ВИЧ-1 протеаза
| Протеаза ВИЧ-1 (ретропепсин) | |||
|---|---|---|---|
 протеазы ВИЧ-1 Димер выделен белым и серым цветом, пептидный субстрат показан черным, а активного центра боковые цепи аспартата показаны красным. ( PDB : 1KJF ) | |||
| Идентификаторы | |||
| Номер ЕС. | 3.4.23.16 | ||
| Номер CAS. | 144114-21-6 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Протеаза ВИЧ-1 или PR — это ретровирусная аспартилпротеаза (ретропепсин), фермент , участвующий в гидролизе пептидных связей в ретровирусах, который необходим для жизненного цикла ВИЧ , ретровируса , вызывающего СПИД . [ 1 ] [ 2 ] PR ВИЧ-1 расщепляет вновь синтезированные полипротеины (а именно Gag и Gag- Pol). [ 3 ] ) в девяти сайтах расщепления для создания зрелых белковых компонентов вириона ВИЧ , инфекционной формы вируса вне клетки-хозяина. [ 4 ] Без эффективного PR ВИЧ-1 вирионы ВИЧ остаются неинфекционными. [ 5 ] [ 6 ]
Структура
[ редактировать ]
Зрелая протеаза ВИЧ существует в виде гомодимера массой 22 кДа , каждая субъединица которого состоит из 99 аминокислот. [ 1 ] Один активный сайт расположен между идентичными субъединицами и имеет характерную последовательность Asp - Thr - Gly (Asp25, Thr26 и Gly27), каталитическую триадную общую для аспарагиновых протеаз. [ 8 ] Поскольку PR ВИЧ-1 может функционировать только в виде димера, зрелая протеаза содержит две аминокислоты Asp25, по одной от каждого мономера, которые действуют вместе друг с другом в качестве каталитических остатков. [ 9 ] Кроме того, протеаза ВИЧ имеет два молекулярных «лоскута», которые перемещаются на расстояние до 7 Å , когда фермент связывается с субстратом. [ 10 ] Это можно визуализировать с помощью анимации открытия и закрытия створок .
Биосинтез
[ редактировать ]
Предшественник
[ редактировать ]Полипротеин Gag-Pol, который содержит преждевременно кодирующие белки, в том числе PR ВИЧ-1. [ 9 ] PR расположен между обратной транскриптазой (которая находится на С-конце PR) и p6. pol (который находится на N-конце PR) трансфреймовой области (TFR). [ 11 ]
Чтобы этот предшественник стал функциональным белком, каждый мономер должен ассоциироваться с другим мономером PR ВИЧ-1 с образованием функционального каталитического активного сайта, каждый из которых вносит Asp25 в свои соответствующие каталитические триады. [ 9 ]
Механизм синтеза
[ редактировать ]Когда вирусная РНК ВИЧ попадает в клетку, это сопровождается обратной транскриптазой , интегразой и зрелым PR ВИЧ-1. Обратная транскриптаза преобразует вирусную РНК в ДНК, облегчая роль интегразы во включении вирусной генетической информации в ДНК клетки-хозяина. [ 2 ] Вирусная ДНК может либо оставаться в спящем состоянии в ядре, либо транскрибироваться в мРНК и транслироваться клеткой-хозяином в полипротеин Gag-Pol, который затем расщепляется на отдельные функциональные белки (включая вновь синтезированный PR ВИЧ-1) зрелыми клетками. ВИЧ-1 ПР. [ 9 ]
Предшественник PR ВИЧ-1 катализирует собственное производство, способствуя его отщеплению от полипротеина Gag-Pol по механизму, известному как аутопроцессинг. Аутопроцессинг PR ВИЧ-1 характеризуется двумя последовательными этапами: (1) внутримолекулярное расщепление N-конца на уровне p6. pol -сайт расщепления протеазы, который служит для завершения процессинга PR и повышения ферментативной активности с вновь образованным промежуточным продуктом PR-обратной транскриптазы, и (2) межмолекулярного расщепления С-конца в сайте расщепления протеазой-обратной транскриптазой, что приводит к сборке двух субъединиц PR в зрелые димеры. [ 12 ] [ 13 ] Димеризация двух субъединиц позволяет сформировать полностью функциональный комбинированный активный центр, характеризующийся двумя каталитическими остатками Asp25 (по одному от каждого мономера). [ 14 ]


Функция
[ редактировать ]ПР по ВИЧ-1 служит двойной цели. PR-предшественник ВИЧ-1 отвечает за катализацию собственного производства в зрелые ферменты PR посредством автопроцессинга PR. [ 15 ] Зрелая протеаза способна гидролизовать пептидные связи полипротеинов Gag-Pol в девяти специфических сайтах, превращая полученные субъединицы в зрелые, полностью функциональные белки. Эти расщепленные белки, включая обратную транскриптазу, интегразу и РНКазу H, кодируются компонентами кодирующей области, необходимыми для репликации вируса. [ 4 ]
Механизм
[ редактировать ]Как аспарагиновая протеаза, димеризованный PR ВИЧ-1 действует через комплекс аспартильных групп, осуществляя гидролиз. Из двух остатков Asp25 в объединенном каталитическом активном сайте PR ВИЧ-1 один депротонирован, а другой протонирован из-за различий pKa от микроокружения. [ 16 ]
В общем механизме аспарагиновой протеазы, как только субстрат правильно связывается с активным центром фермента, депротонированная каталитическая аминокислота Asp25 подвергается основному катализу, делая поступающую молекулу воды лучшим нуклеофилом за счет ее депротонирования. Образующийся гидроксильный ион атакует карбонильный углерод пептидной связи, образуя промежуточный продукт с временным оксианионом, который стабилизируется первоначально протонированным Asp25. Оксианион повторно образует двойную связь, что приводит к разрыву пептидной связи между двумя аминокислотами, в то время как первоначально депротонированный Asp25 подвергается кислотному катализу, отдавая свой протон аминогруппе, что делает аминогруппу лучшей уходящей группой для полного разрыв пептидной связи и возвращение в исходное депротонированное состояние. [ 2 ] [ 17 ]
Хотя PR ВИЧ-1 имеет многие из тех же характеристик, что и невирусная аспарагиновая протеаза, некоторые данные показали, что PR ВИЧ-1 согласованно катализирует гидролиз; другими словами, нуклеофильная молекула воды и протонированный Asp25 одновременно атакуют разрезаемую пептидную связь во время катализа. [ 17 ] [ 18 ]

Как мишень для наркотиков
[ редактировать ]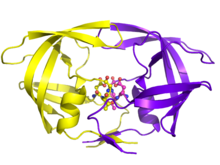
Благодаря своей неотъемлемой роли в репликации ВИЧ, протеаза ВИЧ стала основной мишенью для лекарственной терапии. Ингибиторы ВИЧ-протеазы действуют путем специфического связывания с активным центром, имитируя тетраэдрический промежуточный продукт его субстрата и, по сути, «застревают», выводя из строя фермент. После сборки и отпочкования вирусные частицы, лишенные активной протеазы, не могут созреть в инфекционные вирионы. Несколько ингибиторов протеазы были лицензированы для терапии ВИЧ. [ 19 ]
одобрено десять ингибиторов PR ВИЧ-1 В настоящее время Управление по санитарному надзору за качеством пищевых продуктов и медикаментов : индинавир , саквинавир , ритонавир , нелфинавир , лопинавир , ампренавир , фосампреневир , атазанавир , типранавир и дарунавир . Многие ингибиторы имеют разные молекулярные компоненты и, следовательно, разные механизмы действия, такие как блокирование активного центра. Их функциональная роль также распространяется на влияние на концентрацию в крови других ингибиторов (ритонавир) и их использование только при определенных обстоятельствах, при которых вирус проявляет толерантность к другим ингибиторам (типранавиру). [ 4 ] [ 20 ]
Эволюция и сопротивление
[ редактировать ]Из-за высокой частоты мутаций ретровирусов, особенно из-за мутационно-чувствительных областей (особенно области, содержащей последовательность каталитической триады), а также учитывая, что изменения нескольких аминокислот в протеазе ВИЧ могут сделать ее гораздо менее заметной для ингибитора, активный Сайт этого фермента может быстро меняться под селективным давлением препаратов, ингибирующих репликацию. [ 21 ] [ 22 ]
Два типа мутаций обычно связаны с повышением лекарственной устойчивости: «основные» мутации и «вторичные» мутации. Основные мутации включают мутацию в активном сайте PR ВИЧ-1, не позволяющую селективным ингибиторам связывать его. Вторичные мутации относятся к молекулярным изменениям на периферии фермента из-за длительного воздействия аналогичных химических веществ, потенциально влияющих на специфичность ингибитора PR ВИЧ-1. [ 3 ]
Один из подходов к минимизации развития лекарственной устойчивости ВИЧ заключается в назначении комбинации препаратов , которые одновременно ингибируют несколько ключевых аспектов цикла репликации ВИЧ, а не по одному препарату за раз. Другие цели лекарственной терапии включают обратную транскриптазу , прикрепление вируса, слияние мембран, интеграцию кДНК и сборку вириона. [ 23 ] [ 24 ]
См. также
[ редактировать ]Внешние ссылки
[ редактировать ]- Онлайн-база данных MEROPS . по пептидазам и их ингибиторам: A02.001
- Proteopedia ВИЧ-1_протеаза - структура протеазы ВИЧ-1 в интерактивном 3D.
- Proteopedia Flaps_Morph_for_HIV_Protease — Анимация открытия и закрытия створок на основе рентгеновских кристаллических структур.
- ВИЧ-1+протеаза в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Дэвис Д.Р. (1990). «Структура и функции аспарагиновых протеиназ» . Ежегодный обзор биофизики и биофизической химии . 19 (1): 189–215. дои : 10.1146/annurev.bb.19.060190.001201 . ПМИД 2194475 .
- ^ Перейти обратно: а б с Брик А., Вонг CH (январь 2003 г.). «Протеаза ВИЧ-1: механизм и открытие лекарства». Органическая и биомолекулярная химия . 1 (1): 5–14. дои : 10.1039/b208248a . ПМИД 12929379 .
- ^ Перейти обратно: а б Хуанг X, Бритто, доктор медицинских наук, Кир-Скотт Дж.Л., Бун К.Д., Рокка-младший, Зиммерлинг С., Маккенна Р., Биери М., Гули PR, Данн Б.М., Фануччи Дж.Е. (июнь 2014 г.). «Роль избранных полиморфизмов подтипов в конформационной выборке и динамике протеазы ВИЧ-1» . Журнал биологической химии . 289 (24): 17203–14. дои : 10.1074/jbc.M114.571836 . ПМК 4059161 . ПМИД 24742668 .
- ^ Перейти обратно: а б с Лев З, Чу Ю, Ван Ю (апрель 2015 г.). «Ингибиторы протеазы ВИЧ: обзор молекулярной селективности и токсичности» . ВИЧ/СПИД: исследования и паллиативная помощь . 7 : 95–104. дои : 10.2147/hiv.s79956 . ПМК 4396582 . ПМИД 25897264 .
- ^ Краусслих Х.Г., Ингрэм Р.Х., Скуг М.Т., Виммер Э., Паллай П.В., Картер Калифорния (февраль 1989 г.). «Активность очищенной биосинтетической протеиназы вируса иммунодефицита человека на природных субстратах и синтетических пептидах» . Труды Национальной академии наук Соединенных Штатов Америки . 86 (3): 807–11. Бибкод : 1989ПНАС...86..807К . дои : 10.1073/pnas.86.3.807 . ПМК 286566 . ПМИД 2644644 .
- ^ Коль Н.Е., Эмини Э.А., Шляйф В.А., Дэвис Л.Дж., Хаймбах Дж.К., Диксон Р.А., Сколник Э.М., Сигал И.С. (июль 1988 г.). «Для вирусной инфекционности необходима активная протеаза вируса иммунодефицита человека» . Труды Национальной академии наук Соединенных Штатов Америки . 85 (13): 4686–90. Бибкод : 1988PNAS...85.4686K . дои : 10.1073/pnas.85.13.4686 . ПМК 280500 . ПМИД 3290901 .
- ^ Перриман А.Л., Лин Дж.Х., Маккаммон Дж.А. (апрель 2004 г.). «Молекулярная динамика протеазы ВИЧ-1 дикого типа и мутанта V82F/I84V: возможный вклад в устойчивость к лекарствам и потенциальный новый целевой сайт для лекарств» (PDF) . Белковая наука . 13 (4): 1108–23. дои : 10.1110/ps.03468904 . ПМК 2280056 . ПМИД 15044738 . Архивировано из оригинала (PDF) 16 декабря 2008 г.
- ^ Чаттерджи А., Мридула П., Мишра Р.К., Миттал Р., Хосур Р.В. (март 2005 г.). «Фолдинг регулирует аутопроцессинг предшественника протеазы ВИЧ-1» . Журнал биологической химии . 280 (12): 11369–78. дои : 10.1074/jbc.M412603200 . ПМИД 15632156 .
- ^ Перейти обратно: а б с д Петтит С.К., Эверитт Л.Е., Чоудхури С., Данн Б.М., Каплан А.Х. (август 2004 г.). «Первоначальное расщепление предшественника GagPol вируса иммунодефицита человека типа 1 его активированной протеазой происходит по внутримолекулярному механизму» . Журнал вирусологии . 78 (16): 8477–85. doi : 10.1128/JVI.78.16.8477-8485.2004 . ПМК 479095 . ПМИД 15280456 .
- ^ Миллер М., Шнайдер Дж., Сатьянараяна Б.К., Тот М.В., Маршалл Г.Р., Клоусон Л., Селк Л., Кент С.Б., Влодавер А. (декабрь 1989 г.). «Структура комплекса синтетической протеазы ВИЧ-1 с ингибитором на основе субстрата при разрешении 2,3 А». Наука . 246 (4934): 1149–52. дои : 10.1126/science.2686029 . ПМИД 2686029 .
- ^ Луи Дж. М., Клор Г. М., Гроненборн А. М. (сентябрь 1999 г.). «Аутопроцессинг протеазы ВИЧ-1 тесно связан со сворачиванием белка». Структурная биология природы . 6 (9): 868–75. дои : 10.1038/12327 . ПМИД 10467100 . S2CID 6375519 .
- ^ Луи Дж.М., Нэшед Н.Т., Пэррис К.Д., Киммел А.Р., Джерина Д.М. (август 1994 г.). «Кинетика и механизм аутопроцессинга протеазы вируса иммунодефицита человека 1 типа из аналога полипротеина Gag-Pol» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (17): 7970–4. Бибкод : 1994PNAS...91.7970L . дои : 10.1073/pnas.91.17.7970 . ПМК 44526 . ПМИД 8058744 .
- ^ Вондрак Э.М., Нэшед Н.Т., Хабер М.Т., Джерина Д.М., Луи Дж.М. (февраль 1996 г.). «Переходный предшественник протеазы ВИЧ-1. Выделение, характеристика и кинетика созревания» . Журнал биологической химии . 271 (8): 4477–81. дои : 10.1074/jbc.271.8.4477 . ПМИД 8626801 .
- ^ Чжан С., Каплан А.Х., Тропша А. (ноябрь 2008 г.). «Исследование функции и структуры протеазы ВИЧ-1 с использованием упрощенного анализа окрестности метода упаковки белка» . Белки . 73 (3): 742–53. дои : 10.1002/prot.22094 . ПМЦ 2765824 . ПМИД 18498108 .
- ^ Хуан Л., Чен С. (июль 2013 г.). «Понимание автопроцессинга протеазы ВИЧ-1 для разработки новых терапевтических средств» . Будущая медицинская химия . 5 (11): 1215–29. дои : 10.4155/fmc.13.89 . ПМЦ 3826259 . ПМИД 23859204 .
- ^ Смит Р., Бреретон И.М., Чай Р.Ю., Кент С.Б. (ноябрь 1996 г.). «Состояния ионизации каталитических остатков протеазы ВИЧ-1». Структурная биология природы . 3 (11): 946–50. дои : 10.1038/nsb1196-946 . ПМИД 8901873 . S2CID 1076528 .
- ^ Перейти обратно: а б Лю Х, Мюллер-Плате Ф, ван Гюнстерен ВФ (август 1996 г.). «Комбинированное квантово-классическое молекулярно-динамическое исследование каталитического механизма протеазы ВИЧ». Журнал молекулярной биологии . 261 (3): 454–69. дои : 10.1006/jmbi.1996.0476 . ПМИД 8780786 .
- ^ Яскольски М., Томасселли А.Г., Сойер Т.К., Стейплс Д.Г., Хейнриксон Р.Л., Шнайдер Дж., Кент С.Б., Влодавер А. (февраль 1991 г.). «Структура с разрешением 2,5 А химически синтезированной протеазы вируса иммунодефицита человека типа 1 в комплексе с ингибитором на основе гидроксиэтилена». Биохимия . 30 (6): 1600–9. дои : 10.1021/bi00220a023 . ПМИД 1993177 .
- ^ Позвонил HP (2007). Фармакология Ранга и Дейла (6-е изд.). Филадельфия, Пенсильвания, США: Черчилль Ливингстон/Эльзевир. ISBN 9780808923541 .
- ^ Гриффин Л., Аннаерт П., Брауэр К.Л. (сентябрь 2011 г.). «Влияние белков-переносчиков лекарств на фармакокинетику и лекарственное взаимодействие ингибиторов протеазы ВИЧ» . Журнал фармацевтических наук . 100 (9): 3636–54. дои : 10.1002/jps.22655 . ПМЦ 3750718 . ПМИД 21698598 .
- ^ Уоткинс Т., Реш В., Ирлбек Д., Суонстром Р. (февраль 2003 г.). «Выбор высокого уровня устойчивости к ингибиторам протеазы вируса иммунодефицита человека 1 типа» . Антимикробные средства и химиотерапия . 47 (2): 759–69. doi : 10.1128/AAC.47.2.759-769.2003 . ПМК 151730 . ПМИД 12543689 .
- ^ Леб Д.Д., Суонстром Р., Эверитт Л., Манчестер М., Стампер С.Е., Хатчисон, Калифорния (август 1989 г.). «Полный мутагенез протеазы ВИЧ-1». Природа . 340 (6232): 397–400. Бибкод : 1989Natur.340..397L . дои : 10.1038/340397a0 . ПМИД 2666861 . S2CID 4351388 .
- ^ Мур Дж.П., Стивенсон М. (октябрь 2000 г.). «Новые мишени для ингибиторов репликации ВИЧ-1». Обзоры природы. Молекулярно-клеточная биология . 1 (1): 40–9. дои : 10.1038/35036060 . ПМИД 11413488 . S2CID 10811618 .
- ^ Де Клерк Э (декабрь 2007 г.). «Разработка лекарств от ВИЧ и ВГС». Обзоры природы. Открытие наркотиков . 6 (12): 1001–18. дои : 10.1038/nrd2424 . ПМИД 18049474 . S2CID 37859193 .