ВИЧ
| Вирусы иммунодефицита человека | |
|---|---|

| |
| Сканирующая электронная микрофотография ВИЧ-1 (в зеленом) подающении от культивируемых лимфоцитов . Многочисленные круглые неровности на клеточной поверхности представляют участки сборки и начинание вирионов. | |
| Научная классификация | |
| (не вмешательство): | Вирус |
| Область : | Рибовирия |
| Королевство: | Pararnavirae |
| Филум: | Art Verviricota |
| Сорт: | Revtraviricetes |
| Заказ: | Ортервиралес |
| Семья: | Retroviridae |
| Подсемейство: | Орторетровирина |
| Род: | Лентивирус |
| Группы включены | |
| Другие лентивирусы | |
Вирусы иммунодефицита человека ( ВИЧ ) представляют собой два вида лентивируса (подгруппа ретровируса ), которые заражают людей. Со временем они вызывают синдром приобретенного иммунодефицита (СПИД), [ 1 ] [ 2 ] Условие, в котором прогрессивный сбой иммунной системы опасно для жизни оппортунистические инфекции и рак . позволяет процветать [ 3 ] Без лечения среднее время выживаемости после заражения ВИЧ оценивается от 9 до 11 лет, в зависимости от подтипа ВИЧ . [ 4 ]
В большинстве случаев ВИЧ является инфекцией, передаваемой половым путем , и возникает путем контакта с или переносом крови , предварительной эякуляции , спермы и влагалищных жидкостей . [ 5 ] [ 6 ] Несексуальная передача может происходить от зараженной матери до ее младенца во время беременности , во время родов в результате воздействия ее крови или влагалищной жидкости и через грудное молоко . [ 7 ] [ 8 ] [ 9 ] [ 10 ] В этих телесных жидкостях ВИЧ присутствуют как частицы свободного вируса , так и вирус в инфицированных иммунных клетках . Исследования показали (как для однополых, так и противоположных парами), что ВИЧ не заражен во время полового акта без презерватива, если ВИЧ-позитивный партнер имеет неизменно неопределяемую вирусную нагрузку . [ 5 ] [ 6 ]
ВИЧ -инфицирует жизненно важные клетки в иммунной системе человека, таких как TPER T -клетки (в частности CD4 + Т -клетки), макрофаги и дендритные клетки . [ 11 ] ВИЧ -инфекция приводит к низким уровням CD4 + Т -клетки через ряд механизмов, включая пироптоз абортивно инфицированных Т -клеток, [ 12 ] апоптоз неинфицированных клеток свидетеля, [ 13 ] Прямое вирусное убийство инфицированных клеток и убийство инфицированного CD4 + Т -клетки CD8 + цитотоксические лимфоциты , которые распознают инфицированные клетки. [ 14 ] Когда CD4 + Числа Т-клеток снижаются ниже критического уровня, клеточный опосредованный иммунитет теряется, и тело постепенно становится более восприимчивым к оппортунистическим инфекциям, что приводит к развитию СПИДа.
Вирусология
| Разновидность | Вирулентность | Инфекционность | Распространенность | Предполагаемое происхождение |
|---|---|---|---|---|
| ВИЧ-1 | Высокий | Высокий | Глобальный | Общий шимпанзе |
| ВИЧ-2 | Ниже | Низкий | Западная Африка | Сажи Мангаби |
Классификация
ВИЧ является членом рода лентивируса , [ 15 ] Часть семейства Retroviridae . [ 16 ] Лентивирусы имеют много морфологий и биологических общих свойств. Многие виды заражены лентивирусами, которые характерны ответственны за длительные заболевания с длительным периодом инкубации . [ 17 ] Лентивирусы передаются в виде одноцепочечных , положительных , охваченных РНК -вирусов . После входа в клетку-мишени вирусной РНК геном превращается (обратная транскрибируется) в двухцепочечную ДНК , кодируемым вирусным ферментом, обратной транскриптазой , который транспортируется вместе с вирусным геномом в вирусной частице. Полученная вирусная ДНК затем импортируется в клеточное ядро и интегрируется в клеточную ДНК с помощью кодируемых вирусами фермента, интегразы и соавторов- хозяев . [ 18 ] После интеграции вирус может стать скрытым , позволяя вирусу и его клетке -хозяину избежать обнаружения иммунной системой в течение неопределенного времени. [ 19 ] Вирус может оставаться бездействующим в организме человека в течение десяти лет после первичной инфекции; В течение этого периода вирус не вызывает симптомов. Альтернативно, интегрированная вирусная ДНК может быть транскрибирована , продуцируя новые геномы РНК и вирусные белки с использованием клеточных ресурсов хозяина, которые упаковываются и высвобождаются из клетки в качестве новых вирусных частиц, которые начнут цикл репликации заново.
Были охарактеризованы два типа ВИЧ: ВИЧ-1 и ВИЧ-2. ВИЧ-1-это вирус, который был первоначально обнаружен и называется как вирус, связанный с лимфаденопатией (LAV), так и Т-лимфотропный вирус 3 человека (HTLV-III). ВИЧ-1 более вирулентный и более инфекционный, чем ВИЧ-2, [ 20 ] и является причиной большинства ВИЧ -инфекций во всем мире. Более низкая инфекционность ВИЧ-2, по сравнению с ВИЧ-1, подразумевает, что меньшее количество лиц, подвергшихся воздействию ВИЧ-2, будет инфицировано за воздействие. Из-за его относительно плохой способности к передаче ВИЧ-2 в значительной степени ограничен Западной Африкой . [ 21 ]
Структура и геном

ВИЧ по структуре схож с другими ретровирусами. Это примерно сферическое [ 22 ] с диаметром около 120 нм , примерно в 100 000 раз меньше по объему, чем эритроцитная клетка . [ 23 ] Он состоит из двух копий одноцепочечной состоящий РНК вируса, , которая кодирует девять генов заключенные в коническую капсид, из 2000 копий вирусного белка p24 . [ 24 ] Одноцепочечная РНК тесно связана с нуклеокапсидными белками, P7 и ферментами, необходимыми для развития вириона, такого как обратная транскриптаза , протеазы , рибонуклеаза и интеграция . Матрица, состоящая из вирусного белка P17, окружает капсид, обеспечивая целостность частицы вириона. [ 24 ]
Это, в свою очередь, окружено вирусной оболочкой , которая состоит из липидного бислоя, взятого из мембраны клетки -хозяина человека, когда вновь образованные полосы частиц вируса из клетки. Вирусная оболочка содержит белки из клетки -хозяина и относительно мало копий белка огибальной части ВИЧ, [ 24 ] который состоит из крышки из трех молекул, известных как гликопротеин (GP) 120 , и стебель, состоящий из трех молекул GP41 , которые закрепляют структуру в вирусную оболочку. [ 25 ] [ 26 ] Белок оболочки, кодируемый геном ENV ВИЧ , позволяет вирусу прикрепляться к клеткам -мишеням и сливать вирусную оболочку с мембраной клетки -мишени , высвобождающей содержание вируса в клетку и инициируя инфекционный цикл. [ 25 ]

В качестве единственного вирусного белка на поверхности вируса белок оболочки является основной мишенью для усилий против ВИЧ . [ 27 ] Более половины массы тримерного шипа конверта представляет собой n-связанные гликаны . Плотность высока, так как гликаны защищают основной вирусный белок от нейтрализации антителами. Это одна из наиболее плотно известных гликозилированных молекул, и плотность достаточно высока, чтобы предотвратить нормальный процесс созревания гликанов во время биогенеза в эндоплазматическом аппарате и гольджи. [ 28 ] [ 29 ] Таким образом, большинство гликанов останавливаются как незрелые «гликаны с высокой мужской», которые обычно не присутствуют на гликопротеинах человека, которые секретируются или присутствуют на клеточной поверхности. [ 30 ] Необычная обработка и высокая плотность означают, что почти все широко нейтрализующие антитела, которые до сих пор были идентифицированы (от подмножества пациентов, которые были инфицированы в течение многих месяцев до многих лет), с или адаптированы, чтобы справиться с этими гликанами о конверте. [ 31 ]
Молекулярная структура вирусного всплеска теперь определяется рентгеновской кристаллографией [ 32 ] и криогенная электронная микроскопия . [ 33 ] Эти достижения в области структурной биологии стали возможными благодаря развитию стабильных рекомбинантных форм вирусного пика путем введения межсубунитной дисульфидной связи и изолейцина с пролина мутацией ( радикальная замена аминокислоты) в GP41. [ 34 ] Так называемые тримеры SOSIP не только воспроизводят антигенные свойства нативного вирусного всплеска, но также демонстрируют ту же степень незрелых гликанов, что и на нативном вирусе. [ 35 ] Рекомбинантные тримерные вирусные пики являются многообещающими кандидатами на вакцину, поскольку они показывают меньше нететрализирующих эпитопов , чем рекомбинантный мономерный GP120, которые действуют для подавления иммунного ответа на эпитопы-мишени. [ 36 ]

The RNA genome consists of at least seven structural landmarks (LTR, TAR, RRE, PE, SLIP, CRS, and INS), and nine genes (gag, pol, and env, tat, rev, nef, vif, vpr, vpu, and sometimes a tenth tev, which is a fusion of tat, env and rev), encoding 19 proteins. Three of these genes, gag, pol, and env, contain information needed to make the structural proteins for new virus particles.[24] For example, env codes for a protein called gp160 that is cut in two by a cellular protease to form gp120 and gp41. The six remaining genes, tat, rev, nef, vif, vpr, and vpu (or vpx in the case of HIV-2), are regulatory genes for proteins that control the ability of HIV to infect cells, produce new copies of virus (replicate), or cause disease.[24]
The two tat proteins (p16 and p14) are transcriptional transactivators for the LTR promoter acting by binding the TAR RNA element. The TAR may also be processed into microRNAs that regulate the apoptosis genes ERCC1 and IER3.[37][38] The rev protein (p19) is involved in shuttling RNAs from the nucleus and the cytoplasm by binding to the RRE RNA element. The vif protein (p23) prevents the action of APOBEC3G (a cellular protein that deaminates cytidine to uridine in the single-stranded viral DNA and/or interferes with reverse transcription[39]). The vpr protein (p14) arrests cell division at G2/M. The nef protein (p27) down-regulates CD4 (the major viral receptor), as well as the MHC class I and class II molecules.[40][41][42]
Nef also interacts with SH3 domains. The vpu protein (p16) influences the release of new virus particles from infected cells.[24] The ends of each strand of HIV RNA contain an RNA sequence called a long terminal repeat (LTR). Regions in the LTR act as switches to control production of new viruses and can be triggered by proteins from either HIV or the host cell. The Psi element is involved in viral genome packaging and recognized by gag and rev proteins. The SLIP element (TTTTTT) is involved in the frameshift in the gag-pol reading frame required to make functional pol.[24]
Tropism

The term viral tropism refers to the cell types a virus infects. HIV can infect a variety of immune cells such as CD4+ T cells, macrophages, and microglial cells. HIV-1 entry to macrophages and CD4+ T cells is mediated through interaction of the virion envelope glycoproteins (gp120) with the CD4 molecule on the target cells' membrane and also with chemokine co-receptors.[25][43]
Macrophage-tropic (M-tropic) strains of HIV-1, or non-syncytia-inducing strains (NSI; now called R5 viruses[44]) use the β-chemokine receptor, CCR5, for entry and are thus able to replicate in both macrophages and CD4+ T cells.[45] This CCR5 co-receptor is used by almost all primary HIV-1 isolates regardless of viral genetic subtype. Indeed, macrophages play a key role in several critical aspects of HIV infection. They appear to be the first cells infected by HIV and perhaps the source of HIV production when CD4+ cells become depleted in the patient. Macrophages and microglial cells are the cells infected by HIV in the central nervous system. In the tonsils and adenoids of HIV-infected patients, macrophages fuse into multinucleated giant cells that produce huge amounts of virus.
T-tropic strains of HIV-1, or syncytia-inducing strains (SI; now called X4 viruses[44]) replicate in primary CD4+ T cells as well as in macrophages and use the α-chemokine receptor, CXCR4, for entry.[45][46][47]
Dual-tropic HIV-1 strains are thought to be transitional strains of HIV-1 and thus are able to use both CCR5 and CXCR4 as co-receptors for viral entry.
The α-chemokine SDF-1, a ligand for CXCR4, suppresses replication of T-tropic HIV-1 isolates. It does this by down-regulating the expression of CXCR4 on the surface of HIV target cells. M-tropic HIV-1 isolates that use only the CCR5 receptor are termed R5; those that use only CXCR4 are termed X4, and those that use both, X4R5. However, the use of co-receptors alone does not explain viral tropism, as not all R5 viruses are able to use CCR5 on macrophages for a productive infection[45] and HIV can also infect a subtype of myeloid dendritic cells,[48] which probably constitute a reservoir that maintains infection when CD4+ T cell numbers have declined to extremely low levels.
Some people are resistant to certain strains of HIV.[49] For example, people with the CCR5-Δ32 mutation are resistant to infection by the R5 virus, as the mutation leaves HIV unable to bind to this co-receptor, reducing its ability to infect target cells.
Sexual intercourse is the major mode of HIV transmission. Both X4 and R5 HIV are present in the seminal fluid, which enables the virus to be transmitted from a male to his sexual partner. The virions can then infect numerous cellular targets and disseminate into the whole organism. However, a selection process[further explanation needed] leads to a predominant transmission of the R5 virus through this pathway.[50][51][52] In patients infected with subtype B HIV-1, there is often a co-receptor switch in late-stage disease and T-tropic variants that can infect a variety of T cells through CXCR4.[53] These variants then replicate more aggressively with heightened virulence that causes rapid T cell depletion, immune system collapse, and opportunistic infections that mark the advent of AIDS.[54] HIV-positive patients acquire an enormously broad spectrum of opportunistic infections, which was particularly problematic prior to the onset of HAART therapies; however, the same infections are reported among HIV-infected patients examined post-mortem following the onset of antiretroviral therapies.[3] Thus, during the course of infection, viral adaptation to the use of CXCR4 instead of CCR5 may be a key step in the progression to AIDS. A number of studies with subtype B-infected individuals have determined that between 40 and 50 percent of AIDS patients can harbour viruses of the SI and, it is presumed, the X4 phenotypes.[55][56]
HIV-2 is much less pathogenic than HIV-1 and is restricted in its worldwide distribution to West Africa. The adoption of "accessory genes" by HIV-2 and its more promiscuous pattern of co-receptor usage (including CD4-independence) may assist the virus in its adaptation to avoid innate restriction factors present in host cells. Adaptation to use normal cellular machinery to enable transmission and productive infection has also aided the establishment of HIV-2 replication in humans. A survival strategy for any infectious agent is not to kill its host, but ultimately become a commensal organism. Having achieved a low pathogenicity, over time, variants that are more successful at transmission will be selected.[57]
Replication cycle
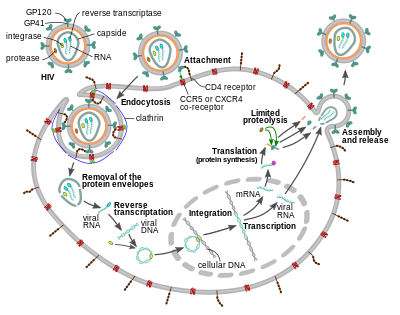
Entry to the cell

The HIV virion enters macrophages and CD4+ T cells by the adsorption of glycoproteins on its surface to receptors on the target cell followed by fusion of the viral envelope with the target cell membrane and the release of the HIV capsid into the cell.[58][59]
Entry to the cell begins through interaction of the trimeric envelope complex (gp160 spike) on the HIV viral envelope and both CD4 and a chemokine co-receptor (generally either CCR5 or CXCR4, but others are known to interact) on the target cell surface.[58][59] Gp120 binds to integrin α4β7 activating LFA-1, the central integrin involved in the establishment of virological synapses, which facilitate efficient cell-to-cell spreading of HIV-1.[60] The gp160 spike contains binding domains for both CD4 and chemokine receptors.[58][59]
The first step in fusion involves the high-affinity attachment of the CD4 binding domains of gp120 to CD4. Once gp120 is bound with the CD4 protein, the envelope complex undergoes a structural change, exposing the chemokine receptor binding domains of gp120 and allowing them to interact with the target chemokine receptor.[58][59] This allows for a more stable two-pronged attachment, which allows the N-terminal fusion peptide gp41 to penetrate the cell membrane.[58][59] Repeat sequences in gp41, HR1, and HR2 then interact, causing the collapse of the extracellular portion of gp41 into a hairpin shape. This loop structure brings the virus and cell membranes close together, allowing fusion of the membranes and subsequent entry of the viral capsid.[58][59]
After HIV has bound to the target cell, the HIV RNA and various enzymes, including reverse transcriptase, integrase, ribonuclease, and protease, are injected into the cell.[58][failed verification] During the microtubule-based transport to the nucleus, the viral single-strand RNA genome is transcribed into double-strand DNA, which is then integrated into a host chromosome.
HIV can infect dendritic cells (DCs) by this CD4-CCR5 route, but another route using mannose-specific C-type lectin receptors such as DC-SIGN can also be used.[61] DCs are one of the first cells encountered by the virus during sexual transmission. They are currently thought to play an important role by transmitting HIV to T cells when the virus is captured in the mucosa by DCs.[61] The presence of FEZ-1, which occurs naturally in neurons, is believed to prevent the infection of cells by HIV.[62]

HIV-1 entry, as well as entry of many other retroviruses, has long been believed to occur exclusively at the plasma membrane. More recently, however, productive infection by pH-independent, clathrin-mediated endocytosis of HIV-1 has also been reported and was recently suggested to constitute the only route of productive entry.[63][64][65][66][67]
Replication and transcription
Shortly after the viral capsid enters the cell, an enzyme called reverse transcriptase liberates the positive-sense single-stranded RNA genome from the attached viral proteins and copies it into a complementary DNA (cDNA) molecule.[68] The process of reverse transcription is extremely error-prone, and the resulting mutations may cause drug resistance or allow the virus to evade the body's immune system. The reverse transcriptase also has ribonuclease activity that degrades the viral RNA during the synthesis of cDNA, as well as DNA-dependent DNA polymerase activity that creates a sense DNA from the antisense cDNA.[69] Together, the cDNA and its complement form a double-stranded viral DNA that is then transported into the cell nucleus. The integration of the viral DNA into the host cell's genome is carried out by another viral enzyme called integrase.[68]
The integrated viral DNA may then lie dormant, in the latent stage of HIV infection.[68] To actively produce the virus, certain cellular transcription factors need to be present, the most important of which is NF-κB (nuclear factor kappa B), which is upregulated when T cells become activated.[70] This means that those cells most likely to be targeted, entered and subsequently killed by HIV are those actively fighting infection.
During viral replication, the integrated DNA provirus is transcribed into RNA. The full-length genomic RNAs (gRNA) can be packaged into new viral particles in a pseudodiploid form. The selectivity in the packaging is explained by the structural properties of the dimeric conformer of the gRNA. The gRNA dimer is characterized by a tandem three-way junction within the gRNA monomer, in which the SD and AUG hairpins, responsible for splicing and translation respectively, are sequestered and the DIS (dimerization initiation signal) hairpin is exposed. The formation of the gRNA dimer is mediated by a 'kissing' interaction between the DIS hairpin loops of the gRNA monomers. At the same time, certain guanosine residues in the gRNA are made available for binding of the nucleocapsid (NC) protein leading to the subsequent virion assembly.[71] The labile gRNA dimer has been also reported to achieve a more stable conformation following the NC binding, in which both the DIS and the U5:AUG regions of the gRNA participate in extensive base pairing.[72]
RNA can also be processed to produce mature messenger RNAs (mRNAs). In most cases, this processing involves RNA splicing to produce mRNAs that are shorter than the full-length genome. Which part of the RNA is removed during RNA splicing determines which of the HIV protein-coding sequences is translated.[73]
Mature HIV mRNAs are exported from the nucleus into the cytoplasm, where they are translated to produce HIV proteins, including Rev. As the newly produced Rev protein is produced it moves to the nucleus, where it binds to full-length, unspliced copies of virus RNAs and allows them to leave the nucleus.[74] Some of these full-length RNAs function as mRNAs that are translated to produce the structural proteins Gag and Env. Gag proteins bind to copies of the virus RNA genome to package them into new virus particles.[75] HIV-1 and HIV-2 appear to package their RNA differently.[76][77] HIV-1 will bind to any appropriate RNA.[78] HIV-2 will preferentially bind to the mRNA that was used to create the Gag protein itself.[79]
Recombination
Two RNA genomes are encapsidated in each HIV-1 particle (see Structure and genome of HIV). Upon infection and replication catalyzed by reverse transcriptase, recombination between the two genomes can occur.[80][81] Recombination occurs as the single-strand, positive-sense RNA genomes are reverse transcribed to form DNA. During reverse transcription, the nascent DNA can switch multiple times between the two copies of the viral RNA. This form of recombination is known as copy-choice. Recombination events may occur throughout the genome. Anywhere from two to 20 recombination events per genome may occur at each replication cycle, and these events can rapidly shuffle the genetic information that is transmitted from parental to progeny genomes.[81]
Viral recombination produces genetic variation that likely contributes to the evolution of resistance to anti-retroviral therapy.[82] Recombination may also contribute, in principle, to overcoming the immune defenses of the host. Yet, for the adaptive advantages of genetic variation to be realized, the two viral genomes packaged in individual infecting virus particles need to have arisen from separate progenitor parental viruses of differing genetic constitution. It is unknown how often such mixed packaging occurs under natural conditions.[83]
Bonhoeffer et al.[84] suggested that template switching by reverse transcriptase acts as a repair process to deal with breaks in the single-stranded RNA genome. In addition, Hu and Temin[80] suggested that recombination is an adaptation for repair of damage in the RNA genomes. Strand switching (copy-choice recombination) by reverse transcriptase could generate an undamaged copy of genomic DNA from two damaged single-stranded RNA genome copies. This view of the adaptive benefit of recombination in HIV could explain why each HIV particle contains two complete genomes, rather than one. Furthermore, the view that recombination is a repair process implies that the benefit of repair can occur at each replication cycle, and that this benefit can be realized whether or not the two genomes differ genetically. On the view that recombination in HIV is a repair process, the generation of recombinational variation would be a consequence, but not the cause of, the evolution of template switching.[84]
HIV-1 infection causes chronic inflammation and production of reactive oxygen species.[85] Thus, the HIV genome may be vulnerable to oxidative damage, including breaks in the single-stranded RNA. For HIV, as well as for viruses in general, successful infection depends on overcoming host defense strategies that often include production of genome-damaging reactive oxygen species. Thus, Michod et al.[86] suggested that recombination by viruses is an adaptation for repair of genome damage, and that recombinational variation is a byproduct that may provide a separate benefit.
Assembly and release
The final step of the viral cycle, assembly of new HIV-1 virions, begins at the plasma membrane of the host cell. The Env polyprotein (gp160) goes through the endoplasmic reticulum and is transported to the Golgi apparatus where it is cleaved by furin resulting in the two HIV envelope glycoproteins, gp41 and gp120.[87] These are transported to the plasma membrane of the host cell where gp41 anchors gp120 to the membrane of the infected cell. The Gag (p55) and Gag-Pol (p160) polyproteins also associate with the inner surface of the plasma membrane along with the HIV genomic RNA as the forming virion begins to bud from the host cell. The budded virion is still immature as the gag polyproteins still need to be cleaved into the actual matrix, capsid and nucleocapsid proteins. This cleavage is mediated by the packaged viral protease and can be inhibited by antiretroviral drugs of the protease inhibitor class. The various structural components then assemble to produce a mature HIV virion.[88] Only mature virions are then able to infect another cell.
Spread within the body

The classical process of infection of a cell by a virion can be called "cell-free spread" to distinguish it from a more recently recognized process called "cell-to-cell spread".[89] In cell-free spread (see figure), virus particles bud from an infected T cell, enter the blood or extracellular fluid and then infect another T cell following a chance encounter.[89] HIV can also disseminate by direct transmission from one cell to another by a process of cell-to-cell spread, for which two pathways have been described. Firstly, an infected T cell can transmit virus directly to a target T cell via a virological synapse.[60][90] Secondly, an antigen-presenting cell (APC), such as a macrophage or dendritic cell, can transmit HIV to T cells by a process that either involves productive infection (in the case of macrophages) or capture and transfer of virions in trans (in the case of dendritic cells).[91] Whichever pathway is used, infection by cell-to-cell transfer is reported to be much more efficient than cell-free virus spread.[92] A number of factors contribute to this increased efficiency, including polarised virus budding towards the site of cell-to-cell contact, close apposition of cells, which minimizes fluid-phase diffusion of virions, and clustering of HIV entry receptors on the target cell towards the contact zone.[90] Cell-to-cell spread is thought to be particularly important in lymphoid tissues, where CD4+ T cells are densely packed and likely to interact frequently.[89] Intravital imaging studies have supported the concept of the HIV virological synapse in vivo.[93] The many dissemination mechanisms available to HIV contribute to the virus' ongoing replication in spite of anti-retroviral therapies.[89][94]
Genetic variability

HIV differs from many viruses in that it has very high genetic variability. This diversity is a result of its fast replication cycle, with the generation of about 1010 virions every day, coupled with a high mutation rate of approximately 3 x 10−5 per nucleotide base per cycle of replication and recombinogenic properties of reverse transcriptase.[95][96][97]
This complex scenario leads to the generation of many variants of HIV in a single infected patient in the course of one day.[95] This variability is compounded when a single cell is simultaneously infected by two or more different strains of HIV. When simultaneous infection occurs, the genome of progeny virions may be composed of RNA strands from two different strains. This hybrid virion then infects a new cell where it undergoes replication. As this happens, the reverse transcriptase, by jumping back and forth between the two different RNA templates, will generate a newly synthesized retroviral DNA sequence that is a recombinant between the two parental genomes.[95] This recombination is most obvious when it occurs between subtypes.[95]
The closely related simian immunodeficiency virus (SIV) has evolved into many strains, classified by the natural host species. SIV strains of the African green monkey (SIVagm) and sooty mangabey (SIVsmm) are thought to have a long evolutionary history with their hosts. These hosts have adapted to the presence of the virus,[98] which is present at high levels in the host's blood, but evokes only a mild immune response,[99] does not cause the development of simian AIDS,[100] and does not undergo the extensive mutation and recombination typical of HIV infection in humans.[101]
In contrast, when these strains infect species that have not adapted to SIV ("heterologous" or similar hosts such as rhesus or cynomologus macaques), the animals develop AIDS and the virus generates genetic diversity similar to what is seen in human HIV infection.[102] Chimpanzee SIV (SIVcpz), the closest genetic relative of HIV-1, is associated with increased mortality and AIDS-like symptoms in its natural host.[103] SIVcpz appears to have been transmitted relatively recently to chimpanzee and human populations, so their hosts have not yet adapted to the virus.[98] This virus has also lost a function of the nef gene that is present in most SIVs. For non-pathogenic SIV variants, nef suppresses T cell activation through the CD3 marker. Nef's function in non-pathogenic forms of SIV is to downregulate expression of inflammatory cytokines, MHC-1, and signals that affect T cell trafficking. In HIV-1 and SIVcpz, nef does not inhibit T-cell activation and it has lost this function. Without this function, T cell depletion is more likely, leading to immunodeficiency.[103][104]
Three groups of HIV-1 have been identified on the basis of differences in the envelope (env) region: M, N, and O.[105] Group M is the most prevalent and is subdivided into eight subtypes (or clades), based on the whole genome, which are geographically distinct.[106] The most prevalent are subtypes B (found mainly in North America and Europe), A and D (found mainly in Africa), and C (found mainly in Africa and Asia); these subtypes form branches in the phylogenetic tree representing the lineage of the M group of HIV-1. Co-infection with distinct subtypes gives rise to circulating recombinant forms (CRFs). In 2000, the last year in which an analysis of global subtype prevalence was made, 47.2% of infections worldwide were of subtype C, 26.7% were of subtype A/CRF02_AG, 12.3% were of subtype B, 5.3% were of subtype D, 3.2% were of CRF_AE, and the remaining 5.3% were composed of other subtypes and CRFs.[107] Most HIV-1 research is focused on subtype B; few laboratories focus on the other subtypes.[108] The existence of a fourth group, "P", has been hypothesised based on a virus isolated in 2009.[109][110] The strain is apparently derived from gorilla SIV (SIVgor), first isolated from western lowland gorillas in 2006.[109]
HIV-2's closest relative is SIVsm, a strain of SIV found in sooty mangabees. Since HIV-1 is derived from SIVcpz, and HIV-2 from SIVsm, the genetic sequence of HIV-2 is only partially homologous to HIV-1 and more closely resembles that of SIVsm.[111][112]
Diagnosis
Many HIV-positive people are unaware that they are infected with the virus.[113] For example, in 2001 less than 1% of the sexually active urban population in Africa had been tested, and this proportion is even lower in rural populations.[113] Furthermore, in 2001 only 0.5% of pregnant women attending urban health facilities were counselled, tested or received their test results.[113] Again, this proportion is even lower in rural health facilities.[113] Since donors may therefore be unaware of their infection, donor blood and blood products used in medicine and medical research are routinely screened for HIV.[114]
HIV-1 testing is initially done using an enzyme-linked immunosorbent assay (ELISA) to detect antibodies to HIV-1. Specimens with a non-reactive result from the initial ELISA are considered HIV-negative, unless new exposure to an infected partner or partner of unknown HIV status has occurred. Specimens with a reactive ELISA result are retested in duplicate.[115] If the result of either duplicate test is reactive, the specimen is reported as repeatedly reactive and undergoes confirmatory testing with a more specific supplemental test (e.g., a polymerase chain reaction (PCR), western blot or, less commonly, an immunofluorescence assay (IFA)). Only specimens that are repeatedly reactive by ELISA and positive by IFA or PCR or reactive by western blot are considered HIV-positive and indicative of HIV infection. Specimens that are repeatedly ELISA-reactive occasionally provide an indeterminate western blot result, which may be either an incomplete antibody response to HIV in an infected person or nonspecific reactions in an uninfected person.[116]
HIV deaths in 2014 excluding the U.S.:[117]
Although IFA can be used to confirm infection in these ambiguous cases, this assay is not widely used. In general, a second specimen should be collected more than a month later and retested for persons with indeterminate western blot results. Although much less commonly available, nucleic acid testing (e.g., viral RNA or proviral DNA amplification method) can also help diagnosis in certain situations.[115] In addition, a few tested specimens might provide inconclusive results because of a low quantity specimen. In these situations, a second specimen is collected and tested for HIV infection.
Modern HIV testing is extremely accurate, when the window period is taken into consideration. A single screening test is correct more than 99% of the time.[118] The chance of a false-positive result in a standard two-step testing protocol is estimated to be about 1 in 250,000 in a low risk population.[119] Testing post-exposure is recommended immediately and then at six weeks, three months, and six months.[120]
Последние рекомендации Центров США по контролю и профилактике заболеваний (CDC) показывают, что тестирование на ВИЧ должно начинаться с комбинированного теста иммуноанализа к ВИЧ-1 и ВИЧ-2 для антител P24 и антигена . Негативный результат исключает воздействие на ВИЧ, в то время как положительный должен сопровождаться иммуноанализом дифференцировки антител в ВИЧ-1/2, чтобы обнаружить, какие антитела присутствуют. Это порождает четыре возможных сценария:
- 1. ВИЧ-1 (+) и ВИЧ-2 (-): ВИЧ-1 обнаружены
- 2. ВИЧ-1 (-) и ВИЧ-2 (+): ВИЧ-2 обнаружены антитела
- 3. ВИЧ-1 (+) и ВИЧ-2 (+): Обнаруженные антитела к ВИЧ-1 и ВИЧ-2
- 4. ВИЧ-1 (-) или неопределенный и ВИЧ-2 (-): тест на нуклеиновую кислоту для обнаружения острой инфекции ВИЧ-1 или его отсутствия. необходимо провести [ 121 ]
Исследовать
Исследования по ВИЧ/СПИДу включают все медицинские исследования , которые пытаются предотвратить, лечить или лечить ВИЧ/СПИД , а также фундаментальные исследования о природе ВИЧ как инфекционного агента и СПИДа как заболевания, вызванного ВИЧ.
Многие правительства и исследовательские институты участвуют в исследованиях по ВИЧ/СПИДу. Это исследование включает в себя поведенческие вмешательства в области здоровья , такие как исследования полового воспитания и разработка лекарств , такие как исследования микробицидов для заболеваний, передаваемых половым путем , вакцин против ВИЧ и антиретровирусные препараты . [ 122 ] Другие области медицинских исследований включают темы профилактики до воздействия , профилактику после воздействия , обрезание и ускоренные эффекты старения .
Обработка и передача
Управление ВИЧ/СПИДом обычно включает использование множества антиретровирусных препаратов . Во многих частях мира ВИЧ стал хроническим состоянием, причем прогрессирование СПИДа все более редко.
Задержка ВИЧ и полученное вирусное резервуар в CD4 + Т -клетки, дендритные клетки и макрофаги являются основным барьером для уничтожения вируса. [ 19 ] [ 123 ]
В то время как ВИЧ очень вирулентный, передача через половой контакт не происходит, когда ВИЧ-позитивный человек поддерживает неизменно неопределяемую вирусную нагрузку (<50 копий/мл) из-за антиретровирусного лечения. Эта концепция была впервые предложена Швейцарской федеральной комиссией по СПИДу/ВИЧ в 2008 году в том, что известно как Швейцарское заявление . [ 124 ] [ 125 ] Хотя изначально последующие исследования подтвердили, что риск передачи ВИЧ посредством пола эффективно нулевой, когда у ВИЧ-позитивного человека постоянно не обнаруживаемая вирусная нагрузка, концепция, широко известная как u = u, или «неопределяемое = невозможное». [ 126 ] [ 127 ]
Исследования, установившие принцип U = U [ 128 ] Партнер 1, [ 129 ] Партнер 2 [ 5 ] [ 130 ] (который был сосредоточен на парах мужского мужчин), [ 131 ] и HPTN052 [ 132 ] (который сосредоточен на гетеросексуальных парах). [ 131 ] Эти исследования включали пары, в которых один партнер был ВИЧ-положительным, а один был ВИЧ-негативным, и включал регулярное тестирование на ВИЧ. В этих четырех исследованиях в общей сложности 4097 пар приняли участие в четырех континентах, сообщив о 151 880 актах без презерватива с нулевым филогенетически связанным с передачей ВИЧ, когда у положительного партнера была неопределяемая вирусная нагрузка. [ 133 ] Следуя этим выводам, консенсус -заявление U = U, выступающее за использование термина «нулевой риск», было одобрено многочисленными людьми и организациями, включая CDC , Британская ассоциация ВИЧ и медицинский журнал Lancet . [ 134 ]
Кроме того, реактивация герпестного вируса-2 (HSV-2) у людей с генитальным герпесом связана с увеличением CCR-5, обогащенного CD4 + Т -клетки и воспалительные дендритные клетки в дерме язвенной генитальной кожи, сохраняющиеся даже после заживления язвы. Тропизм ВИЧ для CCR-5-положительных клеток способствует увеличению риска приобретения ВИЧ в два-три раза у людей с генитальным герпесом. Примечательно, что ежедневные противовирусные препараты, такие как ацикловир , не снижают субклиническое воспаление после реактивации и, следовательно, не снижают риск получения ВИЧ. [ 135 ] [ 136 ]
История
Открытие



Первая новость о «Экзотической новой болезни» появилась 18 мая 1981 года в гей -газете в Нью -Йорке . [ 137 ]
СПИД впервые наблюдался в 1981 году в Соединенных Штатах. [ 138 ] Первоначальные случаи были кластером инъекционных потребителей наркотиков и геев без известной причины нарушения иммунитета, у которых были симптомы пневмоцистисной пневмонии (PCP или PJP, последний термин, признающий, что причинный агент теперь называется Pneumocystis Jirovecii ), редкая оппортуальная инфекция, теперь редкая оппортунистическая инфекция, редкая оппортунистическая инфекция, редкая оппортунистическая инфекция, редкая оппортунистическая инфекция, редкая оппортунистическая инфекция) Известно, что это происходит у людей с очень скомпрометированной иммунной системой. [ 139 ] Вскоре после этого исследователи из Медицинской школы Нью -Йоркского университета изучали гей -мужчины, развивавшие ранее редкий рак кожи под названием саркома Капоси (KS). [ 140 ] [ 141 ] Появились еще много случаев PJP и KS, предупреждающие центры США о контроле и профилактике заболеваний (CDC) и была сформирована целевая группа CDC для мониторинга вспышки. [ 142 ] Считается, что самый ранний ретроспективно описанный случай СПИДа был в Норвегии, начиная с 1966 года. [ 143 ]
Вначале CDC не имел официального названия для этой болезни, часто ссылаясь на нее посредством заболеваний, которые были связаны с ней, например, лимфаденопатию , болезнь, после которой открывники ВИЧ первоначально назвали вирус. [ 144 ] [ 145 ] Они также использовали саркому Капоси и оппортунистические инфекции , название, с помощью которого в 1981 году была создана целевая группа. [ 146 ] В общей прессе была придумана термин сетка , которая стояла за иммунодефицитом, связанного с геями , была придумана. [ 147 ] CDC в поисках имени и смотрев на зараженные сообщества, придумал «болезнь 4H», поскольку оно, казалось, выделяло гомосексуалистов, пользователей героина, гемофиликов и гаитян . [ 148 ] [ 149 ] Однако после определения того, что СПИД не был изолирован для гей -сообщества , [ 146 ] Было понято, что термин сетка вводит в заблуждение, а СПИД был представлен на встрече в июле 1982 года. [ 150 ] К сентябрю 1982 года CDC начал использовать имя СПИДа. [ 151 ]
В 1983 году две отдельные исследовательские группы во главе с американским Робертом Галло и французскими следователями Франсуазой Барре-Синоси и Люком Монтанье независимо заявили, что новый ретровирус, возможно, заразил пациентов СПИДа и опубликовал свои результаты в том же вопросе в журнале науки . [ 152 ] [ 145 ] [ 153 ] Галло утверждал, что вирус, который его группа изолировал от человека со СПИДом, поразительно похож по форме с другими человеческими Т-лимфотропными вирусами (HTLV), его группа была первой, кто изолировал. Галло признал в 1987 году, что вирус, который он утверждал, обнаружил в 1984 году, на самом деле был вирусом, отправленным ему из Франции годом ранее. [ 154 ] Группа Галло назвала их недавно изолированный вирус HTLV-III. Группа Montagnier выделяла вирус от пациента с отеком лимфатических узлов шеи и физической слабости , двух классических симптомов первичной ВИЧ -инфекции. Противоречивая отчету группы Галло, Монтанье и его коллеги показали, что основные белки этого вируса иммунологически отличаются от белков HTLV-I. Группа Montagnier назвала их изолированный вирусный вирус, связанный с лимфаденопатией (LAV). [ 142 ] Поскольку эти два вируса оказались одинаковыми, в 1986 году Лав и HTLV-III были переименованы в ВИЧ. [ 155 ]
Другой группой, работающей одновременно с группами Montagnier и Gallo, была группа Джея А. Леви в Калифорнийском университете в Сан -Франциско . Он самостоятельно обнаружил вирус СПИДа в 1983 году и назвал его связанным с СПИДом ретровирусом (ARV). [ 156 ] Этот вирус сильно отличался от вируса, о котором сообщалось в группах Монтанье и Галло. Штаммы ARV впервые указали на неоднородность изолятов ВИЧ, и некоторые из них остаются классическими примерами вируса СПИДа, обнаруженных в Соединенных Штатах. [ 157 ]
Происхождение
Считается, что как ВИЧ-1, так и ВИЧ-2 возникли у нечеловеческих приматов в Западной Центральной Африке, и, как полагают, передали людям (процесс, известный как зооноз ) в начале 20-го века. [ 158 ] [ 159 ]
ВИЧ-1, по-видимому, возник в южном Камеруне благодаря эволюции SIVCPZ, вируса иммунодефицита обезьян (SIV), который заражает диких шимпанзе (ВИЧ-1 выходит из эндемических SIVCPZ в подвиде шимпанзе, панирующих троглодиты троглодитов ). [ 160 ] [ 161 ] Самым близким родственником ВИЧ-2 является SIVSMM, вирус сажих Мангаби ( Cercocebus atys Atys ), обезьяны Старого Света, живущая в прибрежной Западной Африке (от южного Сенегала до западного Кот-д'Ивуара ). [ 21 ] Обезьяны Нового Света, такие как Обезьяна Совы, устойчивы к инфекции ВИЧ-1, возможно, из-за геномного слияния двух генов устойчивости к вируса. [ 162 ]
Считается, что ВИЧ-1 прыгал на видовый барьер как минимум в трех отдельных случаях, вызывая три группы вируса, M, N и O. [ 163 ]

Существуют доказательства того, что люди, которые участвуют в мероприятиях в буштинге , либо в качестве охотников, либо как продавцы буштирования, обычно приобретают SIV. [ 164 ] Однако SIV является слабым вирусом, и он обычно подавляется иммунной системой человека в течение нескольких недель после инфекции. Считается, что несколько передач вируса от человека к человеку в быстрой последовательности необходимы, чтобы дать ему достаточно времени, чтобы утирать в ВИЧ. [ 165 ] Кроме того, из-за его относительно низкой скорости передачи от человека к человеку он может распространяться только по всей популяции в присутствии одного или нескольких каналов передачи высокого риска, которые, как считается, отсутствовали в Африке до 20-го века.
Специфические предложенные каналы передачи высокого риска, позволяющие вирусу адаптироваться к людям и распространяться по всему обществу, зависят от предлагаемого времени пересечения животного к человеку. Генетические исследования вируса предполагают, что самый последний общий предок группы ВИЧ-1 М восходит к C. 1910 . [ 166 ] Сторонники этого знакомства связывают эпидемию ВИЧ с появлением колониализма и роста крупных колониальных африканских городов, что приводит к социальным изменениям, включая различные модели сексуального контакта (особенно множественные, одновременные партнерские отношения), распространение проституции и сопутствующую высокую частоту генитальных язвенных заболеваний (таких как сифилис ) в зарождающихся колониальных городах. [ 167 ] В то время как скорости передачи ВИЧ во время вагинального полового акта, как правило, низкие, они увеличиваются многократно, если один из партнеров имеет инфекцию, передаваемую половым путем , приводя к генитальным язвам. В начале 1900 -х годов колониальные города были известны своей высокой распространенностью проституции и генитальных язв до той степени, в которой по состоянию на 1928 год 45% женщин -жителей Восточного Леопольдвилля (в настоящее время Киншаса) были проститутками и в 1933 году около 15%. Из всех жителей одного города были заражены одной из форм сифилиса . [ 167 ]
Самый ранний, хорошо документированный случай ВИЧ в человеческом датировании 1959 года в Бельгийском Конго . [ 168 ] Вирус, возможно, присутствовал в Соединенных Штатах еще в середине и конце 1960-х годов, когда шестнадцатилетний мужчина по имени Роберт Рэйфорд с симптомами в 1966 году умер в 1969 году. [ 169 ]
Альтернативная и, вероятно, дополнительная гипотеза указывает на широкое использование небезопасных медицинских практик в Африке в течение многих лет после Второй мировой войны, таких как неофиливое повторное использование одноразовых шприцев во время массовых вакцинаций, антибиотиков и антималарийских лечения. [ 165 ] [ 170 ] [ 171 ] Исследования по срокам самого последнего общего предка для групп ВИЧ-1 M и O, а также о группах ВИЧ-2 A и B указывают на то, что SIV породил трансмиссируемые линии ВИЧ на протяжении всего двадцатого века. [ 172 ] Распределенное время этих передач с людьми подразумевает, что для объяснения перекрестной передачи ВИЧ не требуется единого внешнего фактора. Это наблюдение согласуется с обоими двумя преобладающими взглядами на происхождение эпидемий ВИЧ, а именно передачи SIV для людей во время бойни или заканчивающего инфицированными приматами, и колониальной экспансии африканских городов Африки к югу от Сахары. [ 172 ]
Смотрите также
- Антивирусное препарат
- Открытие и развитие ингибиторов ВИЧ-протеазы
- ВИЧ/ОТКРЫТИЕ СПИДА
- Hivtoolbox
- Всемирный день СПИДа
Ссылки
- ^ Вайс Р.А. (май 1993). «Как ВИЧ вызывает СПИД?». Наука . 260 (5112): 1273–9. Bibcode : 1993sci ... 260.1273W . doi : 10.1126/science.8493571 . PMID 8493571 .
- ^ Douek DC, Roederer M, Koup RA (2009). «Новые концепции в иммунопатогенезе СПИДа» . Ежегодный обзор медицины . 60 : 471–84. doi : 10.1146/annurev.med.60.041807.123549 . PMC 2716400 . PMID 18947296 .
- ^ Jump up to: а беременный Пауэлл М.К., Бенкова К., Селингер П., Догос М., Кинкорова Лухачкова И., Кутникова Х., Лаштикова Дж., Рубикова А., Сперкова З., Лаклова Л., Эйс В., Шахмат Дж., Хенеберг П. (2016). «Оппортунистические инфекции у ВИЧ-инфицированных пациентов сильно различаются по частотам и спектрам между пациентами с низким количеством CD4+ клеток, изучаемых после обмотки, и пациентов с компенсацией исследовали антреморт независимо от HAART» . Plos один . 11 (9): E0162704. BIBCODE : 2016PLOSO..1162704P . Doi : 10.1371/journal.pone.0162704 . PMC 5017746 . PMID 27611681 .
- ^ Unaids, кто (декабрь 2007 г.). «Обновление эпидемии СПИДа 2007 года» (PDF) . п. 16
- ^ Jump up to: а беременный в Rodger AJ, Cambiano V, Bruun T, Vernazza P, Collins S, Degen O, et al. (Июнь 2019). «Риск передачи ВИЧ с помощью секса без презерватива у серодифферентных гей-пар, а ВИЧ-положительный партнер принимает подавляющую антиретровирусную терапию (партнер): окончательные результаты многоцентрового, проспективного, наблюдательного исследования» . Лансет . 393 (10189): 2428–2438. doi : 10.1016/s0140-6736 (19) 30418-0 . PMC 6584382 . PMID 31056293 .
- ^ Jump up to: а беременный Eisinger RW, Dieffenbach CW, Fauci As (февраль 2019 г.). «ВИЧ -вирусная нагрузка и трансмиссия ВИЧ -инфекции: неопределяемое равняется невозможным». Джама . 321 (5): 451–452. doi : 10.1001/Jama.2018.21167 . PMID 30629090 . S2CID 58599661 .
- ^ Mabuka J, Nduati R, Odem-Davis K, Peterson D, Overbough J (2012). Desrosiers RC (ред.). «ВИЧ-специфические антитела, способные к ADCC, распространены в грудном молоке и связаны со снижением риска передачи у женщин с высокими вирусными нагрузками» . PLO -патогены . 8 (6): E1002739. doi : 10.1371/journal.ppat.1002739 . PMC 3375288 . PMID 22719248 .
- ^ Hahn Ra, Inhorn MC, eds. (2009). Антропология и общественное здравоохранение: соединение различий в культуре и обществе (2 -е изд.). Оксфорд: издательство Оксфордского университета. п. 449. ISBN 978-0-19-537464-3 Полем OCLC 192042314 .
- ^ Мид М.Н. (2008). «Загрязнители в человеческом молоке: взвесить риски против преимуществ грудного вскармливания» . Перспективы здоровья окружающей среды . 116 (10): A426–34. doi : 10.1289/ehp.116-A426 . PMC 2569122 . PMID 18941560 . Архивировано из оригинала 6 ноября 2008 года.
- ^
 Эта статья включает текст из этого источника, который находится в общественном доступе : «Предотвращение передачи от ВИЧ от матери-ребенка» . ВИЧ.ГОВ . 15 мая 2017 года . Получено 8 декабря 2017 года .
Эта статья включает текст из этого источника, который находится в общественном доступе : «Предотвращение передачи от ВИЧ от матери-ребенка» . ВИЧ.ГОВ . 15 мая 2017 года . Получено 8 декабря 2017 года .
- ^ Каннингем А.Л., Донахи Х., Харман А.Н., Ким М., Турвилль С.Г. (август 2010 г.). «Манипулирование функцией дендритных клеток вирусами». Текущее мнение о микробиологии . 13 (4): 524–9. doi : 10.1016/j.mib.2010.06.002 . PMID 20598938 .
- ^ Doitsh G, Galloway NL, Geng X, Yang Z, Monroe KM, Zepeda O, et al. (Январь 2014). «Клеточная гибель путем пироптоза приводит к истощению CD4 Т-клеток при ВИЧ-1 инфекции» . Природа . 505 (7484): 509–14. Bibcode : 2014natur.505..509d . doi : 10.1038/nature12940 . PMC 4047036 . PMID 24356306 .
- ^ Гарг Х., Моль Дж., Джоши А (ноябрь 2012 г.). «ВИЧ-1 вызвал апоптоз свидетеля» . Вирусы . 4 (11): 3020–43. doi : 10.3390/v4113020 . PMC 3509682 . PMID 23202514 .
- ^ Кумар V (2012). Роббинс Базовая патология (9 -е изд.). Elsevier Health Sciences. п. 147. ISBN 978-1-4557-3787-1 .
- ^ Международный комитет по таксономии вирусов (2002). "61.0.6. лентивирус" . Национальные институты здравоохранения . Архивировано из оригинала 14 октября 2006 года . Получено 28 февраля 2006 г.
{{cite web}}: Cs1 maint: непредвзятый URL ( ссылка ) - ^ Международный комитет по таксономии вирусов (2002). "61. Retroviridae" . Национальные институты здравоохранения. Архивировано из оригинала 2 октября 2006 года . Получено 28 февраля 2006 г.
{{cite web}}: Cs1 maint: непредвзятый URL ( ссылка ) - ^ Леви JA (ноябрь 1993). «Патогенез ВИЧ и долгосрочное выживание». СПИД . 7 (11): 1401–10. doi : 10.1097/00002030-199311000-00001 . PMID 8280406 .
- ^ Смит Дж.А., Даниэль Р (май 2006 г.). «Следуя пути вируса: эксплуатация механизмов репарации ДНК хозяина ретровирусами». ACS Химическая биология . 1 (4): 217–26. doi : 10.1021/cb600131q . PMID 17163676 .
- ^ Jump up to: а беременный Силисиано Р.Ф., Грин, WC (сентябрь 2011 г.). «Задержка ВИЧ» . Перспективы Cold Spring Harbor в медицине . 1 (1): A007096. doi : 10.1101/cshperspect.a007096 . PMC 3234450 . PMID 22229121 .
- ^ Гилберт П.Б., МакКиг И.В., Эйзен Г., Маллинс С., Гея-Ндиай А., Мбуп С., Канки П.Дж. (28 февраля 2003 г.). «Сравнение инфекционности ВИЧ-1 и ВИЧ-2 из проспективного когортного исследования в Сенегале». Статистика в медицине . 22 (4): 573–593. doi : 10.1002/sim.1342 . PMID 12590415 . S2CID 28523977 .
- ^ Jump up to: а беременный Reeves JD, Doms RW (2002). «Вирус иммунодефицита человека тип 2» . Журнал общей вирусологии . 83 (Pt 6): 1253–65. doi : 10.1099/0022-1317-83-6-1253 . PMID 12029140 .
- ^ McGovern SL, Caselli E, Grigorieff N, Shoichet BK (2002). «Общий механизм, лежащий в основе беспорядочных ингибиторов из виртуального и высокопроизводительного скрининга». Журнал лекарственной химии . 45 (8): 1712–22. doi : 10.1021/jm010533y . HDL : 11380/977912 . PMID 11931626 .
- ^ По сравнению с обзором в: Фишер Б., Харви Р.П., Чампе ПК (2007). Иллюстрированные обзоры Липпинкотта: Микробиология . Hagerstown, MD: Lippincott Williams & Wilkins. п. 3. ISBN 978-0-7817-8215-9 .
- ^ Jump up to: а беременный в дюймовый и фон глин Различные (2008). Сборник ВИЧ Compendium 2008 ВВЕДЕНИЕ (PDF) . Получено 31 марта 2009 г.
- ^ Jump up to: а беременный в Chan DC, Fass D, Berger JM, Kim PS (апрель 1997 г.). «Основная структура GP41 из гликопротеина огибания ВИЧ» (PDF) . Клетка . 89 (2): 263–73. doi : 10.1016/s0092-8674 (00) 80205-6 . PMID 9108481 . S2CID 4518241 .
- ^ Klein JS, Bjorkman PJ (май 2010 г.). «Несколько и далеко между ними: как ВИЧ может уклоняться от обнурания антител» . PLO -патогены . 6 (5): E1000908. doi : 10.1371/journal.ppat.1000908 . PMC 2877745 . PMID 20523901 .
- ^ Национальный институт здравоохранения (17 июня 1998 г.). «Кристаллическая структура ключевого белка ВИЧ выявляет новую профилактику, мишени для лечения» (пресс -высвобождение). Архивировано из оригинала 19 февраля 2006 года . Получено 14 сентября 2006 г.
- ^ Behrens AJ, Vasiljevic S, Pritchard LK, Harvey DJ, Andev RS, Krumm SA, et al. (Март 2016 г.). «Композиция и антигенные эффекты отдельных гликановых участков тримерного гликопротеина огибального ВИЧ-1» . Сотовые отчеты . 14 (11): 2695–706. doi : 10.1016/j.celrep.2016.02.058 . PMC 4805854 . PMID 26972002 .
- ^ Причард Л.К., Спенсер Д.И., Ройл Л., Бонолли С., Сибрайт Г.Е., Беренс А.Дж. и др. (Июнь 2015 г.). «Гликан кластеризация стабилизирует маннозу ВИЧ-1 и сохраняет уязвимость для широкого нейтрализации антител» . Природная связь . 6 : 7479. Bibcode : 2015natco ... 6.7479p . doi : 10.1038/ncomms8479 . PMC 4500839 . PMID 26105115 .
- ^ Причард Л.К., Харви Д.Дж., Бонолли С., Криспин М., Дверей К.Дж. (сентябрь 2015 г.). «Клеточно-белковая гликозилирование нативного расщепленного огибала ВИЧ-1» . Журнал вирусологии . 89 (17): 8932–44. doi : 10.1128/jvi.01190-15 . PMC 4524065 . PMID 26085151 .
- ^ Crispin M, Doores KJ (апрель 2015 г.). «Нацеливание на гликаны, полученные из хозяина на охваченные вирусы для конструкции вакцины на основе антител» . Текущее мнение о вирусологии . 11 : 63–9. doi : 10.1016/j.coviro.2015.02.002 . PMC 4827424 . PMID 25747313 .
- ^ Julien JP, Cupo A, Sok D, Stanfield RL, Lyumkis D, Deller MC, et al. (Декабрь 2013). «Кристаллическая структура растворимой тримера растворимой расщепленной ВИЧ-1» . Наука . 342 (6165): 1477–83. Bibcode : 2013sci ... 342.1477j . doi : 10.1126/science.1245625 . PMC 3886632 . PMID 24179159 .
- ^ Lyumkis D, Julien JP, De Val N, Cupo A, Potter CS, Klasse PJ, et al. (Декабрь 2013). «Крио-эм структура полностью гликозилированной растворимой расщепленной тример огибала ВИЧ-1» . Наука . 342 (6165): 1484–90. Bibcode : 2013sci ... 342.1484L . doi : 10.1126/science.1245627 . PMC 3954647 . PMID 24179160 .
- ^ Sanders RW, Derking R, Cupo A, Julien JP, Yasmeen A, De Val N, et al. (Сентябрь 2013 г.). «Растворимый растворимый растворимый ВИЧ-1 Env Trimer, BG505 Sosip.664 GP140, экспрессирует множественные эпитопы для широко нейтрализующих, но не нейтрализующих антител» . PLO -патогены . 9 (9): E1003618. doi : 10.1371/journal.ppat.1003618 . PMC 3777863 . PMID 24068931 .
- ^ Причард Л.К., Васильжевич С., Озоровский Г., Сибрайт Г.Е., Купочка А., Ринге Р. и др. (Июнь 2015 г.). «Структурные ограничения определяют гликозилирование тримеров оболочки ВИЧ-1» . Сотовые отчеты . 11 (10): 1604–13. doi : 10.1016/j.celrep.2015.05.017 . PMC 4555872 . PMID 26051934 .
- ^ De Taeye SW, Ozorowski G, Torrents de La Peña A, Guttman M, Julien JP, Van Den Kerkhof TL, et al. (Декабрь 2015). «Иммуногенность стабилизированных тримеров оболочки ВИЧ-1 с уменьшенным воздействием нететрализующих эпитопов» . Клетка . 163 (7): 1702–15. doi : 10.1016/j.cell.2015.11.056 . PMC 4732737 . PMID 26687358 .
- ^ Ouellet DL, Plante I, Landry P, Barat C, Janelle ME, Flamand L, Tremblay MJ, Provost P (апрель 2008 г.). «Идентификация функциональных микроРНК, высвобождаемого посредством асимметричной обработки элемента ВИЧ-1 TAR» . Исследование нуклеиновых кислот . 36 (7): 2353–65. doi : 10.1093/nar/gkn076 . PMC 2367715 . PMID 18299284 .
- ^ Гейдарит Р., Гейдарян М., Фу С., МакКаффри Т. "ВИЧ-1 смол . Рировирология . 6 (1): / 18. 1742-4690-6-1 10.1186 PMC 2654423 . 19220914PMID
- ^ Vasudevan AA, Smits SH, Höppner A, Häussinger D, Koenig BW, Münk C (ноябрь 2013 г.). «Структурные особенности антивирусных ДНК -цитидина деминаз» . Биологическая химия (представленная рукопись). 394 (11): 1357–70. doi : 10.1515/hsz-2013-0165 . PMID 23787464 . S2CID 4151961 .
- ^ Garcia JV, Miller AD (апрель 1991 г.). «Сериновая фосфорилирование-независимая подавляющая регуляция клеточной поверхности CD4 с помощью NEF». Природа . 350 (6318): 508–11. Bibcode : 1991natur.350..508G . doi : 10.1038/350508A0 . PMID 2014052 . S2CID 1628392 .
- ^ Schwartz O, Maréchal V, Le Gall S, Lemonnier F, Hered JM (март 1996 г.). «Эндоцитоз основных молекул комплекса гистосовместимости индуцируется белком NEF ВИЧ-1». Природная медицина . 2 (3): 338–42. doi : 10.1038/nm0396-338 . PMID 8612235 . S2CID 7461342 .
- ^ Stumptner-Cuvelette P, Morchoisne S, Dugast M, Le Gall S, Raposo G, Schwartz O, Benaroch P (октябрь 2001 г.). «NEF ВИЧ-1 ухудшает представление антигена II класса MHC и выражение поверхности» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (21): 12144–9. Bibcode : 2001pnas ... 9812144S . doi : 10.1073/pnas.221256498 . PMC 59782 . PMID 11593029 .
- ^ Аррильдт К.Т., Джозеф С.Б., Суонстром Р (март 2012 г.). «Белок Env ВИЧ-1: слой многих цветов» . Текущие отчеты о ВИЧ/СПИДе . 9 (1): 52–63. doi : 10.1007/s11904-011-0107-3 . PMC 3658113 . PMID 22237899 .
- ^ Jump up to: а беременный Berger EA, Doms RW, Fenyö EM, Korber BT, Littman DR, Moore JP, SchuteTemaker H, Sodroski J, Weiss RA (1998). "Новая классификация для ВИЧ-1 " Природа 391 (6664): Bibcode : 1998natur.391..240b 240. Doi : 10.1038/ 34571 9440686PMID 2159146S2CID
- ^ Jump up to: а беременный в Коакли Э., Петропулос К.Дж., Уиткомб Дж. М. (2005). «Оценка использования CH vbgemokine Co-рецептора при ВИЧ». Текущее мнение инфекционных заболеваний . 18 (1): 9–15. doi : 10.1097/00001432-200502000-00003 . PMID 15647694 . S2CID 30923492 .
- ^ Дэн Х, Лю Р., Эллмейер В., Чо С., Унутмаз Д., Буркхарт М., Ди Марцио П., Мармон С., Саттон Р.Е., Хилл С.М., Дэвис К.Б., Пейпер С.К., Шалл Т.Дж., Литтман Д.Р., Ландау Н.Р. (1996). «Идентификация основного корецептора для первичных изолятов ВИЧ-1». Природа . 381 (6584): 661–6. Bibcode : 1996natur.381..661d . doi : 10.1038/381661A0 . PMID 8649511 . S2CID 37973935 .
- ^ Feng Y, Broder CC, Kennedy PE, Berger EA (1996). «ВИЧ-1 кофактор: функциональное клонирование кДНК семилетнего рецептора, связанное с белком,» . Наука . 272 (5263): 872–7. Bibcode : 1996sci ... 272..872f . doi : 10.1126/science.272.5263.872 . PMC 3412311 . PMID 8629022 . S2CID 44455027 .
- ^ Knight SC, Macatonia SE, Patterson S (1990). «ВИЧ I Инфекция дендритных клеток». Международный обзор иммунологии . 6 (2–3): 163–75. doi : 10.3109/08830189009056627 . PMID 2152500 .
- ^ Тан Дж, Каслоу Р.А. (2003). «Влияние генетики хозяина на ВИЧ -инфекцию и прогрессирование заболеваний в эпоху высоко активной антиретровирусной терапии» . СПИД . 17 (Suppl 4): S51 - S60. doi : 10.1097/00002030-200317004-00006 . PMID 15080180 .
- ^ Zhu T, Mo H, Wang N, Nam DS, Cao Y, Koup RA, Ho DD (1993). «Генотипическая и фенотипическая характеристика пациентов с ВИЧ-1 с первичной инфекцией». Наука . 261 (5125): 1179–81. Bibcode : 1993sci ... 261.1179z . doi : 10.1126/science.8356453 . PMID 8356453 .
- ^ от wout ab, coatstra na, muller-kampinga ga, lbreight nent n, serpie hj, veenstra j, feenstra j, coutinho ra, cutteriner h (1994). «Макрогаж-тропический вариант инициирует вирус вируса иммунодефицита человека типа 1 и вертикальную передачу» . Журнал или клиническая инсатизация . 94 (5): 2060–7. doi : 10 1172/JCI17560 . PMC 29462 . PMID 796252 .
- ^ Чжу Т., Ван Н., Карр А., Нам Д.С., Мур-Янковский Р., Купер Д.А., Хо Д.Д. (1996). «Генетическая характеристика вируса иммунодефицита человека типа 1 в секреции крови и половых органов: доказательства вирусной компартментализации и отбора во время сексуальной передачи» . Журнал вирусологии . 70 (5): 3098–107. doi : 10.1128/JVI.70.5.3098-3107.1996 . PMC 190172 . PMID 8627789 .
- ^ Clevestig P, Maljkovic I, Casper C, Carlenor E, Lindgren S, Navér L, Bohlin AB, Fenyö EM, Leitner T, Ehrnst A (2005). «Фенотип X4 ВИЧ типа 1 развивается от R5 у двух детей матерей, несущих X4 и не связан с передачей». Исследование СПИДа и ретровирусы человека . 21 (5): 371–8. doi : 10.1089/aid.2005.21.371 . PMID 15929699 .
- ^ Мур JP (1997). «Корецепторы: последствия для патогенеза ВИЧ и терапии». Наука . 276 (5309): 51–2. doi : 10.1126/science.276.5309.51 . PMID 9122710 . S2CID 33262844 .
- ^ Карлссон А., Парсмир К., Аперия К., Сандстрем Е., Фениё Э.М., Альберт Дж. (1994). «Тропизм клеток MT-2 изолятов вируса иммунодефицита человека типа 1 в качестве маркера ответа на лечение и развитие лекарственной устойчивости». Журнал инфекционных заболеваний . 170 (6): 1367–75. doi : 10.1093/infdis/170.6.1367 . PMID 7995974 .
- ^ Koot M, Van 'T Wout AB, Kootstra NA, De Goede Re, Tersmette M, Schuitemaker H (1996). «Связь между изменениями в клеточной нагрузке, эволюцией вирусного фенотипа и клональным составом популяций вируса в ходе инфекции вируса иммунодефицита человека типа 1 типа» . Журнал инфекционных заболеваний . 173 (2): 349–54. doi : 10.1093/infdis/173.2.349 . PMID 8568295 .
- ^ Чейни К, Макнайт А (2010). «Тропизм и болезнь ВИЧ-2». Лентивирусы и макрофаги: молекулярные и клеточные взаимодействия . Caister Academic Press . ISBN 978-1-904455-60-8 . [ страница необходима ]
- ^ Jump up to: а беременный в дюймовый и фон глин Chan DC, Kim PS (1998). «Вход на ВИЧ и его торможение» . Клетка . 93 (5): 681–4. doi : 10.1016/s0092-8674 (00) 81430-0 . PMID 9630213 . S2CID 10544941 .
- ^ Jump up to: а беременный в дюймовый и фон Wyatt R, Sodroski J (1998). «Гликопротеины конверта ВИЧ-1: фусогены, антигены и иммуногены». Наука . 280 (5371): 1884–8. Bibcode : 1998sci ... 280.1884W . doi : 10.1126/science.280.5371.1884 . PMID 9632381 .
- ^ Jump up to: а беременный Arthos J, Cicala C, Martinelli E, Macleod K, Van Ryk D, Wei D, Xiao Z, Veenstra TD, Conrad TP, Lempicki RA, McLaughlin S, Pascuccio M, Gopaul R, McNally J, Cruz CC, Censoplano N, Chung E, Reitano KN, Kottilil S, Goode DJ, Fauci AS (2008). «Белок огибала ВИЧ-1 связывается и сигнализирует с помощью бета-бета (4) интегрина (4) (7), рецептора дома кишечника для периферических Т-клеток». Природа иммунология . 9 (3): 301–9. doi : 10.1038/ni1566 . PMID 18264102 . S2CID 205361178 .
- ^ Jump up to: а беременный Папа М, Хааз в (2003). «Передача, острая инфекция ВИЧ-1 и поиск стратегий для предотвращения инфекции» . Природная медицина . 9 (7): 847–52. doi : 10.1038/nm0703-847 . PMID 12835704 . S2CID 26570505 .
- ^ Haedicke J, Brown C, Naghavi MH (август 2009 г.). «Специфичный фактор мозга FEZ1 является детерминантом восприимчивости нейронов к инфекции ВИЧ-1» . Труды Национальной академии наук . 106 (33): 14040–14045. Bibcode : 2009pnas..10614040H . doi : 10.1073/pnas.0900502106 . PMC 2729016 . PMID 19667186 .
- ^ Daecke J, Fackler OT, Dittmar MT, Kräusslich Hg (2005). «Вовлечение клатрин-опосредованного эндоцитоза в проникновении вируса иммунодефицита человека типа 1 типа» . Журнал вирусологии . 79 (3): 1581–1594. doi : 10.1128/JVI.79.3.1581-1594.2005 . PMC 544101 . PMID 15650184 .
- ^ Мияучи К., Ким Ю., Латинович О., Морозов В., Меликян Г.Б. (2009). «ВИЧ входит в клетки через эндоцитоз и динамин-зависимое слияние с эндосомами» . Клетка . 137 (3): 433–444. doi : 10.1016/j.cell.2009.02.046 . PMC 2696170 . PMID 19410541 .
- ^ Кох П., Лампе М., Годинез В.Дж., Мюллер Б., Рор К., Краусслих Х.Г., Леманн М.Дж. (2009). «Визуализация слияния псевдотипированных частиц ВИЧ-1 в реальном времени с помощью живой клеточной микроскопии» . Ретровирология . 6 : 84. DOI : 10.1186/1742-4690-6-84 . PMC 2762461 . PMID 19765276 .
- ^ Thorley JA, McKeating JA, Rappoport JZ (2010). «Механизмы вирусного входа: красть в входной двери» . Протоплазма . 244 (1–4): 15–24. doi : 10.1007/s00709-010-0152-6 . PMC 3038234 . PMID 20446005 .
- ^ Permanyer M, Ballana E, Esté JA (2010). «Эндоцитоз ВИЧ: все идет». Тенденции в микробиологии . 18 (12): 543–551. doi : 10.1016/j.tim.2010.09.003 . PMID 20965729 .
- ^ Jump up to: а беременный в Zheng YH, Lovsin N, Peterlin BM (2005). «Недавно идентифицированные факторы хоста модулируют репликацию ВИЧ». Иммунологические письма . 97 (2): 225–34. doi : 10.1016/j.imlet.2004.11.026 . PMID 15752562 .
- ^ «IV. Вирусы> F. Жизненные циклы вируса животных> 3. Жизненный цикл ВИЧ» . Док Кайзерс на домашней странице микробиологии . Общественный колледж округа Балтимор. Январь 2008 года. Архивировано с оригинала 26 июля 2010 года.
- ^ Hiscott J, Kwon H, Génin P (2001). «Враждебные поглощения: вирусное присвоение пути NF-KB» . Журнал клинических исследований . 107 (2): 143–151. doi : 10.1172/jci11918 . PMC 199181 . PMID 11160127 .
- ^ Keane SC, Heng X, Lu K, Kharytonchyk S, Ramakrishnan V, Carter G, Barton S, Hosic A, Florwick A, Santos J, Bolden NC (22 мая 2015 г.). «Структура сигнала упаковки ВИЧ-1 РНК» . Наука . 348 (6237): 917–921. Bibcode : 2015sci ... 348..917K . doi : 10.1126/science.aaa9266 . ISSN 0036-8075 . PMC 4492308 . PMID 25999508 .
- ^ Keane SC, Van V, Frank HM, Sciandra CA, McCowin S, Santos J, Heng X, Summers MF (10 октября 2016 г.). «ЯМР обнаружение мест межмолекулярного взаимодействия в димерном 5'-лидере генома ВИЧ-1» . Труды Национальной академии наук . 113 (46): 13033–13038. BIBCODE : 2016PNAS..11313033K . doi : 10.1073/pnas.1614785113 . ISSN 0027-8424 . PMC 5135362 . PMID 27791166 .
- ^ Ocwieja Ke, Sherrill-Mix S, Mukherjee R, Custers-Allen R, David P, Brown M, et al. (Ноябрь 2012). «Динамическая регуляция популяций мРНК ВИЧ-1, проанализированные с помощью одномолекулярной обогащения и длинного чткого секвенирования» . Исследование нуклеиновых кислот . 40 (20): 10345–55. doi : 10.1093/nar/gks753 . PMC 3488221 . PMID 22923523 .
- ^ Поллард В.В., Малим М.Х. (1998). «Белок rev-1 ВИЧ-1». Ежегодный обзор микробиологии . 52 : 491–532. doi : 10.1146/annurev.micro.52.1.491 . PMID 9891806 .
- ^ Butsch M, Boris-Lawrie K (апрель 2002 г.). "Судьба не сплачивающейся ретровирусной РНК: рибосома и/или вирион?" Полем Журнал вирусологии . 76 (7): 3089–94. doi : 10.1128/jvi.76.7.3089-3094.2002 . PMC 136024 . PMID 11884533 .
- ^ Hellmund C, Lever Am (июль 2016 г.). «Координация упаковки геномной РНК с вирусной сборкой при ВИЧ-1» . Вирусы . 8 (7): 192. doi : 10.3390/v8070192 . PMC 4974527 . PMID 27428992 .
- ^ Soto-Rifo R, Limousin T, Rubilar PS, Ricci EP, Decime D, Moncorgé O и Al. (Март 2012 г.). Структурированная смолу и трансляция нуклеиновых кислот геномной РНК ВИЧ-2 Исследование 40 (6): 2653–67. два 10.1093/nar/gkr1093: 3315320PMC 22121214PMID
- ^ Saad JS, Muriaux DM (28 июля 2015 г.). Роль липидов в сборке вируса . Frontiers Media SA. ISBN 978-2-88919-582-4 .
- ^ Ricci EP, Herbreateau CH, Decimo D, Schaupp A, Datta SA, Rein A, et al. (Июль 2008 г.). «Экспрессия in vitro геномной РНК ВИЧ-2 контролируется тремя различными внутренними сегментами ввода рибосом, которые регулируются протеазой ВИЧ и полипротеином GAG» . РНК . 14 (7): 1443–55. doi : 10.1261/rna.813608 . PMC 2441975 . PMID 18495939 .
- ^ Jump up to: а беременный Hu WS, Temin HM (1990). «Ретровирусная рекомбинация и обратная транскрипция». Наука . 250 (4985): 1227–33. Bibcode : 1990sci ... 250.1227H . doi : 10.1126/science.1700865 . PMID 1700865 .
- ^ Jump up to: а беременный Charpentier C, Nora T, Tenaillon O, Clavel F, Hance AJ (2006). «Обширная рекомбинация среди кеазиспезии вируса иммунодефицита человека типа 1 вносит важный вклад в разнообразие вируса у отдельных пациентов» . Журнал вирусологии . 80 (5): 2472–82. doi : 10.1128/jvi.80.5.2472-2482.2006 . PMC 1395372 . PMID 16474154 .
- ^ Нора Т., Чарпентье С., Тенайлон О., Ходе С., Клавел Ф., Ханс А.Дж. (2007). «Вклад рекомбинации в эволюцию вирусов иммунодефицита человека, экспрессирующих устойчивость к антиретровирусным лечению» . Журнал вирусологии . 81 (14): 7620–8. doi : 10.1128/jvi.00083-07 . PMC 1933369 . PMID 17494080 .
- ^ Chen J, Powell D, Hu WS (2006). «Высокая частота генетической рекомбинации является общей чертой репликации лентивируса приматов» . Журнал вирусологии . 80 (19): 9651–8. doi : 10.1128/jvi.00936-06 . PMC 1617242 . PMID 16973569 .
- ^ Jump up to: а беременный Bonhoeffer S, Chappey C, Parkin NT, Whitcomb JM, Petropoulos CJ (2004). «Свидетельство положительного эпистаза в ВИЧ-1». Наука . 306 (5701): 1547–50. Bibcode : 2004sci ... 306.1547b . doi : 10.1126/science.1101786 . PMID 15567861 . S2CID 45784964 .
- ^ Исраэль Н., Гугеро-Поцидало М.А. (1997). «Окислительный стресс при инфекции вируса иммунодефицита человека» . Клеточные и молекулярные науки о жизни . 53 (11–12): 864–70. doi : 10.1007/s000180050106 . PMC 11147326 . PMID 9447238 . S2CID 22663454 .
- ^ Мишод Р.Е., Бернштейн Х, Недельку А.М. (май 2008 г.). «Адаптивная ценность пола в микробных патогенах» (PDF) . Инфекция, генетика и эволюция . 8 (3): 267–85. Bibcode : 2008infge ... 8..267m . doi : 10.1016/j.meegid.2008.01.002 . PMID 18295550 . Архивировано из оригинала (PDF) 16 мая 2017 года . Получено 10 мая 2013 года .
- ^ Hallenberger S, Bosch V, Angliker H, Shaw E, Klenk HD, Garden W (26 ноября 1992 г.). «Ингибирование опосредованной фун-опосредованной активацией расщепления ВИЧ-1 гликопротеина GP160». Природа . 360 (6402): 358–61. Bibcode : 1992natur.360..358h . Doi : 10.1038/360358A0 . PMID 1360148 . S2CID 4306605 .
- ^ Gelderblom HR (1997). «Прекрасная структура ВИЧ и SIV» (PDF) . В Лос -Аламос Национальной лаборатории (ред.). Сборник ВИЧ -последовательности . Лос -Аламос Национальная лаборатория . С. 31–44.
- ^ Jump up to: а беременный в дюймовый Zhang C, Zhou S, Groppelli E, Pellegrino P, Williams I, Bourn P, Chain Bm, Jolly C (2015). «Гибридные механизмы распространения и активация Т-клеток формируют динамику ВИЧ-1 инфекции» . PLOS Computational Biology . 11 (4): E1004179. Arxiv : 1503.08992 . BIBCODE : 2015PLSCB..11E4179Z . doi : 10.1371/journal.pcbi.1004179 . PMC 4383537 . PMID 25837979 .
- ^ Jump up to: а беременный Jolly C, Kashefi K, Hollinshead M, Sattentau QJ (2004). «Клетка ВИЧ-1 к переносу клеток через индуцированный ENV, актин-зависимый синапс» . Журнал экспериментальной медицины . 199 (2): 283–293. doi : 10.1084/jem.20030648 . PMC 2211771 . PMID 14734528 .
- ^ Sattentau Q (2008). «Избегание пустоты: клеточный распространение вирусов человека» . Nature Reviews Microbiology . 6 (11): 815–826. doi : 10.1038/nrmicro1972 . PMID 18923409 . S2CID 20991705 .
- ^ Дункан К.Дж., Рассел Р.А., Саттентау QJ (2013). «Высокая множественность ВИЧ-1-клеточной передачи от макрофагов к CD4+ T-клеткам ограничивает антиретровирусную эффективность» . СПИД . 27 (14): 2201–2206. doi : 10.1097/QAD.0B013E3283632EC4 . PMC 4714465 . PMID 24005480 .
- ^ Sewald X, Gonzalez DG, Haberman AM, Mothes W (2012). «Визуализация вирусологических синапсов in vivo» . Природная связь . 3 : 1320. Bibcode : 2012natco ... 3.1320S . doi : 10.1038/ncomms2338 . PMC 3784984 . PMID 23271654 .
- ^ Сигал А., Ким Д.Т., Балазс А.Б., Декель Э., Майо А., Мило Р., Балтимор Д. (2011). «Клеточно-клеточное распространение ВИЧ позволяет постоянной репликации, несмотря на антиретровирусную терапию» . Природа . 477 (7362): 95–98. Bibcode : 2011natur.477 ... 95 с . doi : 10.1038/nature10347 . PMID 21849975 . S2CID 4409389 .
- ^ Jump up to: а беременный в дюймовый Робертсон Д.Л., Хан Б.Х., Шарп П.М. (1995). «Рекомбинация в вирусах СПИДа» . Журнал молекулярной эволюции . 40 (3): 249–59. Bibcode : 1995jmole..40..249r . doi : 10.1007/bf00163230 . PMID 7723052 . S2CID 19728830 .
- ^ Rambaut A, Posada D, Crandall KA, Holmes EC (январь 2004 г.). «Причины и последствия эволюции ВИЧ» . Nature Reviews Genetics . 5 (52–61): 52–61. doi : 10.1038/nrg1246 . PMID 14708016 . S2CID 5790569 .
- ^ Перельсон А.С., Рибейро Р.М. (октябрь 2008 г.). «Оценка эффективности лекарственного средства и вирусных динамических параметров: ВИЧ и ВГС» . Статистика в медицине . 27 (23): 4647–57. doi : 10.1002/sim.3116 . PMID 17960579 . S2CID 33662579 .
- ^ Jump up to: а беременный Содора Д.Л., Аллан Дж.С., Апетрей С., Бренчли Дж. М., Доук Д.К., Истот Дж.Г., Эстес Дж. Д., Хан Б.Х., Хирш В.М., Каур А., Кирххофф Ф., Мюллер-Трутвин М., Пандреа I, Шмитц Дж., Силвестри Г. (2009). «На пути к вакцине со СПИДом: уроки от природных вирусных инфекций вируса об одномине иммунодефицита африканских хозяев нечеловеческих приматов» . Природная медицина . 15 (8): 861–865. doi : 10.1038/nm.2013 . PMC 2782707 . PMID 19661993 .
- ^ Хольсаммер С., Хольцнагель Э., Каул А., Курт Р., Норли С. (2001). «Высокие вирусные нагрузки в естественном и экспериментально инфицированном SIVAGM Африканские зеленые обезьяны» . Вирусология . 283 (2): 324–31. doi : 10.1006/viro.2001.0870 . PMID 11336557 .
- ^ Курт Р., Норли С. (1996). «Почему природные хозяева SIV не разрабатывают Semian SIDS?». Журнал NIH Research . 8 : 33–37.
- ^ Baier M, Dittmar MT, Cichutek K, Kurth R (1991). «Разработка Vivo генетической изменчивости вируса иммунодефицита обезьян» » . Труды Национальной академии наук Соединенных Штатов Америки . 88 (18): 8126–30. Bibcode : 1991pnas ... 88.8126b . doi : 10.1073/pnas.88.18.8126 . PMC 52459 . PMID 1896460 .
- ^ Даниэль М.Д., Кинг Н.В., Летвин Н.Л., Хант Р.Д., Сегал П.К., Десрозье Р.К. (1984). «Новый ретровирус типа D, выделенный из макак с синдромом иммунодефицита». Наука . 223 (4636): 602–5. Bibcode : 1984sci ... 223..602d . doi : 10.1126/science.6695172 . PMID 6695172 .
- ^ Jump up to: а беременный Кил Б.Ф., Джонс Дж. Х., Терио К.А., Эстес Д.Д., Рудиселл Р.С., Уилсон М.Л., Ли Й., Г.Х., Бизли Т.М., Шумахер-Станки Дж., Ворбски Е., Моссер А., Рафаэль Дж., Каменя С., Лонсдорф Е.В. Mlengeya T, Kinsel MJ, Else JG, Silvestri G, Goodall J, Sharp PM, Shaw GM, Pusey AE, Hahn BH (2009). «Увеличение смертности и иммунопатологии, подобной СПИДу, у диких шимпанзе, инфицированных SIVCPZ» . Природа . 460 (7254): 515–519. Bibcode : 2009natur.460..515K . doi : 10.1038/nature08200 . PMC 2872475 . PMID 19626114 .
- ^ Schindler M, Münch J, Kutsch O, Li H, Santiago Ml, Bibollet-ruche F, Müller-Trutn BH, Kirchhoff F (2006). «NEF-опосредованное подавление активации Т-клеток было потеряно в лентивирусной линии, которая породила ВИЧ-1» . Клетка . 125 (6): 1055–67. doi : 10.1016/j.cell.2006.04.033 . PMID 16777597 . S2CID 15132918 .
- ^ Томсон М.М., Перес-Альварес Л., Наджера Р. (2002). «Молекулярная эпидемиология генетических форм ВИЧ-1 и ее значение для развития вакцины и терапии». Lancet инфекционные заболевания . 2 (8): 461–471. doi : 10.1016/s1473-3099 (02) 00343-2 . PMID 12150845 .
- ^ Карр JK, BT, Leitner T, Salmone M, Kongyan F (1998). . PDF В Национальной лаборатории Аламоса (ред. ВИЧ -сексекский сборник Лос -Аламос, Нью -Мексико : Лос -Аламос Национальный . стр. 10–19.
- ^ Osmanov S, Pattou C, Walker N, Schwardländer B, Esparza J (2002). «Расчетное глобальное распределение и региональное распространение генетических подтипов ВИЧ-1 в 2000 году». Журнал приобретенных синдромов иммунной дефицита . 29 (2): 184–190. doi : 10.1097/00042560-200202010-00013 . PMID 11832690 . S2CID 12536801 .
- ^ Perrin L, Kaiser L, Yerly S (2003). «Путешествие и распространение генетических вариантов ВИЧ-1». Lancet инфекционные заболевания . 3 (1): 22–27. doi : 10.1016/s1473-3099 (03) 00484-5 . PMID 12505029 .
- ^ Jump up to: а беременный Plineier JC, Leoz M, Dickerson JE, De Oliveira F, Cordonnier F, Lemée V, Damond F, Robertson DL, Simon F (август 2009 г.). «Новый вирус иммуновидности человека, полученный из горилл». Природная медицина . 15 (8): 871–2. Doi : 10.1038/nm.2016 . PMID 19648927 . S2CID 76837833 .
- ^ Смит Л (3 августа 2009 г.). «Женщина обнаружила новую напряжение ВИЧ от горилл» . Независимый . Получено 27 ноября 2015 года .
- ^ Sharp PM, Hahn BH (август 2010 г.). «Эволюция ВИЧ-1 и происхождение СПИДа» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 365 (1552): 2487–94. doi : 10.1098/rstb.2010.0031 . PMC 2935100 . PMID 20643738 .
- ^ Keele Bf, Van Heuverswyn F, Li Y, Bailes E, Takehisa J, Santiago ML, et al. (Июль 2006 г.). «Шимпанзе водохранилища пандемии и непенемического ВИЧ-1» . Наука . 313 (5786): 523–6. Bibcode : 2006sci ... 313..523K . doi : 10.1126/science.1126531 . PMC 2442710 . PMID 16728595 .
- ^ Jump up to: а беременный в дюймовый Kumaranayake L, Watts C (2001). «Распределение ресурсов и приоритетная настройка вмешательств ВИЧ/СПИДа: решение обобщенной эпидемии в странах Африки к югу от Сахары». Журнал международного развития . 13 (4): 451–466. doi : 10.1002/jid.797 .
- ^ Kleinman S (сентябрь 2004 г.). «Информация о пациенте: донорство крови и переливание» . До настоящего времени. Архивировано с оригинала 12 апреля 2008 года.
- ^ Jump up to: а беременный Центры по контролю и профилактике заболеваний (2001). «Пересмотренные руководящие принципы по консультированию, тестированию и направлению ВИЧ». Рекомендации и отчеты MMWR . 50 (RR - 19): 1–57. PMID 11718472 .
- ^ Celum CL, Coombs RW, Lafferty W, Inui TS, Louie PH, Gates CA, McCreedy BJ, Egan R, Grove T, Alexander S (1991). «Неопределенные вирусы иммунодефицита человека Вестерн 1 типа 1: риск сероконверсии, специфичность дополнительных тестов и алгоритм для оценки». Журнал инфекционных заболеваний . 164 (4): 656–664. doi : 10.1093/infdis/164.4.656 . PMID 1894929 .
- ^ «Сравнение страны :: ВИЧ/СПИД - смерти» . Мировой факт, Центральное разведывательное агентство. Архивировано с оригинала 30 апреля 2017 года . Получено 22 ноября 2015 года .
- ^ Chou R, Selph S, Dana T, Bougatsos C, Zakher B, Blazina I, Korthuis Pt (ноябрь 2012 г.). «Скрининг на ВИЧ: систематический обзор для обновления рекомендации Целевой группы по профилактическим службам США» 2005 года » . Анналы внутренней медицины . 157 (10): 706–18. doi : 10.7326/0003-4819-157-10-201211200-00007 . PMID 23165662 . S2CID 27494096 .
- ^ Chou R, Huffman LH, Fu R, Smits AK, Korthuis PT (июль 2005 г.). «Скрининг на ВИЧ: обзор доказательств Целевой группы по профилактическим услугам США» . Анналы внутренней медицины . 143 (1): 55–73. doi : 10.7326/0003-4819-143-1-200507050-00010 . PMID 15998755 . S2CID 24086322 .
- ^ Толле Ма, Шварцвальд Хл (июль 2010 г.). «Профилактика постэкспозиции против вируса иммунодефицита человека» . Американский семейный врач . 82 (2): 161–6. PMID 20642270 . Архивировано с оригинала 28 ноября 2023 года.
- ^ «Быстрое справочное руководство - лабораторное тестирование для диагностики ВИЧ -инфекции: обновленные рекомендации» (PDF) . Центры для контроля и профилактики заболеваний . Департамент здравоохранения штата Нью -Йорк. 27 июня 2014 года. С. 1–2. Архивировано из оригинала (PDF) 2 марта 2017 года . Получено 13 апреля 2017 года .
- ^ «Лечение ВИЧ: одобренные FDA лекарства от ВИЧ» . AIDSINFO. Архивировано из оригинала 23 февраля 2017 года . Получено 7 октября 2016 года .
- ^ Rodari A, Darcis G, Van Lint CM (29 сентября 2021 г.). «Текущее состояние агентов по обращению задержки для ремиссии ВИЧ-1» . Ежегодный обзор вирусологии . 8 (1): 491–514. doi : 10.1146/annurev-virology-091919-103029 . ISSN 2327-056X . PMID 34586875 .
- ^ Швейцарская национальная комиссия по СПИДу (15 октября 2016 г.). «Швейцарское заявление» . ВИЧ I-база . Получено 2 апреля 2019 года .
- ^ Vernazza P, Bernard EJ (29 января 2016 г.). «ВИЧ не передается при полностью подавляющей терапии: швейцарское заявление - восемь лет спустя» . Швейцарский медицинский недель . 146 : W14246. doi : 10.4414/smw.2016.14246 . PMID 26824882 .
- ^ Ланцетный ВИЧ (ноябрь 2017 г.). "U = u взлетает в 2017 году" . Редакционная статья. Lancet. ВИЧ . 4 (11): E475. doi : 10.1016/s2352-3018 (17) 30183-2 . PMID 29096785 .
- ^ «Не могу пройти его» . Терренс Хиггинс доверие . 2019. Архивировано с оригинала 7 апреля 2019 года . Получено 2 апреля 2019 года .
- ^ Bavinton BR, Pinto AN, Phanuphak N, Grinsztejn B, Prestage GP, Zablotska-Manos IB, et al. (Август 2018). «Вирусное подавление и передача ВИЧ в серодискордантных мужских парах: международное, проспективное, наблюдательное, когортное исследование». Lancet. ВИЧ . 5 (8): E438 - E447. doi : 10.1016/s2352-3018 (18) 30132-2 . PMID 30025681 . S2CID 51702998 .
- ^ Роджер А.Дж., Камбиано В., Брун Т., Вернацца П., Коллинз С., Ван Лунзен Дж. И др. (Июль 2016 г.). «Сексуальная активность без презервативов и риск передачи ВИЧ в серодифферентных парах, когда ВИЧ-положительный партнер использует подавляющую антиретровирусную терапию» . Джама . 316 (2): 171–81. doi : 10.1001/Jama.2016.5148 . PMID 27404185 .
- ^ Роджер A ((июль 2018 г.). исследование Partner2 расширило результаты у гея . парных парах с супрессивным искусством : сексу без презерватива в Риск передачи ВИЧ через
- ^ Jump up to: а беременный Хоффман Х (10 января 2019 г.). «Наука ясна: с ВИЧ, неопределяемой равным невозможным» (пресс -релиз). Национальные институты здравоохранения . Национальный институт аллергии и инфекционных заболеваний . Получено 3 мая 2019 года .
Директор NIAID Энтони С. Фаучи, MD и коллеги суммируют результаты крупных клинических испытаний и когортных исследований, подтверждающих u = u. Клиническое исследование, финансируемое NIH, финансируемой NIH 052, показало, что не было связанных передачи ВИЧ среди серодифферентных гетеросексуальных пар ВИЧ, когда партнер, живущий с ВИЧ, имел более сильную вирусную нагрузку. Впоследствии партнер и противоположности привлекают исследования, подтвердили эти результаты и распространили их на пар мужчины. ... Успех u = u как метод профилактики ВИЧ зависит от достижения и поддержания неопределяемой вирусной нагрузки, принимая искусство ежедневно в соответствии с указаниями.
- ^ Cohen MS , Chen YQ, McCauley M, Gamble T, Hosseinipour MC, Kumarasamy N, et al. (Сентябрь 2016 г.). «Антиретровирусная терапия для профилактики передачи ВИЧ-1» . Новая Англия Журнал медицины . 375 (9): 830–9. doi : 10.1056/nejmoa1600693 . PMC 5049503 . PMID 27424812 .
- ^ Ходсон М (17 ноября 2017 г.). U = u: разговор с пациентами о риске передачи (PDF) . Британская ассоциация ВИЧ Осенняя конференция 2017 . Получено 3 мая 2019 года . ( Аннотация для презентации от имени NAM / SIDSMAP )
- ^ «Консенсусное заявление: риск сексуальной передачи ВИЧ от человека, живущего с ВИЧ, у которого не обнаруживается вирусная нагрузка» . Кампания доступа к профилактике . 21 июля 2016 года . Получено 2 апреля 2019 года . Примечание . Когда было получено заявление и список одобрений, в последний раз он был обновлен 23 августа 2018 года и включал в себя «более 850 организаций из почти 100 стран».
- ^ Zhu J, Hladik F, Woodward A, Klock A, Peng T, Johnston C, et al. (Август 2009 г.). «Стабильность рецептор-позитивных клеток ВИЧ-1 после реактивации HSV-2 является потенциальным механизмом повышенного приобретения ВИЧ-1» . Природная медицина . 15 (8): 886–92. doi : 10.1038/nm.2006 . PMC 2723183 . PMID 19648930 .
- ^ Looker KJ, Elmes JA, Gottlieb SL, Schiffer JT, Vickerman P, Turner KM, Boily MC (декабрь 2017 г.). «Влияние инфекции HSV-2 на последующее приобретение ВИЧ: обновленный систематический обзор и метаанализ» . Lancet. Заразительные заболевания . 17 (12): 1303–1316. doi : 10.1016/s1473-3099 (17) 30405-x . PMC 5700807 . PMID 28843576 .
- ^ "В этот день". Новости и запись . 18 мая 2020 года. С. 2а.
- ^ Манделл Г.Л., Беннетт Дж., Долин Р., ред. (2010). «Глава 169». Манделл, Дуглас и принципы Беннетта и практика инфекционных заболеваний (7 -е изд.). Филадельфия: Черчилль Ливингстон/Эльмвиер. ISBN 978-0-443-06839-3 . [ страница необходима ]
- ^ Gottlieb MS (2006). «Pneumocystis Pneumonia - Los Angeles. 1981» . Американский журнал общественного здравоохранения . 96 (6): 980–1, обсуждение 982–3. doi : 10.2105/ajph.96.6.980 . PMC 1470612 . PMID 16714472 . Архивировано с оригинала 22 апреля 2009 года.
- ^ Фридман-Киен А.Е. (октябрь 1981 г.). «Распространенный синдром саркомы Капоси у молодых гомосексуальных мужчин». Журнал Американской академии дерматологии . 5 (4): 468–71. doi : 10.1016/s0190-9622 (81) 80010-2 . PMID 7287964 .
- ^ Hymes KB, Cheung T, Greene JB, Prose NS, Marcus A, Ballard H, William DC, Laubenstein LJ (сентябрь 1981). «Саркома Капоси у гомосексуальных мужчин - отчет о восьми случаях». Lancet . 2 (8247): 598–600. doi : 10.1016/s0140-6736 (81) 92740-9 . PMID 6116083 . S2CID 43529542 .
- ^ Jump up to: а беременный Basavapatruni A, Anderson KS (декабрь 2007 г.). «Обратная транскрипция пандемии ВИЧ-1» . Журнал FASEB . 21 (14): 3795–3808. doi : 10.1096/fj.07-8697rev . PMID 17639073 . S2CID 24960391 .
- ^ Ледерберг Дж, изд. (2000). Энциклопедия микробиологии (2 -е изд.). Берлингтон: Elsevier. п. 106. ISBN 978-0-08-054848-7 Полем Получено 9 июня 2016 года .
- ^ Центры по борьбе с заболеваниями (1982). «Постоянная, генерализованная лимфаденопатия среди гомосексуальных мужчин» . Еженедельный отчет о заболеваемости и смертности . 31 (19): 249–251. PMID 6808340 .
- ^ Jump up to: а беременный Barré-Sinesi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, Dauguet C, Axler-Blin C, Vézinet-Brun F, Rouzioux C, Rozenbaum W, Montagnier L (1983). «Выделение T-лимфотропного ретровируса от пациента, подвергающегося риску синдрома приобретенного иммунного дефицита (СПИД)». Наука . 220 (4599): 868–871. Bibcode : 1983sci ... 220..868b . doi : 10.1126/science.6189183 . PMID 6189183 . S2CID 390173 .
- ^ Jump up to: а беременный Центры по борьбе с заболеваниями (1982). «Оппортунистические инфекции и саркома Капоси среди гаитян в Соединенных Штатах» . Еженедельный отчет о заболеваемости и смертности . 31 (26): 353–354, 360–361. PMID 6811853 .
- ^ Альтман Л.К. (11 мая 1982 г.). «Новые гомосексуальные расстройства беспокоятся о чиновниках здравоохранения» . New York Times . Получено 31 августа 2011 года .
- ^ Гилман С.Л. (1987). Гилмансл (ред.). «СПИД и сифилис: иконография болезней». Октябрь 43 : 87–107. doi : 10.2307/3397566 . JSTOR 3397566 .
- ^ «Приготовление вперед при уравновешенных обстоятельствах» (PDF) . Американская ассоциация по развитию науки . 28 июля 2006 года. Архивировано из оригинала (PDF) 24 июня 2008 года . Получено 23 июня 2008 года .
- ^ Хер У (27 июля 1982 г.). «Имя чумы» . Время . Архивировано из оригинала 7 марта 2008 года . Получено 10 марта 2008 года .
- ^ Центры по борьбе с заболеваниями (1982). «Обновление на синдроме приобретенного иммунного дефицита (СПИД) - государства». Еженедельный отчет о заболеваемости и смертности . 31 (37): 507–508, 513–514. PMID 6815471 .
- ^ Галло Р.К., Сарин П.С., Гелманн Э.П., Роберт-Гурофф М., Ричардсон Э., Калянараман В.С., Манн Д., Сидху Г.Д., Шталь Р.Е., Золла-Пазнер С., Лейбович Дж., Попович М. (1983). «Выделение вируса Т-клеточного лейкоза человека при синдроме приобретенного иммунного дефицита (СПИД)». Наука . 220 (4599): 865–867. Bibcode : 1983sci ... 220..865G . doi : 10.1126/science.6601823 . PMID 6601823 .
- ^ «Нобелевская премия 2008 года в области физиологии или медицины - пресс -релиз» . www.nobelprize.org . Получено 28 января 2018 года .
- ^ Crewdson J (30 мая 1991 г.). «Галло признает, что Френч обнаружил вирус СПИДа» . Чикаго Трибьюн . Получено 25 апреля 2020 года .
- ^ Олдрич Р., Уотерспун Г., ред. (2001). Кто есть кто в истории геев и лесбиянок . Лондон: Routledge. п. 154. ISBN 978-0-415-22974-6 .
- ^ Леви Дж.А. и др. (1984). «Выделение лимфоцитопатических ретровирусов от пациентов Сан -Франциско со СПИДом». Наука . 225 (4664): 840–842. Bibcode : 1984sci ... 225..840L . doi : 10.1126/science.6206563 . PMID 6206563 .
- ^ Леви Дж.А., Камински Л.С., Морроу В.Дж., Стеймер К., Люси П., Дина Д., Хокси Дж., Оширо Л. (1985). «Инфекция ретровирусом, связанным с синдромом приобретенного иммунодефицита». Анналы внутренней медицины . 103 (5): 694–699. doi : 10.7326/0003-4819-103-5-694 . PMID 2996401 .
- ^ Sharp PM, Hahn BH (2011). «Происхождение ВИЧ и пандемии СПИДа» . Перспективы Cold Spring Harbor в медицине . 1 (1): A006841. doi : 10.1101/cshperspect.a006841 . PMC 3234451 . PMID 22229120 .
- ^ Фария Н.Р., Рамбаут А., Сучерд М.А., Баеле Г., Бедфорд Т., Уорд М.Дж., Татем А.Дж., Соуза Д.Д., Аринаминпати Н., Пепин Дж., Посада Д., Питер М., Пибус Ог, Лемей П (2014). «Раннее распространение и эпидемическое воспламенение ВИЧ-1 в популяциях человека» . Наука . 346 (6205): 56–61. Bibcode : 2014sci ... 346 ... 56f . doi : 10.1126/science.1256739 . PMC 4254776 . PMID 25278604 .
- ^ Gao F, Bailes E, Robertson DL, Chen Y, Rodenburg CM, Michael SF, Cummins LB, Arthur LO, Peeters M, Shaw GM, Sharp PM, Hahn BH (1999). «Происхождение ВИЧ-1 в шимпанзе Пан Троглодиты Троглодиты» . Природа . 397 (6718): 436–41. Bibcode : 1999natur.397..436G . doi : 10.1038/17130 . PMID 9989410 . S2CID 4432185 .
- ^ Keele Bf, Van Heuverswyn F, Li Y, Bailes E, Takehisa J, Santiago Ml, Bibollet-ruche F, Chen Y, Wain Lv, Liegeois F, Loul S, Ngole EM, Bienvenue Y, Delaporte E, Brookfield JF, Sharp PM , Shaw GM, Peeters M, Hahn BH (2006). «Шимпанзе водохранилища пандемии и непенемического ВИЧ-1» . Наука . 313 (5786): 523–6. Bibcode : 2006sci ... 313..523K . doi : 10.1126/science.1126531 . PMC 2442710 . PMID 16728595 .
- ^ Goodier JL, Kazazian HH (2008). «Ретротранспозоны пересмотрены: сдержанность и реабилитация паразитов» . Клетка . 135 (1): 23–35. doi : 10.1016/j.cell.2008.09.022 . PMID 18854152 . S2CID 3093360 .
- ^ Sharp PM, Bailes E, Chaudhuri RR, Rodenburg CM, Santiago MO, Hahn BH (2001). «Происхождение вирусов синдрома приобретенного иммунного дефицита: где и когда?» Полем Философские транзакции Королевского общества б . 356 (1410): 867–76. doi : 10.1098/rstb.2001.0863 . PMC 1088480 . PMID 11405934 .
- ^ Калиш М.Л., Вулф Н.Д., Ндонгмо К.Б., Макнихолл Дж., Роббинс К.Е., Эйду М., Фонджунго П.Н., Алеммджи Г., Зех С., Джоко К.Ф., Мподи-Нголе Е., Берк Д.С., Релькс Т.М. (2005). «Охотники за центральной африкой подвергаются воздействию вируса иммунодефицита об одномине» . Возникающие инфекционные заболевания . 11 (12): 1928–30. doi : 10.3201/eid1112.050394 . PMC 3367631 . PMID 16485481 .
- ^ Jump up to: а беременный Marx PA, Alcabes PG, Drucker E (2001). «Серийный проход человека вируса иммунодефицита об одномине с помощью нестерильных инъекций и появление эпидемического вируса иммунодефицита человека в Африке» . Философские транзакции Королевского общества б . 356 (1410): 911–20. doi : 10.1098/rstb.2001.0867 . PMC 1088484 . PMID 11405938 .
- ^ Worobey M, Gemmel M, Teuwen DE, Haselkorn T, Kunstman K, Bunce M, Muyembe JJ, Kabongo JM, Kalengayi RM, Van Marck E, Gilbert MT, Wolinsky SM (2008). «Прямые доказательства обширного разнообразия ВИЧ-1 в Киншасе к 1960 году» . Природа . 455 (7213): 661–4. Bibcode : 2008natur.455..661W . doi : 10.1038/nature07390 . PMC 3682493 . PMID 18833279 .
- ^ Jump up to: а беременный De Sousa JD, Müller V, Lemey P, Vandamme AM (2010). Мартин Д.П. (ред.). «Высокая заболеваемость GUD в начале 20 -го века создало особенно допустимое временное окно для происхождения и начального распространения эпидемических штаммов ВИЧ» . Plos один . 5 (4): E9936. Bibcode : 2010ploso ... 5,9936S . doi : 10.1371/journal.pone.0009936 . PMC 2848574 . PMID 20376191 .
- ^ Zhu T, Korber BT, Nahmias AJ, Hooper E, Sharp PM, Ho DD (1998). «Африканская последовательность ВИЧ-1 с 1959 года и последствия для происхождения эпидемии» . Природа . 391 (6667): 594–7. Bibcode : 1998natur.391..594Z . doi : 10.1038/35400 . PMID 9468138 . S2CID 4416837 .
- ^ Kolata G (28 октября 1987 г.). «Смерть мальчика 1969 года предполагает, что СПИД вторгалась на нас несколько раз» . New York Times . Получено 11 февраля 2009 г.
- ^ Chitnis A, Rawls D, Moore J (январь 2000 г.). «Происхождение ВИЧ типа 1 в колониальной французской экваториальной Африке?». Исследование СПИДа и ретровирусы человека . 16 (1): 5–8. doi : 10.1089/088922200309548 . PMID 10628811 . S2CID 17783758 .
- ^ McNeil D Jr (16 сентября 2010 г.). «Предшественник ВИЧ был в обезьянах на протяжении тысячелетий» . New York Times . Архивировано из оригинала 3 января 2022 года . Получено 17 сентября 2010 года .
Доктор Маркс считает, что важным событием было введение в Африку миллионов недорогих, массовых продуцированных шприцев в 1950-х годах. ... подозревает, что рост колониальных городов виноват. До 1910 года ни в одном из центральной африканской города не было более 10 000 человек. Но городская миграция выросла, увеличивая сексуальные контакты и приводя к районам красного света.
- ^ Jump up to: а беременный Wertheim Jo, Worobey M (май 2009 г.). «Датируется возрастом линий SIV, которые породили ВИЧ-1 и ВИЧ-2» . PLOS Computational Biology . 5 (5): E1000377. BIBCODE : 2009PLSCB ... 5E0377W . doi : 10.1371/journal.pcbi.1000377 . PMC 2669881 . PMID 19412344 .
Дальнейшее чтение
- Berlier W, Bourlet T, Lawrence P, Hamzeh H, Lambert C, Genin C, Verrier B, Diu-Nosjean MC, Pozzetto B, Delezay O (2005). «Селективная секвестрация изолятов x4 генитальными эпителиальными клетками человека: значение для процесса отбора вируса тропизма во время сексуальной передачи ВИЧ». Журнал медицинской вирусологии . 77 (4): 465–74. doi : 10.1002/jmv.20478 . PMID 16254974 . S2CID 25762969 .
- Объединенная программа Организации Объединенных Наций по ВИЧ/СПИДу (UNAIDS) (2011). Глобальная реакция на ВИЧ/СПИД, обновление эпидемии и сектор здравоохранения прогрессирует в направлении универсального доступа (PDF) . Объединенная программа Организации Объединенных Наций по ВИЧ/СПИДу.
- Muciaccia B, Padula F, Vicini E, Gandini L, Lenzi A, Stefanini M (2005). «Рецепторы бета-хемокинов 5 и 3 экспрессируются в области головы человеческого сперматозона» . Журнал FASEB . 19 (14): 2048–50. doi : 10.1096/fj.05-3962fje . HDL : 11573/361629 . PMID 16174786 . S2CID 7928126 .
Внешние ссылки
 Определения из Wiktionary
Определения из Wiktionary  СМИ из Commons
СМИ из Commons  Новости от Wikinews
Новости от Wikinews  Учебники из Wikibooks
Учебники из Wikibooks