ 2 ] с общей структурной архитектурой. Липополисахариды ( LPS ) представляют собой большие молекулы, состоящие из трех частей: полисахарид наружного ядра , называемый о- антигеном внутреннего ядра , олигосахарид и липид (из которого в значительной степени получена токсичность), все ковалентно связаны. В текущей терминологии термин эндотоксин часто используется синонимом LPS, хотя существует несколько эндотоксинов (в исходном смысле токсинов , которые находятся внутри бактериальной клетки, которые высвобождаются, когда клетка распадается), которые не связаны с LP, например Так называемые дельта-эндотоксины белки, продуцируемые Bacillus thuringiensis . [ 3 ]
Липополисахариды могут оказывать существенное влияние на здоровье человека, в первую очередь посредством взаимодействия с иммунной системой. LPS является мощным активатором иммунной системы и является пирогеном (агент, который вызывает лихорадку). [ 4 ] В тяжелых случаях LPS может вызвать оживленный ответ хоста и несколько типов острой органа. [ 5 ] что может привести к септическому шоку . [ 6 ] На более низких уровнях и в течение более длительного периода времени есть доказательства, которые LPs могут играть важную и вредную роль в аутоиммунитете , ожирении , депрессии и клеточном старении . [ 7 ] [ 8 ] [ 9 ] [ 10 ]
Открытие
[ редактировать ]Токсическая активность LPS была впервые обнаружена и названа эндотоксином Ричардом Фридрихом Йоханнесом Пфайффером . Он различал экзотоксины , токсины, которые высвобождаются бактериями в окружающую среду, и эндотоксинами, которые являются токсинами »в« бактериальной клетке и высвобождаются только после разрушения внешней мембраны бактерий. [ 11 ] Последующая работа показала, что высвобождение ЛП из грамотрицательных микробов не обязательно требует разрушения бактериальной клеточной стенки, а скорее, LPS секретируется как часть нормальной физиологической активности переноса мембранных пузырьков в форме везикул бактериальной внешней мембраны (OMVS (OMVS (OMVS ) , что также может содержать другие факторы вирулентности и белки. [ 12 ] [ 2 ]
Функции в бактериях
[ редактировать ]LPS является основным компонентом внешней клеточной мембраны грамотрицательных бактерий , внося в значительной степени структурную целостность бактерий и защита мембраны от определенных видов химической атаки. LPS является наиболее распространенным антигеном на клеточной поверхности большинства грамотрицательных бактерий, внося до 80% внешней мембраны E. coli и сальмонеллы . [ 2 ] LPS увеличивает отрицательный заряд клеточной мембраны и помогает стабилизировать общую мембранную структуру. Это имеет решающее значение для многих грамотрицательных бактерий, которые умирают, если гены, кодирующие его, мутированы или удалены. Тем не менее, кажется, что LPS является несущественным, по крайней мере, в некоторых грамотрицательных бактериях, таких как Neisseria meningitidis , Moraxella Catarrhalis и Acinetobacter baumannii . [ 13 ] Он также участвовал в непатогенных аспектах бактериальной экологии, включая поверхностную адгезию, чувствительность бактериофагов и взаимодействие с хищниками, такими как амебы . LPS также необходим для функционирования Omptins , класса бактериальной протеазы. [ 14 ]
Композиция
[ редактировать ]
LPS являются амфипатическими и состоит из трех частей: антиген O (или O -полисахарид), который является гидрофильным, основным олигосахаридом (также гидрофильным) и липидом A , гидрофобным доменом.
O-Antigen
[ редактировать ]Повторяющийся гликана полимер , содержащийся в LPS, называется O антиген , полисахарид или O-боковой цепь бактерий. А антиген прикреплен к основному олигосахариду и включает в себя самый внешний домен молекулы LPS. Структура и состав цепи o сильно варьируются от штамма к деформации, определяя серологическую специфичность родительского бактериального штамма; [ 15 ] Существует более 160 различных структур антигенов, продуцируемых различными штаммами E. coli . [ 16 ] Наличие или отсутствие цепей O определяют, считается ли LPS «грубым» или «гладким». Полноразмерные O-цепи будут делать LPS гладкими, тогда как отсутствие или уменьшение O-цепей сделают LPS грубыми. [ 17 ] Бактерии с грубыми LPS обычно имеют более проницательные клеточные мембраны для гидрофобных антибиотиков, поскольку грубая LPS является более гидрофобным . [ 18 ] O Антиген подвергается воздействию на самой внешней поверхности бактериальной клетки и, как следствие, является мишенью для распознавания антителами хозяина .
Основной
[ редактировать ]Основной домен всегда содержит олигосахаридный компонент, который непосредственно присоединяется к липиду А и обычно содержит сахары , такие как гептоза и 3-дезокси-D-манно-окт-2-мулосоновая кислота (также известная как KDO, кето-дезоксиоктулозонат). [ 19 ] Олигосахарид основного олигосахарида в своей структуре и составе является менее изменчивым, причем данная структура ядра является общей для больших групп бактерий. [ 15 ] Ядра LPS многих бактерий также содержат негледратные компоненты, такие как фосфат, аминокислоты и заместители этаноламина.
Липид а
[ редактировать ]Липид А, в нормальных обстоятельствах, является фосфорилированным глюкозамина дисахаридом , украшенным несколькими жирными кислотами . Эти гидрофобные цепи жирной кислоты привязывают LPS в бактериальную мембрану, а остальная часть LPS проецирует с клеточной поверхности. Домен липидов является наиболее биологическим и ответственным за большую часть токсичности грамотрицательных бактерий . Когда бактериальные клетки лизируются иммунной системой , фрагменты мембраны, содержащих липид А, могут высвобождаться в кровообращение, вызывая лихорадку, диарею и возможный смертельный эндотоксический септический шок (форма септического шока ). Липид А. Форт является очень консервативным компонентом ЛПС. [ 20 ] Однако липидная структура варьируется среди бактериальных видов. Липид А Структура в значительной степени определяет степень и характер общей иммунной активации хозяина. [ 21 ]
Липолигосахариды
[ редактировать ]«Грубая форма» LPS имеет более низкую молекулярную массу из -за отсутствия полисахарида O. Вместо этого-короткий олигосахарид: эта форма известна как липолигосахарид (LOS) и является гликолипидом, обнаруженным во внешней мембране некоторых типов грамотрицательных бактерий , таких как Neisseria spp. и Haemophilus spp. [ 7 ] [ 22 ] LOS играет центральную роль в поддержании целостности и функциональности внешней мембраны грамма -негативной огибающей клеток. LOS играет важную роль в патогенезе определенных бактериальных инфекций, потому что они способны действовать в качестве иммуностимуляторов и иммуномодуляторов. [ 7 ] Кроме того, молекулы LOS ответственны за способность некоторых бактериальных штаммов демонстрировать молекулярную мимикурию и антигенное разнообразие , помогая в уклонке иммунной защиты хозяина и, таким образом, способствуя вирулентности этих бактериальных штаммов . В случае Neisseria meningitidis , липидная часть молекулы имеет симметричную структуру, а внутреннее ядро состоит из 3-дезокси-D-манно-2-октулосоновой кислоты (KDO) и гластской (HEP). Олигосахаридная цепь Внешнего ядра варьируется в зависимости от бактериального штамма . [ 7 ] [ 22 ]
Детоксикация LPS
[ редактировать ]Высококонсервативный фермент хозяина, называемый ацилоксиацил гидролазой (AOAH), может детоксифицировать LPS, когда он входит или продуцируется в тканях животных. Это также может преобразовать LPS в кишечник в ингибитор LPS. Нейтрофилы, макрофаги и дендритные клетки продуцируют эту липазу, которая инактивирует LP, удаляя две вторичные ацильные цепь из липидов A, чтобы получить тетраацил LPS. Если мышам дают LPS парентерально, у тех, у кого не хватает AOAH, развиваются высокие титры неспецифических антител, развивают длительную гепатомегалию и испытывают длительную толерантность к эндотоксинам. Инактивация LPS может потребоваться для животных для восстановления гомеостаза после парентерального воздействия LPS. [ 23 ] Хотя у мышей есть много других механизмов для ингибирования передачи сигналов LPS, ни один из них не может предотвратить эти изменения у животных, которым не хватает AOAH.
Dephosphorylation of LPS by intestinal alkaline phosphatase can reduce the severity of Salmonella tryphimurium and Clostridioides difficile infection restoring normal gut microbiota.[24] Alkaline phosphatase prevents intestinal inflammation (and "leaky gut") from bacteria by dephosphorylating the Lipid A portion of LPS.[25][26][27]
Biosynthesis and transport
[edit]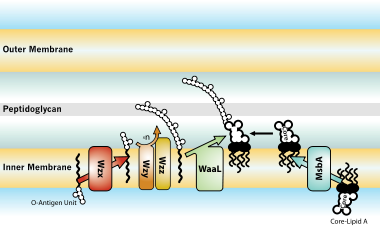
The entire process of making LPS starts with a molecule called lipid A-Kdo2, which is first created on the surface of the bacterial cell's inner membrane. Then, additional sugars are added to this molecule on the inner membrane before it's moved to the space between the inner and outer membranes (periplasmic space) with the help of a protein called MsbA. The O-antigen, another part of LPS, is made by special enzyme complexes on the inner membrane. It is then moved to the outer membrane through three different systems: one is Wzy-dependent, another relies on ABC transporters, and the third involves a synthase-dependent process.[30]
Ultimately, LPS is transported to the outer membrane by a membrane-to-membrane bridge of lipolysaccharide transport (Lpt) proteins.[29][31] This transporter is a potential antibiotic target.[32][33]
Biological effects on hosts infected with Gram-negative bacteria
[edit]LPS storage in the body
[edit]The human body carries endogenous stores on LPS.[34] The epithelial surfaces are colonized by a complex microbial flora (including gram-negative bacteria), which outnumber human cells by a factor of 10 to 1. Gram-negative bacterial will shed endotoxins. This host-microbial interaction is a symbiotic relationship which plays a critical role in systemic immunologic homeostasis. When this is disrupted, it can lead to disease such as endotoxemia and endotoxic septic shock.
Immune response
[edit]LPS acts as the prototypical endotoxin because it binds the CD14/TLR4/MD2 receptor complex in many cell types, but especially in monocytes, dendritic cells, macrophages and B cells, which promotes the secretion of pro-inflammatory cytokines, nitric oxide, and eicosanoids.[35] Bruce Beutler was awarded a portion of the 2011 Nobel Prize in Physiology or Medicine for his work demonstrating that TLR4 is the LPS receptor.[36][37]
As part of the cellular stress response, superoxide is one of the major reactive oxygen species induced by LPS in various cell types that express TLR (toll-like receptor).[38] LPS is also an exogenous pyrogen (fever-inducing substance).[4]
LPS function has been under experimental research for several years due to its role in activating many transcription factors. LPS also produces many types of mediators involved in septic shock. Of mammals, humans are much more sensitive to LPS than other primates,[39] and other animals as well (e.g., mice). A dose of 1 μg/kg induces shock in humans, but mice will tolerate a dose up to a thousand times higher.[40] This may relate to differences in the level of circulating natural antibodies between the two species.[41][42] It may also be linked to multiple immune tactics against pathogens, and part of a multi-faceted anti-microbial strategy that has been informed by human behavioral changes over our species' evolution (e.g., meat eating, agricultural practices, and smoking).[39] Said et al. showed that LPS causes an IL-10-dependent inhibition of CD4 T-cell expansion and function by up-regulating PD-1 levels on monocytes which leads to IL-10 production by monocytes after binding of PD-1 by PD-L1.[43]
Endotoxins are in large part responsible for the dramatic clinical manifestations of infections with pathogenic Gram-negative bacteria, such as Neisseria meningitidis, the pathogens that causes meningococcal disease, including meningococcemia, Waterhouse–Friderichsen syndrome, and meningitis.
Portions of the LPS from several bacterial strains have been shown to be chemically similar to human host cell surface molecules; the ability of some bacteria to present molecules on their surface which are chemically identical or similar to the surface molecules of some types of host cells is termed molecular mimicry.[44] For example, in Neisseria meningitidis L2,3,5,7,9, the terminal tetrasaccharide portion of the oligosaccharide (lacto-N-neotetraose) is the same tetrasaccharide as that found in paragloboside, a precursor for ABH glycolipid antigens found on human erythrocytes.[7] In another example, the terminal trisaccharide portion (lactotriaose) of the oligosaccharide from pathogenic Neisseria spp. LOS is also found in lactoneoseries glycosphingolipids from human cells.[7] Most meningococci from groups B and C, as well as gonococci, have been shown to have this trisaccharide as part of their LOS structure.[7] The presence of these human cell surface 'mimics' may, in addition to acting as a 'camouflage' from the immune system, play a role in the abolishment of immune tolerance when infecting hosts with certain human leukocyte antigen (HLA) genotypes, such as HLA-B35.[7]
LPS can be sensed directly by hematopoietic stem cells (HSCs) through the bonding with TLR4, causing them to proliferate in reaction to a systemic infection. This response activate the TLR4-TRIF-ROS-p38 signaling within the HSCs and through a sustained TLR4 activation can cause a proliferative stress, leading to impair their competitive repopulating ability.[45] Infection in mice using S. typhimurium showed similar results, validating the experimental model also in vivo.
Effect of variability on immune response
[edit]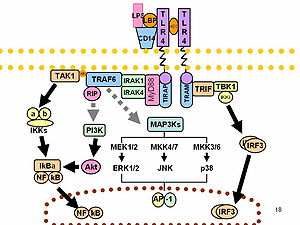
O-antigens (the outer carbohydrates) are the most variable portion of the LPS molecule, imparting antigenic specificity. In contrast, lipid A is the most conserved part. However, lipid A composition also may vary (e.g., in number and nature of acyl chains even within or between genera). Some of these variations may impart antagonistic properties to these LPS. For example, diphosphoryl lipid A of Rhodobacter sphaeroides (RsDPLA) is a potent antagonist of LPS in human cells, but is an agonist in hamster and equine cells.[46]
It has been speculated that conical lipid A (e.g., from E. coli) is more agonistic, while less conical lipid A like that of Porphyromonas gingivalis may activate a different signal (TLR2 instead of TLR4), and completely cylindrical lipid A like that of Rhodobacter sphaeroides is antagonistic to TLRs.[47][48] In general, LPS gene clusters are highly variable between different strains, subspecies, species of bacterial pathogens of plants and animals.[49][50]
Normal human blood serum contains anti-LOS antibodies that are bactericidal and patients that have infections caused by serotypically distinct strains possess anti-LOS antibodies that differ in their specificity compared with normal serum.[51] These differences in humoral immune response to different LOS types can be attributed to the structure of the LOS molecule, primarily within the structure of the oligosaccharide portion of the LOS molecule.[51] In Neisseria gonorrhoeae it has been demonstrated that the antigenicity of LOS molecules can change during an infection due to the ability of these bacteria to synthesize more than one type of LOS,[51] a characteristic known as phase variation. Additionally, Neisseria gonorrhoeae, as well as Neisseria meningitidis and Haemophilus influenzae,[7] are capable of further modifying their LOS in vitro, for example through sialylation (modification with sialic acid residues), and as a result are able to increase their resistance to complement-mediated killing [51] or even down-regulate complement activation[7] or evade the effects of bactericidal antibodies.[7] Sialylation may also contribute to hindered neutrophil attachment and phagocytosis by immune system cells as well as a reduced oxidative burst.[7] Haemophilus somnus, a pathogen of cattle, has also been shown to display LOS phase variation, a characteristic which may help in the evasion of bovine host immune defenses.[52] Taken together, these observations suggest that variations in bacterial surface molecules such as LOS can help the pathogen evade both the humoral (antibody and complement-mediated) and the cell-mediated (killing by neutrophils, for example) host immune defenses.
Non-canonical pathways of LPS recognition
[edit]Recently, it was shown that in addition to TLR4 mediated pathways, certain members of the family of the transient receptor potential ion channels recognize LPS.[53] LPS-mediated activation of TRPA1 was shown in mice[54] and Drosophila melanogaster flies.[55] At higher concentrations, LPS activates other members of the sensory TRP channel family as well, such as TRPV1, TRPM3 and to some extent TRPM8.[56] LPS is recognized by TRPV4 on epithelial cells. TRPV4 activation by LPS was necessary and sufficient to induce nitric oxide production with a bactericidal effect.[57]
Testing
[edit]Lipopolysaccharide is a significant factor that makes bacteria harmful, and it helps categorize them into different groups based on their structure and function. This makes LPS a useful marker for telling apart various Gram-negative bacteria. Swiftly identifying and understanding the types of pathogens involved is crucial for promptly managing and treating infections. Since LPS is the main trigger for the immune response in our cells, it acts as an early signal of an acute infection. Therefore, LPS testing is more specific and meaningful than many other serological tests.[58]
The current methods for testing LPS are quite sensitive, but many of them struggle to differentiate between different LPS groups. Additionally, the nature of LPS, which has both water-attracting and water-repelling properties (amphiphilic), makes it challenging to develop sensitive and user-friendly tests.[58]
The typical detection methods rely on identifying the lipid A part of LPS because Lipid A is very similar among different bacterial species and serotypes. LPS testing techniques fall into six categories, and they often overlap: in vivo tests, in vitro tests, modified immunoassays, biological assays, and chemical assays.[58]
Endotoxin Activity Assay
[edit]Because the LPS is very difficult to measure in whole blood and because most LPS is bound to proteins and complement, the Endotoxin Activity Assay (EAA™) was developed and cleared by the US FDA in 2003. EAA is a rapid in vitro chemiluminescent immunodiagnostic test. It utilizes a specific monoclonal antibody to measure the endotoxin activity in EDTA whole blood specimens. This assay uses the biological response of the neutrophils in a patient’s blood to an immunological complex of endotoxin and exogenous antibody – the chemiluminescent reaction formed creates an emission of light. The amount of chemiluminescence is proportional to the logarithmic concentration of LPS in the sample and is a measure of the endotoxin activity in the blood.[59] The assay reacts specifically with the Lipid A moiety of LPS of Gram-negative bacteria and does not cross-react with cell wall constituents of Gram-positive bacteria and other microorganisms.
Pathophysiology
[edit]LPS is a powerful toxin that, when in the body, triggers inflammation by binding to cell receptors. Excessive LPS in the blood, endotoxemia, may cause a highly lethal form of sepsis known as endotoxic septic shock.[5] This condition includes symptoms that fall along a continuum of pathophysiologic states, starting with a systemic inflammatory response syndrome (SIRS) and ending in multiorgan dysfunction syndrome (MODS) before death. Early symptoms include rapid heart rate, quick breathing, temperature changes, and blood clotting issues, resulting in blood vessels widening and reduced blood volume, leading to cellular dysfunction.[58]
Recent research indicates that even small LPS exposure is associated with autoimmune diseases and allergies. High levels of LPS in the blood can lead to metabolic syndrome, increasing the risk of conditions like diabetes, heart disease, and liver problems.[58]
LPS also plays a crucial role in symptoms caused by infections from harmful bacteria, including severe conditions like Waterhouse-Friderichsen syndrome, meningococcemia, and meningitis. Certain bacteria can adapt their LPS to cause long-lasting infections in the respiratory and digestive systems.[58]
Recent studies have shown that LPS disrupts cell membrane lipids, affecting cholesterol and metabolism, potentially leading to high cholesterol, abnormal blood lipid levels, and non-alcoholic fatty liver disease. In some cases, LPS can interfere with toxin clearance, which may be linked to neurological issues.[58]
Health effects
[edit]In general the health effects of LPS are due to its abilities as a potent activator and modulator of the immune system, especially its inducement of inflammation. LPS is directly cytoxic and is highly immunostimulatory – as host immune cells recognize LPS, complement are strongly activated. Complement activation and a rising anti-inflammatory response can lead to immune cell dysfunction, immunosuppression, widespread coagulopathy, serious tissue damage and can progress to multi-system organ failure and death.[39]
Endotoxemia
[edit]The presence of endotoxins in the blood is called endotoxemia. High level of endotoxemia can lead to septic shock,[60] or more specifically endotoxic septic shock,[5] while lower concentration of endotoxins in the bloodstream is called metabolic endotoxemia.[61] Endotoxemia is associated with obesity, diet,[62] cardiovascular diseases,[62] and diabetes,[61] while also host genetics might have an effect.[63]
Moreover, endotoxemia of intestinal origin, especially, at the host-pathogen interface, is considered to be an important factor in the development of alcoholic hepatitis,[64] which is likely to develop on the basis of the small bowel bacterial overgrowth syndrome and an increased intestinal permeability.[65]
Lipid A may cause uncontrolled activation of mammalian immune systems with production of inflammatory mediators that may lead to endotoxic septic shock.[22][5] This inflammatory reaction is primarily mediated by Toll-like receptor 4 which is responsible for immune system cell activation.[22] Damage to the endothelial layer of blood vessels caused by these inflammatory mediators can lead to capillary leak syndrome, dilation of blood vessels and a decrease in cardiac function and can further worsen shock.[66] LPS is also a potent activator of complemen.[66] Uncontrolled complement activation may trigger destructive endothelial damage leading to disseminated intravascular coagulation (DIC), or atypical hemolytic uremic syndrome (aHUS) with injury to various organs such as including kidneys and lungs.[67] The skin can show the effects of vascular damage often coupled with depletion of coagulation factors in the form of petechiae, purpura and ecchymoses. The limbs can also be affected, sometimes with devastating consequences such as the development of gangrene, requiring subsequent amputation.[66] Loss of function of the adrenal glands can cause adrenal insufficiency and additional hemorrhage into the adrenals causes Waterhouse-Friderichsen syndrome, both of which can be life-threatening.
It has also been reported that gonococcal LOS can cause damage to human fallopian tubes.[51]
Treatment of Endotoxemia
[edit]Toraymyxin is a widely used extracorporeal endotoxin removal therapy through direct hemoadsorption (also referred to as hemoperfusion). It is a polystyrene-derived cartridge with molecules of polymyxin B (PMX-B) covalently bound to mesh fibers contained within it. Polymyxins are cyclic cationic polypeptide antibiotics derived from Bacillus polymyxa with an effective antimicrobial activity against Gram-negative bacteria, but their intravenous clinical use has been limited due to their nephrotoxicity and neurotoxicity side effects.[68] The extracorporeal use of the Toraymyxin cartridge allows PMX-B to bind lipid A with a very stable interaction with its hydrophobic residues thereby neutralizing endotoxins as the blood is filtered through the extracorporeal circuit inside the cartridge, thus reversing endotoxemia and avoiding its toxic systemic effects.[69]
Auto-immune disease
[edit]The molecular mimicry of some LOS molecules is thought to cause autoimmune-based host responses, such as flareups of multiple sclerosis.[7][44] Other examples of bacterial mimicry of host structures via LOS are found with the bacteria Helicobacter pylori and Campylobacter jejuni, organisms which cause gastrointestinal disease in humans, and Haemophilus ducreyi which causes chancroid. Certain C. jejuni LPS serotypes (attributed to certain tetra- and pentasaccharide moieties of the core oligosaccharide) have also been implicated with Guillain–Barré syndrome and a variant of Guillain–Barré called Miller-Fisher syndrome.[7]
Link to obesity
[edit]Epidemiological studies have shown that increased endotoxin load, which can be a result of increased populations of endotoxin-producing bacteria in the intestinal tract, is associated with certain obesity-related patient groups.[8][70][71] Other studies have shown that purified endotoxin from Escherichia coli can induce obesity and insulin-resistance when injected into germ-free mouse models.[72] A more recent study has uncovered a potentially contributing role for Enterobacter cloacae B29 toward obesity and insulin resistance in a human patient.[73] The presumed mechanism for the association of endotoxin with obesity is that endotoxin induces an inflammation-mediated pathway accounting for the observed obesity and insulin resistance.[72] Bacterial genera associated with endotoxin-related obesity effects include Escherichia and Enterobacter.
Depression
[edit]There is experimental and observational evidence that LPS might play a role in depression. Administration of LPS in mice can lead to depressive symptoms, and there seem to be elevated levels of LPS in some people with depression. Inflammation may sometimes play a role in the development of depression, and LPS is pro-inflammatory.[9]
Cellular senescence
[edit]Inflammation induced by LPS can induce cellular senescence, as has been shown for the lung epithelial cells and microglial cells (the latter leading to neurodegeneration).[10]
Role as contaminant in biotechnology and research
[edit]Lipopolysaccharides are frequent contaminants in plasmid DNA prepared from bacteria or proteins expressed from bacteria, and must be removed from the DNA or protein to avoid contaminating experiments and to avoid toxicity of products manufactured using industrial fermentation.[74]
Ovalbumin is frequently contaminated with endotoxins. Ovalbumin is one of the extensively studied proteins in animal models and also an established model allergen for airway hyper-responsiveness (AHR). Commercially available ovalbumin that is contaminated with LPS can falsify research results, as it does not accurately reflect the effect of the protein antigen on animal physiology.[75]
In pharmaceutical production, it is necessary to remove all traces of endotoxin from drug product containers, as even small amounts of endotoxin will cause illness in humans. A depyrogenation oven is used for this purpose. Temperatures in excess of 300 °C are required to fully break down LPS.[76]
The standard assay for detecting presence of endotoxin is the Limulus Amebocyte Lysate (LAL) assay, utilizing blood from the Horseshoe crab (Limulus polyphemus).[77] Very low levels of LPS can cause coagulation of the limulus lysate due to a powerful amplification through an enzymatic cascade. However, due to the dwindling population of horseshoe crabs, and the fact that there are factors that interfere with the LAL assay, efforts have been made to develop alternative assays, with the most promising ones being ELISA tests using a recombinant version of a protein in the LAL assay, Factor C.[78]
See also
[edit]- Bioaerosol
- Depyrogenation
- Host-pathogen interface
- Mucopolysaccharide
- Nesfatin-1
- Schwartzman reaction
- AOAH
References
[edit]- ^ Rietschel ET, Kirikae T, Schade FU, Mamat U, Schmidt G, Loppnow H, et al. (February 1994). "Bacterial endotoxin: molecular relationships of structure to activity and function". FASEB Journal. 8 (2): 217–225. doi:10.1096/fasebj.8.2.8119492. PMID 8119492. S2CID 28156137.
- ^ Jump up to: a b c Avila-Calderón ED, Ruiz-Palma MD, Aguilera-Arreola MG, Velázquez-Guadarrama N, Ruiz EA, Gomez-Lunar Z, et al. (2021). "Outer Membrane Vesicles of Gram-Negative Bacteria: An Outlook on Biogenesis". Frontiers in Microbiology. 12: 557902. doi:10.3389/fmicb.2021.557902. PMC 7969528. PMID 33746909.
- ^ Höfte H, de Greve H, Seurinck J, Jansens S, Mahillon J, Ampe C, et al. (December 1986). "Structural and functional analysis of a cloned delta endotoxin of Bacillus thuringiensis berliner 1715". European Journal of Biochemistry. 161 (2): 273–280. doi:10.1111/j.1432-1033.1986.tb10443.x. PMID 3023091.
- ^ Jump up to: a b Roth J, Blatteis CM (October 2014). "Mechanisms of fever production and lysis: lessons from experimental LPS fever". Comprehensive Physiology. 4 (4): 1563–1604. doi:10.1002/cphy.c130033. ISBN 978-0-470-65071-4. PMID 25428854.
- ^ Jump up to: a b c d Kellum JA, Ronco C (October 2023). "The role of endotoxin in septic shock". Crit Care. 27 (1): 400. doi:10.1186/s13054-023-04690-5. PMC 10585761. PMID 37858258.
- ^ Dellinger RP, Levy MM, Rhodes A, Annane D, Gerlach H, Opal SM, et al. (February 2013). "Surviving sepsis campaign: international guidelines for management of severe sepsis and septic shock: 2012". Critical Care Medicine. 41 (2): 580–637. doi:10.1097/CCM.0b013e31827e83af. PMID 23353941. S2CID 34855187.
- ^ Jump up to: a b c d e f g h i j k l m n Moran AP, Prendergast MM, Appelmelk BJ (December 1996). "Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease". FEMS Immunology and Medical Microbiology. 16 (2): 105–115. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.
- ^ Jump up to: a b Lasselin J, Schedlowski M, Karshikoff B, Engler H, Lekander M, Konsman JP (August 2020). "Comparison of bacterial lipopolysaccharide-induced sickness behavior in rodents and humans: Relevance for symptoms of anxiety and depression". Neuroscience and Biobehavioral Reviews. 115: 15–24. doi:10.1016/j.neubiorev.2020.05.001. PMID 32433924. S2CID 218665128.
- ^ Jump up to: a b Wei W, Ji S (December 2018). "Cellular senescence: Molecular mechanisms and pathogenicity". Journal of Cellular Physiology. 233 (12): 9121–9135. doi:10.1002/jcp.26956. PMID 30078211. S2CID 51924586.
- ^ Parija SC (1 January 2009). Textbook of Microbiology & Immunology. India: Elsevier. ISBN 978-8131221631.
- ^ Kulp A, Kuehn MJ (2010). "Biological functions and biogenesis of secreted bacterial outer membrane vesicles". Annual Review of Microbiology. 64: 163–184. doi:10.1146/annurev.micro.091208.073413. PMC 3525469. PMID 20825345.
- ^ Zhang G, Meredith TC, Kahne D (December 2013). "On the essentiality of lipopolysaccharide to Gram-negative bacteria". Current Opinion in Microbiology. 16 (6): 779–785. doi:10.1016/j.mib.2013.09.007. PMC 3974409. PMID 24148302.
- ^ Kukkonen M, Korhonen TK (July 2004). "The omptin family of enterobacterial surface proteases/adhesins: from housekeeping in Escherichia coli to systemic spread of Yersinia pestis". International Journal of Medical Microbiology. 294 (1): 7–14. doi:10.1016/j.ijmm.2004.01.003. PMID 15293449.
- ^ Jump up to: a b Galanos C, Freudenberg MA (1993). "Bacterial endotoxins: biological properties and mechanisms of action". Mediators Inflamm. 2 (7): S11-16. doi:10.1155/S0962935193000687. PMC 2365449. PMID 18475562.
- ^ Raetz CR, Whitfield C (2002). "Lipopolysaccharide endotoxins". Annual Review of Biochemistry. 71: 635–700. doi:10.1146/annurev.biochem.71.110601.135414. PMC 2569852. PMID 12045108.
- ^ Rittig MG, Kaufmann A, Robins A, Shaw B, Sprenger H, Gemsa D, et al. (December 2003). "Smooth and rough lipopolysaccharide phenotypes of Brucella induce different intracellular trafficking and cytokine/chemokine release in human monocytes". Journal of Leukocyte Biology. 74 (6): 1045–1055. doi:10.1189/jlb.0103015. PMID 12960272.
- ^ Tsujimoto H, Gotoh N, Nishino T (December 1999). "Diffusion of macrolide antibiotics through the outer membrane of Moraxella catarrhalis". Journal of Infection and Chemotherapy. 5 (4): 196–200. doi:10.1007/s101560050034. PMID 11810516. S2CID 2742306.
- ^ Hershberger C, Binkley SB (April 1968). "Chemistry and metabolism of 3-deoxy-D-mannooctulosonic acid. I. Stereochemical determination". The Journal of Biological Chemistry. 243 (7): 1578–1584. doi:10.1016/S0021-9258(18)93581-7. PMID 4296687.
- ^ Tzeng YL, Datta A, Kolli VK, Carlson RW, Stephens DS (May 2002). "Endotoxin of Neisseria meningitidis composed only of intact lipid A: inactivation of the meningococcal 3-deoxy-D-manno-octulosonic acid transferase". Journal of Bacteriology. 184 (9): 2379–2388. doi:10.1128/JB.184.9.2379-2388.2002. PMC 134985. PMID 11948150.
- ^ Khan MM, Ernst O, Sun J, Fraser ID, Ernst RK, Goodlett DR, Nita-Lazar A (August 2018). "Mass Spectrometry-based Structural Analysis and Systems Immunoproteomics Strategies for Deciphering the Host Response to Endotoxin". Journal of Molecular Biology. 430 (17): 2641–2660. doi:10.1016/j.jmb.2018.06.032. PMID 29949751. S2CID 49481716.
- ^ Jump up to: a b c d Kilár A, Dörnyei Á, Kocsis B (2013). "Structural characterization of bacterial lipopolysaccharides with mass spectrometry and on- and off-line separation techniques". Mass Spectrometry Reviews. 32 (2): 90–117. Bibcode:2013MSRv...32...90K. doi:10.1002/mas.21352. PMID 23165926.
- ^ Munford R, Lu M, Varley AW (2009). Chapter 2 Kill the Bacteria…and Also Their Messengers?. Advances in Immunology. Vol. 103. pp. 29–48. doi:10.1016/S0065-2776(09)03002-8. ISBN 9780123748324. PMC 2812913. PMID 19755182.
- ^ Bilski J, Mazur-Bialy A, Wojcik D, Zahradnik-Bilska J, Brzozowski B, Magierowski M, et al. (2017). "The Role of Intestinal Alkaline Phosphatase in Inflammatory Disorders of Gastrointestinal Tract". Mediators of Inflammation. 2017: 9074601. doi:10.1155/2017/9074601. PMC 5339520. PMID 28316376.
- ^ Bates JM, Akerlund J, Mittge E, Guillemin K (December 2007). "Intestinal alkaline phosphatase detoxifies lipopolysaccharide and prevents inflammation in zebrafish in response to the gut microbiota". Cell Host & Microbe. 2 (6): 371–382. doi:10.1016/j.chom.2007.10.010. PMC 2730374. PMID 18078689.
- ^ Alam SN, Yammine H, Moaven O, Ahmed R, Moss AK, Biswas B, et al. (April 2014). "Intestinal alkaline phosphatase prevents antibiotic-induced susceptibility to enteric pathogens". Annals of Surgery. 259 (4): 715–722. doi:10.1097/sla.0b013e31828fae14. PMC 3855644. PMID 23598380.
- ^ Lallès JP (February 2014). "Intestinal alkaline phosphatase: novel functions and protective effects". Nutrition Reviews. 72 (2): 82–94. doi:10.1111/nure.12082. PMID 24506153.
- ^ Wang X, Quinn PJ (April 2010). "Lipopolysaccharide: Biosynthetic pathway and structure modification". Progress in Lipid Research. 49 (2): 97–107. doi:10.1016/j.plipres.2009.06.002. PMID 19815028.
- ^ Jump up to: a b Ruiz N, Kahne D, Silhavy TJ (September 2009). "Transport of lipopolysaccharide across the cell envelope: the long road of discovery". Nature Reviews. Microbiology. 7 (9): 677–683. doi:10.1038/nrmicro2184. PMC 2790178. PMID 19633680.
- ^ Romano KP, Hung DT (March 2023). "Targeting LPS biosynthesis and transport in gram-negative bacteria in the era of multi-drug resistance". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1870 (3): 119407. doi:10.1016/j.bbamcr.2022.119407. PMC 9922520. PMID 36543281.
- ^ Sherman DJ, Xie R, Taylor RJ, George AH, Okuda S, Foster PJ, et al. (February 2018). "Lipopolysaccharide is transported to the cell surface by a membrane-to-membrane protein bridge". Science. 359 (6377): 798–801. Bibcode:2018Sci...359..798S. doi:10.1126/science.aar1886. PMC 5858563. PMID 29449493.
- ^ Pahil KS, Gilman MS, Baidin V, Clairfeuille T, Mattei P, Bieniossek C, et al. (January 2024). "A new antibiotic traps lipopolysaccharide in its intermembrane transporter". Nature. 625 (7995): 572–577. Bibcode:2024Natur.625..572P. doi:10.1038/s41586-023-06799-7. PMC 10794137. PMID 38172635.
- ^ Zampaloni C, Mattei P, Bleicher K, Winther L, Thäte C, Bucher C, et al. (January 2024). "A novel antibiotic class targeting the lipopolysaccharide transporter". Nature. 625 (7995): 566–571. Bibcode:2024Natur.625..566Z. doi:10.1038/s41586-023-06873-0. PMC 10794144. PMID 38172634.
- ^ Marshall JC (2005). "Lipopolysaccharide: an endotoxin or an exogenous hormone?". Clin Infect Dis. 41 (S7): S470–80. doi:10.1086/432000. PMID 16237650.
- ^ Abbas A (2006). Basic Immunology. Elsevier. ISBN 978-1-4160-2974-8.
- ^ Poltorak A, He X, Smirnova I, Liu MY, Van Huffel C, Du X, et al. (December 1998). "Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene". Science. 282 (5396): 2085–2088. Bibcode:1998Sci...282.2085P. doi:10.1126/science.282.5396.2085. PMID 9851930.
- ^ "The 2011 Nobel Prize in Physiology or Medicine - Press Release". www.nobelprize.org. Archived from the original on 23 March 2018. Retrieved 28 April 2018.
- ^ Li Y, Deng SL, Lian ZX, Yu K (June 2019). "Roles of Toll-Like Receptors in Nitroxidative Stress in Mammals". Cells. 8 (6): 576. doi:10.3390/cells8060576. PMC 6627996. PMID 31212769.
- ^ Jump up to: a b c Brinkworth JF, Valizadegan N (2021). "Sepsis and the evolution of human increased sensitivity to lipopolysaccharide". Evol Anthropol. 30 (2): 141–157. doi:10.1002/evan.21887. PMID 33689211.
- ^ Warren HS, Fitting C, Hoff E, Adib-Conquy M, Beasley-Topliffe L, Tesini B, et al. (Январь 2010). «Устойчивость к бактериальной инфекции: разница между видами может быть связана с белками в сыворотке» . Журнал инфекционных заболеваний . 201 (2): 223–232. doi : 10.1086/649557 . PMC 2798011 . PMID 20001600 .
- ^ Рейд Р.Р., Продоус А.П., Хан В., Хсу Т., Розен Ф.С., Кэрролл М.К. (июль 1997 г.). «Эндотоксин-шок у мышей с дефицитом антител: раскрытие роли природного антитела и комплемента в клиренсе липополисахарида» . Журнал иммунологии . 159 (2): 970–975. doi : 10.4049/jimmunol.159.2.970 . PMID 9218618 .
- ^ Boes M, Prodeus AP, Schmidt T, Carroll MC, Chen J (декабрь 1998 г.). «Критическая роль естественного иммуноглобулина М в немедленной защите от системной бактериальной инфекции» . Журнал экспериментальной медицины . 188 (12): 2381–2386. doi : 10.1084/jem.188.12.2381 . PMC 2212438 . PMID 9858525 .
- ^ Сказал EA, Dupuy FP, Trautmann L, Zhang Y, Shi Y, El-Far M, et al. (Апрель 2010). «Программированная производство интерлейкина-10, индуцированное DEATH-1 с помощью моноцитов, нарушает активацию CD4+ Т-клеток во время ВИЧ-инфекции» . Природная медицина . 16 (4): 452–459. doi : 10.1038/nm.2106 . PMC 4229134 . PMID 20208540 .
- ^ Jump up to: а беременный Chastain EM, Miller SD (январь 2012 г.). «Молекулярная мимика как индуцирующая триггер для аутоиммунного демиелинизирующего заболевания ЦНС» . Иммунологические обзоры . 245 (1): 227–238. doi : 10.1111/j.1600-065x.2011.01076.x . PMC 3586283 . PMID 22168423 .
- ^ Takizawa H, Fritch K, Kovtonyuk LV, Saito Y, Yakkala C, Jacobs K, et al. (Август 2017). «Индуцированная патогеном TLR4-триф врожденная иммунная передача сигналов в гематопоэтических стволовых клетках способствует пролиферации, но снижает конкурентную подготовку» . Клеточная стволовая клетка . 21 (2): 225–240.e5. doi : 10.1016/j.stem.2017.06.013 . PMID 28736216 .
- ^ Lohmann KL, Vandenplas ML, Barton MH, Bryant CE, Moore JN (2007). «Комплекс лошадей TLR4/MD-2 опосредует распознавание липополисахарида из Rhodobacter sphaeroides в качестве агониста». Журнал исследований эндотоксинов . 13 (4): 235–242. doi : 10.1177/0968051907083193 . PMID 17956942 . S2CID 36784237 .
- ^ Netea MG, Van Deuren M, Kullberg BJ, Cavaillon JM, Van Der Meer JW (март 2002 г.). «Определяет ли форма липидов взаимодействие LP с рецепторами, похожими на Toll?». Тенденции в иммунологии . 23 (3): 135–139. doi : 10.1016/s1471-4906 (01) 02169-x . PMID 11864841 .
- ^ Сейдель У, Оикава М., Фукасе К., Кусумото С., Бранденбург К (май 2000). «Внутренняя конформация липида A отвечает за агонистическую и антагонистическую активность». Европейский журнал биохимии . 267 (10): 3032–3039. doi : 10.1046/j.1432-1033.2000.01326.x . PMID 10806403 .
- ^ Ривз П.П., Ван Л. (2002). «Геномная организация LPS-специфических локусов». В Hacker J, Kaper JB (ред.). Острова патогенности и эволюция патогенных микробов . Текущие темы в области микробиологии и иммунологии. Тол. 2. С. 109–35. doi : 10.1007/978-3-642-56031-6_7 . ISBN 978-3-540-42682-0 Полем PMID 12014174 . 264
- ^ Patil PB, Sonti RV (октябрь 2004 г.). «Изменение, наводящее на мысль о горизонтальном переносе генов в липополисахаридном (LPS) биосинтетическом локусе в Xanthomonas oryzae pv. Oryzae, бактериальный патоген листьев риса» . BMC Микробиология . 4 : 40. DOI : 10.1186/1471-2180-4-40 . PMC 524487 . PMID 15473911 .
- ^ Jump up to: а беременный в дюймовый и Yamasaki R, Kerwood DE, Schneider H, Quinn KP, Griffiss JM, Mandrell Re (декабрь 1994 г.). «Структура липоолгосахарида, продуцируемая Neisseria gonorrhoeae, штамм 15253, изолированной от пациента с диссеминированной инфекцией. Свидетельство о новом пути гликозилирования гонококкового липолигосахарида» . Журнал биологической химии . 269 (48): 30345–30351. doi : 10.1016/s0021-9258 (18) 43819-7 . PMID 7982947 .
- ^ Говард MD, Cox AD, Weiser JN, Schurig GG, Inzana TJ (декабрь 2000 г.). «Антигеновое разнообразие Haemophilus somnus lipooligosacharichride: ориентированная на фаза доступность фосфорилхолинового эпитопа» . Журнал клинической микробиологии . 38 (12): 4412–4419. doi : 10.1128/jcm.38.12.4412-4419.2000 . PMC 87614 . PMID 11101573 .
- ^ Boonen B, Alpizar YA, Meseguer VM, Talavera K (август 2018 г.). «TRP каналы как датчики бактериальных эндотоксинов» . Токсины . 10 (8): 326. doi : 10.3390/toxins10080326 . PMC 6115757 . PMID 30103489 .
- ^ Meseguer V, Alpizar YA, Luis E, Tajada S, Denlinger B, Fajardo O, et al. (20 января 2014 г.). «Каналы TRPA1 опосредуют острое нейрогенное воспаление и боль, продуцируемое бактериальными эндотоксинами» . Природная связь . 5 : 3125. Bibcode : 2014natco ... 5.3125m . doi : 10.1038/ncomms4125 . PMC 3905718 . PMID 24445575 .
- ^ Soldano, Alpizar YA, Boonen B, Franco L, López-Requena, Liu G, et et 1. (Июнь 2016 г.). «Бого, которое избегает бактериальных липопополисахаридов через активацию TRPA1 у дрозофилы » Элиф 5 Doi : 10.7554/ elife.1 4907694PMC PMID 27296646
- ^ Boonen B, Alpizar YA, Sanchez A, López-Requena A, Voets T, Talavera K (июль 2018 г.). «Дифференциальное влияние липополисахарида на сенсорные каналы TRP мыши». Клеточный кальций . 73 : 72–81. doi : 10.1016/j.ceca.2018.04.004 . PMID 29689522 . S2CID 13681499 .
- ^ Alpizar YA, Boonen B, Sanchez A, Jung C, López-Requena A, Naert R, et al. (Октябрь 2017). «Активация TRPV4 запускает защитные реакции на бактериальные липополисахариды в эпителиальных клетках дыхательных путей» . Природная связь . 8 (1): 1059. Bibcode : 2017natco ... 8.1059a . doi : 10.1038/s41467-017-01201-3 . PMC 5651912 . PMID 29057902 .
- ^ Jump up to: а беременный в дюймовый и фон глин Page MJ, Kell DB, Pretorius E (2022). «Роль липополисахарид-индуцированной передачи сигналов клеток при хроническом воспалении» . Хронический стресс . 6 : 24705470221076390. DOI : 10.1177/24705470221076390 . PMC 8829728 . PMID 35155966 .
- ^ Romaschin AD, Harris DM, Riberio MB, Paice J, Foster DM, Walker PM, Marshall JC (1998). «Быстрый анализ эндотоксина в цельной крови с использованием аутологичной нейтрофильной зависимой хемилюминесценции». J Immunol Methods . 212 (2): 169–185. doi : 10.1016/s0022-1759 (98) 00003-9 . PMID 9672205 .
- ^ Opal SM (2010). «Эндотоксины и другие триггеры сепсиса». Эндотоксемия и эндотоксин шок . Вклад в нефрологию. Тол. 167. С. 14–24. doi : 10.1159/000315915 . ISBN 978-3-8055-9484-4 Полем PMID 20519895 .
- ^ Jump up to: а беременный Gomes JM, Costa JA, Alfenas RC (март 2017 г.). «Метаболическая эндотоксемия и сахарный диабет: систематический обзор». Метаболизм . 68 : 133–144. doi : 10.1016/j.metabol.2016.12.009 . PMID 28183445 .
- ^ Jump up to: а беременный Kallio Ka, Hätönen Ka, Lehto M, Salomaa V, Männistö S, Pussinen PJ (апрель 2015 г.). «Эндотокемия, питание и кардиометаболические расстройства». Acta Diabetologica . 52 (2): 395-404. Doi : 10.1007/s00592-014-0662-3 . PMID 25326898 . S2CID 24020127 .
- ^ Leskelä J, Toppila I, Härma Mon, Palviainen T, Salminen A, Sandholm N, et al. (Ноябрь 2021 г.). «Генетический профиль эндотоксемии выявляет связь с тромбоэмболией и инсультом» . Журнал Американской кардиологической ассоциации . 10 (21): E022482. Doi : 10.1161/jaha.121.022482 . PMC 8751832 . PMID 34668383 .
- ^ Ceccanti M, Attili A, Balducci G, Attilia F, Giacomelli S, Rotondo C, et al. (Octaber 2006). «Острый алкогольный гепатит». Журнал клинической гастроэнтерологии . 40 (9): 833–841. Doi : 10.1097/01.mcg.0000225570.04773.5d . PMID 17016141 .
- ^ Parlesak A, Schäfer C, Schütz T, Bode JC, Bode C (май 2000). «Повышенная проницаемость кишечника для макромолекул и эндотоксемии у пациентов с хроническим злоупотреблением алкоголем на разных стадиях вызванного алкоголем заболевания печени». Журнал гепатологии . 32 (5): 742–747. doi : 10.1016/s0168-8278 (00) 80242-1 . PMID 10845660 .
- ^ Jump up to: а беременный в Стивенс Д.С., Гринвуд Б., Брандцаг П (июнь 2007 г.). «Эпидемический менингит, менингококемия и Neisseria meningitidis». Лансет . 369 (9580): 2196–2210. doi : 10.1016/s0140-6736 (07) 61016-2 . PMID 17604802 . S2CID 16951072 .
- ^ Kellum JA, Formeck CL, Kernan KF, Gomez H, Carcillo JA (2022). «Подтипы и имитируют сепсис». Клиники Crit Care . 38 (2): 195–211. doi : 10.1016/j.ccc.2021.11.013 . PMID 35369943 .
- ^ Li J, Nation RL, Turnidge JD, Milne RW, Coulthard K, Rayner CR, Paterson DL (2006). «Колистин: восстановление антибиотика для устойчивых к грамотрицательным бактериальным инфекциям». Lancet Infect Dis . 6 (9): 589–601. doi : 10.1016/s1473-3099 (06) 70580-1 . PMID 16931410 .
- ^ Virzi GM, Mattiotti M, De Cal M, Ronco C, Zanella M, Derosa S (2022). «Эндотоксин при сепсисе: методы обнаружения LPS и использование методов OMIC» . Диагностика . 13 (1): 79. doi : 10.3390/Diagnostics13010079 . PMC 9818564 . PMID 36611371 .
- ^ Леппер П.М., Шуман С., Три) К., Раше Ф.М., Шустер Т., Фрэнк Х и др. (Июль 2007 г.). «Ассоциация липополисахарид-связывающих белок и ишемической болезни артерий у мужчин». Журнал Американского колледжа кардиологии . 50 (1): 25–31. doi : 10.1016/j.jacc.2007.02.070 . PMID 17601541 . S2CID 12136094 .
- ^ Ruiz AG, Casafont F, Crespo J, Cayón A, Mayorga M, Estebanez A, et al. (Октябрь 2007 г.). «Липополисахарид-связывающие белковые уровни плазмы и экспрессия гена TNF-альфа печени у пациентов с ожирением: свидетельство потенциальной роли эндотоксина в патогенезе неалкогольного стеатогепатита». Хирургия ожирения . 17 (10): 1374–1380. doi : 10.1007/s11695-007-9243-7 . PMID 18000721 . S2CID 44494003 .
- ^ Jump up to: а беременный Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. (Июль 2007 г.). «Метаболическая эндотоксемия инициирует ожирение и резистентность к инсулину» . Диабет . 56 (7): 1761–1772. doi : 10.2337/db06-1491 . PMID 17456850 .
- ^ Fei N, Zhao L (апрель 2013 г.). «Оппортунистический патоген, выделенный из кишечника ожирением, вызывает ожирение у мышей без зародышей» . Журнал ISME . 7 (4): 880–884. Bibcode : 2013ismej ... 7..880f . doi : 10.1038/ismej.2012.153 . PMC 3603399 . PMID 23235292 .
- ^ Викс И.П., Хауэлл М.Л., Хэнкок Т., Кохсака Х., Оли Т., Карсон Д.А. (март 1995 г.). «Бактериальные липополисахаридные связываются с плазмидной ДНК: последствия для животных моделей и генной терапии человека». Генная терапия человека . 6 (3): 317–323. doi : 10.1089/hum.1995.6.3-317 . PMID 7779915 .
- ^ Ватанабе Дж., Миядзаки Ю., Циммерман Г.А., Альбертин К.Х., Макинтайр Т.М. (октябрь 2003 г.). «Загрязнение эндотоксина овальбумина подавляет мышиные иммунологические реакции и развитие гиперреактивности дыхательных путей» . Журнал биологической химии . 278 (43): 42361–42368. doi : 10.1074/jbc.m307752200 . PMID 12909619 .
- ^ Комски Л (16 декабря 2014 г.). «Обнаружение эндотоксинов с помощью теста LAL, хромогенный метод» . Wako Chemicals USA, Inc. Архивирована из оригинала 29 марта 2015 года . Получено 14 марта 2015 года .
- ^ Iwanaga S (май 2007). «Биохимический принцип теста Limulus для обнаружения бактериальных эндотоксинов» . Материалы Японской академии. Серия B, физические и биологические науки . 83 (4): 110–119. Bibcode : 2007pjab ... 83..110i . doi : 10.2183/pjab.83.110 . PMC 3756735 . PMID 24019589 .
- ^ Ding JL, Ho B (август 2001 г.). «Новая эра в тестировании пирогена» (PDF) . Тенденции в биотехнологии . 19 (8): 277–281. doi : 10.1016/s0167-7799 (01) 01694-8 . PMID 11451451 . Архивировано из оригинала (PDF) 2 января 2014 года . Получено 2 января 2014 года .
Внешние ссылки
[ редактировать ]- Липополисахариды в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)