G белок
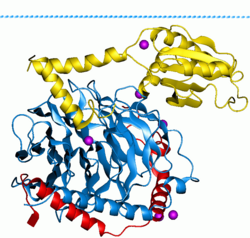
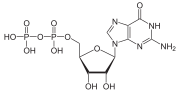

G-белки , также известные как гуаниновые нуклеотидсвязывающие белки , представляют собой семейство белков , которые действуют как молекулярные переключатели внутри клеток, и участвуют в передаче сигналов из различных стимулов вне клетки в его интерьер. Их активность регулируется факторами, которые контролируют их способность связываться и гидролизовать гуанозин -трифосфат (GTP) к гуанозин -дифхосфату (ВВП). Когда они связаны с GTP, они «включены», и, когда они связаны с ВВП, они «выключены». G -белки принадлежат к более крупной группе ферментов, называемых GTPases .
Есть два класса белка G. Первая функция как мономерные небольшие GTPases (небольшие G-белки), в то время как вторая функция как гетеротримерные комплексы G-белков . состоит из альфа (G α ), бета (G β ) и гамма (G γ ) Последний класс комплексов . [ 1 ] Кроме того, субъединицы бета и гамма могут образовывать стабильный димерный комплекс, называемый комплексом бета-гамма Полем [ 2 ]
Гетеротримерные G-белки, расположенные в клетках, активируются с помощью G-белковых рецепторов (GPCR), которые охватывают клеточную мембрану . [ 3 ] Сигнальные молекулы связываются с доменом GPCR, расположенной вне клетки, и внутриклеточный домен GPCR, а затем, в свою очередь, активирует конкретный G -белок. Было также показано, что некоторые GPCR активного состояния являются «предварительно связанными» с G-белками, тогда как в других случаях считается, что механизм связывания столкновения. [ 4 ] [ 5 ] [ 6 ] G -белок запускает каскад дальнейших сигнальных событий , которые в конечном итоге приводят к изменению функции клеток. G-белковые рецепторы и G-белки, работающие вместе передают сигналы из многих гормонов , нейротрансмиттеров и других сигнальных факторов. [ 7 ] G -белки регулируют метаболические ферменты , ионные каналы , транспортерные белки и другие части клеточного механизма, контролируя транскрипцию , подвижность , сократимость и секрецию , которые, в свою очередь, регулируют различные системные функции, такие как эмбриональное развитие , обучение и память, а также гомеостаз . [ 8 ]
История
[ редактировать ]G -белки были обнаружены в 1980 году, когда Альфред Г. Гилман и Мартин Родбелл исследовали стимуляцию клеток с помощью адреналина . Они обнаружили, что когда адреналин связывается с рецептором, рецептор не стимулирует ферменты (внутри клетки) напрямую. Вместо этого рецептор стимулирует G -белок, который затем стимулирует фермент. Примером является аденилатциклаза , которая производит Messenger второй циклический AMP . [ 9 ] Для этого открытия они получили Нобелевскую премию 1994 года по физиологии или медицине . [ 10 ]
Нобелевские призы были награждены за многие аспекты передачи сигналов G -белками и GPCR. К ним относятся антагонисты рецепторов , нейротрансмиттеры нейротрансмиттера , обратный захват , G-белковые рецепторы , G-белки, вторые мессенджеры , ферменты, которые запускают фосфорилирование белка в ответ на цАМФ и последующие метаболические процессы, такие как гликогенолиз .
Выдающиеся примеры включают (в хронологическом порядке награждения):
- Нобелевская премия 1947 года за физиологию или медицину для Карла Кори , Герти Кори и Бернардо , за их обнаружение того, как гликоген разбит на глюкозу и восстанавливается в организме, для использования в качестве магазина и источника энергии. Гликогенолиз стимулируется многочисленными гормонами и нейротрансмиттерами, включая адреналин .
- Нобелевская премия 1970 года по физиологии или медицине Джулиусу Аксельроду , Бернарду Кацу и Ульф фон Эйлеру за работу по выпуску обращению нейротрансмиттеров и .
- Нобелевская премия 1971 года по физиологии или медицине графу Сазерленду за обнаружение ключевой роли аденилатциклазы , которая производит второй циклический усилитель . [ 9 ]
- Нобелевская премия 1988 года по физиологии или медицине Джорджу Х. Хитчингсу , сэру Джеймсу Блэку и Гертруде Элион за их открытиями важных принципов для лечения наркотиков », нацеленных на GPCR.
- Нобелевская премия 1992 года по физиологии или медицине Эдвину Г. Кребсу и Эдмонду Х. Фишеру за описание того, как обратимое фосфорилирование работает как переключатель для активации белков , и для регулирования различных клеточных процессов, включая гликогенолиз . [ 11 ]
- Нобелевская премия 1994 года по физиологии или медицине Альфреду Г. Гилману и Мартину Родбеллу за их открытие «G-белков и роль этих белков в трансдукции сигнала в клетках». [ 12 ]
- Нобелевская премия 2000 года по физиологии или медицине Эрику Канделю , Арвиду Карлссону и Полу Грингарду , для исследований нейротрансмиттеров, таких как дофамин , которые действуют через GPCR.
- Нобелевская премия 2004 года по физиологии или медицине Ричарду Акселю и Линде Б. Бак за их работу над обонятельными рецепторами, связанными с G, G. [ 13 ]
- Нобелевская премия 2012 года по химии Брайану Кобильке и Роберту Лефковицу за работу по функции GPCR. [ 14 ]
Функция
[ редактировать ]G -белки являются важными молекулами передачи сигнала в клетках. «Неисправность сигнальных путей GPCR [G-белкового рецептора] участвует во многих заболеваниях, таких как диабет , слепота, аллергия, депрессия, сердечно-сосудистые дефекты и некоторые формы рака . По оценкам, около 30% современных препаратов ' Коточные мишени являются GPCR. " [ 15 ] Человеческий геном кодирует примерно 800 [ 16 ] G-белок-связанные рецепторы , которые обнаруживают фотоны света, гормонов, факторов роста, лекарств и других эндогенных лигандов . Приблизительно 150 GPCR, обнаруженных в геноме человека, все еще имеют неизвестные функции.
Принимая во внимание, что G-белки активируются с помощью G-белка-рецепторов , они инактивируются белками RGS (для «регулятора передачи сигналов G-белка»). Рецепторы стимулируют связывание GTP (включение G -белка). Белки RGS стимулируют гидролиз GTP (создавая ВВП, таким образом выключая G -белок).
Разнообразие
[ редактировать ]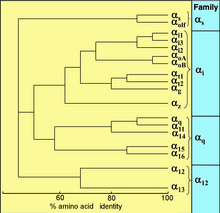
Все эукариоты используют G -белки для передачи сигналов и развили большое разнообразие G -белков. Например, люди кодируют 18 различных G α белков , 5 г β -белков и 12 г γ -белков. [ 17 ]
Сигнализация
[ редактировать ]G белок может относиться к двум различным семействам белков. Гетеротримерные G-белки , иногда называемые «большими» G-белками, активируются рецепторами G-белков и состоят из субъединиц альфа (α), бета (β) и гамма (γ) . «Маленькие» G-белки (20-25 кДа) принадлежат к RAS суперсемейству небольших GTPases . Эти белки гомологичны субъединице альфа (α), обнаруженной в гетеротримерах, но на самом деле являются мономерными, состоящие только из одной единицы. Однако, как и их более крупные родственники, они также связывают GTP и ВВП и участвуют в трансдукции сигнала .
Гетеротримерный
[ редактировать ]Различные типы гетеротримерных G -белков имеют общий механизм. Они активируются в ответ на конформационное изменение в GPCR, обмену GDP на GTP и диссоциируя для активации других белков в конкретном пути передачи сигнала . [ 18 ] Специфические механизмы, однако, различаются между типами белков.
Механизм
[ редактировать ]
Рецептор-активированные G-белки связаны с внутренней поверхностью клеточной мембраны . Они состоят из G α и тесно связанных субъединиц G βγ . Существует четыре основных семейства субъединиц G α : Gα S (G (G -стимулирующий), Gα I (G -ингибирование), Gα Q/11 и Gα 12/13 . [ 20 ] [ 21 ] Они ведут себя по -разному в распознавании эффекторной молекулы, но имеют аналогичный механизм активации.
Активация
[ редактировать ]Когда лиганд активирует рецептор, связанный с G-белком , он индуцирует конформационное изменение рецептора, которое позволяет рецептору функционировать как коэффициент обмена нуклеотидов гуанина (GEF), который обменивает ВВП на GTP. GTP (или GDP) связан с субъединицей G α в традиционном взгляде на гетеротримерную активацию GPCR. Этот обмен вызывает диссоциацию субъединицы G α (которая связана с GTP) из димера G βγ и рецептора в целом. Тем не менее, модели, которые предполагают молекулярную перестройку, реорганизацию и предварительную компетенцию эффекторных молекул, начинают принимать. [ 4 ] [ 22 ] [ 23 ] И G α -GTP, так и G βγ затем могут активировать различные сигнальные каскады (или мессенджера вторые путями ) и эффекторные белки, в то время как рецептор способен активировать следующий G -белок. [ 24 ]
Завершение
[ редактировать ]Субъединица G α в конечном итоге будет гидролизовать прикрепленный GTP к GDP по его присущей ферментативной активности, позволяя ему повторно ассоциироваться с G βγ и запустить новый цикл. Группа белков, называемых регулятором передачи сигналов G-белка (RGSS), действуя как активирующие GTPase белки (промежутки), специфичны для G α субъединиц . Эти белки ускоряют гидролиз GTP до ВВП, таким образом заканчивая передачу сигнала. эффектор В некоторых случаях сам может обладать внутренней активностью разрыва, которая затем может помочь отключить путь. Это верно в случае фосфолипазы C -бета, которая обладает активностью GAP в его C -концевой области. Это альтернативная форма регуляции для субъединицы G α . Такие промежутки G α не имеют каталитических остатков (специфических аминокислотных последовательностей) для активации белка G α . Вместо этого они работают, снижая необходимую энергию активации для реализации. [ 25 ]
Конкретные механизмы
[ редактировать ]G αS
[ редактировать ]G αS активирует CAMP-зависимый путь , стимулируя производство циклического AMP (CAMP) из АТФ . Это достигается путем прямой стимуляции мембраны, ассоциированной с ферментом, аденилатциклазы . Затем лагерь может действовать как второй посланник, который продолжает взаимодействовать и активировать протеинкиназу A (PKA). PKA CAN фосфорилирует бесчисленные нисходящие цели.
КАМФ -зависимый путь используется в качестве пути передачи сигнала для многих гормонов, включая:
- ADH - способствует задержке воды почками ( созданным магноцеллюлярными нейросекреторными клетками заднего гипофиза )
- GHRH - стимулирует синтез и высвобождение GH ( соматотропные клетки переднего гипофиза )
- Ghih - ингибирует синтез и высвобождение GH (соматотропные клетки переднего гипофиза)
- CRH - стимулирует синтез и высвобождение АКТГ (передний гипофиз)
- ACTH - стимулирует синтез и высвобождение кортизола ( Zona Fasciculata в коры надпочечников надпочечниках)
- TSH - стимулирует синтез и высвобождение большинства T4 (щитовидная железа)
- LH - стимулирует фолликулярное созревание и овуляцию у женщин; или продукция тестостерона и сперматогенез у мужчин
- FSH - стимулирует развитие фолликула у женщин; или сперматогенез у мужчин
- ПТГ - повышает уровень кальция в крови . Это достигается с помощью рецептора паращитовидного гормона 1 (PTH1) в почках и костях, или через рецептор паращитовидного гормона 2 (PTH2) в центральной нервной системе и мозге, а также кости и почки.
- Кальцитонин - снижает уровень кальция в крови (через рецептор кальцитонина в кишечнике, костях, почках и мозге)
- Глюкагон - стимулирует распад гликогена в печени
- ХГЧ - способствует клеточной дифференцировке и потенциально участвует в апоптозе . [ 26 ]
- Адреналин - выделяется медедом надпочечников во время состояния голодания, когда тело находится в метаболическом принуждении. Он стимулирует гликогенолиз , в дополнение к действию глюкагона .
G αi
[ редактировать ]G αi ингибирует производство лагеря из АТФ. Например, соматостатин, простагландины
G αQ/11
[ редактировать ]G αQ/11 стимулирует мембраносвязанную фосфолипазу C , которая затем расщепляет фосфатидилинозитол 4,5-бисфосфат (PIP 2 ) на два вторых мессенджера, иннозитол трипросфат (IP 3 ) и диацилглицерин (DAG). IP 3 индуцирует высвобождение кальция из эндоплазматической ретикулумы . DAG активирует протеинкиназу c . В качестве пути трансдукции сигнала для многих гормонов используется путем, зависящий от фосфолипидов.
- Адреналин
- ADH ( вазопрессин /AVP) - индуцирует синтез и высвобождение глюкокортикоидов ( Zona Fasciculata ) коры надпочечников ; Индуцирует вазоконстрикцию (клетки V1 заднего гипофиза )
- TRH - индуцирует синтез и высвобождение TSH ( передняя гипофиза )
- TSH - индуцирует синтез и высвобождение небольшого количества T4 ( щитовидная железа )
- Ангиотензин II - индуцирует синтез и высвобождение альдостерона ( Zona Glomerulosa коры надпочечников в почках)
- GNRH - индуцирует синтез и высвобождение FSH и LH (передний гипофиз)
G A12/13
[ редактировать ]- G α12/13 участвует в передаче сигналов GTPase Family RHO (см. Семейство GTPase RHO ). Это происходит через суперсемейство Rhogef с участием домена Rhogef структур белков). Они участвуют в контроле ремоделирования клеточного цитоскелета и, следовательно, в регуляции миграции клеток.
G B , G
[ редактировать ]- Комплексы G βγ иногда также имеют активные функции. Примеры включают в себя связь и активирование G-белка, связанного с внутренним реекцией калия .
Маленькие GTPases
[ редактировать ]Небольшие GTPases, также известные как небольшие G-белок, также связывают GTP и GDP, и участвуют в трансдукции сигнала . Эти белки гомологичны субъединице альфа (α), обнаруженной в гетеротримерах, но существуют в виде мономеров. Это небольшие белки (от 20 кДа до 25 кДа) , которые связываются с гуанозин-трифосфатом ( GTP ). Это семейство белков гомологично для GTPases Ras и также называется GTPases Superbamily RAS .
Липидация
[ редактировать ]Чтобы связаться с внутренней листочной листовой плазматической мембраной, многие G -белки и небольшие GTPases липидируются [ Цитация необходима ] , то есть ковалентно модифицированным с помощью липидов. Они могут быть миристоилированными , пальмитоилированными или пренилированными .
Ссылки
[ редактировать ]- ^ Hurowitz EH, Melnyk JM, Chen YJ, Kouros-Mehr H, Simon Mi, Shizuya H (апрель 2000 г.). «Геномная характеристика человеческого гетеротримерного G -белка Alpha, бета и гамма -субъединичных генов» . ДНК -исследования . 7 (2): 111–20. doi : 10.1093/dnares/7.2.111 . PMID 10819326 .
- ^ Клэпхэм Д.Е., Ниер Э.Дж. (1997). «G белок бета -гамма -субъединицы». Ежегодный обзор фармакологии и токсикологии . 37 : 167–203. doi : 10.1146/annurev.pharmtox.37.1.167 . PMID 9131251 .
- ^ «Семь трансмембранных рецепторов: Роберт Лефковиц» . 9 сентября 2012 года . Получено 11 июля 2016 года .
- ^ Jump up to: а беременный Цинь К, Донг С., Ву Г, Ламберт На (август 2011 г.). «Неактивная предварительная символа G (Q)-связанных рецепторов и гетеротримеров G (Q)» . Природная химическая биология . 7 (10): 740–7. doi : 10.1038/nchembio.642 . PMC 3177959 . PMID 21873996 .
- ^ Толковский А.М., Левицки А. (1978). «Режим связи между бета-адренергическим рецептором и аденилатциклазой в эритроцитах индейки». Биохимия . 17 (18): 3795–3810. doi : 10.1021/bi00611a020 . PMID 212105 .
- ^ Boltz HH, Sirbu A, Stelzer N, De Lanerolle P, Winkelmann S, Annibale P (2022). «Влияние диффузии мембранного белка на передачу сигналов GPCR» . Ячейки 11 (10): 1660. doi : 10.3390/cells11101660 . PMC 9139411 . PMID 35626696 .
- ^ Reece J, CN (2002). Биология . Сан -Франциско: Бенджамин Каммингс. ISBN 0-8053-6624-5 .
- ^ Neves Sr, Ram Pt, Iyengar R (май 2002). "G белковые пути". Наука . 296 (5573): 1636–9. Bibcode : 2002sci ... 296.1636n . doi : 10.1126/science.1071550 . PMID 12040175 . S2CID 20136388 .
- ^ Jump up to: а беременный Нобелевская премия по физиологии или медицине 1994 , иллюстрированная лекция.
- ^ Пресс-релиз: Нобелевская ассамблея в Институте Каролински решила вручить Нобелевскую премию по физиологии или медицине в 1994 году совместно с Альфредом Г. Гилманом и Мартином Родбеллом за их открытие «G-белков и роль этих белков в трансдукции сигнала в клетки ». 10 октября 1994
- ^ «Нобелевская премия в области физиологии или медицины 1992 года» . Нобелевское собрание в Институте Каролинской . Получено 21 августа 2013 года .
- ^ Пресс -релиз
- ^ «Пресс -релиз: Нобелевская премия 2004 года по физиологии или медицине» . Nobelprize.org . Получено 8 ноября 2012 года .
- ^ Королевская шведская академия наук (10 октября 2012 г.). «Нобелевская премия по химии 2012 Роберт Дж. Лефковиц, Брайан К. Кобилка» . Получено 10 октября 2012 года .
- ^ Bosch DE, Siderovski DP (март 2013 г.). «G -белок передача сигналов в паразите entamoeba histolytica» . Экспериментальная и молекулярная медицина . 45 (1038): E15. doi : 10.1038/emm.2013.30 . PMC 3641396 . PMID 23519208 .
- ^ Baltoumas FA, Theodoropoulou MC, Hamodrakas SJ (июнь 2013 г.). «Взаимодействия α-субъединиц гетеротримерных G-белков с GPCR, эффекторами и белками RGS: критический обзор и анализ взаимодействующих поверхностей, конформационные сдвиги, структурное разнообразие и электростатические потенциалы». Журнал структурной биологии . 182 (3): 209–18. doi : 10.1016/j.jsb.2013.03.004 . PMID 23523730 .
- ^ Jump up to: а беременный Syrovatkina V, Alegre Ko, Dey R, Huang XY (сентябрь 2016 г.). «Регуляция, передача сигналов и физиологические функции G-белков» . Журнал молекулярной биологии . 428 (19): 3850–68. doi : 10.1016/j.jmb.2016.08.002 . PMC 5023507 . PMID 27515397 .
- ^ Лим, Венделл (2015). Сигнализация ячейки: принципы и механизмы . Брюс Майер, Т. Поусон. Нью-Йорк. ISBN 978-0-8153-4244-1 Полем OCLC 868641565 .
{{cite book}}: CS1 Maint: местоположение отсутствует издатель ( ссылка ) - ^ Стюарт, Адель; Фишер, Рори А. (2015). Прогресс в молекулярной биологии и трансляционной науке . Тол. 133. Elsevier. С. 1–11. doi : 10.1016/bs.pmbts.2015.03.002 . ISBN 9780128029381 Полем PMID 26123299 .
- ^ Сироваткина, Виктория; Алегре, Камела О.; Дей, Раджа; Хуан, Синь-Юн (25 сентября 2016 г.). «Регуляция, передача сигналов и физиологические функции G-белков» . Журнал молекулярной биологии . 428 (19): 3850–3868. doi : 10.1016/j.jmb.2016.08.002 . ISSN 0022-2836 . PMC 5023507 . PMID 27515397 .
- ^ "InterPro" . www.ebi.ac.uk. Получено 25 мая 2023 года .
- ^ Digby GJ, Lober RM, Sethi PR, Lambert NA (ноябрь 2006 г.). «Некоторые гетеротримеры G -белка физически диссоциации в живых клетках» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (47): 17789–94. Bibcode : 2006pnas..10317789d . doi : 10.1073/pnas.0607116103 . PMC 1693825 . PMID 17095603 .
- ^ Khafizov K, Lattanzi G, Carloni P (июнь 2009 г.). «G -белок неактивные и активные формы, исследуемые с помощью методов моделирования». Белки . 75 (4): 919–30. doi : 10.1002/prot.22303 . PMID 19089952 . S2CID 23909821 .
- ^ Юэн Дж.В., Пун Л.С., Чан А.С., Ю Ф.В., Ло Рк, Вонг Й.Х. (июнь 2010 г.). «Активация STAT3 специфическими субъединицами Galpha и множественными димерами GbetAgamma». Международный журнал биохимии и клеточной биологии . 42 (6): 1052–9. doi : 10.1016/j.biocel.2010.03.017 . PMID 20348012 .
- ^ Sprang Sr, Chen Z, Du X (2007). «Структурная основа регуляции эффекторной регуляции и прекращения сигнала в гетеротримерных белках Gα». Структурная основа регулирования эффекторной и прекращения сигнала в гетеротримерных белках галфа . Достижения в химии белка. Тол. 74. С. 1–65. doi : 10.1016/s0065-3233 (07) 74001-9 . ISBN 978-0-12-034288-4 Полем PMID 17854654 .
- ^ Коул Ла (август 2010 г.). «Биологические функции молекул, связанных с HCG и HCG» . Репродуктивная биология и эндокринология . 8 (1): 102. doi : 10.1186/1477-7827-8-102 . PMC 2936313 . PMID 20735820 .
Внешние ссылки
[ редактировать ]- GTP-связывающие белки в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)
| Базы данных управления авторитетом : национальный |
|---|