С1ПР1
| С1ПР1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | S1PR1 , CD363, CHEDG1, D1S3362, ECGF1, EDG-1, EDG1, S1P1, рецептор сфингозин-1-фосфата 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 601974 ; МГИ : 1096355 ; Гомологен : 1071 ; Генные карты : S1PR1 ; ОМА : S1PR1 – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





Рецептор 1 сфингозин-1-фосфата (рецептор 1 S1P или S1PR1), также известный как ген 1 эндотелиальной дифференцировки (EDG1), представляет собой белок , который у человека кодируется S1PR1 геном . S1PR1 представляет собой рецептор, связанный с G-белком , который связывает биоактивную сигнальную молекулу сфингозин-1-фосфат (S1P). S1PR1 принадлежит к подсемейству рецепторов сфингозин-1-фосфата, состоящему из пяти членов (S1PR1-5). [ 5 ] S1PR1 первоначально был идентифицирован как обильный транскрипт в эндотелиальных клетках. [ 6 ] и он играет важную роль в регуляции структуры цитоскелета эндотелиальных клеток , миграции, образования капилляроподобной сети и созревания сосудов. [ 7 ] [ 8 ] Кроме того, передача сигналов S1PR1 важна в регуляции лимфоцитов . созревания, миграции и транспорта [ 9 ] [ 10 ]
Структура
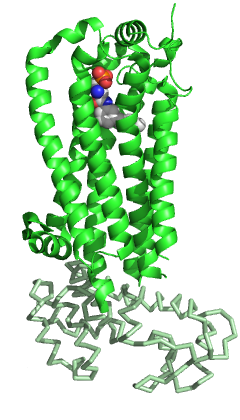
[ редактировать ]S1PR1, как и другие члены семейства GPCR, состоит из семи трансмембранных спиралей, собранных в структурно консервативный пучок. [ 5 ] Как и другие GPCR, во внеклеточной области S1PR1 состоит из трех петель: ECL1 между спиралями II и III, ECL2 между спиралями IV и V и ECL3 между спиралями VI и VII. По сравнению с другими представителями семейства S1PR1 имеет некоторые особенности. белка N-конец сворачивается в виде спиральной кепки над вершиной рецептора и, следовательно, ограничивает доступ лигандов к амфипатическому связывающему карману. Эта выраженная амфипатичность действительно согласуется с цвиттер-ионной природой S1P. Кроме того, спирали ECL1 и ECL2 плотно прилегают к N-концевой спирали, дополнительно перекрывая доступ лиганда из внеклеточного пространства. Аналоги S1P или S1P, вероятно, достигают связывающего кармана изнутри клеточной мембраны, а не из внеклеточного пространства, и могут происходить через отверстие между спиралями I и VII. По сравнению с другими GPCR эта область более открыта из-за различного расположения спиралей I и II по отношению к спирали III. [ 5 ] Эта окклюзия пространства доступа лиганда из внеклеточного пространства может также объяснить медленное насыщение связывания рецептора в присутствии избытка лиганда. [ 11 ]
Функция
[ редактировать ]Как и другие члены семейства GPCR, S1PR1 воспринимает свой лиганд снаружи клетки и активирует внутриклеточные сигнальные пути, которые в конечном итоге приводят к клеточным ответам. Сигнал передается посредством ассоциации рецептора с различными G-белками , что задействует ряд систем для последующей амплификации сигнала. [ 12 ]
Иммунная система
[ редактировать ]Торговля иммунными клетками
[ редактировать ]S1P и его рецепторы играют ключевую роль в регулировании движения иммунных клеток, образуя градиенты, которые направляют иммунные клетки между тканями и сосудами. S1PR1 играет решающую роль в обеспечении выхода Т-клеток из лимфоидных органов , в то время как изменения уровней S1P могут влиять на миграцию и расположение иммунных клеток в лимфоидных и нелимфоидных тканях во время воспаления или иммунного надзора. [ 13 ]
S1PR1, в первую очередь расположенный на клеточной мембране большинства лимфоцитов, связывается с распространенным лигандом S1P в кровотоке, способствуя выходу лимфоцитов из лимфоидных органов, позволяя им перемещаться к пораженным тканям. S1PR1 реагирует на градиент S1P между лимфоидными тканями (низкий S1P) и лимфой (высокий S1P), способствуя движению Т-клеток через эндотелиальный барьер. [ 14 ] Однако после активации Т-клеток в лимфоидных органах посредством передачи сигналов цитокинов и Т-клеточных рецепторов белковый кластер дифференцировки 69 ( CD69 ) экспрессируется и образует комплекс с S1PR1. Это взаимодействие, включающее трансмембранный домен CD69 и спираль-4 S1PR1, приводит к интернализации и деградации S1PR1, предотвращая связывание S1P и передачу сигналов ниже по ходу процесса. [ 15 ] Этот механизм приводит к временной задержке лимфоцитов в лимфатических органах, повышая шансы на успешную активацию лимфоцитов, особенно если первоначальный сигнал активации был слабым. При встрече с антигеном или стимуляции интерфероном I типа в лимфоидных органах экспрессия S1PR1 снижается за счет взаимодействия CD69 и подавления транскрипционного фактора Kruppel-подобного фактора 2. [ 16 ] Эффекторные Т-клетки в конечном итоге повторно экспрессируют S1PR1, выходя из лимфатического узла и проникая в периферические ткани. Однако повышенные уровни S1P в лимфоидных тканях из-за ингибирования лиазы S1P, воспаления или синтетических лигандов S1PR1, таких как FTY720 , могут блокировать выход Т-клеток путем рассеивания градиента S1P, индуцирования интернализации S1PR1 и усиления контактов эндотелиальных соединений для закрытия выходных портов. [ 16 ]
Регуляция иммунных клеток
[ редактировать ]Активация S1PR1 активно участвует в иммунных клеток регуляции и развитии . Рецептор сфингозин-1-фосфата 1 также участвует в иммуномодуляции и непосредственно участвует в подавлении врожденного иммунного ответа Т-клеток. [ 17 ] белка G в сочетании с S1PR1 достигаются разнообразные клеточные эффекты: Gαi В зависимости от и Gαo модулируют клеточное выживание, пролиферацию и подвижность ; Gα12 и и Gα13 модулируют ремоделирование цитоскелета формы клеток, а Gαq изменения модулирует некоторые эффекторные функции клеток. [ 12 ] Все внутриклеточные функции осуществляются посредством взаимодействия с Gαi и Gαo: эти два белка рекрутируют другие белки для последующей амплификации сигнала. [ 12 ] Основные нижестоящие эффекторные функции системы S1P-S1PR1 следующие:
- Сигнальный путь фосфатидилинозитол -3-киназы (PI3K) и липид-зависимой протеинкиназы B (PKB) увеличивает выживаемость лимфоцитов и других иммунных клеток путем ингибирования апоптоза .
- Фосфоинозитид-3-киназа (PI3K) и ГТФаза RAC ответственны за миграцию лимфоцитов и их взаимодействие с другими клетками или с поверхностями соединительной ткани. [ 12 ] Тимоциты с дефицитом S1PR1 не эмигрируют из тимуса, что приводит к увеличению количества зрелых тимоцитов в тимусе и медуллярной гиперплазии, а небольшое количество Т-клеток с дефицитом S1PR1 можно обнаружить в крови, лимфатических узлах , селезенке или нелимфоидных органах. в этих моделях мышей. [ 9 ] [ 10 ] Пролиферация иммунных клеток происходит за счет S1P-опосредованных сигналов через ГТФазу RAS и киназу, регулируемую внеклеточными сигналами (ERK). IV) Вызванное фосфолипазой C (PLC) увеличение внутриклеточного уровня кальция способствует секреции цитокинов и других иммунных медиаторов. [ 12 ]
Васкулогенез
[ редактировать ]S1PR1 является одним из основных рецепторов, ответственных за рост и развитие сосудов , по крайней мере, во время эмбриогенеза . [ 18 ] В эндотелиальных клетках сосудов связывание S1P с S1PR1 индуцирует миграцию, пролиферацию, выживание клеток и морфогенез в капилляроподобные структуры. [ 19 ] Более того, связывание S1P с S1PR1 участвует в формировании межклеточных адгезионных соединений , тем самым ингибируя параклеточную проницаемость растворенных веществ и макромолекул . [ 20 ] [ 21 ] Также было показано in vivo, что S1P взаимодействует с ангиогенными факторами, такими как FGF-2 и VEGF, индуцируя ангиогенез и созревание сосудов посредством S1PR1. [ 21 ] [ 22 ] показали, что мыши S1PR1-KO погибли во время развития из-за дефекта стабилизации сосудов, что указывает на то, что этот рецептор важен для развития сосудов. В заключение, несколько данных подтверждают, что S1P через S1PR1 является мощным регулятором роста и развития сосудов, по крайней мере, во время эмбриогенеза. [ 18 ]
Клиническое значение
[ редактировать ]Рак
[ редактировать ]S1PR1 участвует в подвижности раковых клеток при стимуляции S1P. Сигнальный путь включает RAC-CDC42 и коррелирует с активацией ERK1 и ERK2 . Путь RAC-CDC42 приводит к миграции клеток, тогда как путь ERK приводит к пролиферации и неоваскуляризации. [ 23 ] [ 24 ] продемонстрировали, что S1PR1 сильно индуцируется в эндотелиальных клетках во время опухолевого ангиогенеза, а миРНК против S1PR1 способна ингибировать ангиогенез и рост опухоли. S1PR1 также участвует в других типах рака: клетки фибросаркомы мигрируют при активации S1PR1 с помощью S1P по RAC1-CDC42-зависимому пути. [ 25 ] [ 26 ] а инвазия раковых клеток яичников включает S1PR1 или S1PR3 и мобилизацию кальция. [ 27 ]
Рассеянный склероз
[ редактировать ]S1PR1 участвует в рассеянном склерозе . Финголимод , препарат, который интернализует рецептор, одобрен в качестве модифицирующего заболевание средства при рассеянном склерозе. Существуют и другие модуляторы рецепторов сфингозин-1-фосфата . Ван Доорн и др. (2010) [ 28 ] наблюдали сильное увеличение экспрессии S1PR1 (и S1PR3) в гипертрофических астроцитах как в активных, так и в неактивных поражениях рассеянного склероза у пациентов по сравнению с незатронутыми пациентами.
Эволюция
[ редактировать ]Паралоги S1PR1 гена [ 29 ]
[ редактировать ]- С1ПР3
- С1ПР5
- С1ПР2
- С1ПР4
- LPAR1
- LPAR2
- LPAR3
- CNR1
- GPR6
- ГПР12
- MC4R
- MC5R
- CNR2
- MC3R
- Георадар3
- MC2R
- MC1R
- ГПР119
Взаимодействия
[ редактировать ]Было показано, что S1PR1 взаимодействует с рецептором 5-HT1A . [ 30 ] ГНАИ1 , [ 31 ] и ГНАИ3 . [ 31 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000170989 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: Ensembl, выпуск 89: ENSMUSG00000045092 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б с Хэнсон М.А., Рот С.Б., Джо Э., Гриффит М.Т., Скотт Ф.Л., Рейнхарт Г. и др. (февраль 2012 г.). «Кристаллическая структура рецептора, связанного с липидом G-белком» . Наука . 335 (6070): 851–855. Бибкод : 2012Sci...335..851H . дои : 10.1126/science.1215904 . ПМЦ 3338336 . ПМИД 22344443 .
- ^ Хла Т, Масиаг Т (июнь 1990 г.). «Обильный транскрипт, индуцируемый при дифференцировке эндотелиальных клеток человека, кодирует полипептид со структурным сходством с рецепторами, связанными с G-белком» . Журнал биологической химии . 265 (16): 9308–9313. дои : 10.1016/S0021-9258(19)38849-0 . ПМИД 2160972 .
- ^ Ли М.Дж., Ван Броклин Дж.Р., Тангада С., Лю Ч.Х., Хэнд А.Р., Мензелеев Р. и др. (март 1998 г.). «Сфингозин-1-фосфат как лиганд для рецептора, связанного с G-белком EDG-1». Наука . 279 (5356): 1552–1555. Бибкод : 1998Sci...279.1552L . дои : 10.1126/science.279.5356.1552 . ПМИД 9488656 .
- ^ Лю Ч., Тангада С., Ли М.Дж., Ван Броклин-младший, Шпигель С., Хла Т. (апрель 1999 г.). «Индуцированный лигандом трафик сфингозин-1-фосфатного рецептора EDG-1» . Молекулярная биология клетки . 10 (4): 1179–1190. дои : 10.1091/mbc.10.4.1179 . ПМК 25247 . ПМИД 10198065 .
- ^ Jump up to: а б Альенде М.Л., Драйер Дж.Л., Мандала С., Пройа Р.Л. (апрель 2004 г.). «Экспрессия рецептора сфингозин-1-фосфата, S1P1, на Т-клетках контролирует эмиграцию тимуса» . Журнал биологической химии . 279 (15): 15396–15401. дои : 10.1074/jbc.M314291200 . ПМИД 14732704 .
- ^ Jump up to: а б Матлубиан М., Ло К.Г., Синамон Г., Леснески М.Дж., Сюй Ю., Бринкманн В. и др. (январь 2004 г.). «Выход лимфоцитов из тимуса и периферических лимфоидных органов зависит от рецептора S1P 1». Природа . 427 (6972): 355–360. Бибкод : 2004Natur.427..355M . дои : 10.1038/nature02284 . ПМИД 14737169 . S2CID 4371877 .
- ^ Розен Х., Гонсалес-Кабрера П.Дж., Санна М.Г., Браун С. (2009). «Передача сигналов рецептора сфингозин-1-фосфата». Ежегодный обзор биохимии . 78 : 743–768. doi : 10.1146/annurev.biochem.78.072407.103733 . ПМИД 19231986 .
- ^ Jump up to: а б с д и Розен Х. (сентябрь 2005 г.). «Химические подходы к лизофосфолипидным рецепторам». Простагландины и другие липидные медиаторы . 77 (1–4): 179–184. doi : 10.1016/j.prostaglandins.2004.09.011 . ПМИД 16099402 .
- ^ Гаррис К.С., Блахо В.А., Хла Т., Хан М.Х. (июль 2014 г.). «Передача сигнала сфингозин-1-фосфатного рецептора 1 в Т-клетках: торговля и не только» . Иммунология . 142 (3): 347–353. дои : 10.1111/imm.12272 . ПМК 4080950 . ПМИД 24597601 .
- ^ Чи Х (январь 2011 г.). «Сфингозин-1-фосфат и иммунная регуляция: торговля и не только» . Тенденции в фармакологических науках . 32 (1): 16–24. дои : 10.1016/j.tips.2010.11.002 . ПМК 3017656 . ПМИД 21159389 .
- ^ Сибриан Д., Санчес-Мадрид Ф (июнь 2017 г.). «CD69: от маркера активации к метаболическому привратнику» . Европейский журнал иммунологии . 47 (6): 946–953. дои : 10.1002/eji.201646837 . ПМЦ 6485631 . ПМИД 28475283 .
- ^ Jump up to: а б Ривера Дж., Пройа Р.Л., Оливера А. (октябрь 2008 г.). «Альянс сфингозин-1-фосфата и его рецепторов в иммунитете» . Обзоры природы. Иммунология . 8 (10): 753–763. дои : 10.1038/nri2400 . ПМК 2600775 . ПМИД 18787560 .
- ^ Шарма Н., Ахаде А.С., Кадри А. (апрель 2013 г.). «Сфингозин-1-фосфат подавляет TLR-индуцированную секрецию CXCL8 из Т-клеток человека». Журнал биологии лейкоцитов . 93 (4): 521–528. дои : 10.1189/jlb.0712328 . ПМИД 23345392 . S2CID 21897008 .
- ^ Jump up to: а б Чае С.С., Пайк Дж.Х., Альенде М.Л., Пройа Р.Л., Хла Т (апрель 2004 г.). «Регуляция развития конечностей с помощью рецептора сфингозин-1-фосфата S1p1/EDG-1 происходит через ось гипоксия/VEGF» . Биология развития . 268 (2): 441–447. дои : 10.1016/j.ydbio.2004.01.001 . ПМИД 15063179 .
- ^ Ли М.Дж., Тангада С., Клаффи К.П., Анселлин Н., Лю Ч., Клюк М. и др. (октябрь 1999 г.). «Сборка и морфогенез слипчивых эндотелиальных клеток сосудов, индуцированные сфингозин-1-фосфатом» . Клетка . 99 (3): 301–312. doi : 10.1016/S0092-8674(00)81661-X . ПМИД 10555146 . S2CID 1126846 .
- ^ Санчес Т., Эстрада-Эрнандес Т., Пайк Дж.Х., Ву М.Т., Венкатараман К., Бринкманн В. и др. (ноябрь 2003 г.). «Фосфорилирование и действие иммуномодулятора FTY720 ингибируют проницаемость сосудов, вызванную фактором роста эндотелиальных клеток сосудов» . Журнал биологической химии . 278 (47): 47281–47290. дои : 10.1074/jbc.M306896200 . ПМИД 12954648 .
- ^ Jump up to: а б Гарсия Дж.Г., Лю Ф., Верин А.Д., Бирюкова А., Дечерт М.А., Гертоффер В.Т. и др. (сентябрь 2001 г.). «Сфингозин-1-фосфат способствует целостности барьера эндотелиальных клеток за счет Edg-зависимой перестройки цитоскелета» . Журнал клинических исследований . 108 (5): 689–701. дои : 10.1172/JCI12450 . ПМК 209379 . ПМИД 11544274 .
- ^ Лю Ю, Вада Р., Ямасита Т, Ми Ю, Дэн СХ, Хобсон Дж. П. и др. (октябрь 2000 г.). «Edg-1, связанный с G-белком рецептор сфингозин-1-фосфата, необходим для созревания сосудов» . Журнал клинических исследований . 106 (8): 951–961. дои : 10.1172/JCI10905 . ПМК 314347 . ПМИД 11032855 .
- ^ Пайн, Нью-Джерси, Пайн С. (июль 2010 г.). «Сфингозин-1-фосфат и рак» (PDF) . Обзоры природы. Рак . 10 (7): 489–503. дои : 10.1038/nrc2875 . ПМИД 20555359 . S2CID 32955497 .
- ^ Чае С.С., Пайк Дж.Х., Фюрно Х., Хла Т. (октябрь 2004 г.). «Потребность в рецепторе сфингозин-1-фосфата-1 при опухолевом ангиогенезе, продемонстрированная с помощью РНК-интерференции in vivo» . Журнал клинических исследований . 114 (8): 1082–1089. дои : 10.1172/JCI22716 . ПМК 522258 . ПМИД 15489955 .
- ^ Фишер К.Э., Поп А., Ко В., Антис Нью-Джерси, Сондерс В.Б., Дэвис Дж.Э. (декабрь 2006 г.). «Инвазия опухолевых клеток в коллагеновые матрицы требует координации передачи сигналов, индуцированной липидными агонистами G-белком, и матриксной металлопротеиназы-1 мембранного типа» . Молекулярный рак . 5:69 . дои : 10.1186/1476-4598-5-69 . ПМК 1762019 . ПМИД 17156449 .
- ^ Ньялендо С., Мишо М., Болье Е., Роги С., Мерфи Г., Гинграс Д. и др. (май 2007 г.). «Src-зависимое фосфорилирование мембранной матриксной металлопротеиназы I типа на цитоплазматическом тирозине 573: роль в миграции эндотелиальных и опухолевых клеток» . Журнал биологической химии . 282 (21): 15690–15699. дои : 10.1074/jbc.M608045200 . ПМИД 17389600 .
- ^ Пак К.С., Ким М.К., Ли Х.И., Ким С.Д., Ли С.И., Ким Дж.М. и др. (апрель 2007 г.). «S1P стимулирует хемотаксическую миграцию и инвазию в клетки рака яичников OVCAR3». Связь с биохимическими и биофизическими исследованиями . 356 (1): 239–244. дои : 10.1016/j.bbrc.2007.02.112 . ПМИД 17349972 .
- ^ Ван Доорн Р., Ван Хорссен Дж., Верзийл Д., Витте М., Ронкен Э., Ван Хет Хоф Б. и др. (сентябрь 2010 г.). «Рецепторы сфингозин-1-фосфата 1 и 3 активируются при поражениях рассеянного склероза». Глия . 58 (12): 1465–1476. дои : 10.1002/glia.21021 . ПМИД 20648639 . S2CID 26000783 .
- ^ «GeneCards®: База данных генов человека» .
- ^ Салим К., Фентон Т., Бача Дж., Уриен-Родригес Х., Боннерт Т., Скайннер Х.А. и др. (май 2002 г.). «Олигомеризация рецепторов, связанных с G-белком, показанная с помощью селективной коиммунопреципитации» . Журнал биологической химии . 277 (18): 15482–15485. дои : 10.1074/jbc.M201539200 . ПМИД 11854302 .
- ^ Jump up to: а б Ли М.Дж., Эванс М., Хла Т. (май 1996 г.). «Индуцибельный рецептор edg-1, связанный с G-белком, передает сигнал по пути G(i)/митоген-активируемой протеинкиназы» . Журнал биологической химии . 271 (19): 11272–11279. дои : 10.1074/jbc.271.19.11272 . ПМИД 8626678 .
Внешние ссылки
[ редактировать ]- «Лизофосфолипидные рецепторы: S 1 P 1 » . База данных IUPHAR по рецепторам и ионным каналам . Международный союз фундаментальной и клинической фармакологии. Архивировано из оригинала 7 января 2015 года . Проверено 5 декабря 2008 г.
- Лизофосфолипид + рецепторы Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .