BRAF (ген)
| Брут | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | BRAF , B-RAF1, BRAF1, NS7, RAFB1, B-RAF, B-RAF Protoncogene, Serine/Threoninine Kinase | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Омим : 164757 ; MGI : 88190 ; Гомологен : 3197 ; Genecards BRAF ; OMA : BRAF - ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||




BRAF -это человеческий ген , который кодирует белок, называемый B-RAF. Ген также называется протоонкогеном B-RAF и V-RAF мышиной саркомы-саркомы B-гомолога B , в то время как белок более формально известен как серин/треонин-белка киназа B-RAF . [ 5 ] [ 6 ]
Белок B-RAF участвует в отправке сигналов внутри клеток, которые участвуют в направлении роста клеток . В 2002 году было показано, что он мутировал при некоторых видах рака человека . [ 7 ]
Некоторые другие унаследованные мутации BRAF вызывают врожденные дефекты.
лекарства, которые лечат рак, вызванные мутациями BRAF Были разработаны . Два из этих препаратов, Вемурафениб [ 8 ] и дабрафениб , одобрены FDA для лечения меланомы поздней стадии. Vemurafenib был первым утвержденным препаратом, выходящим из фрагментных лекарств . [ 9 ]
Функция
[ редактировать ]B-RAF является членом семейства RAF-киназы протеинкиназ передачи сигнала трансдукции . Этот белок играет роль в регуляции MAP -киназы / ERKS сигнального пути , который влияет на деление клеток , дифференцировку и секрецию. [ 10 ]
Структура
[ редактировать ]B-RAF является 766- аминокислотой , регулируемой передачей сигнала сериновой/треониновой специфической протеинкиназы . Вообще говоря, он состоит из трех консервативных доменов, характерных для семейства киназы RAF : консервативный регион 1 (CR1), RAS - GTP -связывание [ 11 ] Саморегуляторный домен, консервативная область 2 (CR2), релишная петля и консервативная область 3 (CR3), домен каталитической протеинкиназы , который фосфорилирует консенсусную последовательность на белковых субстратах. [ 12 ] В своей активной конформации B-RAF образует димеры посредством водородного связующего и электростатических взаимодействий своих киназных доменов. [ 13 ]
CR1
[ редактировать ]Консервативный регион 1 (CR1) аутоингибирует киназный домен B-RAF (CR3), так что передача сигналов B-RAF регулируется, а не конститутивная. [ 12 ] Остатки 155–227 [ 14 ] Составьте домен -связывающий RAS доменом RAS -GTP, (RBD), который связывается с эффекторным чтобы высвободить ингибирование киназы CR1 и останавливается. Остатки 234–280 составляют фарболом / DAG с мотив цинкового пальца , который участвует в стыковке мембраны B-RAF после RAS-связывания. [ 14 ] [ 15 ]
CR2
[ редактировать ]Консервативный регион 2 (CR2) предоставляет гибкий линкер, который соединяет CR1 и CR3 и действует как шарнир. [ 16 ]
CR3
[ редактировать ]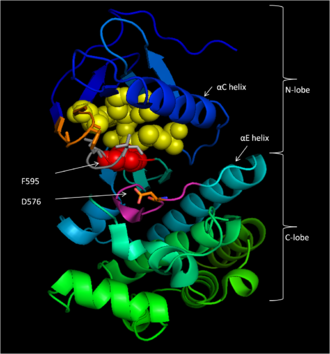
Консервативный регион 3 (CR3), остатки 457–717, [ 14 ] Создает ферментативный киназный домен B-Raf. Эта в основном консервативная структура [ 17 ] это биолобал, соединен с помощью короткой области петли. [ 18 ] Меньший N -Lobe (остатки 457–530) в первую очередь ответственна за связывание АТФ , в то время как более крупный C -лоб (остатки 535–717) связывает субстратные белки. [ 17 ] Активный сайт представляет собой расщелину между двумя долями, а каталитический остаток ASP 576 расположен на C-болоте, обращаясь к внутренней части этой расщелины. [ 14 ] [ 17 ]
Субрегионы
[ редактировать ]P-петля
[ редактировать ]P -петля B-RAF (остатки 464–471) стабилизирует непередаваемые фосфатные группы АТФ во время ферментного АТФ-связывания. В частности, S 467, F 468 и G 469 BACKBONE AMIDES -связи водорода в β-фосфат АТФ для закрепления молекулы. Функциональные мотивы B-RAF были определены путем анализа гомологии PKA, проанализированной Hanks и Hunter в домен B-Raf-киназы. [ 17 ]
Нуклеотидсвязывающий карман
[ редактировать ]V 471, C 532, W 531, T 529, L 514 и 481 образуют гидрофобный карман, в котором аденин АТФ закрепляется через аттракционы Ван -дер -Ваальса при связывании АТФ. [ 17 ] [ 19 ]
Каталитическая петля
[ редактировать ]Остатки 574–581 сочиняют участок киназного домена, ответственный за поддержку переноса γ-фосфата АТФ в белковый субстрат B-RAF. В частности, D 576 действует как акцептор протона для активации нуклеофильного гидроксилового кислорода на остатках серина или треонина субстрата, что позволяет возникать реакция переноса фосфата, опосредованную основным катализом . [ 17 ]
DFG Motif
[ редактировать ]D 594, F 595 и G 596 составляют мотив, центральный для функции B-RAF как в его неактивном, так и в активном состоянии. В неактивном состоянии F595 занимает нуклеотидсвязывающий карман, запрещая АТФ входить и уменьшать вероятность катализа фермента. [ 13 ] [ 19 ] [ 20 ] В активном состоянии D594 хелатирует дивалентный , который стабилизирует β- и γ - магния катион фосфатные группы АТФ, ориентируя γ-фосфат для переноса. [ 17 ]
Петля активации
[ редактировать ]Остатки 596–600 образуют сильные гидрофобные взаимодействия с P-петли в неактивной конформации киназы, блокируя киназу в ее неактивном состоянии до тех пор, пока цикл активации не будет фосфорилирован, дестабилизируя эти взаимодействия с присутствием негативного заряда. Это вызывает переход к активному состоянию киназы. В частности, L597 и V600 петли активации взаимодействуют с G466, F468 и V471 P-петли, чтобы сохранить киназный домен неактивным, пока он не будет фосфорилирован. [ 18 ]
Фермология
[ редактировать ]B-RAF является специфичной для серина/треонина протеинкиназы . Таким образом, он катализирует фосфорилирование остатков серина и треонина в консенсусной последовательности на целевых белках с помощью АТФ , давая ADP и фосфорилированный белок в качестве продуктов. [ 17 ] Поскольку это высокорегулируемая передачи сигнала киназа , B -RAF должен сначала связывать Ras - GTP, прежде чем стать активным в качестве фермента. [ 15 ] Как только B-RAF активируется, консервативная протеинкиназа каталитическая ядра фосфорилирует протеиновые субстраты, способствуя нуклеофильным атаке активированного субстрата серина или треонинового гидроксильного атома кислорода на γ-фосфатную группу АТФ посредством бимолекулярного нуклеофильного замещения . [ 17 ] [ 21 ] [ 22 ] [ 23 ]
Активация
[ редактировать ]Облегчение автоингибирования CR1
[ редактировать ]Домен киназы (CR3) человеческих RAF -киназ ингибируется двумя механизмами: аутоингибирование помощью собственного регуляторного домена RAS -GTP с -связывания CR1 и отсутствие посттрансляционного фосфорилирования ключевых остатков серине и тирозина (S338 и Y341 для c -raf ) в шарнирной области CR2. домен белка в белках Во время активации B-RAF в сначала эффекторный домен RAS-GTP связывает эффекторный с RAS-связывающим доменом CR1 (RBD), чтобы высвободить домен киназы CR3, как и другие члены семейства RAF-киназы человека . Взаимодействие CR1 -RAS позже укрепляется посредством связывания поддожа, ришанного цистеина (CRD) CR1 с Ras и мембранными фосфолипидами . [ 12 ] В отличие от A-RAF и C-RAF , которые должны быть фосфорилированы на гидроксилсодержащих остатках CR2, прежде чем полностью выпустить CR1, чтобы стать активным, B-RAF конфигуративно фосфорилируется на CR2 S445. [ 24 ] Это позволяет отрицательно заряженному фосфосерину немедленно отталкивать CR1 через стерические и электростатические взаимодействия после того, как регуляторный домен не является несвязанным, освобождая домен киназы CR3 для взаимодействия с субстратными белками.
Активация домена CR3
[ редактировать ]После высвобождения регуляторного домена CR1, домен киназы B -RAF CR3 должен измениться на его ATP -связывающий активное конформер , прежде чем он сможет катализировать фосфорилирование белка . В неактивной конформации F595 мотива DFG блокирует гидрофобный адениновый связывающий карман, в то время как остатки петли активации образуют гидрофобные взаимодействия с P-петли, останавливая ATP от доступа к сайту связывания. Когда петля активации фосфорилируется, отрицательный заряд фосфата нестабилен в гидрофобной среде P-петли. В результате цикл активации изменяет конформацию , растянувшись через C-блоки киназного домена. В этом процессе он образует стабилизацию β-листовых взаимодействий с цепью β6. Между тем, фосфорилированный остаток приближается к K507, образуя стабилизирующий соляный мост, чтобы закрепить петлю активации. Мотив DFG изменяет конформацию с петлей активации, заставляя F595 выходить из сайта связывания адениновых нуклеотидов в гидрофобный карман, граничащий с αc и αe -спирали . Вместе DFG и движение петли активации при фосфорилировании открывают сайт связывания АТФ . Поскольку все другие субстрат-связывающие и каталитические домены уже находятся на месте, фосфорилирование одной петли активации только активирует киназный домен B-RAF посредством цепной реакции, которая по существу удаляет крышку из активного сайта, не подготовленного. [ 18 ]
Механизм катализа
[ редактировать ]
Чтобы эффективно катализировать фосфорилирование белка посредством бимолекулярной замены сериновых и треониновых остатков ADP в качестве уходной группы , B-RAF должен сначала связывать АТФ, а затем стабилизировать переходное состояние при передаче γ-фосфата АТФ. [ 17 ]
АТФ -связывание
[ редактировать ]B-RAF связывает АТФ, закрепляя адениновый нуклеотид в неполярном кармане (желтый, рис. 1) и ориентируя молекулу посредством водородного связывания и электростатических взаимодействий с фосфатными группами. В дополнение к связыванию фосфатов мотива P-петли и DFG, описанного выше, K483 и E501 играют ключевую роль в стабилизации непередаваемых фосфатных групп. Положительный заряд на первичный амин K483 позволяет ему стабилизировать отрицательный заряд на ATP α- и β-фосфатных группах, когда АТФ связывается. Когда АТФ отсутствует, отрицательный заряд карбоксильной группы E501 уравновешивает этот заряд. [ 17 ] [ 18 ]
Фосфорилирование
[ редактировать ]Как только АТФ связан с доменом киназы B-Raf, D576 каталитической петли активирует гидроксильную группу субстрата, увеличивая его нуклеофильность, чтобы кинетически привести к реакции фосфорилирования, в то время как другие остатки каталитической петли стабилизируют переходное состояние (рис. 2). N581 хелатирует дивалентный катион магния, связанный с АТФ, чтобы помочь ориентировать молекулу для оптимальной замены. K578 нейтрализует отрицательный заряд в γ-фосфатной группе АТФ, так что активированный остаток субстрата Ser/THR не будет испытывать столько электрон-электронного отталкивания при атаке фосфата. После переноса фосфатной группы выпускаются ADP и новый фосфопротеин. [ 17 ]
Ингибиторы
[ редактировать ]Поскольку конститутивно активные мутанты B-RAF обычно вызывают рак (см. Клиническую значимость) из-за чрезмерных сигнальных клеток для роста, были разработаны ингибиторы B-RAF как для неактивных, так и для активных конформаций киназного домена в качестве кандидатов на рак. [ 18 ] [ 19 ] [ 20 ]
Сарафениб
[ редактировать ]
Bay43-9006 ( Sorafenib , Nexavar)-это мутант B-RAF и ингибитор C-RAF V600E , одобренный FDA для лечения первичного рака печени и почек . Bay43-9006 отключает домен B-Raf- киназы , блокируя фермент в его неактивной форме. Ингибитор выполняет это, блокируя карман связывания АТФ через высокую аффинность к киназному домену. Затем он связывает остатки ключевой петли активации и мотива DFG, чтобы остановить движение цикла активации и мотива DFG к активной конформации. Наконец, трифторметиловый фенильный фрагмент стерически блокирует мотив DFG и петлю активации активного конформации, что делает невозможным, чтобы киназный домен мог сдвинуть конформацию, чтобы стать активной. [ 18 ]
Дистальное пиридиловое кольцо Bay43-9006 прикрепляет в гидрофобном нуклеотид-связывающем кармане киназы N-лобовой, взаимодействующей с W531, F583 и F595. Гидрофобные взаимодействия с каталитической петлей F583 и DFG Motif F595 стабилизируют неактивную конформацию этих структур, уменьшая вероятность активации фермента. Дальнейшее гидрофобное взаимодействие K483, L514 и T529 с центральным фенильным кольцом увеличивает аффинность киназного домена к ингибитору. Гидрофобное взаимодействие F595 с центральным кольцом также снижает энергетическую благоприятность переключателя конформации DFG. Наконец, полярные взаимодействия Bay43-9006 с киназной доменом продолжают эту тенденцию повышения сродства фермента к ингибитору и стабилизации остатков DFG в неактивной конформации. Водородная связь E501 и C532 Мочевина и пиридильные группы ингибитора соответственно, в то время как мочевины карбонил азота D594, принимает водородную связь из амидного чтобы заблокировать мотив DFG на месте. [ 18 ]
Трифторметиловый фенил-фрагмент цементирует термодинамическую благоприятность неактивной конформации, когда киназа связан с Bay43-9006, стерически блокируя гидрофобный карман между αC и αE-спирациями, что DFG-мотиф и петля активации живет при переключении на свои локации в Активная конформация белка. [ 18 ]
Рэмпхейниби
[ редактировать ]
PLX4032 ( Vemurafenib )-это ингибитор B-RAF мутанта V600 , одобренный FDA для лечения меланомы поздней стадии . [ 13 ] В отличие от Bay43-9006 , которая ингибирует неактивную форму киназного домена, Vemurafenib ингибирует активную форму «DFG-In» киназы, [ 19 ] [ 20 ] твердо прикрепляя себя на АТФ-связывающем сайте. Ингибируя только активную форму киназы, Вемурафениб избирательно ингибирует пролиферацию клеток с нерегулируемым B-RAF, обычно те, которые вызывают рак .
Поскольку Vemurafenib отличается только от своего предшественника PLX4720, в фенильном кольце, добавленном по фармакокинетическим причинам, [ 20 ] Режим действия PLX4720 эквивалентен Vemurafenib. PLX4720 обладает хорошим сродством к сайту связывания АТФ частично потому, что его якорная область, индольная бициклика с 7-азами , отличается только от естественного аденина, который занимает место в двух местах, где атомы азота были заменены углеродом. Это обеспечивает сильные межмолекулярные взаимодействия, такие как водородная связь N7 с водородной связью C532 и N1 с Q530, чтобы сохранить. Отличная посадка в гидрофобном кармане АТФ (C532, W531, T529, L514, A481) также увеличивает аффинность связывания. Кетононовой линкер водородной связи с водой и дифторо-фенилом во втором гидрофобном кармане (A481, V482, K483, V471, I527, T529, L514 и F583) способствуют исключительно высокому аффинту связывания в целом. Селективное связывание с активным RAF осуществляется терминальной пропильной группой, которая связывается с RAF-селективным карманом, созданным путем смещения спирали αC. Селективность для активной конформации киназы дополнительно увеличивается путем чувствительного к рН депротонированным Сульфонамидная группа, которая стабилизируется водородной связью с основным пептидом NH D594 в активном состоянии. В неактивном состоянии группа сульфонамидов ингибитора взаимодействует с основным карбонилом этого остатка, создавая отталкивание. Таким образом, Вемурафениб связывается преимущественно с активным состоянием киназного домена B-RAF. [ 19 ] [ 20 ]
Клиническое значение
[ редактировать ]Мутации в гене BRAF могут вызвать заболевание двумя способами. Во -первых, мутации могут быть унаследованы и вызвать врожденные дефекты. Во -вторых, мутации могут появиться позже в жизни и вызвать рак, как онкоген .
Унаследованные мутации в этом геном вызывают кардиофциококожный синдром , болезнь, характеризующееся дефектами сердца, умственной отсталости и отличительным внешним видом лица. [ 25 ]
Мутации в этом гене были обнаружены при раке, включая неходжкинскую лимфому , колоректальный рак , злокачественную меланому , папиллярную карциному щитовидной железы , немелкоклеточную карциному легких и плеоморфную ксантоастому, а также интуируют, а также интуматорома, а также как интуматорома, так же как интуматорома , а также интуматорома и плеоморфы, так же как интуматорома, а также интуируют, а также интуированные , а также интубационные , а также , аденокарциному легкого, опухоли мозга, включая глиобластому интуматорома и плеоморфная ксантоастома, а также интуируют и плеоморфные ксантоастроки , а также интуирующие Такие заболевания, как болезнь Эрдгейма - Честер . [ 10 ]
Мутация V600E гена BRAF была связана с лейкозом волосатой клеток в многочисленных исследованиях и была предложена для применения при скрининге на синдром Линча , чтобы уменьшить количество пациентов, перенесших ненужное секвенирование MLH1 . [ 26 ] [ 27 ]
Мутанты
[ редактировать ]более 30 мутаций гена BRAF Было выявлено , связанных с раком человека. Частота мутаций BRAF широко варьируется при раке человека, от более чем 80% в меланомах и невии , до всего лишь 0–18% в других опухолях , таких как 1–3% при раке легких и 5% при колоректальном раке . [ 28 ] В 90% случаев тимин заменяется аденином в нуклеотиде 1799. Это приводит к тому, что валин (v) заменяется глутаматом (E) в кодоне 600 (в настоящее время называется V600E ) в сегменте активации, который был обнаружен в человеческий рак. [ 29 ] Эта мутация широко наблюдалась при папиллярной карциноме щитовидной железы , колоректальном раке, меланоме и немелкоклеточном раке легких . [ 30 ] [ 31 ] [ 32 ] [ 33 ] [ 34 ] [ 35 ] [ 36 ] Мутация BRAF-V600E присутствует у 57% пациентов с гистиоцитозом клеток Лангерганса. [ 37 ] Мутация V600E является вероятной мутацией водителя в 100% случаев лейкоза волосатых клеток . [ 38 ] Высокая частота мутаций BRAF V600E была обнаружена при амелобластоме , доброкачественной, но местно -инфильтративной одонтогенной новообразовании. [ 39 ] Мутация V600E также может быть связана, как мутация одного водителя (генетическое «курящее оружие») с определенными случаями развития папиллярной черепниофарингиомы . [ 40 ]
Other mutations which have been found are R461I, I462S, G463E, G463V, G465A, G465E, G465V, G468A, G468E, G469R, N580S, E585K, D593V, F594L, G595R, L596V, T598I, V599D, V599E, V599K, V599R, V600K , A727V и т. Д. И большинство из этих мутаций кластеризованы до двух областей: богатая глицином P-петлей N доли и сегмента активации и фланкирующих областей. [ 18 ] Эти мутации изменяют сегмент активации от неактивного состояния в активное состояние, например, в предыдущей цитируемой статье сообщалось, что алифатическая боковая цепь Val599 взаимодействует с фенильным кольцом PHE467 в петле P. Замена гидрофобной боковой цепи среднего класса среднего размера большим и заряженным остатком, как можно было бы привести к раку человека (Glu, ASP, Lys или Arg) Сегмент активации в активную позицию. В зависимости от типа мутации активность киназы в отношении MEK также может различаться. Большинство мутантов стимулируют повышенную активность B-Raf- Kinase по отношению к MEK. Однако несколько мутантов действуют через другой механизм, потому что, хотя их активность в отношении MEK снижается, они принимают конформацию, которая активирует C-RAF дикого типа, который затем сигнализирует ERK .
NICE-V600E
[ редактировать ]- BRAF V600E является детерминантом чувствительности к ингибиторам протеасом . Уязвимость к ингибиторам протеасомы зависит от постоянной передачи сигналов BRAF, потому что блокада BRAF-V600E с помощью PLX4720 обратной чувствительности к карфилзомибу в клетках колоректального рака BRAF-мутант. Ингибирование протеасомы может представлять собой ценную стратегию нацеливания в колоректальных опухолях BRAF V600E-мутант. [ 41 ]
Ингибиторы BRAF
[ редактировать ]Как упомянуто выше, некоторые фармацевтические фирмы разрабатывают специфические ингибиторы мутированного белка B-RAF для противоопухолевого использования, поскольку BRAF является хорошо понятной, высокой мишенью. [ 19 ] [ 42 ] Vemurafenib США (RG7204 или PLX4032) был лицензирован Управлением по контролю за продуктами и лекарствами в качестве Zelboraf для лечения метастатической меланомы в августе 2011 года на основе клинических данных фазы III. Улучшенная выживаемость была замечена, а также уровень ответа на лечение 53% по сравнению с 7–12% с первым лучшим химиотерапевтическим лечением, дакарбазином . [ 43 ] В клинических испытаниях B-RAF увеличил вероятность выживания пациента с метастатической меланомой. Несмотря на высокую эффективность препарата, у 20% опухолей все еще развивается устойчивость к лечению. У мышей 20% опухолей становятся устойчивыми через 56 дней. [ 44 ] Хотя механизмы этого сопротивления все еще оспариваются, некоторые гипотезы включают сверхэкспрессию B-RAF для компенсации высоких концентраций Vemurafenib [ 44 ] и повышенная активация передачи сигналов роста. [ 45 ]
Более общие ингибиторы B-RAF включают GDC-0879, PLX-4720, Sorafenib , Dabrafenib и Encorafenib .
ингибиторы панрафа
[ редактировать ]Бельварафениб классифицируется как ингибитор Панрафа. Ингибитор Panraf блокирует каталитическую функцию обоих белков в димере. [ 46 ]
Взаимодействия
[ редактировать ]Было показано, что BRAF (ген) взаимодействует :
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в GRCH38: Ensembl Release 89: ENSG00000157764 - ENSEMBL , май 2017 г.
- ^ Jump up to: а беременный в GRCM38: Ensembl Release 89: Ensmusg00000002413 - Ensembl , май 2017 г.
- ^ «Человеческая PubMed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «Мышь Pubmed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ Ситханандам Г., Колч В., Дух Ф.М., Рапп Ур (декабрь 1990). «Полная кодирующая последовательность кДНК B-RAF человека и обнаружение протеинкиназы B-RAF с изозимными специфическими антителами». Онкоген . 5 (12): 1775–1780. PMID 2284096 .
- ^ Ситханандам Г., Druck T, Cannizzaro LA, Leuzzi G, Huebner K, Rapp UR (апрель 1992 г.). «B-RAF и псевдоген B-RAF расположены на 7Q в человеке». Онкоген . 7 (4): 795–799. PMID 1565476 .
- ^ Дэвис Х., Бигнелл Г.Р., Кокс С., Стивенс П., Эдканс С., Клегг С. и др. (Июнь 2002 г.). «Мутации гена BRAF при раке человека» (PDF) . Природа . 417 (6892): 949–954. Bibcode : 2002natur.417..949d . doi : 10.1038/nature00766 . PMID 12068308 . S2CID 3071547 .
- ^ «FDA одобряет Zelboraf (Vemurafenib) и компаньон-диагностику для MRAF-позитивной метастатической меланомы, смертельной формы рака кожи» (пресс-релиз). Genentech . Получено 2011-08-17 .
- ^ Erlanson DA, Fesik SW, Hubbard RE, Jahnke W, Jhoti H (сентябрь 2016 г.). «Двадцать лет спустя: влияние фрагментов на обнаружение наркотиков». Природные обзоры. Открытие наркотиков . 15 (9): 605–619. doi : 10.1038/nrd.2016.109 . PMID 27417849 . S2CID 19634793 .
- ^ Jump up to: а беременный "Entrez Gene: BRAF" .
- ^ Daum G, Eisenmann-Tappe I, Fries HW, Troppmair J, Rapp Ur (ноябрь 1994 г.). «Входы и выходы RAF -киназы». Тенденции в биохимических науках . 19 (11): 474–480. doi : 10.1016/0968-0004 (94) 90133-3 . PMID 7855890 .
- ^ Jump up to: а беременный в Катлер Р.Е., Стивенс Р.М., Сарачино М.Р., Моррисон Д.К. (август 1998 г.). «Авторегуляция RAF-1 сериновой/треонинкиназы» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (16): 9214–9219. Bibcode : 1998pnas ... 95.9214c . doi : 10.1073/pnas.95.16.9214 . PMC 21318 . PMID 9689060 .
- ^ Jump up to: а беременный в Bollag G, Tsai J, Zhang J, Zhang C, Ibrahim P, Nolop K, et al. (Ноябрь 2012). «Vemurafenib: первый препарат, одобренный для рака Braf-мутант». Природные обзоры. Открытие наркотиков . 11 (11): 873–886. doi : 10.1038/nrd3847 . PMID 23060265 . S2CID 9337155 .
- ^ Jump up to: а беременный в дюймовый «Серин/треонин протеинкиназа B-RAF» . Получено 4 марта 2013 года .
- ^ Jump up to: а беременный Моррисон Д.К., Катлер Р.Е. (апрель 1997 г.). «Сложность регулирования RAF-1». Современное мнение в клеточной биологии . 9 (2): 174–179. doi : 10.1016/s0955-0674 (97) 80060-9 . PMID 9069260 .
- ^ Хан П.С., Раджеш П., Раджендра П., Часкар М.Г., Рохидас А., Джайпракаш С (март 2022 г.). «Недавние достижения в ингибиторах B-RAF как противоопухолевые агенты». Биоорганическая химия . 120 . Elsevier BV: 105597. DOI : 10.1016/j.bioorg.2022.105597 . PMID 35033817 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л Хэнкс С.К., Охотник Т (май 1995 г.). «Протеинкиназы 6. Суперсемейство эукариотической протеинкиназы: киназа (каталитическая) структура и классификация» . FASEB Journal . 9 (8): 576–596. doi : 10.1096/fasebj.9.8.7768349 . PMID 7768349 . S2CID 21377422 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Wan PT, Garnett MJ, Roe SM, Lee S, Niculescu-Duvaz D, Good VM, et al. (Март 2004 г.). «Механизм активации сигнального пути RAF-ERK путем онкогенных мутаций B-RAF» . Клетка . 116 (6). Проект генома рака: 855–867. doi : 10.1016/s0092-8674 (04) 00215-6 . PMID 15035987 . S2CID 126161 .
- ^ Jump up to: а беременный в дюймовый и фон Tsai J, Lee JT, Wang W, Zhang J, Cho H, Mamo S, et al. (Февраль 2008 г.). «Открытие селективного ингибитора онкогенной B-Raf-киназы с активной активностью антимеланомы» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (8): 3041–3046. Bibcode : 2008pnas..105.3041t . doi : 10.1073/pnas.0711741105 . PMC 2268581 . PMID 18287029 .
- ^ Jump up to: а беременный в дюймовый и Bollag G, Hirth P, Tsai J, Zhang J, Ibrahim PN, Cho H, et al. (Сентябрь 2010). «Клиническая эффективность ингибитора RAF нуждается в широкой целевой блокаде при мутантной меланоме BRAF» . Природа . 467 (7315): 596–599. Bibcode : 2010natur.467..596b . doi : 10.1038/nature09454 . PMC 2948082 . PMID 20823850 .
- ^ Хэнкс С.К., Куинн А.М., Хантер Т (июль 1988 г.). «Семейство протеинкиназы: консервативные особенности и выведенная филогения каталитических доменов». Наука . 241 (4861): 42–52. Bibcode : 1988sci ... 241 ... 42H . doi : 10.1126/science.3291115 . PMID 3291115 .
- ^ Хэнкс С.К. (июнь 1991 г.). «Эукариотические протеинкиназы». Карт Мнение Структура Биол . 1 (3): 369–383. doi : 10.1016/0959-440x (91) 90035-R .
- ^ Хэнкс С.К., Куинн А.М. (1991). «[2] База данных последовательностей каталитического домена протеинкиназы: идентификация консервативных особенностей первичной структуры и классификации членов семейства». База данных последовательностей каталитического домена протеинкиназы: идентификация консервативных особенностей первичной структуры и классификации членов семьи . Методы в фермере. Тол. 200. С. 38–62. doi : 10.1016/0076-6879 (91) 00126-H . ISBN 978-0-12-182101-2 Полем PMID 1956325 .
- ^ Мейсон CS, Springer CJ, Cooper RG, Superti-Furga G, Marshall CJ, Marais R (апрель 1999 г.). «Фосфорилирование серина и тирозина сотрудничают в RAF-1, но не активации B-RAF» . Embo Journal . 18 (8): 2137–2148. doi : 10.1093/emboj/18.8.2137 . PMC 1171298 . PMID 10205168 .
- ^ Робертс А., Аллансон Дж., Джадико С.К., Кавамура М.И., Нунан Дж., Опиц Дж. М. и др. (Ноябрь 2006 г.). «Кардиофакофукожный синдром» . Журнал медицинской генетики . 43 (11): 833–842. doi : 10.1136/jmg.2006.042796 . PMC 2563180 . PMID 16825433 .
- ^ Ewalt M, Nandula S, Phillips A, Alobeid B, Murty VV, Mansukhani MM, et al. (Декабрь 2012 г.). «Анализ на основе ПЦР в реальном времени мутации BRAF V600E в лимфомах низкого и промежуточного уровня подтверждает частое возникновение при волосатой клеточной лейкозе». Гематологическая онкология . 30 (4): 190–193. doi : 10.1002/hon.1023 . PMID 22246856 . S2CID 204843221 .
- ^ Palomaki GE, McClain MR, Melillo S, Hampel HL, Thibodeau SN (январь 2009 г.). «Egapp Дополнительные данные обзора: стратегии тестирования ДНК, направленные на снижение заболеваемости и смертности от синдрома Линча» . Генетика в медицине . 11 (1): 42–65. doi : 10.1097/gim.0b013e31818fa2db . PMC 2743613 . PMID 19125127 .
- ^ Намба Х, Накашима М., Хаяси Т., Хаясида Н., Маэда С., Рогунович Т.И. и др. (Сентябрь 2003 г.). «Клиническое значение мутации BRAF Hot Spot, V599E, при папиллярном раке щитовидной железы» . Журнал клинической эндокринологии и метаболизма . 88 (9): 4393–4397. doi : 10.1210/jc.2003-030305 . PMID 12970315 .
- ^ Tan YH, Liu Y, Eu KW, Ang PW, Li WQ, Salto-Tellez M, et al. (Апрель 2008 г.). «Обнаружение мутации BRAF V600E путем пиросеквенирования». Патология . 40 (3): 295–298. doi : 10.1080/00313020801911512 . PMID 18428050 . S2CID 32051681 .
- ^ Li WQ, Kawakami K, Ruszkiewicz A, Bennett G, Moore J, Iacopetta B (январь 2006 г.). «Мутации BRAF связаны с отличительными клиническими, патологическими и молекулярными особенностями колоректального рака независимо от состояния нестабильности микросателлита» . Молекулярный рак . 5 (1): 2. DOI : 10.1186/1476-4598-5-2 . PMC 1360090 . PMID 16403224 .
- ^ Benlloch S, Payá A, Alenda C, Bessa X, Andreu M, Jover R, et al. (Ноябрь 2006 г.). «Обнаружение мутации BRAF V600E при колоректальном раке: сравнение автоматического секвенирования и методологии химии в реальном времени» . Журнал молекулярной диагностики . 8 (5): 540–543. doi : 10.2353/jmoldx.2006.060070 . PMC 1876165 . PMID 17065421 .
- ^ Денг Г., Белл I, Кроули С., Гум Дж., Тердиман Дж.П., Аллен Б.А. и др. (Январь 2004 г.). «Мутация BRAF часто присутствует при спорадическом колоректальном раке с метилированным HMLH1, но не при наследственном неполипозе колоректального рака» . Клиническое исследование рака . 10 (1 пт 1): 191–195. doi : 10.1158/1078-0432.ccr-1118-3 . PMID 14734469 .
- ^ Gear H, Williams H, Kemp EG, Roberts F (август 2004 г.). «Мутации BRAF в меланоме конъюнктивации» . Расследование офтальмология и визуальная наука . 45 (8): 2484–2488. doi : 10.1167/iovs.04-0093 . PMID 15277467 .
- ^ Maldonado JL, Fridlyand J, Patel H, Jain AN, Busam K, Kageshita T, et al. (Декабрь 2003 г.). «Дерманты мутаций BRF в первичных меланомах » Журнал Национального института рака 95 (24): 1878–1890. Doi : 10.1093/ jnci/ djg1 14679157PMID
- ^ Puxeddu E, Moretti S, Elisei R, Romei C, Pascucci R, Martinelli M, et al. (Май 2004 г.). «Мутация BRAF (V599E) является ведущим генетическим событием во взрослых спорадических папиллярных карциномах щитовидной железы» . Журнал клинической эндокринологии и метаболизма . 89 (5): 2414–2420. doi : 10.1210/jc.2003-031425 . PMID 15126572 .
- ^ Elisei R, Ugolini C, Viola D, Lupi C, Biagini A, Giannini R, et al. (Октябрь 2008 г.). «Мутация BRAF (V600E) и исход пациентов с папиллярной карциномой щитовидной железы: 15-летнее среднее последующее исследование» . Журнал клинической эндокринологии и метаболизма . 93 (10): 3943–3949. doi : 10.1210/jc.2008-0607 . PMID 18682506 .
- ^ Badalian-Every G, Verglio JA, Degar BA, Rodriguguez-Galindo C, Rollins BJ (январь 2012 г.). «Последние достижения в понимании гистоцитоза клеток Лангерганса » Британский журнал гематологии 156 (2): 163–1 Doi : 10.1111/ j.1365-2141.2011.08915.x 22017623PMID 34922416S2CID
- ^ Tiacci E, Trifonov V, Schiavoni G, Holmes A, Kern W, Martelli MP, et al. (Июнь 2011 г.). «Мутации BRAF при волосатой лейкемии» . Новая Англия Журнал медицины . 364 (24): 2305–2315. doi : 10.1056/nejmoa1014209 . PMC 3689585 . PMID 21663470 . *Lay Support в: «Исследования по лейкемии волосатую ячейки показывают обещание новых технологий сканирования ДНК» . Рак исследований Великобритания . 11 июня 2011 г.
- ^ Курппа К.Дж., Катон Дж., Морган П.Р., Ристимаки А., Рухин Б., Келлокоски Дж. И др. (Апрель 2014). «Высокая частота мутаций BRAF V600E при амелобластоме» . Журнал патологии . 232 (5): 492-498. Doi : 10.1002/path.4317 . PMC 4255689 . PMID 24374844 .
- ^ Brastianos PK, Taylor-Weiner A, Manley PE, Jones RT, Dias-Santagata D, Thorner AR, et al. (Февраль 2014 г.). «Секвенирование экзома идентифицирует мутации BRAF в папиллярных краниофрингиомах» . Природа генетика . 46 (2): 161–165. doi : 10.1038/ng.2868 . PMC 3982316 . PMID 24413733 . *Lay Support в: Лия Эйзенштадт (30 января 2014 г.). «Мутация одиночной драйверы, обнаруженная в редкой опухоли головного мозга» . Широкий институт .
- ^ Zecchin D, Boscaro V, Medico E, Barault L, Martini M, Arena S, et al. (Декабрь 2013). «BRAF V600E является определяющим фактором чувствительности к ингибиторам протеасом» . Молекулярная терапия рака . 12 (12): 2950–2961. doi : 10.1158/1535-7163.mct-13-0243 . HDL : 2318/140935 . PMID 24107445 . S2CID 17012966 .
- ^ Король А.Дж., Патрик Д.Р., Батерский Р.С., Хо М.Л., До Х.Т., Чжан Си, и др. (Декабрь 2006 г.). «Демонстрация генетического терапевтического индекса для опухолей, экспрессирующих онкогенный BRAF ингибитором киназы SB-590885» . РАНКА . 66 (23): 11100–11105. doi : 10.1158/0008-5472.can-06-2554 . PMID 17145850 .
- ^ Chapman PB, Hauschild A, Robert C, Haanen JB, Ascierto P, Larkin J, et al. (Июнь 2011 г.). «Улучшена выживаемость с Vemurafenib при меланоме с мутацией BRAF V600E» . Новая Англия Журнал медицины . 364 (26). Исследовательская группа BRIM-3: 2507–2516. doi : 10.1056/nejmoa1103782 . PMC 3549296 . PMID 21639808 .
- ^ Jump up to: а беременный Das Thakur M, Salangsang F, Landman AS, Sellers WR, Pryer NK, Levesque MP, et al. (Февраль 2013 г.). «Моделирование устойчивости Вемурафениба при меланоме выявляет стратегию предотвращения лекарственной устойчивости» . Природа . 494 (7436): 251–255. Bibcode : 2013natur.494..251d . doi : 10.1038/nature11814 . PMC 3930354 . PMID 23302800 .
- ^ Назарян Р., Ши Х, Ван К, Конг Х, Коя Р.К., Ли Х. и др. (Декабрь 2010). «Меланомы приобретают устойчивость к ингибированию B-RAF (V600E) с помощью активации RTK или N-RAS» . Природа . 468 (7326): 973–977. Bibcode : 2010natur.468..973n . doi : 10.1038/nature09626 . PMC 3143360 . PMID 21107323 .
- ^ Degirmenci U, Yap J, Sim YR, Qin S, Hu J (2021). «Устойчивость к лекарственной устойчивости при целевой терапии рака с ингибиторами RAF» . Раковая лекарственная устойчивость . 4 (3): 665–683. doi : 10.20517/cdr.2021.36 . PMC 9094075 . PMID 35582307 .
- ^ Guan KL, Figueroa C, Brtva TR, Zhu T, Taylor J, Barber TD, et al. (Сентябрь 2000). «Негативная регуляция сериновой/треонинкиназы B-RAF от AKT» . Журнал биологической химии . 275 (35): 27354–27359. doi : 10.1074/jbc.m004371200 . PMID 10869359 .
- ^ Вебер К.К., Слюпский младший, Калмес Х.А., Рапп Ур (май 2001 г.). «Активный RAS вызывает гетеродимеризацию CRAF и BRAF». РАНКА . 61 (9): 3595–3598. PMID 11325826 .
- ^ Stang S, Bottorff D, Stone JC (июнь 1997 г.). «Взаимодействие активированных RA с RAF-1 может быть достаточным для трансформации клеток Rat2» . Молекулярная и клеточная биология . 17 (6): 3047–3055. doi : 10.1128/mcb.17.6.3047 . PMC 232157 . PMID 9154803 .
- ^ Reuter CW, Catling AD, Jelinek T, Weber MJ (март 1995 г.). «Биохимический анализ активации MEK в фибробластах NIH3T3. Идентификация B-RAF и других активаторов» . Журнал биологической химии . 270 (13): 7644–7655. doi : 10.1074/jbc.270.13.7644 . PMID 7706312 .
- ^ Юинг Р.М., Чу П., Элизма Ф., Ли Х, Тейлор П., Клили С. и др. (2007). «Крупномасштабное картирование взаимодействия белка белка человека с помощью масс-спектрометрии» . Биология молекулярных систем . 3 (1): 89. doi : 10.1038/msb4100134 . PMC 1847948 . PMID 17353931 .
- ^ Qiu W, Zhuang S, Von Lintig FC, Boss GR, Pilz RB (октябрь 2000 г.). «Специфичная клетка регуляция B-Raf-киназы с помощью CAMP и 14-3-3 белков» . Журнал биологической химии . 275 (41): 31921–31929. doi : 10.1074/jbc.m003327200 . PMID 10931830 .
Дальнейшее чтение
[ редактировать ]- Гарнетт М.Дж., Марайс Р. (октябрь 2004 г.). «Виновен как обвинение: B-Raf-это человеческий онкоген» . Раковая клетка . 6 (4): 313–319. doi : 10.1016/j.ccr.2004.09.022 . PMID 15488754 .
- Quiros RM, Ding Hg, Gattuso P, Prinz RA, Xu X (июнь 2005 г.). «Доказательства того, что одна подмножество анапластических карцином щитовидной железы получена из папиллярных карциномов из -за мутаций BRAF и p53» . Рак . 103 (11): 2261–2268. doi : 10.1002/cncr.21073 . PMID 15880523 . S2CID 29665029 .
- Karbowniczek M, Henske EP (ноябрь 2005 г.). «Роль туберина в клеточной дифференцировке: участвуют ли B-RAF и MAPK?». Анналы нью -йоркской академии наук . 1059 (1): 168–173. Bibcode : 2005nyasa1059..168k . doi : 10.1196/annals.1339.045 . PMID 16382052 . S2CID 39146204 .
- Ciampi R, Nikiforov ye (март 2007 г.). «Перестройки RET/PTC и мутации BRAF в онкогенезе щитовидной железы» . Эндокринология . 148 (3): 936–941. doi : 10.1210/en.2006-0921 . PMID 16946010 .
- Порчия Л., Рингель, MD (январь "BRAF при раке щитовидной железы " Британский журнал рака 96 (1): 16–2 doi : 10.1038/sj.bjc . PMC 2360215 . 17179987PMID
- Аллансон Дж., Робертс А.Е. (8 августа 2019 г.). «Синдром Нунана» . В Pagon RA, Bird TD, Dolan CR, et al. (ред.). GenereViews [Интернет] . Сиэтл Вашингтон: Университет Вашингтона, Сиэтл. PMID 20301303 .
- Рауэн Ка (3 марта 2016 г.) [18 января 2007 г.]. «Кардиоофакокожный синдром» . В Pagon RA, Bird TD, Dolan CR (ред.). GenereViews [Интернет] . Сиэтл Вашингтон: Университет Вашингтона, Сиэтл. PMID 20301365 .
- Gelb BD, Tartaglia M (14 мая 2015 г.) [30 ноября 2007 г.]. «Синдром леопарда» . В Pagon RA, Bird TD, Dolan CR (ред.). GenereViews [Интернет] . Сиэтл Вашингтон: Университет Вашингтона, Сиэтл. PMID 20301557 .
Внешние ссылки
[ редактировать ]- «Ген BRAF» . NCI Словарь раковых терминов . Получено 2007-11-25 .
- Поиск недостатков в BRAF Archived 2018-08-19 в The Wayback Machine -рак исследования в Великобритании в блоге об обнаружении мутаций BRAF, вызывающих рак (incl video)
- человека Местоположение генома BRAF и страница детали гена BRAF в браузере генома UCSC .
![]() Эта статья включает в себя материал общественного достояния из Словарь раковых терминов . Национальный институт рака США .
Эта статья включает в себя текст из Национальной медицины Соединенных Штатов , которая находится в общественном достоянии .
Эта статья включает в себя материал общественного достояния из Словарь раковых терминов . Национальный институт рака США .
Эта статья включает в себя текст из Национальной медицины Соединенных Штатов , которая находится в общественном достоянии .
PDB Галерея |
|---|