Структура и геном ВИЧ
Геном ВИЧ и белки (вирус иммунодефицита были предметом обширных исследований с человека) момента открытия вируса в 1983 году. [ 1 ] [ 2 ] «В поисках причинного агента первоначально считалось, что вирус является формой вируса люкоза человека (HTLV) человека, который в то время был известен, чтобы поразить иммунную систему человека и вызвать определенные лейкемии. Однако,, Исследователи из Института Пастера в Париже выделяли ранее неизвестный и генетически различный ретровирус у пациентов со СПИДом, который впоследствии был назван ВИЧ ». [ 3 ] Каждый вирион содержит вирусную оболочку и связанную матрицу, окружающую капсид , который сам окладывает две копии одноцепочечного генома РНК и несколько ферментов . Открытие самого вируса произошло через два года после отчета о первых основных случаях заболеваний, связанных с СПИДом. [ 4 ] [ 5 ]
Структура
[ редактировать ]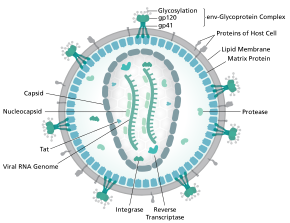

Полная последовательность генома ВИЧ-1, извлеченную из инфекционных вирионов, была решена до однобуклеотидного разрешения . [ 6 ] Геном ВИЧ кодирует небольшое количество вирусных белков , неизменно устанавливая кооперативные ассоциации между белками ВИЧ и между белками ВИЧ и хозяином, чтобы вторгаться в клетки -хозяина и захватить их внутренние механизмы. [ 7 ] ВИЧ отличается по структуре от других ретровирусов . ВИЧ -вирион диаметром ~ 100 нм. в форме конуса Его внутренняя область состоит из ядра , которое включает в себя две копии (положительного смысла) генома SSRNA , ферменты обратной транскриптазы , интегразу и протеазы , некоторые незначительные белки и основной ядро белок. [ 8 ] Геном вируса иммунодефицита человека (ВИЧ) кодирует 8 вирусных белков, играющих важные роли в течение жизненного цикла ВИЧ. [ 7 ]
ВИЧ-1 состоит из двух копий нековалентно связанного , не сплачированной одноцепочечной РНК, заключенной в коническую капсид, состоящий из вирусного белка p24 , типичного для лентивирусов . [ 9 ] [ 10 ] Два РНК часто идентичны, но они не являются независимыми, но образуют компактный димер внутри вириона. [ 11 ] Несколько причин того, почему были предложены две копии РНК, а не только одна, в том числе, вероятно, комбинация этих преимуществ: одно преимущество заключается Обратная транскрипция репликации вируса, тем самым увеличивая генетическое разнообразие. [ 11 ] Другое преимущество заключается в том, что наличие двух копий РНК позволит обратной транскриптазе переключать шаблоны при столкновении с разрывом в вирусной РНК, что завершает обратную транскрипцию без потери генетической информации. [ 11 ] Еще одна причина заключается в том, что димерная природа генома РНК вируса может играть структурную роль в репликации вируса. [ 11 ] Сдерживание двух копий одноцепочечной РНК в вирионе, но выработка только одного провируса ДНК называется псевдодиплоидией. [ 12 ] РНК -компонент составляет 9749 нуклеотидов длиной [ 13 ] [ 14 ] и носит 5 'колпачок (GPPP), 3' поли (а) хвост и множество открытых кадров считывания (ORF). [ 15 ] Вирусные структурные белки кодируются длинными ORF, тогда как меньшие ORF кодируют регуляторы жизненного цикла вируса: привязанность, мембранное слияние, репликация и сборка. [ 15 ]
Одноцепочечная РНК тесно связана с белками нуклеокапсид P7 , белкам позднего сборки P6 и ферментами, необходимыми для развития вириона, таких как обратная транскриптаза и интеграция . ЛНК-лизин является праймером магния-зависимой обратной транскриптазы. [ 9 ] Нуклеокапсид ассоциируется с геномной РНК (одна молекула на гексамер) и защищает РНК от расщепления нуклеазами . В частица вириона также находятся VIF , VPR , NEF и вирусная протеаза . [ Цитация необходима ] Оболочка . вириона образуется плазматической мембраной происхождения клеток -хозяина, которая подтверждается матрицей, состоящей из вирусного белка p17, обеспечивающего целостность частицы вириона На поверхности вириона можно найти ограниченное количество гликопротеина оболочки ( ENV) ВИЧ, тримера, образованного гетеродимерами GP120 и GP41 . ENV отвечает за связывание с его первичным рецептором хозяина, CD4 и его совместным рецептором (в основном CCR5 или CXCR4 ), что приводит к проникновению вируса в клетку-мишени. [ 16 ]
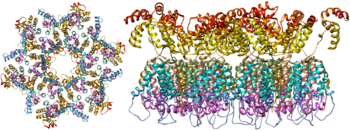
Как единственные белки на поверхности вируса, гликопротеины оболочки (GP120 и GP41) являются основными мишенями для усилий по вакцинам ВИЧ . [ 17 ] Более половины массы тримерного шипа конверта представляет собой n-связанные гликаны . Плотность высока, как гликаны , лежащий в основе вирусного белка от нейтрализации антителами . Это одна из наиболее плотно известных гликозилированных молекул, и плотность достаточно высока, чтобы предотвратить нормальный процесс созревания гликанов во время биогенеза в эндоплазматическом ретикулуме и аппарате Гольги . [ 18 ] [ 19 ] Следовательно, большинство гликанов останавливаются как незрелые гликаны с высокой маннозой , которые обычно не присутствуют на секретируемой или клеточной поверхности гликопротеинов человека. [ 20 ] Необычная обработка и высокая плотность означают, что почти все широко нейтрализующие антитела, которые до сих пор были идентифицированы (от подмножества пациентов, которые были инфицированы в течение многих месяцев до годов), с или, адаптированы, чтобы справиться с этими гликанами конверта. [ 21 ]
Молекулярная структура вирусного всплеска теперь определяется рентгеновской кристаллографией [ 22 ] и крио-электронная микроскопия . [ 23 ] Эти достижения в области структурной биологии стали возможными благодаря развитию стабильных рекомбинантных форм вирусного пика путем введения межсубунитной дисульфидной связи и изолейцина с мутацией пролина в GP41. [ 24 ] Так называемые тримеры SOSIP не только воспроизводят антигенные свойства нативного вирусного всплеска, но и демонстрируют ту же степень незрелых гликанов, что и на нативном вирусе. [ 25 ] Рекомбинантные тримерные вирусные пики являются многообещающими кандидатами на вакцину, поскольку они показывают меньше нететрализирующих эпитопов , чем рекомбинантный мономерный GP120, которые действуют для подавления иммунного ответа на эпитопы-мишени. [ 26 ]
Геном организация
[ редактировать ]
ВИЧ имеет несколько основных генов, кодирующих структурные белки, которые обнаружены во всех ретровирусах, а также несколько неструктурных («аксессуаров») генов, уникальных для ВИЧ. [ 27 ] Геном ВИЧ содержит девять генов, которые кодируют пятнадцать вирусных белков. [ 28 ] Они синтезируются как полипротеины, которые производят белки для внутреннего вириона, называемого GAG, специфического антигена группы; Вирусные ферменты (POL, полимераза) или гликопротеины вирионной ENV (оболочка). [ 29 ] В дополнение к этому, ВИЧ кодирует белки, которые также имеют определенные регуляторные и вспомогательные функции. [ 29 ] ВИЧ-1 имеет два важных регуляторных элемента: TAT и Rev и несколько важных аксессуаров, таких как NEF, VPR, VIF и VPU, которые не являются необходимыми для репликации в определенных тканях. [ 29 ] Ген GAG обеспечивает основную физическую инфраструктуру вируса, а POL обеспечивает базовый механизм, с помощью которого ретровирусы воспроизводятся, в то время как другие помогают ВИЧ войти в клетку -хозяина и усилить его размножение. Хотя они могут быть изменены мутацией, все эти гены, кроме TEV, существуют во всех известных вариантах ВИЧ; См. Генетическая изменчивость ВИЧ . [ Цитация необходима ]
ВИЧ использует сложную систему дифференциального сплайсинга РНК для получения девяти различных генных продуктов от генома менее 10 КБ. [ 30 ] ВИЧ имеет геномный транскрипт 9,2 КБ, который кодирует для предшественников GAG и Pol; Полично сплайсированный, 4,5 т.п.н. Кодирование для ENV, VIF, VPR и VPU и сплайсированной сплайсированной мРНК 2 КБ для TAT, REV и NEF. [ 30 ]
| Сорт | Имя гена | Первичные белковые продукты | Обработанные белковые продукты |
|---|---|---|---|
| Вирусные структурные белки | кляп | GAG Polyprotein | MA, CA, SP1, NC, SP2, P6 |
| pol | Полипотеин | Rt, rnase h, in, pr | |
| эн | GP160 | GP120, GP41 | |
| Основные регулирующие элементы | давать | Давать | |
| преподобный | Преподобный | ||
| Аксессуальные регуляторные белки | нефт | Нефт | |
| vpr | Vpr | ||
| Vif | Vif | ||
| ВПУ | ВПУ |
Вирусные структурные белки
[ редактировать ]
- GAG (групповой антиген) коды для полипротеина GAG-предшественника , который обрабатывается вирусной протеазой во время созревания до MA ( матричный белок , p17); CA (капсидный белок, P24 ); SP1 (спейсер пептид 1, P2); NC ( нуклеокапсидный белок , P7); SP2 (спейсер -пептид 2, P1) и белок P6. [ 31 ]
- Коды POL для вирусных ферментов обратная транскриптаза (RT) и RNASE H , интеграция (In) и ВИЧ -протеаза (PR). [ 29 ] ВИЧ-протеаза необходима для расщепления полипротеина GAG-предшественника для получения структурных белков, RT требуется для транскрибирования ДНК из матрицы РНК, и в нем необходимо интегрировать двухцепочечную вирусную ДНК в геном хозяина. [ 27 ]
- Env (для «конверта») коды для GP160 , который расщепляется протеазой -хозяином, фурин , в рамках эндоплазматической ретикулумы клетки -хозяина. Посттрансляционная обработка продуцирует поверхностный гликопротеин, GP120 или SU, который прикрепляется к рецепторам CD4 , присутствующим на лимфоцитах, и GP41 или TM, который внедряет в вирусную конверту, чтобы вирус прикреплялся к клеткам-мишеням и образуется. [ 27 ] [ 31 ]
Основные регулирующие элементы
[ редактировать ]- TAT (ВИЧ-транс-активатор) играет важную роль в регуляции обратной транскрипции РНК вирусного генома, обеспечивая эффективный синтез вирусных мРНК и регуляции высвобождения вирионов из инфицированных клеток. [ 29 ] TAT экспрессируется как 72- аминокислотный одноэкзонный тат , а также двухэкзонный тат 86–101-аминокислота и играет важную роль в начале ВИЧ-инфекции. TAT (14–15 кДа) связывается с вторичной структурой ствола с стеблями выпиченной геномной РНК вблизи 5- дюймовой области, образующей элемент отклика транса-активации (TAR) . [ 9 ] [ 29 ]
- Rev (Регулятор экспрессии белков вириона): белок Rev связывается с вирусным геномом через аргининовый РНК -связывающий мотив, который также действует как NLS ( сигналы ядерной локализации ), необходимый для транспортировки Rev в ядро из цитозола Во время репликации вируса. [ 29 ] REV распознает сложную структуру ствола env мРНК, расположенную в интроне, разделяющем кодирующий экзон TAT и Rev, известный как элемент ответа на ВИЧ (RRE). [ 9 ] [ 29 ] Преподобный важен для синтеза основных вирусных белков и, следовательно, необходимо для репликации вируса . [ Цитация необходима ]
Аксессуальные регуляторные белки
[ редактировать ]- VPR ( белок лентивируса R): VPR является вирионом, ассоциированным с нуклеоцитоплазматическим регуляторным белком . [ 29 ] Считается, что он играет важную роль в репликации вируса, в частности, ядерного импорта предпринятого комплекса . VPR также, по -видимому, заставляет его клетки -хозяина остановить свой клеточный цикл в фазе G2 . Этот арест активирует механизм восстановления ДНК -хозяина, которая может обеспечить интеграцию вирусной ДНК. [ 9 ] ВИЧ-2 и SIV кодируют дополнительный VPR, связанный с VPR, называемый VPX, который функционирует в связи с VPR. [ 29 ]
- VIF -VIF-это высококонсервативный , 23 кДа фосфопротеин, важный для инфективности вирионов ВИЧ-1 в зависимости от типа клеток. [ 9 ] Было обнаружено, что ВИЧ-1 требует, чтобы VIF синтезировал инфекционные вирусы в лимфоцитах , макрофагах и некоторых клеточных линиях человека . Похоже, что он не требует VIF для того же процесса в клетках HeLa или COS -клеток , среди прочего. [ 29 ]
- NEF -NEF, отрицательный фактор, является N-концевым миристоилированным мембранным фосфопротеином. Он участвует в нескольких функциях во время цикла репликации вируса. Считается, что он играет важную роль в апоптозе клеток и повышает инфекционность вируса . [ 29 ]
- VPU (вирусный белок U)-VPU специфичен для ВИЧ-1. Это олигомерный интегральный мембранный фосфопротеин класса I с многочисленными биологическими функциями. VPU участвует в CD4 деградации путь убиквитина , включающего протеасомный , а также в успешном высвобождении вирионов из инфицированных клеток. [ 9 ] [ 29 ]
- TEV : Этот ген присутствует только в нескольких изолятах ВИЧ-1. Это слияние частей генов TAT , ENV и Rev , и кодирует белок с некоторыми свойствами TAT или ни один из свойств Rev. , но мало [ 32 ]
РНК вторичная структура
[ редактировать ]| HIV pol-1 stem loop | |
|---|---|
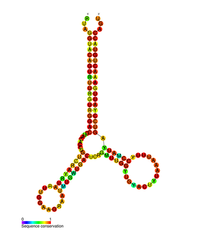 Прогнозируемая вторичная структура петли ствола ВИЧ Pol-1 | |
| Идентификаторы | |
| Символ | pol |
| RFAM | RF01418 |
| Другие данные | |
| РНК тип | Цис-рег |
| PDB Структуры | PDBE |
Несколько консервативных элементов вторичной структуры были идентифицированы в геноме РНК ВИЧ . Структуры ВИЧ -вирусной РНК регулируют прогрессирование обратной транскрипции. [ 33 ] Структура 5'UTR состоит из серии структур петли стеблей, соединенных небольшими линкерами. [ 10 ] Эти петли стеблей (от 5 'до 3') включают элемент транса-активационной области (TAR), 5 ' Полиаденилирующий сигнал [Poly (a)], PBS, DIS, основной SD и ψ Шпилька , расположенная внутри 5 'конец генома и элемент ответа на ВИЧ Rev (RRE) в гене ENV. [ 10 ] [ 34 ] [ 35 ] Другая структура РНК, которая была идентифицирована, - это GAG STEM -петля 3 (GSL3) , которая, как считается, участвует в вирусной упаковке. [ 36 ] [ 37 ] Было предложено вторичные структуры РНК, влияющие на жизненный цикл ВИЧ путем изменения функции ВИЧ -протеазы и обратной транскриптазы , хотя не всем идентифицированным элементам была назначена функция. [ Цитация необходима ]
Вторичная структура РНК, определяемая анализом формы , содержит три петли ствола и расположена между генами ВИЧ -протеазы и обратной транскриптазы. эта цис -регуляторная РНК сохраняется по всей семье ВИЧ и, как считается, влияет на жизненный цикл вируса. Было показано, что [ 38 ]
V3 Loop
[ редактировать ]Третья переменная петля или петля V3 - это часть или область вируса иммунодефицита человека . Петля V3 гликопротеина огибала Viron, GP120 , позволяет ему инфицировать иммунные клетки человека путем связывания с рецептором цитокинов на мишени иммунной клетки человека, таких как клетка CCR5 или клетки CXCR4 , в зависимости от штамма ВИЧ . [ 39 ] Гликопротеин конверта (ENV) GP 120/41 необходим для вступления ВИЧ-1 в клетки. ENV служит молекулярной мишенью лекарства, лечащего людей с ВИЧ-1 инфекцией, и источником иммуногена для развития вакцины СПИДа. Тем не менее, структура функционального тримера Env остается неуловимой. [ 40 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Barré-Sinesi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, Dauguet C, Axler-Blin C, Vézinet-Brun F, Rouzioux C, Rozenbaum W, Montagnier L (май 1983 г.). «Выделение T-лимфотропного ретровируса от пациента, подвергающегося риску синдрома приобретенного иммунного дефицита (СПИД)». Наука . 220 (4599): 868–71. Bibcode : 1983sci ... 220..868b . doi : 10.1126/science.6189183 . PMID 6189183 . S2CID 390173 .
- ^ Галло Р.К., Сарин П.С., Гелманн Э.П., Роберт-Гурофф М., Ричардсон Э., Калянараман В.С., Манн Д., Сидху Г.Д., Шталь Р.Е., Золла-Пазнер С., Лейбович Дж., Попович М (май 1983). «Выделение вируса Т-клеточного лейкоза человека при синдроме приобретенного иммунного дефицита (СПИД)». Наука . 220 (4599): 865–7. Bibcode : 1983sci ... 220..865G . doi : 10.1126/science.6601823 . PMID 6601823 .
- ^ Churri C, Ross MW (2015). "ВИЧ/СПИД " WHELEHAN P, BOLIN A (Eds.). Человек Уайли. ISBN 9781405190060 Полем OCLC 949701914 .
- ^ Центры по борьбе с заболеваниями (июнь 1981 г.). «Pneumocystis pneumonia-Los Angeles». Ммвр. Еженедельный отчет о заболеваемости и смертности . 30 (21): 250–2. PMID 6265753 .
- ^ Центры по контролю заболеваний (CDC) (июль 1981 г.). «Саркома и пневмоцистисная пневмония Капоси среди гомосексуальных мужчин - Нью -Йорк и Калифорния» (PDF) . Ммвр. Еженедельный отчет о заболеваемости и смертности . 30 (25): 305–8. PMID 6789108 . Архивировано из оригинала 22 октября 2012 года . Получено 15 сентября 2017 года .
{{cite journal}}: Cs1 maint: непредвзятый URL ( ссылка ) - ^ Уоттс Дж. М., Данг К.К., Горелик Р.Дж., Леонард С.В., Бесс Дж.В., Свонстром Р., Берч К.Л., недели Км (август 2009 г.). «Архитектура и вторичная структура всего РНК-генома ВИЧ-1» . Природа . 460 (7256): 711–6. Bibcode : 2009natur.460..711w . doi : 10.1038/nature08237 . PMC 2724670 . PMID 19661910 .
- ^ Jump up to: а беременный Li G, de Clercq E (сентябрь 2016 г.). «Ассоциации белков в масштабах ВИЧ-генома: обзор 30 лет исследований» . Микробиология и молекулярная биология обзоры . 80 (3): 679–731. doi : 10.1128/mmbr.00065-15 . PMC 4981665 . PMID 27357278 .
- ^ Singleton P, Sainsbury D, Eds. (2006). "ВИЧ" . Словарь микробиологии и молекулярной биологии (3 -е изд.). Хобокен, Нью -Джерси: Уайли. ISBN 9780470035450 Полем OCLC 71223221 .
- ^ Jump up to: а беременный в дюймовый и фон глин Montagnier L (1999). "Retrovidae" Вирусология (2 -е изд.). стр. 763–774.
- ^ Jump up to: а беременный в Lu K, Heng X, Summers MF (июль 2011 г.). «Структурные детерминанты и механизм упаковки генома ВИЧ-1» . Журнал молекулярной биологии . 410 (4): 609–33. doi : 10.1016/j.jmb.2011.04.029 . PMC 3139105 . PMID 21762803 .
- ^ Jump up to: а беременный в дюймовый Мур, Майкл Д.; Ху, Вэй Шау (2009). «Димеризация РНК ВИЧ-1: для танго требуется два» . Обзоры СПИДа . 11 (2): 91–102. ISSN 1139-6121 . PMC 3056336 . PMID 19529749 .
- ^ Hwang CK, Svarovskaia ES, Pathak VK (октябрь 2001 г.). «Динамический выбор копий: устойчивое состояние между полимеразой вируса мышиного лейкоза и активностью RNASE-H-зависимой полимеразы определяет частоту переключения шаблона in vivo» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (21): 12209–14. Bibcode : 2001pnas ... 9812209h . doi : 10.1073/pnas.221289898 . PMC 59793 . PMID 11593039 .
- ^ Wain-Hobson S, Sonigo P, Danos O, Cole S, Alizon M (январь 1985 г.). «Нуклеотидная последовательность вируса СПИДа, Lav». Клетка . 40 (1): 9–17. doi : 10.1016/0092-8674 (85) 90303-4 . PMID 2981635 . S2CID 33055050 .
- ^ Ратнер Л., Хасельтин В., Патарка Р., Ливак К.Дж., Старчич Б., Джозефс С.Ф., Доран Э.Р., Рафальски Дж.А., Уайтхорн Э.А., Бауместер К. (1985). «Полная нуклеотидная последовательность вируса СПИДа, HTLV-III». Природа . 313 (6000): 277–84. Bibcode : 1985natur.313..277r . doi : 10.1038/313277A0 . PMID 2578615 . S2CID 4316242 .
- ^ Jump up to: а беременный JC, Levy A (2002). "ВИЧ. Энциклопедия рака Тол. 2 (2 -е изд.). стр. 407–415.
- ^ CHECKLY MA, Freed EO (22 июля 2011 г.). «Биосинтез Гликопротеина ВИЧ-1 Гликопротеина, торговля и включение» . Журнал молекулярной биологии . 410 (4): 582–608. doi : 10.1016/j.jmb.2011.04.042 . PMC 3139147 . PMID 21762802 .
- ^ Национальный институт здравоохранения (17 июня 1998 г.). «Кристаллическая структура ключевого белка ВИЧ выявляет новую профилактику, мишени для лечения» (пресс -высвобождение). Архивировано из оригинала 19 февраля 2006 года . Получено 14 сентября 2006 г.
- ^ Behrens AJ, Vasiljevic S, Pritchard LK, Harvey DJ, Andev RS, Krumm SA, Struwe WB, Cupo A, Kumar A, Zitzmann N, Seabright GE, Kramer HB, Spencer DI, Royle L, Lee JH, Klasse PJ, Burton Dr, DR, Royle L, Lee JH, Klasse PJ, Burton Dr, Spencer DI, Royle L, Lee JH, Klasse PJ, Burton Dr, Spencer DI, Royle L, Lee JH, Klass , Wilson IA, Ward AB, Sanders RW, Moore JP, Doores KJ, Crispin M (март 2016 г.). «Композиция и антигенные эффекты отдельных гликановых участков тримерного гликопротеина огибального ВИЧ-1» . Сотовые отчеты . 14 (11): 2695–706. doi : 10.1016/j.celrep.2016.02.058 . PMC 4805854 . PMID 26972002 .
- ^ Причард Л.К., Спенсер Д.И., Ройл Л., Бонолли С., Сибрайт Г.Е., Беренс А.Дж., Кульп Д.В., Менис С., Крумм С.А., Данлоп Д.К., Криспин Д.Дж (Июнь 2015 г.). «Гликан кластеризация стабилизирует маннозу ВИЧ-1 и сохраняет уязвимость для широкого нейтрализации антител» . Природная связь . 6 : 7479. Bibcode : 2015natco ... 6.7479p . doi : 10.1038/ncomms8479 . PMC 4500839 . PMID 26105115 .
- ^ Причард Л.К., Харви Д.Дж., Бонолли С., Криспин М., Дверей К.Дж. (сентябрь 2015 г.). «Клеточно-белковая гликозилирование нативного расщепленного огибала ВИЧ-1» . Журнал вирусологии . 89 (17): 8932–44. doi : 10.1128/jvi.01190-15 . PMC 4524065 . PMID 26085151 .
- ^ Crispin M, Doores KJ (апрель 2015 г.). «Нацеливание на гликаны, полученные из хозяина на охваченные вирусы для конструкции вакцины на основе антител» . Текущее мнение о вирусологии . Вирусный патогенез • Профилактические и терапевтические вакцины. 11 : 63–9. doi : 10.1016/j.coviro.2015.02.002 . PMC 4827424 . PMID 25747313 .
- ^ Жюльен Дж.П., Купе А., Сок Д., Стэнфилд Р.Л., Люмкис Д., Деллер М.К., Классе П.Дж., Бертон Д.Р., Сандерс Р.В., Мур Дж.П., Уорд А.Б., Уилсон И.А. (декабрь 2013 г.). «Кристаллическая структура растворимой тримера растворимой расщепленной ВИЧ-1» . Наука . 342 (6165): 1477–83. Bibcode : 2013sci ... 342.1477j . doi : 10.1126/science.1245625 . PMC 3886632 . PMID 24179159 .
- ^ Lyumkis D, Julien JP, De Val N, Cupo A, Potter CS, Klasse PJ, Burton DR, Sanders RW, Moore JP, Carragher B, Wilson IA, Ward AB (декабрь 2013 г.). «Крио-эм структура полностью гликозилированной растворимой расщепленной тример огибала ВИЧ-1» . Наука . 342 (6165): 1484–90. Bibcode : 2013sci ... 342.1484L . doi : 10.1126/science.1245627 . PMC 3954647 . PMID 24179160 .
- ^ Сандерс Р.В., Деркинг Р., Куп -А., Жюльен Дж.П., Ясмин А., де Валь Н., Ким Х.Дж , Wilson IA, Ward AB, Klasse PJ, Moore JP (сентябрь 2013 г.). «Растворимый растворимый растворимый ВИЧ-1 Env Trimer, BG505 Sosip.664 GP140, экспрессирует множественные эпитопы для широко нейтрализующих, но не нейтрализующих антител» . PLO -патогены . 9 (9): E1003618. doi : 10.1371/journal.ppat.1003618 . PMC 3777863 . PMID 24068931 .
- ^ Притчард Л.К., Васильжевич С., Озоровски Г., Сибрайт Г.Е., Куп -А., Ринг Р., Ким Х.Дж., Сандерс Р.В., Дверей К.Дж., Бертон Д.А., Уилсон И.А., Уорд А.Б., Мур Дж.П., Криспин М (июнь 2015 г.). «Структурные ограничения определяют гликозилирование тримеров оболочки ВИЧ-1» . Сотовые отчеты . 11 (10): 1604–13. doi : 10.1016/j.celrep.2015.05.017 . PMC 4555872 . PMID 26051934 .
- ^ De Taeye SW, Ozorowski G, Torrents de la Peña A, Guttman JP, Van Den Kekhof TL, Burger JA, Pritchard LK, Pugach P, Yasmeen A, Crampton J, Hu J, Bontjer I, Torres JL, Aredt H, Destefano J, , Koff WC, Schuitemaker H, Eggink D, Berkhout B, Dean H, Labranche C, Crottty S, Crottty S, Crottty S, Crottty M, Montefiori DC, Klasse PJ, Lee KK, Moore JP, Wilson IA, Wilson IA, слова RW (декабрь 2015 г.). «Иммуногенность стабилизированных охватов ВИЧ-1 с уменьшенным воздействием нететрализующих эпитопов » Ячейка 163 (7): 1702–1 Doi : 10.1016/ j.cell.2015.11.0 4732737PMC 26687358PMID
- ^ Jump up to: а беременный в Мушахвар Ик (2007). «Вирусы иммунодефицита человека: молекулярная вирусология, патогенез, диагностика и лечение». Перспективы в медицинской вирусологии . 13 : 75–87. doi : 10.1016/s0168-7069 (06) 13005-0 . ISBN 9780444520739 .
- ^ Li G, Piampongsant S, Faria NR, Voet A, Pineda-Peña AC, Khori R, Lemey P, Vandamme Am, The The K (февраль 2015 г.). «Интегрированная карта вариации по всему геному ВИЧ с точки зрения популяции» . Ретровирология . 12 (1): 18. doi : 10.1186/s12977-015-0148-6 . PMC 4358901 . PMID 25808207 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м Votteler J, Schubert U (2008). «Вирусы иммунодефицита человека: молекулярная биология». Энциклопедия вирусологии (3 -е изд.). С. 517–525.
- ^ Jump up to: а беременный Файнберг Марк Б., Грин Уорнер С. (1992). «Молекулярное понимание патогенеза вируса иммунодефицита человека типа». Текущее мнение в иммунологии . 4 (4): 466–474. doi : 10.1016/s0952-7915 (06) 80041-5 . PMID 1356348 .
- ^ Jump up to: а беременный Король Стивен Р. (1994). «ВИЧ: вирусология и механизмы болезней». Анналы неотложной медицины . 24 (3): 443–449. doi : 10.1016/s0196-0644 (94) 70181-4 . PMID 7915889 .
- ^ Бенко Д.М., Шварц С., Павлакис Г.Н., Фелбер Б.К. (июнь 1990 г.). «Новый белок вируса иммунодефицита человека типа 1, TEV, разделяет последовательности с белками TAT, ENV и Rev» . Журнал вирусологии . 64 (6): 2505–18. doi : 10.1128/jvi.64.6.2505-2518.1990 . PMC 249426 . PMID 2186172 .
- ^ Krupkin M, Jackson LN, Ha B, Puglisi EV (декабрь 2020 г.). «Достижения в понимании инициации обратной транскрипции ВИЧ-1» . Curr Mind Struct Biol . 65 : 175–183. doi : 10.1016/j.sbi.2020.07.005 . PMC 9973426 . PMID 32916568 . S2CID 221636459 .
- ^ Беркхаут Б (январь 1992 г.). «Структурные особенности в TAR РНК вирусов иммунодефицита человека и обезьяны: филогенетический анализ» . Исследование нуклеиновых кислот . 20 (1): 27–31. doi : 10.1093/nar/20.1.27 . PMC 310321 . PMID 1738599 .
- ^ Paillart JC, Skripkin E, Ehresmann B, Ehresmann C, Marquet R (февраль 2002 г.). «Доказательства in vitro для дальнего псевдокнота в 5'-добываемых и матричных областях кодирующих геномной РНК ВИЧ-1» . Журнал биологической химии . 277 (8): 5995–6004. doi : 10.1074/jbc.m108972200 . PMID 11744696 .
- ^ Damgarard CK, Andersen ES, Knudsen B, Gorodkin J, Kjems J (февраль 2004 г.). «РНК -взаимодействия в 5 'геноме регионального состояния» Журнал молекулярной биологии 336 (2): 369–7 Doi : 10.1016/ jmb.2003.12.0 14757051PMID
- ^ Ронг Л., Рассел Р.С., Ху Дж, Лафреа М., Уэйнберг М.А., Лян С. (сентябрь 2003 г.). «Удаление стволового петли 3 компенсируется мутациями второго места в белке GAG вируса иммунодефицита человека типа 1». Вирусология . 314 (1): 221–8. doi : 10.1016/s0042-6822 (03) 00405-7 . PMID 14517075 .
- ^ Wang Q, Barr I, Guo F, Lee C (декабрь 2008 г.). «Свидетельство новой вторичной структуры РНК в кодирующей области гена Pol ВИЧ-1» . РНК . 14 (12): 2478–88. doi : 10.1261/rna.1252608 . PMC 2590956 . PMID 18974280 .
- ^ «Взаимодействие петли GP120 V3 различных штаммов ВИЧ-1 с мощным анти-ВИЧ-моноклональным антителом 447-52D» . Институт науки Вайзмана: кафедра структурной биологии . Архивировано из оригинала 2007-07-18 . Получено 2017-04-18 .
- ^ Такеда С., Такизава М., Мияучи К., Урано Е., Фудзино М., Мураками Т., Мураками Т., Комано Дж (июнь 2016 г.). «Конформационные свойства третьей переменной петли гликопротеина огибания ВИЧ-1AD8 в условиях лиговых условий». Биохимическая и биофизическая исследовательская коммуникация . 475 (1): 113–8. doi : 10.1016/j.bbrc.2016.05.051 . PMID 27178216 .
Внешние ссылки
[ редактировать ]- Вход RFAM для петли STEM по ВИЧ POL-1
- 3D -модель полного вириона ВИЧ1
- Лю Дж., Райт Э.Р., Винклер Х. (2010). «3D -визуализация вирионов ВИЧ с помощью криоэлектронной томографии». Крио-Эм, часть C: анализ, интерпретация и тематические исследования . Методы в фермере. Тол. 483. С. 267–90. doi : 10.1016/s0076-6879 (10) 83014-9 . ISBN 9780123849939 Полем PMC 3056484 . PMID 20888479 .