Структура и геном ВИЧ
Геном ВИЧ и белки иммунодефицита ( вируса человека) стали предметом обширных исследований с момента открытия вируса в 1983 году. [ 1 ] [ 2 ] «При поиске возбудителя изначально предполагалось, что вирус представляет собой форму вируса Т-клеточного лейкоза человека (HTLV), который, как было известно в то время, влияет на иммунную систему человека и вызывает некоторые лейкозы. Однако исследователи из Института Пастера в Париже выделили ранее неизвестный и генетически отличающийся ретровирус у пациентов со СПИДом, который позже был назван ВИЧ». [ 3 ] Каждый вирион состоит из вирусной оболочки и связанного с ней матрикса, содержащего капсид , который в свою очередь содержит две копии генома одноцепочечной РНК и несколько ферментов . Открытие самого вируса произошло через два года после сообщения о первых крупных случаях заболеваний, связанных со СПИДом. [ 4 ] [ 5 ]
Структура
[ редактировать ]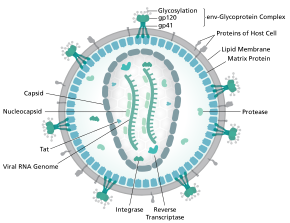

Полная последовательность генома ВИЧ-1, извлеченная из инфекционных вирионов, была решена с точностью до одного нуклеотида . [ 6 ] Геном ВИЧ кодирует небольшое количество вирусных белков , неизменно устанавливая кооперативные ассоциации между белками ВИЧ, а также между ВИЧ и белками хозяина, чтобы проникнуть в клетки-хозяева и похитить их внутренние механизмы. [ 7 ] ВИЧ отличается по структуре от других ретровирусов . Вирион ВИЧ имеет диаметр ~100 нм. Его самая внутренняя область состоит из конусообразного ядра (положительного смысла) , которое включает две копии генома оцРНК , ферменты обратной транскриптазы , интегразы и протеазы , некоторые второстепенные белки и основной ядерный белок. [ 8 ] Геном вируса иммунодефицита человека (ВИЧ) кодирует 8 вирусных белков, играющих важную роль в жизненном цикле ВИЧ. [ 7 ]
ВИЧ-1 состоит из двух копий нековалентно связанных , несплайсированных , с положительным смыслом, одноцепочечных РНК заключенных в конический капсид, состоящий из вирусного белка p24 , типичного для лентивирусов . [ 9 ] [ 10 ] Две РНК часто идентичны, но они не независимы, а образуют компактный димер внутри вириона. [ 11 ] Было предложено несколько причин того, почему упаковываются две копии РНК, а не только одна, включая, возможно, комбинацию этих преимуществ. Одним из преимуществ является то, что две копии нитей РНК жизненно важны для содействия рекомбинации ВИЧ-1, которая происходит во обратная транскрипция репликации вируса, тем самым увеличивая генетическое разнообразие. [ 11 ] Еще одним преимуществом является то, что наличие двух копий РНК позволит обратной транскриптазе переключать матрицы при обнаружении разрыва вирусной РНК, тем самым завершая обратную транскрипцию без потери генетической информации. [ 11 ] Еще одна причина заключается в том, что димерная природа РНК-генома вируса может играть структурную роль в репликации вируса. [ 11 ] Содержание двух копий одноцепочечной РНК внутри вириона, но образование только одной ДНК-провируса, называется псевдодиплоидией. [ 12 ] Компонент РНК имеет 9749 нуклеотидов. длину [ 13 ] [ 14 ] и имеет 5'-шапку (Gppp), 3'- поли(А)-хвост и множество открытых рамок считывания (ORF). [ 15 ] Вирусные структурные белки кодируются длинными ORF, тогда как более мелкие ORF кодируют регуляторы жизненного цикла вируса: прикрепление, слияние мембран, репликацию и сборку. [ 15 ]
Одноцепочечная РНК прочно связана с белками нуклеокапсида p7 , белком поздней сборки p6 и ферментами, необходимыми для развития вириона, такими как обратная транскриптаза и интеграза . Лизиновая тРНК является праймером магнийзависимой обратной транскриптазы. [ 9 ] Нуклеокапсид связывается с геномной РНК (одна молекула на гексамер) и защищает РНК от расщепления нуклеазами . Внутри частицы вириона также заключены Vif , Vpr , Nef и вирусная протеаза . [ нужна ссылка ] Оболочка . вириона образована плазматической мембраной клеточного происхождения, которая поддерживается матриксом, состоящим из вирусного белка p17, обеспечивающего целостность частицы вириона На поверхности вириона можно обнаружить ограниченное количество оболочечного гликопротеина (Env) ВИЧ, тримера, образованного гетеродимерами gp120 и gp41 . Env отвечает за связывание со своим первичным рецептором хозяина, CD4, и его корецептором (в основном CCR5 или CXCR4 ), что приводит к проникновению вируса в клетку-мишень. [ 16 ]
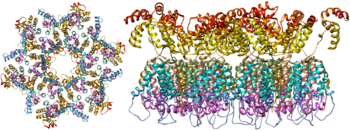
Гликопротеины оболочки (gp120 и gp41), единственные белки на поверхности вируса, являются основными мишенями для разработки вакцин против ВИЧ . [ 17 ] Более половины массы шипа тримерной оболочки составляют N-связанные гликаны . Плотность высока, поскольку гликаны защищают лежащий в основе вирусный белок от нейтрализации антителами . Это одна из наиболее плотно гликозилированных известных молекул, плотность которой достаточно высока, чтобы предотвратить нормальный процесс созревания гликанов во время биогенеза в эндоплазматическом ретикулуме и аппарате Гольджи . [ 18 ] [ 19 ] Таким образом, большинство гликанов остаются незрелыми гликанами с высоким содержанием маннозы, которые обычно не присутствуют на секретируемых или клеточных поверхностных гликопротеинах человека. [ 20 ] Необычная обработка и высокая плотность означают, что почти все идентифицированные к настоящему времени нейтрализующие антитела широкого спектра действия (у части пациентов, инфицированных в течение многих месяцев или лет) связываются с этими гликанами оболочки или адаптированы для борьбы с ними. [ 21 ]
Молекулярная структура вирусного шипа теперь определена с помощью рентгеновской кристаллографии. [ 22 ] и криоэлектронная микроскопия . [ 23 ] Эти достижения в структурной биологии стали возможными благодаря разработке стабильных рекомбинантных форм вирусного спайка путем введения межсубъединичной дисульфидной связи и мутации изолейцина в пролин в gp41. [ 24 ] Так называемые тримеры SOSIP не только воспроизводят антигенные свойства нативного вирусного шипа, но также демонстрируют ту же степень незрелых гликанов, что и нативный вирус. [ 25 ] Рекомбинантные тримерные вирусные шипы являются многообещающими кандидатами на вакцины, поскольку они содержат меньше ненейтрализующих эпитопов , чем рекомбинантный мономерный gp120, который подавляет иммунный ответ на целевые эпитопы. [ 26 ]
Организация генома
[ редактировать ]
ВИЧ имеет несколько основных генов, кодирующих структурные белки, которые встречаются во всех ретровирусах, а также несколько неструктурных («дополнительных») генов, уникальных для ВИЧ. [ 27 ] Геном ВИЧ содержит девять генов, кодирующих пятнадцать вирусных белков. [ 28 ] Они синтезируются в виде полипротеинов, которые производят белки для внутренней части вириона, называемые Gag, группоспецифический антиген; вирусные ферменты (Pol, полимераза) или гликопротеины env (оболочки) вириона. [ 29 ] Помимо этого, ВИЧ кодирует белки, которые также выполняют определенные регуляторные и вспомогательные функции. [ 29 ] ВИЧ-1 имеет два важных регуляторных элемента: Tat и Rev, а также несколько важных вспомогательных белков, таких как Nef, Vpr, Vif и Vpu, которые не необходимы для репликации в определенных тканях. [ 29 ] Ген gag обеспечивает базовую физическую инфраструктуру вируса, а pol обеспечивает основной механизм размножения ретровирусов, в то время как другие помогают ВИЧ проникнуть в клетку-хозяина и усилить ее размножение. Хотя они могут быть изменены в результате мутации, все эти гены, кроме tev, существуют во всех известных вариантах ВИЧ; см. Генетическая изменчивость ВИЧ . [ нужна ссылка ]
ВИЧ использует сложную систему дифференциального сплайсинга РНК для получения девяти различных генных продуктов из генома размером менее 10 т.п.н. [ 30 ] ВИЧ имеет несплайсированный геномный транскрипт размером 9,2 КБ, который кодирует предшественники gag и pol; однократно сплайсированная мРНК размером 4,5 т.п.н., кодирующая env, Vif, Vpr и Vpu, и мРНК с множественным сплайсингом длиной 2 т.п.н., кодирующая Tat, Rev и Nef. [ 30 ]
| Сорт | Имя гена | Первичные белковые продукты | Переработанные белковые продукты |
|---|---|---|---|
| Вирусные структурные белки | кляп | Кляпный полипротеин | МА, КА, СП1, НК, СП2, П6 |
| pol | Пол полипротеин | RT, РНКаза H, ИН, PR | |
| окружение | gp160 | гп120, гп41 | |
| Основные регуляторные элементы | отдал | Гат | |
| оборот | Преподобный | ||
| Акцессорные регуляторные белки | неф | Неф | |
| впр | Впр | ||
| виф | Виф | ||
| ВПУ | Впу |
Вирусные структурные белки
[ редактировать ]
- gag (группоспецифический антиген) кодирует полипротеин- предшественник gag , который процессируется вирусной протеазой во время созревания до MA ( матричный белок , p17); СА (капсидный белок, р24 ); SP1 (спейсерный пептид 1, р2); NC ( белок нуклеокапсида , p7); SP2 (спейсерный пептид 2, p1) и белок P6. [ 31 ]
- pol кодирует вирусные ферменты обратной транскриптазы (RT) и РНКазы H , интегразы (IN) и протеазы ВИЧ (PR). [ 29 ] Протеаза ВИЧ необходима для расщепления полипротеина-предшественника Gag для производства структурных белков, RT необходима для транскрипции ДНК с матрицы РНК, а IN необходима для интеграции двухцепочечной вирусной ДНК в геном хозяина. [ 27 ]
- env (от «конверт») кодирует gp160 , который расщепляется протеазой хозяина, фурином , внутри эндоплазматической сети клетки-хозяина. В результате посттрансляционного процессинга образуется поверхностный гликопротеин gp120 или SU, который прикрепляется к рецепторам CD4 , присутствующим на лимфоцитах, и gp41 или TM, который встраивается в вирусную оболочку, позволяя вирусу прикрепляться к клеткам-мишеням и сливаться с ними. [ 27 ] [ 31 ]
Основные регуляторные элементы
[ редактировать ]- tat (транс-активатор ВИЧ) играет важную роль в регуляции обратной транскрипции РНК вирусного генома, обеспечивая эффективный синтез вирусных мРНК и регуляцию высвобождения вирионов из инфицированных клеток. [ 29 ] Tat экспрессируется как Tat из 72 аминокислот одноэкзонный , а также двухэкзонный Tat из 86–101 аминокислот и играет важную роль на ранних стадиях ВИЧ-инфекции. Tat (14–15 кДа) связывается с выпуклой вторичной структурой «стебель-петля» геномной РНК вблизи 5'- области LTR, образуя элемент ответа трансактивации (TAR) . [ 9 ] [ 29 ]
- rev (регулятор экспрессии белков вириона): белок Rev связывается с вирусным геномом посредством богатого аргинином РНК-связывающего мотива, который также действует как NLS ( сигналы ядерной локализации ), необходимые для транспортировки Rev в ядро из цитозоля. во время репликации вируса. [ 29 ] Rev распознает сложную структуру «стебель-петля» мРНК env, расположенную в интрон -разделяющем кодирующем экзоне Tat и Rev, известную как элемент ответа Rev ВИЧ (RRE). [ 9 ] [ 29 ] Rev важен для синтеза основных вирусных белков и, следовательно, необходим для репликации вируса . [ нужна ссылка ]
Акцессорные регуляторные белки
[ редактировать ]- vpr ( белок R лентивируса ): Vpr представляет собой связанный с вирионом нуклеоцитоплазматического челнока регуляторный белок . [ 29 ] Считается, что он играет важную роль в репликации вируса, в частности, в ядерном импорте преинтеграционного комплекса . Vpr также, по-видимому, заставляет клетки-хозяева останавливать клеточный цикл в фазе G2 . Этот арест активирует механизм восстановления ДНК хозяина, что может обеспечить интеграцию вирусной ДНК. [ 9 ] ВИЧ-2 и SIV кодируют дополнительный белок, родственный Vpr, называемый Vpx, который функционирует совместно с Vpr. [ 29 ]
- vif – Vif представляет собой высококонсервативный фосфопротеин 23 кДа, массой важный для инфекционности вирионов ВИЧ-1 в зависимости от типа клеток. [ 9 ] Было обнаружено, что ВИЧ-1 требует Vif для синтеза инфекционных вирусов в лимфоцитах , макрофагах и некоторых клеточных линиях человека . Похоже, что Vif не требуется для того же процесса в HeLa клетках или клетках COS и других. [ 29 ]
- nef – Nef, отрицательный фактор, представляет собой N-концевой миристоилированный мембраносвязанный фосфопротеин. Он участвует во многих функциях во время цикла репликации вируса. Считается, что он играет важную роль в апоптозе клеток и повышении инфекционности вируса . [ 29 ]
- vpu (вирусный белок U) – Vpu специфичен для ВИЧ-1. Это олигомерный интегральный мембранный фосфопротеин класса I с многочисленными биологическими функциями. Vpu участвует в CD4 деградации пути убиквитина с участием протеасомного , а также в успешном высвобождении вирионов из инфицированных клеток. [ 9 ] [ 29 ]
- tev : Этот ген присутствует только в нескольких изолятах ВИЧ-1. Это слияние частей генов tat , env и rev и кодирует белок с некоторыми свойствами tat , но с небольшим количеством свойств rev или вообще без них . [ 32 ]
Вторичная структура РНК
[ редактировать ]| HIV pol-1 stem loop | |
|---|---|
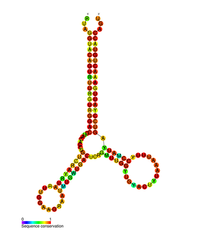 Прогнозируемая вторичная структура стволовой петли ВИЧ pol-1 | |
| Идентификаторы | |
| Символ | pol |
| Рфам | RF01418 |
| Другие данные | |
| РНК Тип | СНГ-рег. |
| PDB Структуры | ПДБе |
несколько консервативных элементов вторичной структуры РНК ВИЧ было идентифицировано В геноме . Структуры РНК вируса ВИЧ регулируют развитие обратной транскрипции. [ 33 ] Структура 5'UTR состоит из ряда структур «стебель-петля», соединенных небольшими линкерами. [ 10 ] Эти петли-стебли (от 5' до 3') включают элемент области транс-активации (TAR), 5'- сигнал полиаденилирования [поли(А)], PBS, DIS, основную SD и структуру шпильки ψ , расположенную внутри 5'-конец генома и элемент ответа ВИЧ Rev (RRE) в гене env. [ 10 ] [ 34 ] [ 35 ] Другая идентифицированная структура РНК — это петля стебля gag 3 (GSL3) , которая, как полагают, участвует в упаковке вируса. [ 36 ] [ 37 ] Было высказано предположение, что вторичные структуры РНК влияют на жизненный цикл ВИЧ, изменяя функцию протеазы ВИЧ и обратной транскриптазы , хотя не всем идентифицированным элементам была присвоена определенная функция. [ нужна ссылка ]
Было показано , что вторичная структура РНК, определенная с помощью анализа SHAPE, содержит три петли ствола и расположена между генами протеазы ВИЧ и генами обратной транскриптазы. эта цис- регуляторная РНК консервативна во всем семействе ВИЧ и, как полагают, влияет на жизненный цикл вируса. Было показано, что [ 38 ]
Петля V3
[ редактировать ]Третья переменная петля или петля V3 является частью или областью вируса иммунодефицита человека . Петля V3 гликопротеина оболочки вирона, gp120 , позволяет ему инфицировать иммунные клетки человека путем связывания с рецептором цитокина на целевой иммунной клетке человека, такой как клетка CCR5 или клетка CXCR4 , в зависимости от штамма ВИЧ . [ 39 ] Гликопротеин оболочки (Env) gp 120/41 необходим для проникновения ВИЧ-1 в клетки. Env служит молекулярной мишенью для лекарств, лечащих людей с ВИЧ-1, и источником иммуногена для разработки вакцины против СПИДа. Однако структура функционального тримера Env остается неясной. [ 40 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Барре-Синусси Ф, Шерманн Х.К., Рей Ф., Нугейр М.Т., Шамаре С., Грюст Дж., Доге С., Экслер-Блен С., Везине-Брюн Ф., Рузиу К., Розенбаум В., Монтанье Л. (май 1983 г.). «Выделение Т-лимфотропного ретровируса от пациента с риском развития синдрома приобретенного иммунодефицита (СПИД)». Наука . 220 (4599): 868–71. Бибкод : 1983Sci...220..868B . дои : 10.1126/science.6189183 . ПМИД 6189183 . S2CID 390173 .
- ^ Галло Р.К., Сарин П.С., Гельманн Е.П., Роберт-Гурофф М., Ричардсон Э., Кальянараман В.С., Манн Д., Сидху Г.Д., Шталь Р.Э., Золла-Пазнер С., Лейбович Дж., Попович М. (май 1983 г.). «Выделение вируса Т-клеточного лейкоза человека при синдроме приобретенного иммунодефицита (СПИД)». Наука . 220 (4599): 865–7. Бибкод : 1983Sci...220..865G . дои : 10.1126/science.6601823 . ПМИД 6601823 .
- ^ Чури С., Росс М.В. (2015). «ВИЧ/СПИД» . В Уилехан П., Болин А. (ред.). Международная энциклопедия сексуальности человека . Уайли. ISBN 9781405190060 . OCLC 949701914 .
- ^ Центры по контролю заболеваний (июнь 1981 г.). «Пневмоцистная пневмония - Лос-Анджелес». ММВР. Еженедельный отчет о заболеваемости и смертности . 30 (21): 250–2. ПМИД 6265753 .
- ^ Центры по контролю заболеваний (CDC) (июль 1981 г.). «Саркома Капоши и пневмоцистная пневмония среди гомосексуальных мужчин – Нью-Йорк и Калифорния» (PDF) . ММВР. Еженедельный отчет о заболеваемости и смертности . 30 (25): 305–8. ПМИД 6789108 . Архивировано из оригинала 22 октября 2012 года . Проверено 15 сентября 2017 г.
{{cite journal}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Уоттс Дж. М., Данг К. К., Горелик Р. Дж., Леонард К. В., Бесс Дж. В., Суонстром Р., Берч К. Л., Уикс К. М. (август 2009 г.). «Архитектура и вторичная структура всего генома РНК ВИЧ-1» . Природа . 460 (7256): 711–6. Бибкод : 2009Natur.460..711W . дои : 10.1038/nature08237 . ПМЦ 2724670 . ПМИД 19661910 .
- ^ Перейти обратно: а б Ли Дж., Де Клерк Э. (сентябрь 2016 г.). «Полногеномные белковые ассоциации ВИЧ: обзор 30-летних исследований» . Обзоры микробиологии и молекулярной биологии . 80 (3): 679–731. дои : 10.1128/MMBR.00065-15 . ПМЦ 4981665 . ПМИД 27357278 .
- ^ Синглтон П., Сэйнсбери Д., ред. (2006). "ВИЧ" . Словарь микробиологии и молекулярной биологии (3-е изд.). Хобокен, Нью-Джерси: Уайли. ISBN 9780470035450 . OCLC 71223221 .
- ^ Перейти обратно: а б с д и ж г Монтанье Л. (1999). «Вирусы иммунодефицита человека (Retroviridae)». Энциклопедия вирусологии (2-е изд.). стр. 763–774.
- ^ Перейти обратно: а б с Лу К., Хэн Икс, Саммерс М.Ф. (июль 2011 г.). «Структурные детерминанты и механизм упаковки генома ВИЧ-1» . Журнал молекулярной биологии . 410 (4): 609–33. дои : 10.1016/j.jmb.2011.04.029 . ПМК 3139105 . ПМИД 21762803 .
- ^ Перейти обратно: а б с д Мур, Майкл Д.; Ху, Вэй Шау (2009). «Димеризация РНК ВИЧ-1: для танго нужны двое» . Обзоры по СПИДу . 11 (2): 91–102. ISSN 1139-6121 . ПМК 3056336 . ПМИД 19529749 .
- ^ Хван С.К., Сваровская Е.С., Патхак В.К. (октябрь 2001 г.). «Выбор динамической копии: устойчивое состояние между полимеразой вируса мышиного лейкоза и полимеразно-зависимой активностью РНКазы H определяет частоту переключения матрицы in vivo» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (21): 12209–14. Бибкод : 2001PNAS...9812209H . дои : 10.1073/pnas.221289898 . ПМК 59793 . ПМИД 11593039 .
- ^ Уэйн-Хобсон С., Сониго П., Данос О., Коул С., Ализон М. (январь 1985 г.). «Нуклеотидная последовательность вируса СПИДа, LAV». Клетка . 40 (1): 9–17. дои : 10.1016/0092-8674(85)90303-4 . ПМИД 2981635 . S2CID 33055050 .
- ^ Ратнер Л., Хазелтин В., Патарка Р., Ливак К.Дж., Старчич Б., Джозефс С.Ф., Доран Э.Р., Рафальски Дж.А., Уайтхорн Э.А., Баумайстер К. (1985). «Полная нуклеотидная последовательность вируса СПИДа, HTLV-III». Природа . 313 (6000): 277–84. Бибкод : 1985Natur.313..277R . дои : 10.1038/313277a0 . ПМИД 2578615 . S2CID 4316242 .
- ^ Перейти обратно: а б Кастелли Дж.К., Леви А. (2002). «ВИЧ (вирус иммунодефицита человека)». Энциклопедия рака . Том. 2 (2-е изд.). стр. 407–415.
- ^ Чекли М.А., Фрид Э.О. (22 июля 2011 г.). «Биосинтез, торговля и внедрение гликопротеина оболочки ВИЧ-1» . Журнал молекулярной биологии . 410 (4): 582–608. дои : 10.1016/j.jmb.2011.04.042 . ПМК 3139147 . ПМИД 21762802 .
- ^ Национальный институт здравоохранения (17 июня 1998 г.). «Кристаллическая структура ключевого белка ВИЧ открывает новые цели профилактики и лечения» (пресс-релиз). Архивировано из оригинала 19 февраля 2006 года . Проверено 14 сентября 2006 г.
- ^ Беренс А.Дж., Василевич С., Притчард Л.К., Харви Д.Дж., Андев Р.С., Крумм С.А., Струве В.Б., Купо А., Кумар А., Зитцманн Н., Сибрайт Г.Е., Крамер Х.Б., Спенсер Д.И., Ройл Л., Ли Дж.Х., Класс П.Дж., Бертон Д.Р. , Уилсон И.А., Уорд А.Б., Сандерс Р.В., Мур Дж.П., Дорес К.Дж., Криспин М. (март 2016 г.). «Состав и антигенные эффекты отдельных гликановых участков тримерного гликопротеина оболочки ВИЧ-1» . Отчеты по ячейкам . 14 (11): 2695–706. дои : 10.1016/j.celrep.2016.02.058 . ПМЦ 4805854 . ПМИД 26972002 .
- ^ Притчард Л.К., Спенсер Д.И., Ройл Л., Бономелли С., Сибрайт Дж.Е., Беренс А.Дж., Кулп Д.В., Менис С., Крамм С.А., Данлоп Д.С., Криспин Д.Дж., Боуден Т.А., Сканлан К.Н., Уорд А.Б., Шиф У.Р., Дорес К.Дж., Криспин М. (июнь 2015 г.). «Кластеризация гликанов стабилизирует маннозный участок ВИЧ-1 и сохраняет уязвимость к нейтрализующим антителам широкого спектра действия» . Природные коммуникации . 6 : 7479. Бибкод : 2015NatCo...6.7479P . дои : 10.1038/ncomms8479 . ПМЦ 4500839 . ПМИД 26105115 .
- ^ Притчард Л.К., Харви DJ, Бономелли С., Криспин М., Дорес К.Дж. (сентябрь 2015 г.). «Клеточно- и белково-направленное гликозилирование нативной расщепленной оболочки ВИЧ-1» . Журнал вирусологии . 89 (17): 8932–44. дои : 10.1128/JVI.01190-15 . ПМК 4524065 . ПМИД 26085151 .
- ^ Криспин М., Дорес К.Дж. (апрель 2015 г.). «Нацеливание гликанов, полученных от хозяина, на вирусы с оболочкой для разработки вакцин на основе антител» . Современное мнение в вирусологии . Вирусный патогенез • Профилактические и лечебные вакцины. 11 : 63–9. дои : 10.1016/j.coviro.2015.02.002 . ПМЦ 4827424 . ПМИД 25747313 .
- ^ Жюльен Дж.П., Купо А., Сок Д., Стэнфилд Р.Л., Люмкис Д., Деллер М.К., Класс П.Дж., Бертон Д.Р., Сандерс Р.В., Мур Дж.П., Уорд А.Б., Уилсон И.А. (декабрь 2013 г.). «Кристаллическая структура растворимого расщепленного тримера оболочки ВИЧ-1» . Наука . 342 (6165): 1477–83. Бибкод : 2013Sci...342.1477J . дои : 10.1126/science.1245625 . ПМЦ 3886632 . ПМИД 24179159 .
- ^ Люмкис Д., Жюльен Дж.П., де Валь Н., Купо А., Поттер К.С., Класс П.Дж., Бертон Д.Р., Сандерс Р.В., Мур Дж.П., Каррагер Б., Уилсон И.А., Уорд А.Б. (декабрь 2013 г.). «Крио-ЭМ структура полностью гликозилированного растворимого расщепленного тримера оболочки ВИЧ-1» . Наука . 342 (6165): 1484–90. Бибкод : 2013Sci...342.1484L . дои : 10.1126/science.1245627 . ПМЦ 3954647 . ПМИД 24179160 .
- ^ Сандерс Р.В., Деркинг Р., Купо А., Жюльен Дж.П., Ясмин А., де Валь Н., Ким Х.Дж., Блаттнер С., де ла Пенья А.Т., Корсун Дж., Голабек М., де Лос Рейес К., Кетас Т.Дж., ван Гилс М.Дж., Кинг Ч.Р. , Уилсон И.А., Уорд А.Б., Класс П.Дж., Мур Дж.П. (сентябрь 2013 г.). «Расщепленный растворимый тример Env ВИЧ-1 нового поколения, BG505 SOSIP.664 gp140, экспрессирует множество эпитопов для широко нейтрализующих, но не ненейтрализующих антител» . ПЛОС Патогены . 9 (9): e1003618. дои : 10.1371/journal.ppat.1003618 . ПМЦ 3777863 . ПМИД 24068931 .
- ^ Притчард Л.К., Василевич С., Озоровский Г., Сибрайт Дж.Е., Купо А., Ринг Р., Ким Х.Дж., Сандерс Р.В., Дорес К.Дж., Бертон Д.Р., Уилсон И.А., Уорд А.Б., Мур Дж.П., Криспин М. (июнь 2015 г.). «Структурные ограничения определяют гликозилирование тримеров оболочки ВИЧ-1» . Отчеты по ячейкам . 11 (10): 1604–13. дои : 10.1016/j.celrep.2015.05.017 . ПМЦ 4555872 . ПМИД 26051934 .
- ^ де Тайе С.В., Озоровский Г., Торрентс оф Рок А., Гуттман М., Жюльен Дж.П., ван ден Керкхоф Т.Л., Бургер Дж.А., Притчард Л.К., Пугач П., Ясмин А., Крэмптон Дж., Ху Дж., Бонтьер И., Торрес Дж.Л., Арендт Х. Посмотреть статью PubMed PubMed Central Google Scholar, ДеСтефано Дж., Кофф В.К., Шуйтемейкер Х., Эггинка Д., Беркхаут Б., Дин. Х., ЛаБранш С., Кротти С., Криспин М., Монтефиори Д.С., Класс П.Дж., Ли К.К., Мур Дж.П., Уилсон И.А., Уорд А.Б., Сандерс Р.В. (декабрь 2015 г.). «Иммуногенность стабилизированных тримеров оболочки ВИЧ-1 с уменьшенным воздействием ненейтрализующих эпитопов» . Ячейка 163 (7): 1702–15. дои : 10.1016/j.cell.2015.11.056 . ПМЦ 4732737 . ПМИД 26687358 .
- ^ Перейти обратно: а б с Мушахвар ИК (2007). «Вирусы иммунодефицита человека: молекулярная вирусология, патогенез, диагностика и лечение». Перспективы медицинской вирусологии . 13 : 75–87. дои : 10.1016/S0168-7069(06)13005-0 . ISBN 9780444520739 .
- ^ Ли Дж., Пьямпонгсант С., Фариа Н.Р., Воет А., Пинеда-Пенья А.С., Хури Р., Леми П., Вандамм А.М., Тейс К. (февраль 2015 г.). «Интегрированная карта вариаций всего генома ВИЧ с популяционной точки зрения» . Ретровирусология . 12 (1): 18. дои : 10.1186/s12977-015-0148-6 . ПМЦ 4358901 . ПМИД 25808207 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Воттелер Дж, Шуберт У (2008). «Вирусы иммунодефицита человека: молекулярная биология». Энциклопедия вирусологии (3-е изд.). стр. 517–525.
- ^ Перейти обратно: а б Фейнберг Марк Б., Грин Уорнер С. (1992). «Молекулярный взгляд на патогенез вируса иммунодефицита человека типа 1». Современное мнение в иммунологии . 4 (4): 466–474. дои : 10.1016/s0952-7915(06)80041-5 . ПМИД 1356348 .
- ^ Перейти обратно: а б Кинг Стивен Р. (1994). «ВИЧ: вирусология и механизмы заболевания». Анналы неотложной медицины . 24 (3): 443–449. дои : 10.1016/s0196-0644(94)70181-4 . ПМИД 7915889 .
- ^ Бенко Д.М., Шварц С., Павлакис Г.Н., Фельбер Б.К. (июнь 1990 г.). «Новый белок типа 1 вируса иммунодефицита человека, tev, имеет общие последовательности с белками tat, env и rev» . Журнал вирусологии . 64 (6): 2505–18. doi : 10.1128/JVI.64.6.2505-2518.1990 . ПМК 249426 . ПМИД 2186172 .
- ^ Крупкин М., Джексон Л.Н., Ха Б., Пуглиси Е.В. (декабрь 2020 г.). «Достижения в понимании инициации обратной транскрипции ВИЧ-1» . Curr Opin Struct Biol . 65 : 175–183. дои : 10.1016/j.sbi.2020.07.005 . ПМЦ 9973426 . ПМИД 32916568 . S2CID 221636459 .
- ^ Берхаут Б (январь 1992 г.). «Структурные особенности TAR РНК вирусов иммунодефицита человека и обезьян: филогенетический анализ» . Исследования нуклеиновых кислот . 20 (1): 27–31. дои : 10.1093/нар/20.1.27 . ПМК 310321 . ПМИД 1738599 .
- ^ Пайяр Дж.К., Скрипкин Э., Эресманн Б., Эресманн С., Марке Р. (февраль 2002 г.). «Доказательства in vitro существования псевдоузла большого радиуса действия в 5'-нетранслируемых и матричных кодирующих областях геномной РНК ВИЧ-1» . Журнал биологической химии . 277 (8): 5995–6004. дои : 10.1074/jbc.M108972200 . ПМИД 11744696 .
- ^ Дамгаард К.К., Андерсен Э.С., Кнудсен Б., Городкин Дж., Кьемс Дж. (февраль 2004 г.). «Взаимодействия РНК в 5'-области генома ВИЧ-1» Журнал молекулярной биологии . 336 (2): 369–79. дои : 10.1016/j.jmb.2003.12.010 . ПМИД 14757051 .
- ^ Ронг Л., Рассел Р.С., Ху Дж., Логри М., Вайнберг М.А., Лян С. (сентябрь 2003 г.). «Удаление стволовой петли 3 компенсируется мутациями второго сайта в белке Gag вируса иммунодефицита человека типа 1». Вирусология . 314 (1): 221–8. дои : 10.1016/S0042-6822(03)00405-7 . ПМИД 14517075 .
- ^ Ван Ц, Барр И, Го Ф, Ли С (декабрь 2008 г.). «Доказательства новой вторичной структуры РНК в кодирующей области гена pol ВИЧ-1» . РНК . 14 (12): 2478–88. дои : 10.1261/rna.1252608 . ПМК 2590956 . ПМИД 18974280 .
- ^ «Взаимодействие петли V3 gp120 различных штаммов ВИЧ-1 с мощным человеческим моноклональным антителом против ВИЧ 447-52D» . Научный институт Вейцмана: Отдел структурной биологии . Архивировано из оригинала 18 июля 2007 г. Проверено 18 апреля 2017 г.
- ^ Такеда С., Такидзава М., Мияучи К., Урано Э., Фуджино М., Мураками Т., Мураками Т., Комано Дж. (июнь 2016 г.). «Конформационные свойства третьей переменной петли оболочечного гликопротеина ВИЧ-1AD8 в лигандированных условиях». Связь с биохимическими и биофизическими исследованиями . 475 (1): 113–8. дои : 10.1016/j.bbrc.2016.05.051 . ПМИД 27178216 .
Внешние ссылки
[ редактировать ]- Запись Rfam для стволовой петли ВИЧ pol-1
- 3D-модель полного вириона ВИЧ1
- Лю Дж., Райт Э.Р., Винклер Х. (2010). «3D-визуализация вирионов ВИЧ методом криоэлектронной томографии». Крио-ЭМ, Часть C: Анализ, интерпретация и тематические исследования . Методы энзимологии. Том. 483. стр. 267–90. дои : 10.1016/S0076-6879(10)83014-9 . ISBN 9780123849939 . ПМК 3056484 . ПМИД 20888479 .