CD86
| CD86 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CD86 , B7-2, B7.2, B70, CD28LG2, LAB72, молекула CD86 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 601020 ; МГИ : 101773 ; Гомологен : 10443 ; Генные карты : CD86 ; ОМА : CD86 – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





Кластер дифференциации 86 (также известный как CD86 и B7-2 ) представляет собой белок, конститутивно экспрессируемый на дендритных клетках , клетках Лангерганса , макрофагах , B-клетках (включая B-клетки памяти ) и на других антигенпрезентирующих клетках . [ 5 ] Наряду с CD80 , CD86 обеспечивает костимулирующие сигналы , необходимые для Т-клеток активации и выживания . В зависимости от связанного лиганда CD86 может сигнализировать о саморегуляции и межклеточной ассоциации или об ослаблении регуляции и межклеточной диссоциации. [ 6 ]
Ген CD86 , кодирует мембранный белок типа I который является членом суперсемейства иммуноглобулинов . [ 7 ] Альтернативный сплайсинг приводит к образованию двух вариантов транскриптов, кодирующих разные изоформы . Описаны дополнительные варианты транскриптов, но их полноразмерные последовательности не определены. [ 8 ]
Структура
[ редактировать ]CD86 принадлежит к семейству B7 суперсемейства иммуноглобулинов. [ 9 ] Это массой 70 кДа, гликопротеин состоящий из 329 аминокислот . И CD80, и CD86 имеют общий консервативный аминокислотный мотив, который образует их лигандсвязывающий домен . [ 10 ] CD86 состоит из Ig-подобных внеклеточных доменов (одного вариабельного и одного константного), трансмембранной области и короткого цитоплазматического домена, который длиннее, чем у CD80. [ 11 ] [ 12 ] костимулирующие лиганды CD80 и CD86 можно обнаружить на профессиональных антигенпредставляющих клетках, таких как моноциты , дендритные клетки и даже активированные B-клетки. Их также можно индуцировать на других типах клеток, например на Т-клетках. [ 13 ] Экспрессия CD86 более выражена по сравнению с CD80, и при его активации CD86 увеличивается быстрее, чем CD80. [ 14 ]
На уровне белка CD86 на 25% идентичен CD80. [ 15 ] и оба закодированы на человеческой хромосоме 3q13.33q21. [ 16 ]
Роль в совместной стимуляции, активации и ингибировании Т-клеток
[ редактировать ]CD86 и CD80 связываются в качестве лигандов с костимулирующей молекулой CD28 на поверхности всех наивных Т-клеток . [ 17 ] и к ингибирующему рецептору CTLA-4 (цитотоксический Т-лимфоцитарный антиген-4, также известный как CD152). [ 18 ] [ 19 ] CD28 и CTLA-4 играют важную, но противоположную роль в стимуляции Т-клеток. Связывание с CD28 способствует Т-клеточным ответам, а связывание с CTLA-4 ингибирует их. [ 20 ]
Взаимодействие между CD86 ( CD80 ), экспрессируемым на поверхности антигенпредставляющей клетки, с CD28 на поверхности зрелой, наивной Т-клетки, необходимо для активации Т-клеток. [ 21 ] Чтобы активироваться, лимфоцит должен задействовать как антиген, так и костимулирующий лиганд на одной и той же антигенпрезентирующей клетке. Т-клеточный рецептор (TCR) взаимодействует с молекулами главного комплекса гистосовместимости (MHC) класса II . [ 13 ] и эта сигнализация должна сопровождаться костимулирующими сигналами, обеспечиваемыми костимулирующим лигандом. Эти костимулирующие сигналы необходимы для предотвращения анергии и обеспечиваются взаимодействием между CD80/CD86 и костимулирующей молекулой CD28. [ 22 ] [ 23 ]
Это белковое взаимодействие также важно для получения Т-лимфоцитами полного сигнала активации, что, в свою очередь, приводит к дифференцировке и делению Т-клеток, выработке интерлейкина 2 и клональной экспансии. [ 9 ] [ 22 ] Взаимодействие между CD86 и CD28 активирует митоген-активируемую протеинкиназу и фактор транскрипции nf-κB в Т-клетке. Эти белки усиливают выработку CD40L (используемого при активации B-клеток), IL-21 и IL-21R (используемого для деления/пролиферации) и IL-2 , а также других цитокинов. [ 21 ] Взаимодействие также регулирует аутотолерантность, поддерживая гомеостаз CD4+CD25+ Tregulatory клеток , также известных как Treg. [ 9 ]
CTLA-4 представляет собой коингибирующую молекулу, которая индуцируется активированными Т-клетками. Взаимодействие CTLA-4 и CD80/CD86 приводит к доставке негативных сигналов в Т-клетки и уменьшению количества костимулирующих молекул на поверхности клетки. Он также может запускать сигнальный путь, ответственный за экспрессию фермента IDO (индоламин-2,3-диоксигеназы). Этот фермент может метаболизировать аминокислоту триптофан , которая является важным компонентом успешной пролиферации и дифференцировки Т-лимфоцитов. IDO снижает концентрацию триптофана в окружающей среде, тем самым подавляя активацию обычных Т-клеток, а также стимулируя функцию регуляторных Т-клеток. [ 24 ] [ 25 ]
И CD80, и CD86 связывают CTLA-4 с более высоким сродством, чем CD28. Это позволяет CTLA-4 конкурировать с CD28 за связывание CD80/CD86. [ 23 ] [ 26 ] Между CD80 и CD86 CD80, по-видимому, имеет более высокое сродство как к CTLA-4, так и к CD28, чем к CD86. Это позволяет предположить, что CD80 является более мощным лигандом, чем CD86. [ 15 ] но исследования с использованием мышей с нокаутом CD80 и CD86 показали, что CD86 более важен в активации Т-клеток, чем CD80. [ 27 ]
Трег-посредничество
[ редактировать ]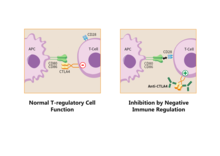
Пути семейства B7:CD28 играют ключевую роль в регуляции активации и толерантности Т-клеток. Их отрицательные вторые сигналы ответственны за подавление клеточных ответов. По всем этим причинам эти пути считаются терапевтическими мишенями. [ 9 ]
Регуляторные Т-клетки производят CTLA-4 . Благодаря взаимодействию с CD80/CD86 Treg могут конкурировать с обычными Т-клетками и блокировать их костимулирующие сигналы. Экспрессия Treg CTLA-4 может эффективно подавлять как CD80, так и CD86 на APC. [ 28 ] подавляют иммунный ответ и приводят к усилению анергии . [ 6 ] Поскольку CTLA-4 связывается с CD86 с более высоким сродством, чем с CD28, это также влияет на костимуляцию, необходимую для правильной активации Т-клеток. [ 29 ] В работе группы Сагурачи было показано, что Treg-клетки способны подавлять CD80 и CD86, но не CD40 или MHC класса II на DC, способом, который зависит от адгезии. Понижающая регуляция блокировалась антителом против CTLA-4 и отменялась, если Treg-клетки имели дефицит CTLA-4. [ 30 ]
Связанный с CTLA-4, CD86 может быть удален с поверхности APC и на Treg-клетку в процессе, называемом трогоцитозом . [ 6 ] Блокирование этого процесса с помощью антител против CTLA-4 полезно для определенного типа иммунотерапии рака, называемого «терапия рака путем ингибирования негативной иммунной регуляции». [ 31 ] Японский иммунолог Тасуку Хондзё и американский иммунолог Джеймс П. Эллисон получили Нобелевскую премию по физиологии и медицине в 2018 году за свою работу по этой теме.
Роль в патологии
[ редактировать ]Роли CD80 и CD86 изучаются в контексте многих патологий. Селективное ингибирование костимулирующих ингибиторов исследовали на модели аллергического воспаления легких и гиперреактивности дыхательных путей (AHR). [ 32 ] Поскольку первоначальный ответ хозяина на Staphylococcus aureus , особенно иммунный ответ, основанный на Т-клетках, является фактором, способствующим патогенезу острой пневмонии , была исследована роль пути CD80/CD86 в патогенезе. [ 33 ] Костимулирующие молекулы также исследовались в контексте бронхиальной астмы . [ 34 ] Трег при раке , [ 35 ] и иммунотерапия . [ 36 ]
См. также
[ редактировать ]- Кластер дифференциации
- CD80
- CD28
- КТЛА-4
- Список человеческих кластеров дифференцировки для списка молекул CD
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000114013 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000022901 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Леншоу Д.Д., Су Г.Х., Цукерман Л.А., Набави Н., Джеллис К.Л., Грей Г.С. и др. (декабрь 1993 г.). «Экспрессия и функциональное значение дополнительного лиганда для CTLA-4» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (23): 11054–8. Бибкод : 1993PNAS...9011054L . дои : 10.1073/pnas.90.23.11054 . ПМК 47920 . ПМИД 7504292 .
- ^ Перейти обратно: а б с Оуэ Ю., Нисикава Х. (июль 2019 г.). «Регуляторные Т-(Treg)-клетки при раке: могут ли Treg-клетки стать новой терапевтической мишенью?» . Раковая наука . 110 (7): 2080–2089. дои : 10.1111/cas.14069 . ПМК 6609813 . ПМИД 31102428 .
- ^ Чен С., Голт А., Шен Л., Набави Н. (май 1994 г.). «Молекулярное клонирование и экспрессия ранней костимулирующей молекулы Т-клеток-1 и ее характеристика как молекулы B7-2» . Журнал иммунологии . 152 (10): 4929–36. дои : 10.4049/jimmunol.152.10.4929 . ПМИД 7513726 . S2CID 22260156 .
- ^ «Ген Энтрез: молекула CD86 CD86» .
- ^ Перейти обратно: а б с д Гринвальд Р.Дж., Фриман Г.Дж., Шарп А.Х. (2005). «Возвращение к семье B7». Ежегодный обзор иммунологии . 23 : 515–48. doi : 10.1146/annurev.immunol.23.021704.115611 . ПМИД 15771580 .
- ^ Ю С., Соннен А.Ф., Джордж Р., Дессайи Б.Х., Стэгг Л.Дж., Эванс Э.Дж. и др. (февраль 2011 г.). «Распознавание лиганда с жестким телом приводит к срабатыванию рецептора цитотоксического антигена Т-лимфоцитов 4 (CTLA-4)» . Журнал биологической химии . 286 (8): 6685–96. дои : 10.1074/jbc.M110.182394 . ПМК 3057841 . ПМИД 21156796 .
- ^ Фриман Г.Дж., Борриелло Ф., Ходес Р.Дж., Райзер Х., Хэткок К.С., Ласло Г. и др. (ноябрь 1993 г.). «Обнаружение функционального альтернативного контррецептора CTLA-4 у мышей с дефицитом B7». Наука . 262 (5135): 907–9. Бибкод : 1993Sci...262..907F . дои : 10.1126/science.7694362 . ПМИД 7694362 .
- ^ Шарп А.Х., Фриман Г.Дж. (февраль 2002 г.). «Суперсемейство B7-CD28». Обзоры природы. Иммунология . 2 (2): 116–26. дои : 10.1038/nri727 . ПМИД 11910893 . S2CID 205492817 .
- ^ Перейти обратно: а б Мерфи К., Уивер С., Джейнвей С. (2017). Иммунобиология Джейнвей (9-е изд.). Нью-Йорк. ISBN 978-0-8153-4505-3 . OCLC 933586700 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Сансом ДМ (октябрь 2000 г.). «CD28, CTLA-4 и их лиганды: кто, что и с кем делает?» . Иммунология . 101 (2): 169–77. дои : 10.1046/j.1365-2567.2000.00121.x . ПМК 2327073 . ПМИД 11012769 .
- ^ Перейти обратно: а б Коллинз А.В., Броди Д.В., Гилберт Р.Дж., Ябони А., Мансо-Санчо Р., Уолс Б. и др. (август 2002 г.). «Возврат к свойствам взаимодействия костимулирующих молекул» . Иммунитет . 17 (2): 201–10. дои : 10.1016/s1074-7613(02)00362-x . ПМИД 12196291 .
- ^ Мир МА (25 мая 2015 г.). Разработка костимулирующих молекул для иммунотерапии заболеваний . Лондон. ISBN 978-0-12-802675-5 . OCLC 910324332 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Линсли П.С., Брэди В., Гросмэр Л., Аруффо А., Дамл Н.К., Ледбеттер Дж.А. (март 1991 г.). «Связывание антигена активации В-клеток B7 с CD28 костимулирует пролиферацию Т-клеток и накопление мРНК интерлейкина 2» . Журнал экспериментальной медицины . 173 (3): 721–30. дои : 10.1084/jem.173.3.721 . ПМК 2118836 . ПМИД 1847722 .
- ^ Лим Т.С., Го Дж.К., Мортелларо А., Лим К.Т., Хеммерлинг Г.Дж., Риккарди-Кастаньоли П. (2012). «CD80 и CD86 по-разному регулируют механические взаимодействия Т-клеток с антигенпредставляющими дендритными клетками и В-клетками» . ПЛОС ОДИН . 7 (9): е45185. Бибкод : 2012PLoSO...745185L . дои : 10.1371/journal.pone.0045185 . ПМЦ 3443229 . ПМИД 23024807 .
- ^ Линсли П.С., Брэди В., Урнес М., Гросмэр Л.С., Дамл Н.К., Ледбеттер Дж.А. (сентябрь 1991 г.). «CTLA-4 является вторым рецептором антигена активации B7 B7» . Журнал экспериментальной медицины . 174 (3): 561–9. дои : 10.1084/jem.174.3.561 . ПМК 2118936 . ПМИД 1714933 .
- ^ Сансом Д.М., Манзотти С.Н., Чжэн Ю (июнь 2003 г.). «В чем разница между CD80 и CD86?». Тенденции в иммунологии . 24 (6): 314–9. дои : 10.1016/s1471-4906(03)00111-x . ПМИД 12810107 .
- ^ Перейти обратно: а б Дайк Л., Миллс К.Х. (май 2017 г.). «Иммунные контрольные точки и их ингибирование при раке и инфекционных заболеваниях» . Европейский журнал иммунологии . 47 (5): 765–779. дои : 10.1002/eji.201646875 . ПМИД 28393361 .
- ^ Перейти обратно: а б Койл А.Дж., Гутьеррес-Рамос Дж.К. (март 2001 г.). «Расширяющееся суперсемейство B7: возрастающая сложность костимулирующих сигналов, регулирующих функцию Т-клеток». Природная иммунология . 2 (3): 203–9. дои : 10.1038/85251 . ПМИД 11224518 . S2CID 20542148 .
- ^ Перейти обратно: а б Гаус В.К., Урбан Дж.Ф., Линсли П., Лу П. (1995). «Роль передачи сигналов B7 в дифференцировке наивных CD4 + Т-клеток в эффекторные Т-хелперные клетки, продуцирующие интерлейкин-4» . Иммунологические исследования . 14 (3): 176–88. дои : 10.1007/BF02918215 . ПМИД 8778208 . S2CID 20098311 .
- ^ Чен Л., Flies DB (апрель 2013 г.). «Молекулярные механизмы костимуляции и коингибирования Т-клеток» . Обзоры природы. Иммунология . 13 (4): 227–42. дои : 10.1038/nri3405 . ПМЦ 3786574 . ПМИД 23470321 .
- ^ Манн Д.Х., Шарма, доктор медицинских наук, Меллор А.Л. (апрель 2004 г.). «Лигирование B7-1/B7-2 CD4+ Т-клетками человека запускает активность индоламин-2,3-диоксигеназы в дендритных клетках» . Журнал иммунологии . 172 (7): 4100–10. дои : 10.4049/jimmunol.172.7.4100 . ПМИД 15034022 .
- ^ Уокер Л.С., Сансом DM (ноябрь 2011 г.). «Новая роль CTLA4 как внешнего регулятора Т-клеточных ответов». Обзоры природы. Иммунология . 11 (12): 852–63. дои : 10.1038/nri3108 . ПМИД 22116087 . S2CID 9617595 .
- ^ Борриелло Ф., Сетна М.П., Бойд С.Д., Швейцер А.Н., Тивол Э.А., Джейкоби Д. и др. (март 1997 г.). «B7-1 и B7-2 играют критически важную роль в переключении классов иммуноглобулинов и формировании зародышевых центров» . Иммунитет . 6 (3): 303–13. дои : 10.1016/s1074-7613(00)80333-7 . ПМИД 9075931 .
- ^ Уокер Л.С., Сансом Д.М. (февраль 2015 г.). «Сбивающие с толку сигналы: недавний прогресс в биологии CTLA-4» . Тенденции в иммунологии . 36 (2): 63–70. дои : 10.1016/j.it.2014.12.001 . ПМЦ 4323153 . ПМИД 25582039 .
- ^ Лайтман С.М., Атли А., Ли КП (3 мая 2019 г.). «Выживание долгоживущих плазматических клеток (LLPC): собираем головоломку» . Границы в иммунологии . 10 :965. дои : 10.3389/fimmu.2019.00965 . ПМК 6510054 . ПМИД 31130955 .
- ^ Ониши И., Фехервари З., Ямагути Т., Сакагути С. (июль 2008 г.). «Естественные регуляторные Т-клетки Foxp3+ преимущественно образуют агрегаты на дендритных клетках in vitro и активно ингибируют их созревание» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (29): 10113–8. Бибкод : 2008PNAS..10510113O . дои : 10.1073/pnas.0711106105 . ПМЦ 2481354 . ПМИД 18635688 .
- ^ Чен Р., Ганесан А., Окойе И., Арутюнова Е., Элахи С., Лемье М.Дж. и др. (март 2020 г.). «Нацеливание на B7-1 в иммунотерапии». Обзоры медицинских исследований . 40 (2): 654–682. дои : 10.1002/мед.21632 . PMID 31448437 . S2CID 201748060 .
- ^ Марк Д.А., Донован С.Э., Де Санктис Г.Т., Кринцман С.Дж., Кобзик Л., Линсли П.С. и др. (ноябрь 1998 г.). «И CD80, и CD86 костимулирующие молекулы регулируют аллергическое воспаление легких» . Международная иммунология . 10 (11): 1647–55. дои : 10.1093/интимм/10.11.1647 . ПМИД 9846693 .
- ^ Паркер Д. (июль 2018 г.). «Передача сигналов CD80/CD86 способствует провоспалительной реакции золотистого стафилококка в дыхательных путях» . Цитокин . 107 : 130–136. дои : 10.1016/j.cyto.2018.01.016 . ПМК 5916031 . ПМИД 29402722 .
- ^ Чен Ю., Ши Х.З. (январь 2006 г.). «Костимулирующий путь CD28/CTLA-4-CD80/CD86 и ICOS-B7RP-1 при бронхиальной астме». Аллергия . 61 (1): 15–26. дои : 10.1111/j.1398-9995.2006.01008.x . ПМИД 16364152 . S2CID 23564785 .
- ^ Оуэ Ю., Нисикава Х. (июль 2019 г.). «Регуляторные Т-(Treg)-клетки при раке: могут ли Treg-клетки стать новой терапевтической мишенью?» . Раковая наука . 110 (7): 2080–2089. дои : 10.1111/cas.14069 . ПМК 6609813 . ПМИД 31102428 .
- ^ Бурк Дж., Хавигер Д. (2018). «Иммуномодулирующие связи партнерства между дендритными клетками и Т-клетками» . Критические обзоры по иммунологии . 38 (5): 379–401. doi : 10.1615/CritRevImmunol.2018026790 . ПМК 6380512 . ПМИД 30792568 .
Внешние ссылки
[ редактировать ]Дальнейшее чтение
[ редактировать ]- Давила С., Фрёлинг Ф.Е., Тан А., Боннард С., Боланд Г.Дж., Сниппе Х. и др. (апрель 2010 г.). «Новые генетические ассоциации обнаружены в исследовании реакции организма на вакцину против гепатита В» . Гены и иммунитет . 11 (3): 232–8. дои : 10.1038/gen.2010.1 . ПМИД 20237496 .
- Чиллаг А., Болдох И., Пазманди К., Мадьярикс З., Гоголак П., Сур С. и др. (март 2010 г.). «Вызванный пыльцой окислительный стресс влияет как на врожденный, так и на адаптивный иммунный ответ посредством изменения функций дендритных клеток» . Журнал иммунологии . 184 (5): 2377–85. doi : 10.4049/jimmunol.0803938 . ПМК 3028537 . ПМИД 20118277 .
- Боссе Ю., Лемир М., Пун А.Х., Дейли Д., Хе Дж.К., Сэндфорд А. и др. (октябрь 2009 г.). «Астма и гены, кодирующие компоненты пути витамина D» . Респираторные исследования . 10 (1): 98. дои : 10.1186/1465-9921-10-98 . ПМК 2779188 . ПМИД 19852851 .
- Мосбругер Т.Л., Дуггал П., Годерт Дж.Дж., Кирк Г.Д., Хутс В.К., Тоблер Л.Х. и др. (май 2010 г.). «Крупномасштабный анализ генов-кандидатов спонтанного клиренса вируса гепатита С» . Журнал инфекционных болезней . 201 (9): 1371–80. дои : 10.1086/651606 . ПМЦ 2853721 . ПМИД 20331378 .
- Буджен Л., Даллман М.Дж. (октябрь 2000 г.). «Костимуляция Т-клеток». Американский журнал респираторной медицины и медицины интенсивной терапии . 162 (4 ч. 2): S164-8. doi : 10.1164/ajrccm.162.supplement_3.15tac5 . ПМИД 11029388 .
- Пан ХМ, Гао ЛБ, Лян ВБ, Лю Ю, Чжу Ю, Тан М и др. (июль 2010 г.). «Полиморфизм CD86 +1057 G/A и риск колоректального рака». ДНК и клеточная биология . 29 (7): 381–6. дои : 10.1089/dna.2009.1003 . ПМИД 20380573 .
- Далла-Коста Р., Пинсерати М.Р., Бельтраме М.Х., Малхейрос Д., Петцль-Эрлер М.Л. (август 2010 г.). «Полиморфизмы в областях хромосом 2q33 и 3q21, включая корецепторы Т-клеток и гены-лиганды, могут влиять на восприимчивость к листовидной пузырчатке». Иммунология человека . 71 (8): 809–17. дои : 10.1016/j.humimm.2010.04.001 . ПМИД 20433886 .
- Талмуд П.Дж., Дренос Ф., Шах С., Шах Т., Палмен Дж., Верзилли С. и др. (ноябрь 2009 г.). «Сигналы геноцентрической ассоциации липидов и аполипопротеинов, идентифицированные с помощью HumanCVD BeadChip» . Американский журнал генетики человека . 85 (5): 628–42. дои : 10.1016/j.ajhg.2009.10.014 . ПМЦ 2775832 . ПМИД 19913121 .
- Карреньо Л.Дж., Пачеко Р., Гутьеррес М.А., Якобелли С., Калергис А.М. (ноябрь 2009 г.). «Активность заболевания при системной красной волчанке связана с измененной экспрессией низкоаффинных гамма-рецепторов Fc и костимулирующих молекул на дендритных клетках» . Иммунология . 128 (3): 334–41. дои : 10.1111/j.1365-2567.2009.03138.x . ПМК 2770681 . ПМИД 20067533 .
- Коясу С. (апрель 2003 г.). «Роль PI3K в иммунных клетках». Природная иммунология . 4 (4): 313–9. дои : 10.1038/ni0403-313 . ПМИД 12660731 . S2CID 9951653 .
- Ким Ш., Ли Дж.Э., Ким Ш., Джи Ю.К., Ким Ю.К., Пак Х.С. и др. (декабрь 2009 г.). «Аллельные варианты генов CD40 и CD40L взаимодействуют, вызывая кожные аллергические реакции, вызванные антибиотиками». Клиническая и экспериментальная аллергия . 39 (12): 1852–6. дои : 10.1111/j.1365-2222.2009.03336.x . ПМИД 19735272 . S2CID 26024387 .
- Лю Ю, Лян ВБ, Гао ЛБ, Пань ХМ, Чен Тюй, Ван Юй и др. (ноябрь 2010 г.). «Полиморфизм генов CTLA4 и CD86 и предрасположенность к хронической обструктивной болезни легких». Иммунология человека . 71 (11): 1141–6. дои : 10.1016/j.humimm.2010.08.007 . ПМИД 20732370 .
- Ма С.Н., Ван Х., Ян Ю.И., Ян Л., Чжан Д.Л., Шэн Х. и др. (июнь 2010 г.). «Отсутствие связи между полиморфизмом CD86 + 1057G/A и ишемической болезнью сердца». ДНК и клеточная биология . 29 (6): 325–8. дои : 10.1089/dna.2009.0987 . ПМИД 20230296 .
- Ишизаки Ю, Юкая Н, Кусухара К, Кира Р, Торису Х, Ихара К и др. (апрель 2010 г.). «PD1 как общий ген-кандидат предрасположенности к подострому склерозирующему панэнцефалиту». Генетика человека . 127 (4): 411–9. дои : 10.1007/s00439-009-0781-z . ПМИД 20066438 . S2CID 12633836 .
- Чанг Т.Т., Качру В.К., Шарп А.Х. (2002). «Роль пути B7-CD28/CTLA-4 при аутоиммунных заболеваниях». Современные направления в области аутоиммунитета . 5 : 113–30. дои : 10.1159/000060550 . ISBN 3-8055-7308-1 . ПМИД 11826754 .
- Груич М., Бартольди С., Реми М., Пинщевер Д.Д., Кристенсен Дж.П., Томсен А.Р. (август 2010 г.). «Роль CD80/CD86 в генерации и поддержании функциональных вирус-специфичных CD8+ Т-клеток у мышей, инфицированных вирусом лимфоцитарного хориоменингита» . Журнал иммунологии . 185 (3): 1730–43. doi : 10.4049/jimmunol.0903894 . ПМИД 20601595 .
- Куаранта М.Г., Маттиоли Б., Джордани Л., Виора М. (ноябрь 2006 г.). «Имуннорегулирующее действие ВИЧ-1 Nef на дендритные клетки и патогенез СПИДа» . Журнал ФАСЭБ . 20 (13): 2198–208. дои : 10.1096/fj.06-6260рев . ПМИД 17077296 . S2CID 3111709 .
- Шуурхоф А., Бонт Л., Зизен К.Л., Ходемакерс Х., ван Хаувелинген Х.К., Кимман Т.Г. и др. (июнь 2010 г.). «Полиморфизм интерлейкина-9 у детей грудного возраста с респираторно-синцитиальной вирусной инфекцией: противоположный эффект у мальчиков и девочек». Детская пульмонология . 45 (6): 608–13. дои : 10.1002/ppul.21229 . ПМИД 20503287 . S2CID 24678182 .
- Бэйли С.Д., Се С., До Р., Монтпети А., Диас Р., Мохан В. и др. (октябрь 2010 г.). «Изменения в локусе NFATC2 увеличивают риск возникновения отека, вызванного тиазолидиндионом, в исследовании по оценке снижения диабета с помощью рамиприла и препарата росиглитазона (DREAM)» . Уход при диабете . 33 (10): 2250–3. дои : 10.2337/dc10-0452 . ПМЦ 2945168 . ПМИД 20628086 .
- Радзиевич Х., Ибегбу К.С., Хон Х., Бедар Н., Брюно Дж., Ворковски К.А. и др. (март 2010 г.). «Транзиторная экспрессия CD86 на CD8+ Т-клетках, специфичных для вируса гепатита С, при острой инфекции связана с достаточной передачей сигналов IL-2» . Журнал иммунологии . 184 (5): 2410–22. doi : 10.4049/jimmunol.0902994 . ПМЦ 2924663 . ПМИД 20100932 .
галерея PDB |
|---|


Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .