Икра – это яйца и сперматозоиды , выделяемые или попадающие в воду водными животными . Глагол « нереститься » относится к процессу свободного выброса яиц и спермы в водоем (пресную или морскую); физический акт известен как нерест . Подавляющее большинство водных и амфибийных животных размножаются нерестом. К ним относятся следующие группы:
Кораллы — это живые колонии крошечных водных организмов, а не растений, как их иногда считают. Кораллы, хотя по своей природе кажутся малоподвижными или ботаническими, на самом деле нерестятся, выпуская облака сперматозоидов и яйцеклеток в толщу воды, где они смешиваются.
Spawn consists of the reproductive cells (gametes) of many aquatic animals, some of which will become fertilized and produce offspring. The process of spawning typically involves females releasing ova (unfertilized eggs) into the water, often in large quantities, while males simultaneously or sequentially release spermatozoa (milt) to fertilize the eggs.[1][2][3]
The fungi (mushrooms), are also said to "spawn" when they release a white, ‘fibrous’ matter, forming the matrix from-which they grow.[citation needed]
There are many variations in the way spawning occurs, depending on sexual differences in anatomy, how the sexes relate to each other, where and how the spawn is released and whether or how the spawn is subsequently guarded.
Pacific salmon are semelparous or "big bang" spawners, which means they die shortly after spawningThe pickled and dehydrated roe of mullet
Marine animals, and particularly bony fish, commonly reproduce by broadcast spawning. This is an external method of reproduction where the female releases many unfertilised eggs into the water. At the same time, a male or many males release a lot of sperm into the water which fertilises some of these eggs. The eggs contain a drop of nutrient oil to sustain the embryo as it develops inside the egg case. The oil also provides buoyancy, so the eggs float and drift with the current. The strategy for survival of broadcast spawning is to disperse the fertilised eggs, preferably away from the coast into the relative safety of the open ocean. There the larvae develop as they consume their fat stores, and eventually hatch from the egg capsule into miniature versions of their parents.
To survive, they must then become miniature predators themselves, feeding on plankton. Fish eventually encounter others of their own kind (conspecifics), where they form aggregations and learn to school.
Internally, the sexes of most marine animals can be determined by looking at the gonads. For example, male testes of spawning fish are smooth and white and account for up to 12% of the mass of the fish, while female ovaries are granular and orange or yellow, accounting for up to 70% of the fish's mass. Male lampreys, hagfish and salmon discharge their sperm into the body cavity where it is expelled through pores in the abdomen. Male sharks and rays can pass sperm along a duct into a seminal vesicle, where they store it for a while before it is expelled, while teleosts usually employ separate sperm ducts.[4]: 141
Externally, many marine animals, even when spawning, show little sexual dimorphism (difference in body shape or size) or little difference in colouration. Where species are dimorphic, such as sharks or guppies, the males often have penis-like intromittent organs in the form of a modified fin.[4]: 141
A species is semelparous if its individuals spawn only once in their lifetime, and iteroparous if its individuals spawn more than once. The term semelparity comes from the Latinsemel, once, and pario, to beget, while iteroparity comes from itero, to repeat, and pario, to beget.
Semelparity is sometimes called "big bang" reproduction, since the single reproductive event of semelparous organisms is usually large and fatal to the spawners.[5] The classic example of a semelparous animal is the Pacific salmon, which lives for many years in the ocean before swimming to the freshwater stream of its birth, spawning, and then dying. Other spawning animals which are semelparous include mayflies, squid, octopus, smelt, capelin and some amphibians.[6] Semelparity is often associated with r-strategists. However, most fish and other spawning animals are iteroparous.
When the internal ovaries or egg masses of fish and certain marine animals are ripe for spawning they are called roe. Roe from certain species, such as shrimp, scallop, crab and sea urchins, are sought as human delicacies in many parts of the world. Caviar is a name for the processed, salted roe of non-fertilized sturgeon. The term soft roe or white roe denotes fish milt. Lobster roe is called coral because it turns bright red when cooked. Roe (reproductive organs) are usually eaten either raw or briefly cooked.
"The reproductive behaviour of fishes is remarkably diversified: they may be oviparous (lay eggs), ovoviviparous (retain the eggs in the body until they hatch), or viviparous (have a direct tissue connection with the developing embryos and give birth to live young). All cartilaginous fishes—the elasmobranches (e.g., sharks, rays, and skates)—employ internal fertilization and usually lay large, heavy-shelled eggs or give birth to live young. The most characteristic features of the more primitive bony fishes is the assemblage of polyandrous (many males) breeding aggregations in open water and the absence of parental care..."[7]
There are two main reproduction methods in fish. The first method is by laying eggs and the second by live-bearing (producing their young alive).
In the first method, the female fish lays eggs either on the sea floor or on the leaves of an aquatic plant. A male fish fertilizes the eggs, and both then work together to protect the eggs/babies from danger until they can defend themselves.
In the second method, the male fish uses its anal fin to transmit sperm into the female fish and fertilize the fish eggs. Later, the female gives live birth to her fry.
Cutthroat trout are monogamous pair spawnersThe anglerfish Haplophryne mollis is polyandrous. This female is trailing the atrophied remains of males she has encountered
Monogamy occurs when one male mates with one female exclusively. This is also called pair spawning.[9] Most fish are not monogamous, and when they are, they often alternate with non-monogamous behaviours. Monogamy can occur when feeding and breeding grounds are small, when it is difficult for fish to find partners, or when both sexes look after the young.[8] Many tropical cichlids, which rear their young together in locations where they must fiercely defend against competitors and predators are monogamous.[10] "In some pipefishes and seahorses, development of eggs takes a long time before the female can place them in the brood pouch of a male, where they are fertilized. While the male is pregnant, the female starts a new batch of eggs, which are ready at about the same time that the male gives birth to the young from the previous mating. This close timing of development promotes monogamy, especially if the likelihood of encountering another potential mate is low."[8]
Polygyny occurs when one male gets exclusive mating rights with multiple females. In polygyny, a large conspicuous male usually defends females from other males or defends a breeding site.[8] The females choose large males that are successfully defending prime breeding sites which the females find attractive. For example, sculpin males defend "caves" underneath rocks which are suitable for the incubation of embryos.
Another way males get to mate with several females is through the use of leks. Leks are places where many fish come together, and the males display to each other. Based on these displays, each female then selects the male they want to be their mate. For example, among the cichlid Cyrtocara eucinostomus in Lake Malawi, up to 50,000 large and colourful males display together on a lek four kilometres long. The females, which are mouth brooders, choose which male they want to fertilize their eggs.[11]
Polyandry occurs when one female gets exclusive mating rights with multiple males. This happens among fish like clownfish that change their sex. It can also happen when males do the brooding but cannot handle all the eggs the female produce, such as with some pipefish.[4]: 161
The males in some deep sea anglerfishes are much smaller than the females. When they find a female they bite into her skin, releasing an enzyme that digests the skin of their mouth and her body and fusing the pair down to the blood-vessel level. The male then slowly atrophies, losing first his digestive organs, then his brain, heart, and eyes, ending as nothing more than a pair of gonads, which release sperm in response to hormones in the female's bloodstream indicating egg release. This ensures that, when the female is ready to spawn, she has a mate immediately available.[12] A single anglerfish female can "mate" with many males in this manner.
Polygynandry occurs when multiple males mate indiscriminately with multiple females. This mutual promiscuity is the approach most commonly used by spawning animals, and is perhaps the "original fish mating system."[4]: 161 Common examples are forage fish, such as herrings, which form huge mating shoals in shallow water. The water becomes milky with sperm and the bottom is draped with millions of fertilized eggs.[4]: 161
Small male bluegill sunfishes cuckold large males by adopting sneaker or satellite strategiesFemale groupers change their sex to male if no male is availableAn anemone fish couple guarding their anemone. If the female dies, a juvenile male moves in, and the resident male changes sex.
Alternate male strategies which allow small males to engage in cuckoldry can develop in species where spawning is dominated by large and aggressive males. Cuckoldry is a variant of polyandry, and can occur with sneak spawners (sometimes called streak spawners). A sneak spawner is a male that rushes in to join the spawning rush of a spawning pair.[13] A spawning rush occurs when a fish makes a burst of speed, usually on a near vertical incline, releasing gametes at the apex, followed by a rapid return to the lake or sea floor or fish aggregation.[14] Sneaking males do not take part in courtship. In salmon and trout, for example, jack males are common. These are small silvery males that migrate upstream along with the standard, large, hook-nosed males and that spawn by sneaking into a redd (spawning nest) to release sperm simultaneously with a mated pair. This behaviour is an evolutionarily stable strategy for reproduction, because it is favoured by natural selection just like the "standard" strategy of large males.[15]
Cuckoldry occurs in many fish species, including dragonets, parrotfishes and wrasses on tropical reefs and the bluegill sunfish in fresh water. Sneaker males that become too large to hide effectively become satellite males. With bluegill sunfish, satellite males mimic the behaviour and colouration of the females. They hover over a nest containing a pair of courting sunfish, and gradually descend to reach the pair just as they spawn. Males may need to be 6 or 7 years old to function capably as parental males, but may be able to function as sneaker or satellite males when they are as young as 2 or 3 years old. The smaller satellite and sneaker males may get mauled by the more powerful parental males, but they spawn when they are younger and they do not put energy into parental care.[4]: 161–2 [16]
Hermaphroditism occurs when a given individual in a species possesses both male and female reproductive organs, or can alternate between possessing first one, and then the other. Hermaphroditism is common in invertebrates but rare in vertebrates. It can be contrasted with gonochorism, where each individual in a species is either male or female, and remains that way throughout their lives. Most fish are gonochorists, but hermaphroditism is known to occur in 14 families of teleost fishes.[17]
Usually hermaphrodites are sequential, meaning they can switch sex, usually from female to male (protogyny). This can happen if a dominant male is removed from a group of females. The largest female in the harem can switch sex over a few days and replace the dominant male.[17] This is found amongst coral reef fishes such as groupers, parrotfishes and wrasses. It is less common for a male to switch to a female (protandry).[4]: 162 As an example, most wrasses are protogynous hermaphrodites within a haremic mating system.[18][19] Hermaphroditism allows for complex mating systems. Wrasses exhibit three different mating systems: polygynous, lek-like, and promiscuous mating systems.[20] Group spawning and pair spawning occur within mating systems. The type of spawning that occurs depends on male body size.[19] Labroids typically exhibit broadcast spawning, releasing high amounts of planktonic eggs, which are broadcast by tidal currents; adult wrasses have no interaction with offspring.[21] Wrasse of a particular subgroup of the family Labridae, Labrini, do not exhibit broadcast spawning.
Less commonly hermaphrodites can be synchronous, meaning they simultaneously possess both ovaries and testicles and can function as either sex at any one time. Black hamlets "take turns releasing sperm and eggs during spawning. Because such egg trading is advantageous to both individuals, hamlets are typically monogamous for short periods of time–an unusual situation in fishes."[22] The sex of many fishes is not fixed, but can change with physical and social changes to the environment where the fish lives.[23]
Particularly among fishes, hermaphroditism can pay off in situations where one sex is more likely to survive and reproduce, perhaps because it is larger.[24]Anemone fishes are sequential hermaphrodites which are born as males, and become females only when they are mature. Anemone fishes live together monogamously in an anemone, protected by the anemone stings. The males do not have to compete with other males, and female anemone fish are typically larger. When a female dies a juvenile (male) anemone fish moves in, and "the resident male then turns into a female and reproductive advantages of the large female–small male combination continue".[25] In other fishes sex changes are reversible. For example, if some gobies are grouped by sex (male or female), some will switch sex.[4]: 164 [24]
Unisexuality occurs when a species is all-male or all-female. Unisexuality occurs in some fish species, and can take complex forms. Squalius alburnoides, a minnow found in several river basins in Portugal and Spain, appears to be an all-male species. The existence of this species illustrates the potential complexity of mating systems in fish. The species originated as a hybrid between two species, and is diploid, but not hermaphroditic. It can have triploid and tetraploid forms, including all-female forms that reproduce mainly through hybridogenesis.[26]
It is rare to find true parthenogenesis in fishes, where females produce female offspring with no input from males. All-female species include the Texas silverside, Menidia clarkhubbsi[27] as well as a complex of Mexican mollies.[4]: 162 Parthenogenesis has been recently observed in hammerhead sharks[28] and blacktip sharks.[29] It is also known to occur in crayfish[30][31] and amphibians.[32][33]
This section is patterned after a classification of the spawning behaviours of fish by Balon (1975, 1984) into reproductive guilds. This classification is based on how the eggs are fertilized (internal or external spawners), where the eggs are deposited (pelagic or benthic spawners), and whether and how the parents look after the eggs after spawning (bearers, guarders and nonguarders).[34]
Bitterlings transfer responsibility for the care of their young to mussels. This male bitterling is exhibiting spawning colours
Open substrate spawners scatter their eggs in the environment. They usually spawn in shoals without complex courtship rituals, and males outnumber females.
Broadcast spawners: release their gametes (sperm and eggs) into open water for external fertilisation. There is no subsequent parental care.[35] About 75% of coral species are broadcasters, the majority of which are hermatypic, or reef-building corals.[36]
Pelagic spawners: a type of broadcast spawners, spawn in the open sea, mostly near the surface. They are usually pelagic fish such as tuna and sardines. Some demersal fish leave the bottom to spawn pelagically, particularly coral reef fish such as parrotfish and wrasses. Pelagic spawning means water currents widely disperse the young. The eggs, embryos and larvae of pelagic spawners contain oil globules or have a high water content. As a result, they are buoyant and are widely dispersed by currents. The downside is that mortality is high, because they can be eaten so easily by pelagic predators or they can drift into unsuitable areas. Females compensate by spawning large numbers of eggs and extending their spawning periods. Pelagic spawners that live in or around coral reefs can spawn a small number of eggs almost daily over a period of months. These fishes have complex breeding behaviours including sex changes, harems, leks and territoriality.[4]: 143
Benthic spawners: deposit their spawn on or near the bottom of the sea (or lake). They are usually demersal fish such as cod and flatfish. These species typically spawn without ceremony; they do not engage in elaborate courtship rituals. Each female is usually followed by several males who fertilize the eggs as they are released. Various strategies ensure the eggs and embryos remain in place, and do not drift with the current. The eggs can adhere to other eggs or to whatever they are deposited on, or the eggs can be laid in long strings which are wrapped around plants or rocks. Some eggs take on water after they are released, so they can be dropped into cracks where they swell and wedge themselves in place.
Egg scatterers: scatter adhesive or non-adhesive eggs to fall to the substrate, into plants, or float to the surface. These species do not look after their brood and even eat their own eggs. These are often schooling fish which spawn in groups or pairs, often laying a large number of small eggs. The fry hatch quickly.
Egg depositors: deposit eggs on a substrate (tank glass, wood, rocks, plants). Egg depositors usually lay fewer eggs than egg-scatterers, although the eggs are larger. Egg depositors fall into two groups: those that care for their eggs, and those that do not. Among egg depositors that care for their eggs are cichlids and some catfish. Egg depositors that care for their young can be divided into two groups: cavity spawners and open spawners.
Cavity spawners: lay eggs in a cave or cavity. These fish form pairs and have advanced brood care where the eggs are defended and cleaned. The eggs take a few days to hatch, and the fry are often guarded by the parents. Various catfish, Cyprinidae, and killifish make up the majority. Cavity spawners can be contrasted with open (shelter) spawners, which lay their eggs on an open surface.
Brood hiders hide their eggs but do not give parental care after they have hidden them. Brood hiders are mostly benthic spawners that bury the fertilized eggs. For example, among salmon and trout the female digs a nest with her tail in gravel. These nests are called redds. The female then lays her eggs while the male fertilizes them, while both fish defend the redd if necessary from other members of the same species. Then the female buries the nest, and the nest site is abandoned. In North America, some minnows build nests out of piles of stones rather than dig holes. The minnow males have tubercles on their head and body which they use to help them defend the nest site.[4]: 145
Egg buriers - can inhabit waters that dry up at some time of the year. An example are annual killifish which lay their eggs in mud. The parents mature quickly and lay their eggs before dying when the water dries up. The eggs remain in a dormant stage until rains stimulate hatching.
Bitterlings have a remarkable reproduction strategy where parents transfer responsibility for the care of their young to mussels. The female extends her ovipositor into the mantle cavity of the mussel and deposits her eggs between the gill filaments. The male then ejects his sperm into the mussel's inhalant water current and fertilization takes place within the gills of the host. The same female may use a number of mussels, and she deposits only one or two yellow, oval eggs into each. Early developmental stages are protected from predation within the body of the mussel. After 3 to 4 weeks larvae swim away from the host to continue life on their own.
The stickleback glues plant material to make its nest
Guarders protect their eggs and offspring after spawning by practicing parental care (also called brood care). Parental care is an "investment by parents in offspring that increases the offspring's chances of surviving (and hence reproducing). In fish, parental care can take a variety of forms including guarding, nest building, fanning, splashing, removal of dead eggs, retrieval of straying fry, external egg carrying, egg burying, moving eggs or young, ectodermal feeding, oral brooding, internal gestation, brood-pouch egg carrying, etc."[37]
Territorial behaviour is generally necessary for guarders, and the embryos are almost always guarded by males (apart from cichlids). There is a need to be territorial because looking after embryos usually includes defending the site where they are being looked after. It also often means there is competition for the best egg-laying sites. Elaborate courtship behaviour is usual among guarders.[4]: 145
Guarding males keep the embryos safe from predators, keep oxygen levels high by fanning water currents, and keep the area free from dead embryos and debris. They protect the embryos until they hatch, and often look after the larval stages as well. The time spent guarding can range from a few days to several months.[4]: 145
Some guarders build nests (nest spawners) and some do not (substrate spawners), though the difference between the two groups can be small.[4]: 142 Substrate spawners clean off a suitable area of surface suitable for egg laying, and look after the area, but they do not actively build a nest.
Mouth brooders - carry eggs or larvae in their mouth. Mouth brooders can be ovophiles or larvophiles. Ovophile or egg-loving mouth-brooders lay their eggs in a pit, which are sucked up into the mouth of the female. The small number of large eggs hatch in the mother's mouth, and the fry remain there for a period of time. Fertilization often occurs with the help of egg-spots, which are colorful spots on the anal fin of the male. When the female sees these spots, she tries to pick up the egg-spots, but instead gets sperm that fertilizes the eggs in her mouth. Many cichlids and some labyrinth fish are ovophile mouthbrooders. Larvophile or larvae-loving mouth-brooders lay their eggs on a substrate and guard them until the eggs hatch. After hatching, the female picks up the fry and keeps them in her mouth. When the fry can fend for themselves, they are released. Some eartheaters are larvophile mouthbrooders.
The beginning of the evolutionary process of livebearing starts with facultative (optional) internal bearing. The process occurs in several species of oviparous (egg-laying) killifishes which spawn in the normal way on the substrate, but in the process accidentally fertilize eggs that the female retains and does not spawn. These eggs are spawned later, usually without allowing much time for embryonic development.[4]: 147
The next step in the evolution of livebearing is obligate (by necessity) internal bearing, where the female retains all the embryos. "The only source of nutrition for these embryos, however, is the egg yolk, as in externally spawned eggs. This situation, also referred to as ovoviviparity, is characteristic of marine rock fishes and the Lake Baikal sculpins. This strategy allows these fish to have fecundities approaching those of pelagic fish with external fertilization, but it also enables them to protect the young during their most vulnerable stage of development. By contrast, sharks and rays using this strategy produce a relatively small number of embryos and retain them for a few weeks to 16 months or longer. The shorter times spans are characteristic of species that eventually deposit their embryos in the environment, surrounded by a horny capsule; whereas the longer periods are characteristic of sharks that retain the embryos until they are ready to emerge as actively swimming young."[4]: 147 [38]
However, some fish do not fit these categories. The livebearing largespring gambusia (Gambusia geiseri) was thought to be ovoviviparous until it was shown in 2001 that the embryos received nutrients from the mother.[39]
Spawning grounds are the areas of water where aquatic animals spawn, or produce their eggs. After spawning, the spawn may or may not drift to new grounds which become their nursery grounds. Many species undertake migrations each year, and sometimes great migrations, to reach their spawning grounds. For example, lakes and river watersheds can be major spawning grounds for anadromous fish such as salmon. These days, it is often necessary to construct fish ladders and other bypass systems so salmon can navigate their way past hydroelectric dams or other obstructions such as weirs on their way to spawning grounds.[40][41]Coastal fish often use mangroves and estuaries as spawning grounds, while reef fish can find adjacent seagrass meadows that make good spawning grounds. Short-finned eels can travel anything up to three or four thousand kilometres to their spawning ground in deep water somewhere in the Coral Sea.
Forage fish often make great migrations between their spawning, feeding and nursery grounds. Schools of a particular stock usually travel in a triangle between these grounds. For example, one stock of herrings have their spawning ground in southern Norway, their feeding ground in Iceland, and their nursery ground in northern Norway. Wide triangular journeys such as these may be important because forage fish, when feeding, cannot distinguish their own offspring.[42]
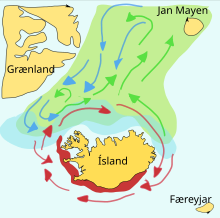
Мойва – кормовая рыба семейства корюшка , обитающая в Атлантическом и Северном Ледовитом океанах. Летом они пасутся в густых стаях планктона на краю шельфового ледника. Более крупная мойва также поедает криль и других ракообразных . Мойва большими стаями перемещается к берегу для нереста и миграции весной и летом, чтобы накормиться в богатых планктоном районах между Исландией , Гренландией и Ян-Майеном . На миграцию влияют океанские течения . В окрестностях Исландии созревающая мойва весной и летом совершает крупные кормовые миграции на север. Обратная миграция происходит в сентябре-ноябре. Нерестовая миграция начинается к северу от Исландии в декабре или январе. [ 43 ]
На схеме справа показаны основные нерестилища и пути перемещения личинок . Мойва на пути к местам нагула окрашена в зеленый цвет, мойва на обратном пути – в синий, а места размножения – в красный. [ 43 ] В статье, опубликованной в 2009 году, исследователи из Исландии рассказывают о своем применении модели взаимодействующих частиц к запасам мойвы вокруг Исландии, успешно предсказывая маршрут нерестовой миграции на 2008 год. [ 44 ]
Названный «величайшим косяком на земле», забег сардин происходит, когда миллионы сардин мигрируют со своих нерестилищ к югу от южной оконечности Африки на север вдоль береговой линии Восточного Кейпа. Чавыча совершает самую длинную пресноводную миграцию среди всех лососей: более 3000 километров (1900 миль) вверх по реке Юкон к нерестилищам выше по течению от Уайтхорса , Юкон. Некоторые зеленые морские черепахи проплывают более 2600 километров (1600 миль), чтобы добраться до мест нереста.
Самки золотых рыбок откладывают (выбрасывают) икру в воду, поощряемые самцами золотой рыбки, которые одновременно выделяют сперму, которая оплодотворяет икру извне. В течение двух-трех дней из уязвимых икринок золотой рыбки вылупляются личинки , из которых быстро развиваются мальки.
Золотые рыбки , как и все карповые , являются яйцекладущими. Обычно они приступают к размножению после значительного изменения температуры, часто весной. Самцы преследуют самок, побуждая их выпустить яйца, ударяя и подталкивая их. Пока самка золотой рыбки выметывает икру, самец остается рядом и оплодотворяет ее. Их яйца клейкие и прикрепляются к водной растительности. Яйца вылупляются в течение 48–72 часов. Примерно через неделю мальки начинают принимать окончательную форму, хотя может пройти год, прежде чем они приобретут зрелый цвет золотой рыбки; до тех пор они имеют металлически-коричневый цвет, как и их дикие предки. В первые недели жизни мальки растут быстро — адаптация, вызванная высоким риском быть съеденными взрослой золотой рыбкой.
Карп обычно нерестится весной и летом, в зависимости от климата и условий. Уровень кислорода в воде, наличие пищи, размер каждой рыбы, возраст, количество раз, когда рыба нерестилась раньше, и температура воды — все это факторы, которые, как известно, влияют на то, когда и сколько икринок каждый карп будет нерестить одновременно. [ 45 ]
Перед нерестом самцы сиамских бойцовских рыб строят пузырьковые гнезда на поверхности воды разного размера. Когда самец начинает интересоваться самкой, он раздувает жабры, крутит тело и расправляет плавники. Самка темнеет и изгибает тело вперед и назад. Акт нереста происходит в «брачных объятиях», когда самец обвивает свое тело вокруг самки, каждое объятие приводит к высвобождению 10–40 яиц, пока у самки не закончатся яйца. Самец со своей стороны выпускает молоки в воду и оплодотворение происходит снаружи. Во время и после нереста самец ртом извлекает тонущую икру и откладывает ее в пузырьковое гнездо (во время спаривания самка иногда помогает партнеру, но чаще она просто пожирает всю икру, которую ей удается поймать). Как только самка выпустила все яйца, ее прогоняют с территории самца, так как вполне вероятно, что она съест яйца из-за голода. [ 46 ] Затем яйца остаются на попечении самца. Он держит их в пузырьковом гнезде, следя за тем, чтобы ни один из них не упал на дно, и при необходимости ремонтирует гнездо. Инкубация длится 24–36 часов, а только что вылупившиеся личинки остаются в гнезде следующие 2–3 дня, пока их желточные мешки полностью не рассасываются. После этого мальки покидают гнездо и наступает стадия свободного плавания. [ 47 ]
Веслоногие ракообразные — это крошечные ракообразные , которые обычно размножаются либо разбросанным нерестом , либо нерестом в мешочках . Вещательные копеподы разбрасывают икру в воду, а производители мешков откладывают икру в яйцевидный мешок. Производители мешков выметывают немного, но относительно крупных икринок, которые развиваются медленно. Напротив, широковещательные производители выметывают множество маленьких икринок, которые быстро развиваются. [ 48 ] Однако более короткое время вылупления, возникающее в результате разброса, недостаточно, чтобы компенсировать более высокую смертность по сравнению с производителями мешков. Чтобы произвести заданное количество вылупившихся яиц, разбрасыватели должны выметать больше яиц, чем производители мешков. [ 49 ]
После спаривания оплодотворенные яйца калифорнийского лангуста самки переносятся на плеоподах до тех пор, пока они не вылупятся, причем одна самка переносит от 120 000 до 680 000 икринок. [ 50 ] Яйца начинаются с кораллово-красного цвета, но по мере развития становятся темно-бордовыми. [ 51 ] Когда она вынашивает яйца, о самке говорят, что она «ягодная». Яйца готовы к вылуплению через 10 недель. [ 50 ] нерест происходит с мая по август. [ 52 ] личинками филлосомы Вылупившиеся личинки (называемые . ) не похожи на взрослых особей Вместо этого это плоские, прозрачные животные длиной около 14 мм (0,55 дюйма), но тонкие, как лист бумаги. [ 53 ] Личинки питаются планктоном , [ 51 ] и вырастают через десять линек до десяти дальнейших личиночных стадий, последняя из которых имеет длину около 30–32 мм (1,2–1,3 дюйма). [ 53 ] Полная серия личиночной линьки занимает около 7 месяцев, и когда линяет последняя стадия, она превращается в пуэрулюсное состояние, которое представляет собой ювенильную форму взрослой особи, хотя и все еще прозрачную. [ 53 ] Личинки puerulus оседают на морское дно, когда температура воды приближается к максимальной, что в Нижней Калифорнии приходится на осень. [ 54 ]
омаров, несущих икру, Самки мигрируют к берегу из более глубоких вод, чтобы высидеть икру, хотя у них нет конкретных нерестилищ. [ 55 ] [ 56 ] Эти миграции омаров могут происходить в виде тесных одиночных группировок «поездов омаров».
Устрицы являются широковещательными производителями , то есть икра и сперма выпускаются в открытую воду, где происходит оплодотворение. Они протандрические ; в течение первого года жизни они нерестятся как самцы, выпуская сперму в воду. По мере того, как они растут в течение следующих двух или трех лет и развивают большие запасы энергии, они нерестятся как самки, выпуская яйца . Нерестятся заливные устрицы обычно к концу июня. Повышение температуры воды побуждает к нересту нескольких устриц. Это вызывает нерест у остальных, замутняя воду миллионами яиц и сперматозоидов. Одна самка устрицы может производить до 100 миллионов яиц в год. Яйца оплодотворяются в воде и быстро развиваются в планктонных личинок . которые в конечном итоге находят подходящие места, например, раковину другой устрицы, где могут поселиться. Прикрепившиеся личинки устриц называются спатом . Спат — это устрицы длиной менее 25 миллиметров (0,98 дюйма).
Тихоокеанская устрица обычно раздельнополая. Их пол можно определить путем исследования гонад , и он может меняться из года в год, обычно в зимние месяцы. В определенных условиях окружающей среды один пол имеет преимущество над другим. Протандрия предпочтительна в районах с высоким изобилием пищи, а протогиния возникает в областях с низким изобилием пищи. В местах обитания с высоким запасом пищи соотношение полов во взрослом населении имеет тенденцию отдавать предпочтение самкам, а в районах с низким изобилием пищи, как правило, наблюдается большая доля взрослых самцов. Нерест тихоокеанских устриц происходит при температуре 20 ° C (68 ° F). Этот вид очень плодовит : самки выпускают около 50–200 миллионов икринок через равные промежутки времени (со скоростью 5–10 раз в минуту) за один нерест. После выхода из гонад яйца проходят через наджаберные камеры ( жабры ), затем через жаберные отверстия попадают в мантийную камеру и, наконец, выбрасываются в воду, образуя небольшое облако. У самцов сперма высвобождается на противоположном конце устрицы вместе с нормальной выдыхаемой струей воды. [ 57 ] Считается, что повышение температуры воды является основным сигналом к началу нереста, поскольку повышение температуры воды летом приводит к более раннему нересту тихоокеанских устриц. [ 58 ]
Личинки тихоокеанской устрицы планктотрофны , их размер на стадии продиссоконха 1 составляет около 70 мкм. Личинки перемещаются по толще воды с помощью личиночной стопы, чтобы найти подходящие места для поселения. На этом этапе они могут провести несколько недель, что зависит от температуры воды, солености и наличия пищи. За эти недели личинки могут рассеиваться на большие расстояния с помощью потоков воды, прежде чем они метаморфизируются и оседают в виде небольших брызг. Как и у других видов устриц, как только личинки тихоокеанских устриц находят подходящую среду обитания, они прикрепляются к ней навсегда, используя цемент, выделяемый железой на их стопе. После расселения личинки превращаются в молодь. Темпы роста очень высоки в оптимальных условиях окружающей среды, а объем рынка может быть достигнут за 18–30 месяцев. [ 59 ]
Головоногие моллюски , такие как кальмары и осьминоги, имеют выдающуюся голову и набор рук ( щупалец ), видоизмененных из примитивной стопы моллюсков. Все головоногие имеют половой диморфизм . Однако у них отсутствуют внешние половые признаки, поэтому они используют цветовое общение. Ухаживающий самец приближается к вероятно выглядящей партнерше, сверкая своими самыми яркими цветами, часто в виде ряби. Если другое головоногие - самка и восприимчива, ее кожа изменит цвет и станет бледной, и произойдет спаривание. Если другое головоногие моллюски остаются ярко окрашенными, это воспринимается как предупреждение. [ 60 ]
Все головоногие моллюски размножаются нерестовой икрой . Большинство головоногих моллюсков используют полувнутреннее оплодотворение, при котором самец помещает свои гаметы в мантийную полость самки , чтобы оплодотворить яйцеклетку самки в единственном яичнике . [ 61 ] «Пенис» у большинства самцов головоногих моллюсков представляет собой длинный и мускулистый конец гонодода, используемый для переноса сперматофоров в модифицированную несущую сперму руку, называемую гектокотилем . Это, в свою очередь, используется для передачи сперматофоров самке. У видов, у которых гектокотиль отсутствует, «пенис» длинный, может выходить за пределы мантийной полости и переносить сперматофоры непосредственно самке. У многих головоногих спаривание происходит «голова к голове», и самец может просто передать сперму самке. Другие могут отсоединить руку, несущую сперму, и оставить ее прикрепленной к самке. Глубоководные кальмары имеют самую большую известную длину пениса по отношению к размеру тела среди всех подвижных животных, уступая только некоторым сидячим ракушкам во всем животном мире . Удлинение пениса у большого крючковатого кальмара может привести к тому, что длина пениса будет равна длине мантии, головы и рук вместе взятых. [ 62 ] [ 63 ]
Некоторые виды высиживают оплодотворенные яйца: самки бумажных наутилусов строят убежища для детенышей, а гонатиидные кальмары несут нагруженную личинками оболочку с крючков на своих руках. [ 64 ] Другие головоногие откладывают детенышей под камни и проветривают их вылупляющимися щупальцами. В основном яйца предоставлены сами себе; Многие кальмары откладывают яйцеобразные сгустки, похожие на колбаски, в расщелинах или иногда на морском дне. Каракатицы откладывают икру отдельно в чехлы и прикрепляют их к коралловым или водорослевым листьям. [ 65 ] Как и тихоокеанский лосось, головоногие моллюски в основном являются семеплавильными , выметывая за одну порцию множество мелких икринок, а затем умирают. Головоногие моллюски обычно живут быстро и умирают молодыми. Большая часть энергии, получаемой из их пищи, используется для роста, и они быстро созревают до взрослого размера. Некоторые набирают до 12% массы тела каждый день. Большинство из них живут один-два года, размножаются, а затем вскоре умирают. [ 66 ] [ 67 ]
Иглокожие – морские животные, широко распространенные во всех океанах, но не встречающиеся в пресных водах. Прямо под их кожей находится эндоскелет, состоящий из известковых пластинок или косточек .
Морские ежи — это колючие иглокожие со сферическими телами, которые обычно содержат пять гонад . Они передвигаются медленно, питаются в основном морскими водорослями и играют важную роль в рационе каланов . Морские ежи раздельнополы , имеют отдельные мужской и женский пол, хотя, как правило, отличить их нелегко. Гонады покрыты мышцами под брюшиной , что позволяет животному выдавливать свои гаметы через проток в окружающую морскую воду, где происходит оплодотворение. [ 68 ] Их икра (мужские и женские половые железы) мягкая и тающая, ее цвет варьируется от оранжевого до бледно-желтого, и во многих частях мира ее используют как человеческий деликатес.
Морские огурцы — кожистые иглокожие с удлиненным телом, содержащим одну разветвленную гонаду. Они встречаются на морском дне по всему миру и в большом количестве встречаются на глубоководном морском дне, где они часто составляют большую часть биомассы животных . [ 69 ] Они питаются планктоном и разлагающимися органическими отходами, обнаруженными на морском дне, ловя протекающую мимо пищу раскрытыми щупальцами или просеивая донные отложения . Как и морские ежи, большинство морских огурцов размножаются путем выпуска спермы и яйцеклеток в океанскую воду. В зависимости от условий один организм может производить тысячи гамет .
Морские огурцы обычно раздельнополые , с отдельными мужскими и женскими особями. Репродуктивная система состоит из единственной гонады , состоящей из скопления канальцев, впадающих в один проток, открывающийся на верхней поверхности животного, рядом со щупальцами. [ 68 ] Многие виды оплодотворяют яйца внутренне. Оплодотворенная яйцеклетка развивается в мешочке на теле взрослой особи и в конечном итоге вылупляется как молодой морской огурец. [ 70 ] Некоторые виды вынашивают детенышей внутри полости тела, рожая через небольшой разрыв в стенке тела рядом с анусом. Остальные виды развивают яйца в свободно плавающую личинку обычно примерно через три дня развития. Эта личинка плавает с помощью длинного пучка ресничек, обернутого вокруг ее тела. По мере роста личинка превращается в бочкообразное тело с тремя-пятью отдельными кольцами ресничек. Щупальца обычно появляются первыми у взрослых особей, прежде чем обычные трубчатые ножки. [ 68 ]
Земноводные успешно решили большинство проблем, связанных с воздействием воздуха. Но их репродуктивная система была и остается связана с водой и остается очень похожей на рыбью. Почти все земноводные нерестятся в воде и откладывают большое количество мелких яиц, из которых быстро вылупляются плавающие личинки. Яйца не нуждаются в какой-либо комплексной защите от высыхания, поскольку при высыхании среды личинки обречены так же, как и яйца. Таким образом, отбор способствовал выбору подходящих мест для откладки яиц, а не подходящих устройств для защиты яиц. И рыбы, и амфибии могут мигрировать на большие расстояния в поисках нереста, и предпочтительные места часто активно оспариваются.
Обыкновенные лягушки сортируют свою икру Икра лягушки вблизи Развитие лягушачьей икры
Земноводные обитают в пресноводных озерах и прудах и вокруг них, но не в морской среде. Примерами являются лягушки и жабы, саламандры , тритоны и червяги (напоминающие змей). Это хладнокровные животные, которые превращаются из юной вододышащей формы, обычно во взрослую воздушно-дышащую форму, хотя грязевые щенки сохраняют молодые жабры и во взрослом возрасте.
Самки лягушек и жаб обычно выметывают студенистые яичные массы, содержащие тысячи икринок в воде. Различные виды откладывают яйца разными и узнаваемыми способами. Например, американская жаба откладывает длинные ниточки яиц. Яйца очень уязвимы для хищников , поэтому лягушки разработали множество методов, обеспечивающих выживание следующего поколения. В более холодных регионах эмбрион черный, чтобы поглощать больше солнечного тепла, что ускоряет развитие. Чаще всего это предполагает синхронное воспроизведение . Многие особи будут размножаться одновременно, подавляя действия хищников; большая часть потомства все равно умрет из-за хищников, но есть больше шансов, что некоторые выживут. Другой способ, которым некоторые виды избегают хищников и яиц болезнетворных микроорганизмов, которым подвергаются яйца в прудах, - это откладывать яйца на листьях над прудом с желеобразным покрытием, предназначенным для удержания влаги. У этих видов головастики после вылупления падают в воду. Яйца некоторых видов, отложенные из воды, могут улавливать вибрации находящихся поблизости хищных ос или змей и вылупляются рано, чтобы их не съели. [ 72 ]
Хотя продолжительность стадии яйца зависит от вида и условий окружающей среды, водные яйца обычно вылупляются в течение одной недели. В отличие от саламандр и тритонов, лягушки и жабы никогда не достигают половой зрелости, находясь еще на личиночной стадии. Вылупившиеся яйца продолжают жизнь в виде головастиков , которые обычно имеют овальные тела и длинные, сплюснутые по вертикали хвосты. Как правило, свободноживущие личинки полностью водны. У них отсутствуют веки, они имеют хрящевой скелет, систему боковой линии , жабры для дыхания (сначала наружные, позже внутренние) и хвост с дорсальными и брюшными складками кожи для плавания. [ 73 ] У них быстро развивается жаберный мешок, закрывающий жабры и передние ноги; легкие также развиваются на ранней стадии как вспомогательный орган дыхания. У некоторых видов, которые проходят метаморфоз внутри яйца и вылупляются в маленьких лягушек, никогда не развиваются жабры; вместо этого есть специальные участки кожи, которые отвечают за дыхание. У головастиков также отсутствуют настоящие зубы, но челюсти у большинства видов обычно имеют два удлиненных параллельных ряда небольших ороговевших структур, называемых керадонтами, в верхней челюсти, тогда как нижняя челюсть имеет три ряда керадонтов, окруженных роговым клювом , но количество рядов может быть ниже (иногда ноль) или значительно выше. [ 74 ] Головастики питаются водорослями , в том числе диатомовыми водорослями , фильтрующимися из воды через жабры . Некоторые виды на стадии головастика плотоядны и поедают насекомых, более мелких головастиков и рыбу. каннибализм Среди головастиков наблюдался . Ранние развивающиеся особи, получившие ноги, могут быть съедены другими, поэтому поздно расцветшие выживут дольше. [ 75 ]
Морские черепахи — земноводные рептилии, но не земноводные . Рептилии относятся к классу Рептилии , а земноводные — к классу Амфибии . Это две отдельные таксономические группы. У рептилий чешуя и кожистая кожа, а у амфибий кожа гладкая и пористая. В отличие от лягушек, яйца морских черепах имеют прочную кожистую скорлупу, которая позволяет им выживать на суше, не высыхая.
Некоторые морские черепахи мигрируют на большие расстояния между местами кормления и нереста. Зеленые черепахи имеют места кормления вдоль побережья Бразилии. Каждый год тысячи этих черепах мигрируют на расстояние около 2300 километров (1400 миль) к месту своего нереста, острову Вознесения в Атлантике, острову шириной всего 11 километров (6,8 миль). Каждый год возвращающиеся черепахи выкапывают от 6 000 до 15 000 гнезд, часто возвращаясь на тот же пляж, откуда они вылупились. Самки обычно спариваются каждые два-четыре года. С другой стороны, самцы каждый год посещают места размножения, пытаясь спариваться. [ 76 ] Спаривание зеленых морских черепах похоже на спаривание других морских черепах. Самки черепах контролируют процесс. Некоторые популяции практикуют полиандрию , хотя это, похоже, не приносит пользы птенцам. [ 77 ] После спаривания в воде самка поднимается над линией прилива на пляже, выкапывает яму задними ластами и откладывает икру. Размер помета зависит от возраста самки и вида, но количество кладок зеленой черепахи колеблется от 100 до 200. Затем она засыпает гнездо песком и возвращается в море. [ 78 ]
Примерно через 45–75 дней яйца вылупляются ночью, и птенцы инстинктивно направляются прямо в воду. Это самое опасное время в жизни черепахи. Пока они идут, их хватают хищники, такие как чайки и крабы . Значительный процент никогда не добирается до океана. Мало что известно о первоначальной истории жизни только что вылупившихся морских черепах. [ 79 ] Молодые особи проводят от трех до пяти лет в открытом океане, прежде чем еще незрелыми молодыми особями освоятся и начнут вести постоянный образ жизни на мелководье. [ 80 ] [ 81 ] Предполагается, что им требуется от двадцати до пятидесяти лет, чтобы достичь половой зрелости . Особи живут до восьмидесяти лет в дикой природе. [ 78 ] Они относятся к числу самых крупных морских черепах, многие из которых имеют длину более метра и вес до 300 килограммов (660 фунтов). [ 82 ]
водные насекомые Нерестятся и . Подёнки «славятся своей короткой взрослой жизнью. У некоторых видов есть меньше часа, чтобы спариваться и откладывать яйца, прежде чем они умрут. Их предвзрослая стадия, известная как субимаго, может быть еще короче — возможно, она длится всего несколько минут, прежде чем они умрут. линяют во взрослую форму, поэтому подёнка проводит большую часть своей жизни в виде нимфы, спрятанной от глаз под водой». [ 83 ]
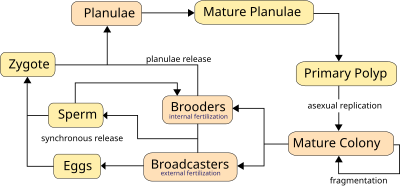
Кораллы могут быть как гонохорическими (однополыми), так и гермафродитными , каждый из которых может размножаться половым и бесполым путем. Размножение также позволяет кораллам заселять новые территории.
Кораллы размножаются преимущественно половым путем . 25% герматипических кораллов (каменистых кораллов) образуют однополые ( гонохористические ) колонии, остальные — гермафродиты . [ 36 ] Около 75% всех герматипических кораллов «транслируют икру», выпуская гаметы — яйца и сперму — в воду для распространения потомства. Гаметы сливаются во время оплодотворения, образуя микроскопическую личинку , называемую планулой , обычно розового цвета и эллиптической формы. Типичная колония кораллов образует несколько тысяч личинок в год, чтобы преодолеть препятствия к образованию новой колонии. [ 84 ]
Планулы демонстрируют положительный фототаксис , плывя к свету, чтобы достичь поверхностных вод, где они дрейфуют и растут, прежде чем спуститься в поисках твердой поверхности, к которой они могут прикрепиться и основать новую колонию. Они также демонстрируют положительный сонотаксис , двигаясь навстречу звукам, исходящим от рифа, и вдали от открытой воды. [ 85 ] Высокая частота неудач затрагивает многие стадии этого процесса, и хотя каждая колония высвобождает миллионы гамет, образуется очень мало новых колоний. Время от нереста до заселения обычно составляет 2–3 дня, но может достигать 2 месяцев. [ 86 ] Личинка превращается в полип и в конечном итоге становится коралловой головой в результате бесполого почкования и роста.
Синхронный нерест очень типичен для коралловых рифов, и часто, даже если несколько видов присутствует , все кораллы нерестятся в одну и ту же ночь. Эта синхронность необходима для того, чтобы мужские и женские гаметы могли встретиться. Кораллам приходится полагаться на сигналы окружающей среды, варьирующиеся от вида к виду, чтобы определить подходящее время для выпуска гамет в воду. Сигналы включают лунные изменения, время заката и, возможно, химические сигналы. [ 36 ] Синхронный нерест может образовывать гибриды и, возможно, участвует в видообразовании кораллов . [ 87 ] В некоторых местах нерест может быть визуально эффектным: обычно прозрачная вода замутняется гаметами, обычно ночью.
Кораллы используют два метода полового размножения, которые различаются тем, выделяются ли женские гаметы:
Вещатели , большинство из которых массово нерестятся, в значительной степени полагаются на сигналы окружающей среды, поскольку они выпускают в воду как сперму, так и яйцеклетки. Кораллы используют долгосрочные сигналы, такие как продолжительность дня , температура воды и/или скорость изменения температуры. Краткосрочным сигналом чаще всего является лунный цикл с закатом, сигнализирующим об освобождении. [ 36 ] Около 75% видов кораллов являются трансляторами, большинство из которых являются герматипическими кораллами, или кораллами, строящими рифы. [ 36 ] Положительно плавучие гаметы всплывают к поверхности, где в результате оплодотворения образуются личинки планулы . Личинки плывут к поверхности света и попадают в течения, где обычно остаются в течение двух дней, но иногда до трех недель, а в одном известном случае - два месяца. [ 86 ] после чего они расселяются и превращаются в полипы и образуют колонии.
Брудеры чаще всего являются агерматипическими (постройками без рифов) в районах с сильным течением или волнением. Брудеры выделяют только сперму, которая имеет отрицательную плавучесть и может содержать неоплодотворенные икринки в течение нескольких недель, что снижает потребность в массовом синхронном нересте, который иногда случается. [ 36 ] После оплодотворения кораллы выпускают личинки планулы , готовые к оседанию. [ 88 ]
Сбор урожая вешенки Pleurotus ostreatus , выращенной с использованием икры, помещенной в смесь опилок , помещенную в пластиковые контейнеры.
Грибы не являются растениями, и для оптимального роста им требуются разные условия. Растения развиваются посредством фотосинтеза – процесса, который превращает углекислый газ из атмосферы в углеводы , особенно в целлюлозу . В то время как солнечный свет является источником энергии для растений, грибы получают всю свою энергию и материалы для роста из среды роста посредством биохимического разложения процессов . Это не означает, что свет является ненужным требованием, поскольку некоторые грибы используют свет как сигнал, вызывающий плодоношение. Однако все материалы для роста уже должны присутствовать в питательной среде. Вместо семян грибы размножаются половым путем при подземном росте и бесполым путем спорами . Любой из них может быть заражен переносимыми по воздуху микроорганизмами, которые будут мешать росту грибов и препятствовать получению здорового урожая. Мицелий , или активно растущая грибная культура, помещается на субстрат для посева или внесения грибов для выращивания на субстрате . Это также известно как прививка, нерест или добавление икры. Его основные преимущества заключаются в снижении вероятности заражения и придании грибам твердого начала. [ 89 ] [ 90 ]
^ "Нерест". Британская энциклопедия. Британская энциклопедия Интернет. Британская энциклопедия, 2011. Интернет. 03 февраля 2011 г. < «Нерест | биология | Британника» . Архивировано из оригинала 2 ноября 2012 г. Проверено 5 февраля 2011 г. >
^ Гросс МР (1982). «Кроссовки, спутники и родители: стратегии полиморфного спаривания североамериканских солнечных рыб». Журнал психологии животных . 60 :1-26. дои : 10.1111/j.1439-0310.1982.tb01073.x .
^ Перейти обратно: а б Шапиро Д.Ю. (1984) «Реверсия пола и социально-демографические процессы у коралловых рифовых рыб», страницы 103–116 в Г.В. Поттсе и Р.К. Вутуне, ред., Размножение рыб: стратегии и тактика , Academic Press.
^ Робертсон, доктор медицинских наук; Р. Р. Уорнер (1978). «Половые особенности лаброидов Западного Карибского бассейна II: рыбы-попугаи (Scaridae)». Смитсоновский вклад в зоологию . 255 (255): 1–26. дои : 10.5479/si.00810282.255 .
^ Чан СТХ и Юнг ВСБ (1983) «Контроль над сексом и смена пола у рыб в естественных условиях». Страницы 171–222 в WS Hoar, DJ Randall и EM Donaldson, ред., Физиология рыб 9B: Размножение, поведение и контроль фертильности . Академическая пресса.
^ Перейти обратно: а б Вильялмссон, Х (2002). «Мойва (Mallotus villosus) в экосистеме Исландия – Восточная Гренландия – Ян-Майен». Журнал морских наук ICES . 59 (5): 870–883. дои : 10.1006/jmsc.2002.1233 .
^ Бранч Джордж, Бранч, Марго и Баннистер, Энтони (1981). Живые берега Южной Африки . Кейптаун: К. Струик. ISBN 978-0-86977-115-0 . {{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка )
Arc.Ask3.Ru Номер скриншота №: 75c6c97cd24d2a362fe686f617359e5e__1721276880 URL1:https://arc.ask3.ru/arc/aa/75/5e/75c6c97cd24d2a362fe686f617359e5e.html Заголовок, (Title) документа по адресу, URL1: Spawn (biology) - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)












 Baby paradise fish just hatched, gathered under the surface of a bubble nest
Baby paradise fish just hatched, gathered under the surface of a bubble nest Anemone fish nest in an anemone. Here a male is protecting spawn produced by his partner.
Anemone fish nest in an anemone. Here a male is protecting spawn produced by his partner.

Mouthbrooding tilapia — YouTube






 Сиамские бойцовые рыбы строят пузырьковые гнезда разного размера.
Сиамские бойцовые рыбы строят пузырьковые гнезда разного размера. Пара сиамских бойцовских рыб, нерестящихся под своим пузырьковым гнездом.
Пара сиамских бойцовских рыб, нерестящихся под своим пузырьковым гнездом. Однодневные личинки сиамских боевых рыб в пузырьковом гнезде — их желточные мешки еще не рассосались
Однодневные личинки сиамских боевых рыб в пузырьковом гнезде — их желточные мешки еще не рассосались 15-дневный малек сиамской бойцовой рыбы, свободно плавающий.
15-дневный малек сиамской бойцовой рыбы, свободно плавающий.


















 Нерест хрупкой звезды
Нерест хрупкой звезды Голова самки криля с выводковым мешком
Голова самки криля с выводковым мешком Нерест нерки
Нерест нерки Мертвый лосось после нереста
Мертвый лосось после нереста Эти лагуны, соединенные с рекой Тис, обеспечивают тихую заводь для нереста рыбы и убежища во время паводка.
Эти лагуны, соединенные с рекой Тис, обеспечивают тихую заводь для нереста рыбы и убежища во время паводка. В середине этой плотины находится рыбная лестница , которая позволяет форели и лососю пройти через плотину и подняться вверх по реке на нерест.
В середине этой плотины находится рыбная лестница , которая позволяет форели и лососю пройти через плотину и подняться вверх по реке на нерест.







