Морские микроорганизмы по -разному составляют около 70%, [ 4 ] или около 90%, [ 5 ] [ 6 ] биомассы в океане . Взятые вместе они образуют морской микробиом . Через миллиарды лет этот микробиом развил много стилей жизни и адаптации и участвует в глобальном велосипеде практически всех химических элементов. [ 7 ] Микроорганизмы имеют решающее значение для утилизации питательных веществ в экосистемах , поскольку они действуют как декомпозиер . Они также ответственны за почти весь фотосинтез , который происходит в океане, а также на велосипеде углерода , азота , фосфора и других питательных веществ и следовых элементов. [ 8 ] Морские микроорганизмы изобилуют большие количества углерода и производят большую часть мирового кислорода.
Небольшая часть морских микроорганизмов является патогенными , вызывая заболевания и даже смерть у морских растений и животных. [ 9 ] Однако морские микроорганизмы перерабатывают основные химические элементы , которые производят и потребляют около половины всего органического вещества, генерируемого на планете каждый год. Как жители крупнейшей среды на Земле, микробные морские системы ведут изменения в каждой глобальной системе.
В июле 2016 года ученые сообщили о выявлении набора из 355 генов из последнего универсального общего предка (Лука) всей жизни на планете, включая морские микроорганизмы. [ 10 ] Несмотря на свое разнообразие, микроскопическая жизнь в океанах все еще плохо изучена. Например, роль вирусов в морских экосистемах едва изучена даже в начале 21 -го века. [ 11 ]
Микроорганизмы имеют решающее значение для утилизации питательных веществ в экосистемах , поскольку они действуют как декомпозиер . Некоторые микроорганизмы являются патогенными , вызывающие заболевание и даже смерть у растений и животных. [ 9 ] Как жители крупнейшей среды на Земле, микробные морские системы ведут изменения в каждой глобальной системе. Микробы отвечают практически за весь фотосинтез , который происходит в океане, а также на велосипеде углерода , азота , фосфора и других питательных веществ и следовых элементов. [ 8 ]
Хотя недавние технологические разработки и научные открытия были существенными, нам по -прежнему не хватает особого понимания на всех уровнях основных экологических вопросов в отношении микроорганизмов в наших морях и океанах. Эти фундаментальные вопросы:
1. Что там? Какие микроорганизмы присутствуют в наших морях и океанах и в каких числах
они встречаются?
2. Что они делают? Какие функции выполняют каждый из этих микроорганизмов в морской среде и как они способствуют глобальным циклам энергии и материи?
3. Каковы факторы, которые определяют наличие или отсутствие микроорганизма и как они влияют на биоразнообразие и функции и наоборот?
Диапазон размеров, показанных прокариотами (бактериями и археей) и вирусами относительно видов других организмов и биомолекул Под увеличением, всплеск морской воды замирает микроскопической жизнью
Микроскопическая жизнь подводная жизнь разнообразна и все еще плохо изучена, например, для роли вирусов в морских экосистемах. [ 13 ] Большинство морских вирусов - это бактериофаги , которые безвредны для растений и животных, но имеют важное значение для регуляции соленой и пресноводных экосистем. [ 14 ] Они заражают и разрушают бактерии в водных микробных сообществах и являются наиболее важным механизмом утилизации углерода в морской среде. Органические молекулы, высвобождаемые из мертвых бактериальных клеток, стимулируют рост свежих бактерий и водорослей. [ 15 ] способствовать биологическому насосу , процесс, посредством которого углерод изолируется Вирусная активность также может в глубоком океане. [ 16 ]
Морский спрей, содержащий морские микроорганизмы, можно охватить высоко в атмосферу, где они становятся аэропланктоном , и могут путешествовать по миру, прежде чем упасть на Землю.
Поток воздушных микроорганизмов окружает планету над погодными системами, но ниже коммерческих воздушных дорожек. [ 17 ] Некоторые перипатетические микроорганизмы охвачены от наземных пыльных бури, но большинство из них происходят из морских микроорганизмов при морском спрея . В 2018 году ученые сообщили, что сотни миллионов вирусов и десятков миллионов бактерий ежедневно откладываются на каждом квадратном метре вокруг планеты. [ 18 ] [ 19 ]
Микроскопические организмы живут на протяжении всей биосферы . Масса прокариот -микроорганизмов, которые включают бактерии и археи, но не зародышевые микроорганизмы эукариот -микроорганизмов , может составлять до 0,8 триллионов углерода (общей биосферной массы , оцениваемой в 1-4 триллиона тонн). [ 20 ] Одноклеточные барофильные морские микробы были обнаружены на глубине 10 900 м (35 800 футов) в траншеи Мариан , самого глубокого места в океанах Земли. [ 21 ] [ 22 ] Микроорганизмы живут в скалах 580 м (1900 футов) ниже морского пола под 2590 м (8500 футов) океана у побережья северо -запада Соединенных Штатов , [ 21 ] [ 23 ] а также 2400 м (7900 футов; 1,5 мили) под морским дном у Японии. [ 24 ] Наибольшая известная температура, при которой может существовать срок службы микробов, составляет 122 ° C (252 ° F) ( Methanopyrus kandleri ). [ 25 ] В 2014 году ученые подтвердили существование микроорганизмов, живущих на 800 м (2600 футов) ниже льда Антарктиды . [ 26 ] [ 27 ] По словам одного исследователя, «вы можете найти микробы повсюду - они чрезвычайно адаптируются к условиям и выживают, где бы они ни находились». [ 21 ] Морские микроорганизмы служат «основой всех морских пищевых сетей, утилизации основных элементов и производства и потребления около половины органического вещества, создаваемого на земле каждый год». [ 28 ] [ 29 ]
Когда не внутри зараженной клетки или в процессе инфекции клетки, вирусы существуют в форме независимых частиц. Эти вирусные частицы, также известные как вирионы , состоят из двух или трех частей: (i) генетический материал (геном), изготовленный из ДНК или РНК , длинных молекул , которые несут генетическую информацию; (ii) белковое пальто, называемое капсидом , которое окружает и защищает генетический материал; и в некоторых случаях (iii) оболочка липидов , которая окружает белковое пальто, когда они находятся за пределами клетки. Формы этих вирусных частиц варьируются от простых спиральных и икосаэдрических форм для некоторых видов вирусов до более сложных структур для других. Большинство видов вирусов имеют вирионы, которые слишком малы, чтобы их можно было увидеть с помощью оптического микроскопа . Средний вирион составляет около одного сотого размера средней бактерии .
Происхождение вирусов в эволюционной истории жизни неясно: некоторые, возможно, развивались из плазмид - части ДНК, которые могут перемещаться между клетками, в то время как другие могли развиваться из бактерий. В эволюции вирусы являются важным средством горизонтального переноса генов , что увеличивает генетическое разнообразие . [ 32 ] Некоторые считают вирусами жизненной формой, потому что они несут генетический материал, воспроизводят и развиваются посредством естественного отбора . Тем не менее, им не хватает ключевых характеристик (таких как структура клеток), которые обычно считаются необходимыми для сведения как жизнь. Поскольку они обладают некоторыми, но не всеми такими качествами, вирусы были описаны как «организмы на грани жизни» [ 33 ] и как репликаторы. [ 34 ]
Бактериофаги (фаги)
Несколько фагов, прикрепленных к бактериальной клеточной стенке при увеличении 200 000 раз
Вирусы встречаются везде, где бывает жизнь и, вероятно, существовали с тех пор, как живые клетки впервые развивались. [ 35 ] Происхождение вирусов неясно, потому что они не образуют окаменелости, поэтому молекулярные методы использовались для сравнения ДНК или РНК вирусов и являются полезным средством изучения того, как они возникли. [ 36 ]
Вирусы теперь признаются древними и имеют происхождение, которое предварительно дает дивергенцию жизни в три области . [ 37 ]
Мнения отличаются от того, являются ли вирусы формой жизни или органических структур, которые взаимодействуют с живыми организмами. [ 34 ] Некоторые считают, что они являются жизненной формой, потому что они несут генетический материал, воспроизводясь, создавая несколько копий самих себя посредством самосборки и развиваются посредством естественного отбора . Однако им не хватает ключевых характеристик, таких как клеточная структура, как правило, считается необходимым для считать жизнь. Поскольку они обладают некоторыми, но не всеми такими качествами, вирусы были описаны как репликаторы [ 34 ] и как «организмы на краю жизни». [ 33 ]
Бактериофаги , часто называемые фагами , являются вирусами, которые паразиты бактерии и археи. Морские фаги паразит морские бактерии и археи, такие как цианобактерии . [ 38 ] Они являются общей и разнообразной группой вирусов и являются наиболее распространенной биологической сущностью в морской среде, потому что их хозяева, бактерии, обычно являются численной доминирующей клеточной жизнью в море. Как правило, в каждой мл морской воды насчитывается от 1 миллиона до 10 миллионов вирусов, или примерно в десять раз больше двухцепочечных ДНК-вирусов, чем клеточные организмы, [ 39 ] [ 40 ] Хотя оценки вирусного изобилия в морской воде могут варьироваться в широком диапазоне. [ 41 ] [ 42 ]
В течение долгого времени хвостовые фаги ордена Каудовиралс, по -видимому, доминировали в морских экосистемах по количеству и разнообразию организмов. [ 38 ]
Однако в результате более недавних исследований не хвост вирусы, по-видимому, доминируют по нескольким глубинах и океаническим регионам, за которыми следуют семьи каудовирал миовирусов, подовирусов и сифавирусов. [ 43 ]
Фаги, принадлежащие семьям:
Corticoviridae , [ 44 ] Inoviridae , [ 45 ] Microviridae , [ 46 ]
и Autolykiviridae [ 47 ] [ 48 ] [ 49 ] [ 50 ]
Также известно, что заражают разнообразные морские бактерии.
Существуют также архейские вирусы, которые реплицируются в археи : это двухцепочечные ДНК-вирусы с необычными, а иногда и уникальными формами. [ 51 ] [ 52 ] Эти вирусы были изучены наиболее подробно в термофильной археи, в частности, заказы сульфобалы и термопротеалы . [ 53 ]
Микроорганизмы составляют около 70% морской биомассы. [ 4 ] По оценкам, вирусы убивают 20% этой биомассы каждый день, и что в океанах в 15 раз больше вирусов, чем бактерии и археи. Вирусы являются основными агентами, ответственными за быстрое разрушение вредных цветов водорослей , [ 40 ] которые часто убивают другую морскую жизнь. [ 54 ]
Количество вирусов в океанах уменьшается дальше от берега и глубже в воду, где меньше организмов хозяина. [ 16 ]
Вирусы являются важным естественным средством передачи генов между различными видами, что увеличивает генетическое разнообразие и развивает эволюцию. [ 32 ] Считается, что вирусы сыграли центральную роль в ранней эволюции, до диверсификации бактерий, археи и эукариот, во время последнего универсального общего предка жизни на Земле. [ 55 ] Вирусы по -прежнему являются одним из крупнейших резервуаров неисследованного генетического разнообразия на Земле. [ 16 ]
Вирусы обычно варьируются в длину от 20 до 300 нанометров. Это может быть контрастировано с длиной бактерий, которая начинается с 400 нанометров. Существуют также гигантские вирусы , часто называемые прожилками , как правило, около 1000 нанометров (один микрон) в длину.
Все гигантские вирусы принадлежат к филому нуклеоцитовирикоты (NCLDV), вместе с оспами .
Самым крупным известным из них является Tupanvirus . Этот род гигантского вируса был обнаружен в 2018 году в глубоком океане, а также в газированном озере, и может достигать 2,3 микрона общей длины. [ 56 ]
Открытие и последующая характеристика гигантских вирусов вызвали некоторые дебаты, касающиеся их эволюционного происхождения. [ 57 ] Две основные гипотезы для их происхождения заключаются в том, что либо они эволюционировали из небольших вирусов, забирая ДНК из организмов-хозяев, либо эволюционировали из очень сложных организмов в нынешнюю форму, которая не является самодостаточной для размножения. [ 58 ] Из каких сложных вирусов гиганта организма могли бы расходиться, также является темой дебатов. Одним из предложений является то, что точка происхождения фактически представляет четвертую домену жизни, [ 59 ] [ 60 ] Но это было в значительной степени дисконтировано. [ 61 ] [ 62 ]
Предки современных бактерий были одноклеточными микроорганизмами, которые были первыми формами жизни , появившимися на Земле, около 4 миллиардов лет назад. В течение примерно 3 миллиардов лет большинство организмов были микроскопическими, а бактерии и археи были доминирующими формами жизни. [ 65 ] [ 66 ] Хотя существуют бактериальные окаменелости , такие как строматолиты , их отсутствие отличительной морфологии не позволяет их использовать для изучения истории эволюции бактерий или до настоящего времени времени происхождения конкретных видов бактерий. Тем не менее, последовательности генов могут использоваться для реконструкции бактериальной филогении , и эти исследования показывают, что бактерии смещались в первую очередь от архейной/эукариотической линии. [ 67 ]
Бактерии также участвовали во второй великой эволюционной дивергенции, у археи и эукариот. Здесь эукариоты были получены в результате входа в древние бактерии в эндосимбиотические ассоциации с предками эукариотических клеток, которые сами по себе были связаны с архей . [ 68 ] [ 69 ] Это включало в себя поглощение протоукариотическими клетками альфапротеобактериальных симбионтов с образованием либо митохондрий , либо водородных средств , которые все еще обнаруживаются во всех известных эукарья. Позже некоторые эукариоты, которые уже содержали митохондрии, также охватили цианобактериальные организмы. Это привело к образованию хлоропластов в водорослях и растениях. Есть также некоторые водоросли, которые возникли даже из -за более поздних эндосимбиотических событий. Здесь эукариоты охватили эукариотические водоросли, которые превратились в пластид «второго поколения». [ 70 ] [ 71 ] Это известно как вторичный эндосимбиоз .
Pelagibacter Ubique и его родственники могут быть самыми распространенными организмами в океане, и было утверждено, что они, возможно, являются самыми распространенными бактериями в мире. Они составляют около 25% всех клеток микробного планктона , и летом они могут объяснить примерно половину клеток, присутствующих в поверхностных водах умеренного океана. Общая численность P. ubique и родственников оценивается около 2 × 10 28 Микробы. [ 73 ] HTVC010P бактериофаг Тем не менее, в феврале 2013 года сообщалось, что , который атакует P. ubique , был обнаружен, и «вероятно, это действительно самый распространенный организм на планете». [ 74 ] [ 75 ]
Археи первоначально рассматривались как экстремофилы, живущие в суровых условиях, такие как желтая археа, изображенная здесь в горячем источнике , но с тех пор они были обнаружены в гораздо более широком диапазоне среды обитания . [ 78 ]
Археи были первоначально классифицированы как бактерии , но эта классификация устарела. [ 80 ] Архаальные клетки обладают уникальными свойствами, отделяющими их от двух других доменов жизни, бактерий и эукариоты . Археи дополнительно разделены на множественные признанные фила . Классификация сложна, потому что большинство не были изолированы в лаборатории и были обнаружены только путем анализа их нуклеиновых кислот в образцах из окружающей среды.
Археи особенно многочисленны в океанах, и археи в планктоне может быть одной из самых распространенных групп организмов на планете. Археи являются основной частью жизни Земли и могут играть роль как в углеродном цикле , так и в цикле азота . Термопротеота (также известная как эоциты или Crenarchaeota) представляют собой тип археи, который считается очень распространенным в морской среде и одним из основных участников фиксации углерода. [ 82 ]
Эоциты могут быть наиболее распространенными из морской археи
Halobacteria , обнаруженные в воде, почти насыщенные солью, теперь признаются археей.
Все живые организмы могут быть сгруппированы как прокариоты или эукариоты . Жизнь возникла как одноклеточные прокариоты, а затем превратилась в более сложные эукариоты. В отличие от прокариотических клеток, эукариотические клетки высокоорганизованы. Прокариоты являются бактериями и археей, в то время как эукариоты являются другими формами жизни - протистами , растениями, грибами и животными. Протисты обычно одноклеточны, в то время как растения, грибы и животные обычно многоклеточны .
Кажется очень правдоподобным, что корень эукариот лежит в археи; Самыми ближайшими родственниками в настоящее время известны, могут быть филом Heimdallarchaeota предлагаемого суперфилума Asgard . Эта теория является современной версией сценария, первоначально предложенного в 1984 году как гипотеза эоцитов , когда термопротета были самыми близкими известными архейными родственниками эукариот.
Возможная переходная форма микроорганизма между прокариотом и эукариотом была обнаружена в 2012 году японскими учеными. Parakaryon Myojinensis - это уникальный микроорганизм, больше, чем типичный прокариот, но с ядерным материалом, заключенным в мембрану, как в эукариоте, и наличие эндосимбионтов . Это является первой правдоподобной эволюционной формой микроорганизма, демонстрирующей стадию развития от прокариота до эукариоте. [ 83 ] [ 84 ]
Протисты - это эукариоты, которые не могут быть классифицированы как растения, грибы или животные. Обычно они одноклеточные и микроскопические. Жизнь возникла как одноклеточные прокариоты (бактерии и археи), а затем превратились в более сложные эукариоты . Эукариоты - более развитые формы жизни, известные как растения, животные, грибы и протисты. Термин «Протист» исторически использовался в качестве условия удобства для эукариот, которые не могут быть строго классифицированы как растения, животные или грибы. Они не являются частью современной кладистики, потому что они парафилетические (отсутствуют общего предка).
Протисты могут быть в целом разделены на четыре группы в зависимости от того, является ли их питание, похожее на животные, похожие на грибковые, похожие на грибку, [ 85 ] или смесь из них. [ 86 ]
Протисты в соответствии с тем, как они получают еду
Протисты представляют собой очень разнообразные организмы, в настоящее время организованные в 18 Phyla, но их нелегко классифицировать. [ 89 ] [ 90 ] Исследования показали, что высокое разнообразие протиста существует в океанах, глубоких морских и речных отложениях, что позволяет предположить, что большое количество эукариотических микробных сообществ еще не обнаружено. [ 91 ] [ 92 ] Было мало исследований по миксотрофным протистам, но недавние исследования в морских средах показали, что миксотрофные протесты вносят значительный вклад в профистную биомассу . [ 87 ] Поскольку протисты являются эукариотами, которые они обладают в своей клетке, по крайней мере, в одном ядре , а также органелле, таких как митохондрии и тела Гольджи . Протисты бесполые, но могут быстро воспроизводить через митоз или фрагментацию .
Одноклеточные и микроскопические протисты
Диатомы являются основной группой водорослей, генерирующей около 20% мировой продукции кислорода. [ 93 ]
В отличие от клеток прокариот, клетки эукариот очень организованы. Растения, животные и грибы обычно являются многоклеточными и обычно макроскопические . Большинство протистов являются одноклельными и микроскопическими. Но есть исключения. Некоторые одноклеточные морские протисты макроскопичны. Некоторые морские плесень слизи имеют уникальные жизненные циклы, которые включают переключение между одноклеточными, колониальными и многоклеточными формами. [ 95 ] Другие морские протесты не являются ни одноклельными, ни микроскопическими, такими как морские водоросли .
Протисты были описаны как таксономический мешок с закусками, где может быть размещено все, что не вписывается в одно из основных биологических королевств . [ 98 ] Некоторые современные авторы предпочитают исключать многоклеточные организмы из традиционного определения протиста, ограничивающего протисты одноклеточными организмами. [ 99 ] [ 100 ] Это более ограниченное определение исключает много коричневых , многоклеточных красных и зеленых водорослей и слизи . [ 101 ]
Другой способ классификации протистов - в соответствии с их способом передвижения. Многие одноклеточные протисты, особенно простейшие, являются подвижными и могут генерировать движение, используя жгутиков , ресничек или псевдопод . Клетки, которые используют жгутиков для движения, обычно называют жгутиками , клетки, которые используют реснички, обычно называют ресничку , а клетки, которые используют псевдопод, обычно называют амебой или амебоидами . Другие протисты не являются подвижными и, следовательно, не имеют механизма движения.
Плачениц )-это придаток , (латынь для кнута похожий на ресниц, который выступает из клеточного тела некоторых протистов (а также некоторых бактерий). Плаговки используют от одного до нескольких жгутиков для передвижения, а иногда в качестве кормления и сенсорной органеллы .
Ресничка ( латынь для ресниц ) - это крошечный жгутичок. Инрустры используют несколько ресничек, которые могут насмехаться за многие сотни, чтобы питать себя через воду.
Плаговки включают бактерии, а также протисты. Модель роторного двигателя, используемая бактериями, использует протоны электрохимического градиента для перемещения их жгутиков. Крутящий момент в жгутиках бактерий создается частицами, которые проводят протоны вокруг основания жгутика. Направление вращения жгутиков в бактериях происходит от занятости протонных каналов вдоль периметра жгутикового двигателя. [ 107 ]
Цилиаты, как правило, имеют сотни до тысяч ресничек, которые плотно упакованы вместе в массивах. Во время движения индивидуальный ресничек деформируется с использованием силового хода с высоким содержанием фарки с последующим ударом восстановления с низким содержанием фонаря. Поскольку на отдельном организме есть несколько ресничек, они демонстрируют коллективное поведение в метахрональном ритме . Это означает, что деформация одной реснички находится в фазе с деформацией его соседа, вызывая волны деформации, которые распространяются вдоль поверхности организма. Эти распространяющиеся волны ресничек - это то, что позволяет организму использовать реснички скоординированным образом для движения. Типичным примером ресничного микроорганизма является парамеция , одноклеточный ресничный протозой, покрытый тысячами ресничек. Избиение ресничек вместе позволяют парамециуму простираться через воду со скоростью 500 микрометров в секунду. [ 108 ]
Более 1500 видов грибов известны из морской среды. [ 109 ] Они паразита на морских водорослях или животных, или являются сапробами, питающимися мертвым органическим веществом из водорослей, кораллов, простейших кист, морских трав и других субстратов. [ 110 ] Споры многих видов имеют специальные придатки, которые облегчают привязанность к субстрату. [ 111 ] Морские грибы также можно найти в морской пене и вокруг гидротермальных районов океана. [ 112 ] разнообразные необычные вторичные метаболиты . Морские грибы производят [ 113 ]
Типичный миллилитр морской воды содержит около 10 3 до 10 4 Грибковые клетки. [ 119 ] Это число больше в прибрежных экосистемах и устьях из -за стока питания от наземных сообществ. Более высокое разнообразие микопланктона встречается вокруг побережья и в поверхностных водах до 1000 метров, с вертикальным профилем , который зависит от того, насколько обильным фитопланктоном является. [ 120 ] [ 121 ] Этот профиль изменяется между сезонами из -за изменений в доступности питательных веществ. [ 122 ] Морские грибы выживают в постоянной среде с дефицитом кислорода и, следовательно, зависят от диффузии кислорода с помощью турбулентности и кислорода, генерируемых фотосинтетическими организмами . [ 123 ]
Лишайники - это взаимные ассоциации между грибом, обычно аскомицетом , и водорослей или цианобактерией . Несколько лишайников встречаются в морской среде. [ 124 ] Многие другие встречаются в зоне всплеска , где они занимают разные вертикальные зоны в зависимости от того, насколько они терпимы к погружению. [ 125 ] Некоторые лишайники живут долго; Один вид был датирован на 8600 лет. [ 126 ] Однако их продолжительность жизни трудно измерить, потому что то, что определяет один и тот же лишайник, не точнее. [ 127 ] Лишайники растут путем вегетативно разбивающегося кусочка, который может быть или не быть определен как один и тот же лишайник, и две лишайники разных возрастов могут слиться, поднимая проблему того, является ли это одним и тем же лишайником. [ 127 ]
Морская улитка Tittoraria Irrorata наносит ущерб растениям Спартина в морских болотах, где она живет, что позволяет спорам литоральных ашо -амортизированных грибов колонизировать растение. Улитка тогда ест грибковой рост в предпочтениях самой траве. [ 128 ]
Согласно Fossil Records, грибы датируются покойной протерозойской эпохой 900-570 миллионов лет назад. В Китае было обнаружено, что ископаемые морские лишайны 600 миллионов лет были обнаружены. [ 129 ] Было выдвинуто предположение, что микопланктон развивался из наземных грибов, вероятно, в палеозойскую эпоху (390 миллионов лет назад). [ 130 ]
Композитное изображение, показывающее глобальное распределение фотосинтеза, включая как океанического фитопланктона , так и наземная растительность . Темно-красный и сине-зеленый указывает на области высокой фотосинтетической активности в океане и на земле, соответственно.
Основными производителями являются организмы AutoTroph , которые делают свою собственную пищу вместо того, чтобы употреблять другие организмы. Это означает, что основные производители становятся отправной точкой в пищевой цепи для гетеротрофных организмов, которые едят другие организмы. Некоторые морские первичные производители являются специализированными бактериями и архей, которые являются хемотрофами , делая собственную пищу, собираясь вокруг гидротермальных вентиляционных отверстий и прохладных просачиваний и используя хемосинтез . Однако большая часть морской первичной продукции поступает от организмов, которые используют фотосинтез на углекисленном диоксиде, растворенной в воде. Этот процесс использует энергию от солнечного света для преобразования воды и углекислого газа [ 133 ] : 186–187 в сахара, которые можно использовать как в качестве источника химической энергии, так и органических молекул, которые используются в структурных компонентах клеток. [ 133 ] : 1242 Морские первичные производители важны, потому что они лежат в основе практически всей жизни морских животных, генерируя большую часть кислорода и пищи, которые предоставляют другие организмы с химической энергией, которую им необходимо существовать.
Цианобактерии были первыми организмами, которые развивали способность превращать солнечный свет в химическую энергию. Они образуют тип (разделение) бактерий, которые варьируются от одноклеточного до нитевидного и включают колониальные виды . Они встречаются почти везде на Земле: в влажной почве, как в пресной, так и в морской среде, и даже на антарктических породах. [ 136 ] В частности, некоторые виды встречаются как дрейфующие клетки, плавающие в океане, и как таковые были одними из первых из фитопланктона .
Первыми основными производителями, которые использовали фотосинтез, были океанические цианобактерии около 2,3 миллиардов лет назад. [ 137 ] [ 138 ] Высвобождение молекулярного кислорода цианобактериями в качестве побочного продукта фотосинтеза вызвало глобальные изменения в окружающей среде Земли. Поскольку кислород был токсичным для большинства жизни на Земле в то время, это привело к почти взысканию организмов-непереносимости кислорода , драматическом изменении , которое перенаправляло эволюцию основных видов животных и растений. [ 139 ]
Крошечный (0,6 мкм ) морской цианобактерий Prochlorococcus , обнаруженный в 1986 году, сегодня является важной частью основания океанической пищевой цепи и объясняет большую часть фотосинтеза открытого океана [ 140 ] и, по оценкам, 20% кислорода в атмосфере Земли. [ 141 ] Возможно, это самый обильный род на Земле: один миллилитр поверхностной морской воды может содержать 100 000 клеток или более. [ 142 ]
Первоначально биологи считали, что цианобактерии были водоросли, и называли его «сине-зелеными водорослями». Более позднее мнение состоит в том, что цианобактерии являются бактериями, и, следовательно, даже не в том же королевстве, что и водоросли. Большинство властей исключают все прокариоты и, следовательно, цианобактерии из определения водорослей. [ 143 ] [ 144 ]
Динофлагелляты и диатомовые средства являются важными компонентами морских водорослей и имеют свои собственные участки ниже. Евгленфиты представляют собой тип одноклеточных жгутиков с несколькими морскими членами.
Не все водоросли микроскопичны. Зеленые, красные и коричневые водоросли имеют многоклеточные макроскопические формы, которые составляют знакомые морские водоросли . Зеленые водоросли , неформальная группа, содержит около 8000 признанных видов. [ 145 ] Многие виды живут большую часть своей жизни как отдельные клетки или являются нитевидными, в то время как другие образуют колонии, состоящие из длинных цепей клеток, или являются высоко дифференцированными макроскопическими морскими водорослями. Красные водоросли , а (оспариваемый) филом содержит около 7000 признанных видов, [ 146 ] В основном многоклеточные и включающие много известных морских водорослей. [ 146 ] [ 147 ] Коричневые водоросли образуют класс , содержащий около 2000 признанных видов, [ 148 ] В основном многоклеточные и включающие много морских водорослей, таких как водоросли .
В отличие от более высоких растений, водорослей не хватает корней, стеблей или листьев. Они могут быть классифицированы по размеру как микроводоросли или макроводоросли .
Микроводоросли - это микроскопические виды водорослей, не видимые невооруженным глазом. Это в основном одноклеточные виды, которые существуют в качестве индивидуумов или в цепях или группах, хотя некоторые из них являются многоклеточными . Микроводоросли являются важными компонентами морских протистов, обсуждаемых выше , а также фитопланктона, обсуждаемых ниже . Они очень разнообразны . Было подсчитано, что было описано 200 000-800 000 видов, из которых было описано около 50 000 видов. [ 149 ] В зависимости от вида их размеры варьируются от нескольких микрометров (мкМ) до нескольких сотен микрометров. Они специально адаптированы к окружающей среде, в которой преобладают вязкие силы.
Одноклеточные организмы обычно являются микроскопическими. Есть исключения. Вейнглас «Русалочка» , родом субтропических зеленых водорослей , представляет собой одноклеточные, но удивительно большие и сложные по форме с одним большим ядром, что делает его образцовым организмом для изучения клеточной биологии . [ 150 ] Другие одноклеточные водоросли, Caulerpa Taxifolia , имеют появление сосудистого растения, в том числе «листья», устроенные аккуратно вверх по стеблям, как папоротник. Селективное размножение в аквариумах для производства более твердых штаммов привело к случайному выбросу в Средиземноморье, где он стал инвазивным видом, известным в разговорной речи как убийственных водорослей . [ 151 ]
Chlamydomonas globosa , одноклеточная зеленая водоросли с двумя жгутиками , только что видимыми внизу слева
Макроводоросли являются более крупными, многоклеточными и более заметными типами водорослей, обычно называемых морских водорослей . Морские водоросли обычно растут в неглубоких прибрежных водах, где они привязаны к морскому дну . Как и микроводоросли, макроводоросли (морские водоросли) можно рассматривать как морские протисты, поскольку они не являются настоящими растениями. Но они не микроорганизмы, поэтому они не находятся в рамках этой статьи.
Планктон - это дрейфующие или плавающие организмы, которые не могут эффективно плавать против течения, и включают организмы из большинства областей жизни: бактерии , археи , водоросли , простейшие и животные . Красный, оранжевый, желтый и зеленый представляют участки, где расцветает водоросль. Голубые области представляют собой зоны с питательными веществами, где фитопланктон существует в более низких концентрациях.
Планктон (от греческого для странников ) представляют собой разнообразную группу организмов, которые живут в толще воды из больших водоемов, но не могут плавать против течения. В результате они бродят или дрейфуют с течениями. [ 153 ] Планктон определяется их экологической нишей , а не какой -либо филогенетической или таксономической классификацией. Они являются важным источником пищи для многих морских животных, от кормовой рыбы до китов . Планктон можно разделить на растение, похожий на растение, и компонент животных.
Phytoplankton - например, эта колония Chaetoceros socialis - естественно испускает красное флуоресцентное свет, который рассеивает избыточную солнечную энергию, которую они не могут употреблять с помощью фотосинтеза. Это свечение может быть обнаружена спутниками как показатель того, насколько эффективно океанский фитопланктон является фотосинтезированием. [ 154 ] [ 155 ]
Фитопланктон -это растения, подобные компонентам сообщества планктона («Фито» происходит от греческого для растения ). Они являются автотрофными (самосыпляющимися), что означает, что они генерируют свою собственную еду и не должны потреблять другие организмы.
Фитопланктон выполняет три важные функции: они генерируют почти половину мирового кислорода в атмосфере, они регулируют уровни углекислого газа в океане и атмосфере и образуют основание морской пищевой сети . Когда условия правы, цветы фитопланктонов водорослей могут происходить в поверхностных водах. Фитопланктон-это R-стратегии , которые быстро растут и могут удвоить свою популяцию каждый день. Цветы могут стать токсичными и истощать воду кислорода. Тем не менее, числа фитопланктона обычно контролируются фитопланктоном, исчерпавающим доступные питательные вещества и за счет выпаса зоопланктона. [ 156 ]
Диатомы образуют (оспариваемый) филим, содержащий около 100 000 распознанных видов в основном одноклеточных водорослей. Диатомовые средства генерируют около 20 процентов кислорода, производимого на планете каждый год, [ 93 ] брать более 6,7 миллиардов метрических тонн кремния каждый год из вод, в которых они живут, [ 159 ] и внести почти половину органического материала, найденного в океанах.
Диатомоты являются одним из наиболее распространенных типов фитопланктона
Их защитные раковины (достоинства) изготовлены из кремния
Диатомовые средства имеют раковину кремнезема ( надушка ) с радиальной (центрической) или двусторонней (трудовой) симметрией
Диатомы закрыты в оболочках защитного кремнезема (стекло), называемых ящиками . Каждая ямочка изготовлена из двух взаимосвязанных деталей, покрытых крошечными отверстиями, через которые диатома обменивается питательными веществами и отходами. [ 156 ] Престолы мертвых диатомов дрейфовали на дно океана, где в течение миллионов лет они могут нарастать до половины мили глубиной . [ 160 ]
Кокколитофоры - это мельчайшие одноклеточные фотосинтетические протисты с двумя жгутиками для локомоции. Большинство из них защищены оболочкой, покрытой богато украшенными круглыми пластинами или чешуйками, называемыми кокколитами . Кокколиты изготовлены из карбоната кальция. Оболочки кальцита важны для морского углеродного цикла. [ 163 ] Термин кокколитофор вытекает с греческого языка для семян, несущего камень , ссылаясь на их небольшой размер и камни кокколита, которые они несут. При правильных условиях они цветут, как и другие фитопланктоны, и могут повернуть молочный белый. [ 164 ]
Модель механизма генерирования энергии у морских бактерий
(1) Когда солнечный свет поражает молекулу родопсина (2) Он изменяет свою конфигурацию, поэтому протон исключен из ячейки (3) Химический потенциал приводит к тому, что протон вытекает обратно в клетку (4) таким образом генерируя энергию (5) в форме аденозин трихосфат . [ 165 ]
Фототрофический метаболизм основан на одном из трех энергетических пигментов: хлорофилл , бактериохлорофилл и сетчатка . Сетчатка является хромофором, обнаруженным в родопсинах . Значение хлорофилла при преобразовании световой энергии было написано на протяжении десятилетий, но фототрофия, основанная на пигментах сетчатки, только начинает изучаться. [ 166 ]
В 2000 году команда микробиологов во главе с Эдвардом Делонгом сделала решающее открытие в понимании морских углеродных и энергетических циклов. Они обнаружили ген у нескольких видов бактерий [ 168 ] [ 169 ] Отвечает за продукцию белкового родопсина , ранее неслыханный у бактерий. Эти белки, обнаруженные в клеточных мембранах, способны преобразовать энергию света в биохимическую энергию из -за изменения конфигурации молекулы родопсина, когда солнечный свет наносит ее, вызывая накапливание протона изнутри и последующее воспаление, которое генерирует энергию. [ 170 ] Архаальные родопсины впоследствии были обнаружены среди разных таксонов, протистов, а также у бактерий и археи, хотя они редки в сложных многоклеточных организмах . [ 171 ] [ 172 ] [ 173 ]
Исследования в 2019 году показывают, что эти «защехающие солнечные бактерии» более распространены, чем предполагалось ранее, и могут изменить то, как глобальное потепление влияет на океаны. «Результаты преодолевают традиционную интерпретацию морской экологии, обнаруженной в учебниках, в которой говорится, что почти весь солнечный свет в океане захватывается хлорофиллом в водорослях. Вместо этого, оснащенные родопсином бактерий, такие как гибридные автомобили, приведенные в действии органическим веществом. Большинство бактерий - и под солнечным светом, когда питательные вещества мало ». [ 174 ] [ 166 ]
В течение 1930 -х годов Альфред С. Редфилд обнаружил сходство между составом элементов в фитопланктоне и основными растворенными питательными веществами в глубоком океане. [ 177 ] Редфилд предположил, что отношение углерода к азоту к фосфору (106: 16: 1) в океане контролировалось требованиями фитопланктона, поскольку фитопланктон впоследствии высвобождает азот и фосфор, когда они повсеместны. Это соотношение стало известно как соотношение Редфилда и используется в качестве фундаментального принципа при описании стехиометрии эволюции морской воды и фитопланктона. [ 178 ]
Тем не менее, отношение Редфилда не является универсальным значением и может измениться с такими вещами, как географическая широта. [ 179 ] Основываясь на распределении ресурсов, Phytoplankton может быть классифицирован на три различных стратегии роста: выживший, Bloomer и генерал. Survivalist Phytoplankton имеет высокое соотношение N: P (> 30) и содержит множество механизма приобретения ресурсов для поддержания роста в условиях дефицитных ресурсов. Bloomer Phytoplankton имеет низкое соотношение N: P (<10), содержит высокую долю механизма роста и адаптирован к экспоненциальному росту. Общественный фитопланктон имеет аналогичное соотношение N: P к соотношению Redfield и содержит относительно равный механизм приобретения ресурсов и роста. [ 178 ]
Зопланктон - это компонент животных планктонного сообщества («зоопарк» происходит от греческого для животного ). Они гетеротрофны (в другом кормлении), что означает, что они не могут производить свою собственную пищу и должны потреблять вместо других растений или животных в качестве пищи. В частности, это означает, что они едят фитопланктон.
Радиоларии являются одноклеточными хищными протистами, заключенными в сложные шаровые раковины, обычно изготовленные из кремнезема и пронзили отверстия. Их имя происходит от латыни для "радиуса". Они ловят добычу, расширяя части своего тела через отверстия. Как и в случае с кремнеземами ятомов, радиолоальные раковины могут погрузиться в дно океана, когда радиолориалы умирают и становятся сохраняемыми как часть океанского отложения . Эти останки, как микрофоссии , предоставляют ценную информацию о прошлых океанических условиях. [ 185 ]
Как диатомовые, радиолориалы бывают разных форм
Также, как диатомовые ткатовые средства, радиояловые раковины обычно изготавливаются из силиката
Подобно радиолььериям, фораминифераны ( Forams для коротких) представляют собой одноклеточные хищные протисты, также защищенные оболочками, которые имеют отверстия в них. Их имя происходит от латыни для «носителей отверстий». Их раковины, часто называемые тестами , находятся в камере (Формам добавляют больше камер по мере роста). Оболочки обычно изготовлены из кальцита, но иногда изготавливаются из агглютинированных частиц отложений или хитона и (редко) кремнезема. Большинство Forams являются бентическими, но около 40 видов являются планктическими. [ 187 ] Они широко исследованы с хорошо известными ископаемыми записями, которые позволяют ученым многое сделать в прошлой среде и климате. [ 185 ]
Фораминиферанс
... может иметь более одного ядра
... и защитные шипы
Фораминифераны являются важными одноклеточными протистами зоопланктона , с тестами кальция
Миксотроф - это организм, который может использовать смесь различных источников энергии и углерода , вместо того, чтобы иметь единый трофический режим на континууме от полной автотрофии на одном конце до гетеротрофии в другом. Предполагается, что миксотрофы составляют более половины всего микроскопического планктона. [ 192 ] Существуют два типа эукариотических миксотрофов: те, которые имеют свои хлоропласты , и те, у кого есть эндосимбионты - и другие, которые приобретают их через клептопластику или порабощают всю фототрофическую клетку. [ 193 ]
Недавние исследования морского микроазурзопарина показали, что 30–45% численности киляграммы были миксотрофическими, а до 65% амебоидной, форум и радиоловой биомассы была миксотрофической. [ 87 ]
Phaeocystis является важным родом водорослей, обнаруженным как часть морского фитопланктона по всему миру. Он имеет полиморфный жизненный цикл, начиная от свободных клеток до больших колоний. [ 196 ] Он обладает способностью образовывать плавающие колонии, где сотни клеток встроены в гелевую матрицу, которая может массово увеличиваться в размерах во время цветов . [ 197 ] В результате, Phaeocystis вносит важное значение для морского углерода [ 198 ] и циклы серы . [ 199 ] Виды Phaeocystis представляют собой эндосимбионты для акантарских радиолариалов. [ 200 ] [ 201 ]
Миксотрофный планктон, который сочетает в себе фототрофию и гетеротрофию - таблица, основанная на Stoecker et al., 2017 [ 202 ]
Называется конститутивными миксотрофами Mitra et al., 2016. [ 203 ] Фитопланктон, которые едят: фотосинтетические протисты с унаследованными пластидами и способность принимать добычу.
Называется не совместимыми миксотрофами Mitra et al., 2016. [ 203 ] Зопланктон, которые являются фотосинтетическими: Microzooplankton или зоопланктон Metazoan, которые приобретают фототрофию через удержание хлоропластов а или поддержание эндосимбионтов водорослей.
Универсалы
Протисты, которые сохраняют хлоропласты и редко другие органеллы от многих таксонов водорослей
Большинство олиготрихских цилиат, которые сохраняют пластиды а
Специалисты
1. Протисты, которые сохраняют хлоропласты, а иногда и другие органеллы из одного вида водорослей или очень тесно связанных видов водорослей
а Хлоропласт (или пластид) удержание = секвестрация = порабощение. Некоторые виды, поддерживающие пластид, также сохраняют другие органеллы и добычу цитоплазму.
Динофлагелляты являются частью группы водорослей и образуют тип одноклеточных жгутиков с около 2000 морских видов. [ 204 ] Название происходит от греческого «диноми», означающих вихрь и латинский «жгутичок», означающий кнут или ресницы . Это относится к двум привязанным к кнуту (жгутичье), используемые для движения вперед. Большинство динофлагеллятов защищены красной коричневой, целлюлозной броней. Как и другие фитопланктоны, динофлагелляты являются R-стратегистами , которые в правильных условиях могут цветут и создавать красные приливы . Раскопки могут быть самой базальной жгутиковой линией. [ 102 ]
По трофической ориентации динофлагелляты не могут быть равномерно классифицированы. Известно, что некоторые динофлагелляты являются фотосинтетическими , но большая часть из них на самом деле сочетая миксотрофическая , сочетающая фотосинтез с приглашением добычи ( фаготрофия ). [ 205 ] Некоторые виды представляют собой эндосимбионты морских животных и других протистов и играют важную роль в биологии коралловых рифов . Другие предшествуют другим простейшим, и несколько форм являются паразитными. Многие динофлагелляты являются миксотрофными и также могут быть классифицированы как фитопланктон.
Токсичный динофлагеллятный динофиз Acuta приобретает хлоропласты от своей добычи. «Он не может уловить криптофиты сами по себе, и вместо этого опирается на употребление ресничек, таких как красно -мирионекта , которая секвеста своих хлоропластов из определенного криптофита клады (Geminigera/Plagioselmis/Teleaulax)». [ 202 ]
Гиродиния , один из немногих обнаженных динофлагеллятов, в которых отсутствует броня
Dinoflagellate Protoperidinium вытягивает большую кормление, чтобы захватить добычу
Населларийские радиольолярии могут быть в симбиозе с динофлагеллятами
Традиционно динофлагелляты были представлены как бронированные или безвольные
Динофлагелляты часто живут в симбиозе с другими организмами. Многие населларийские радиолярийские дома находятся своих в тестах. [ 207 ] Nassellarian обеспечивает аммоний и углекислый газ для динофлагеллята, в то время как динофлагеллят обеспечивает населларию слизистую мембрану, полезную для охоты и защиты от вредных захватчиков. [ 208 ] есть данные о Из анализа ДНК том, что динофлагеллятный симбиоз с радиояльцами развивался независимо от других динофлагелляционных симбиозов, таких как с фораминифера . [ 209 ]
Некоторые динофлагелляты биолюминесцентные . Ночью вода океана может освещаться внутренне и сверкать синим светом из -за этих динофлагеллятов. [ 210 ] [ 211 ] Биолюминесцентные динофлагелляты обладают сцинтиллонами , отдельными цитоплазматическими телами, которые содержат динофлагеллят люциферазу , основной фермент, участвующий в люминесценции. Люминесценция, иногда называемая фосфоресценцией моря , возникает в виде коротких (0,1 с) синих вспышек или искры, когда индивидуальные сцинтиллоны стимулируются, как правило, механическими нарушениями, например, лодки или пловца или серфинга. [ 212 ]
Каменный кинжал Отси Ледяной , который жил в медь -эпоху . Лезвие изготовлено из черта , содержащих радиолььеры, кальсферы, кальпионеллиды и несколько спикул губки. Присутствие кальпионеллидов , которые вымерли, использовалось для встречи с этим кинжалом. [ 216 ]
Отложения в нижней части океана имеют два основных происхождения, терригенные и биогенные.
Терригенные отложения составляют около 45% от общего морского осадка и возникают в эрозии пород на суше, транспортируемых реками и земельными стоками, ветроэнергетической пылью, вулканами или шлифованием ледниками.
Биогенные отложения учитывают остальные 55% от общего отложения и происходят в скелетных остатках морских протистов (одноклеточные микроорганизмы планктона и бентоса). Также могут присутствовать гораздо меньшие количества осажденных минералов и метеорической пыли. Происхождение , в контексте морского осадка, относится не к консистенции осадка, а к его биологическому происхождению. Термин «слизь» первоначально использовался Джоном Мюрреем , «отцом современной океанографии», который предложил термин радиояловый исток для отложений кремнезема радиоальных раковинов, приведенных на поверхность во время экспедиции Челленджер . [ 217 ] Биогенная слива - это пелагический осадок, содержащий не менее 30 процентов от скелетных остатков морских организмов.
Кокколитофоры являются крупнейшим глобальным источником биогенного карбоната кальция и значительно способствуют глобальному углеродному циклу. [ 221 ] Они являются основной составляющей меловых месторождений, таких как белые скалы Дувра .
Распределение типов отложений на морском дне В каждой цветной области показанный тип материала - это то, что доминирует, хотя, вероятно, присутствуют другие материалы. Для получения дополнительной информации см. Здесь
Морские микробентосы - это микроорганизмы, которые живут в бентической зоне океана - которые живут вблизи или на морском дне, или внутри или на поверхностных отложениях морского дна. Слово бентос происходит от греческого, что означает «глубина моря». Microbenthos встречаются повсюду на морском дне континентальных полков или вокруг, а также в более глубоких водах, с большим разнообразием в отложениях морского дна. В мелководье луга из морских трав , коралловые рифы и водоросли обеспечивают особенно богатые места обитания. В фотоамериканских зонах . В литоральных зонах изменение прилива сильно контролируют возможности для микробентоса.
Elphidium широко распространенный род бентических фурамов
Как фораминифера, так и диатомовые средства имеют планктонные и бентовые формы, то есть они могут дрейфовать в толще воды или жить на отложениях на дне океана. В любом случае, их снаряды оказываются на морском дне после смерти. Эти раковины широко используются в качестве климатических прокси . Химический состав оболочек является следствием химического состава океана в то время, когда были образованы раковины. Прошлые температуры воды также могут быть выведены из соотношений стабильных изотопов кислорода в раковинах, поскольку более легкие изотопы более легко испаряются в более теплой воде, оставляя более тяжелые изотопы в раковинах. Информация о прошлом климате может быть выведена дальше из изобилия фурматов и диатомовых ткатоев, поскольку они, как правило, более распространены в теплой воде. [ 223 ]
Внезапное событие вымирания , которое убило динозавров 66 миллионов лет назад, также вызвало вымершие три четверти всех других видов животных и растений. Однако глубоководные бентические фарамы процветали после последствий. В 2020 году сообщалось, что исследователи изучили химический состав тысяч образцов этих бентических фурамов и использовали свои результаты для создания наиболее подробной климатической записи Земли. [ 224 ] [ 225 ]
Некоторые эндолиты имеют чрезвычайно долгую жизнь. В 2013 году исследователи сообщили о доказательствах эндолитов в дне океана, возможно, миллионы лет, и время поколения составило 10 000 лет. [ 226 ] Они медленно метаболизируются, а не в состоянии покоя. Некоторым актиномицетота , обнаруженным в Сибири , по оценкам, полмиллиона лет. [ 227 ] [ 228 ] [ 229 ]
(А) Микробные взаимодействия варьируются от взаимовыгодного до вредного для одного или нескольких партнеров. Синие стрелки с двойными головами подчеркивают, что отношения могут двигаться между классификациями, на которые часто влияют условия окружающей среды. (B) Симбиозы в хосте-микробах следует учитывать в контексте микробных сообществ, где хозяин участвует в многочисленных и часто разных симбиотических отношениях. (C) Микробные сообщества подвержены влиянию различных симбиозов микроботового микроба, начиная от сотрудничества (например, синтрофия или совместного метаболизма) до конкуренции. Стрелки изображают в целом полезные (синие) и вредные (красные) результаты для одного (одиночная стрелка) или обоих (двойная стрелка). Примечание, как и в случае с симбиозами-хозяином-микробом, эти отношения можно рассматривать как жидкость и под влиянием условий окружающей среды. [ 230 ]
Концепция Holobiont была первоначально определена доктором Линн Маргулисом в ее книге 1991 года как источник эволюционного инновации как собрания хозяина и многих других видов, живущих в нем или вокруг него, которые вместе образуют отдельную экологическую единицу . [ 231 ] Компонентами голобиона являются отдельные виды или бионты , в то время как комбинированный геном всех бионтов является гологеномом . [ 232 ]
Концепция впоследствии развивалась с момента этого оригинального определения, [ 233 ] с фокусом, перемещающимся к микробным видам, связанным с хозяином. Таким образом, Holobiont включает в себя хозяин, вирус , микробиом и другие участники, каждый из которых каким -то образом способствует функции целого. [ 234 ] [ 235 ] Холобионт обычно включает в себя эукариота хозяин и все симбиотические вирусы , бактерии , грибы и т. Д., Которые живут на или внутри него. [ 236 ]
Тем не менее, существует противоречия по поводу того, можно ли рассматривать Holobionts как отдельные эволюционные единицы. [ 237 ]
Путь вирусного шунтирования представляет собой механизм, который предотвращает микробных частиц миграцию органического вещества (POM) морских (POM) путем переработки их в растворенное органическое вещество (DOM), которое может быть легко занято микроорганизмами. [ 244 ] Вирусная шунтирование помогает поддерживать разнообразие в микробной экосистеме, предотвращая доминирование в микро-среде. [ 245 ] DOM, переработанный вирусным шунтированным путем, сопоставим с количеством, генерируемым другими основными источниками морского DOM. [ 246 ]
Морские микробные сообщества (SIMCO) относятся к группам микроорганизмов, живущих внутри и в интерфейсах морского льда на полюсах. Мягкая матрица, в которой они обитают, имеет сильные вертикальные градиенты солености, света, температуры и питательных веществ. На химию морского льда больше всего влияет соленость рассола, которая влияет на рН и концентрацию растворенных питательных веществ и газов. Рассох , образованный во время таящего морского льда, создает поры и каналы в морском льду, в которых могут жить эти микробы. В результате этих градиентов и динамических условий в нижнем слое льда обнаружено более высокое содержание микробов, хотя некоторые из них находятся в среднем и верхнем слоях. [ 251 ]
Гидротермальные вентиляционные отверстия расположены там, где тектонические пластины перемещаются и распространяются. Это позволяет воде из океана войти в кору Земли, где она нагревается магмой. Увеличивающее давление и температура вынуждают воду из этих отверстий, на выходе вода накапливает растворенные минералы и химические вещества из пород, с которыми она сталкивается. Вентиляционные отверстия можно охарактеризовать температурой и химическим составом как диффузные вентиляционные отверстия , которые высвобождают прозрачную относительно прохладную воду, обычно ниже 30 ° C, как белые курильщики , которые излучают молочную воду при более теплых температурах, около 200-330 ° C, и как черные курильщики , которые испускают воду затемнены накапливаемыми осадками сульфида при горячих температурах, около 300-400 ° C. [ 252 ]
Микробные сообщества гидротермических вентиляционных средств представляют собой микроскопические одноклеточные организмы, которые живут и размножаются в химически отдельной области вокруг гидротермальных вентиляционных отверстий. К ним относятся организмы в микробных матах , свободные плавающие клетки и бактерии в эндосимбиотических отношениях с животными. Поскольку на этих глубинах нет солнечного света, хемосинтез обеспечивается энергией , где симбиотические бактерии и археи образуют дно пищевой цепи и способны поддерживать различные организмы, такие как гигантские трубные черви и черви Pompeii . Эти организмы используют эту симбиотическую связь, чтобы использовать и получить химическую энергию, которая выделяется в этих областях гидротермальных вентиляционных средств. [ 253 ] Хемолитоавтотрофные бактерии могут получать питательные вещества и энергию от геологической активности при гидротермальном вентиляционном отверстии, чтобы исправить углерод в органические формы. [ 254 ]
Вирусы также являются частью гидротермального вентиляционного микробного сообщества, и их влияние на микробную экологию в этих экосистемах является растущей областью исследований. [ 255 ] Вирусы - самая распространенная жизнь в океане, содержит величайший резервуар генетического разнообразия. [ 256 ] Поскольку их инфекции часто являются смертельными, они представляют собой значительный источник смертности и, следовательно, оказывают широкое влияние на биологические океанографические процессы, эволюцию и биогеохимический велосипед в океане. [ 257 ] Однако были обнаружены доказательства того, что вирусы, обнаруженные в вентиляционных местах обитания, приняли более взаимную, чем паразитическую эволюционную стратегию, чтобы пережить экстремальную и нестабильную среду, в которой они существуют. [ 258 ] Было обнаружено, что глубоководные гидротермальные вентиляционные отверстия имеют большое количество вирусов, указывающих на высокую вирусную продукцию. [ 259 ] Как и в других морских средах, глубоководные гидротермальные вирусы влияют на изобилие и разнообразие прокариот и, следовательно, влияют на микробейный биогеохимический велосипед, лизируя своих хозяев. [ 260 ] Тем не менее, в отличие от их роли в качестве источника смертности и контроля населения, были также постулированы вирусы для повышения выживаемости прокариот в экстремальных средах, действующих в качестве резервуаров генетической информации. Поэтому считается, что взаимодействие виросферы с микроорганизмами при стрессах окружающей среды помогают выживанию микроорганизмов посредством рассеивания генов хозяина посредством горизонтального переноса генов . [ 261 ]
Флуоресцентная микроскопия различных пикопланктона в океане, большая часть которой не может быть эффективно изучена, потому что они сопротивляются попыткам лабораторной культуры
- Глубокая биосфера та часть биосферы , которая находится ниже первых нескольких метров поверхности. Он простирается как минимум на 5 километров ниже континентальной поверхности и 10,5 километров ниже поверхности моря, температура, которые могут превышать 100 ° C.
Над поверхностными живыми организмами потребляют органическое вещество и кислород. Ниже, они недоступны, поэтому они используют «пищевые» ( доноры электронов ), такие как водород, высвобождаемый из пород, различными химическими процессами, метаном, уменьшенными соединениями серы и аммония. Они «дышат» электронами, такие как нитраты и нитриты, оксиды марганца и железа, окисленные соединения серы и диоксид углерода.
На большей глубине очень мало энергии, и метаболизм может быть в миллион раз медленнее, чем на поверхности. Клетки могут жить в течение тысячелетий, прежде чем разделиться, и нет известных пределов их возраста. На подповерхностном приходится около 90% биомассы у бактерий и археи, а 15% от общей биомассы для биосферы. Эукариоты также обнаружены, в основном микроскопических, но включают в себя некоторую многоклеточную жизнь. Вирусы также присутствуют и заражают микробы.
Подповерхностные жизненные среды
В 2018 году исследователи из глубокой углеродной обсерватории объявили, что формы жизни , в том числе 70% бактерий и археи на Земле, общая биомасса в 23 миллиарда тонн углерода , живая до 4,8 км (3,0 миль) в глубине под землей, в том числе 2,5 км ( 1,6 мили) ниже морского дна. [ 262 ] [ 263 ] [ 264 ] В 2019 году были обнаружены микробные организмы, живущие на 7 900 футов (2400 м) ниже поверхности, дыхание серы и употребление в скалах, таких как пирит , как их обычный источник пищи. [ 265 ] [ 266 ] [ 267 ] Это открытие произошло в самой старой известной воде на земле. [ 268 ]
Эти аэробные микроорганизмы, обнаруженные глубоко в органически плохих отложениях, находятся в квазиспендированной анимации в течение 100 миллионов лет
Неопознанный сферический микроорганизм водорослей, часть микробной темной материи
На сегодняшний день биологи не смогли культивировать в лаборатории подавляющее большинство микроорганизмов. Это особенно относится к бактериям и археи и связано с отсутствием знаний или способности обеспечивать необходимые условия роста. [ 271 ] [ 272 ] Термин «Микробная темная материя» стал использовать для описания микроорганизмов, которые ученые знают, есть ли существуют, но не смогли культивировать, и чьи свойства остаются неуловимыми. [ 271 ] Микробная темная материя не связана с темной веществом физики и космологии, но так называется сложностью в эффективном изучении ее. Трудно оценить его относительную величину, но принятая грубая оценка заключается в том, что менее одного процента микробных видов в данной экологической нише культурируют. В последние годы усилия предпринимаются для расшифровки большего количества микробной темной материи путем изучения генома последовательности ДНК из образцов окружающей среды [ 273 ] а затем, получая информацию об их метаболизме от их секвенированного генома, способствуя знаниям, необходимым для их выращивания.
Оценки микробных видов считаются в трех доменах жизни
Бактерии - самая старая и самая биоративная группа, за которой следуют археа и грибы (самые последние группы). В 1998 году, до того, как началась осознание степени микробной жизни, Роберт М. Мэй [ 274 ] По оценкам, на планете было 3 миллиона видов живых организмов. Но в 2016 году Лоси и Леннон [ 275 ] По оценкам, количество видов микроорганизма может достигать 1 триллиона. [ 276 ]
Микробное разнообразие
Сравнительное представление известного и предполагаемого (маленького ящика) и еще неизвестного (большой коробки) микробного разнообразия, которое относится как к морским, так и к наземным микроорганизмам. Текстовые поле относятся к факторам, которые отрицательно влияют на знание микробного разнообразия, которое существует на планете. [ 276 ]
Стратегии для отбора проб планктона по классам и численности Синий фон указывает отфильтрованный объем, необходимый для получения достаточных чисел организма для анализа. Фактические объемы, из которых отображаются организмы, всегда регистрируются. [ 277 ]
Традиционно была выведена филогения микроорганизмов, и их таксономия была установлена на основе исследований морфологии . Тем не менее, разработки в молекулярной филогенетике позволили установить эволюционную связь видов путем анализа более глубоких характеристик, таких как их последовательности ДНК и белков , например, рибосомальная ДНК . [ 278 ] Отсутствие легкодоступных морфологических особенностей, таких как те, которые присутствуют у животных и растений , особенно затрудненные ранние усилия по классификации бактерий и археи. Это привело к ошибочной, искаженной и запутанной классификации, примером которой, отмеченный Карл Возе , является Pseudomonas , этимология которых по иронии судьбы соответствовала его таксономии, а именно «ложная единица». [ 279 ] Многие бактериальные таксоны были реклассифицированы или пересмотрены с использованием молекулярной филогенетики.
Мариномонас Арктика , бактерия, которая растет внутри арктического морского ледя
Было бы трудно последовательно отделить эти два микроба, используя только изображения. Однако, если их штрих -коды выровнены друг с другом, и их основания окрашены, чтобы увидеть их более четко, становится легко видеть, какие основания различаются между этими двумя микробами. Таким образом, миллионы различных видов микробов можно различить. [ 280 ]
Три различные возможности для обработки выборки - это необработанные образцы, фиксацию или сохранение и фильтрацию. Для микроскопии и проточной цитометрии сырые образцы либо измеряются немедленно, либо должны быть зафиксированы для последующих измерений. Поскольку молекулярные методы, пигментный анализ и обнаружение молекулярных трассеров обычно требуют концентрированных клеток, остатки фильтров служат для измерений фитопланктона. Молекулярная характеристика и количественная оценка молекул трассировки выполняются с использованием хроматографии , масс -спектрометрии и ядерной магнитно -резонансной спектроскопии. [ 281 ]
Recent developments in molecular sequencing have allowed for the recovery of genomesin situ, directly from environmental samples and avoiding the need for culturing. This has led for example, to a rapid expansion in knowledge of the diversity of bacterial phyla. These techniques are genome-resolved metagenomics and single-cell genomics.
The new sequencing technologies and the accumulation of sequence data have resulted in a paradigm shift, highlighted both the ubiquity of microbial communities in association within higher organisms and the critical roles of microbes in ecosystem health.[283] These new possibilities have revolutionized microbial ecology, because the analysis of genomes and metagenomes in a high-throughput manner provides efficient methods for addressing the functional potential of individual microorganisms as well as of whole communities in their natural habitats.[284][285][286]
DNA sequencing technologies used in marine metagenomics
The discovery process involves marine sampling, DNA sequencing and contig generation. Previously unknown genes, pathways and even whole genomes are being discovered. These genome-editing technologies are used to retrieve and modify valuable microorganisms for production, particularly in marine metagenomics. Organisms may be cultivable or uncultivable. Metagenomics is providing especially valuable information for uncultivable samples.[287]
Omics is a term used informally to refer to branches of biology whose names end in the suffix -omics, such as genomics, proteomics, metabolomics, and glycomics. Marine Omics has recently emerged as a field of research of its own.[288] Omics aims at collectively characterising and quantifying pools of biological molecules that translate into the structure, function, and dynamics of an organism or organisms. For example, functional genomics aims at identifying the functions of as many genes as possible of a given organism. It combines different
-omics techniques such as transcriptomics and proteomics with saturated mutant collections.[289][290]
Many omes beyond the original genome have become useful and have been widely adopted in recent years by research scientists. The suffix -omics can provide an easy shorthand to encapsulate a field; for example, an interactomics study is reasonably recognisable as relating to large-scale analyses of gene-gene, protein-protein, or protein-ligand interactions, while proteomics has become established as a term for studying proteins on a large scale.
Any given omics technique, used just by itself, cannot adequately disentangle the intricacies of a host microbiome. Multi-omics approaches are needed to satisfactorily unravel the complexities of the host-microbiome interactions.[291] For instance, metagenomics, metatranscriptomics, metaproteomics and metabolomics methods are all used to provide information on the metagenome.[292]
Meta-omics data based biogeochemical modeling [293]
A schematic conceptual framework for marine biogeochemical modeling from environmental, imaging, and meta-omics data.[294] A semi-automatic computational pipeline is schematized for combining biomarkers with biogeochemical data [295] that can be incorporated into classic biogeochemical models [296] for creating a next generation of biogeochemical trait-based meta-omics models by considering their respective traits. Such novel meta-omics-enabled approaches aim to improve the monitoring and prediction of ocean processes while respecting the complexity of the planktonic system.[297][298]
As an example of how omics data can be used with marine phytoplankton to inform Earth system science, metatranscriptome sequences from natural phytoplankton communities were used to help identify physiological traits (cellular concentration of ribosomes and their rRNAs) underpinning adaptation to environmental conditions (temperature). A mechanistic phytoplankton cell model was used to test the significance of the identified physiological trait for cellular stoichiometry. Environmental selection in a trait‐based global marine ecosystem model was then linking emergent growth and cellular allocation strategies to large‐scale patterns in light, nutrients and temperature in the surface marine environment. Global predictions of cellular resource allocation and stoichiometry (N:P ratio) were consistent with patterns in metatranscriptome data[300] and latitudinal patterns in the elemental ratios of marine plankton and organic matter.[301] The three‐dimensional view of ribosome shows rRNA in dark blue and dark red. Lighter colours represent ribosomal proteins. Bands above show temperature‐dependent abundance of the eukaryotic ribosomal protein S14.[300]
Microorganisms and climate change in marine and terrestrial biomes[6]
In marine environments, microbial primary production contributes substantially to CO2 sequestration. Marine microorganisms also recycle nutrients for use in the marine food web and in the process release CO2 to the atmosphere. Microbial biomass and other organic matter (remnants of plants and animals) are converted to fossil fuels over millions of years. By contrast, burning of fossil fuels liberates greenhouse gases in a small fraction of that time. As a result, the carbon cycle is out of balance, and atmospheric CO2 levels will continue to rise as long as fossil fuels continue to be burnt.[6]
Microorganisms have key roles in carbon and nutrient cycling, animal (including human) and plant health, agriculture and the global food web. Microorganisms live in all environments on Earth that are occupied by macroscopic organisms, and they are the sole life forms in other environments, such as the deep subsurface and ‘extreme’ environments. Microorganisms date back to the origin of life on Earth at least 3.8 billion years ago, and they will likely exist well beyond any future extinction events... Unless we appreciate the importance of microbial processes, we fundamentally limit our understanding of Earth's biosphere and response to climate change and thus jeopardize efforts to create an environmentally sustainable future.[6]
Marine microorganisms known as cyanobacteria first emerged in the oceans during the Precambrian era roughly 2 billion years ago. Over eons, the photosynthesis of marine microorganisms generated by oxygen has helped shape the chemical environment in the evolution of plants, animals and many other life forms. Marine microorganisms were first observed in 1675 by Dutch lensmaker Antonie van Leeuwenhoek.
^ Jump up to: abRybicki EP (1990). "The classification of organisms at the edge of life, or problems with virus systematics". South African Journal of Science. 86: 182–6. ISSN0038-2353.
^Bolhuis, H. and Cretoiu, M.S. (2016) "What is so special about marine microorganisms?". In: L. J. Stal and M. S. Cretoiu (Eds.) The Marine Microbiome, pages 3–20, Springer. ISBN9783319330006
^ Jump up to: abCanchaya C, Fournous G, Chibani-Chennoufi S, Dillmann ML, Brüssow H (2003). "Phage as agents of lateral gene transfer". Current Opinion in Microbiology. 6 (4): 417–24. doi:10.1016/S1369-5274(03)00086-9. PMID12941415.
^ Jump up to: abRybicki EP (1990). "The classification of organisms at the edge of life, or problems with virus systematics". South African Journal of Science. 86: 182–186.
^ Ямагучи, Масаши; и др. (1 декабря 2012 г.). «Прокариот или эукариот? Уникальный микроорганизм из глубокого моря». Журнал электронной микроскопии . 61 (6): 423–431. doi : 10.1093/jmicro/dfs062 . PMID 23024290 .
^ Гудэй, AJ; Аранда да Силва, а.; Pawlowski, J. (1 декабря 2011 г.). «Ксенофиофоры (Rhizaria, Foraminifera) из Каньона Назаре (португальская маржа, NE Atlantic)». Глубоководные исследования, часть II: Актуальные исследования в области океанографии . Геология, геохимия и биология подводных каньонов к западу от Португалии. 58 (23–24): 2401–2419. Bibcode : 2011dsrii..58.2401g . doi : 10.1016/j.dsr2.2011.04.005 .
^ Синглтон, Пол (2006). Словарь микробиологии и молекулярной биологии, 3 -е издание, пересмотренное . Чичестер, Великобритания: Джон Вили и сыновья. п. 32. ISBN 978-0-470-03545-0 .
^ Гутьеррес, Марсело Х; Pantoja, Silvio; Хиноны, Ренато А и Гонсалес, Родриго Р. Первая запись о мягких грибах в прибрежной экосистеме у центра Чили. Гаяна (Консеп.) [Онлайн]. 2010, Vol.74, N.1, pp. 66-73. ISSN 0717-6538 .
^ Jump up to: а беременный Шридхар, К.Р. (2009). «10. Водные грибы - они планктоничны?». Динамика Планктона индийских вод . Джайпур, Индия: Pratiksha Publications. С. 133–148.
^ "Лишайники" . Служба национальных парков, Министерство внутренних дел США, правительство Соединенных Штатов. 22 мая 2016 года . Получено 4 апреля 2018 года .
^ Jump up to: а беременный Кэмпбелл, Нил А.; Рис, Джейн Б.; Урри, Лиза Андреа; Каин, Майкл Л.; Вассерман, Стивен Александр; Минорский, Питер V.; Джексон, Роберт Брэдли (2008). Биология (8 -е изд.). Сан -Франциско: Пирсон - Бенджамин Каммингс. ISBN 978-0-321-54325-7 .
^ Старккс, Сенн (31 октября 2012 г.). «Место на солнце» . Фландрия сегодня . Архивировано с оригинала 4 марта 2016 года . Получено 8 декабря 2012 года . Водоросли - это урожай будущего, по словам исследователей в Гиле
^ Мандоли Д.Ф. (1998). «Разработка плана тела и изменения фазы во время развития вертебуляции: как создается сложная архитектура гигантского Unicell?». Ежегодный обзор физиологии растений и молекулярной биологии растений . 49 : 173–198. doi : 10.1146/annurev.arplant.49.1.173 . PMID 15012232 . S2CID 6241264 .
^ Rost, B. and Riebesell, U. (2004) «Кокколитофоры и биологический насос: ответы на изменения окружающей среды». В кн.: Кокколитофоры: от молекулярных процессов до глобального воздействия , страницы 99–125, Springer. ISBN 9783662062784 .
^ Wassilieff, Maggy (2006) «Кокколитофор» , Te Ara - Энциклопедия Новой Зеландии . Доступ: 2 ноября 2019 года.
^ Sparks, William B.; Dassarma, S.; Рейд, в (декабрь 2006 г.). «Эволюционная конкуренция между примитивными фотосинтетическими системами: существование ранней фиолетовой земли?». Американское астрономическое общество встречается тезисами . 38 : 901. Bibcode : 2006aas ... 209.0605s .
^ Редфилд, Альфред С. (1934). «О пропорциях органических производных в морской воде и их отношении к составу планктона». В Джонстоне, Джеймс; Даниэль, Ричард Джеллико (ред.). Мемориал Джеймса Джонстона . Ливерпуль: Университетская издательство Ливерпуля. С. 176–92. OCLC 13993674 .
^ Finlay, BJ (2013). «Микроаэрофилия» . В Маршалле, KC (ред.). Достижения в области микробной экологии . Тол. 11. Springer Science & Business Media. п. 22. ISBN 978-1-4684-7612-5 .
^ Schoemann, Véronique; Becquevort, Sylvie; Стефельс, Жаклин; Руссо, Вероник; Ланселот, Кристиан (1 января 2005 г.). «Phaeocystis цветет в глобальном океане и их контролирующие механизмы: обзор». Журнал морских исследований . Железные ресурсы и океанические питательные вещества - продвижение глобального экологического моделирования. 53 (1–2): 43–66. Bibcode : 2005jsr .... 53 ... 43 с . Citeseerx 10.1.1.319.9563 . doi : 10.1016/j.seares.2004.01.008 .
^ Buskey, EJ (1995). «Рост и биолюминесценция Noctiluca Scintillans на различных диете водорослей». Журнал Plankton Research . 17 (1): 29–40. doi : 10.1093/plankt/17.1.29 .
^ Грет Р. Хасл; Эрик Э. Сивертсен; Карен А. Стейдингер; Карл Танген (25 января 1996 г.). «Морские диатомовые» . В Кармело Р. Томас (ред.). Идентификация морских диатомовых и динофлагеллятов . Академическая пресса. С. 5–385. ISBN 978-0-08-053441-1 Полем Получено 13 ноября 2013 года .
^ Вестер, Томас; Марван, Норберт; Друри, Анна Джой; Либранд, Дидерик; Агнини, Клаудия; Anannostou, Eleni; Барнет, Джеймс С.К.; Бохати, Стивен М.; Де Влешвер, Дэвид; Флоридо, Фабио; Фредерихс, Томас; Ходелл, Дэвид А.; Holbourn, Ann E.; Крун, Дик; Лауретано, Виттория; Литтлер, Кейт; Lourens, Lucas J.; Лайл, Митчелл; Пяковы, Хейко; Röhl, Урсула; живот, июнь; Уилкенс, Рой Х.; Уилсон, Пол А.; Зачос, Джеймс С. (11 сентября 2020 г.). «Астрономическая учет климата Земли и его предсказуемости за последние 66 миллионов лет » Наука 369 (6509): 1383–1 Bibcode : 2020sci ... 369.1 Doi : 10.1126/ science.ab6 HDL : 11577/3 PMID 32913105 S2CID 221593388
^ This, KR, Dheilly, NM, Klassen, JL, Brucker, RM, Baines, JF, Bosch, TC, Cryan, JF, Gilbert, SF, Goodnight, CJ, Lloyd, EA and Sapp, J. (2016) " Концепция Hologenome Right: экоэволюционная структура для хозяев и их микробиомов ». Msystems , 1 (2). doi : 10.1128/msystems.00028-16 . Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
^ Bulgarelli, D., Schlaeppi, K., Spaepen, S., van Themaat, EVL и Schulze-Lefert, P. (2013) «Структура и функции бактериальной микробиоты растений». Ежегодный обзор биологии растений , 64 : 807–838. doi : 10.1146/annurev-arplant-050312-120106 .
^ Sánchez-Cañizares, C., Jorrín, B., Poole, PS и Tkacz, A. (2017) «Понимание голобионта: взаимозависимость растений и их микробиом». Текущее мнение в микробиологии , 38 : 188–196. Два : 10.1016/j.mib.2017.07.001 .
^ Douglas, AE и Werren, JH (2016) «Отверстия в голокоме: почему симбиозы хозяина-микроба не являются голобионтами». Mbio , 7 (2). Doi : 10.1128/race.02099-15 .
^ Knowlton, N. and Rohwer, F. (2003) «Многоспективные микробные взаимные взаимности на коралловых рифах: хозяин как среда обитания». Американский натуралист , 162 (S4): S51-S62. doi : 10.1086/378684 .
^ Thompson, JR, Rivera, HE, Closek, CJ и Medina, M. (2015) «Микробы в коралловом холобионте: партнеры посредством эволюции, развития и экологических взаимодействий». Границы в клеточной и инфекционной микробиологии , 4 : 176. Два : 10.3389/fcimb.2014.00176 .
^ Cavalcanti, GS, Shukla, P., Morris, M., Ribeiro, B., Foley, M., Doane, MP, Thompson, CC, Edwards, MS, Dinsdale, EA и Thompson, FL (2018). В изменяющемся океане: взаимодействия с хозяином-микробами опосредуют устойчивость кораллических водорослей при подкислении океана ». BMC Genomics , 19 (1): 1–13. Два : 10.1186/S12864-018-5064-4 . Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
^ Робинсон, Кэрол и Нагаппа Рамая. «Микробные гетеротрофные скорости метаболизма ограничивают микробного углеродного насоса». Американская ассоциация по развитию науки, 2011.
^ Саймон, М., Гроссарт Х., Швейцер Б. и Плуг, Х. (2002) «Микробная экология органических агрегатов в водных экосистемах». Водная микробная экология , 28 : 175–211. два : 10.3354/ame028175 .
^ Boetius, Antje; Anesio, Александр М.; Деминг, Джоди В.; Микаки, Джилл А.; Рапп, Жозефина З. (1 ноября 2015 г.). «Микробная экология криосферы: морской лед и ледниковые места обитания». Nature Reviews Microbiology . 13 (11): 677–690. doi : 10.1038/nrmicro3522 . PMID 26344407 . S2CID 20798336 .
^ Кадар, Эник; Коста, Валентина; Сантос, Рикардо С.; Пауэлл, Джонатан Дж. (Июль 2006 г.). «Разделение тканей микроэффективных металлов в вентиляционных двустворчатых велосипедах батимодиола азорикуса и связанных с ними организмов (бактерии эндосимбионта и полихет паразита) от геохимически отличных вентиляционных вентиляционных технологий среднего атлантического хребта». Журнал морских исследований . 56 (1): 45–52. Bibcode : 2006jsr .... 56 ... 45K . doi : 10.1016/j.seares.2006.01.002 . ISSN 1385-1101 .
^ Ортманн, Алиса С.; Саттл, Кертис А. (август 2005 г.). «Высокое содержание вирусов в глубоководной гидротермальной вентиляционной системе указывает на вирусную микробную смертность». Deep Sea Research Part I: океанографические исследовательские работы . 52 (8): 1515–1527. Bibcode : 2005dsri ... 52.1515o . doi : 10.1016/j.dsr.2005.04.002 . ISSN 0967-0637 .
^ Lollar, Garnet S.; Уор, Оливер; Рассказ, Джон; Осберн, Магдалена Р.; Шервуд Лоллар, Барбара (2019). « Следуйте за водой»: гидрогеохимические ограничения на микробных исследованиях на 2,4 км ниже поверхности в глубокой жидкости и глубокой жизни на обсерватории и глубокой жизни ». Журнал геомикробиологии . 36 (10): 859–872. Bibcode : 2019gmbj ... 36..859L . doi : 10.1080/01490451.2019.1641770 . S2CID 199636268 .
^ Lozupone, CA, Stombaugh, Ji, Gordon, Ji, Jansson, JK and Knight, R. (2012) «Разнообразие, стабильность и устойчивость человеческой кишечной микробиоты». Nature , 489 (7415): 220–230. doi : 10.1038/nature11550 .
^ Venter, JC, Remington, K., Heidelberg, JF, Halpern, AL, Rusch, D., Eisen, JA, Wu, D., Paulsen, I., Nelson, KE, Nelson, W. и Fouts, de (DE (DE 2004) «Секвенирование ружья экологического генома моря Саргассо». Science , 304 (5667): 66–74. doi : 10.1126/science.1093857 .
^ Liu, L., Li, Y., Li, S., Hu, N., He, Y., Pong, R., Lin, D., Lu, L. and Law, M. (2012) "Сравнение систем секвенирования следующего поколения ». Biomed Research International , 2012 : 251364. doi : 10.1155/2012/251364 .
^ Медведь, Габриэль ; Дория Рыбоксова, Дорин Фиан, Томислав Чернава, Мари-Чристин Чампумер Верс, Тревор Чарльз, Сяоюлон, Кей Коко, Кзоу, Линдо Казу, Линдо Казу, Линия Ланге, Линия Ланге, Нельсон Лайма, Эмманлле Магюн, Тим Маучлайн, Рян Макайр, Рейн Мака. , Биргит Миттер, Мэтью Райан, Инга Серанд, Хауке Смидт, Хьюго Рум, Г. Сегал Киран, Джозеф Селвиан, Эйр, Эйр, Лиуза, Портуач Лео из Овербеля, Борбор, Борбор Борбор, Брайех К. Сингх К. Сингх К. Сингх. Микробийны , 8 (103): 1–22. Два : 10.1186/S40168-020-00875-0 . Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
^ Zhang, W., Li, F. and Nie, L. (2010) «Интеграция множественного анализа« OMIC »для микробной биологии: применение и методологии». Микробиология , 156 (2): 287–301. два : 10.1099/small.0.034793-0 .
^ Capotondi, A., Jacox, M., Bowler, C., Kavanaugh, M., Lehodey, P., Barrie, D., Brodie, S., Chaffron, S., Cheng, W., Dias, DF, EVEILLARD, D. и 20 других (2019) «Наблюдательные потребности в поддержке моделирования и прогнозирования морских экосистем: от глобального океана до региональных и прибрежных систем». Границы в морской науке , 6 : 623. Два : 10.3389/fmars.2019.00623 . Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
^ Sunagawa, S., Coelho, LP, Chaffron, S., Kultima, JR, Labadie, K., Salazar, G., et al. (2015). «Ocean Plankton. Структура и функция глобального океанского микробиома». Наука , 348 : 1261359. doi : 10.1126/science.1261359 .
^ Guidi, L., Chaffron, S., Bittner, L., Eveillard, D., Larhlimi, A., Roux, S., et al. (2016). «Планктонные сети, управляющие экспортом углерода в олиготрофном океане». Природа , 532 : 465–470. doi : 10.1038/nature16942 .
^ Guyennon, A., Baklouti, M., Diaz, F., Palmieri, J., Beuvier, J., Lebaupin-Brossier, C., et al. (2015). «Новое понимание экспорта органического углерода в Средиземном море из трехмерного моделирования». Biogeosciences , 11 : 425–442. Два : 10.5194/BG-12-7025-2015 .
^ Лука С., Парфри, Л.В. и Доубели, М. (2016). «Функция развязки и таксономия в глобальном океанском микробиоме». Наука , 353 : 1272–1277. doi : 10.1126/science.aaf4507 .
^ Coles, VJ, Stukel, MR, Brooks, MT, Burd, A., Crump, BC, Moran, MA, et al. (2017). «Океанская биогеохимия смоделирована с геномикой на основе признаков». Наука , 358 : 1149–1154. doi : 10.1126/science.aan5712 .
^ Mock, T., Daines, SJ, Geider, R., Collins, S., Metodiev, M., Millar, AJ, Moulton, V. and Lenton, TM (2016). » Чтобы лучше понять, как изменения окружающей среды влияют на морские микробы ». Глобальная биология изменений , 22 (1): 61–75. Doi : 10.1111 / gcb.12983 . Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
^ Jump up to: а беременный Toseland A, Daines SJ и Clark Jr et al. (2013) «Влияние температуры на распределение ресурсов и метаболизма морского фитопланктона». Изменение климата природы , 3 : 979–984. два : 10.1038/климат1989 .
^ Martiny AC, Pham CTA, Primeau FW, Vrugt JA, Moore JK, Levin SA и Lomas MW (2013) «Сильные широтные закономерности в элементарных соотношениях морского планктона и органического вещества». Nature Geoscience , 6 : 279–283. два : 10.1038/ngeo1757 .
Arc.Ask3.Ru Номер скриншота №: 7a6ca26b6a9dead4008f14eb96dd4eed__1717309860 URL1:https://arc.ask3.ru/arc/aa/7a/ed/7a6ca26b6a9dead4008f14eb96dd4eed.html Заголовок, (Title) документа по адресу, URL1: Marine microorganisms - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)
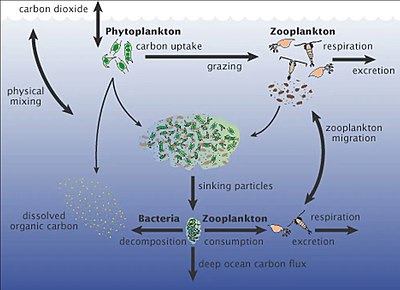



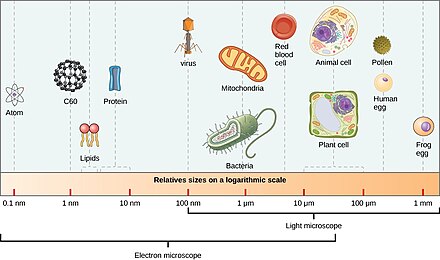




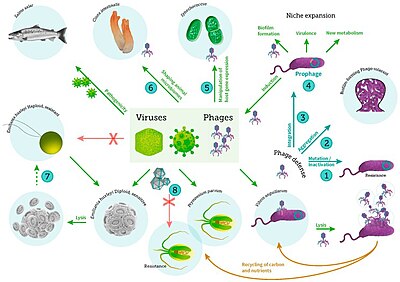
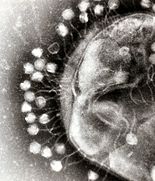
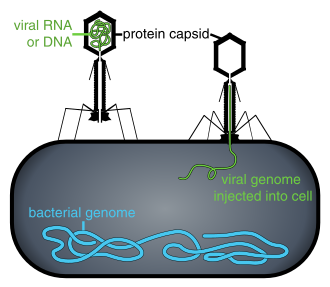
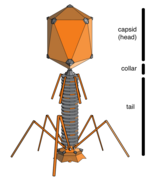


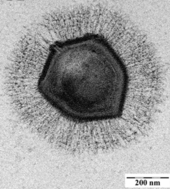



 Marine Thiomargarita namibiensis , крупнейшая известная бактерия
Marine Thiomargarita namibiensis , крупнейшая известная бактерия
![Хлоропласты глаукофитов имеют слой пептидогликана, свидетельствующие о их эндосимбиотическом происхождении из цианобактерий. [72]](//upload.wikimedia.org/wikipedia/commons/thumb/c/c9/Glaucocystis_sp.jpg/280px-Glaucocystis_sp.jpg) Хлоропласты , глаукофитов из имеют слой пептидогликана подтверждающие их эндосимбиотическое происхождение цианобактерий . [ 72 ]
Хлоропласты , глаукофитов из имеют слой пептидогликана подтверждающие их эндосимбиотическое происхождение цианобактерий . [ 72 ] Бактерия мариномонас Арктика растет внутри арктического морского ледя
Бактерия мариномонас Арктика растет внутри арктического морского ледя

![Хлоропласты глаукофитов имеют слой пептидогликана, свидетельствующие о их эндосимбиотическом происхождении из цианобактерий. [72]](http://upload.wikimedia.org/wikipedia/commons/thumb/c/c9/Glaucocystis_sp.jpg/280px-Glaucocystis_sp.jpg)


 Эоциты могут быть наиболее распространенными из морской археи
Эоциты могут быть наиболее распространенными из морской археи Halobacteria , обнаруженные в воде, почти насыщенные солью, теперь признаются археей.
Halobacteria , обнаруженные в воде, почти насыщенные солью, теперь признаются археей. Плоские, квадратные ячейки архей Haloquadratum walsbyi
Плоские, квадратные ячейки архей Haloquadratum walsbyi Methanosarcina Barkeri , морская архея, которая производит метатан
Methanosarcina Barkeri , морская архея, которая производит метатан Термофилы , такие как Pyrolobus fumarii , выживают более 100 ° C
Термофилы , такие как Pyrolobus fumarii , выживают более 100 ° C









![Диатомы являются основной группой водорослей, генерирующей около 20% мировой продукции кислорода. [93]](//upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg)
![Диатомовые средства имеют стекло, как клеточные стены из кремнезема, и называемые фрукторами. [94]](//upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg)
 Ископаемое диатомовое лечение от 32 до 40 млн лет
Ископаемое диатомовое лечение от 32 до 40 млн лет
 Один клетчатая водоросли, Gephyrocapsa Oceanica
Один клетчатая водоросли, Gephyrocapsa Oceanica Два динофлагеллята
Два динофлагеллята
 Zooxanthellae - это фотосинтетические водоросли, которые живут внутри хозяев, таких как Coral
Zooxanthellae - это фотосинтетические водоросли, которые живут внутри хозяев, таких как Coral
 Этот ресничный день переваривает цианобактерии . Цитостом . или рот внизу справа
Этот ресничный день переваривает цианобактерии . Цитостом . или рот внизу справа![Диатомы являются основной группой водорослей, генерирующей около 20% мировой продукции кислорода. [93]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg)
![Диатомовые средства имеют стекло, как клеточные стены из кремнезема, и называемые фрукторами. [94]](http://upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg)








 Одноклеточная гигантская амеба имеет до 1000 ядер и достигает длины 5 мм
Одноклеточная гигантская амеба имеет до 1000 ядер и достигает длины 5 мм![Gromia Sphaerica - это большое сферическое испытания амебы, которая делает грязевые тропы. Его диаметр до 3,8 см. [96]](//upload.wikimedia.org/wikipedia/commons/thumb/9/99/Gromia_in_situ_closeup.png/266px-Gromia_in_situ_closeup.png) Gromia Sphaerica - это большое сферическое испытания амебы , которая делает грязевые тропы. Его диаметр до 3,8 см. [ 96 ]
Gromia Sphaerica - это большое сферическое испытания амебы , которая делает грязевые тропы. Его диаметр до 3,8 см. [ 96 ] Spiculosiphon Oceana , одноклеточный фораминиферан с внешностью и образу жизни, который имитирует губку , растет до 5 см в длину.
Spiculosiphon Oceana , одноклеточный фораминиферан с внешностью и образу жизни, который имитирует губку , растет до 5 см в длину.![Ксенофиофор, еще один одноклетний фораманиферан, живет в зонах абиссалов. Он имеет гигантскую оболочку до 20 см в поперечнике. [97]](//upload.wikimedia.org/wikipedia/commons/thumb/5/5d/Xenophyophore.jpg/292px-Xenophyophore.jpg)
 Гигантская водоросли , коричневые водоросли , не является настоящим растением, но он многоклеточный и может расти до 50 м
Гигантская водоросли , коричневые водоросли , не является настоящим растением, но он многоклеточный и может расти до 50 м
![Gromia Sphaerica - это большое сферическое испытания амебы, которая делает грязевые тропы. Его диаметр до 3,8 см. [96]](http://upload.wikimedia.org/wikipedia/commons/thumb/9/99/Gromia_in_situ_closeup.png/266px-Gromia_in_situ_closeup.png)

![Ксенофиофор, еще один одноклетний фораманиферан, живет в зонах абиссалов. Он имеет гигантскую оболочку до 20 см в поперечнике. [97]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/5d/Xenophyophore.jpg/292px-Xenophyophore.jpg)








 Бактериальный жгутичок вращается молекулярным двигателем у его основания
Бактериальный жгутичок вращается молекулярным двигателем у его основания лосося Сперматозоид
лосося Сперматозоид
 Килиат Окситрича трифаллакс с ресничками четко видна
Килиат Окситрича трифаллакс с ресничками четко видна Амеба с проглатываемыми диатомогами
Амеба с проглатываемыми диатомогами






 Более 10 000 морских видов - копепод , маленькие, часто микроскопические ракообразные
Более 10 000 морских видов - копепод , маленькие, часто микроскопические ракообразные Фотография Даркфилда Гастротриха отложений , длиной 0,06-3,0 мм, животное, похожее на червя, живет между частицами
Фотография Даркфилда Гастротриха отложений , длиной 0,06-3,0 мм, животное, похожее на червя, живет между частицами Armored Pliciloricus engmaticus , длиной около 0,2 мм, живет в пространствах между морским гравием
Armored Pliciloricus engmaticus , длиной около 0,2 мм, живет в пространствах между морским гравием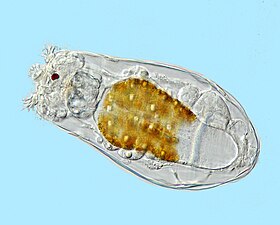 Враншивки , как правило, 0,1–0,5 мм, могут выглядеть как протисты, но многоклеточные и принадлежат Animalia
Враншивки , как правило, 0,1–0,5 мм, могут выглядеть как протисты, но многоклеточные и принадлежат Animalia Тардиграды (водные медведи), длиной около 0,5 мм, являются одними из самых устойчивых животных, известных
Тардиграды (водные медведи), длиной около 0,5 мм, являются одними из самых устойчивых животных, известных





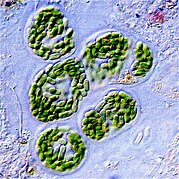
 Chlamydomonas globosa , одноклеточная зеленая водоросли с двумя жгутиками , только что видимыми внизу слева
Chlamydomonas globosa , одноклеточная зеленая водоросли с двумя жгутиками , только что видимыми внизу слева![Chlorella vulgaris, общие зеленые микроводоросли, в эндосимбиозе с ресничной частью [152]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b6/%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg/338px-%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg)
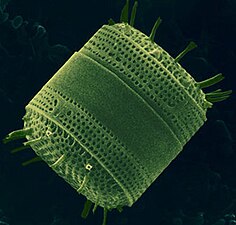 Центричный диатом
Центричный диатом Dinoflagellates
Dinoflagellates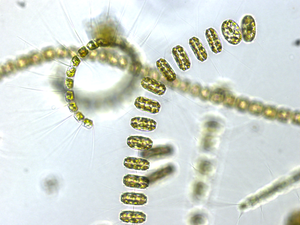 Колониальные цепочки водорослей
Колониальные цепочки водорослей
![Chlorella vulgaris, общие зеленые микроводоросли, в эндосимбиозе с ресничной частью [152]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b6/%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg/338px-%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg)





 Phytoplankton являются основой океанской пищевой цепи
Phytoplankton являются основой океанской пищевой цепи Они бывают разных форм и размеров.
Они бывают разных форм и размеров. Колониальный фитопланктон
Колониальный фитопланктон Cyanobacterium prochlorococcus составляет большую часть основного производства океана
Cyanobacterium prochlorococcus составляет большую часть основного производства океана Зеленая цианобактерийная мрака вымыта на скале в Калифорнии
Зеленая цианобактерийная мрака вымыта на скале в Калифорнии






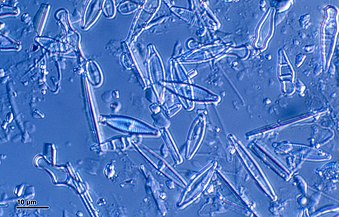 Диатомоты являются одним из наиболее распространенных типов фитопланктона
Диатомоты являются одним из наиболее распространенных типов фитопланктона Их защитные раковины (достоинства) изготовлены из кремния
Их защитные раковины (достоинства) изготовлены из кремния
 Они бывают разных форм и размеров
Они бывают разных форм и размеров



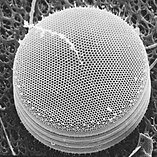
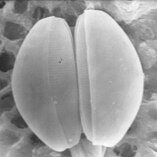
![Guinardia delicatula, диатомо, ответственный за цветение водорослей в Северном море и Английском канале [161]](//upload.wikimedia.org/wikipedia/commons/thumb/4/43/Fjouenne_sbrmvr012w_20070924163039_small.jpg/260px-Fjouenne_sbrmvr012w_20070924163039_small.jpg) Guinardia Delicatula , диатомо, ответственный за цветение водорослей в Северном море и Английском канале [ 161 ]
Guinardia Delicatula , диатомо, ответственный за цветение водорослей в Северном море и Английском канале [ 161 ] Ископаемое диатомо
Ископаемое диатомо Существует более 100 000 видов диатомовых заболеваний , которые составляют 50% основного производства океана
Существует более 100 000 видов диатомовых заболеваний , которые составляют 50% основного производства океана![Guinardia delicatula, диатомо, ответственный за цветение водорослей в Северном море и Английском канале [161]](http://upload.wikimedia.org/wikipedia/commons/thumb/4/43/Fjouenne_sbrmvr012w_20070924163039_small.jpg/260px-Fjouenne_sbrmvr012w_20070924163039_small.jpg)


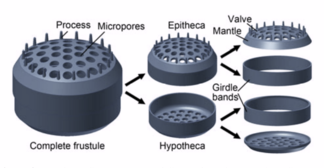
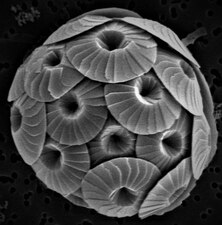 Кокколитофоры имеют тарелки, называемые кокколитами
Кокколитофоры имеют тарелки, называемые кокколитами Coccolithus pelagicus ssp. Braarudii
Coccolithus pelagicus ssp. Braarudii

 Блум водоросли Эмилиани Хакслии у южного побережья Англии
Блум водоросли Эмилиани Хакслии у южного побережья Англии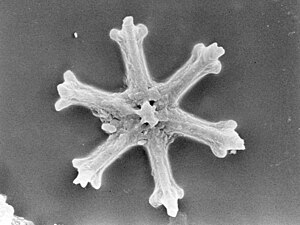 Вымерший ископаемый
Вымерший ископаемый





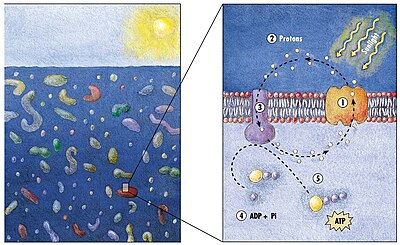



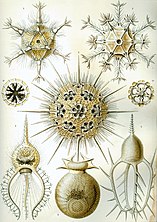
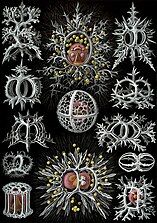
 Как диатомовые, радиолориалы бывают разных форм
Как диатомовые, радиолориалы бывают разных форм Также, как диатомовые ткатовые средства, радиояловые раковины обычно изготавливаются из силиката
Также, как диатомовые ткатовые средства, радиояловые раковины обычно изготавливаются из силиката Однако у акантарских радиолььеров есть раковины, изготовленные из сульфата стронция кристаллов
Однако у акантарских радиолььеров есть раковины, изготовленные из сульфата стронция кристаллов Схематическая схема сферической радиолоальной раковины
Схематическая схема сферической радиолоальной раковины Cladococcus abietinus
Cladococcus abietinus




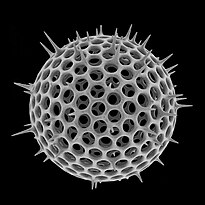




 раздел, показывающий камеры спирального фурама
раздел, показывающий камеры спирального фурама Живой аммиак тепида потоковой гранулированной эктоплазмы для ловли пищи
Живой аммиак тепида потоковой гранулированной эктоплазмы для ловли пищи Группа планктонных фурамов
Группа планктонных фурамов Ископаемые номерулитидные отверстия различных размеров от эоцена
Ископаемые номерулитидные отверстия различных размеров от эоцена![Египетские пирамиды были построены из известняка, в котором содержались нумерулиты. [188]](//upload.wikimedia.org/wikipedia/commons/thumb/a/af/All_Gizah_Pyramids.jpg/325px-All_Gizah_Pyramids.jpg)




![Египетские пирамиды были построены из известняка, в котором содержались нумерулиты. [188]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/af/All_Gizah_Pyramids.jpg/325px-All_Gizah_Pyramids.jpg)
 Testate Amoeba , Cyphoderia sp.
Testate Amoeba , Cyphoderia sp. Оболочка или проверка тестирования амебы , Arcella sp.
Оболочка или проверка тестирования амебы , Arcella sp. Ксеногенное испытания амеба, покрытая диатомогами (из коллекции амебы Пенард )
Ксеногенное испытания амеба, покрытая диатомогами (из коллекции амебы Пенард ) Голая амеба, хаос sp.
Голая амеба, хаос sp. Обнаженная амеба, показывающая продовольственную вакуоли и проглатывается диатомо
Обнаженная амеба, показывающая продовольственную вакуоли и проглатывается диатомо




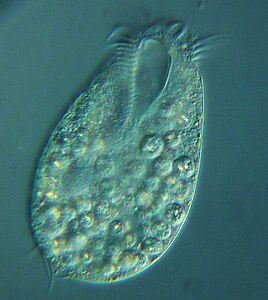

 Голофира Яйцо
Голофира Яйцо![Месодиниевый рубрим производит глубокие красные цветы, используя растворенные хлоропласты из их добычи водорослей [191]](//upload.wikimedia.org/wikipedia/commons/thumb/d/df/Mesodinium_rubrum.jpg/300px-Mesodinium_rubrum.jpg) Мезодиниевый рубрим производит глубокие красные цветы, используя растворенные хлоропласты из их добычи водорослей [ 191 ]
Мезодиниевый рубрим производит глубокие красные цветы, используя растворенные хлоропласты из их добычи водорослей [ 191 ]


![Месодиниевый рубрим производит глубокие красные цветы, используя растворенные хлоропласты из их добычи водорослей [191]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/df/Mesodinium_rubrum.jpg/300px-Mesodinium_rubrum.jpg)

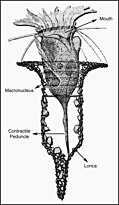
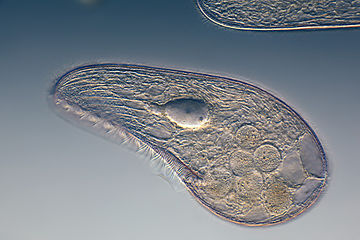







 Тинтиннид Ресничная Фавелла
Тинтиннид Ресничная Фавелла Euglena -изменение , фотосинтетический жгутичок
Euglena -изменение , фотосинтетический жгутичок Zoochlorellae (зеленый) Жизнь в ресничной Stichotricha Secunda
Zoochlorellae (зеленый) Жизнь в ресничной Stichotricha Secunda


 Гиродиния , один из немногих обнаженных динофлагеллятов, в которых отсутствует броня
Гиродиния , один из немногих обнаженных динофлагеллятов, в которых отсутствует броня Dinoflagellate Protoperidinium вытягивает большую кормление, чтобы захватить добычу
Dinoflagellate Protoperidinium вытягивает большую кормление, чтобы захватить добычу Населларийские радиольолярии могут быть в симбиозе с динофлагеллятами
Населларийские радиольолярии могут быть в симбиозе с динофлагеллятами Динофлагеллятный динофиз acuta
Динофлагеллятный динофиз acuta




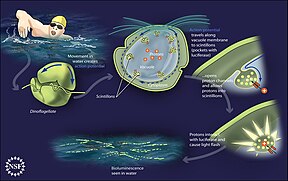


 Tripos Muelleri узнаваемы по его образным рогам
Tripos Muelleri узнаваемы по его образным рогам![Ooodinium, род паразитических динофлагеллятов, вызывает бархатную болезнь у рыб [213]](//upload.wikimedia.org/wikipedia/commons/thumb/8/87/Archives_de_zoologie_exp%C3%A9rimentale_et_g%C3%A9n%C3%A9rale_%281920%29_%2820299351186%29.jpg/260px-Archives_de_zoologie_exp%C3%A9rimentale_et_g%C3%A9n%C3%A9rale_%281920%29_%2820299351186%29.jpg)
![Karenia Brevis производит красные приливы, высокотоксичные для людей [214]](//upload.wikimedia.org/wikipedia/commons/thumb/a/a0/Karenia_brevis.jpg/214px-Karenia_brevis.jpg) Karenia Brevis производит красные приливы, высоко токсичные для людей [ 214 ]
Karenia Brevis производит красные приливы, высоко токсичные для людей [ 214 ]
![Noctiluca Scintillans, биолюминесцентный динофлагеллят [215]](//upload.wikimedia.org/wikipedia/commons/thumb/6/62/Noctiluca_scintillans_unica.jpg/196px-Noctiluca_scintillans_unica.jpg) Noctiluca Scintillans , биолюминесцентный динофлагеллят [ 215 ]
Noctiluca Scintillans , биолюминесцентный динофлагеллят [ 215 ]
![Ooodinium, род паразитических динофлагеллятов, вызывает бархатную болезнь у рыб [213]](http://upload.wikimedia.org/wikipedia/commons/thumb/8/87/Archives_de_zoologie_exp%C3%A9rimentale_et_g%C3%A9n%C3%A9rale_%281920%29_%2820299351186%29.jpg/260px-Archives_de_zoologie_exp%C3%A9rimentale_et_g%C3%A9n%C3%A9rale_%281920%29_%2820299351186%29.jpg)
![Karenia Brevis производит красные приливы, высокотоксичные для людей [214]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/a0/Karenia_brevis.jpg/214px-Karenia_brevis.jpg)

![Noctiluca Scintillans, биолюминесцентный динофлагеллят [215]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/62/Noctiluca_scintillans_unica.jpg/196px-Noctiluca_scintillans_unica.jpg)








 Диатоматическая земля - это мягкая, кремнистая , осадочная порода, состоящая из микрофоссилий в виде ящиков (оболочки) одноклеточных диатомовых диатомов (нажмите, чтобы увеличить)
Диатоматическая земля - это мягкая, кремнистая , осадочная порода, состоящая из микрофоссилий в виде ящиков (оболочки) одноклеточных диатомовых диатомов (нажмите, чтобы увеличить) Известные микрофоссили из морских осадков, состоящих в основном из звездных диско-атмосферов с разбрызгиванием кокколитов
Известные микрофоссили из морских осадков, состоящих в основном из звездных диско-атмосферов с разбрызгиванием кокколитов Оболочки ( тесты ), обычно изготовленные из карбоната кальция, из фораминиферальной слизки на глубоком дне океана
Оболочки ( тесты ), обычно изготовленные из карбоната кальция, из фораминиферальной слизки на глубоком дне океана Иллюстрация Globigerina ooze
Иллюстрация Globigerina ooze![Мрамор может содержать протеристические микрофоссили фораминиферов, кокколитофоров, известковых нанопланктонов и водорослей, остракодов, птероподов, кальпионеллидов и Bryozoa [222]](//upload.wikimedia.org/wikipedia/commons/thumb/6/61/MarmoCipollino_FustoBasMassenzioRoma.jpg/320px-MarmoCipollino_FustoBasMassenzioRoma.jpg) Мрамор может содержать протеристические микрофоссили фораминиферанов, кокколитофоров, известковых нанопланктонов и водорослей, остракодов , птероподов , кальпионеллидов и Брайозои . [ 222 ]
Мрамор может содержать протеристические микрофоссили фораминиферанов, кокколитофоров, известковых нанопланктонов и водорослей, остракодов , птероподов , кальпионеллидов и Брайозои . [ 222 ]![Опал может содержать протестные микрофоссили диатомовых, радиолььеров, силикофлагеллятов и эбридианцев [222]](//upload.wikimedia.org/wikipedia/commons/thumb/d/d9/Coober_Pedy_Opal_Doublet.jpg/352px-Coober_Pedy_Opal_Doublet.jpg) Опал может содержать протестные микрофоссили диатомовых, радиолориалов, силикофлагеллятов и эбридианцев [ 222 ]
Опал может содержать протестные микрофоссили диатомовых, радиолориалов, силикофлагеллятов и эбридианцев [ 222 ]




![Мрамор может содержать протеристические микрофоссили фораминиферов, кокколитофоров, известковых нанопланктонов и водорослей, остракодов, птероподов, кальпионеллидов и Bryozoa [222]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/61/MarmoCipollino_FustoBasMassenzioRoma.jpg/320px-MarmoCipollino_FustoBasMassenzioRoma.jpg)
![Опал может содержать протестные микрофоссили диатомовых, радиолььеров, силикофлагеллятов и эбридианцев [222]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/d9/Coober_Pedy_Opal_Doublet.jpg/352px-Coober_Pedy_Opal_Doublet.jpg)



 Распределение типов отложений на морском дне
Распределение типов отложений на морском дне



 Elphidium широко распространенный род бентических фурамов
Elphidium широко распространенный род бентических фурамов Гетерогеликс , вымерший род бентических фурамов
Гетерогеликс , вымерший род бентических фурамов

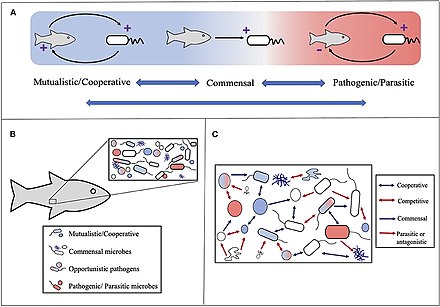
![Coral Holobiont [240]](http://upload.wikimedia.org/wikipedia/commons/thumb/2/26/Trophic_connections_of_the_coral_holobiont_in_the_planktonic_food_web.jpg/237px-Trophic_connections_of_the_coral_holobiont_in_the_planktonic_food_web.jpg)
![Губка Холобонт [241]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/6b/The_sponge_holobiont.webp/304px-The_sponge_holobiont.webp.png)
![Seargass Holobiont [242]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/35/Processes_within_the_seagrass_holobiont.webp/197px-Processes_within_the_seagrass_holobiont.webp.png)
![Изменение климата и родолит Холобионт [243]](http://upload.wikimedia.org/wikipedia/commons/thumb/f/f7/Climate_change_stressors_and_rhodolith_holobiont_fitness.webp/415px-Climate_change_stressors_and_rhodolith_holobiont_fitness.webp.png)
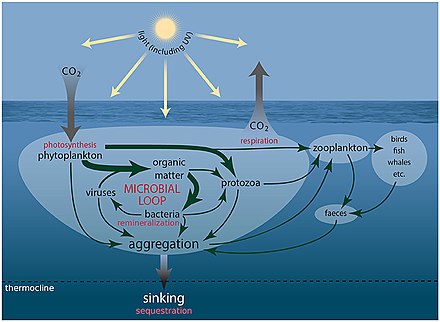

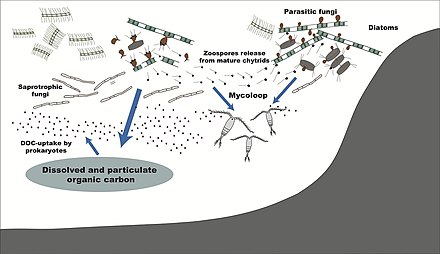

 Pelagibacter Ubique , наиболее распространенные бактерии в океане, играет важную роль в глобальном углеродном цикле .
Pelagibacter Ubique , наиболее распространенные бактерии в океане, играет важную роль в глобальном углеродном цикле .![Морской снег - это душ органических частиц, который падает из верхних вод в глубокий океан [248], это основной экспортер углерода.](//upload.wikimedia.org/wikipedia/commons/thumb/5/59/Marine_snow.jpg/270px-Marine_snow.jpg) Морской снег - это душ органических частиц, который падает из верхних вод в глубокий океан [ 248 ] Это основной экспортер углерода.
Морской снег - это душ органических частиц, который падает из верхних вод в глубокий океан [ 248 ] Это основной экспортер углерода.
![Морской снег - это душ органических частиц, который падает из верхних вод в глубокий океан [248], это основной экспортер углерода.](http://upload.wikimedia.org/wikipedia/commons/thumb/5/59/Marine_snow.jpg/270px-Marine_snow.jpg)




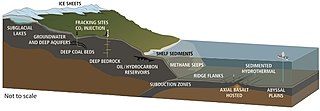
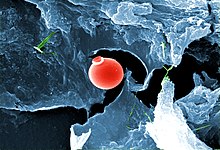
![Оценки подсчета микробных видов в трех доменах жизненных бактерий являются самой старой и наиболее биоразнообразие, за которой следуют археа и грибы (самые последние группы). В 1998 году, до того, как началась осознание степени микробной жизни, Роберт М. Мэй [274] по оценкам, на планете было 3 миллиона видов живых организмов. Но в 2016 году Лоси и Леннон [275] оценили, что число видов микроорганизма может достигать 1 триллиона. [276]](//upload.wikimedia.org/wikipedia/commons/thumb/b/bc/Microbial_species_present_in_the_three_domains_of_life.png/889px-Microbial_species_present_in_the_three_domains_of_life.png) Оценки микробных видов считаются в трех доменах жизниБактерии - самая старая и самая биоративная группа, за которой следуют археа и грибы (самые последние группы). В 1998 году, до того, как началась осознание степени микробной жизни, Роберт М. Мэй [ 274 ] По оценкам, на планете было 3 миллиона видов живых организмов. Но в 2016 году Лоси и Леннон [ 275 ] По оценкам, количество видов микроорганизма может достигать 1 триллиона. [ 276 ]
Оценки микробных видов считаются в трех доменах жизниБактерии - самая старая и самая биоративная группа, за которой следуют археа и грибы (самые последние группы). В 1998 году, до того, как началась осознание степени микробной жизни, Роберт М. Мэй [ 274 ] По оценкам, на планете было 3 миллиона видов живых организмов. Но в 2016 году Лоси и Леннон [ 275 ] По оценкам, количество видов микроорганизма может достигать 1 триллиона. [ 276 ]![Микробное разнообразие Сравнительное представление известного и оцененного (маленького ящика) и еще неизвестного (крупного) микробного разнообразия, которое применяется как к морским, так и к наземным микроорганизмам. Текстовые поле относятся к факторам, которые отрицательно влияют на знание микробного разнообразия, которое существует на планете. [276]](//upload.wikimedia.org/wikipedia/commons/thumb/1/16/Known%2C_estimated_and_unknown_microbial_diversity.webp/661px-Known%2C_estimated_and_unknown_microbial_diversity.webp.png) Микробное разнообразиеСравнительное представление известного и предполагаемого (маленького ящика) и еще неизвестного (большой коробки) микробного разнообразия, которое относится как к морским, так и к наземным микроорганизмам. Текстовые поле относятся к факторам, которые отрицательно влияют на знание микробного разнообразия, которое существует на планете. [ 276 ]
Микробное разнообразиеСравнительное представление известного и предполагаемого (маленького ящика) и еще неизвестного (большой коробки) микробного разнообразия, которое относится как к морским, так и к наземным микроорганизмам. Текстовые поле относятся к факторам, которые отрицательно влияют на знание микробного разнообразия, которое существует на планете. [ 276 ]![Оценки подсчета микробных видов в трех доменах жизненных бактерий являются самой старой и наиболее биоразнообразие, за которой следуют археа и грибы (самые последние группы). В 1998 году, до того, как началась осознание степени микробной жизни, Роберт М. Мэй [274] по оценкам, на планете было 3 миллиона видов живых организмов. Но в 2016 году Лоси и Леннон [275] оценили, что число видов микроорганизма может достигать 1 триллиона. [276]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/bc/Microbial_species_present_in_the_three_domains_of_life.png/889px-Microbial_species_present_in_the_three_domains_of_life.png)
![Микробное разнообразие Сравнительное представление известного и оцененного (маленького ящика) и еще неизвестного (крупного) микробного разнообразия, которое применяется как к морским, так и к наземным микроорганизмам. Текстовые поле относятся к факторам, которые отрицательно влияют на знание микробного разнообразия, которое существует на планете. [276]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/16/Known%2C_estimated_and_unknown_microbial_diversity.webp/661px-Known%2C_estimated_and_unknown_microbial_diversity.webp.png)

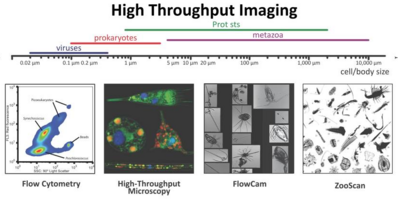
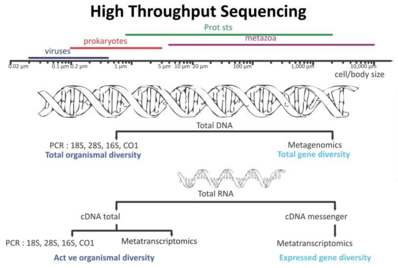


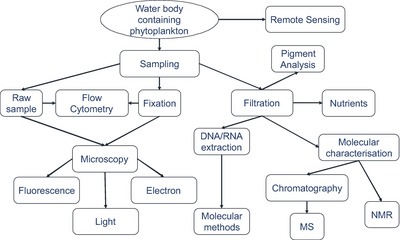
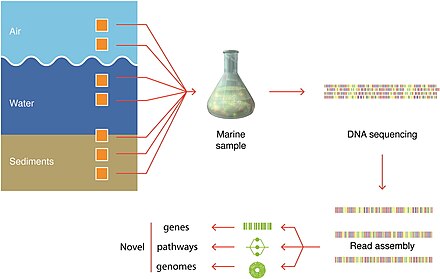
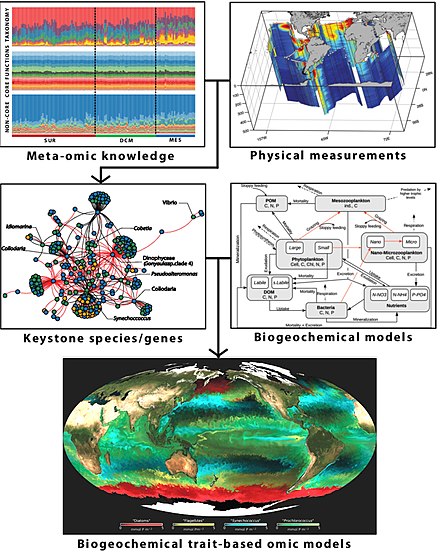

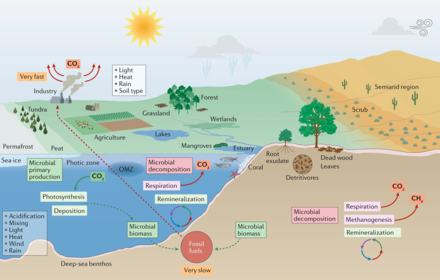
 Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
 Эта статья включает текст из этого источника, который находится в общественном доступе .
Эта статья включает текст из этого источника, который находится в общественном доступе .
