Генетика
| Часть серии о |
| Генетика |
|---|
 |
Генетика это изучение генов , генетических вариаций и наследственности организмов – . [1] [2] [3] Это важная отрасль биологии организмов , поскольку наследственность жизненно важна для эволюции . Грегор Мендель , моравский монах -августинец, работавший в XIX веке в Брно , был первым, кто начал научно изучать генетику. Мендель изучал «наследование признаков» — закономерности передачи признаков от родителей потомству с течением времени. Он заметил, что организмы (растения гороха) наследуют признаки посредством дискретных «единиц наследования». Этот термин, используемый до сих пор, представляет собой несколько двусмысленное определение того, что называется геном.
наследования генов по-прежнему Наследование признаков и механизмы молекулярного остаются основными принципами генетики в 21 веке, но современная генетика расширилась и стала изучать функции и поведение генов. Структура и функция генов, изменчивость и распределение изучаются в контексте клетки , организма (например, доминирование ) и в контексте популяции. Генетика породила ряд подполей, включая молекулярную генетику , эпигенетику и популяционную генетику . Организмы, изучаемые в широкой области, охватывают все области жизни ( археи , бактерии и эукарии ).
Генетические процессы работают в сочетании с окружающей средой и опытом организма, влияя на развитие и поведение , что часто называют « природа против воспитания» . Внутриклеточная . или внеклеточная среда живой клетки или организма может увеличивать или уменьшать транскрипцию генов Классический пример — два семени генетически идентичной кукурузы, одно из которых находится в умеренном климате , а другое — в засушливом (без достаточного количества водопадов и дождей). Хотя средняя высота, до которой могут вырасти два стебля кукурузы, определяется генетически, стебель в засушливом климате вырастает только до половины высоты стебля в умеренном климате из-за нехватки воды и питательных веществ в окружающей среде.
Этимология
[ редактировать ]Слово генетика происходит от древнегреческого γενετικός Genetikos, означающего «родительный падеж»/«генеративный», которое, в свою очередь, происходит от γένεσις Genesis, означающего «происхождение». [4] [5] [6]
История
[ редактировать ]Наблюдение о том, что живые существа наследуют черты своих родителей, использовалось с доисторических времен для улучшения сельскохозяйственных растений и животных посредством селекционного разведения . [7] [8] Современная наука генетика, стремящаяся понять этот процесс, началась с работы монаха- Грегора августинца Менделя в середине 19 века. [9]

До Менделя Имре Фештетич , венгерский дворянин, живший в Кёсеге до Менделя, был первым, кто использовал слово «генетический» в наследственном контексте и считается первым генетиком. Он описал несколько правил биологической наследственности в своей работе «Генетические законы природы» (Die Genetischen Gesetze der Natur, 1819). [10] Его второй закон аналогичен тому, который опубликовал Мендель. [11] В своем третьем законе он разработал основные принципы мутации (его можно считать предшественником Гюго де Фриза ). [12] Фештетич утверждал, что изменения, наблюдаемые в поколении сельскохозяйственных животных, растений и людей, являются результатом научных законов. [13] Фестетик эмпирически пришел к выводу, что организмы наследуют свои характеристики, а не приобретают их. Он признал рецессивные черты и присущие им вариации, постулировав, что черты прошлых поколений могут проявиться позже и организмы могут производить потомство с другими характеристиками. [14] Эти наблюдения представляют собой важную прелюдию к теории Менделя о наследственности частиц, поскольку они отражают переход наследственности от статуса мифа к статусу научной дисциплины, обеспечивая фундаментальную теоретическую основу для генетики в двадцатом веке. [10] [15]

Работе Менделя предшествовали другие теории наследственности. Популярной в XIX веке теорией, выдвинутой Чарльзом Дарвином в 1859 году в книге «Происхождение видов» , была смешанная наследственность : идея о том, что люди наследуют плавное сочетание признаков от своих родителей. [16] Работа Менделя предоставила примеры, когда признаки определенно не смешивались после гибридизации, показывая, что признаки возникают в результате комбинации различных генов, а не в результате непрерывного смешения. Смешение признаков в потомстве теперь объясняется действием множества генов с количественным эффектом . Другая теория, имевшая в то время некоторую поддержку, заключалась в наследственности приобретенных характеристик : вера в то, что люди наследуют черты, закрепленные их родителями. Сейчас известно, что эта теория (обычно связываемая с Жаном-Батистом Ламарком ) ошибочна: опыт отдельных людей не влияет на гены, которые они передают своим детям. [17] Дарвина Другие теории включали пангенезис (который имел как приобретенные, так и унаследованные аспекты) и Фрэнсисом Гальтоном как как частиц, так и унаследованного. переформулировку пангенезиса [18]
Менделевская генетика
[ редактировать ]
Современная генетика началась с исследований Менделя природы наследственности у растений. В своей статье « Versuche über Pflanzenhybriden » (« Опыты по гибридизации растений »), представленной в 1865 году Naturforschender Verein (Обществу исследований природы) в Брно , Мендель проследил закономерности наследования определенных признаков у растений гороха и описал их математически. Хотя этот образец наследования можно было наблюдать только для нескольких признаков, работа Менделя предполагала, что наследственность была частной, а не приобретенной, и что закономерности наследования многих признаков можно объяснить с помощью простых правил и соотношений. [19]
Важность работы Менделя не получила широкого понимания до 1900 года, после его смерти, когда Уго де Врис и другие ученые заново открыли его исследования. Уильям Бейтсон , сторонник работ Менделя, ввёл слово «генетика» в 1905 году. [20] [21] (Прилагательное «генетический» , происходящее от греческого слова «генезис» —γένεσις, «происхождение», предшествует существительному и впервые было использовано в биологическом смысле в 1860 году.) [22] Бейтсон выступал в качестве наставника, и ему в значительной степени помогли работы других ученых из Ньюнхэм-колледжа в Кембридже, в частности работы Бекки Сондерс , Норы Дарвин Барлоу и Мюриэл Уэлдейл Онслоу . [23] Бейтсон популяризировал использование слова «генетика» для описания изучения наследственности в своей инаугурационной речи на Третьей международной конференции по гибридизации растений в Лондоне в 1906 году. [24]
После повторного открытия работы Менделя ученые попытались определить, какие молекулы в клетке отвечают за наследование. В 1900 году Нетти Стивенс начала изучать мучного червя. [25] В течение следующих 11 лет она обнаружила, что у женщин есть только Х-хромосома, а у мужчин — как Х, так и Y-хромосома. [25] Ей удалось сделать вывод, что пол является хромосомным фактором и определяется мужчиной. [25] В 1911 году Томас Хант Морган утверждал, что гены находятся в хромосомах , основываясь на наблюдениях за сцепленной с полом мутацией белого глаза у плодовых мух . [26] В 1913 году его ученик Альфред Стертевант использовал явление генетического сцепления, чтобы показать, что гены расположены в хромосоме линейно. [27]
Молекулярная генетика
[ редактировать ]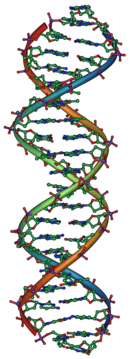
Хотя было известно, что гены существуют в хромосомах, хромосомы состоят как из белка , так и из ДНК, и ученые не знали, какой из двух ответственен за наследование. В 1928 году Фредерик Гриффит открыл феномен трансформации : мертвые бактерии могли передавать генетический материал для «трансформации» других еще живых бактерий. Шестнадцать лет спустя, в 1944 году, эксперимент Эйвери-Маклауда-Маккарти определил ДНК как молекулу, ответственную за трансформацию. [28] Роль ядра как хранилища генетической информации у эукариот была установлена Хеммерлингом в 1943 в его работе над одноклеточной водорослью Acetabularia . [29] Эксперимент Херши -Чейза в 1952 году подтвердил, что ДНК (а не белок) является генетическим материалом вирусов, заражающих бактерии, что предоставило дополнительные доказательства того, что ДНК является молекулой, ответственной за наследование. [30]
Джеймс Уотсон и Фрэнсис Крик определили структуру ДНК в 1953 году, используя по рентгеновской кристаллографии работы Розалинды Франклин и Мориса Уилкинса , которые показали, что ДНК имеет спиральную структуру (то есть имеет форму штопора). [31] [32] Их модель двойной спирали имела две нити ДНК с нуклеотидами, направленными внутрь, каждый из которых соответствовал комплементарному нуклеотиду на другой цепи, образуя что-то, похожее на ступеньки на извилистой лестнице. [33] Эта структура показала, что генетическая информация существует в последовательности нуклеотидов на каждой нити ДНК. Структура также предполагает простой метод репликации : если цепи разделены, для каждой из них можно реконструировать новые партнерские цепи на основе последовательности старой цепи. Именно это свойство придает ДНК ее полуконсервативную природу, когда одна цепь новой ДНК происходит от исходной родительской цепи. [34]
Хотя структура ДНК показала, как работает наследование, до сих пор не было известно, как ДНК влияет на поведение клеток. В последующие годы ученые пытались понять, как ДНК контролирует процесс производства белка . [35] Было обнаружено, что клетка использует ДНК в качестве матрицы для создания соответствующей информационной РНК — молекул с нуклеотидами, очень похожими на ДНК. Нуклеотидная последовательность информационной РНК используется для создания аминокислотной последовательности в белке; этот перевод между последовательностями нуклеотидов и последовательностями аминокислот известен как генетический код . [36]
С новым молекулярным пониманием наследственности произошел взрыв исследований. [37] Известная теория возникла у Томоко Охты в 1973 году, когда она внесла поправки в нейтральную теорию молекулярной эволюции , опубликовав почти нейтральную теорию молекулярной эволюции . В этой теории Охта подчеркивал важность естественного отбора и окружающей среды для скорости генетической эволюции . [38] с обрывом цепи, Одним из важных достижений стало секвенирование ДНК в 1977 году проведенное Фредериком Сэнгером . Эта технология позволяет ученым читать нуклеотидную последовательность молекулы ДНК. [39] В 1983 году Кэри Бэнкс Маллис разработал полимеразную цепную реакцию , обеспечивающую быстрый способ выделения и амплификации определенного участка ДНК из смеси. [40] Усилия проекта «Геном человека » Министерства энергетики НИЗ и параллельные частные усилия Celera Genomics привели к секвенированию генома человека в 2003 году. [41] [42]
Особенности наследования
[ редактировать ]Дискретное наследование и законы Менделя
[ редактировать ]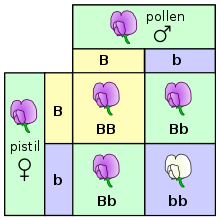
На самом фундаментальном уровне наследование в организмах происходит путем передачи дискретных наследственных единиц, называемых генами , от родителей к потомству. [43] Это свойство было впервые обнаружено Грегором Менделем, который изучал разделение наследственных признаков у растений гороха , показав, например, что цветки на одном растении были либо фиолетовыми, либо белыми, но никогда не были промежуточными между двумя цветами. Дискретные версии одного и того же гена, контролирующие наследственную внешность (фенотипы), называются аллелями . [19] [44]
В случае гороха, который является диплоидным видом, каждое отдельное растение имеет две копии каждого гена, по одной копии, унаследованной от каждого родителя. [45] Многие виды, включая человека, имеют такой тип наследования. Диплоидные организмы с двумя копиями одного и того же аллеля данного гена называются гомозиготными по этому локусу гена , а организмы с двумя разными аллелями данного гена называются гетерозиготными . Совокупность аллелей данного организма называется его генотипом , а наблюдаемые признаки организма — его фенотипом . Когда организмы гетерозиготны по гену, часто один аллель называют доминантным, поскольку его качества доминируют в фенотипе организма, а другой аллель называют рецессивным, поскольку его качества отступают и не наблюдаются. Некоторые аллели не обладают полным доминированием, а вместо этого имеют неполное доминирование за счет выражения промежуточного фенотипа или кодоминирование за счет экспрессии обеих аллелей одновременно. [46]
Когда пара организмов размножается половым путем , их потомство случайным образом наследует один из двух аллелей от каждого родителя. Эти наблюдения дискретного наследования и разделения аллелей известны как первый закон Менделя или закон сегрегации. Однако вероятность превосходства одного гена над другим может меняться из-за доминантных, рецессивных, гомозиготных или гетерозиготных генов. Например, Мендель обнаружил, что если скрещивать гетерозиготные организмы, шансы получить доминантный признак составляют 3:1. Настоящие генетики изучают и рассчитывают вероятности, используя теоретические вероятности, эмпирические вероятности, правило произведения, правило сумм и многое другое. [47]
Обозначения и диаграммы
[ редактировать ]
Генетики используют диаграммы и символы для описания наследования. Ген обозначается одной или несколькими буквами. Часто символ «+» используется для обозначения обычного немутантного аллеля гена. [48]
В экспериментах по оплодотворению и размножению (и особенно при обсуждении законов Менделя) родители называются поколением «P», а потомство - поколением «F1» (первое дочернее). Когда потомство F1 спаривается друг с другом, потомство называется поколением «F2» (второе дочернее поколение). Одной из распространенных диаграмм, используемых для прогнозирования результата скрещивания, является квадрат Пеннета . [49]
При изучении генетических заболеваний человека генетики часто используют таблицы родословных, чтобы представить наследование признаков. [50] Эти диаграммы отображают наследование признака в генеалогическом древе.
Множественные взаимодействия генов
[ редактировать ]
Организмы имеют тысячи генов, и у организмов, размножающихся половым путем, эти гены обычно сортируются независимо друг от друга. Это означает, что наследование аллеля желтого или зеленого цвета горошка не связано с наследованием аллелей белого или фиолетового цветка. Это явление, известное как « второй закон Менделя » или «закон независимого ассортимента», означает, что аллели разных генов перемешиваются между родителями, образуя потомство с множеством различных комбинаций. Разные гены часто взаимодействуют, оказывая влияние на один и тот же признак. У Голубоглазой Мэри ( Omphalodes verna ), например, существует ген с аллелями, определяющими окраску цветков: синюю или пурпурную. Однако другой ген контролирует, имеют ли цветы цвет вообще или они белые. Когда у растения есть две копии этого белого аллеля, его цветы белые — независимо от того, имеет ли первый ген синие или пурпурные аллели. Это взаимодействие между генами называется эпистазом , при этом второй ген эпистатический по отношению к первому. [51]
Многие признаки не являются отдельными признаками (например, фиолетовые или белые цветы), а являются непрерывными признаками (например, рост человека и цвет кожи ). Эти сложные черты являются продуктами многих генов. [52] Влияние этих генов в разной степени опосредовано средой, в которой находится организм. Степень, в которой гены организма способствуют развитию сложного признака, называется наследственностью . [53] Измерение наследственности признака является относительным: в более изменчивой среде среда оказывает большее влияние на общую изменчивость признака. Например, человеческий рост — это черта, имеющая сложные причины. В Соединенных Штатах наследственность составляет 89%. Однако в Нигерии, где люди имеют более вариативный доступ к хорошему питанию и медицинскому обслуживанию , наследственность роста составляет всего 62%. [54]
Молекулярная основа наследственности
[ редактировать ]ДНК и хромосомы
[ редактировать ]
Молекулярной ( ДНК основой генов является дезоксирибонуклеиновая кислота ). ДНК состоит из дезоксирибозы (молекулы сахара), фосфатной группы и основания (аминогруппы). Существует четыре типа оснований: аденин (А), цитозин (С), гуанин (G) и тимин (Т). Фосфаты образуют фосфодиэфирные связи с сахарами, образуя длинные фосфатно-сахарные основы. Базы специально соединяются вместе (T&A, C&G) между двумя магистральными цепями и образуют ступеньки на лестнице. Основания, фосфаты и сахара вместе образуют нуклеотид , который соединяется, образуя длинные цепи ДНК. [55] Генетическая информация существует в последовательности этих нуклеотидов, а гены существуют в виде участков последовательности вдоль цепи ДНК. [56] Эти цепи скручиваются в двойную а-спираль и обволакивают белки, называемые гистонами , которые обеспечивают структурную поддержку. ДНК, обернутая вокруг этих гистонов, называется хромосомами. [57] Вирусы иногда используют аналогичную молекулу РНК вместо ДНК в качестве генетического материала. [58]
ДНК обычно существует в виде двухцепочечной молекулы, свернутой в форме двойной спирали . Каждый нуклеотид в ДНК преимущественно спаривается со своим нуклеотидом-партнером на противоположной цепи: A соединяется с T, а C соединяется с G. Таким образом, в двухцепочечной форме каждая цепь эффективно содержит всю необходимую информацию, дублирующую ее партнерскую цепь. Такая структура ДНК является физической основой наследственности: репликация ДНК дублирует генетическую информацию путем расщепления цепей и использования каждой цепи в качестве матрицы для синтеза новой партнерской цепи. [59]

Гены расположены линейно вдоль длинных цепочек последовательностей пар оснований ДНК. У бактерий каждая клетка обычно содержит один кольцевой генофор , тогда как у эукариотических организмов (таких как растения и животные) ДНК расположена в нескольких линейных хромосомах. Эти нити ДНК часто бывают чрезвычайно длинными; Например, самая большая человеческая хромосома имеет около 247 миллионов пар оснований . длину [60] ДНК хромосомы связана со структурными белками, которые организуют, уплотняют и контролируют доступ к ДНК, образуя материал, называемый хроматином ; у эукариот хроматин обычно состоит из нуклеосом — сегментов ДНК, намотанных вокруг ядер белков- гистонов . [61] Полный набор наследственного материала в организме (обычно совокупность последовательностей ДНК всех хромосом) называется геномом .
ДНК чаще всего встречается в ядре клеток, но Рут Сейгер помогла открыть нехромосомные гены, находящиеся за пределами ядра. [62] У растений они часто обнаруживаются в хлоропластах, а у других организмов - в митохондриях. [62] Эти нехромосомные гены все еще могут передаваться любым партнером при половом размножении, и они контролируют множество наследственных характеристик, которые воспроизводятся и остаются активными на протяжении поколений. [62]
В то время как гаплоидные организмы имеют только одну копию каждой хромосомы, большинство животных и многие растения являются диплоидными , содержащими две копии каждой хромосомы и, следовательно, две копии каждого гена. Две аллели гена расположены в идентичных локусах двух гомологичных хромосом , причем каждая аллель унаследована от разных родителей. [45]
Многие виды имеют так называемые половые хромосомы , определяющие пол каждого организма. [63] У людей и многих других животных Y-хромосома содержит ген, запускающий развитие специфически мужских признаков. В ходе эволюции эта хромосома утратила большую часть своего содержимого, а также большую часть генов, в то время как Х-хромосома похожа на другие хромосомы и содержит множество генов. При этом Мэри Фрэнсис Лайон обнаружила, что во время размножения происходит инактивация Х-хромосомы, чтобы избежать передачи потомству вдвое большего количества генов. [64] Открытие Лиона привело к открытию Х-сцепленных заболеваний. [64]
Воспроизведение
[ редактировать ]
Когда клетки делятся, их полный геном копируется, и каждая дочерняя клетка наследует одну копию. Этот процесс, называемый митозом , является простейшей формой размножения и лежит в основе бесполого размножения. Бесполое размножение также может происходить у многоклеточных организмов, производящих потомство, унаследовавшее геном от одного родителя. Потомство, генетически идентичное своим родителям, называется клонами . [65]
Эукариотические организмы часто используют половое размножение для создания потомства, содержащего смесь генетического материала, унаследованного от двух разных родителей. В процессе полового размножения чередуются формы, содержащие единичные копии генома ( гаплоидные ) и двойные копии ( диплоидные ). [45] Гаплоидные клетки сливаются и объединяют генетический материал, образуя диплоидную клетку с парными хромосомами. Диплоидные организмы образуют гаплоиды путем деления без репликации своей ДНК с образованием дочерних клеток, которые случайным образом наследуют одну из каждой пары хромосом. Большинство животных и многие растения диплоидны на протяжении большей части своей жизни, при этом гаплоидная форма редуцируется до одноклеточных гамет , таких как сперматозоиды или яйцеклетки . [66]
Хотя бактерии не используют гаплоидный/диплоидный метод полового размножения, они обладают множеством способов приобретения новой генетической информации. Некоторые бактерии могут подвергаться конъюгации , передавая небольшой кольцевой фрагмент ДНК другой бактерии. [67] Бактерии также могут поглощать необработанные фрагменты ДНК, обнаруженные в окружающей среде, и интегрировать их в свои геномы — явление, известное как трансформация . [68] Эти процессы приводят к горизонтальному переносу генов , передавая фрагменты генетической информации между организмами, которые в противном случае не были бы связаны между собой. Естественная бактериальная трансформация происходит у многих видов бактерий и может рассматриваться как половой процесс переноса ДНК из одной клетки в другую клетку (обычно того же вида). [69] Трансформация требует воздействия многочисленных продуктов бактериальных генов , и ее основной адаптивной функцией, по-видимому, является восстановление в повреждений ДНК клетке-реципиенте. [69]
Рекомбинация и генетическое сцепление
[ редактировать ]
Диплоидная природа хромосом позволяет генам на разных хромосомах сортироваться независимо или отделяться от гомологичной пары во время полового размножения, при котором образуются гаплоидные гаметы. Таким образом, у потомства спаривающейся пары могут возникать новые комбинации генов. Гены в одной хромосоме теоретически никогда не рекомбинируются. Однако они это делают посредством клеточного процесса хромосомного кроссинговера . Во время скрещивания хромосомы обмениваются участками ДНК, эффективно перемещая аллели генов между хромосомами. [70] Этот процесс хромосомного кроссовера обычно происходит во время мейоза — серии клеточных делений, в результате которых образуются гаплоидные клетки. Мейотическая рекомбинация , особенно у микробных эукариот , по-видимому, выполняет адаптивную функцию восстановления повреждений ДНК. [69]
Первая цитологическая демонстрация кроссинговера была проведена Гарриет Крейтон и Барбарой МакКлинток в 1931 году. Их исследования и эксперименты на кукурузе предоставили цитологические доказательства генетической теории, согласно которой сцепленные гены в парных хромосомах действительно меняют места от одного гомолога к другому. [71]
Вероятность хромосомного кроссинговера, происходящего между двумя заданными точками хромосомы, связана с расстоянием между точками. На сколь угодно большом расстоянии вероятность скрещивания достаточно высока, поэтому наследование генов фактически некоррелировано. [72] Однако для генов, которые расположены ближе друг к другу, более низкая вероятность кроссинговера означает, что гены демонстрируют генетическое сцепление; аллели двух генов, как правило, наследуются вместе. Количество связей между рядом генов можно объединить, чтобы сформировать линейную карту сцепления , которая примерно описывает расположение генов вдоль хромосомы. [73]
Экспрессия генов
[ редактировать ]Генетический код
[ редактировать ]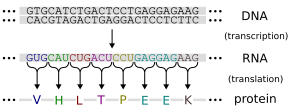
Гены выражают свой функциональный эффект посредством производства белков — молекул, отвечающих за большинство функций в клетке. Белки состоят из одной или нескольких полипептидных цепей, каждая из которых состоит из последовательности аминокислот . Последовательность ДНК гена используется для создания определенной аминокислотной последовательности . Этот процесс начинается с производства молекулы РНК с последовательностью, соответствующей последовательности ДНК гена, — процесс, называемый транскрипцией .
Эта молекула информационной РНК затем служит для производства соответствующей аминокислотной последовательности посредством процесса, называемого трансляцией . Каждая группа из трех нуклеотидов в последовательности, называемая кодоном , соответствует либо одной из двадцати возможных аминокислот в белке, либо команде завершения аминокислотной последовательности ; это соответствие называется генетическим кодом . [74] Поток информации однонаправлен: информация переносится из нуклеотидных последовательностей в аминокислотную последовательность белков, но никогда не переносится из белка обратно в последовательность ДНК — феномен, который Фрэнсис Крик назвал центральной догмой молекулярной биологии . [75]
Особая последовательность аминокислот приводит к уникальной трехмерной структуре этого белка, а трехмерная структура белков связана с их функциями. [76] [77] Некоторые из них представляют собой простые структурные молекулы, такие как волокна, образованные белком коллагеном . Белки могут связываться с другими белками и простыми молекулами, иногда действуя как ферменты, способствуя химическим реакциям внутри связанных молекул (без изменения структуры самого белка). Структура белка динамична; Белок гемоглобин принимает несколько разные формы, поскольку он облегчает захват, транспортировку и высвобождение молекул кислорода в крови млекопитающих. [ нужна ссылка ]
Разница в один нуклеотид в ДНК может вызвать изменение аминокислотной последовательности белка. Поскольку структуры белков являются результатом их аминокислотных последовательностей, некоторые изменения могут резко изменить свойства белка, дестабилизируя структуру или изменяя поверхность белка таким образом, что меняется его взаимодействие с другими белками и молекулами. Например, серповидно-клеточная анемия человека — это генетическое заболевание , которое возникает в результате разницы в одном основании в кодирующей области β-глобинового участка гемоглобина, вызывая изменение одной аминокислоты, которое изменяет физические свойства гемоглобина. [78] Серповидноклеточные версии гемоглобина прилипают друг к другу, образуя волокна, которые искажают форму эритроцитов, несущих белок. Эти серповидные клетки больше не могут плавно проходить через кровеносные сосуды , имея тенденцию закупориваться или разрушаться, что приводит к медицинским проблемам, связанным с этим заболеванием. [ нужна ссылка ]
Некоторые последовательности ДНК транскрибируются в РНК, но не транслируются в белковые продукты — такие молекулы РНК называются некодирующими РНК . В некоторых случаях эти продукты складываются в структуры, которые участвуют в важнейших функциях клетки (например, рибосомальная РНК и транспортная РНК ). РНК также может оказывать регуляторное действие посредством гибридизационного взаимодействия с другими молекулами РНК (такими как микроРНК ). [ нужна ссылка ]
Природа и воспитание
[ редактировать ]
Хотя гены содержат всю информацию, которую организм использует для функционирования, окружающая среда играет важную роль в определении окончательных фенотипов, которые проявляет организм. Фраза « природа и воспитание » относится к этим взаимодополняющим отношениям. Фенотип организма зависит от взаимодействия генов и окружающей среды. Интересным примером является окрас шерсти сиамской кошки . В этом случае температура тела кошки играет роль окружающей среды. Гены кошки кодируют темную шерсть, поэтому клетки, производящие шерсть у кошки, вырабатывают клеточные белки, в результате чего волосы становятся темными. Но эти белки, производящие темную шерсть, чувствительны к температуре (т.е. имеют мутацию, вызывающую температурную чувствительность) и денатурируют в условиях более высокой температуры, неспособные производить пигмент темной шерсти в тех местах, где у кошки более высокая температура тела. Однако в условиях низкой температуры структура белка стабильна и нормально производит пигмент темных волос. Белок остается функциональным на более холодных участках кожи, таких как ноги, уши, хвост и морда, поэтому на конечностях у кошки темная шерсть. [79]
Окружающая среда играет важную роль в развитии генетического заболевания человека фенилкетонурии . Мутация, вызывающая фенилкетонурию, нарушает способность организма расщеплять аминокислоту фенилаланин , вызывая токсичное накопление промежуточной молекулы, которая, в свою очередь, вызывает серьезные симптомы прогрессирующей умственной отсталости и судорог. Однако, если человек с мутацией фенилкетонурии соблюдает строгую диету, избегающую этой аминокислоты, он остается нормальным и здоровым. [80]
Распространенный метод определения того, как гены и окружающая среда («природа и воспитание») способствуют фенотипу, включает изучение однояйцевых и разнояйцевых близнецов или других братьев и сестер, родившихся от нескольких детей . [81] Идентичные братья и сестры генетически одинаковы, поскольку происходят из одной и той же зиготы. Между тем, разнояйцевые близнецы генетически отличаются друг от друга так же, как и обычные братья и сестры. Сравнивая, как часто определенное заболевание возникает у пары однояйцевых близнецов, с тем, как часто оно встречается у пары разнояйцевых близнецов, ученые могут определить, вызвано ли это расстройство генетическими или послеродовыми факторами окружающей среды. Одним из известных примеров было исследование четверных генайнов , которые были идентичными четвероногими, у которых была диагностирована шизофрения . [82]
Генная регуляция
[ редактировать ]Геном данного организма содержит тысячи генов, но не все эти гены должны быть активными в любой данный момент. Ген экспрессируется, когда он транскрибируется в мРНК, и существует множество клеточных методов контроля экспрессии генов, при которых белки производятся только тогда, когда это необходимо клетке. Факторы транскрипции — это регуляторные белки, которые связываются с ДНК, либо стимулируя, либо ингибируя транскрипцию гена. [83] Например, в геноме бактерий Escherichia coli существует ряд генов, необходимых для синтеза аминокислоты триптофана . Однако когда триптофан уже доступен клетке, эти гены для синтеза триптофана больше не нужны. Присутствие триптофана напрямую влияет на активность генов — молекулы триптофана связываются с репрессором триптофана (фактором транскрипции), изменяя структуру репрессора так, что репрессор связывается с генами. Репрессор триптофана блокирует транскрипцию и экспрессию генов, тем самым создавая по отрицательной обратной связи . регуляцию процесса синтеза триптофана [84]
Различия в экспрессии генов особенно очевидны у многоклеточных организмов , где все клетки содержат один и тот же геном, но имеют совершенно разные структуры и поведение из-за экспрессии разных наборов генов. Все клетки многоклеточного организма происходят из одной клетки, дифференцируясь в различные типы клеток в ответ на внешние и межклеточные сигналы и постепенно устанавливая разные модели экспрессии генов, создавая различное поведение. Поскольку ни один ген не отвечает за развитие структур внутри многоклеточных организмов, эти закономерности возникают в результате сложных взаимодействий между многими клетками. [ нужна ссылка ]
Внутри эукариот существуют структурные особенности хроматина , которые влияют на транскрипцию генов, часто в виде модификаций ДНК и хроматина, которые стабильно наследуются дочерними клетками. [85] Эти особенности называются « эпигенетическими », поскольку они существуют «поверх» последовательности ДНК и наследуются от одного поколения клеток к другому. Из-за эпигенетических особенностей разные типы клеток, выращенные в одной и той же среде, могут сохранять совершенно разные свойства. Хотя эпигенетические особенности, как правило, динамичны в ходе развития, некоторые из них, например феномен парамутации , наследуются от нескольких поколений и существуют как редкие исключения из общего правила, согласно которому ДНК является основой наследования. [86]
Генетические изменения
[ редактировать ]Мутации
[ редактировать ]
В процессе репликации ДНК иногда возникают ошибки при полимеризации второй цепи. Эти ошибки, называемые мутациями, могут повлиять на фенотип организма, особенно если они происходят в кодирующей последовательность белка гена. Частота ошибок обычно очень низкая — 1 ошибка на каждые 10–100 миллионов оснований — из-за способности ДНК-полимеразы «корректировать» . [87] [88] Процессы, которые увеличивают скорость изменений в ДНК, называются мутагенными : мутагенные химические вещества способствуют ошибкам в репликации ДНК, часто вмешиваясь в структуру спаривания оснований, в то время как УФ-излучение вызывает мутации, вызывая повреждение структуры ДНК. [89] Химическое повреждение ДНК также происходит естественным путем, и клетки используют механизмы восстановления ДНК для устранения несовпадений и разрывов. Однако восстановление не всегда восстанавливает исходную последовательность. Особенно важным источником повреждений ДНК являются активные формы кислорода. [90] вырабатываются клеточным аэробным дыханием и могут приводить к мутациям. [91]
У организмов, которые используют хромосомный кроссовер для обмена ДНК и рекомбинации генов, ошибки выравнивания во время мейоза также могут вызывать мутации. Ошибки при кроссинговере особенно вероятны, когда сходные последовательности заставляют партнерские хромосомы принимать ошибочное выравнивание; это делает некоторые области генома более склонными к таким мутациям. Эти ошибки создают большие структурные изменения в последовательности ДНК — дупликации , инверсии , делеции целых областей — или случайный обмен целыми частями последовательностей между разными хромосомами, хромосомную транслокацию . [92]

Естественный отбор и эволюция
[ редактировать ]Мутации изменяют генотип организма, и иногда это приводит к появлению разных фенотипов. организма Большинство мутаций мало влияют на фенотип, здоровье или репродуктивную способность . [93] Мутации, которые действительно оказывают влияние, обычно вредны, но иногда некоторые из них могут быть полезны. [94] Исследования на мухе Drosophila melanogaster показывают, что если мутация изменяет белок, вырабатываемый геном, около 70 процентов этих мутаций вредны, а остальные либо нейтральны, либо слабо полезны. [95]

Популяционная генетика изучает распределение генетических различий внутри популяций и то, как это распределение меняется с течением времени. [96] На изменения частоты аллеля в популяции в основном влияет естественный отбор , при котором данный аллель обеспечивает организму селективное или репродуктивное преимущество. [97] а также другие факторы, такие как мутации , генетический дрейф , генетический автостоп , [98] искусственный отбор и миграция . [99]
На протяжении многих поколений геномы организмов могут существенно меняться, что приводит к эволюции. В процессе, называемом адаптацией , отбор полезных мутаций может привести к тому, что вид эволюционирует в формы, более способные выживать в окружающей среде. [100] Новые виды образуются в процессе видообразования , часто вызванного географическим разделением, которое не позволяет популяциям обмениваться генами друг с другом. [101]
Сравнивая гомологию геномов разных видов, можно рассчитать эволюционное расстояние между ними и определить, когда они могли разойтись . Генетические сравнения обычно считаются более точным методом характеристики родства между видами, чем сравнение фенотипических характеристик. Эволюционные расстояния между видами можно использовать для формирования эволюционных деревьев ; эти деревья представляют собой общее происхождение и дивергенцию видов с течением времени, хотя они не показывают передачу генетического материала между несвязанными видами (известный как горизонтальный перенос генов и наиболее распространенный у бактерий). [102]
Исследования и технологии
[ редактировать ]Модельные организмы
[ редактировать ]
Хотя первоначально генетики изучали наследование у самых разных организмов, круг изучаемых видов сузился. Одна из причин заключается в том, что, когда уже проводятся серьезные исследования данного организма, новые исследователи с большей вероятностью выберут его для дальнейшего изучения, и поэтому в конечном итоге несколько модельных организмов стали основой для большинства генетических исследований. Общие темы исследований в области генетики модельных организмов включают изучение регуляции генов и участие генов в развитии и раке . Организмы были выбраны отчасти из соображений удобства: короткое время генерации и простота генетических манипуляций сделали некоторые организмы популярными инструментами генетических исследований. Широко используемые модельные организмы включают кишечную бактерию Escherichia coli , растение Arabidopsis thaliana , пекарские дрожжи ( Saccharomyces cerevisiae ), нематоду Caenorhabditis elegans , обыкновенную плодовую мушку ( Drosophila melanogaster ), рыбку данио ( Danio rerio ) и обыкновенную домовую мышь ( Dario rerio). Мускулистые мышцы ). [103]
Лекарство
[ редактировать ]
Медицинская генетика стремится понять, как генетические вариации связаны со здоровьем и болезнями человека. [104] При поиске неизвестного гена, который может быть вовлечен в заболевание, исследователи обычно используют генетическую связь и диаграммы генетической родословной , чтобы найти место в геноме, связанное с заболеванием. На популяционном уровне исследователи используют менделевскую рандомизацию для поиска участков генома, связанных с заболеваниями. Этот метод особенно полезен для мультигенных признаков, которые четко не определены одним геном. [105] После обнаружения гена-кандидата дальнейшие исследования часто проводятся на соответствующих (или гомологичных ) генах модельных организмов. Помимо изучения генетических заболеваний, возросшая доступность методов генотипирования привела к развитию фармакогенетики : изучению того, как генотип может влиять на реакцию на лекарства. [106]
Люди различаются по наследственной склонности к развитию рака , а рак является генетическим заболеванием. Процесс развития рака в организме представляет собой совокупность событий. Мутации иногда происходят внутри клеток организма во время их деления. Хотя эти мутации не будут унаследованы ни одним потомком, они могут влиять на поведение клеток, иногда заставляя их расти и делиться чаще. Существуют биологические механизмы, которые пытаются остановить этот процесс; Неправильно делящимся клеткам передаются сигналы, которые должны вызвать гибель клеток , но иногда возникают дополнительные мутации, которые заставляют клетки игнорировать эти сообщения. внутренний процесс естественного отбора В организме происходит , и в конечном итоге мутации накапливаются внутри клеток, способствуя их собственному росту, создавая раковую опухоль , которая растет и проникает в различные ткани организма. Обычно клетка делится только в ответ на сигналы, называемые факторами роста , и прекращает рост после контакта с окружающими клетками и в ответ на сигналы, ингибирующие рост. Обычно затем он делится ограниченное число раз и умирает, оставаясь в пределах эпителия , где он не может мигрировать в другие органы. Чтобы стать раковой клеткой, клетка должна накопить мутации в ряде генов (от трех до семи). Раковая клетка может делиться без фактора роста и игнорирует ингибирующие сигналы. Кроме того, он бессмертен и может расти бесконечно даже после контакта с соседними клетками. Он может выйти из эпителия и, в конечном итоге, из первичной опухоли . Затем сбежавшая клетка может пересечь эндотелий кровеносного сосуда и попасть в кровоток, чтобы колонизировать новый орган, образуя смертельные метастазы . Хотя существует некоторая генетическая предрасположенность к небольшой части случаев рака, основная часть обусловлена набором новых генетических мутаций, которые первоначально появляются и накапливаются в одной или небольшом количестве клеток, которые делятся с образованием опухоли и не передаются другим клеткам. потомство ( соматические мутации ). Наиболее частыми мутациями являются утрата функции белка р53 , супрессора опухоли или пути р53, а также мутации усиления функции в Белки Ras или другие онкогены . [107] [108]
Методы исследования
[ редактировать ]
ДНК можно манипулировать в лаборатории. Ферменты рестрикции — это обычно используемые ферменты, которые разрезают ДНК по определенным последовательностям, образуя предсказуемые фрагменты ДНК. [109] Фрагменты ДНК можно визуализировать с помощью гель-электрофореза , который разделяет фрагменты в зависимости от их длины. [110]
Использование ферментов лигирования позволяет соединять фрагменты ДНК. Связывая («лигируя») фрагменты ДНК из разных источников, исследователи могут создавать рекомбинантную ДНК , ДНК, часто связанную с генетически модифицированными организмами . Рекомбинантная ДНК обычно используется в контексте плазмид : коротких кольцевых молекул ДНК с несколькими генами на них. В процессе, известном как молекулярное клонирование , исследователи могут амплифицировать фрагменты ДНК, вставляя плазмиды в бактерии, а затем культивируя их на чашках с агаром (чтобы изолировать клоны бактериальных клеток ). «Клонирование» также может относиться к различным способам создания клонированных («клональных») организмов. [111]
ДНК также можно амплифицировать с помощью процедуры, называемой полимеразной цепной реакцией (ПЦР). [112] Используя специфические короткие последовательности ДНК, ПЦР может изолировать и экспоненциально амплифицировать целевой участок ДНК. Поскольку ПЦР может амплифицироваться на очень небольших количествах ДНК, ее также часто используют для обнаружения присутствия определенных последовательностей ДНК. [113] [114]
Секвенирование ДНК и геномика
[ редактировать ]Секвенирование ДНК, одна из наиболее фундаментальных технологий, разработанных для изучения генетики, позволяет исследователям определять последовательность нуклеотидов во фрагментах ДНК. Техника секвенирования обрыва цепи , разработанная в 1977 году командой под руководством Фредерика Сэнгера , до сих пор регулярно используется для секвенирования фрагментов ДНК. Используя эту технологию, исследователи смогли изучить молекулярные последовательности, связанные со многими заболеваниями человека. [115]
Поскольку секвенирование стало менее дорогостоящим, исследователи секвенировали геномы многих организмов, используя процесс, называемый сборкой генома , который использует вычислительные инструменты для сшивания последовательностей из множества различных фрагментов. [116] Эти технологии использовались для секвенирования генома человека в рамках проекта «Геном человека», завершенного в 2003 году. [41] Новые высокопроизводительные технологии секвенирования резко снижают стоимость секвенирования ДНК, и многие исследователи надеются снизить стоимость повторного секвенирования человеческого генома до тысячи долларов. [117]
Секвенирование следующего поколения (или высокопроизводительное секвенирование) появилось из-за постоянно растущего спроса на недорогое секвенирование. Эти технологии секвенирования позволяют создавать потенциально миллионы последовательностей одновременно. [118] [119] Большое количество доступных данных о последовательностях привело к созданию раздела геномики — исследования, в котором используются вычислительные инструменты для поиска и анализа закономерностей в полных геномах организмов. Геномику можно также считать подобластью биоинформатики , которая использует вычислительные подходы для анализа больших наборов биологических данных . Общая проблема в этих областях исследований заключается в том, как управлять и обмениваться данными, которые касаются людей и информации, позволяющей идентифицировать личность . [ нужна ссылка ]
Общество и культура
[ редактировать ]19 марта 2015 года группа ведущих биологов призвала запретить во всем мире клиническое использование методов, в частности использования CRISPR и цинкового пальца , для редактирования генома человека таким образом, чтобы он мог передаваться по наследству. [120] [121] [122] [123] В апреле 2015 года китайские исследователи сообщили о результатах фундаментального исследования по редактированию ДНК нежизнеспособных эмбрионов человека с помощью CRISPR. [124] [125]
См. также
[ редактировать ]- Размер бактериального генома
- Криоконсервация генетических ресурсов животных
- Евгеника
- Эмбриология
- Генетическое заболевание
- Генетическое разнообразие
- Генная инженерия
- Генетическое улучшение
- Глоссарий генетики (M−Z)
- Указатель статей по генетике
- Медицинская генетика
- Молекулярные инструменты для изучения генов
- Нейроэпигенетика
- Очерк генетики
- Хронология истории генетики
- Генетические ресурсы растений
Ссылки
[ редактировать ]- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Генетика и организм: Введение» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Хартл Д., Джонс Э. (2005)
- ^ «определение генетики» . www.dictionary.com . Проверено 25 октября 2018 г.
- ^ «Генетикос (γενετ-ικός)» . Генри Джордж Лидделл, Роберт Скотт, Греко-английский лексикон . Цифровая библиотека Персея, Университет Тафтса. Архивировано из оригинала 15 июня 2010 года . Проверено 20 февраля 2012 г.
- ^ «Бытие (γένεσις)» . Генри Джордж Лидделл, Роберт Скотт, Греко-английский лексикон . Цифровая библиотека Персея, Университет Тафтса. Архивировано из оригинала 15 июня 2010 года . Проверено 20 февраля 2012 г.
- ^ «Генетический» . Интернет-словарь этимологии . Архивировано из оригинала 23 августа 2011 года . Проверено 20 февраля 2012 г.
- ^ Наука: полное визуальное руководство . Пингвин. 2009. с. 362. ИСБН 978-0-7566-6490-9 .
- ^ Почаи П., Сантьяго-Блей Дж.А. (июль 2022 г.). «Темы биологической наследственности в овцеводстве начала девятнадцатого века, раскрытые Дж. М. Эренфельсом» . Гены . 13 (8): 1311. doi : 10.3390/genes13081311 . ПМЦ 9332421 . ПМИД 35893050 .
- ^ Вейлинг Ф. (июль 1991 г.). «Историческое исследование: Иоганн Грегор Мендель 1822-1884». Американский журнал медицинской генетики . 40 (1): 1–25, обсуждение 26. doi : 10.1002/ajmg.1320400103 . ПМИД 1887835 .
- ^ Перейти обратно: а б Почаи П., Сантьяго-Блей Дж.А. (октябрь 2021 г.). «Принципы и биологические концепции наследственности до Менделя» . Биология Директ . 16 (1): 19. дои : 10.1186/s13062-021-00308-4 . ПМЦ 8532317 . ПМИД 34674746 .
 Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0. Архивировано 16 октября 2017 г. на Wayback Machine .
Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0. Архивировано 16 октября 2017 г. на Wayback Machine . - ^ Сабо А.Т., Почаи П. (июнь 2019 г.). «Появление генетики от овец Фештетича через горох Менделя до цыплят Бейтсона». Журнал генетики . 98 (2): 63. doi : 10.1007/s12041-019-1108-z . hdl : 10138/324962 . ПМИД 31204695 . S2CID 174803150 .
- ^ Почаи П., Белл Н., Хивёнен Дж. (январь 2014 г.). «Имре Фештетич и Моравское общество овцеводов: забытая «исследовательская сеть» Менделя » . ПЛОС Биология . 12 (1): e1001772. дои : 10.1371/journal.pbio.1001772 . ПМЦ 3897355 . ПМИД 24465180 .
- ^ Почаи П. (2022). Наследственность до Менделя: Фестетика и вопрос овечьей шерсти в Центральной Европе . Бока-Ратон, Флорида: CRC Press. п. 113. ИСБН 978-1-032-02743-2 . Проверено 30 августа 2022 г.
- ^ Почаи П., Сантьяго-Блей Х.А., Секерак Дж., Бариска И., Сабо А.Т. (октябрь 2022 г.). «Овца Мимуш и призрак инбридинга: историческая основа органических и генетических законов Фештетича за четыре десятилетия до экспериментов Менделя с горохом» . Журнал истории биологии . 55 (3): 495–536. дои : 10.1007/s10739-022-09678-5 . ПМЦ 9668798 . ПМИД 35670984 . S2CID 249433049 .
- ^ Почаи П., Сантьяго-Блей Дж.А. (2022). «Отколоть старый блок: поколение, развитие и наследственные концепции наследственности» . Границы генетики . 13 : 814436. doi : 10.3389/fgene.2022.814436 . ПМЦ 8959437 . ПМИД 35356423 .
- ^ Гамильтон Х (2011). Популяционная генетика . Джорджтаунский университет. п. 26. ISBN 978-1-4443-6245-9 .
- ^ Ламарк, JB (2008). В Британской энциклопедии . Получено из онлайн-энциклопедии Британской энциклопедии. Архивировано 14 апреля 2020 года в Wayback Machine 16 марта 2008 года.
- ^ Питер Дж. Боулер , Менделевская революция: чрезвычайная ситуация наследственных концепций в современной науке и обществе (Балтимор: издательство Университета Джонса Хопкинса, 1989): главы 2 и 3.
- ^ Перейти обратно: а б Блумберг РБ. «Работа Менделя на английском языке» . Архивировано из оригинала 13 января 2016 года.
- ^ генетика, н. , Оксфордский словарь английского языка , 3-е изд.
- ^ Бейтсон В. «Письмо Уильяма Бейтсона Алану Седжвику в 1905 году» . Центр Джона Иннеса. Архивировано из оригинала 13 октября 2007 года . Проверено 15 марта 2008 г. Письмо было адресовано Адаму Седжвику, зоологу и «читателю по морфологии животных» в Тринити-колледже Кембриджа.
- ^ генетический, прил. , Оксфордский словарь английского языка, 3-е изд.
- ^ Ричмонд, М.Л. (ноябрь 2007 г.). «Возможности для женщин в ранней генетике» . Обзоры природы. Генетика . 8 (11): 897–902. дои : 10.1038/nrg2200 . ПМИД 17893692 . S2CID 21992183 . Архивировано из оригинала 16 мая 2008 года.
- ^ Бейтсон В. (1907). «Прогресс генетических исследований». В Уилксе, В. (ред.). Отчет Третьей Международной конференции по генетике 1906 г.: Гибридизация (скрещивание родов или видов), скрещивание сортов и общая селекция растений . Лондон: Королевское садоводческое общество. : Первоначально называвшаяся «Международная конференция по гибридизации и селекции растений», название было изменено в результате выступления Бейтсона. Видеть: Кок А.Г., Форсдайк Д.Р. (2008). Цените свои исключения: наука и жизнь Уильяма Бейтсона . Спрингер. п. 248 . ISBN 978-0-387-75687-5 .
- ^ Перейти обратно: а б с «Нетти Стивенс: первооткрывательница половых хромосом» . Возбудимый . Природное образование . Проверено 8 июня 2020 г.
- ^ Мур Дж. А. (1983). «Томас Хант Морган – Генетик». Интегративная и сравнительная биология . 23 (4): 855–865. дои : 10.1093/icb/23.4.855 .
- ^ Стертевант А.Х. (1913). «Линейное расположение шести сцепленных с полом факторов у дрозофилы, как показано способом их ассоциации» (PDF) . Журнал экспериментальной биологии . 14 (1): 43–59. Бибкод : 1913JEZ....14...43S . CiteSeerX 10.1.1.37.9595 . дои : 10.1002/jez.1400140104 . S2CID 82583173 . Архивировано (PDF) из оригинала 27 февраля 2008 г.
- ^ Эйвери ОТ, Маклауд СМ, Маккарти М (февраль 1944 г.). «ИССЛЕДОВАНИЕ ХИМИЧЕСКОЙ ПРИРОДЫ ВЕЩЕСТВА, ВЫЗЫВАЮЩЕГО ТРАНСФОРМАЦИЮ ПНЕВМОКОККА: ИНДУКЦИЯ ТРАНСФОРМАЦИИ ФРАКЦИЕЙ ДЕЗОКСИРИБОНКЛЕИНОВОЙ КИСЛОТЫ, ВЫДЕЛЕННОЙ ИЗ ПНЕВМОКОККА III ТИПА» . Журнал экспериментальной медицины . 79 (2): 137–158. дои : 10.1084/jem.79.2.137 . ПМК 2135445 . ПМИД 19871359 . Перепечатка: Эйвери ОТ, Маклауд СМ, Маккарти М (февраль 1979 г.). «Исследование химической природы вещества, индуцирующего трансформацию пневмококков типа. Индукция трансформации фракцией дезоксирибонуклеиновой кислоты, выделенной из пневмококка III типа» . Журнал экспериментальной медицины . 149 (2): 297–326. дои : 10.1084/jem.149.2.297 . ПМК 2184805 . ПМИД 33226 .
- ^ Ханна П. (2008). Клеточная и молекулярная биология . ИК Интернэшнл Пвт Лтд. 221. ИСБН 978-81-89866-59-4 .
- ^ Херши А.Д., Чейз М. (май 1952 г.). «Независимые функции вирусного белка и нуклеиновой кислоты в росте бактериофага» . Журнал общей физиологии . 36 (1): 39–56. дои : 10.1085/jgp.36.1.39 . ПМК 2147348 . ПМИД 12981234 .
- ^ Джадсон Х (1979). Восьмой день творения: творцы революции в биологии . Лабораторный пресс Колд-Спринг-Харбор. стр. 51–169. ISBN 978-0-87969-477-7 .
- ^ Уотсон Дж. Д., Крик Ф. Х. (апрель 1953 г.). «Молекулярная структура нуклеиновых кислот; структура нуклеиновой кислоты дезоксирибозы» (PDF) . Природа . 171 (4356): 737–738. Бибкод : 1953Natur.171..737W . дои : 10.1038/171737a0 . ПМИД 13054692 . S2CID 4253007 . Архивировано (PDF) из оригинала 4 февраля 2007 г.
- ^ Уотсон Дж. Д., Крик Ф. Х. (май 1953 г.). «Генетическое значение структуры дезоксирибонуклеиновой кислоты» (PDF) . Природа . 171 (4361): 964–967. Бибкод : 1953Natur.171..964W . дои : 10.1038/171964b0 . ПМИД 13063483 . S2CID 4256010 . Архивировано (PDF) из оригинала 21 июня 2003 г.
- ^ Стратманн С.А., ван Ойен А.М. (февраль 2014 г.). «Репликация ДНК на уровне одиночной молекулы» (PDF) . Обзоры химического общества . 43 (4): 1201–1220. дои : 10.1039/c3cs60391a . ПМИД 24395040 . S2CID 205856075 . Архивировано (PDF) из оригинала 6 июля 2017 года.
- ^ Фредерик Б. (2010). Управляющая наука: методология и организация исследований . Спрингер. п. 76. ИСБН 978-1-4419-7488-4 .
- ^ Райс С.А. (2009). Энциклопедия эволюции . Издательство информационной базы. п. 134. ИСБН 978-1-4381-1005-9 .
- ^ Саркар С (1998). Генетика и редукционизм . Издательство Кембриджского университета. п. 140. ИСБН 978-0-521-63713-8 .
- ^ Охта Т. (ноябрь 1973 г.). «Слегка вредные мутантные замены в эволюции». Природа . 246 (5428): 96–98. Бибкод : 1973Natur.246...96O . дои : 10.1038/246096a0 . ПМИД 4585855 . S2CID 4226804 .
- ^ Сэнгер Ф., Никлен С., Коулсон А.Р. (декабрь 1977 г.). «Секвенирование ДНК с ингибиторами обрыва цепи» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (12): 5463–5467. Бибкод : 1977PNAS...74.5463S . дои : 10.1073/pnas.74.12.5463 . ПМК 431765 . ПМИД 271968 .
- ^ Сайки Р.К., Шарф С., Фалуна Ф., Муллис К.Б., Хорн Г.Т., Эрлих Х.А. и др. (декабрь 1985 г.). «Ферментативная амплификация геномных последовательностей бета-глобина и анализ сайтов рестрикции для диагностики серповидноклеточной анемии». Наука . 230 (4732): 1350–1354. Бибкод : 1985Sci...230.1350S . дои : 10.1126/science.2999980 . ПМИД 2999980 .
- ^ Перейти обратно: а б «Информация о проекте генома человека» . Проект «Геном человека». Архивировано из оригинала 15 марта 2008 года . Проверено 15 марта 2008 г.
- ^ «Последовательность генома человека». Наука . 291 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Способы наследования: Введение» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Опыты Менделя» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Перейти обратно: а б с Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Менделевская генетика в жизненных циклах эукариот» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Взаимодействия между аллелями одного гена» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ «Вероятности в генетике (статья)» . Ханская академия . Проверено 28 сентября 2022 г.
- ^ Чейни Р.В. «Генетическая нотация» . Университет Кристофера Ньюпорта. Архивировано из оригинала 3 января 2008 года . Проверено 18 марта 2008 г.
- ^ Мюллер-Вилле С., Паролини Дж. (9 декабря 2020 г.). «Квадраты Пеннета и гибридные кресты: как менделисты научились своему ремеслу по книге». Обучение по книге: Учебники и справочники по истории науки . Темы BJHS. Том. 5. Британское общество истории науки / Издательство Кембриджского университета . стр. 149–165. дои : 10.1017/bjt.2020.12 . S2CID 229344415 . Архивировано из оригинала 29 марта 2021 года . Проверено 29 марта 2021 г.
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Генетика человека» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Взаимодействие генов и модифицированные дигибридные соотношения» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Майе Р. (июнь 2005 г.). «Картирование новых границ: сложные генетические нарушения» . Журнал клинических исследований . 115 (6): 1404–1407. дои : 10.1172/JCI25421 . ПМК 1137013 . ПМИД 15931374 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Количественная оценка наследственности» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Люк А., Го X, Адейемо А.А., Уилкс Р., Форрестер Т., Лоу В. и др. (июль 2001 г.). «Наследственность черт, связанных с ожирением, среди нигерийцев, ямайцев и чернокожих жителей США» . Международный журнал ожирения и связанных с ним метаболических нарушений . 25 (7): 1034–1041. дои : 10.1038/sj.ijo.0801650 . ПМИД 11443503 .
- ^ Урри Л., Кейн М., Вассерман С., Минорски П., Рис Дж., Кэмпбелл Н. «Биология Кэмпбелла» . plus.pearson.com . Проверено 28 сентября 2022 г.
- ^ Пирсон Х (май 2006 г.). «Генетика: что такое ген?» . Природа . 441 (7092): 398–401. Бибкод : 2006Natur.441..398P . дои : 10.1038/441398a . ПМИД 16724031 . S2CID 4420674 .
- ^ «Гистон» . Genome.gov . Проверено 28 сентября 2022 г.
- ^ Прескотт Л.М., Харли Дж.П., Кляйн Д.А. (1996). Микробиология (3-е изд.). Вм. К. Браун. п. 343. ИСБН 0-697-21865-1 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Механизм репликации ДНК» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Грегори С.Г., Барлоу К.Ф., Маклей К.Э., Каул Р., Сварбрек Д., Данэм А. и др. (май 2006 г.). «Последовательность ДНК и биологическая аннотация хромосомы 1 человека» . Природа . 441 (7091): 315–321. Бибкод : 2006Natur.441..315G . дои : 10.1038/nature04727 . ПМИД 16710414 .
- ^ Альбертс и др. (2002), II.4. ДНК и хромосомы: хромосомная ДНК и ее упаковка в хроматиновые волокна. Архивировано 18 октября 2007 г. в Wayback Machine.
- ^ Перейти обратно: а б с «Рут Сагер» . Британская энциклопедия . Проверено 8 июня 2020 г.
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Половые хромосомы и наследование, сцепленное с полом» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Перейти обратно: а б Растан С (февраль 2015 г.). «Мэри Ф. Лайон (1925–2014)» . Природа . 518 (7537). Springer Nature Limited: 36. Бибкод : 2015Natur.518...36R . дои : 10.1038/518036а . ПМИД 25652989 . S2CID 4405984 .
- ^ «клон» . Словарь Мерриам-Вебстера . Проверено 13 ноября 2023 г.
- ^ «Гаплоид» . www.genome.gov . Проверено 10 февраля 2024 г.
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Бактериальная конъюгация» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Бактериальная трансформация» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Перейти обратно: а б с Бернштейн Х., Бернштейн К., Мишод Р.Э. (январь 2018 г.). «Пол у микробных патогенов» . Инфекция, генетика и эволюция . 57 : 8–25. Бибкод : 2018InfGE..57....8B . дои : 10.1016/j.meegid.2017.10.024 . ПМИД 29111273 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гельбар, ред. (2000). «Природа кроссинговера» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Крейтон Х.Б., МакКлинток Б. (август 1931 г.). «Корреляция цитологического и генетического кроссинговера у Zea Mays» . Труды Национальной академии наук Соединенных Штатов Америки . 17 (8): 492–497. Бибкод : 1931PNAS...17..492C . дои : 10.1073/pnas.17.8.492 . ПМЦ 1076098 . ПМИД 16587654 .
- ^ Стауб Дж. Э. (1994). Кроссовер: концепции и приложения в генетике, эволюции и селекции . Университет Висконсина Пресс. п. 55. ИСБН 978-0-299-13564-5 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гельбар, ред. (2000). «Карты связей» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Берг Дж.М., Тимочко Дж.Л., Страйер Л., Кларк Н.Д. (2002). «I. 5. ДНК, РНК и поток генетической информации: аминокислоты кодируются группами из трех оснований, начиная с фиксированной точки» . Биохимия (5-е изд.). Нью-Йорк: WH Freeman and Company. Архивировано из оригинала 11 апреля 2006 года.
- ^ Крик Ф. (август 1970 г.). «Центральная догма молекулярной биологии» (PDF) . Природа . 227 (5258): 561–563. Бибкод : 1970Natur.227..561C . дои : 10.1038/227561a0 . ПМИД 4913914 . S2CID 4164029 . Архивировано (PDF) из оригинала 15 февраля 2006 г.
- ^ Альбертс и др. (2002), I.3. Белки: форма и структура белков. Архивировано 1 января 2023 года в Wayback Machine.
- ^ Альбертс и др. (2002), I.3. Белки: функция белка. Архивировано 25 апреля 2006 г. в Wayback Machine.
- ^ «Как серповидноклеточная клетка вызывает заболевание?» . Бригам и женская больница: Информационный центр серповидноклеточных заболеваний и талассемии. 11 апреля 2002 года. Архивировано из оригинала 23 сентября 2010 года . Проверено 23 июля 2007 г.
- ^ Имс Д.Л., Гири Л.А., Гран Р.А., Лайонс, Лос-Анджелес (апрель 2006 г.). «Альбинизм у домашней кошки (Felis catus) связан с мутацией тирозиназы (TYR)» . Генетика животных . 37 (2): 175–178. дои : 10.1111/j.1365-2052.2005.01409.x . ПМЦ 1464423 . ПМИД 16573534 .
- ^ «MedlinePlus: Фенилкетонурия» . НИЗ: Национальная медицинская библиотека. Архивировано из оригинала 25 июля 2008 года . Проверено 15 марта 2008 г.
- ^ Например, Ридли М. (2003). Природа через воспитание: гены, опыт и то, что делает нас людьми . Четвертая власть. п. 73. ИСБН 978-1-84115-745-0 .
- ^ Розенталь Д (1964). «Генейские четверняшки: тематическое исследование и теоретический анализ наследственности и окружающей среды при шизофрении». Поведенческая наука . 9 (4): 371. doi : 10.1002/bs.3830090407 .
- ^ Бриванлу А.Х., Дарнелл Дж.Э. (февраль 2002 г.). «Передача сигнала и контроль экспрессии генов». Наука . 295 (5556): 813–818. Бибкод : 2002Sci...295..813B . CiteSeerX 10.1.1.485.6042 . дои : 10.1126/science.1066355 . ПМИД 11823631 . S2CID 14954195 .
- ^ Альбертс и др. (2002), II.3. Контроль экспрессии генов: репрессор триптофана представляет собой простой переключатель, который включает и выключает гены в бактериях. Архивировано 29 июня 2007 г. на Wayback Machine.
- ^ Джениш Р., Бёрд А (март 2003 г.). «Эпигенетическая регуляция экспрессии генов: как геном объединяет внутренние сигналы и сигналы окружающей среды». Природная генетика . 33 (Приложение): 245–254. дои : 10.1038/ng1089 . ПМИД 12610534 . S2CID 17270515 .
- ^ Чендлер В.Л. (февраль 2007 г.). «Парамутация: от кукурузы до мышей» . Клетка . 128 (4): 641–645. дои : 10.1016/j.cell.2007.02.007 . ПМИД 17320501 . S2CID 6928707 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Спонтанные мутации» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Фрейзингер Э., Гроллман А.П., Миллер Х., Кискер С. (апрель 2004 г.). «Толерантность (не) к повреждениям позволяет лучше понять точность репликации ДНК» . Журнал ЭМБО . 23 (7): 1494–1505. дои : 10.1038/sj.emboj.7600158 . ПМК 391067 . ПМИД 15057282 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Индуцированные мутации» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Кадет Дж, Вагнер-младший (февраль 2013 г.). «Повреждение оснований ДНК активными формами кислорода, окислителями и УФ-излучением» . Перспективы Колд-Спринг-Харбор в биологии . 5 (2): а012559. doi : 10.1101/cshperspect.a012559 . ПМК 3552502 . ПМИД 23378590 .
- ^ Йена Н.Р. (июль 2012 г.). «Повреждение ДНК реактивными видами: механизмы, мутации и репарация». Журнал биологических наук . 37 (3): 503–517. дои : 10.1007/s12038-012-9218-2 . ПМИД 22750987 . S2CID 14837181 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Хромосомная мутация I: Изменения в структуре хромосом: Введение» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Шехтер М (2009). Энциклопедия микробиологии . Академическая пресса. п. 551. ИСБН 978-0-12-373944-5 .
- ^ Калвер М., Лимбери А., МакКомб Дж., Бэмфорд М. (2009). Экологическая биология . Издательство Кембриджского университета. п. 118. ИСБН 978-0-521-67982-4 .
- ^ Сойер С.А., Парш Дж., Чжан З., Хартл Д.Л. (апрель 2007 г.). «Распространенность положительного отбора среди замен почти нейтральных аминокислот у дрозофилы» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (16): 6504–6510. Бибкод : 2007PNAS..104.6504S . дои : 10.1073/pnas.0701572104 . ПМЦ 1871816 . ПМИД 17409186 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Вариация и ее модуляция» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Отбор» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Гиллеспи Дж. Х. (ноябрь 2001 г.). «Влияет ли размер популяции вида на его эволюцию?» . Эволюция; Международный журнал органической эволюции . 55 (11): 2161–2169. дои : 10.1111/j.0014-3820.2001.tb00732.x . ПМИД 11794777 . S2CID 221735887 .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). «Случайные события» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Дарвин С. (1859 г.). О происхождении видов . Лондон: Джон Мюррей. п. 1. ISBN 978-0-8014-1319-3 . Архивировано из оригинала 12 декабря 2006 года.
Ранее аналогичные идеи были признаны в Дарвин С. (1861 г.). О происхождении видов (3-е изд.). Лондон: Джон Мюррей. xiii. ISBN 978-0-8014-1319-3 . Архивировано из оригинала 23 февраля 2011 года. - ^ Гаврилец С (октябрь 2003 г.). «Перспектива: модели видообразования: чему мы научились за 40 лет?». Эволюция; Международный журнал органической эволюции . 57 (10): 2197–2215. дои : 10.1554/02-727 . ПМИД 14628909 . S2CID 198158082 .
- ^ Вольф Ю.И., Рогозин И.Б., Гришин Н.В., Кунин Е.В. (сентябрь 2002 г.). «Геномные деревья и древо жизни». Тенденции в генетике . 18 (9): 472–479. дои : 10.1016/S0168-9525(02)02744-0 . ПМИД 12175808 .
- ^ «Использование модельных организмов в обучении» . Университет Висконсина: Модули информационно-пропагандистских исследований штата Висконсин. Архивировано из оригинала 13 марта 2008 года . Проверено 15 марта 2008 г.
- ^ «NCBI: Гены и болезни» . НИЗ: Национальный центр биотехнологической информации. Архивировано из оригинала 20 февраля 2007 года . Проверено 15 марта 2008 г.
- ^ Смит Г.Д. , Ибрагим С. (февраль 2003 г.). « 'Менделевская рандомизация': может ли генетическая эпидемиология способствовать пониманию экологических детерминант болезней?». Международный журнал эпидемиологии . 32 (1): 1–22. дои : 10.1093/ije/dyg070 . ПМИД 12689998 .
- ^ «Информационный бюллетень по фармакогенетике» . НИЗ: Национальный институт общих медицинских наук. Архивировано из оригинала 12 мая 2008 года . Проверено 15 марта 2008 г.
- ^ Франк С.А. (октябрь 2004 г.). «Генетическая предрасположенность к раку - выводы популяционной генетики». Обзоры природы. Генетика . 5 (10): 764–772. дои : 10.1038/nrg1450 . ПМИД 15510167 . S2CID 6049662 .
- ^ Страчан Т., Рид AP (1999). Молекулярная генетика человека 2 (второе изд.). John Wiley & Sons Inc. Глава 18: Генетика рака. Архивировано 26 сентября 2005 г. в Wayback Machine.
- ^ Лодиш и др. (2000), Глава 7: 7.1. Клонирование ДНК с помощью плазмидных векторов. Архивировано 27 мая 2009 г. в Wayback Machine.
- ^ Тиммс Дж. Ф., Крамер Р. (декабрь 2008 г.). «Разностный гель-электрофорез» . ПРОТЕОМИКА . 8 (23–24): 4886–4897. дои : 10.1002/pmic.200800298 . ISSN 1615-9853 .
- ^ Кифер CL (июль 2015 г.). «Искусственное клонирование домашних животных» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (29): 8874–8878. Бибкод : 2015PNAS..112.8874K . дои : 10.1073/pnas.1501718112 . ПМЦ 4517265 . ПМИД 26195770 .
- ^ Лодиш и др. (2000), Глава 7: 7.7. Полимеразная цепная реакция: альтернатива клонированию
- ^ Чанг Д., Трам К., Ли Б., Фэн К., Шен З., Ли Ч. и др. (8 июня 2017 г.). «Обнаружение ДНК-ампликонов полимеразной цепной реакции с помощью лакмусовой бумажки» . Научные отчеты . 7 (3110): 3110. Бибкод : 2017NatSR...7.3110C . дои : 10.1038/s41598-017-03009-z . ПМЦ 5465217 . ПМИД 28596600 .
- ^ Гарибян Л., Нидхи (март 2013 г.). «Полимеразная цепная реакция» . Журнал исследовательской дерматологии . 133 (3): 1–4. дои : 10.1038/jid.2013.1 . ПМК 4102308 . ПМИД 23399825 . Проверено 27 февраля 2024 г.
- ^ Браун Т.А. (2002). «Раздел 2, глава 6: 6.1. Методика секвенирования ДНК» . Геномы 2 (2-е изд.). Оксфорд: Биос. ISBN 978-1-85996-228-2 .
- ^ Браун (2002), Раздел 2, Глава 6: 6.2. Сборка непрерывной последовательности ДНК. Архивировано 8 февраля 2007 г. в Wayback Machine.
- ^ Служба РФ (март 2006 г.). «Секвенирование генов. Гонка за геном за 1000 долларов». Наука . 311 (5767): 1544–1546. дои : 10.1126/science.311.5767.1544 . ПМИД 16543431 . S2CID 23411598 .
- ^ Зал N (май 2007 г.). «Передовые технологии секвенирования и их более широкое влияние в микробиологии» . Журнал экспериментальной биологии . 210 (Часть 9): 1518–1525. дои : 10.1242/jeb.001370 . ПМИД 17449817 .
- ^ Черч ГМ (январь 2006 г.). «Геномы для всех». Научный американец . 294 (1): 46–54. Бибкод : 2006SciAm.294a..46C . doi : 10.1038/scientificamerican0106-46 . ПМИД 16468433 . S2CID 28769137 . (требуется подписка)
- ^ Уэйд Н. (19 марта 2015 г.). «Ученые добиваются запрета метода редактирования генома человека» . Нью-Йорк Таймс . Архивировано из оригинала 19 марта 2015 года . Проверено 20 марта 2015 г.
- ^ Поллак А (3 марта 2015 г.). «Новый мощный способ редактирования ДНК» . Нью-Йорк Таймс . Архивировано из оригинала 26 марта 2015 года . Проверено 20 марта 2015 г.
- ^ Балтимор Д., Берг П., Ботчан М., Кэрролл Д., Чаро Р.А., Чёрч Дж. и др. (апрель 2015 г.). «Биотехнология. Разумный путь вперед в области геномной инженерии и модификации генов зародышевой линии» . Наука . 348 (6230): 36–38. Бибкод : 2015Sci...348...36B . дои : 10.1126/science.aab1028 . ПМЦ 4394183 . ПМИД 25791083 .
- ^ Ланфьер Э., Урнов Ф., Хеккер С.Е., Вернер М., Смоленский Дж. (март 2015 г.). «Не редактируйте зародышевую линию человека» . Природа . 519 (7544): 410–411. Бибкод : 2015Natur.519..410L . дои : 10.1038/519410a . ПМИД 25810189 .
- ^ Колата Г (23 апреля 2015 г.). «Китайские ученые редактируют гены человеческих эмбрионов, вызывая обеспокоенность» . Нью-Йорк Таймс . Архивировано из оригинала 24 апреля 2015 года . Проверено 24 апреля 2015 г.
- ^ Лян П., Сюй Ю, Чжан Х, Дин С., Хуан Р., Чжан З. и др. (май 2015 г.). «CRISPR/Cas9-опосредованное редактирование генов в трехпронуклеарных зиготах человека» . Белок и клетка . 6 (5): 363–372. дои : 10.1007/s13238-015-0153-5 . ПМЦ 4417674 . ПМИД 25894090 .
Дальнейшее чтение
[ редактировать ]- Альбертс Б., Брэй Д., Хопкин К., Джонсон А., Льюис Дж., Рафф М. и др. (2013). Основная клеточная биология, 4-е издание . Гирляндная наука. ISBN 978-1-317-80627-1 .
- Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт, ред. (2000). Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- Хартл Д., Джонс Э. (2005). Генетика: анализ генов и геномов (6-е изд.). Джонс и Бартлетт. ISBN 978-0-7637-1511-3 .
- Кинг Р.С., Маллиган ПК, Стэнсфилд В.Д. (2013). Генетический словарь (8-е изд.). Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-976644-4 .
- Лодиш Х., Берк А., Зипурски Л.С., Мацудайра П., Балтимор Д., Дарнелл Дж. (2000). Молекулярно-клеточная биология (4-е изд.). Нью-Йорк: Книги Scientific American. ISBN 978-0-7167-3136-8 .
Внешние ссылки
[ редактировать ] Цитаты, связанные с генетикой, в Wikiquote
Цитаты, связанные с генетикой, в Wikiquote  Генетика в Wikibooks
Генетика в Wikibooks - Библиотечные ресурсы в вашей и других библиотеках, посвященные генетике.
- Генетика в наше время на BBC
- Генетика в Керли
