Липополисахарид

Липополисахарид , ныне более известный как эндотоксин , [ 1 ] это собирательный термин для компонентов внешней мембраны клеточной оболочки грамотрицательных бактерий, таких как E. coli и сальмонелла . [ 2 ] с общей структурной архитектурой. Липополисахариды ( ЛПС ) представляют собой большие молекулы, состоящие из трех частей: внешнего ядра полисахарида, называемого О- антигеном , внутреннего ядра олигосахарида и липида А (от которого в значительной степени зависит токсичность), все они ковалентно связаны. В современной терминологии термин «эндотоксин» часто используется как синоним ЛПС, хотя существует несколько эндотоксинов (в первоначальном смысле токсинов , находящихся внутри бактериальной клетки и высвобождающихся при ее распаде), не связанных с ЛПС, например: так называемые дельта-эндотоксина, белки продуцируемые Bacillus thuringiensis . [ 3 ]
Липополисахариды могут оказывать существенное влияние на здоровье человека, прежде всего за счет взаимодействия с иммунной системой. ЛПС является мощным активатором иммунной системы и пирогеном ( агентом, вызывающим лихорадку). [ 4 ] В тяжелых случаях ЛПС может вызвать бурную реакцию организма и множественные типы острой органной недостаточности. [ 5 ] что может привести к септическому шоку . [ 6 ] Есть данные, что на более низких уровнях и в течение более длительного периода времени ЛПС может играть важную и вредную роль в аутоиммунитете , ожирении , депрессии и клеточном старении . [ 7 ] [ 8 ] [ 9 ] [ 10 ]
Открытие
[ редактировать ]Токсическую активность ЛПС впервые обнаружил и назвал эндотоксином Рихард Фридрих Йоханнес Пфайффер . Он различал экзотоксины , токсины, которые выделяются бактериями в окружающую среду, и эндотоксины, которые являются токсинами «внутри» бактериальной клетки и высвобождаются только после разрушения внешней мембраны бактерии. [ 11 ] Последующие работы показали, что высвобождение ЛПС из грамотрицательных микробов не обязательно требует разрушения бактериальной клеточной стенки, а, скорее, ЛПС секретируется как часть нормальной физиологической активности транспорта мембранных везикул в форме везикул внешней мембраны бактерий (OMV). ) , который также может содержать другие факторы вирулентности и белки. [ 12 ] [ 2 ]
Функции у бактерий
[ редактировать ]ЛПС является основным компонентом внешней клеточной мембраны грамотрицательных бактерий , внося значительный вклад в структурную целостность бактерий и защищая мембрану от определенных видов химического воздействия. ЛПС является наиболее распространенным антигеном на поверхности клеток большинства грамотрицательных бактерий, составляя до 80% внешней мембраны E. coli и сальмонелл . [ 2 ] ЛПС увеличивает отрицательный заряд клеточной мембраны и помогает стабилизировать общую структуру мембраны. Это имеет решающее значение для многих грамотрицательных бактерий, которые погибают, если кодирующие их гены мутируют или удаляются. Однако оказывается, что ЛПС не является существенным, по крайней мере, для некоторых грамотрицательных бактерий, таких как Neisseria meningitidis , Moraxella cataralis и Acinetobacter baumannii . [ 13 ] Он также участвует в непатогенных аспектах бактериальной экологии, включая поверхностную адгезию, чувствительность к бактериофагам и взаимодействие с хищниками, такими как амебы . ЛПС также необходим для функционирования омптинов , класса бактериальных протеаз. [ 14 ]
Состав
[ редактировать ]
ЛПС являются амфипатическими и состоят из трех частей: антигена О (или полисахарида О), который является гидрофильным, основного олигосахарида (также гидрофильного) и липида А , гидрофобного домена.
О-антиген
[ редактировать ]Повторяющийся гликановый полимер, содержащийся в ЛПС, называется О- антигеном , О- полисахаридом или О-боковой цепью бактерий. О-антиген прикреплен к центральному олигосахариду и включает самый внешний домен молекулы ЛПС. Структура и состав О-цепи сильно варьируют от штамма к штамму, что определяет серологическую специфичность исходного бактериального штамма; [ 15 ] Существует более 160 различных структур О-антигена, продуцируемых разными штаммами E. coli . [ 16 ] Наличие или отсутствие О-цепей определяет, считается ли ЛПС «шероховатым» или «гладким». Полноразмерные О-цепи сделают ЛПС гладким, тогда как отсутствие или уменьшение количества О-цепей сделает ЛПС грубым. [ 17 ] Бактерии с шероховатым ЛПС обычно имеют клеточные мембраны, более проницаемые для гидрофобных антибиотиков, поскольку шероховатый ЛПС более гидрофобен . [ 18 ] О-антиген экспонируется на самой внешней поверхности бактериальной клетки и, как следствие, является мишенью для распознавания антителами хозяина .
Основной
[ редактировать ]Основной домен всегда содержит олигосахаридный компонент, который прикрепляется непосредственно к липиду А и обычно содержит сахара, такие как гептоза и 3-дезокси-D-манно-окт-2-улозоновая кислота (также известная как KDO, кето-дезоксиоктулозонат). [ 19 ] Ядерный олигосахарид менее изменчив по своей структуре и составу, причем данная структура ядра является общей для больших групп бактерий. [ 15 ] Ядра ЛПС многих бактерий содержат также неуглеводные компоненты, такие как фосфаты, аминокислоты и заместители этаноламина.
Липид А
[ редактировать ]Липид А в нормальных условиях представляет собой фосфорилированный глюкозамина дисахарид , украшенный множеством жирных кислот . Эти гидрофобные цепи жирных кислот закрепляют ЛПС в бактериальной мембране, а остальная часть ЛПС выступает с поверхности клетки. Домен липида А является наиболее биологически активным и ответственен за большую часть токсичности грамотрицательных бактерий . Когда бактериальные клетки лизируются иммунной системой , фрагменты мембран, содержащие липид А, могут попасть в кровоток, вызывая лихорадку, диарею и, возможно, фатальный эндотоксический септический шок (форма септического шока ). Липид А является очень консервативным компонентом ЛПС. [ 20 ] Однако структура липида А варьируется у разных видов бактерий. Структура липида А во многом определяет степень и характер общей иммунной активации организма хозяина. [ 21 ]
Липоолигосахариды
[ редактировать ]«Грубая форма» ЛПС имеет более низкую молекулярную массу из-за отсутствия полисахарида О. На его месте находится короткий олигосахарид: эта форма известна как липоолигосахарид (LOS) и представляет собой гликолипид, обнаруженный во внешней мембране некоторых типов грамотрицательных бактерий , таких как Neisseria spp. и Haemophilus spp. [ 7 ] [ 22 ] ЛОС играет центральную роль в поддержании целостности и функциональности внешней мембраны оболочки грамотрицательных клеток. ЛОС играют важную роль в патогенезе некоторых бактериальных инфекций, поскольку способны действовать как иммуностимуляторы и иммуномодуляторы. [ 7 ] Кроме того, молекулы LOS ответственны за способность некоторых бактериальных штаммов проявлять молекулярную мимикрию и антигенное разнообразие , помогая уклоняться от иммунной защиты хозяина и, таким образом, способствуя вирулентности этих бактериальных штаммов . В случае Neisseria meningitidis липидная часть А молекулы имеет симметричную структуру, а внутреннее ядро состоит из фрагментов 3-дезокси-D-манно-2-октулозоновой кислоты (KDO) и гептозы (Hep). Внешняя сердцевинная олигосахаридная цепь варьируется в зависимости от бактериального штамма . [ 7 ] [ 22 ]
детоксикация ЛПС
[ редактировать ]Высококонсервативный фермент хозяина, называемый ацилоксиацилгидролазой (AOAH), может детоксицировать ЛПС, когда он попадает или вырабатывается в тканях животных. Он также может превращать ЛПС в кишечнике в ингибитор ЛПС. Нейтрофилы, макрофаги и дендритные клетки продуцируют эту липазу, которая инактивирует ЛПС путем удаления двух вторичных ацильных цепей из липида А с образованием тетраацил-ЛПС. Если мышам вводить ЛПС парентерально, у мышей, у которых отсутствует AOAH, развиваются высокие титры неспецифических антител, развивается длительная гепатомегалия и наблюдается длительная толерантность к эндотоксинам. Инактивация ЛПС может потребоваться животным для восстановления гомеостаза после парентерального воздействия ЛПС. [ 23 ] Хотя у мышей имеется множество других механизмов ингибирования передачи сигналов ЛПС, ни один из них не способен предотвратить эти изменения у животных, у которых отсутствует AOAH.
Дефосфорилирование ЛПС кишечной щелочной фосфатазой может снизить тяжесть Salmonella Tryphimurium и Clostridioides difficile инфекции , восстанавливая нормальную микробиоту кишечника. [ 24 ] Щелочная фосфатаза предотвращает воспаление кишечника (и « дырявый кишечник ») из бактерий путем дефосфорилирования липидной части ЛПС. [ 25 ] [ 26 ] [ 27 ]
Биосинтез и транспорт
[ редактировать ]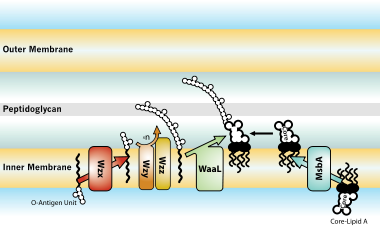
Весь процесс производства ЛПС начинается с молекулы под названием липид A-Kdo2, которая сначала создается на поверхности внутренней мембраны бактериальной клетки. Затем к этой молекуле на внутренней мембране добавляются дополнительные сахара, прежде чем она перемещается в пространство между внутренней и внешней мембранами ( периплазматическое пространство ) с помощью белка под названием MsbA. О-антиген, еще одна часть ЛПС, вырабатывается специальными ферментными комплексами на внутренней мембране. Затем он перемещается на внешнюю мембрану через три разные системы: одна Wzy-зависимая, другая зависит от транспортеров ABC, а третья включает в себя синтазо-зависимый процесс. [ 30 ]
В конечном итоге ЛПС транспортируется на внешнюю мембрану с помощью межмембранного мостика белков-переносчиков липолисахаридов (Lpt). [ 29 ] [ 31 ] Этот транспортер является потенциальной мишенью антибиотиков. [ 32 ] [ 33 ]
Биологическое воздействие на хозяев, инфицированных грамотрицательными бактериями.
[ редактировать ]Хранение ЛПС в организме
[ редактировать ]В организме человека имеются эндогенные запасы ЛПС. [ 34 ] Эпителиальные поверхности колонизированы сложной микробной флорой (включая грамотрицательные бактерии), численность которой превышает численность клеток человека в 10:1. Грамотрицательные бактерии выделяют эндотоксины. Это взаимодействие хозяина и микроба представляет собой симбиотические отношения, которые играют решающую роль в системном иммунологическом гомеостазе. Когда это нарушается, это может привести к таким заболеваниям, как эндотоксемия и эндотоксический септический шок.
Иммунный ответ
[ редактировать ]ЛПС действует как прототип эндотоксина, поскольку он связывает комплекс CD14 / TLR4 / MD2 рецепторный во многих типах клеток, но особенно в моноцитах , дендритных клетках , макрофагах и В-клетках , что способствует секреции провоспалительных цитокинов , оксида азота и эйкозаноидов. . [ 35 ] Брюс Бейтлер был удостоен части Нобелевской премии по физиологии и медицине 2011 года за работу, продемонстрировавшую, что TLR4 является рецептором ЛПС. [ 36 ] [ 37 ]
В рамках клеточной реакции на стресс супероксид является одной из основных активных форм кислорода, индуцируемых ЛПС в различных типах клеток, которые экспрессируют TLR ( толл-подобный рецептор ). [ 38 ] ЛПС также является экзогенным пирогеном (веществом, вызывающим лихорадку). [ 4 ]
Функция ЛПС находится под экспериментальным исследованием в течение нескольких лет из-за его роли в активации многих факторов транскрипции . ЛПС также производит множество типов медиаторов, участвующих в септическом шоке . Из млекопитающих человек гораздо более чувствителен к ЛПС, чем другие приматы. [ 39 ] а также другие животные (например, мыши). Доза 1 мкг/кг вызывает шок у людей, но мыши переносят дозу, в тысячу раз большую. [ 40 ] Это может быть связано с различиями в уровне циркулирующих естественных антител между двумя видами. [ 41 ] [ 42 ] Это также может быть связано с многочисленными иммунными тактиками против патогенов и частью многогранной антимикробной стратегии, основанной на поведенческих изменениях человека в ходе эволюции нашего вида (например, мясоедение, методы ведения сельского хозяйства и курение). [ 39 ] Саид и др. показали, что ЛПС вызывает IL-10- зависимое ингибирование расширения и функционирования Т-клеток CD4 путем повышения уровня PD-1 в моноцитах , что приводит к продукции IL-10 моноцитами после связывания PD-1 с помощью PD-L1 . [ 43 ]
Эндотоксины в значительной степени ответственны за драматические клинические проявления инфекций, вызванных патогенными грамотрицательными бактериями, такими как Neisseria meningitidis , возбудителями, вызывающими менингококковые заболевания , включая менингококцемию , синдром Уотерхауса-Фридериксена и менингит .
Было показано, что части ЛПС из нескольких бактериальных штаммов химически подобны молекулам поверхности клеток человека-хозяина; Способность некоторых бактерий представлять на своей поверхности молекулы, химически идентичные или подобные поверхностным молекулам некоторых типов клеток-хозяев, называется молекулярной мимикрией . [ 44 ] Например, у Neisseria meningitidis L2,3,5,7,9 терминальная тетрасахаридная часть олигосахарида (лакто-N-неотетраоза) представляет собой тот же тетрасахарид, что и параглобозид , предшественник антигенов ABH, гликолипидных обнаруженных на эритроцитах человека. . [ 7 ] В другом примере терминальная трисахаридная часть (лактотриаоза) олигосахарида из патогенных видов Neisseria spp. ЛОС также обнаружен в гликосфинголипидах лактоносерного ряда клеток человека. [ 7 ] большинство менингококков из групп B и C, а также гонококков имеют этот трисахарид как часть своей структуры LOS. Было показано, что [ 7 ] Присутствие этих «имитаторов» поверхности клеток человека может, помимо действия в качестве «камуфляжа» от иммунной системы, играть роль в устранении иммунной толерантности при заражении хозяев определенными генотипами человеческого лейкоцитарного антигена (HLA), такими как HLA-B35 . [ 7 ]
ЛПС может восприниматься непосредственно гемопоэтическими стволовыми клетками (ГСК) посредством связи с TLR4, вызывая их пролиферацию в ответ на системную инфекцию. Этот ответ активирует передачу сигналов TLR4-TRIF-ROS-p38 внутри HSC и посредством устойчивой активации TLR4 может вызвать пролиферативный стресс, приводящий к нарушению их конкурентной способности к репопуляции. [ 45 ] Заражение мышей с помощью S. typhimurium показало аналогичные результаты, что подтверждает экспериментальную модель также и in vivo .
Влияние изменчивости на иммунный ответ
[ редактировать ]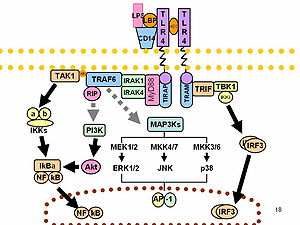
О-антигены (внешние углеводы) представляют собой наиболее изменчивую часть молекулы ЛПС, придающую антигенную специфичность. Напротив, липид А является наиболее консервативной частью. Однако состав липида А также может различаться (например, по количеству и природе ацильных цепей даже внутри или между родами). Некоторые из этих вариаций могут придавать этим ЛПС антагонистические свойства. Например, дифосфориллипид А Rhodobacter sphaeroides (RsDPLA) является мощным антагонистом ЛПС в клетках человека, но является агонистом в клетках хомяка и лошади. [ 46 ]
Было высказано предположение, что конический липид А (например, из E. coli ) является более агонистическим, в то время как менее конический липид А, такой как у Porphyromonas gingivalis, может активировать другой сигнал ( TLR2 вместо TLR4), а полностью цилиндрический липид А, такой как у Rhodobacter sphaeroides является антагонистом TLR. [ 47 ] [ 48 ] В целом кластеры генов ЛПС сильно варьируют между разными штаммами, подвидами, видами бактериальных возбудителей растений и животных. [ 49 ] [ 50 ]
Нормальная сыворотка крови человека содержит антитела против LOS, обладающие бактерицидным действием, а пациенты с инфекциями, вызванными серотипически разными штаммами, обладают антителами против LOS, которые отличаются по своей специфичности по сравнению с нормальной сывороткой. [ 51 ] Эти различия в гуморальном иммунном ответе на разные типы ЛОС можно объяснить структурой молекулы ЛОС, прежде всего структурой олигосахаридной части молекулы ЛОС. [ 51 ] У Neisseria gonorrhoeae было продемонстрировано, что антигенность молекул ЛОС может меняться во время инфекции из-за способности этих бактерий синтезировать более одного типа ЛОС. [ 51 ] характеристика, известная как изменение фазы . Кроме того, Neisseria gonorrhoeae , а также Neisseria meningitidis и Haemophilus influenzae , [ 7 ] способны к дальнейшей модификации своей ЛОС in vitro , например, посредством сиалилирования (модификация остатками сиаловой кислоты), и в результате способны повысить свою устойчивость к опосредованному комплементом уничтожению [ 51 ] или даже подавлять активацию комплемента [ 7 ] или уклоняться от воздействия бактерицидных антител. [ 7 ] Сиалилирование может также способствовать затруднению прикрепления нейтрофилов и фагоцитозу клетками иммунной системы, а также снижению окислительного взрыва. [ 7 ] Было также показано, что Haemophilus somnus , патоген крупного рогатого скота, демонстрирует изменение фазы LOS, характеристику, которая может помочь в уклонении от бычьего хозяина. иммунной защиты [ 52 ] В совокупности эти наблюдения позволяют предположить, что вариации поверхностных молекул бактерий, таких как LOS, могут помочь патогену уклоняться как от гуморальной (опосредованной антителами и комплементом), так и клеточной (например, уничтожение нейтрофилами) иммунной защиты хозяина.
Неканонические пути распознавания ЛПС
[ редактировать ]Недавно было показано, что помимо путей, опосредованных TLR4 , некоторые члены семейства ионных каналов переходного рецепторного потенциала распознают ЛПС. [ 53 ] LPS-опосредованная активация TRPA1 была показана на мышах [ 54 ] и Drosophila melanogaster . мухи [ 55 ] В более высоких концентрациях ЛПС активирует сенсорных каналов TRP и других членов семейства , таких как TRPV1 , TRPM3 и в некоторой степени TRPM8 . [ 56 ] ЛПС распознается TRPV4 на эпителиальных клетках. Активация TRPV4 с помощью ЛПС была необходима и достаточна для индукции продукции оксида азота с бактерицидным эффектом. [ 57 ]
Тестирование
[ редактировать ]Липополисахарид является важным фактором, делающим бактерии вредными, и помогает разделить их на разные группы в зависимости от их структуры и функций. Это делает ЛПС полезным маркером для различия различных грамотрицательных бактерий. Быстрое выявление и понимание типов вовлеченных патогенов имеет решающее значение для оперативного контроля и лечения инфекций. Поскольку ЛПС является основным триггером иммунного ответа в наших клетках, он действует как ранний сигнал острой инфекции. Таким образом, тестирование на ЛПС более специфично и значимо, чем многие другие серологические тесты. [ 58 ]
Современные методы тестирования ЛПС весьма чувствительны, но многие из них с трудом различают разные группы ЛПС. Кроме того, природа ЛПС, который обладает как водопритягивающими, так и водоотталкивающими свойствами (амфифильный), затрудняет разработку чувствительных и удобных для пользователя тестов. [ 58 ]
Типичные методы обнаружения основаны на идентификации липида А, входящего в состав ЛПС, поскольку липид А очень похож среди разных видов и серотипов бактерий. Методы тестирования ЛПС делятся на шесть категорий, и они часто пересекаются: тесты in vivo, тесты in vitro, модифицированные иммуноанализы, биологические анализы и химические анализы. [ 58 ]
Анализ активности эндотоксина
[ редактировать ]Поскольку уровень ЛПС очень трудно измерить в цельной крови, а большая часть ЛПС связана с белками и комплементом, в 2003 году FDA США разработало и одобрило анализ активности эндотоксина (EAA™). EAA — это быстрый хемилюминесцентный иммунодиагностический тест in vitro. . Он использует специфические моноклональные антитела для измерения активности эндотоксина в образцах цельной крови с ЭДТА. В этом анализе используется биологический ответ нейтрофилов в крови пациента на иммунологический комплекс эндотоксина и экзогенных антител – образующаяся хемилюминесцентная реакция создает излучение света. Величина хемилюминесценции пропорциональна логарифмической концентрации ЛПС в образце и является мерой активности эндотоксина в крови. [ 59 ] Анализ специфически реагирует с фрагментом липида А ЛПС грамотрицательных бактерий и не вступает в перекрестную реакцию с компонентами клеточной стенки грамположительных бактерий и других микроорганизмов.
Патофизиология
[ редактировать ]ЛПС — мощный токсин, который при попадании в организм вызывает воспаление, связываясь с клеточными рецепторами. Избыточное содержание ЛПС в крови (эндотоксемия) может вызвать крайне летальную форму сепсиса, известную как эндотоксический септический шок. [ 5 ] Это состояние включает симптомы, которые попадают в континуум патофизиологических состояний, начиная с синдрома системной воспалительной реакции (SIRS) и заканчивая синдромом полиорганной дисфункции (MODS) перед смертью. Ранние симптомы включают учащенное сердцебиение, учащенное дыхание, изменения температуры и проблемы со свертываемостью крови, что приводит к расширению кровеносных сосудов и уменьшению объема крови, что приводит к клеточной дисфункции. [ 58 ]
Недавние исследования показывают, что даже небольшое воздействие ЛПС связано с аутоиммунными заболеваниями и аллергиями. Высокий уровень ЛПС в крови может привести к метаболическому синдрому, увеличивая риск таких заболеваний, как диабет, болезни сердца и проблемы с печенью. [ 58 ]
ЛПС также играет решающую роль в симптомах, вызванных инфекциями, вызываемыми вредными бактериями, включая тяжелые состояния, такие как синдром Уотерхауса-Фридериксена, менингококцемия и менингит. Некоторые бактерии могут адаптировать свой ЛПС, чтобы вызывать длительные инфекции в дыхательной и пищеварительной системах. [ 58 ]
Недавние исследования показали, что ЛПС разрушает липиды клеточных мембран, влияя на уровень холестерина и обмен веществ, что потенциально приводит к повышению уровня холестерина, аномальному уровню липидов в крови и неалкогольной жировой болезни печени. В некоторых случаях ЛПС может препятствовать выведению токсинов, что может быть связано с неврологическими проблемами. [ 58 ]
Влияние на здоровье
[ редактировать ]В целом, влияние ЛПС на здоровье обусловлено его способностью выступать в качестве мощного активатора и модулятора иммунной системы, особенно индуцировать воспаление. ЛПС непосредственно цитотоксичен и обладает сильным иммуностимулирующим действием: поскольку иммунные клетки хозяина распознают ЛПС, комплемент сильно активируется. Активация комплемента и возрастающая противовоспалительная реакция могут привести к дисфункции иммунных клеток, иммуносупрессии, распространенной коагулопатии, серьезному повреждению тканей и могут прогрессировать до полисистемной недостаточности органов и смерти. [ 39 ]
эндотоксемия
[ редактировать ]Наличие эндотоксинов в крови называется эндотоксемией. Высокий уровень эндотоксемии может привести к септическому шоку . [ 60 ] или, более конкретно, эндотоксический септический шок, [ 5 ] тогда как более низкая концентрация эндотоксинов в кровотоке называется метаболической эндотоксемией. [ 61 ] Эндотоксемия связана с ожирением, диетой, [ 62 ] сердечно-сосудистые заболевания, [ 62 ] и диабет, [ 61 ] хотя на это может повлиять и генетика хозяина. [ 63 ]
эндотоксемия кишечного генеза, особенно на границе хозяин-возбудитель . Кроме того, важным фактором развития алкогольного гепатита считается [ 64 ] который, вероятно, развивается на основе синдрома избыточного бактериального роста в тонкой кишке и повышенной кишечной проницаемости . [ 65 ]
Липид А может вызывать неконтролируемую активацию иммунной системы млекопитающих с выработкой медиаторов воспаления , что может привести к эндотоксическому септическому шоку . [ 22 ] [ 5 ] Эта воспалительная реакция в первую очередь опосредуется Toll-подобным рецептором 4 , который отвечает за активацию клеток иммунной системы. [ 22 ] Повреждение эндотелиального слоя кровеносных сосудов, вызванное этими медиаторами воспаления, может привести к синдрому утечки капилляров , расширению кровеносных сосудов и снижению сердечной функции и может еще больше усугубить шок. [ 66 ] ЛПС также является мощным активатором комплемена. [ 66 ] Неконтролируемая активация комплемента может вызвать деструктивное повреждение эндотелия, приводящее к диссеминированному внутрисосудистому свертыванию крови (ДВС-синдром) или атипическому гемолитико-уремическому синдрому (аГУС) с поражением различных органов, в том числе почек и легких. [ 67 ] На коже могут проявляться последствия повреждения сосудов, часто сопровождающиеся истощением факторов свертывания крови в виде петехий , пурпуры и экхимозов . Конечности также могут поражаться, иногда с такими разрушительными последствиями, как развитие гангрены , требующей последующей ампутации . [ 66 ] Потеря функции надпочечников может вызвать надпочечниковую недостаточность , а дополнительное кровоизлияние в надпочечники вызывает синдром Уотерхауса-Фридериксена , оба из которых могут быть опасными для жизни.
Сообщалось также, что гонококковый ЛОС может вызвать повреждение фаллопиевых труб человека . [ 51 ]
Лечение эндотоксемии
[ редактировать ]Toraymyxin© (Toray, Токио, Япония) — широко используемый метод экстракорпоральной терапии удаления эндотоксинов посредством прямой гемоадсорбции (также называемой гемоперфузией). Это картридж на основе полистирола, в котором молекулы полимиксина B (PMX-B) ковалентно связаны с содержащимися в нем сетчатыми волокнами. Полимиксины представляют собой циклические катионные полипептидные антибиотики, полученные из Bacillus Polymyxa, с эффективной антимикробной активностью в отношении грамотрицательных бактерий, но их внутривенное клиническое применение ограничено из-за их нефротоксичности и побочных эффектов нейротоксичности. [ 68 ] Экстракорпоральное использование картриджа Toraymyxin© позволяет PMX-B связывать липид А с очень стабильным взаимодействием с его гидрофобными остатками, тем самым нейтрализуя эндотоксины, когда кровь фильтруется через экстракорпоральный контур внутри картриджа, тем самым обращая вспять эндотоксемию и избегая ее токсических системных эффектов. . [ 69 ]
Аутоиммунное заболевание
[ редактировать ]Считается, что молекулярная мимикрия некоторых молекул ЛОС вызывает аутоиммунные реакции хозяина, такие как обострения рассеянного склероза . [ 7 ] [ 44 ] Другие примеры бактериальной мимикрии структур хозяина посредством LOS обнаруживаются у бактерий Helicobacter pylori и Campylobacter jejuni , организмов, вызывающих желудочно-кишечные заболевания у людей, и Haemophilus ducreyi, вызывающих мягкий шанкр . Некоторые серотипы ЛПС C. jejuni (относящиеся к определенным тетра- и пентасахаридным фрагментам основного олигосахарида) также связаны с синдромом Гийена-Барре и вариантом Гийена-Барре, называемым синдромом Миллера-Фишера . [ 7 ]
Связь с ожирением
[ редактировать ]Эпидемиологические исследования показали, что повышенная эндотоксиновая нагрузка, которая может быть результатом увеличения популяций эндотоксин-продуцирующих бактерий в кишечном тракте, связана с определенными группами пациентов, связанных с ожирением. [ 8 ] [ 70 ] [ 71 ] Другие исследования показали, что очищенный эндотоксин из Escherichia coli может вызывать ожирение и резистентность к инсулину при введении в стерильные мышиные модели . [ 72 ] Более недавнее исследование выявило потенциальную роль Enterobacter cloacae B29 в развитии ожирения и резистентности к инсулину у пациента. [ 73 ] Предполагаемый механизм связи эндотоксина с ожирением заключается в том, что эндотоксин индуцирует воспалительный путь, ответственный за наблюдаемое ожирение и резистентность к инсулину. [ 72 ] Роды бактерий, связанные с эффектами ожирения, связанными с эндотоксинами, включают Escherichia и Enterobacter .
Депрессия
[ редактировать ]Существуют экспериментальные и наблюдательные данные о том, что ЛПС может играть роль в развитии депрессии. Введение ЛПС мышам может привести к симптомам депрессии, и, по-видимому, у некоторых людей с депрессией наблюдается повышенный уровень ЛПС. Иногда воспаление может играть роль в развитии депрессии, а ЛПС является провоспалительным. [ 9 ]
Клеточное старение
[ редактировать ]Воспаление, вызванное ЛПС, может вызвать клеточное старение , как было показано для эпителиальных клеток легких и клеток микроглии (последнее приводит к нейродегенерации ). [ 10 ]
Роль загрязнителя в биотехнологиях и исследованиях
[ редактировать ]Липополисахариды являются частыми примесями плазмидной ДНК, полученной из бактерий, или белков, экспрессируемых бактериями, и их необходимо удалять из ДНК или белка, чтобы избежать экспериментов по загрязнению и токсичности продуктов, произведенных с использованием промышленной ферментации . [ 74 ]
Овальбумин часто загрязнен эндотоксинами. Овальбумин является одним из тщательно изученных белков на животных моделях, а также признанным модельным аллергеном гиперреактивности дыхательных путей (AHR). Коммерчески доступный овальбумин, загрязненный ЛПС, может фальсифицировать результаты исследований, поскольку он неточно отражает влияние белкового антигена на физиологию животных. [ 75 ]
При фармацевтическом производстве необходимо удалять все следы эндотоксина из тары с лекарственными препаратами, поскольку даже небольшие количества эндотоксина вызывают заболевания у человека. депирогенизации . Для этой цели используется печь Для полного разрушения ЛПС необходимы температуры выше 300 °C. [ 76 ]
Стандартным анализом для обнаружения присутствия эндотоксина является анализ лизата амебоцитов Limulus (LAL), в котором используется кровь мечехвоста ( Limulus polyphemus ). [ 77 ] Очень низкие уровни ЛПС могут вызвать коагуляцию лизата лимулюса из-за мощной амплификации через ферментативный каскад. Однако из-за сокращения популяции мечехвостов и того факта, что существуют факторы, которые мешают анализу LAL, были предприняты усилия по разработке альтернативных методов анализа, наиболее многообещающими из которых являются ELISA тесты с использованием рекомбинантной версии белка в LAL-анализ, фактор C. [ 78 ]
См. также
[ редактировать ]- Биоаэрозоль
- Депирогенизация
- Интерфейс хозяин-патоген
- Мукополисахарид
- Несфатин-1
- Реакция Шварцмана
- внешняя реклама
Ссылки
[ редактировать ]- ^ Ритшель Э.Т., Кирика Т., Шаде Ф.У., Мамат У., Шмидт Г., Лоппноу Х. и др. (февраль 1994 г.). «Бактериальный эндотоксин: молекулярные связи структуры с активностью и функцией» . Журнал ФАСЭБ . 8 (2): 217–225. дои : 10.1096/fasebj.8.2.8119492 . ПМИД 8119492 . S2CID 28156137 .
- ^ Jump up to: а б с Авила-Кальдерон Э.Д., Руис-Пальма М.Д., Агилера-Арреола М.Г., Веласкес-Гуадаррама Н., Руис Э.А., Гомес-Лунар З. и др. (2021). «Везикулы наружной мембраны грамотрицательных бактерий: взгляд на биогенез» . Границы микробиологии . 12 : 557902. дои : 10.3389/fmicb.2021.557902 . ПМЦ 7969528 . ПМИД 33746909 .
- ^ Хёфте Х., де Греве Х., Серинк Дж., Янсенс С., Махиллон Дж., Ампе С. и др. (декабрь 1986 г.). «Структурный и функциональный анализ клонированного дельта-эндотоксина Bacillus thuringiensis berliner 1715» . Европейский журнал биохимии . 161 (2): 273–280. дои : 10.1111/j.1432-1033.1986.tb10443.x . ПМИД 3023091 .
- ^ Jump up to: а б Рот Дж., Блаттейс К.М. (октябрь 2014 г.). «Механизмы образования и лизиса лихорадки: уроки экспериментальной лихорадки ЛПС». Комплексная физиология . 4 (4): 1563–1604. дои : 10.1002/cphy.c130033 . ISBN 978-0-470-65071-4 . ПМИД 25428854 .
- ^ Jump up to: а б с д Келлум Дж. А., Ронко С. (октябрь 2023 г.). «Роль эндотоксина при септическом шоке» . Критический уход . 27 (1): 400. дои : 10.1186/s13054-023-04690-5 . ПМЦ 10585761 . ПМИД 37858258 .
- ^ Деллинджер Р.П., Леви М.М., Роудс А., Аннан Д., Герлах Х., Опал С.М. и др. (февраль 2013 г.). «Кампания по выживанию при сепсисе: международные рекомендации по ведению тяжелого сепсиса и септического шока: 2012» . Медицина критических состояний . 41 (2): 580–637. дои : 10.1097/CCM.0b013e31827e83af . ПМИД 23353941 . S2CID 34855187 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Моран А.П., Прендергаст М.М., Аппельмелк Б.Дж. (декабрь 1996 г.). «Молекулярная мимикрия структур хозяина бактериальными липополисахаридами и ее вклад в развитие заболеваний» . ФЭМС Иммунология и медицинская микробиология . 16 (2): 105–115. дои : 10.1016/s0928-8244(96)00072-7 . ПМИД 8988391 .
- ^ Jump up to: а б Ласселин Дж., Щедловски М., Каршикофф Б., Энглер Х., Лекандер М., Консман Дж. П. (август 2020 г.). «Сравнение болезненного поведения, вызванного бактериальными липополисахаридами, у грызунов и людей: актуальность для симптомов тревоги и депрессии» . Неврологические и биоповеденческие обзоры . 115 : 15–24. doi : 10.1016/j.neubiorev.2020.05.001 . ПМИД 32433924 . S2CID 218665128 .
- ^ Jump up to: а б Вэй В, Джи С (декабрь 2018 г.). «Клеточное старение: Молекулярные механизмы и патогенность». Журнал клеточной физиологии . 233 (12): 9121–9135. дои : 10.1002/jcp.26956 . ПМИД 30078211 . S2CID 51924586 .
- ^ Пария СК (1 января 2009 г.). Учебник микробиологии и иммунологии . Индия: Эльзевир. ISBN 978-8131221631 .
- ^ Кулп А., Кюн М.Дж. (2010). «Биологические функции и биогенез секретируемых бактериальных везикул наружной мембраны» . Ежегодный обзор микробиологии . 64 : 163–184. дои : 10.1146/annurev.micro.091208.073413 . ПМЦ 3525469 . ПМИД 20825345 .
- ^ Чжан Г., Мередит Т.К., Кане Д. (декабрь 2013 г.). «О необходимости липополисахаридов для грамотрицательных бактерий» . Современное мнение в микробиологии . 16 (6): 779–785. дои : 10.1016/j.mib.2013.09.007 . ПМЦ 3974409 . ПМИД 24148302 .
- ^ Кукконен М., Корхонен Т.К. (июль 2004 г.). «Семейство энтеробактериальных поверхностных протеаз/адгезинов omptin: от домашнего хозяйства в Escherichia coli до системного распространения Yersinia pestis». Международный журнал медицинской микробиологии . 294 (1): 7–14. дои : 10.1016/j.ijmm.2004.01.003 . ПМИД 15293449 .
- ^ Jump up to: а б Галанос С., Фрейденберг М.А. (1993). «Бактериальные эндотоксины: биологические свойства и механизмы действия» . Медиаторы воспаления . 2 (7): С11-16. дои : 10.1155/S0962935193000687 . ПМЦ 2365449 . ПМИД 18475562 .
- ^ Раец Ч.Р., Уитфилд С. (2002). «Липополисахаридные эндотоксины» . Ежегодный обзор биохимии . 71 : 635–700. doi : 10.1146/annurev.biochem.71.110601.135414 . ПМК 2569852 . ПМИД 12045108 .
- ^ Риттиг М.Г., Кауфманн А., Робинс А., Шоу Б., Шпренгер Х., Гемса Д. и др. (декабрь 2003 г.). «Гладкий и шероховатый фенотипы липополисахаридов бруцелл индуцируют различный внутриклеточный транспорт и высвобождение цитокинов/хемокинов в моноцитах человека» . Журнал биологии лейкоцитов . 74 (6): 1045–1055. дои : 10.1189/jlb.0103015 . ПМИД 12960272 .
- ^ Цудзимото Х., Гото Н., Нишино Т. (декабрь 1999 г.). «Диффузия макролидных антибиотиков через внешнюю мембрану Moraxella cataralis». Журнал инфекции и химиотерапии . 5 (4): 196–200. дои : 10.1007/s101560050034 . ПМИД 11810516 . S2CID 2742306 .
- ^ Хершбергер С., Бинкли С.Б. (апрель 1968 г.). «Химия и метаболизм 3-дезокси-D-маннооктулозоновой кислоты. I. Стереохимическое определение» . Журнал биологической химии . 243 (7): 1578–1584. дои : 10.1016/S0021-9258(18)93581-7 . ПМИД 4296687 .
- ^ Ценг Ю.Л., Датта А., Колли В.К., Карлсон Р.В., Стивенс Д.С. (май 2002 г.). «Эндотоксин Neisseria meningitidis, состоящий только из интактного липида А: инактивация трансферазы менингококковой 3-дезокси-D-маннооктулозоновой кислоты» . Журнал бактериологии . 184 (9): 2379–2388. дои : 10.1128/JB.184.9.2379-2388.2002 . ПМК 134985 . ПМИД 11948150 .
- ^ Хан М.М., Эрнст О., Сан Дж., Фрейзер И.Д., Эрнст Р.К., Гудлетт Д.Р., Нита-Лазар А. (август 2018 г.). «Структурный анализ на основе масс-спектрометрии и стратегии системной иммунопротеомики для расшифровки ответа хозяина на эндотоксин». Журнал молекулярной биологии . 430 (17): 2641–2660. дои : 10.1016/j.jmb.2018.06.032 . ПМИД 29949751 . S2CID 49481716 .
- ^ Jump up to: а б с д Килар А., Дорней А., Кочиш Б. (2013). «Структурная характеристика бактериальных липополисахаридов с помощью масс-спектрометрии, а также методов оперативного и автономного разделения». Обзоры масс-спектрометрии . 32 (2): 90–117. Бибкод : 2013MSRv...32...90K . дои : 10.1002/mas.21352 . ПМИД 23165926 .
- ^ Манфорд Р., Лу М., Варли А.В. (2009). Глава 2 Убить бактерии… а также их посланников? . Достижения иммунологии. Том. 103. С. 29–48. дои : 10.1016/S0065-2776(09)03002-8 . ISBN 9780123748324 . ПМЦ 2812913 . ПМИД 19755182 .
- ^ Бильски Дж., Мазур-Бялы А., Войчик Д., Заградник-Бильска Дж., Бжозовский Б., Магеровски М. и др. (2017). «Роль кишечной щелочной фосфатазы при воспалительных заболеваниях желудочно-кишечного тракта» . Медиаторы воспаления . 2017 : 9074601. doi : 10.1155/2017/9074601 . ПМЦ 5339520 . ПМИД 28316376 .
- ^ Бейтс Дж. М., Акерлунд Дж., Митге Э., Гиймен К. (декабрь 2007 г.). «Кишечная щелочная фосфатаза детоксицирует липополисахарид и предотвращает воспаление у рыбок данио в ответ на микробиоту кишечника» . Клетка-хозяин и микроб . 2 (6): 371–382. дои : 10.1016/j.chom.2007.10.010 . ПМК 2730374 . ПМИД 18078689 .
- ^ Алам С.Н., Яммин Х., Моавен О., Ахмед Р., Мосс А.К., Бисвас Б. и др. (апрель 2014 г.). «Кишечная щелочная фосфатаза предотвращает чувствительность к кишечным патогенам, вызванную антибиотиками» . Анналы хирургии . 259 (4): 715–722. дои : 10.1097/sla.0b013e31828fae14 . ПМЦ 3855644 . ПМИД 23598380 .
- ^ Лаллес ЖП (февраль 2014 г.). «Кишечная щелочная фосфатаза: новые функции и защитные эффекты» . Обзоры питания . 72 (2): 82–94. дои : 10.1111/нуре.12082 . ПМИД 24506153 .
- ^ Ван X, Куинн Пи Джей (апрель 2010 г.). «Липополисахарид: путь биосинтеза и модификация структуры». Прогресс в исследованиях липидов . 49 (2): 97–107. дои : 10.1016/j.plipres.2009.06.002 . ПМИД 19815028 .
- ^ Jump up to: а б Руис Н., Кане Д., Силхави Т.Дж. (сентябрь 2009 г.). «Транспорт липополисахарида через клеточную оболочку: долгий путь открытий» . Обзоры природы. Микробиология . 7 (9): 677–683. дои : 10.1038/nrmicro2184 . ПМК 2790178 . ПМИД 19633680 .
- ^ Романо К.П., Хунг Д.Т. (март 2023 г.). «Нацеливание на биосинтез и транспорт ЛПС у грамотрицательных бактерий в эпоху множественной лекарственной устойчивости» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1870 (3): 119407. doi : 10.1016/j.bbamcr.2022.119407 . ПМЦ 9922520 . ПМИД 36543281 .
- ^ Шерман Д.Д., Се Р., Тейлор Р.Дж., Джордж А.Х., Окуда С., Фостер П.Дж. и др. (февраль 2018 г.). «Липополисахарид транспортируется на поверхность клетки с помощью белкового моста между мембранами» . Наука . 359 (6377): 798–801. Бибкод : 2018Sci...359..798S . doi : 10.1126/science.aar1886 . ПМЦ 5858563 . ПМИД 29449493 .
- ^ Пахил К.С., Гилман М.С., Байдин В., Клерфей Т., Маттеи П., Бьениоссек С. и др. (январь 2024 г.). «Новый антибиотик улавливает липополисахарид в его межмембранном переносчике» . Природа . 625 (7995): 572–577. Бибкод : 2024Natur.625..572P . дои : 10.1038/s41586-023-06799-7 . ПМЦ 10794137 . ПМИД 38172635 .
- ^ Зампалони С., Маттеи П., Блейхер К., Винтер Л., Тете С., Бухер С. и др. (январь 2024 г.). «Новый класс антибиотиков, нацеленный на переносчик липополисахаридов» . Природа . 625 (7995): 566–571. Бибкод : 2024Nature.625..566Z . дои : 10.1038/s41586-023-06873-0 . ПМЦ 10794144 . ПМИД 38172634 .
- ^ Маршалл Дж.К. (2005). «Липополисахарид: эндотоксин или экзогенный гормон?». Клин Инфекционный Дис . 41 (С7): С470–80. дои : 10.1086/432000 . ПМИД 16237650 .
- ^ Аббас А (2006). Основная иммунология . Эльзевир. ISBN 978-1-4160-2974-8 .
- ^ Полторак А., Хе Х, Смирнова И., Лю М.Ю., Ван Хаффель С., Ду Х. и др. (декабрь 1998 г.). «Дефектная передача сигналов LPS у мышей C3H/HeJ и C57BL/10ScCr: мутации в гене Tlr4». Наука . 282 (5396): 2085–2088. Бибкод : 1998Sci...282.2085P . дои : 10.1126/science.282.5396.2085 . ПМИД 9851930 .
- ^ «Нобелевская премия по физиологии и медицине 2011 года — пресс-релиз» . www.nobelprize.org . Архивировано из оригинала 23 марта 2018 года . Проверено 28 апреля 2018 г.
- ^ Ли Ю, Дэн С.Л., Лиан ZX, Ю К (июнь 2019 г.). «Роль Toll-подобных рецепторов в нитроксидативном стрессе у млекопитающих» . Клетки . 8 (6): 576. doi : 10.3390/cells8060576 . ПМК 6627996 . ПМИД 31212769 .
- ^ Jump up to: а б с Бринкворт Дж. Ф., Вализадеган Н. (2021 г.). «Сепсис и эволюция повышенной чувствительности человека к липополисахаридам». Эвол Антропол . 30 (2): 141–157. дои : 10.1002/evan.21887 . ПМИД 33689211 .
- ^ Уоррен Х.С., Фиттинг С., Хофф Э., Адиб-Конки М., Бисли-Топлифф Л., Тезини Б. и др. (январь 2010 г.). «Устойчивость к бактериальной инфекции: разница между видами может быть связана с белками в сыворотке» . Журнал инфекционных болезней . 201 (2): 223–232. дои : 10.1086/649557 . ПМК 2798011 . ПМИД 20001600 .
- ^ Рид Р.Р., Продеус А.П., Хан В., Сюй Т., Розен Ф.С., Кэрролл М.С. (июль 1997 г.). «Эндотоксиновый шок у мышей с дефицитом антител: раскрытие роли природных антител и комплемента в клиренсе липополисахаридов» . Журнал иммунологии . 159 (2): 970–975. дои : 10.4049/jimmunol.159.2.970 . ПМИД 9218618 .
- ^ Боес М., Продеус А.П., Шмидт Т., Кэрролл М.С., Чен Дж. (декабрь 1998 г.). «Критическая роль природного иммуноглобулина М в непосредственной защите от системной бактериальной инфекции» . Журнал экспериментальной медицины . 188 (12): 2381–2386. дои : 10.1084/jem.188.12.2381 . ПМК 2212438 . ПМИД 9858525 .
- ^ Саид Э.А., Дюпюи Ф.П., Траутманн Л., Чжан Ю., Ши Ю., Эль-Фар М. и др. (апрель 2010 г.). «Запрограммированная смерть-1, индуцированная выработкой интерлейкина-10 моноцитами, нарушает активацию CD4+ Т-клеток во время ВИЧ-инфекции» . Природная медицина . 16 (4): 452–459. дои : 10.1038/нм.2106 . ПМЦ 4229134 . ПМИД 20208540 .
- ^ Jump up to: а б Честейн Э.М., Миллер С.Д. (январь 2012 г.). «Молекулярная мимикрия как пусковой механизм аутоиммунного демиелинизирующего заболевания ЦНС» . Иммунологические обзоры . 245 (1): 227–238. дои : 10.1111/j.1600-065X.2011.01076.x . ПМЦ 3586283 . ПМИД 22168423 .
- ^ Такидзава Х., Фрич К., Ковтонюк Л.В., Сайто Ю., Яккала С., Джейкобс К. и др. (август 2017 г.). «Индуцированная патогенами передача врожденных иммунных сигналов TLR4-TRIF в гемопоэтических стволовых клетках способствует пролиферации, но снижает конкурентоспособность» . Клеточная стволовая клетка . 21 (2): 225–240.e5. дои : 10.1016/j.stem.2017.06.013 . ПМИД 28736216 .
- ^ Ломанн К.Л., Ванденплас М.Л., Бартон М.Х., Брайант К.Э., Мур Дж.Н. (2007). «Комплекс TLR4/MD-2 лошадей опосредует распознавание липополисахарида Rhodobacter sphaeroides в качестве агониста». Журнал исследований эндотоксинов . 13 (4): 235–242. дои : 10.1177/0968051907083193 . ПМИД 17956942 . S2CID 36784237 .
- ^ Нетеа М.Г., ван Деурен М., Куллберг Б.Дж., Кавайон Дж.М., Ван дер Меер Дж.В. (март 2002 г.). «Определяет ли форма липида А взаимодействие ЛПС с Toll-подобными рецепторами?». Тенденции в иммунологии . 23 (3): 135–139. дои : 10.1016/S1471-4906(01)02169-X . ПМИД 11864841 .
- ^ Сейдел У, Оикава М, Фукасе К, Кусумото С, Бранденбург К (май 2000 г.). «Внутренняя конформация липида А отвечает за агонистическую и антагонистическую активность». Европейский журнал биохимии . 267 (10): 3032–3039. дои : 10.1046/j.1432-1033.2000.01326.x . ПМИД 10806403 .
- ^ Ривз П.П., Ван Л. (2002). «Геномная организация ЛПС-специфичных локусов». В Hacker J, Kaper JB (ред.). Острова патогенности и эволюция патогенных микробов . Актуальные темы микробиологии и иммунологии. Том. 2. С. 109–35. дои : 10.1007/978-3-642-56031-6_7 . ISBN 978-3-540-42682-0 . ПМИД 12014174 . 264.
- ^ Патил П.Б., Сонти Р.В. (октябрь 2004 г.). «Вариация, свидетельствующая о горизонтальном переносе генов в локусе биосинтеза липополисахаридов (lps) у Xanthomonas oryzae pv. oryzae, возбудителя бактериальной пятнистости листьев риса» . БМК Микробиология . 4:40 . дои : 10.1186/1471-2180-4-40 . ПМК 524487 . ПМИД 15473911 .
- ^ Jump up to: а б с д и Ямасаки Р., Кервуд Д.Э., Шнайдер Х., Куинн К.П., Гриффисс Дж.М., Мандрелл Р.Э. (декабрь 1994 г.). «Структура липоолигосахарида, продуцируемого Neisseria gonorrhoeae, штамм 15253, выделенного от пациента с диссеминированной инфекцией. Доказательства нового пути гликозилирования гонококкового липоолигосахарида» . Журнал биологической химии . 269 (48): 30345–30351. дои : 10.1016/S0021-9258(18)43819-7 . ПМИД 7982947 .
- ^ Ховард М.Д., Кокс А.Д., Вайзер Дж.Н., Шуриг Г.Г., Инзана Т.Дж. (декабрь 2000 г.). «Антигенное разнообразие липоолигосахарида Haemophilus somnus: фазово-переменная доступность эпитопа фосфорилхолина» . Журнал клинической микробиологии . 38 (12): 4412–4419. doi : 10.1128/JCM.38.12.4412-4419.2000 . ПМК 87614 . ПМИД 11101573 .
- ^ Боонен Б., Альпизар Я.А., Месегер В.М., Талавера К. (август 2018 г.). «Каналы TRP как сенсоры бактериальных эндотоксинов» . Токсины . 10 (8): 326. doi : 10.3390/toxins10080326 . ПМК 6115757 . ПМИД 30103489 .
- ^ Месегер В., Альпизар Ю.А., Луис Э., Таджада С., Денлингер Б., Фахардо О. и др. (20 января 2014 г.). «Каналы TRPA1 опосредуют острое нейрогенное воспаление и боль, вызванную бактериальными эндотоксинами» . Природные коммуникации . 5 : 3125. Бибкод : 2014NatCo...5.3125M . дои : 10.1038/ncomms4125 . ПМЦ 3905718 . ПМИД 24445575 .
- ^ Солдано А., Альпизар Ю.А., Боонен Б., Франко Л., Лопес-Рекена А., Лю Г. и др. (июнь 2016 г.). «Вкусоопосредованное избегание бактериальных липополисахаридов посредством активации TRPA1 у дрозофилы» . электронная жизнь 5 . дои : 10.7554/eLife.13133 . ПМЦ 4907694 . ПМИД 27296646 .
- ^ Боонен Б., Альпизар Я.А., Санчес А., Лопес-Рекена А., Воэтс Т., Талавера К. (июль 2018 г.). «Дифференциальное воздействие липополисахарида на сенсорные каналы TRP мыши». Клеточный кальций . 73 : 72–81. дои : 10.1016/j.ceca.2018.04.004 . ПМИД 29689522 . S2CID 13681499 .
- ^ Альпизар Ю.А., Боонен Б., Санчес А., Юнг С., Лопес-Рекена А., Наерт Р. и др. (октябрь 2017 г.). «Активация TRPV4 запускает защитные реакции на бактериальные липополисахариды в эпителиальных клетках дыхательных путей» . Природные коммуникации . 8 (1): 1059. Бибкод : 2017NatCo...8.1059A . дои : 10.1038/s41467-017-01201-3 . ПМК 5651912 . ПМИД 29057902 .
- ^ Jump up to: а б с д и ж г Пейдж М.Дж., Келл Д.Б., Преториус Э. (2022). «Роль индуцированной липополисахаридами клеточной сигнализации при хроническом воспалении» . Хронический стресс . 6 : 24705470221076390. doi : 10.1177/24705470221076390 . ПМЦ 8829728 . ПМИД 35155966 .
- ^ Ромашин А.Д., Харрис Д.М., Риберио М.Б., Пейс Дж., Фостер Д.М., Уокер П.М., Маршалл Дж.К. (1998). «Быстрый анализ эндотоксина в цельной крови с использованием аутологичной нейтрофильно-зависимой хемилюминесценции». J Иммуноловые методы . 212 (2): 169–185. дои : 10.1016/s0022-1759(98)00003-9 . ПМИД 9672205 .
- ^ Опал СМ (2010). «Эндотоксины и другие триггеры сепсиса». Эндотокемия и эндотоксиновый шок . Вклад в нефрологию. Том. 167. стр. 14–24. дои : 10.1159/000315915 . ISBN 978-3-8055-9484-4 . ПМИД 20519895 .
- ^ Jump up to: а б Гомес Х.М., Коста Х.А., Альфенас Р.С. (март 2017 г.). «Метаболическая эндотоксемия и сахарный диабет: систематический обзор». Метаболизм . 68 : 133–144. дои : 10.1016/j.metabol.2016.12.009 . ПМИД 28183445 .
- ^ Jump up to: а б Каллио К.А., Хатонен К.А., Лехто М., Саломаа В., Мяннисто С., Пуссинен П.Ю. (апрель 2015 г.). «Эндотоксемия, питание и кардиометаболические нарушения». Акта Диабетологика . 52 (2): 395–404. дои : 10.1007/s00592-014-0662-3 . ПМИД 25326898 . S2CID 24020127 .
- ^ Лескеля Дж., Топпила И., Хярма М.А., Палвиайнен Т., Салминен А., Сандхольм Н. и др. (ноябрь 2021 г.). «Генетический профиль эндотоксемии обнаруживает связь с тромбоэмболией и инсультом» . Журнал Американской кардиологической ассоциации . 10 (21): e022482. дои : 10.1161/JAHA.121.022482 . ПМЦ 8751832 . ПМИД 34668383 .
- ^ Чекканти М., Аттили А., Бальдуччи Г., Аттилия Ф., Джакомелли С., Ротондо С. и др. (октябрь 2006 г.). «Острый алкогольный гепатит». Журнал клинической гастроэнтерологии . 40 (9): 833–841. дои : 10.1097/01.mcg.0000225570.04773.5d . ПМИД 17016141 .
- ^ Парлесак А., Шефер С., Шютц Т., Боде Дж.К., Боде С. (май 2000 г.). «Повышение проницаемости кишечника для макромолекул и эндотоксемии у больных хроническим злоупотреблением алкоголем на разных стадиях алкогольного заболевания печени». Журнал гепатологии . 32 (5): 742–747. дои : 10.1016/S0168-8278(00)80242-1 . ПМИД 10845660 .
- ^ Jump up to: а б с Стивенс Д.С., Гринвуд Б., Брандцаг П. (июнь 2007 г.). «Эпидемический менингит, менингококцемия и Neisseria meningitidis». Ланцет . 369 (9580): 2196–2210. дои : 10.1016/S0140-6736(07)61016-2 . ПМИД 17604802 . S2CID 16951072 .
- ^ Келлум Дж.А., Формек К.Л., Кернан К.Ф., Гомес Х., Карсильо Дж.А. (2022). «Подтипы и симптомы сепсиса». Клиники интенсивной терапии . 38 (2): 195–211. дои : 10.1016/j.ccc.2021.11.013 . ПМИД 35369943 .
- ^ Ли Дж., Нэйшн Р.Л., Тернидж Дж.Д., Милн Р.В., Култхард К., Рейнер С.Р., Патерсон Д.Л. (2006). «Колистин: новый антибиотик для лечения грамотрицательных бактериальных инфекций с множественной лекарственной устойчивостью». Ланцет Инфекционный Дис . 6 (9): 589–601. дои : 10.1016/S1473-3099(06)70580-1 . ПМИД 16931410 .
- ^ Вирци ГМ, Маттиотти М, де Кал М, Ронко С, Занелла М, ДеРоса С (2022). «Эндотоксин при сепсисе: методы обнаружения ЛПС и использование методов омикса» . Диагностика . 13 (1): 79. doi : 10.3390/diagnostics13010079 . ПМЦ 9818564 . ПМИД 36611371 .
- ^ Леппер П.М., Шуман С., Триантафилу К., Раше Ф.М., Шустер Т., Франк Х. и др. (июль 2007 г.). «Ассоциация липополисахарид-связывающего белка и ишемической болезни сердца у мужчин». Журнал Американского колледжа кардиологов . 50 (1): 25–31. дои : 10.1016/j.jacc.2007.02.070 . ПМИД 17601541 . S2CID 12136094 .
- ^ Руис А.Г., Касафонт Ф., Креспо Х., Кайон А., Майорга М., Эстебанес А. и др. (октябрь 2007 г.). «Уровни липополисахарид-связывающего белка в плазме и экспрессия гена TNF-альфа в печени у пациентов с ожирением: доказательства потенциальной роли эндотоксина в патогенезе неалкогольного стеатогепатита». Хирургия ожирения . 17 (10): 1374–1380. дои : 10.1007/s11695-007-9243-7 . ПМИД 18000721 . S2CID 44494003 .
- ^ Jump up to: а б Кани П.Д., Амар Дж., Иглесиас М.А., Поджи М., Кнауф С., Бастелика Д. и др. (июль 2007 г.). «Метаболическая эндотоксеемия вызывает ожирение и резистентность к инсулину» . Диабет . 56 (7): 1761–1772. дои : 10.2337/db06-1491 . ПМИД 17456850 .
- ^ Фей Н., Чжао Л. (апрель 2013 г.). «Условно-патогенный микроорганизм, выделенный из кишечника человека, страдающего ожирением, вызывает ожирение у стерильных мышей» . Журнал ISME . 7 (4): 880–884. Бибкод : 2013ISMEJ...7..880F . дои : 10.1038/ismej.2012.153 . ПМЦ 3603399 . ПМИД 23235292 .
- ^ Уикс И.П., Хауэлл М.Л., Хэнкок Т., Косака Х., Оли Т., Карсон Д.А. (март 1995 г.). «Бактериальный липополисахарид соочищается с плазмидной ДНК: значение для животных моделей и генной терапии человека». Генная терапия человека . 6 (3): 317–323. дои : 10.1089/hum.1995.6.3-317 . ПМИД 7779915 .
- ^ Ватанабэ Дж., Миядзаки Ю., Циммерман Г.А., Альбертина К.Х., Макинтайр Т.М. (октябрь 2003 г.). «Загрязнение овальбумина эндотоксинами подавляет иммунологические реакции мышей и развитие гиперреактивности дыхательных путей» . Журнал биологической химии . 278 (43): 42361–42368. дои : 10.1074/jbc.M307752200 . ПМИД 12909619 .
- ^ Комский Л. (16 декабря 2014 г.). «Обнаружение эндотоксинов с помощью LAL-теста, хромогенный метод» . Wako Chemicals USA, Inc. Архивировано из оригинала 29 марта 2015 года . Проверено 14 марта 2015 г.
- ^ Иванага С. (май 2007 г.). «Биохимический принцип теста Лимулуса для выявления бактериальных эндотоксинов» . Труды Японской академии. Серия Б, Физические и биологические науки . 83 (4): 110–119. Бибкод : 2007PJAB...83..110I . дои : 10.2183/pjab.83.110 . ПМЦ 3756735 . ПМИД 24019589 .
- ^ Дин Дж.Л., Хо Б. (август 2001 г.). «Новая эра испытаний на пирогенность» (PDF) . Тенденции в биотехнологии . 19 (8): 277–281. дои : 10.1016/s0167-7799(01)01694-8 . ПМИД 11451451 . Архивировано из оригинала (PDF) 2 января 2014 года . Проверено 2 января 2014 г.
Внешние ссылки
[ редактировать ]- Липополисахариды Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)