Абиогенез

Абиогенез — это естественный процесс, в ходе которого жизнь возникает из неживой материи , например простых органических соединений . Преобладающая научная гипотеза состоит в том, что переход от неживых существ к живым на Земле был не единичным событием, а процессом возрастающей сложности, включающим формирование обитаемой планеты , пребиотический синтез органических молекул, молекулярное самовоспроизведение , самовоспроизведение. -сборка , автокатализ и возникновение клеточных мембран . Переход от нежизни к жизни никогда не наблюдался экспериментально, но было сделано множество предположений для различных стадий этого процесса.
Изучение абиогенеза направлено на то, чтобы определить, как химические реакции до жизни привели к возникновению жизни в условиях, разительно отличающихся от тех, которые существуют на Земле сегодня. В основном он использует инструменты биологии и химии , а более поздние подходы пытаются синтезировать многие науки. Жизнь функционирует посредством специализированной химии углерода и воды и в основном строится на четырех ключевых семействах химических веществ: липидах для клеточных мембран, углеводах, таких как сахара, аминокислотах для белкового метаболизма, а также нуклеиновых кислотах ДНК и РНК для механизмов наследственности. Любая успешная теория абиогенеза должна объяснить происхождение и взаимодействие этих классов молекул.
Многие подходы к абиогенезу исследуют, как возникли самовоспроизводящиеся молекулы или их компоненты. Исследователи обычно полагают, что нынешняя жизнь произошла из мира РНК , хотя РНК могли предшествовать и другие самовоспроизводящиеся и самокаталитические молекулы. Другие подходы ( гипотезы «прежде всего метаболизма» ) сосредоточены на понимании того, как катализ в химических системах на ранней Земле мог обеспечить молекулы-предшественники, необходимые для самовоспроизведения. Классический эксперимент Миллера-Юри 1952 года продемонстрировал, что большинство аминокислот, химических составляющих белков , могут быть синтезированы из неорганических соединений в условиях, повторяющих условия ранней Земли . Эти реакции могли быть вызваны внешними источниками энергии, включая молнии , радиацию , попадание в атмосферу микрометеоритов и взрыв пузырей в морских и океанских волнах.
Хотя считается, что последний универсальный общий предок всех современных организмов (LUCA) сильно отличался от происхождения жизни, исследования LUCA могут помочь в исследовании ранних универсальных характеристик. Подход геномики стремился охарактеризовать LUCA путем идентификации генов, общих для архей и бактерий , представителей двух основных ветвей жизни (при этом эукариоты включены в архейную ветвь в двухдоменной системе ). Оказывается, существует 355 генов, общих для всей жизни; их функции подразумевают, что LUCA была анаэробной по пути Вуда-Люнгдала , получая энергию посредством хемиосмоса и сохраняя свой наследственный материал с помощью ДНК, генетического кода и рибосом . Хотя LUCA жили более 4 миллиардов лет назад (4 млрд лет назад ), исследователи полагают, что это была далеко не первая форма жизни. Раньше клетки могли иметь негерметичную мембрану и получать энергию от естественного протонного градиента вблизи глубоководного гидротермального источника белого курильщика .
Земля остается единственным местом во Вселенной, где, как известно, обитает жизнь. Геохимические и ископаемые данные с Земли служат основой для большинства исследований абиогенеза. Земля Западной образовалась в период 4,54 млрд лет назад, а самые ранние свидетельства жизни на Земле датируются как минимум 3,8 млрд лет назад в Австралии . Ископаемые микроорганизмы, по-видимому, жили в осадках гидротермальных источников, датируемых 3,77–4,28 млрд лет назад из Квебека , вскоре после образования океана 4,4 млрд лет назад во время Гадея .
Обзор
[ редактировать ]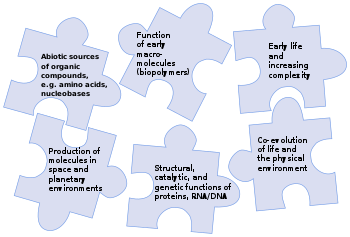
Жизнь состоит из воспроизводства с (наследственными) вариациями. [ 3 ] НАСА определяет жизнь как «самоподдерживающуюся химическую систему, способную к дарвиновской (то есть биологической) эволюции ». [ 4 ] Такая система сложна; последний универсальный общий предок (LUCA), предположительно одноклеточный организм, живший около 4 миллиардов лет назад, уже имел сотни генов, закодированных в ДНК генетическом коде , который сегодня является универсальным. Это, в свою очередь, предполагает наличие набора клеточных механизмов, включающих информационную РНК , транспортную РНК и рибосомы для трансляции кода в белки . Эти белки включали ферменты, обеспечивающие анаэробное дыхание по метаболическому пути Вуда-Люнгдала , и ДНК-полимеразу для репликации генетического материала. [ 5 ] [ 6 ]
Проблема абиогенеза (происхождения жизни) [ 7 ] [ 8 ] [ 9 ] Задача исследователей — объяснить, как такая сложная и тесно взаимосвязанная система могла эволюционно развиваться, ведь на первый взгляд все ее части необходимы для ее функционирования. Например, клетка, будь то LUCA или современный организм, копирует свою ДНК с помощью фермента ДНК-полимеразы, который, в свою очередь, вырабатывается путем трансляции гена ДНК-полимеразы в ДНК. Ни фермент, ни ДНК не могут быть произведены друг без друга. [ 10 ] Эволюционный процесс мог включать молекулярное самовоспроизведение , самосборку, например, клеточных мембран , и автокатализ . [ 5 ] [ 6 ] [ 11 ] Тем не менее, переход нежизни в жизнь экспериментально никогда не наблюдался. [ 12 ]
Предшественники развития живой клетки, такой как LUCA, достаточно ясны, хотя и обсуждаются в деталях: формируется обитаемый мир с запасом минералов и жидкой воды. Синтез пребиотиков создает ряд простых органических соединений, которые собираются в полимеры, такие как белки и РНК. Процесс, произошедший после LUCA, легко понять: биологическая эволюция привела к развитию широкого спектра видов с различными формами и биохимическими способностями. Однако происхождение живых существ, таких как LUCA, из простых компонентов еще далеко не изучено. [ 1 ]
Хотя Земля остается единственным местом, где известна жизнь, [ 13 ] [ 14 ] наука астробиология ищет доказательства существования жизни на других планетах. Стратегия НАСА 2015 года по происхождению жизни была направлена на решение загадки путем выявления взаимодействий, промежуточных структур и функций, источников энергии и факторов окружающей среды, которые способствовали разнообразию, отбору и репликации эволюционирующих макромолекулярных систем. [ 2 ] и составление карты химического ландшафта потенциальных первичных информационных полимеров . Появление полимеров, которые могли реплицироваться, хранить генетическую информацию и проявлять свойства, подлежащие отбору, было, по мнению ученых, скорее всего, решающим шагом в возникновении пребиотической химической эволюции. [ 2 ] Эти полимеры, в свою очередь, происходят из простых органических соединений, таких как нуклеиновые основания , аминокислоты и сахара , которые могли образоваться в результате реакций в окружающей среде. [ 15 ] [ 8 ] [ 16 ] [ 17 ] Успешная теория происхождения жизни должна объяснить, как возникли все эти химические вещества. [ 18 ]
Концептуальная история до 1960-х годов
[ редактировать ]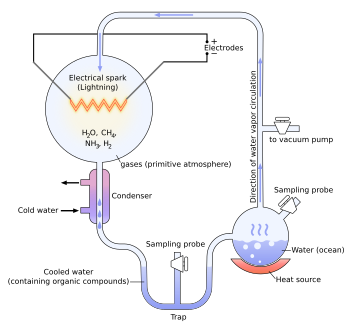
Спонтанное зарождение
[ редактировать ]Одна древняя точка зрения на происхождение жизни, начиная с Аристотеля до XIX века, предполагает самопроизвольное зарождение . [ 19 ] Эта теория утверждала, что «низшие» животные, такие как насекомые, возникли в результате разложения органических веществ и что жизнь возникла случайно. [ 20 ] [ 21 ] Это подвергалось сомнению еще в 17 веке в таких работах, как Томаса Брауна «Эпидемическая псевдодоксия» . [ 22 ] [ 23 ] В 1665 году Роберт Гук опубликовал первые рисунки микроорганизма . В 1676 году Антони ван Левенгук нарисовал и описал микроорганизмы, вероятно, простейшие и бактерии . [ 24 ] Ван Левенгук не соглашался с самопроизвольным зарождением и к 1680-м годам убедился, используя эксперименты, начиная от инкубации мяса в закрытом и открытом помещении и тщательного изучения размножения насекомых, что эта теория неверна. [ 25 ] В 1668 году Франческо Реди показал, что личинки в мясе не появляются, когда мухам не дают откладывать яйца. [ 26 ] К середине XIX века самозарождение считалось опровергнутым. [ 27 ] [ 28 ]
Панспермия
[ редактировать ]Другая древняя идея, восходящая к Анаксагору в V веке до нашей эры, — это панспермия . [ 29 ] идея о том, что жизнь существует во Вселенной , распространяясь метеороидами , астероидами , кометами. [ 30 ] и планетоиды . [ 31 ] Он не пытается объяснить, как зародилась жизнь сама по себе, а переносит зарождение жизни на Земле на другое небесное тело. Преимущество состоит в том, что жизнь не обязательно должна формироваться на каждой планете, на которой она возникает, а скорее в более ограниченном наборе мест или даже в одном месте, а затем распространяться по галактике в другие звездные системы посредством удара кометы или метеорита. [ 32 ] Панспермия не получила особой научной поддержки, поскольку ее в основном использовали для того, чтобы отвлечь внимание от необходимости ответа, а не для объяснения наблюдаемых явлений. Хотя интерес к панспермии возрос, когда исследование метеоритов обнаружило в них следы органических материалов, в настоящее время считается, что жизнь зародилась локально на Земле. [ 33 ]
«Теплый прудик»: первозданный суп
[ редактировать ]Идея о том, что жизнь возникла из неживой материи на медленных стадиях, появилась в книге Герберта Спенсера 1864–1867 годов «Принципы биологии» и в статье Уильяма Тернера Тиселтона-Дайера 1879 года «О спонтанном зарождении и эволюции». 1 февраля 1871 года Чарльз Дарвин написал об этих публикациях Джозефу Хукеру и изложил свои собственные предположения, предполагая, что первоначальная искра жизни могла зародиться в «теплом маленьком пруду со всеми видами аммиака и фосфорных солей , светом, теплом». , электричество и т. д., показывают, что белковое соединение химически образовалось и было готово претерпеть еще более сложные изменения». Далее Дарвин объяснил, что «в наши дни такая материя была бы мгновенно сожрана или поглощена, чего не было до того, как образовались живые существа». [ 34 ] [ 35 ] [ 36 ]
Александр Опарин в 1924 году и Дж. Б. С. Холдейн в 1929 году предположили, что первые молекулы, составляющие самые ранние клетки, медленно самоорганизовались из первичного супа , и эта теория называется гипотезой Опарина-Холдейна . [ 37 ] [ 38 ] Холдейн предположил, что пребиотические океаны Земли представляют собой «горячий разбавленный суп», в котором могли образоваться органические соединения. [ 21 ] [ 39 ] Дж. Д. Бернал показал, что такие механизмы могут образовывать большинство необходимых для жизни молекул из неорганических предшественников. [ 40 ] В 1967 году он предложил три «стадии»: происхождение биологических мономеров ; происхождение биологических полимеров ; и эволюция от молекул к клеткам. [ 41 ] [ 42 ]
Эксперимент Миллера-Юри
[ редактировать ]В 1952 году Стэнли Миллер и Гарольд Юри провели химический эксперимент, чтобы продемонстрировать, как органические молекулы могли спонтанно образоваться из неорганических предшественников в пребиотических условиях, подобных тем, которые постулируются гипотезой Опарина-Холдейна. Он использовал с высокой степенью восстановления смесь газов (без кислорода) — метана , аммиака и водорода , а также водяной пар — для образования простых органических мономеров, таких как аминокислоты . [ 43 ] [ 44 ] Бернал сказал об эксперименте Миллера-Юри, что «недостаточно объяснить образование таких молекул, необходимо физико-химическое объяснение происхождения этих молекул, которое предполагает наличие подходящих источников и поглотителей свободной энергии». ." [ 45 ] Однако современный научный консенсус описывает примитивную атмосферу как слабо восстановительную или нейтральную. [ 46 ] [ 47 ] уменьшение количества и разнообразия аминокислот, которые могут быть произведены. Однако добавление железа и карбонатных минералов, присутствующих в ранних океанах, приводит к образованию разнообразного набора аминокислот. [ 46 ] Более поздние работы были сосредоточены на двух других потенциально восстанавливающих средах: космическом пространстве и глубоководных гидротермальных источниках. [ 48 ] [ 49 ] [ 50 ]
Создание обитаемой Земли
[ редактировать ] происхождения жизни Хронология | ||||||||||||||||||||||||
−13 — – −12 — – −11 — – −10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — |
| |||||||||||||||||||||||
Эволюционная история
[ редактировать ]Ранняя Вселенная с первыми звездами
[ редактировать ]Вскоре после Большого взрыва , который произошел примерно 14 Гя, единственными химическими элементами, присутствующими во Вселенной, были водород , гелий и литий , три самых легких атома в периодической таблице. Эти элементы постепенно срослись и начали вращаться в дисках газа и пыли. Гравитационная аккреция материала в горячих и плотных центрах этих протопланетных дисков образовала звезды в результате синтеза водорода. [ 51 ] Ранние звезды были массивными и недолговечными, производя все более тяжелые элементы посредством звездного нуклеосинтеза . Образование элемента посредством звездного нуклеосинтеза продолжается до его наиболее стабильного элемента Железа -56 . Более тяжелые элементы образовались во время сверхновых в конце жизненного цикла звезды. Углерод , который в настоящее время является четвертым по распространенности химическим элементом во Вселенной (после водорода, гелия и кислорода ), образовался в основном в звездах-белых карликах , особенно в тех, которые больше, чем в два раза массивнее Солнца. [ 52 ] Когда эти звезды достигли конца своего жизненного цикла , они выбросили более тяжелые элементы, в том числе углерод и кислород, по всей Вселенной. Эти более тяжелые элементы позволили сформировать новые объекты, в том числе каменистые планеты и другие тела. [ 53 ] Согласно небулярной гипотезе , формирование и эволюция Солнечной системы начались 4,6 млрд лет назад с гравитационного коллапса небольшой части гигантского молекулярного облака . Большая часть коллапсирующей массы собралась в центре, образовав Солнце , а остальная часть сплюснулась в протопланетный диск, из которого сформировались планеты , луны , астероиды и другие небольшие тела Солнечной системы. [ 54 ]
Появление Земли
[ редактировать ]Возраст Земли составляет 4,54 млрд лет назад, как установлено радиометрическим датированием богатых кальцием и алюминием включений в углеродистых хондритовых метеоритах, старейшем материале в Солнечной системе. [ 55 ] [ 56 ] Гадейская . Земля (с момента ее образования до 4 млрд лет назад) поначалу была негостеприимна для любых живых организмов Во время своего формирования Земля потеряла значительную часть своей первоначальной массы и, как следствие, лишилась гравитации, способной удерживать молекулярный водород и основную часть исходных инертных газов. [ 57 ] Считается , что вскоре после первоначальной аккреции Земли в 4,48 млрд лет назад ее столкновение с Тейей , предполагаемым ударным элементом, привело к образованию выброшенных обломков, которые в конечном итоге сформировали Луну. [ 58 ] Этот удар разрушил бы первичную атмосферу Земли, оставив после себя облака вязких силикатов и углекислого газа. Эта нестабильная атмосфера просуществовала недолго и вскоре после этого конденсировалась, образовав объемную силикатную Землю, оставив после себя атмосферу, состоящую в основном из водяного пара, азота и углекислого газа с меньшим количеством монооксида углерода , водорода и соединений серы . [ 59 ] [ 60 ] Считается, что раствор углекислого газа в воде сделал моря слегка кислыми , с pH около 5,5. [ 61 ]
Предполагается, что конденсация, образующая жидкие океаны , произошла еще во время удара, образовавшего Луну. [ 62 ] [ 63 ] Этот сценарий нашел подтверждение в датировке кристаллов циркона массой 4,404 Гя с высоким δ. 18 Значения O из метаморфизованного кварцита горы Нарриер в Западной Австралии. [ 64 ] [ 65 ] Атмосфера Гадея была охарактеризована как «гигантская, продуктивная химическая лаборатория под открытым небом», похожая на современные вулканические газы, которые все еще поддерживают некоторую абиотическую химию. Несмотря на вероятный рост вулканизма в результате ранней тектоники плит, между 4,4 и 4,3 млрд лет назад Земля могла быть преимущественно водным миром. Спор идет о том, обнажалась ли кора над этим океаном, из-за неуверенности в том, как выглядела ранняя тектоника плит. Обычно считается, что для развития ранней жизни необходимо наличие суши, поэтому этот вопрос важен для определения того, когда в истории Земли возникла жизнь. [ 66 ] После формирования Луны Земля, вероятно, существовала с небольшой континентальной корой или вообще с ней, турбулентной атмосферой и гидросферой, подверженной интенсивному ультрафиолетовому излучению от Солнца в стадии Т Тельца , космическому излучению , а также продолжающимся столкновениям с астероидами и кометами . [ 67 ] Несмотря на все это, в период от позднего хадея до раннего архея на Земле, вероятно, существовала нишевая среда, благоприятная для жизни на Земле.
Гипотеза поздней тяжелой бомбардировки утверждает, что период интенсивного удара произошел примерно на 3,9 млрд лет назад во время Гадея. [ 68 ] [ 69 ] Катастрофическое воздействие могло бы привести к стерилизации всего живого на Земле, улетучивая жидкие океаны и блокируя Солнце, необходимое для фотосинтеза первичных производителей, отодвигая самое раннее возможное появление жизни на период после поздней тяжелой бомбардировки. [ 70 ] Недавние исследования ставят под сомнение как интенсивность поздней тяжелой бомбардировки, так и ее потенциал стерилизации. Неопределенность относительно того, была ли поздняя тяжелая бомбардировка одним гигантским ударом или периодом более высокой интенсивности ударов, сильно изменила значение ее разрушительной силы. [ 71 ] [ 72 ] Дата 3,9 млрд лет является результатом датировки возвратов образцов миссии Аполлон, собранных в основном вблизи бассейна Имбриум , что искажает возраст зарегистрированных столкновений. [ 73 ] Моделирование воздействия на лунную поверхность показывает, что вместо катастрофического события 3,9 млрд лет назад, вероятно, произошли многочисленные мелкомасштабные, кратковременные периоды бомбардировок. [ 74 ] Наземные данные подтверждают эту идею, показывая множественные периоды выбросов в горных породах как до, так и после отметки 3,9 млрд лет, предполагая, что ранняя Земля подвергалась непрерывным воздействиям, которые не оказали бы такого большого влияния на вымирание, как считалось ранее. [ 75 ] Если поздняя тяжелая бомбардировка не произошла, это позволяет предположить, что возникновение жизни произошло намного раньше 3,9 млрд лет назад.
Если бы жизнь развивалась в океане на глубине более десяти метров, она была бы защищена как от поздних ударов, так и от высоких уровней ультрафиолетового излучения Солнца. Геотермически нагретая океаническая кора могла дать гораздо больше органических соединений через глубокие гидротермальные жерла, чем показали эксперименты Миллера-Юри . [ 76 ] Доступная энергия максимизируется при температуре 100–150 °C — температуре, при которой живут гипертермофильные бактерии и термоацидофильные археи . [ 77 ]
Самые ранние свидетельства жизни
[ редактировать ]Точное время возникновения жизни на Земле неизвестно. Оценки минимального возраста основаны на данных геологической летописи горных пород . Самые ранние физические свидетельства жизни, обнаруженные на данный момент, состоят из микробиалитов в Зеленокаменном поясе Нуввуагиттук в Северном Квебеке, в породах полосчатого железа возрастом не менее 3,77, а возможно, и 4,32 млрд лет назад. Микроорганизмы жили в осадках гидротермальных жерл вскоре после формирования океанов 4,4 млрд лет назад во время Гадея. Микробы напоминали современные бактерии гидротермальных источников, что подтверждает точку зрения о том, что абиогенез начался в такой среде. [ 78 ]
Биогенный графит был обнаружен в метаосадочных породах возрастом 3,7 млрд лет на юго-западе Гренландии. [ 79 ] и в окаменелостях микробного мата из кремней 3,49 Гья в регионе Пилбара в Западной Австралии . [ 80 ] Свидетельства ранней жизни в горных породах острова Акилия , недалеко от супракрустального пояса Исуа на юго-западе Гренландии, датируемые 3,7 млрд лет назад, показали наличие биогенных изотопов углерода . [ 81 ] В других частях супракрустального пояса Исуа включения графита, заключенные в кристаллах граната, связаны с другими элементами жизни: кислородом, азотом и, возможно, фосфором в форме фосфата , что является дополнительным доказательством существования жизни 3,7 Гя. [ 82 ] В регионе Пилбара в Западной Австралии убедительные доказательства ранней жизни были обнаружены в пиритсодержащем песчанике на окаменелом пляже с округлыми трубчатыми клетками, которые окисляли серу путем фотосинтеза в отсутствие кислорода. [ 83 ] [ 84 ] Соотношения изотопов углерода в графитовых включениях из цирконов Джек-Хиллз позволяют предположить, что жизнь могла существовать на Земле начиная с 4,1 млрд лет назад. [ 85 ]
В регионе Пилбара в Западной Австралии находится формация Дрессер с породами возрастом 3,48 млрд лет назад, включая слоистые структуры, называемые строматолитами . Их современные аналоги созданы фотосинтезирующими микроорганизмами, включая цианобактерии . [ 86 ] Они залегают в недеформированных гидротермально-осадочных толщах; их текстура указывает на биогенное происхождение. В некоторых частях формации Дрессер на суше сохранились горячие источники , но другие регионы, похоже, представляли собой мелководные моря. [ 87 ] Анализ молекулярных часов предполагает, что LUCA возникла до поздней тяжелой бомбардировки (3,9 млрд лет назад). [ 88 ]
-
 Строматолиты в формации Сие, Национальный парк Глейшер , датированные 3,5 млрд лет назад, что делает их одними из самых ранних форм жизни.
Строматолиты в формации Сие, Национальный парк Глейшер , датированные 3,5 млрд лет назад, что делает их одними из самых ранних форм жизни. -
 Современные строматолиты в заливе Шарк , созданные фотосинтезирующими цианобактериями.
Современные строматолиты в заливе Шарк , созданные фотосинтезирующими цианобактериями.


Молекулы-производители: синтез пребиотиков
[ редактировать ]Все химические элементы, за исключением водорода и гелия, образуются в результате звездного нуклеосинтеза. Основные химические ингредиенты жизни – молекула углерода-водорода (CH), положительный ион углерода-водорода (CH+) и ион углерода (C+) – были созданы ультрафиолетовым светом звезд. [ 89 ] Сложные молекулы, в том числе органические, образуются естественным путем как в космосе, так и на планетах. [ 90 ] Органические молекулы на ранней Земле могли иметь либо земное происхождение, при этом синтез органических молекул был вызван ударными ударами, либо другими источниками энергии, такими как ультрафиолетовый свет, окислительно-восстановительная связь или электрические разряды; или внеземного происхождения ( псевдопанспермия ), когда органические молекулы образуются в межзвездных пылевых облаках, падающих на планету. [ 91 ] [ 92 ]
Наблюдаемые внеземные органические молекулы
[ редактировать ]Органическое соединение – это химическое вещество, молекулы которого содержат углерод. Углерод в изобилии содержится на Солнце, звездах, кометах и в атмосферах большинства планет. [ 93 ] Органические соединения относительно распространены в космосе, образуются на «фабриках сложного молекулярного синтеза», которые встречаются в молекулярных облаках и околозвездных оболочках , и химически развиваются после того, как реакции инициируются в основном ионизирующим излучением . [ 90 ] [ 94 ] [ 95 ] пуриновые и пиримидиновые нуклеиновые основания, в том числе гуанин , аденин , цитозин , урацил и тимин обнаружены В метеоритах . Они могли предоставить материалы для ДНК и РНК формирования на ранней Земле . [ 96 ] Аминокислота глицин была обнаружена в материале, выброшенном кометой Wild 2 ; ранее он был обнаружен в метеоритах. [ 97 ] Кометы покрыты темным материалом, предположительно смолоподобным органическим веществом, образовавшимся из простых соединений углерода под действием ионизирующего излучения. Дождь из кометного материала мог принести на Землю такие сложные органические молекулы. [ 98 ] [ 99 ] [ 60 ] Подсчитано, что во время поздней тяжелой бомбардировки метеориты могли доставлять на Землю до пяти миллионов тонн органических пребиотических элементов в год. [ 60 ]
Мировая гипотеза ПАУ
[ редактировать ]
Зеленые области показывают области, где излучение горячих звезд сталкивается с большими молекулами и мелкими пылинками, называемыми « полициклическими ароматическими углеводородами » (ПАУ), вызывая их флуоресценцию . Космический телескоп «Спитцер» , 2018 год.
Полициклические ароматические углеводороды (ПАУ) являются наиболее распространенными и многочисленными многоатомными молекулами в наблюдаемой Вселенной и являются основным хранилищем углерода. [ 93 ] [ 100 ] [ 101 ] [ 102 ] Судя по всему, они образовались вскоре после Большого взрыва. [ 103 ] [ 101 ] [ 102 ] и связаны с новыми звездами и экзопланетами . [ 93 ] Вероятно, они являются составной частью первобытного моря Земли. [ 103 ] [ 101 ] [ 102 ] ПАУ были обнаружены в туманностях . [ 104 ] и в межзвездной среде , в кометах и метеоритах. [ 93 ]
Гипотеза мира ПАУ предполагает, что ПАУ являются предшественниками мира РНК. [ 105 ] Звезда HH 46-IR, напоминающая Солнце в начале своей жизни, окружена диском материала, который содержит молекулы, в том числе цианидные соединения, углеводороды и окись углерода. ПАУ в межзвездной среде могут трансформироваться посредством гидрирования , оксигенации и гидроксилирования в более сложные органические соединения, используемые в живых клетках. [ 106 ]
Нуклеиновые основания и нуклеотиды
[ редактировать ]Большинство органических соединений, привнесенных на Землю частицами межзвездной пыли, помогли сформировать сложные молекулы благодаря своей особой поверхностно-каталитической активности. [ 107 ] [ 108 ] Исследования 12 С/ 13 C Соотношения изотопов органических соединений в метеорите Мерчисон позволяют предположить, что урацил, компонент РНК, и родственные ему молекулы, включая ксантин , образовались внеземным путем. [ 109 ] Исследования метеоритов НАСА позволяют предположить, что все четыре азотистых основания ДНК (аденин, гуанин и родственные им органические молекулы) образовались в космическом пространстве. [ 107 ] [ 110 ] [ 111 ] Космическая пыль, пронизывающая Вселенную, содержит сложную органику («аморфные органические твердые вещества со смешанной ароматико - алифатической структурой»), которая могла бы быстро создаваться звездами. [ 112 ] Гликолальдегид , молекула сахара и предшественник РНК, был обнаружен в регионах космоса, в том числе вокруг протозвезд и на метеоритах. [ 113 ] [ 114 ]
Лабораторный синтез
[ редактировать ]Еще в 1860-х годах эксперименты продемонстрировали, что биологически значимые молекулы могут быть получены путем взаимодействия простых источников углерода с многочисленными неорганическими катализаторами. Спонтанное образование сложных полимеров из абиотически генерируемых мономеров в условиях, сформулированных теорией «супа», не является простым. в высоких концентрациях также образовывались соединения, которые препятствовали бы образованию полимеров Помимо необходимых основных органических мономеров, во время экспериментов Миллера-Юри и Джоан Оро . [ 115 ] В биологии для кодирования белковых ферментов используются по существу 20 аминокислот, что представляет собой очень небольшую подгруппу структурно возможных продуктов. Поскольку жизнь имеет тенденцию использовать все, что доступно, необходимо объяснение, почему используемый набор настолько мал. [ 116 ] Формамид привлекателен как среда, потенциально являющаяся источником производных аминокислот из простого альдегидного и нитрильного сырья. [ 117 ]
Сахара
[ редактировать ]
Александр Бутлеров показал в 1861 году, что в результате формозной реакции образуются сахара, включая тетрозы, пентозы и гексозы, когда формальдегид нагревается в основных условиях с ионами двухвалентных металлов, таких как кальций. В 1959 г. Р. Бреслоу предположил, что реакция носит автокаталитический характер. [ 118 ]
Нуклеиновые основания
[ редактировать ]Нуклеиновые основания, такие как гуанин и аденин, можно синтезировать из простых источников углерода и азота, таких как цианистый водород (HCN) и аммиак. [ 119 ] Формамид производит все четыре рибонуклеотида при нагревании с земными минералами. Формамид широко распространен во Вселенной и образуется в результате реакции воды и HCN. Его можно сконцентрировать путем испарения воды. [ 120 ] [ 121 ] HCN ядовит только для аэробных организмов ( эукариотов и аэробных бактерий), которых еще не существовало. Он может играть роль в других химических процессах, таких как синтез аминокислоты глицина. [ 60 ]
Компоненты ДНК и РНК, включая урацил, цитозин и тимин, можно синтезировать в условиях космического пространства с использованием исходных химических веществ, таких как пиримидин, обнаруженный в метеоритах. Пиримидин мог образоваться в звездах красных гигантов или в межзвездных пылевых и газовых облаках. [ 122 ] Все четыре основания РНК могут быть синтезированы из формамида в ходе событий с высокой плотностью энергии, таких как инопланетные удары. [ 123 ]
Сообщалось о других путях синтеза оснований из неорганических материалов. [ 124 ] Температуры замерзания выгодны для синтеза пуринов из-за концентрирующего эффекта ключевых предшественников, таких как цианистый водород. [ 125 ] Однако, в то время как аденин и гуанин для синтеза требуют условий замораживания, цитозину и урацилу могут потребоваться температуры кипения. [ 126 ] Семь аминокислот и одиннадцать типов нуклеиновых оснований образовались во льду, когда аммиак и цианид хранились в морозильной камере в течение 25 лет. [ 127 ] [ 128 ] S- триазины (альтернативные нуклеиновые основания), пиримидины, включая цитозин и урацил, и аденин можно синтезировать, подвергая раствор мочевины циклам замораживания-оттаивания в восстановительной атмосфере с использованием искровых разрядов в качестве источника энергии. [ 129 ] Объяснением необычной скорости этих реакций при такой низкой температуре является эвтектическое замерзание , при котором примеси скапливаются в микроскопические карманы жидкости внутри льда, заставляя молекулы чаще сталкиваться. [ 130 ]
Пептиды
[ редактировать ]Предполагается, что синтез пребиотических пептидов происходил несколькими возможными путями. Некоторые сосредотачиваются на условиях высокой температуры/концентрации, при которых конденсация становится энергетически выгодной, в то время как другие сосредотачиваются на доступности вероятных пребиотических конденсирующих агентов. [ 131 ] [ нужны дальнейшие объяснения ]
Экспериментальные данные об образовании пептидов в средах с уникальной концентрацией подкреплены работами, предполагающими, что циклы «влажно-сухо» и присутствие специфических солей могут значительно увеличить спонтанную конденсацию глицина в полиглициновые цепи. [ 132 ] Другая работа предполагает, что, хотя минеральные поверхности, такие как пирит, кальцит и рутил, катализируют конденсацию пептидов, они также катализируют их гидролиз. Авторы предполагают, что для получения пептидов в достаточных концентрациях потребуется дополнительная химическая активация или связывание. Таким образом, катализ минеральной поверхности, хотя и важен, сам по себе недостаточен для синтеза пептидов. [ 133 ]
Было идентифицировано множество пребиотически вероятных конденсирующих/активирующих агентов, в том числе следующие: цианамид, дицианамид, дициандиамид, диаминомалеонитрил, мочевина, триметафосфат, NaCl, CuCl 2 , (Ni,Fe)S, CO, карбонилсульфид (COS), сероуглерод ( CS 2 ) , SO 2 и диаммонийфосфат (ДАФ). [ 131 ]
В эксперименте, о котором сообщалось в 2024 году, использовался саффировый субстрат с сетью тонких трещин под тепловым потоком, аналогичный среде глубоководных океанических жерл , в качестве механизма для отделения и концентрирования пребиотически важных строительных блоков из разбавленной смеси, очищая их концентрацию путем до трех порядков. Авторы предлагают это как правдоподобную модель происхождения сложных биополимеров. [ 134 ] Это представляет собой еще один физический процесс, который позволяет концентрированным предшественникам пептидов объединяться в правильных условиях. Аналогичная роль увеличения концентрации аминокислот была предложена и для глин. [ 135 ]
Хотя все эти сценарии включают конденсацию аминокислот, пребиотический синтез пептидов из более простых молекул, таких как CO, NH 3 и C, минуя стадию образования аминокислот, очень эффективен. [ 136 ] [ 137 ]
Производство подходящих везикул
[ редактировать ]
Самый большой вопрос эволюции, оставшийся без ответа, заключается в том, как простые протоклетки впервые возникли и изменились в репродуктивном вкладе в следующее поколение, положив тем самым начало эволюции жизни. Теория липидного мира постулирует, что первый самовоспроизводящийся объект был липидоподобным . [ 138 ] [ 139 ] образуют в воде липидные бислои Фосфолипиды при перемешивании — ту же структуру, что и в клеточных мембранах. Этих молекул не было на ранней Земле, но другие амфифильные длинноцепочечные молекулы также образуют мембраны. Эти тельца могут расширяться за счет внедрения дополнительных липидов и могут спонтанно разделяться на два потомка одинакового размера и состава. Липидные тела, возможно, служили защитной оболочкой для хранения информации, позволяя развиваться и сохранять полимеры, такие как РНК, которые хранят информацию. Изучены только один или два типа амфифилов, которые могли привести к развитию везикул. [ 140 ] Существует огромное количество возможных вариантов расположения липидных бислойных мембран, и те из них, которые обладают лучшими репродуктивными характеристиками, сходятся к реакции гиперцикла. [ 141 ] [ 142 ] положительная обратная связь, состоящая из двух взаимных катализаторов, представленных мембранным участком и специфическим соединением, захваченным в пузырьке. Такие пары сайт/соединение передаются дочерним пузырькам, что приводит к появлению отдельных линий везикул, что сделало бы возможным естественный отбор . [ 143 ]
Протоклетка . — это самоорганизованная, самоупорядоченная сферическая совокупность липидов, предложенная в качестве трамплина на пути к зарождению жизни [ 140 ] Функциональная протоклетка (по состоянию на 2014 год) еще не создана в лабораторных условиях. [ 144 ] [ 145 ] [ 146 ] Самособирающиеся везикулы являются важными компонентами примитивных клеток. [ 140 ] Теория классической необратимой термодинамики рассматривает самосборку под действием обобщенного химического потенциала в рамках диссипативных систем . [ 147 ] [ 148 ] [ 149 ] Второй закон термодинамики требует, чтобы общая энтропия возрастала, однако жизнь отличается высокой степенью организации. Поэтому необходима граница, чтобы отделить упорядоченные жизненные процессы от хаотичной неживой материи. [ 150 ]
Ирен Чен и Джек В. Шостак предполагают, что элементарные протоклетки могут вызывать клеточное поведение, включая примитивные формы дифференциального воспроизводства, конкуренции и накопления энергии. [ 145 ] Конкуренция за мембранные молекулы будет благоприятствовать стабилизированным мембранам, что предполагает селективное преимущество для эволюции поперечно-сшитых жирных кислот и даже фосфолипидов . современных [ 145 ] Такая микроинкапсуляция позволит осуществлять метаболизм внутри мембраны и обмениваться небольшими молекулами, сохраняя при этом крупные биомолекулы внутри. Такая мембрана необходима клетке для создания собственного электрохимического градиента для хранения энергии путем перекачки ионов через мембрану. [ 151 ] [ 152 ] Везикулы жирных кислот в условиях щелочных гидротермальных источников могут быть стабилизированы изопреноидами, которые синтезируются по формозной реакции; Преимущества и недостатки изопреноидов, включенных в липидный бислой в разных микроокружениях, могли привести к расхождению мембран архей и бактерий. [ 153 ]
Лабораторные эксперименты показали, что везикулы могут подвергаться эволюционному процессу в условиях циклического изменения давления. [ 154 ] Моделирование системной среды в зонах тектонических разломов в земной коре циклическое изменение давления приводит к периодическому образованию пузырьков. [ 155 ] В тех же условиях случайные пептидные формируются цепи, которые непрерывно отбираются по способности интегрироваться в мембрану везикул. Дальнейший отбор везикул по их стабильности потенциально приводит к созданию функциональных пептидных структур, [ 156 ] [ 157 ] [ 158 ] связано с увеличением выживаемости везикул.
Производство биологии
[ редактировать ]Энергия и энтропия
[ редактировать ]Жизнь требует потери энтропии или беспорядка, поскольку молекулы организуются в живую материю. В то же время возникновение жизни связано с образованием структур, выходящих за определенный порог сложности . [ 159 ] Появление жизни с возрастающим порядком и сложностью не противоречит второму закону термодинамики, который гласит, что общая энтропия никогда не уменьшается, поскольку живой организм создает порядок в некоторых местах (например, в своем живом теле) за счет увеличения энтропии в других местах. (например, производство тепла и отходов). [ 160 ] [ 161 ] [ 162 ]
На ранней Земле для химических реакций было доступно множество источников энергии. Тепло геотермальных процессов является стандартным источником энергии для химии. Другие примеры включают солнечный свет, молнию, [ 60 ] попадание в атмосферу микрометеоритов, [ 163 ] и взрыв пузырей в морских и океанских волнах. [ 164 ] Это подтверждено экспериментами [ 165 ] [ 166 ] и симуляции. [ 167 ] Неблагоприятные реакции могут быть вызваны весьма благоприятными, как в случае химии железа с серой. Например, это, вероятно, было важно для фиксации углерода . [ а ] Фиксация углерода путем реакции CO 2 с H 2 S посредством химии железа и серы является благоприятной и происходит при нейтральном pH и 100 °C. Железо-серные поверхности, которых много вблизи гидротермальных источников, могут стимулировать производство небольших количеств аминокислот и других биомолекул. [ 60 ]
хемиосмос
[ редактировать ]
В 1961 году Питер Митчелл предложил хемиосмос как основную систему преобразования энергии клетки. Этот механизм, ныне повсеместно распространенный в живых клетках, обеспечивает преобразование энергии в микроорганизмах и митохондриях эукариот , что делает его вероятным кандидатом на раннюю жизнь. [ 168 ] [ 169 ] Митохондрии производят аденозинтрифосфат (АТФ), энергетическую валюту клетки, используемую для запуска клеточных процессов, таких как химический синтез. Механизм синтеза АТФ включает закрытую мембрану, в которую АТФ-синтаза встроен фермент . Энергия, необходимая для высвобождения прочносвязанного АТФ, исходит от протонов , которые движутся через мембрану. [ 170 ] В современных клетках эти движения протонов вызваны перекачкой ионов через мембрану, поддерживая электрохимический градиент. У первых организмов градиент мог быть обеспечен разницей в химическом составе потока из гидротермального источника и окружающей морской воды. [ 152 ] или, возможно, метеорные хиноны, которые способствовали развитию хемиосмотической энергии через липидные мембраны, если они имели земное происхождение. [ 171 ]

Мир РНК
[ редактировать ]Гипотеза мира РНК описывает раннюю Землю с самовоспроизводящейся и каталитической РНК, но без ДНК и белков. [ 172 ] Многие исследователи сходятся во мнении, что мир РНК, должно быть, предшествовал жизни, основанной на ДНК, которая сейчас доминирует. [ 173 ] Однако жизнь, основанная на РНК, возможно, не была первой. [ 174 ] [ 175 ] Другая модель повторяет дарвиновский «маленький теплый пруд» с циклами увлажнения и высыхания. [ 176 ]
РНК играет центральную роль в процессе трансляции. Малые РНК могут катализировать все химические группы и передачу информации, необходимые для жизни. [ 175 ] [ 177 ] РНК выражает и сохраняет генетическую информацию в современных организмах; а химические компоненты РНК легко синтезируются в условиях, приближенных к ранней Земле, которые сильно отличались от тех, которые преобладают сегодня. Структуру рибосомы назвали «дымящимся ружьем»: с центральным ядром РНК и отсутствием боковых цепей аминокислот в пределах 18 Å от активного сайта , который катализирует образование пептидных связей. [ 178 ] [ 174 ] [ 179 ]
Концепцию мира РНК предложил в 1962 году Александр Рич . [ 180 ] Этот термин был придуман Уолтером Гилбертом в 1986 году. [ 175 ] [ 181 ] Первоначально возникли трудности с объяснением абиотического синтеза нуклеотидов цитозина и урацила. [ 182 ] Последующие исследования показали возможные пути синтеза; например, формамид производит все четыре рибонуклеотида и другие биологические молекулы при нагревании в присутствии различных земных минералов. [ 120 ] [ 121 ]
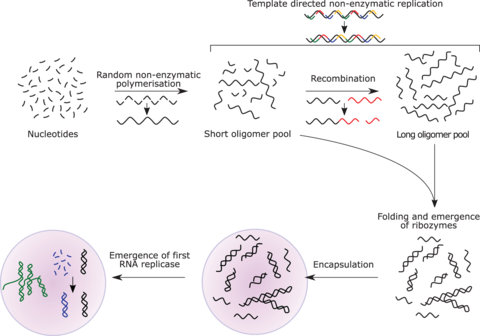
РНК-репликаза может функционировать как код, так и катализатор дальнейшей репликации РНК, т. е. она может быть автокаталитической. Джек Шостак показал, что некоторые каталитические РНК могут объединять более мелкие последовательности РНК, создавая потенциал для саморепликации. Системы репликации РНК, включающие два рибозима, катализирующие синтез друг друга, показали время удвоения продукта около часа и подверглись естественному отбору в условиях эксперимента. [ 183 ] [ 184 ] [ 174 ] Если бы такие условия присутствовали на ранней Земле, то естественный отбор способствовал бы распространению таких автокаталитических наборов , к которым можно было бы добавить дополнительные функциональные возможности. [ 185 ] [ 186 ] [ 187 ] Самосборка РНК может происходить спонтанно в гидротермальных источниках. [ 188 ] [ 189 ] [ 190 ] Предварительная форма тРНК могла бы собраться в такую молекулу-репликатор. [ 191 ]
Возможные предшественники синтеза белка включают синтез коротких пептидных кофакторов или самокатализирующееся дублирование РНК. Вполне вероятно, что предковая рибосома полностью состояла из РНК, хотя некоторые функции с тех пор взяли на себя белки. Основные оставшиеся вопросы по этой теме включают определение силы отбора, способствующей эволюции рибосомы, и определение того, как возник генетический код. [ 192 ]
Юджин Кунин утверждал, что «в настоящее время не существует убедительных сценариев возникновения репликации и трансляции, ключевых процессов, которые вместе составляют ядро биологических систем и очевидную предпосылку биологической эволюции. Концепция мира РНК может предложить лучший шанс для разрешение этой загадки, но до сих пор не может адекватно объяснить появление эффективной РНК-репликазы или системы трансляции». [ 193 ]
От РНК к направленному синтезу белка
[ редактировать ]В соответствии с гипотезой мира РНК, большая часть шаблонного биосинтеза белка в современной биологии осуществляется молекулами РНК, а именно тРНК и рибосомой (состоящей как из белковых компонентов, так и из рРНК). Считается, что наиболее центральная реакция синтеза пептидной связи осуществляется посредством катализа оснований доменом V 23S рРНК. [ 194 ] Экспериментальные данные продемонстрировали успешный синтез ди- и трипептидов с помощью системы, состоящей только из аминоацилфосфатных адаптеров и проводников РНК, что может стать возможным переходным этапом между миром РНК и современным синтезом белков. [ 194 ] [ 195 ] Рибозимы аминоацилирования, которые могут заряжать тРНК родственными им аминокислотами, также были отобраны в экспериментах in vitro. [ 196 ] Авторы также тщательно картировали ландшафты приспособленности в пределах своей выборки и обнаружили, что случайное появление активных последовательностей более важно, чем оптимизация последовательностей. [ 196 ]
Ранние функциональные пептиды
[ редактировать ]Первые белки должны были бы возникнуть без полноценной системы биосинтеза белков. Как обсуждалось выше, существуют многочисленные механизмы пребиотического синтеза полипептидов. Однако эти пептиды со случайной последовательностью, скорее всего, не имели биологической функции. Таким образом, были проведены значительные исследования по изучению того, как ранние функциональные белки могли возникнуть из случайных последовательностей. Во-первых, некоторые данные о скорости гидролиза показывают, что абиотически вероятные пептиды, вероятно, содержат значительные отклонения от «ближайших соседей». [ 197 ] Это могло оказать некоторое влияние на разнообразие ранних последовательностей белков. В другой работе Энтони Кифа и Джека Шостака отбор мРНК осуществляется в библиотеке 6*10. 12 80-меры использовали для поиска последовательностей, обладающих АТФ-связывающей активностью. Они пришли к выводу, что примерно 1 из 10 11 случайные последовательности имели функцию связывания АТФ. [ 198 ] Хотя это единственный пример функциональной частоты в пространстве случайных последовательностей, эта методология может служить мощным инструментом моделирования для понимания ранней эволюции белков. [ 199 ]
Филогения и LUCA
[ редактировать ]Начиная с работы Карла Везе в 1977 году, исследования в области геномики поместили последнего универсального общего предка (LUCA) всех современных форм жизни между бактериями и кладой, образованной археями и эукариотами, на филогенетическом древе жизни. Он прожил более 4 Гя. [ 200 ] [ 201 ] Меньшее количество исследований поместило LUCA в бактерии, предполагая, что археи и эукариоты эволюционно произошли из эубактерий; [ 202 ] Томас Кавальер-Смит предположил в 2006 году, что фенотипически разнообразный бактериальный тип Chloroflexota содержит LUCA. [ 203 ]
-
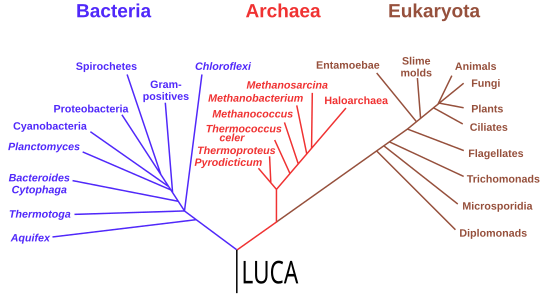 Филогенетическое дерево, показывающее последнего универсального общего предка (LUCA) в корне. Основными кладами являются бактерии, с одной стороны, и археи и эукариоты , с другой.
Филогенетическое дерево, показывающее последнего универсального общего предка (LUCA) в корне. Основными кладами являются бактерии, с одной стороны, и археи и эукариоты , с другой.

В 2016 году был идентифицирован набор из 355 генов, вероятно присутствующих в LUCA. Всего было секвенировано 6,1 миллиона прокариотических генов бактерий и архей, в результате чего было идентифицировано 355 белковых кластеров из 286 514 белковых кластеров, которые, вероятно, были общими для LUCA. Результаты показывают, что LUCA была анаэробной с путем Вуда-Люнгдала (восстановительный ацетил-КоА), фиксирующей азот и углерод, термофильной. Его кофакторы предполагают зависимость от окружающей среды, богатой водородом, углекислым газом, железом и переходными металлами . Его генетическим материалом, вероятно, была ДНК, требующая 4-нуклеотидного генетического кода, информационной РНК, транспортной РНК и рибосом для трансляции кода в белки, такие как ферменты. LUCA, вероятно, обитала в анаэробных гидротермальных источниках в геохимически активной среде. Очевидно, это уже был сложный организм и, должно быть, имел предшественников; это было не первое живое существо. [ 10 ] [ 204 ] Физиология LUCA вызывает споры. [ 205 ] [ 206 ] [ 207 ]
-
![Системы и окружающая среда LUCA включали путь Вуда-Люнгдала.[10]](//upload.wikimedia.org/wikipedia/commons/thumb/e/ed/LUCA_systems_and_environment.svg/622px-LUCA_systems_and_environment.svg.png) Системы и окружающая среда LUCA включали путь Вуда-Люнгдала . [ 10 ]
Системы и окружающая среда LUCA включали путь Вуда-Люнгдала . [ 10 ]
![Системы и окружающая среда LUCA включали путь Вуда-Люнгдала.[10]](http://upload.wikimedia.org/wikipedia/commons/thumb/e/ed/LUCA_systems_and_environment.svg/622px-LUCA_systems_and_environment.svg.png)
Лесли Оргел утверждал, что ранние механизмы перевода генетического кода были подвержены катастрофическим ошибкам . Джеффри Хоффманн, однако, показал, что такое оборудование может стабильно функционировать вопреки «парадоксу Оргеля». [ 208 ] [ 209 ] [ 210 ] Метаболические реакции, которые также были выявлены при LUCA, включают неполный обратный цикл Кребса , глюконеогенез , пентозофосфатный путь , гликолиз , восстановительное аминирование и трансаминирование . [ 211 ] [ 212 ]
Подходящие геологические условия
[ редактировать ]различные были предложены Для возникновения жизни геологические и экологические условия. Эти теории часто конкурируют друг с другом, поскольку существует множество различных взглядов на доступность пребиотических соединений, геофизические условия и характеристики раннего возраста. Первый организм на Земле, вероятно, выглядел иначе, чем LUCA . Между первым появлением жизни и началом разветвления всех современных филогений прошло неизвестное количество времени, с неизвестным переносом генов, вымиранием и эволюционной адаптацией к различным экологическим нишам. [ 213 ] Считается, что одним из основных сдвигов является переход от мира РНК к миру РНК-ДНК-белков. Современные филогении предоставляют более подходящие генетические данные о LUCA, чем о его предшественниках. [ 214 ]
Самыми популярными гипотезами происхождения жизни являются глубоководные гидротермальные источники и поверхностные водоемы. Поверхностные воды можно разделить на горячие источники, озера и пруды с умеренной температурой и холодные воды.
Глубоководные гидротермальные источники
[ редактировать ]Горячие жидкости
[ редактировать ]
Ранние микроископаемые, возможно, произошли из горячего мира газов, таких как метан, аммиак, углекислый газ и сероводород , токсичных для большинства современных форм жизни. [ 215 ] Анализ древа жизни помещает термофильные и гипертермофильные бактерии и археи ближе всего к корню, что позволяет предположить, что жизнь могла развиваться в жаркой среде. [ 216 ] Теория глубоководных или щелочных гидротермальных источников утверждает, что жизнь зародилась в подводных гидротермальных источниках. [ 217 ] [ 218 ] Уильям Мартин и Майкл Рассел предположили, что «жизнь развивалась в структурированных осадках моносульфида железа в гидротермальном холме на месте просачивания при градиенте окислительно-восстановительного потенциала, pH и температуры между богатой сульфидами гидротермальной жидкостью и железо(II)-содержащими водами дна Гадейского океана. Возникающее естественным путем трехмерное разделение, наблюдаемое в окаменелых осадках сульфидов металлов в местах просачивания, указывает на то, что эти неорганические отсеки были предшественниками клеточных стенок и мембран, обнаруженных у свободноживущих прокариот. Известная способность FeS и NiS катализировать синтез. ацетилметилсульфид из монооксида углерода и метилсульфида, составляющих гидротермальную жидкость, указывает на то, что пребиотический синтез происходил на внутренних поверхностях этих отсеков со стенками из сульфида металлов». [ 219 ]
Они образуются там, где богатые водородом флюиды выходят из-под морского дна в результате серпентинизации ультраосновного с морской водой оливина и взаимодействия pH с богатой углекислым газом океанской водой. Вентиляционные отверстия образуют постоянный источник химической энергии, получаемый в результате окислительно-восстановительных реакций, в которых доноры электронов (молекулярный водород) реагируют с акцепторами электронов (диоксид углерода); см. мировую теорию железа и серы . Это экзотермические реакции . [ 217 ] [ б ]
Хемиосмотический градиент
[ редактировать ]
Рассел продемонстрировал, что щелочные источники создают абиогенный хемиосмотический градиент протондвижущей силы . [ 219 ] идеален для абиогенеза. Их микроскопические отсеки «обеспечивают естественные средства концентрации органических молекул», состоящие из железо-серных минералов, таких как макинавит , наделившие эти минеральные клетки каталитическими свойствами, предусмотренными Гюнтером Вехтерсхойзером . [ 220 ] Такое движение ионов через мембрану зависит от сочетания двух факторов:
- Сила диффузии , вызванная градиентом концентрации: все частицы, включая ионы, имеют тенденцию диффундировать от более высокой концентрации к более низкой.
- Электростатическая сила, вызванная градиентом электрического потенциала - катионы, такие как протоны H + имеют тенденцию диффундировать вниз по электрическому потенциалу анионы в противоположном направлении.
Эти два градиента, взятые вместе, можно выразить как электрохимический градиент, обеспечивающий энергию для абиогенного синтеза. Движущую силу протонов можно описать как меру потенциальной энергии, запасенной в виде комбинации градиентов протонов и напряжения на мембране (разницы в концентрации протонов и электрическом потенциале). [ 152 ]
Поверхности минеральных частиц внутри глубоководных гидротермальных источников обладают каталитическими свойствами, подобными свойствам ферментов, и могут создавать простые органические молекулы, такие как метанол (CH 3 OH) и муравьиную , уксусную и пировиноградную кислоты, из растворенного CO 2 в вода, если приводится в действие приложенным напряжением или реакцией с H 2 или H 2 S. [ 221 ] [ 222 ]
Исследование, опубликованное Мартином в 2016 году, подтверждает тезис о том, что жизнь зародилась в гидротермальных источниках. [ 223 ] [ 224 ] что спонтанная химия в земной коре, вызванная взаимодействием камня и воды в условиях неравновесия, термодинамически лежит в основе возникновения жизни. [ 225 ] [ 226 ] и что первоначальные линии архей и бактерий были H 2 -зависимыми автотрофами, которые использовали CO 2 в качестве терминального акцептора в энергетическом метаболизме. [ 227 ] На основании этих данных Мартин предполагает, что выживание LUCA «могло сильно зависеть от геотермальной энергии источника». [ 10 ] Предполагается, что поры глубоководных гидротермальных источников были заняты мембраносвязанными отсеками, которые способствовали биохимическим реакциям. [ 228 ] [ 229 ] Метаболические промежуточные продукты в цикле Кребса, глюконеогенезе, путях биосинтеза аминокислот, гликолизе, пентозофосфатном пути, включая сахара, такие как рибоза, и предшественники липидов, могут возникать неферментативно в условиях, характерных для глубоководных щелочных гидротермальных источников. [ 230 ]
Если глубоководные гидротермальные условия были местом зарождения жизни, то абиогенез мог произойти уже в 4,0-4,2 млрд лет назад. Если бы жизнь развивалась в океане на глубине более десяти метров, она была бы защищена как от ударов, так и от тогдашнего высокого уровня ультрафиолетового излучения Солнца. Доступная энергия в гидротермальных источниках максимальна при температуре 100–150 ° C — температуре, при которой живут гипертермофильные бактерии и термоацидофильные археи . [ 231 ] [ 232 ] Аргументы против гидротермального происхождения жизни утверждают, что гипертермофилия была результатом конвергентной эволюции бактерий и архей и что мезофильная среда. более вероятной была бы [ 233 ] [ 234 ] Эта гипотеза, выдвинутая в 1999 году Галтье, была выдвинута за год до открытия гидротермального поля Затерянного города, где средняя температура гидротермальных жерл белого курильщика составляет ~45-90 °C. [ 235 ] Умеренные температуры и щелочная морская вода в Затерянном городе теперь являются излюбленным местом гидротермальных источников в отличие от кислых, высокотемпературных (~ 350 ° C) черных курильщиков.
Аргументы против установки вентиляции
[ редактировать ]Производство пребиотических органических соединений в гидротермальных источниках оценивается в 1x10 8 кг/год −1 . [ 236 ] Хотя в жерлах обнаружено большое количество ключевых пребиотических соединений, таких как метан, их концентрации гораздо ниже, чем оценки в среде эксперимента Миллера-Юри. В случае метана скорость производства в жерлах примерно на 2-4 порядка ниже, чем прогнозируемые количества в приземной атмосфере эксперимента Миллера-Юри . [ 236 ] [ 237 ]
Другие аргументы против зарождения жизни в океанических жерлах включают неспособность концентрировать пребиотические материалы из-за сильного разбавления морской водой. В этой открытой системе происходит циклическое соединение минералов, образующих жерла, оставляя мало времени для накопления. [ 238 ] Все современные клетки полагаются на фосфаты и калий для формирования нуклеотидного остова и белка соответственно, что делает вероятным, что первые формы жизни также разделяли эти функции. Эти элементы не были доступны в больших количествах в архейских океанах, поскольку оба в основном образуются в результате выветривания континентальных пород на суше, вдали от жерловых жерл. Подводные гидротермальные источники не способствуют реакциям конденсации, необходимым для полимеризации с образованием макромолекул. [ 239 ] [ 240 ]
Более старый аргумент заключался в том, что ключевые полимеры инкапсулировались в везикулы после конденсации, чего предположительно не могло произойти в соленой воде из-за высоких концентраций ионов. Однако, хотя и верно, что соленость ингибирует образование везикул из смесей жирных кислот с низким разнообразием, [ 241 ] Образование везикул из более широкой и реалистичной смеси жирных кислот и 1-алканолов более устойчиво. [ 242 ] [ 241 ]
Поверхностные водоемы
[ редактировать ]Поверхностные водоемы создают среду, способную высыхать и повторно увлажняться. Продолжительные циклы влажно-сухой позволяют концентрировать пребиотические соединения и проводить реакции конденсации для полимеризации макромолекул. Более того, озера и пруды на суше допускают попадание обломков в результате выветривания континентальных пород, содержащих апатит , наиболее распространенный источник фосфатов, необходимых для нуклеотидных остовов. Количество обнаженной континентальной коры в Гадее неизвестно, но модели ранних глубин океана и скорости роста океанских островов и континентальной коры позволяют предположить, что существовала обнаженная земля. [ 243 ] Еще одним доказательством начала жизни на поверхности является необходимость УФ-излучения для функционирования организма. УФ необходим для образования пары нуклеотидных оснований U+C путем частичного гидролиза и потери азотистых оснований. [ 244 ] В то же время УФ может быть вредным и стерилизовать жизнь, особенно для простых ранних форм жизни с небольшой способностью восстанавливать радиационные повреждения. Уровни радиации от молодого Солнца, вероятно, были выше, и при отсутствии озонового слоя вредные коротковолновые УФ-лучи достигли бы поверхности Земли. Чтобы жизнь зародилась, необходима защищенная среда с притоком источников, подвергающихся воздействию УФ-излучения, чтобы одновременно принести пользу и защитить от УФ-излучения. Защита подо льдом, жидкой водой, минеральными поверхностями (например, глиной) или реголитом возможна в различных условиях поверхностных вод. В то время как глубоководные жерла могут быть источником дождя из материалов, обнаженных на поверхность, вероятность концентрации снижается из-за открытой системы океана. [ 245 ]
Горячие источники
[ редактировать ]Большинство ветвящихся филогений являются термофильными или гипертермофильными, что позволяет предположить, что Последний универсальный общий предок (LUCA) и предыдущие формы жизни были столь же термофильными. Горячие источники образуются в результате нагрева грунтовых вод за счет геотермальной активности. Это пересечение обеспечивает приток материала из глубоко проникающих вод и поверхностного стока, переносящего размытые континентальные отложения. Взаимосвязанные системы подземных вод создают механизм распространения жизни на более широкую территорию. [ 246 ]
Мулкиджанян и соавторы утверждают, что морская среда не обеспечивает ионный баланс и состав, повсеместно встречающийся в клетках, или ионы, необходимые для основных белков и рибозимов, особенно в отношении высокого содержания калия. + / Что + соотношение, Мн 2+ , Зн 2+ и концентрации фосфатов. Они утверждают, что единственная среда, имитирующая необходимые условия на Земле, — это горячие источники, подобные Камчатским. [ 247 ] Минеральные отложения в этих средах в бескислородной атмосфере будут иметь подходящий pH (в то время как нынешние бассейны в насыщенной кислородом атмосфере не будут иметь этого), содержать осадки фотокаталитических сульфидных минералов, которые поглощают вредное ультрафиолетовое излучение, иметь циклы влажно-высыхания, которые концентрируют растворы субстрата до концентраций, поддающихся воздействию спонтанное образование биополимеров [ 248 ] [ 249 ] Создаваемые как химическими реакциями в гидротермальной среде, так и воздействием ультрафиолетового света во время транспортировки из жерл в соседние водоемы, которые способствуют образованию биомолекул. [ 250 ] Предполагаемая предбиотическая среда похожа на гидротермальные источники с дополнительными компонентами, которые помогают объяснить особенности LUCA. [ 247 ] [ 171 ]
Филогеномный и геохимический анализ белков, вероятно связанных с LUCA, показывает, что ионный состав его внутриклеточной жидкости идентичен составу горячих источников. Рост LUCA, вероятно, зависел от синтезированного органического вещества. [ 247 ] Эксперименты показывают, что РНК-подобные полимеры можно синтезировать с помощью циклического влажно-сухого цикла и воздействия УФ-излучения. Эти полимеры после конденсации инкапсулировались в везикулы. [ 241 ] Потенциальными источниками органики в горячих источниках могли быть перенос частиц межпланетной пыли, внеземные снаряды или атмосферный или геохимический синтез. Во время Гадея на вулканических массивах могло быть много горячих источников. [ 171 ]
Умеренные поверхностные водоемы
[ редактировать ]Гипотеза мезофильного гипотезы Опарина - начала в поверхностных водоемах возникла из концепции Дарвина о « маленьком теплом пруду » и Холдейна . Пресноводные водоемы в умеренном климате могут накапливать пребиотические материалы, обеспечивая при этом подходящие условия окружающей среды, способствующие развитию простых форм жизни. Климат во время архея до сих пор остается широко обсуждаемой темой, поскольку существует неопределенность относительно того, как тогда выглядели континенты, океаны и атмосфера. Реконструкции атмосферы архея на основе геохимических данных и моделей показывают, что присутствовало достаточное количество парниковых газов для поддержания температуры поверхности в пределах 0–40 ° C. Согласно этому предположению, существует большее количество ниш с умеренной температурой, в которых могла бы зародиться жизнь. [ 251 ]
Веские доказательства мезофилии, полученные в результате биомолекулярных исследований, включают нуклеотидный термометр G+C Галтье . G+C более распространены у термофилов из-за дополнительной стабильности дополнительной водородной связи, отсутствующей между нуклеотидами A+T. Секвенирование рРНК на различных современных формах жизни показывает, что реконструированное содержание G+C в LUCA , вероятно, характерно для умеренных температур. [ 234 ]
Хотя большинство современных филогений являются термофильными или гипертермофильными, возможно, что их широкое разнообразие сегодня является продуктом конвергентной эволюции и горизонтального переноса генов, а не унаследованным признаком от LUCA. [ 252 ] Обратная гиразная топоизомераза обнаружена исключительно у термофилов и гипертермофилов, поскольку она позволяет скручивать ДНК. [ 253 ] Для функционирования фермента обратной гиразы требуется АТФ , оба из которых представляют собой сложные биомолекулы. Если предположить, что происхождение жизни связано с простым организмом, у которого еще не развилась мембрана, не говоря уже об АТФ, это сделало бы существование обратной гиразы маловероятным. Более того, филогенетические исследования показывают, что обратная гираза имела архейное происхождение и передалась бактериям путем горизонтального переноса генов. Это означает, что обратная гираза не присутствовала в LUCA. [ 254 ]
Ледяная поверхность водоемов
[ редактировать ]Теории происхождения жизни из холодного старта основаны на идее, что на ранней Земле могли быть достаточно холодные регионы, чтобы можно было найти большой ледяной покров. Модели звездной эволюции предсказывают, что светимость Солнца была примерно на 25% слабее, чем сегодня. Фьюлнер утверждает, что, хотя это значительное снижение солнечной энергии привело бы к образованию ледяной планеты, существуют убедительные доказательства присутствия жидкой воды, возможно, вызванной парниковым эффектом. Это создало бы раннюю Землю с жидкими океанами и ледяными полюсами. [ 255 ]
Таяние льда, образующегося на ледяных щитах, или таяние ледников создает бассейны с пресной водой — еще одну нишу, способную испытывать циклы влажно-высыхания. Хотя эти водоемы, существующие на поверхности, будут подвергаться интенсивному УФ-излучению, водоемы внутри и подо льдом достаточно защищены, оставаясь при этом связанными с участками, подверженными УФ-излучению, через ледяные трещины. Предположения о воздействии таяния льда позволяют пресной воде в сочетании с метеоритными примесями стать популярным сосудом для пребиотических компонентов. [ 256 ] Обнаружено, что уровни хлорида натрия в морской воде дестабилизируют самосборку мембран жирных кислот, что делает пресноводные условия привлекательными для ранней мембранной жизни. [ 257 ]
В ледяной среде более высокая скорость реакции, которая происходит в теплой среде, будет заменена повышенной стабильностью и накоплением более крупных полимеров. [ 258 ] Эксперименты, имитирующие условия Европы при температуре ~ 20 ° C, позволили синтезировать аминокислоты и аденин, показывая, что синтез типа Миллера-Юри все еще может происходить при низких температурах. [ 259 ] В мире РНК рибозим имел бы даже больше функций, чем в более позднем мире ДНК-РНК-белок. Чтобы РНК функционировала, она должна иметь возможность сворачиваться, а этому процессу препятствует температура выше 30 °C. Хотя сворачивание РНК у психрофильных организмов происходит медленнее, этот процесс протекает более успешно, поскольку гидролиз также происходит медленнее. Более короткие нуклеотиды не пострадают от более высоких температур. [ 260 ] [ 261 ]
Внутри континентальной коры
[ редактировать ]Альтернативная геологическая среда была предложена геологом Ульрихом Шрайбером и физиком-химиком Кристианом Майером: континентальная кора . [ 262 ] Зоны тектонических разломов могут представлять собой стабильную и хорошо защищенную среду для долгосрочной пребиотической эволюции. Внутри этих систем трещин и полостей основными растворителями являются вода и углекислый газ. Их фазовое состояние будет зависеть от местных условий температуры и давления и может варьироваться от жидкого, газообразного и сверхкритического . При образовании двух отдельных фаз (например, жидкой воды и сверхкритического диоксида углерода на глубинах чуть более 1 км) система обеспечивает оптимальные условия для реакций фазового переноса . Одновременно в состав зон тектонических разломов поступает множество неорганических продуктов (например, оксид углерода, водород, аммиак, цианистый водород, азот и даже фосфат из растворенного апатита) и простых органических молекул, образующихся в результате гидротермальной химии (например, аминокислоты, длинноцепочечные амины, жирные кислоты, длинноцепочечные альдегиды). [ 263 ] [ 264 ] Наконец, обилие минеральных поверхностей обеспечивает богатый выбор каталитической активности.
Особенно интересный участок зон тектонических разломов расположен на глубине около 1000 м. Для углекислотной части объемного растворителя он обеспечивает температурно-барические условия вблизи точки фазового перехода между сверхкритическим и газообразным состоянием. Это приводит к образованию естественной зоны накопления липофильных органических молекул , хорошо растворяющихся в сверхкритическом CO 2 , но не в газообразном состоянии, что приводит к их локальному осаждению. [ 265 ] Периодические изменения давления, например, вызванные деятельностью гейзеров или приливными воздействиями, приводят к периодическим фазовым переходам, удерживая местную реакционную среду в постоянном неравновесном состоянии . В присутствии амфифильных соединений (таких как длинноцепочечные амины и жирные кислоты, упомянутые выше) образуются последующие поколения везикул. [ 266 ] которые постоянно и эффективно отбираются из-за их стабильности. [ 267 ] Полученные структуры могут стать гидротермальными источниками, а также горячими источниками с сырьем для дальнейшего развития.
Гомохиральность
[ редактировать ]
Гомохиральность — это геометрическая однородность материалов, состоящих из хиральных (незеркально-симметричных) единиц. В живых организмах используются молекулы, имеющие одинаковую хиральность (руконаправленность): почти без исключений, [ 269 ] аминокислоты являются левосторонними, а нуклеотиды и сахара — правосторонними. Хиральные молекулы могут быть синтезированы, но в отсутствие хирального источника или хирального катализатора они образуются в виде 50/50 (рацемической) смеси обеих форм . Известные механизмы получения нерацемических смесей из рацемических исходных материалов включают: асимметричные физические законы, такие как электрослабое взаимодействие ; асимметричная среда, например, вызванная круговой поляризацией света, кристаллами кварца или вращением Земли, статистические колебания во время рацемического синтеза, [ 268 ] и спонтанное нарушение симметрии . [ 270 ] [ 271 ] [ 272 ]
После установления будет выбрана хиральность. [ 273 ] Небольшое смещение ( энантиомерный избыток ) в популяции может быть усилено до большого с помощью асимметричного автокатализа , например, в реакции Соаи . [ 274 ] В асимметричном автокатализе катализатором является хиральная молекула, что означает, что хиральная молекула катализирует собственное производство. Первоначальный избыток энантиомеров, например, который может быть вызван поляризованным светом, затем позволяет более распространенному энантиомеру вытеснить другой. [ 275 ]
Гомохиральность, возможно, началась в космическом пространстве, так как на Мерчисонском метеорите аминокислота L-аланин (левая) встречается более чем в два раза чаще, чем ее D (правая) форма, а L-глутаминовая кислота более чем в три раза чаще встречается. так же многочисленн, как и его D-двойник. [ 276 ] [ 277 ] Аминокислоты из метеоритов демонстрируют левостороннее смещение, тогда как сахара демонстрируют преимущественно правостороннее смещение: это то же предпочтение, которое наблюдается в живых организмах, что позволяет предположить абиогенное происхождение этих соединений. [ 278 ]
В эксперименте Роберта Рут-Бернштейна 2010 года «были синтезированы два D-РНК-олигонуклеотида, имеющие инверсные последовательности оснований (D-CGUA и D-AUGC) и соответствующие им L-РНК-олигонуклеотиды (L-CGUA и L-AUGC). их сродство определено к Gly и одиннадцати парам L- и D-аминокислот». Результаты показывают, что гомохиральность, включая направленность кодонов, могла «возникнуть как функция происхождения генетического кода». [ 279 ]
См. также
[ редактировать ]- Аутопоэзис
- Химия пребиотиков на основе формамида - научные усилия, направленные на реконструкцию зарождения жизни.
- Протометаболизм
- Гипотеза GADV-белкового мира – Гипотетическая стадия абиогенеза
- Генетическая рекомбинация - производство потомства с комбинациями признаков, отличающихся от тех, которые обнаружены у любого из родителей.
- Теневая биосфера - Гипотетическая биосфера Земли.
- Марганцевые металлические конкреции
Примечания
[ редактировать ]- ^ Реакции:
- FeS + H 2 S → FeS 2 + 2H + + 2е −
- FeS + H 2 S + CO 2 → FeS 2 + HCOOH
- ^ Реакции:
Реакция 1 : Фаялит + вода → магнетит + водный кремнезем + водород.- 3Fe 2 SiO 4 + 2H 2 O → 2Fe 3 O 4 + 3SiO 2 + 2H 2
- 3Mg 2 SiO 4 + SiO 2 + 4H 2 O → 2Mg 3 Si 2 O 5 (OH) 4
- 2Mg 2 SiO 4 + 3H 2 O → Mg 3 Si 2 O 5 (OH) 4 + Mg(OH) 2
- 2 Ca 2 SiO 4 + 4 H 2 O → 3 CaO · 2 SiO 2 · 3 H 2 O + Ca(OH) 2
Ссылки
[ редактировать ]- ^ Jump up to: а б Уокер, Сара И.; Паккард, Н.; Коди, Джорджия (13 ноября 2017 г.). «Переосмысление происхождения жизни» . Философские труды Королевского общества А. 375 (2109): 20160337. Бибкод : 2017RSPTA.37560337W . дои : 10.1098/rsta.2016.0337 . ПМЦ 5686397 . ПМИД 29133439 .
- ^ Jump up to: а б с «Астробиологическая стратегия НАСА» (PDF) . НАСА . 2015. Архивировано из оригинала (PDF) 22 декабря 2016 года . Проверено 24 сентября 2017 г.
- ^ Трифонов, Эдвард Н. (17 марта 2011 г.). «Словарь определений жизни подсказывает определение» . Журнал биомолекулярной структуры и динамики . 29 (2): 259–266. дои : 10.1080/073911011010524992 . ISSN 0739-1102 . ПМИД 21875147 . S2CID 38476092 .
- ^ Войтек, Мэри А. (6 марта 2021 г.). «Об обнаружении жизни» . НАСА . Архивировано из оригинала 16 августа 2021 года . Проверено 8 марта 2021 г.
- ^ Jump up to: а б Вицани, Гюнтер (2016). «Важные шаги к жизни: от химических реакций к кодированию с использованием агентов» (PDF) . БиоСистемы . 140 : 49–57. Бибкод : 2016BiSys.140...49Вт . doi : 10.1016/j.biosystems.2015.12.007 . ПМИД 26723230 . S2CID 30962295 . Архивировано (PDF) из оригинала 31 октября 2018 г. Проверено 30 октября 2018 г.
- ^ Jump up to: а б Хауэлл, Элизабет (8 декабря 2014 г.). «Как жизнь стала сложной и могла ли она произойти за пределами Земли?» . Журнал астробиологии . Архивировано из оригинала 15 февраля 2018 года . Проверено 14 апреля 2022 г.
{{cite web}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Опарин, Александр Иванович (2003) [1938]. Происхождение жизни Перевод Моргулиса Сергия (2-е изд.). Минеола, Нью-Йорк: Курьер. ISBN 978-0-486-49522-4 . Архивировано из оригинала 2 апреля 2023 года . Проверено 16 июня 2018 г.
- ^ Jump up to: а б Перето, Джули (2005). «Споры о происхождении жизни» (PDF) . Международная микробиология . 8 (1): 23–31. ПМИД 15906258 . Архивировано из оригинала (PDF) 24 августа 2015 года . Проверено 1 июня 2015 г.
- ^ Сравните: Шарф, Калеб; и др. (18 декабря 2015 г.). «Стратегия исследования происхождения жизни» . Астробиология . 15 (12): 1031–1042. Бибкод : 2015AsBio..15.1031S . дои : 10.1089/ast.2015.1113 . ПМЦ 4683543 . ПМИД 26684503 .
Что мы подразумеваем под происхождением жизни (OoL)? ...С начала 20 века словосочетание ОоЛ использовалось для обозначения событий, произошедших при переходе от неживых к живым системам на Земле, т. е. зарождении земной биологии (Опарин, 1924; Холдейн, 1929). . Этот термин в значительной степени заменил более ранние концепции, такие как абиогенез (Kamminga, 1980; Fry, 2000).
- ^ Jump up to: а б с д Вайс, MC; Соуза, Флорида; Мрнявац, Н.; Нойкирхен, С.; Реттгер, М.; Нельсон-Сати, С.; Мартин, ВФ (2016). «Физиология и среда обитания последнего всеобщего общего предка» (PDF) . Природная микробиология . 1 (9): 16116. doi : 10.1038/NMICROBIOL.2016.116 . ПМИД 27562259 . S2CID 2997255 . Архивировано (PDF) из оригинала 29 января 2023 года . Проверено 21 сентября 2022 г.
- ^ Тирар, Стефан (20 апреля 2015 г.). "Абиогенез". Энциклопедия астробиологии . п. 1. дои : 10.1007/978-3-642-27833-4_2-4 . ISBN 978-3-642-27833-4 .
Томас Хаксли (1825–1895) использовал термин абиогенез в важном тексте, опубликованном в 1870 году. Он строго проводил различие между самопроизвольным зарождением, которое он не принимал, и возможностью эволюции материи из инертной в живую без какого-либо влияния. жизни. ... С конца девятнадцатого века эволюционный абиогенез означает увеличение сложности и эволюцию материи от инертного состояния к живому в абиотическом контексте эволюции примитивной Земли.
- ^ Луизи, Пьер Луиджи (2018). Возникновение жизни: от химического происхождения к синтетической биологии . Издательство Кембриджского университета . п. 416. ИСБН 9781108735506 .
Однако поворотный момент от нежизни к жизни никогда не был отражен ни в одной экспериментальной установке. Существует, конечно, несколько гипотез, и такое обилие идей уже означает, что убедительной одной из них у нас нет.
- ^ Грэм, Роберт В. (февраль 1990 г.). «Внеземная жизнь во Вселенной» (PDF) . НАСА (Технический меморандум НАСА 102363). Исследовательский центр Льюиса , Кливленд, Огайо. Архивировано (PDF) из оригинала 3 сентября 2014 года . Проверено 2 июня 2015 г.
- ^ Альтерманн 2009 , с. XVII
- ^ Опарин 1953 , с. Ви
- ^ Уормфлэш, Дэвид; Вармфлэш, Бенджамин (ноябрь 2005 г.). «Жизнь пришла из другого мира?». Научный американец . 293 (5): 64–71. Бибкод : 2005SciAm.293e..64W . doi : 10.1038/scientificamerican1105-64 . ПМИД 16318028 .
- ^ Yarus 2010 , p. 47
- ^ Уорд, Питер; Киршвинк, Джо (2015). Новая история жизни: радикальные открытия о происхождении и эволюции жизни на Земле . Блумсбери Пресс . стр. 39–40. ISBN 978-1-60819-910-5 .
- ^ Шелдон 2005
- ^ Леннокс 2001 , стр. 229–258.
- ^ Jump up to: а б Бернал 1967 г.
- ^ Бальме, DM (1962). «Развитие биологии у Аристотеля и Теофраста: теория спонтанного зарождения». Фронезис . 7 (1–2): 91–104. дои : 10.1163/156852862X00052 .
- ^ Росс 1652 г.
- ^ Добелл 1960
- ^ Фермер 1999 г.
- ^ Левин, Р.; Эверс, К. «Медленная смерть спонтанного зарождения (1668-1859)» . Архивировано из оригинала 26 апреля 2008 года . Проверено 18 апреля 2013 г.
- ^ Опарин 1953 , с. 196
- ^ Тиндаль 1905 , IV, XII (1876), XIII (1878)
- ^ Хорнек, Герда; Клаус, Дэвид М.; Манчинелли, Рокко Л. (март 2010 г.). «Космическая микробиология» . Обзоры микробиологии и молекулярной биологии . 74 (1): 121–156. Бибкод : 2010MMBR...74..121H . дои : 10.1128/MMBR.00016-09 . ПМЦ 2832349 . ПМИД 20197502 .
- ^ Викрамасингхе, Чандра (2011). «Бактериальная морфология, подтверждающая кометную панспермию: переоценка». Международный журнал астробиологии . 10 (1): 25–30. Бибкод : 2011IJAsB..10...25W . CiteSeerX 10.1.1.368.4449 . дои : 10.1017/S1473550410000157 . S2CID 7262449 .
- ^ Рампелотто, PH (2010). «Панспермия: перспективное направление исследований». В: Научная конференция по астробиологии. Абс 5224.
- ^ Чанг, Кеннет (12 сентября 2016 г.). «Видения жизни на Марсе в глубинах Земли» . Нью-Йорк Таймс . Архивировано из оригинала 12 сентября 2016 года . Проверено 12 сентября 2016 г.
- ^ Агилера Мочон, Хуан Антонио (2016). Происхождение жизни на Земле [ Происхождение жизни на Земле ] (на испанском языке). Испания: РБА. ISBN 978-84-473-8386-3 .
- ^ «Письмо № 7471 Чарльза Дарвина Джозефу Далтону Хукеру, 1 февраля (1871 г.)» . Дарвиновский заочный проект . Архивировано из оригинала 7 июля 2020 года . Проверено 7 июля 2020 г.
- ^ Приску, Джон К. «Происхождение и эволюция жизни на замерзшей Земле» . Округ Арлингтон, Вирджиния: Национальный научный фонд . Архивировано из оригинала 18 декабря 2013 года . Проверено 1 марта 2014 г.
- ^ Маршалл, Майкл (11 ноября 2020 г.). «Предположение Чарльза Дарвина о молодости, вероятно, было верным» . Новости Би-би-си . Архивировано из оригинала 11 ноября 2020 года . Проверено 11 ноября 2020 г.
- ^ Бахадур, Кришна (1973). «Фотохимическое образование самоподдерживающихся коацерватов» (PDF) . Труды Индийской национальной академии наук . 39 (4): 455–467. дои : 10.1016/S0044-4057(75)80076-1 . ПМИД 1242552 . Архивировано из оригинала (PDF) 19 октября 2013 года.
- ^ Бахадур, Кришна (1975). «Фотохимическое образование самоподдерживающихся коацерватов» . Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene (Центральный журнал бактериологии, паразитологии, инфекционных заболеваний и гигиены) . 130 (3): 211–218. дои : 10.1016/S0044-4057(75)80076-1 . OCLC 641018092 . ПМИД 1242552 . Архивировано из оригинала 13 декабря 2022 года . Проверено 13 декабря 2022 г.
- ^ Брайсон 2004 , стр. 300–302.
- ^ Бернал 1951
- ^ Мартин, Уильям Ф. (январь 2003 г.). «О происхождении клеток: гипотеза эволюционных переходов от абиотической геохимии к хемоавтотрофным прокариотам и от прокариотов к ядросодержащим клеткам» . Фил. Пер. Р. Сок. Лонд. А. 358 (1429): 59–83. дои : 10.1098/rstb.2002.1183 . ПМК 1693102 . ПМИД 12594918 .
- ^ Бернал, Джон Десмонд (сентябрь 1949 г.). «Физическая основа жизни». Труды Физического общества, раздел А. 62 (9): 537–558. Бибкод : 1949PPSA...62..537B . дои : 10.1088/0370-1298/62/9/301 . S2CID 83754271 .
- ^ Миллер, Стэнли Л. (15 мая 1953 г.). «Производство аминокислот в возможных условиях примитивной Земли». Наука . 117 (3046): 528–529. Бибкод : 1953Sci...117..528M . дои : 10.1126/science.117.3046.528 . ПМИД 13056598 .
- ^ Паркер, Эрик Т.; Кливс, Хендерсон Дж.; Дворкин, Джейсон П.; и др. (5 апреля 2011 г.). «Первоначальный синтез аминов и аминокислот в H 2 эксперименте Миллера 1958 года с искровым разрядом, богатым S» . ПНАС . 108 (14): 5526–5531. Бибкод : 2011PNAS..108.5526P . дои : 10.1073/pnas.1019191108 . ПМК 3078417 . ПМИД 21422282 .
- ^ Бернал 1967 , с. 143
- ^ Jump up to: а б Кливс, Х. Джеймс; Чалмерс, Джон Х.; Ласкано, Антонио ; и др. (апрель 2008 г.). «Переоценка пребиотического органического синтеза в нейтральных планетарных атмосферах». Происхождение жизни и эволюция биосфер . 38 (2): 105–115. Бибкод : 2008OLEB...38..105C . дои : 10.1007/s11084-007-9120-3 . ПМИД 18204914 . S2CID 7731172 .
- ^ Чиба, Кристофер Ф. (13 мая 2005 г.). «Переосмысление ранней атмосферы Земли». Наука . 308 (5724): 962–963. дои : 10.1126/science.1113157 . ПМИД 15890865 . S2CID 93303848 .
- ^ Бартон и др. 2007 , стр. 93–95
- ^ Бада и Ласкано 2009 , стр. 56–57
- ^ Бада, Джеффри Л .; Ласкано, Антонио (2 мая 2003 г.). «Суп из пребиотиков – новый взгляд на эксперимент Миллера» (PDF) . Наука . 300 (5620): 745–746. дои : 10.1126/science.1085145 . ПМИД 12730584 . S2CID 93020326 . Архивировано (PDF) из оригинала 4 марта 2016 года . Проверено 13 июня 2015 г.
- ^ Мадау, Пьеро; Дикинсон, Марк (18 августа 2014 г.). «История космического звездообразования» . Ежегодный обзор астрономии и астрофизики . 52 (1): 415–486. arXiv : 1403.0007 . Бибкод : 2014ARA&A..52..415M . doi : 10.1146/annurev-astro-081811-125615 . S2CID 658354 . Архивировано из оригинала 1 июля 2022 года . Проверено 8 декабря 2023 г.
- ^ Мариго, Паола; и др. (6 июля 2020 г.). «Образование углеродных звезд, как видно через немонотонное соотношение начальных и конечных масс» . Природная астрономия . 152 (11): 1102–1110. arXiv : 2007.04163 . Бибкод : 2020НатАс...4.1102М . дои : 10.1038/s41550-020-1132-1 . S2CID 220403402 . Архивировано из оригинала 16 февраля 2023 года . Проверено 7 июля 2020 г.
- ^ «WMAP-Жизнь во Вселенной» . Архивировано из оригинала 29 января 2023 года . Проверено 27 сентября 2019 г.
- ^ «Формирование солнечных систем: теория солнечных небуляров» . Массачусетский университет в Амхерсте . Архивировано из оригинала 27 сентября 2019 года . Проверено 27 сентября 2019 г.
- ^ «Возраст Земли» . Геологическая служба США . 9 июля 2007 года. Архивировано из оригинала 23 декабря 2005 года . Проверено 10 января 2006 г.
- ^ Далримпл 2001 , стр. 205–221.
- ^ Fesenkov 1959 , p. 9
- ^ Боттке, ВФ; Вокруглицкий, Д.; Марчи, С.; Мошенничество, Т.; Скотт, ERD; Вейрих, младший; Левисон, Х. (17 апреля 2015 г.). «Датирование удара по образованию Луны с астероидными метеоритами» . Наука . 348 (6232): 321–323. Бибкод : 2015Sci...348..321B . дои : 10.1126/science.aaa0602 . ПМИД 25883354 . S2CID 206632612 .
- ^ Кастинг, Джеймс Ф. (12 февраля 1993 г.). «Ранняя атмосфера Земли» (PDF) . Наука . 259 (5097): 920–926. Бибкод : 1993Sci...259..920K . дои : 10.1126/science.11536547 . ПМИД 11536547 . S2CID 21134564 . Архивировано из оригинала (PDF) 10 октября 2015 года . Проверено 28 июля 2015 г.
- ^ Jump up to: а б с д и ж Фоллманн, Хартмут; Браунсон, Кэрол (ноябрь 2009 г.). «Возвращение к маленькому теплому пруду Дарвина: от молекул к происхождению жизни». Naturwissenschaften . 96 (11): 1265–1292. Бибкод : 2009NW.....96.1265F . дои : 10.1007/s00114-009-0602-1 . ПМИД 19760276 . S2CID 23259886 .
- ^ Морс, Джон (сентябрь 1998 г.). «Геохимия карбонатов Гадейского океана». Водная геохимия . 4 (3/4): 301–319. Бибкод : 1998MinM...62.1027M . дои : 10.1023/А:1009632230875 . S2CID 129616933 .
- ^ Спи, Норман Х.; Занле, Кевин Дж.; Лупу, Роксана Э. (13 сентября 2014 г.). «Земные последствия удара, образовавшего Луну» . Философские труды Королевского общества A: Математические, физические и технические науки . 372 (2024): 20130172. Бибкод : 2014RSPTA.37230172S . дои : 10.1098/rsta.2013.0172 . ПМИД 25114303 . S2CID 6902632 .
- ^ Морс, Джон В.; Маккензи, Фред Т. (1998). «[Название не найдено]» . Водная геохимия . 4 (3/4): 301–319. дои : 10.1023/А:1009632230875 . S2CID 129616933 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Кроули, Джеймс Л.; Майерс, Джон С.; Сильвестр, Пол Дж; Кокс, Ричард А. (май 2005 г.). «Обломочный циркон из Джек-Хиллз и горы Нарриер, Западная Австралия: свидетельства разнообразия материнских пород > 4,0 млрд лет» . Журнал геологии . 113 (3): 239–263. Бибкод : 2005JG....113..239C . дои : 10.1086/428804 . S2CID 140715676 . Архивировано из оригинала 16 декабря 2023 года . Проверено 8 декабря 2023 г.
- ^ Уайльд, Саймон А.; Вэлли, Джон В.; Пек, Уильям Х.; Грэм, Колин М. (11 января 2001 г.). «Свидетельства обломочных цирконов о существовании континентальной коры и океанов на Земле 4,4 миллиарда лет назад» (PDF) . Природа . 409 (6817): 175–178. Бибкод : 2001Natur.409..175W . дои : 10.1038/35051550 . ПМИД 11196637 . S2CID 4319774 . Архивировано (PDF) из оригинала 5 июня 2015 года . Проверено 3 июня 2015 г.
- ^ Коренага, июнь (декабрь 2008 г.). «Тектоника плит, паводковые базальты и эволюция океанов Земли» . Терра Нова . 20 (6): 419–439. Бибкод : 2008TeNov..20..419K . дои : 10.1111/j.1365-3121.2008.00843.x . S2CID 36766331 .
- ^ Розинг, Миник Т.; Берд, Деннис К.; Спи, Норман Х.; и др. (22 марта 2006 г.). «Возникновение континентов – Очерк геологических последствий фотосинтеза» . Палеогеография, Палеоклиматология, Палеоэкология . 232 (2–4): 99–113. Бибкод : 2006PPP...232...99R . дои : 10.1016/j.palaeo.2006.01.007 . Архивировано (PDF) из оригинала 14 июля 2015 года . Проверено 8 июня 2015 г.
- ^ Тера, Фуад; Папанастассиу, Д.А.; Вассербург, GJ (апрель 1974 г.). «Изотопные доказательства заключительного лунного катаклизма» . Письма о Земле и планетологии . 22 (1): 1–21. Бибкод : 1974E&PSL..22....1T . дои : 10.1016/0012-821x(74)90059-4 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Стоффлер, Д. (1 января 2006 г.). «История кратеров и лунная хронология» . Обзоры по минералогии и геохимии . 60 (1): 519–596. Бибкод : 2006RvMG...60..519S . дои : 10.2138/rmg.2006.60.05 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Спи, Норман Х.; Занле, Кевин Дж.; Кастинг, Джеймс Ф.; Моровиц, Гарольд Дж. (декабрь 1989 г.). «Уничтожение экосистем в результате воздействия крупных астероидов на раннюю Землю» . Природа . 342 (6246): 139–142. Бибкод : 1989Natur.342..139S . дои : 10.1038/342139a0 . ПМИД 11536616 . S2CID 1137852 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Фассетт, Калеб И.; Минтон, Дэвид А. (23 июня 2013 г.). «Ударная бомбардировка планет земной группы и ранняя история Солнечной системы» . Природа Геонауки . 6 (7): 520–524. Бибкод : 2013NatGe...6..520F . дои : 10.1038/ngeo1841 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Абрамов Олег; Мойзис, Стивен Дж. (май 2009 г.). «Микробная обитаемость Гадейской Земли во время поздней сильной бомбардировки» . Природа . 459 (7245): 419–422. Бибкод : 2009Natur.459..419A . дои : 10.1038/nature08015 . ПМИД 19458721 . S2CID 3304147 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Бенке, Патрик; Харрисон, Т. Марк (12 сентября 2016 г.). «Иллюзорные поздние тяжелые бомбардировки» . Труды Национальной академии наук . 113 (39): 10802–10806. Бибкод : 2016PNAS..11310802B . дои : 10.1073/pnas.1611535113 . ПМК 5047187 . ПМИД 27621460 .
- ^ Зеллнер, Николь Э.Б. (3 мая 2017 г.). «Катаклизма больше нет: новые взгляды на время и доставку лунных ударников» . Происхождение жизни и эволюция биосфер . 47 (3): 261–280. arXiv : 1704.06694 . Бибкод : 2017OLEB...47..261Z . дои : 10.1007/s11084-017-9536-3 . ПМК 5602003 . ПМИД 28470374 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Лоу, Дональд Р.; Байерли, Гэри Р. (1 апреля 2018 г.). «Земные записи поздней тяжелой бомбардировки» . Новые обзоры астрономии . 81 : 39–61. Бибкод : 2018НовыйAR..81...39L . дои : 10.1016/j.newar.2018.03.002 .
- ^ Дэвис 1999 , с. 155
- ^ Бок и Гуд 1996
- ^ Jump up to: а б Додд, Мэтью С.; Папино, Доминик; Гренне, Тор; и др. (1 марта 2017 г.). «Доказательства ранней жизни в старейших гидротермальных источниках Земли выпадают» . Природа . 543 (7643): 60–64. Бибкод : 2017Natur.543...60D . дои : 10.1038/nature21377 . ПМИД 28252057 . Архивировано из оригинала 8 сентября 2017 года . Проверено 2 марта 2017 г.
- ^ Отомо, Йоко; Какегава, Такеши; Исида, Акизуми; и др. (январь 2014 г.). «Доказательства наличия биогенного графита в метаосадочных породах раннего архейского периода Иисуса». Природа Геонауки . 7 (1): 25–28. Бибкод : 2014NatGe...7... 25O дои : 10.1038/ngeo2025 .
- ^ Ноффке, Нора ; Кристиан, Дэниел; Уэйси, Дэвид; Хейзен, Роберт М. (16 ноября 2013 г.). «Микробно-индуцированные осадочные структуры, фиксирующие древнюю экосистему в возраст около формации Гё Дрессер, Пилбара, Западная Австралия, 3,48 года» . Астробиология . 13 (12): 1103–1124. Бибкод : 2013AsBio..13.1103N . дои : 10.1089/ast.2013.1030 . ПМК 3870916 . ПМИД 24205812 .
- ^ Дэвис 1999
- ^ Хасенкам, Т.; Андерссон, член парламента; Далби, КНЦ; Маккензи, DMA; Розинг, Монтана (2017). «Элементы эоархейской жизни, заключенные в минеральных включениях». Природа . 548 (7665): 78–81. Бибкод : 2017Natur.548...78H . дои : 10.1038/nature23261 . ПМИД 28738409 . S2CID 205257931 .
- ^ О'Донохью, Джеймс (21 августа 2011 г.). «Самые старые достоверные окаменелости показывают, что ранняя жизнь была на пляже» . Новый учёный . 211 : 13. дои : 10.1016/S0262-4079(11)62064-2 . Архивировано из оригинала 30 июня 2015 года.
- ^ Уэйси, Дэвид; Килберн, Мэтт Р.; Сондерс, Мартин; и др. (октябрь 2011 г.). «Микроокаменелости клеток, метаболизирующих серу, в породах Западной Австралии возрастом 3,4 миллиарда лет». Природа Геонауки . 4 (10): 698–702. Бибкод : 2011NatGe...4..698W . дои : 10.1038/ngeo1238 .
- ^ Белл, Элизабет А.; Бенке, Патрик; Харрисон, Т. Марк; Мао, Венди Л. (24 ноября 2015 г.). «Потенциально биогенный углерод сохранился в цирконе возрастом 4,1 миллиарда лет» . Труды Национальной академии наук . 112 (47): 14518–14521. Бибкод : 2015PNAS..11214518B . дои : 10.1073/pnas.1517557112 . ПМЦ 4664351 . ПМИД 26483481 .
- ^ Баумгартнер, Рафаэль; Ван Кранендонк, Мартин; Уэйси, Дэвид; и др. (2019). «Нанопористый пирит и органическое вещество в строматолитах возрастом 3,5 миллиарда лет свидетельствуют о первозданной жизни» (PDF) . Геология . 47 (11): 1039–1043. Бибкод : 2019Geo....47.1039B . дои : 10.1130/G46365.1 . S2CID 204258554 . Архивировано (PDF) из оригинала 5 декабря 2020 года . Проверено 10 января 2021 г.
- ^ Джокич, Тара; Ван Кранендонк, Мартин Дж.; Кэмпбелл, Кэтлин А.; Уолтер, Малкольм Р.; Уорд, Колин Р. (9 мая 2017 г.). «Самые ранние признаки жизни на суше сохранились в отложениях горячих источников примерно 3,5 Гао» . Природные коммуникации . 8 : 15263. Бибкод : 2017NatCo...815263D . дои : 10.1038/ncomms15263 . ПМК 5436104 . ПМИД 28486437 .
- ^ Беттс, Холли С.; Путтик, Марк Н.; Кларк, Джеймс В.; Уильямс, Том А.; Донохью, Филип CJ; Пизани, Давиде (20 августа 2018 г.). «Комплексные геномные и ископаемые данные проливают свет на раннюю эволюцию жизни и происхождение эукариот» . Экология и эволюция природы . 2 (10): 1556–1562. Бибкод : 2018NatEE...2.1556B . дои : 10.1038/s41559-018-0644-x . ПМК 6152910 . ПМИД 30127539 .
- ^ Ландау, Элизабет (12 октября 2016 г.). «Строительные блоки жизни» исходят из звездного света . НАСА . Архивировано из оригинала 13 октября 2016 года . Проверено 13 октября 2016 г.
- ^ Jump up to: а б Эренфройнд, Паскаль; Ками, Ян (декабрь 2010 г.). «Космическая химия углерода: от межзвездной среды до ранней Земли» . Перспективы Колд-Спринг-Харбор в биологии . 2 (12): а002097. doi : 10.1101/cshperspect.a002097 . ПМЦ 2982172 . ПМИД 20554702 .
- ^ Гебалле, Томас Р.; Нахарро, Франциско; Файгер, Дональд Ф .; и др. (10 ноября 2011 г.). «Инфракрасные диффузные межзвездные полосы в районе центра Галактики». Природа . 479 (7372): 200–202. arXiv : 1111.0613 . Бибкод : 2011Natur.479..200G . дои : 10.1038/nature10527 . ПМИД 22048316 . S2CID 17223339 .
- ^ Клайце 2001
- ^ Jump up to: а б с д Гувер, Рэйчел (21 февраля 2014 г.). «Нужно отслеживать органические наночастицы по всей Вселенной? У НАСА есть для этого приложение» . Исследовательский центр Эймса . НАСА . Архивировано из оригинала 6 сентября 2015 года . Проверено 22 июня 2015 г.
- ^ Гончарук Владислав В.; Зуй, О.В. (февраль 2015 г.). «Вода и углекислый газ как основные предшественники органического вещества на Земле и в космосе». Журнал химии воды и технологий . 37 (1): 2–3. Бибкод : 2015JWCT...37....2G . дои : 10.3103/S1063455X15010026 . S2CID 97965067 .
- ^ Абу Мрад, Нинетт; Виноградов, Василиса; Дюверне, Фабрис; и др. (2015). «Лабораторное экспериментальное моделирование: химическая эволюция органического вещества из аналогов межзвездного и кометного льда» . Бюллетень Королевского общества наук Льежа . 84 : 21–32. Бибкод : 2015BSRSL..84...21A . Архивировано из оригинала 13 апреля 2015 года . Проверено 6 апреля 2015 г.
- ^ Оба, Ясухиро; и др. (26 апреля 2022 г.). «Идентификация широкого разнообразия внеземных пуриновых и пиримидиновых азотистых оснований в углеродистых метеоритах» . Природные коммуникации . 13 (2008): 2008. Бибкод : 2022NatCo..13.2008O . дои : 10.1038/s41467-022-29612-x . ПМЦ 9042847 . ПМИД 35473908 . S2CID 248402205 .
- ^ « В комете обнаружен «химикат жизни»» . Новости Би-би-си . Лондон. 18 августа 2009 года. Архивировано из оригинала 25 мая 2015 года . Проверено 23 июня 2015 г.
- ^ Томпсон, Уильям Рид; Мюррей, Б.Г.; Кхаре, Бишун Нараин ; Саган, Карл (30 декабря 1987 г.). «Окраска и потемнение клатрата метана и других льдов в результате облучения заряженными частицами: применение во внешней солнечной системе». Журнал геофизических исследований . 92 (А13): 14933–14947. Бибкод : 1987JGR....9214933T . дои : 10.1029/JA092iA13p14933 . ПМИД 11542127 .
- ^ Гольдман, Нир; Тэмблин, Исаак (20 июня 2013 г.). «Химия пребиотиков в простой ледяной смеси» . Журнал физической химии А. 117 (24): 5124–5131. Бибкод : 2013JPCA..117.5124G . дои : 10.1021/jp402976n . ПМИД 23639050 . S2CID 5144843 . Архивировано из оригинала 21 июля 2018 года . Проверено 29 августа 2019 г.
- ^ «База данных ИК-спектроскопии НАСА Эймса PAH» . НАСА. Архивировано из оригинала 29 июня 2015 года . Проверено 17 июня 2015 г.
- ^ Jump up to: а б с Хаджинс, Дуглас М.; Баушлихер, Чарльз В. младший; Аламандола, Луи Дж. (10 октября 2005 г.). «Вариации положения пика особенности межзвездного излучения 6,2 мкм: индикатор N в межзвездной популяции полициклических ароматических углеводородов». Астрофизический журнал . 632 (1): 316–332. Бибкод : 2005ApJ...632..316H . CiteSeerX 10.1.1.218.8786 . дои : 10.1086/432495 . S2CID 7808613 .
- ^ Jump up to: а б с Де Марэ, Дэвид Дж.; Аламандола, Луи Дж.; Сэндфорд, Скотт ; и др. (2009). «Космическое распределение химической сложности» . Исследовательский центр Эймса . Маунтин-Вью, Калифорния: НАСА. Архивировано из оригинала 27 февраля 2014 года . Проверено 24 июня 2015 г.
- ^ Jump up to: а б Кэри, Бьорн (18 октября 2005 г.). «Строительные блоки жизни «в изобилии в космосе» » . Space.com . Уотсонвилл, Калифорния: Имагинова . Архивировано из оригинала 26 июня 2015 года . Проверено 23 июня 2015 г.
- ^ Гарсия-Эрнандес, воскресенье. А.; В пятнах, Артур; Гарсиа-Ларио, Питер; и др. (20 ноября 2010 г.). «Образование фуллеренов в H-содержащих планетарных туманностях». Письма астрофизического журнала . 724 (1):L39–L43. arXiv : 1009.4357 . Бибкод : 2010ApJ...724L..39G . дои : 10.1088/2041-8205/724/1/L39 . S2CID 119121764 .
- ^ д'Искья, Марко; Манини, Паола; Мораччи, Марко; и др. (21 августа 2019 г.). «Астрохимия и астробиология: материаловедение в стране чудес?» . Международный журнал молекулярных наук . 20 (17): 4079. doi : 10.3390/ijms20174079 . ПМК 6747172 . ПМИД 31438518 .
- ^ Гудипати, Мурти С.; Ян, Жуй (1 сентября 2012 г.). «Изучение на месте радиационно-индуцированной обработки органических веществ в астрофизических аналогах льда - новые времяпролетные масс-спектроскопические исследования с лазерной десорбцией и лазерной ионизацией». Письма астрофизического журнала . 756 (1): Л24. Бибкод : 2012ApJ...756L..24G . дои : 10.1088/2041-8205/756/1/L24 . S2CID 5541727 .
- ^ Jump up to: а б Галлори, Энцо (июнь 2011 г.). «Астрохимия и происхождение генетического материала». Линцианские счета . 22 (2): 113–118. дои : 10.1007/s12210-011-0118-4 . S2CID 96659714 . «Доклад, представленный на симпозиуме «Астрохимия: молекулы в пространстве и времени» (Рим, 4–5 ноября 2010 г.), спонсируемом Фондом «Гвидо Донегани», Национальной академией Линчеи».
- ^ Мартинс, Зита (февраль 2011 г.). «Органическая химия углеродистых метеоритов». Элементы . 7 (1): 35–40. Бибкод : 2011Элеме...7...35М . дои : 10.2113/gselements.7.1.35 .
- ^ Мартинс, Зита; Ботта, Оливер; Фогель, Мэрилин Л.; и др. (15 июня 2008 г.). «Внеземные азотистые основания в метеорите Мерчисон». Письма о Земле и планетологии . 270 (1–2): 130–136. arXiv : 0806.2286 . Бибкод : 2008E&PSL.270..130M . дои : 10.1016/j.epsl.2008.03.026 . S2CID 14309508 .
- ^ Каллахан, Майкл П.; Смит, Карен Э.; Кливс, Х. Джеймс II; и др. (23 августа 2011 г.). «Углеродистые метеориты содержат широкий спектр внеземных азотистых оснований» . ПНАС . 108 (34): 13995–13998. Бибкод : 2011PNAS..10813995C . дои : 10.1073/pnas.1106493108 . ПМК 3161613 . ПМИД 21836052 .
- ^ Штайгервальд, Джон (8 августа 2011 г.). «Исследователи НАСА: строительные блоки ДНК можно создавать в космосе» . Центр космических полетов Годдарда . НАСА. Архивировано из оригинала 23 июня 2015 года . Проверено 23 июня 2015 г.
- ^ Квок, Вс ; Чжан, Юн (3 ноября 2011 г.). «Смешанные ароматико-алифатические органические наночастицы как носители неидентифицированных свойств инфракрасного излучения». Природа . 479 (7371): 80–83. Бибкод : 2011Природа.479...80К . дои : 10.1038/nature10542 . ПМИД 22031328 . S2CID 4419859 .
- ^ Йоргенсен, Джес К.; Фавр, Сесиль; Бишоп, Сюзанна Э.; и др. (2012). «Обнаружение простейшего сахара, гликоляльдегида, в протозвезде солнечного типа с ALMA» (PDF) . Письма астрофизического журнала . 757 (1): Л4. arXiv : 1208.5498 . Бибкод : 2012ApJ...757L...4J . дои : 10.1088/2041-8205/757/1/L4 . S2CID 14205612 . Архивировано (PDF) из оригинала 24 сентября 2015 г. Проверено 23 июня 2015 г.
- ^ Фурукава, Ёсихиро; Чикараиси, Ёсито; Окоучи, Наахико; и др. (13 ноября 2019 г.). «Внеземные рибоза и другие сахара в примитивных метеоритах» . ПНАС . 116 (49): 24440–24445. Бибкод : 2019PNAS..11624440F . дои : 10.1073/pnas.1907169116 . ПМК 6900709 . ПМИД 31740594 .
- ^ Оро, Джоан; Кимбалл, Обри П. (февраль 1962 г.). «Синтез пуринов в возможных примитивных земных условиях: II. Пуриновые полупродукты из цианида водорода». Архив биохимии и биофизики . 96 (2): 293–313. дои : 10.1016/0003-9861(62)90412-5 . ПМИД 14482339 .
- ^ Кливс II, Хендерсон (2010). «Происхождение биологически кодируемых аминокислот». Журнал теоретической биологии . 263 (4): 490–498. Бибкод : 2010JThBi.263..490C . дои : 10.1016/j.jtbi.2009.12.014 . ПМИД 20034500 .
- ^ Грин, Нью-Джерси, Рассел, Д.А., Таннер, С.Х., Сазерленд, Дж.Д. (2023). «Пребиотический синтез N-формиламинонитрилов и их производных в формамиде» . Журнал Американского химического общества . 145 (19): 10533–10541. дои : 10.1021/jacs.2c13306 . ПМЦ 10197134 . ПМИД 37146260 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бреслоу, Р. (1959). «О механизме формозной реакции». Буквы тетраэдра . 1 (21): 22–26. дои : 10.1016/S0040-4039(01)99487-0 .
- ^ Оро, Жоан (16 сентября 1961 г.). «Механизм синтеза аденина из циановодорода в возможных условиях примитивной Земли». Природа . 191 (4794): 1193–1194. Бибкод : 1961Natur.191.1193O . дои : 10.1038/1911193a0 . ПМИД 13731264 . S2CID 4276712 .
- ^ Jump up to: а б Саладино, Рафаэле; Крестини, Клаудия; Пино, Саманта; и др. (март 2012 г.). «Формамид и происхождение жизни» (PDF) . Обзоры физики жизни . 9 (1): 84–104. Бибкод : 2012PhLRv...9...84S . дои : 10.1016/j.plrev.2011.12.002 . hdl : 2108/85168 . ПМИД 22196896 . Архивировано (PDF) из оригинала 27 января 2023 года . Проверено 29 августа 2019 г.
- ^ Jump up to: а б Саладино, Рафаэле; Ботта, Джорджия; Пино, Саманта; и др. (июль 2012 г.). «От одноуглеродного амид-формамида до РНК все этапы возможны с пребиотической точки зрения». Биохимия . 94 (7): 1451–1456. дои : 10.1016/j.biochi.2012.02.018 . hdl : 11573/515604 . ПМИД 22738728 .
- ^ Марлер, Рут, изд. (3 марта 2015 г.). «НАСА Эймс воспроизводит строительные блоки жизни в лаборатории» . Исследовательский центр Эймса . НАСА. Архивировано из оригинала 5 марта 2015 года . Проверено 5 марта 2015 г.
- ^ Ферус, Мартин; Несворный, Давид; Шпонер, Иржи; и др. (2015). «Химия высоких энергий формамида: единый механизм образования нуклеиновых оснований» . ПНАС . 112 (3): 657–662. Бибкод : 2015PNAS..112..657F . дои : 10.1073/pnas.1412072111 . ПМЦ 4311869 . ПМИД 25489115 .
- ^ Базиль, Бренда; Ласкано, Антонио ; Оро, Джоан (1984). «Пребиотические синтезы пуринов и пиримидинов». Достижения в космических исследованиях . 4 (12): 125–131. Бибкод : 1984АдСпР...4л.125Б . дои : 10.1016/0273-1177(84)90554-4 . ПМИД 11537766 .
- ^ Оргел, Лесли Э. (август 2004 г.). «Возвращение к пребиотику аденину: эвтектика и фотохимия». Происхождение жизни и эволюция биосфер . 34 (4): 361–369. Бибкод : 2004OLEB...34..361O . doi : 10.1023/B:ORIG.0000029882.52156.c2 . ПМИД 15279171 . S2CID 4998122 .
- ^ Робертсон, Майкл П.; Миллер, Стэнли Л. (29 июня 1995 г.). «Эффективный пребиотический синтез цитозина и урацила». Природа . 375 (6534): 772–774. Бибкод : 1995Natur.375..772R . дои : 10.1038/375772a0 . ПМИД 7596408 . S2CID 4351012 .
- ^ Фокс, Дуглас (1 февраля 2008 г.). «Развилась ли жизнь во льду?» . Обнаружить . Архивировано из оригинала 30 июня 2008 года . Проверено 3 июля 2008 г.
- ^ Леви, Мэтью; Миллер, Стэнли Л.; Бринтон, Карен; Бада, Джеффри Л. (июнь 2000 г.). «Пребиотический синтез аденина и аминокислот в условиях Европы». Икар . 145 (2): 609–613. Бибкод : 2000Icar..145..609L . дои : 10.1006/icar.2000.6365 . ПМИД 11543508 .
- ^ Менор-Сальван, Сезар; Руис-Бермехо, Марта; Гусман, Марсело И.; и др. (20 апреля 2009 г.). «Синтез пиримидинов и триазинов во льду: значение для пребиотической химии азотистых оснований». Химия: Европейский журнал . 15 (17): 4411–4418. дои : 10.1002/chem.200802656 . ПМИД 19288488 .
- ^ Рой, Деджани; Наджафян, Катаюн; фон Раге Шлейер, Пол (30 октября 2007 г.). «Химическая эволюция: Механизм образования аденина в пребиотических условиях» . ПНАС . 104 (44): 17272–17277. Бибкод : 2007PNAS..10417272R . дои : 10.1073/pnas.0708434104 . ПМК 2077245 . ПМИД 17951429 .
- ^ Jump up to: а б Френкель-Пинтер, Моран; Саманта, Моусуми; Ашкенаси, Гонен; Леман, Люк Дж. (10 июня 2020 г.). «Пребиотические пептиды: молекулярные центры происхождения жизни» . Химические обзоры . 120 (11): 4707–4765. doi : 10.1021/acs.chemrev.9b00664 . ISSN 0009-2665 . ПМИД 32101414 .
- ^ Кэмпбелл, Томас Д.; февраль, Рио; Маккарти, Джек Т.; Кляйншмидт, Холли Э.; Форсайт, Джей Г.; Брейчер, Пол Дж. (2019). «Конденсация пребиотиков посредством циклического цикла влажно-сухой жизни, регулируемого расплыванием» . Природные коммуникации . 10 (1): 4508. Бибкод : 2019NatCo..10.4508C . дои : 10.1038/s41467-019-11834-1 . ПМК 6778215 . ПМИД 31586058 .
- ^ Маршалл-Боуман, Карина; Охара, Сёхей; Сверженский, Дмитрий А.; Хейзен, Роберт М.; Кливс, Х. Джеймс (октябрь 2010 г.). «Каталитический гидролиз пептидов минеральной поверхностью: значение для химии пребиотиков» . Geochimica et Cosmochimica Acta . 74 (20): 5852–5861. Бибкод : 2010GeCoA..74.5852M . дои : 10.1016/j.gca.2010.07.009 . ISSN 0016-7037 .
- ^ Матрё, Томас; Айккила, Паула; Шой, Беттина; Браун, Дитер; Маст, Кристоф Б. (апрель 2024 г.). «Тепловые потоки обогащают пребиотические строительные блоки и повышают их реакционную способность» . Природа . 628 (8006): 110–116. Бибкод : 2024Natur.628..110M . дои : 10.1038/s41586-024-07193-7 . ISSN 1476-4687 . ПМЦ 10990939 . ПМИД 38570715 .
- ^ Пахт-Горовиц, Мелла (1 января 1974 г.). «Возможная роль глин в синтезе пребиотических пептидов» . Истоки жизни . 5 (1): 173–187. Бибкод : 1974OrLi....5..173P . дои : 10.1007/BF00927022 . ISSN 1573-0875 . ПМИД 4842069 .
- ^ Краснокутский С.А.; Чуанг, К.-Дж.; Ягер, К.; Юбершаар, Н.; Хеннинг, Т. (10 февраля 2022 г.). «Путь к пептидам в космосе через конденсацию атомарного углерода» . Природная астрономия . 6 (3): 381–386. arXiv : 2202.12170 . Бибкод : 2022НатАс...6..381К . дои : 10.1038/s41550-021-01577-9 . ISSN 2397-3366 .
- ^ Краснокутский, Серж А.; Ягер, Корнелия; Хеннинг, Томас; Жеффруа, Клод; Ремори, Квентин Б.; Пуано, Полина (19 апреля 2024 г.). «Образование внеземных пептидов и их производных» . Достижения науки . 10 (16): eadj7179. arXiv : 2405.00744 . Бибкод : 2024SciA...10J7179K . дои : 10.1126/sciadv.adj7179 . ISSN 2375-2548 . ПМК 11023503 . ПМИД 38630826 .
- ^ Ланцет, Дорон (30 декабря 2014 г.). «Системная пребиология-Исследование происхождения жизни» . Лаборатория «Ланцет» . Реховот, Израиль: Отдел молекулярной генетики; Институт науки Вейцмана . Архивировано из оригинала 26 июня 2015 года . Проверено 26 июня 2015 г.
- ^ Сегре, Даниэль; Бен-Эли, Дафна; Димер, Дэвид В.; Ланцет, Дорон (февраль 2001 г.). «Липидный мир» (PDF) . Происхождение жизни и эволюция биосфер . 31 (1–2): 119–145. Бибкод : 2001OLEB...31..119S . дои : 10.1023/А:1006746807104 . ПМИД 11296516 . S2CID 10959497 . Архивировано (PDF) из оригинала 26 июня 2015 года.
- ^ Jump up to: а б с Чен, Ирен А.; Вальде, Питер (июль 2010 г.). «От самособирающихся везикул к протоклеткам» . Перспективы Колд-Спринг-Харбор в биологии . 2 (7): а002170. doi : 10.1101/cshperspect.a002170 . ПМК 2890201 . ПМИД 20519344 .
- ^ Эйген, Манфред ; Шустер, Питер (ноябрь 1977 г.). «Гиперцикл. Принцип естественной самоорганизации. Часть A: Возникновение гиперцикла» (PDF) . Naturwissenschaften . 64 (11): 541–65. Бибкод : 1977NW.....64..541E . дои : 10.1007/bf00450633 . ПМИД 593400 . S2CID 42131267 . Архивировано из оригинала (PDF) 3 марта 2016 года.
- Эйген, Манфред ; Шустер, Питер (1978). «Гиперцикл. Принцип естественной самоорганизации. Часть B: абстрактный гиперцикл» (PDF) . Naturwissenschaften . 65 (1): 7–41. Бибкод : 1978NW.....65....7E . дои : 10.1007/bf00420631 . S2CID 1812273 . Архивировано из оригинала (PDF) 3 марта 2016 года.
- Эйген, Манфред ; Шустер, Питер (июль 1978 г.). «Гиперцикл. Принцип естественной самоорганизации. Часть C: Реалистичный гиперцикл» (PDF) . Naturwissenschaften . 65 (7): 341–369. Бибкод : 1978NW.....65..341E . дои : 10.1007/bf00439699 . S2CID 13825356 . Архивировано из оригинала (PDF) 16 июня 2016 года.
- ^ Маркович, Омер; Ланцет, Дорон (лето 2012 г.). «Для эффективной эволюции необходим избыточный взаимный катализ» . Искусственная жизнь . 18 (3): 243–266. дои : 10.1162/artl_a_00064 . ПМИД 22662913 . S2CID 5236043 .
- ^ Тессера, Марк (2011). «Происхождение эволюции против происхождения жизни: сдвиг парадигмы» . Международный журнал молекулярных наук . 12 (6): 3445–3458. дои : 10.3390/ijms12063445 . ПМК 3131571 . ПМИД 21747687 . Спецвыпуск: «Происхождение жизни 2011».
- ^ «Протоклетки» . Исследование истоков жизни: виртуальная выставка . Округ Арлингтон, Вирджиния: Национальный научный фонд . Архивировано из оригинала 28 февраля 2014 года . Проверено 13 сентября 2023 г.
- ^ Jump up to: а б с Чен, Ирен А. (8 декабря 2006 г.). «Появление клеток в процессе зарождения жизни» . Наука . 314 (5805): 1558–1559. дои : 10.1126/science.1137541 . ПМИД 17158315 .
- ^ Циммер, Карл (26 июня 2004 г.). «Что было до ДНК?» . Обнаружить . Архивировано из оригинала 19 марта 2014 года.
- ^ Онсагер, Ларс (1931). «Взаимные связи в необратимых процессах I» . Физический обзор . 37 (4): 405. Бибкод : 1931ФРв...37..405О . дои : 10.1103/PhysRev.37.405 .
- ^ Онсагер, Ларс (1931). «Взаимные отношения в необратимых процессах II» . Физический обзор . 38 (12): 2265. Бибкод : 1931PhRv...38.2265O . дои : 10.1103/PhysRev.38.2265 .
- ^ Пригожин, Илья (1967). Введение в термодинамику необратимых процессов . Нью-Йорк: Уайли .
- ^ Шапиро, Роберт (июнь 2007 г.). «Простое происхождение жизни» . Научный американец . 296 (6): 46–53. Бибкод : 2007SciAm.296f..46S . doi : 10.1038/scientificamerican0607-46 . ПМИД 17663224 . Архивировано из оригинала 14 июня 2015 года.
- ^ Чанг 2007
- ^ Jump up to: а б с д Лейн, Ник (2015). Жизненно важный вопрос: почему жизнь такая, какая она есть? . Профильные книги . стр. 129–140. ISBN 978-1-78125-036-5 .
- ^ Джордан, Шон Ф.; Нет, Элоиза; Лейн, Ник (6 декабря 2019 г.). «Изопреноиды повышают стабильность мембран жирных кислот при зарождении жизни, что потенциально может привести к раннему разделению липидов» . Фокус на интерфейсе . 9 (6): 20190067. doi : 10.1098/rsfs.2019.0067 . ПМК 6802135 . ПМИД 31641436 .
- ^ Майер, Кристиан; Шрайбер, Ульрих; Давила, Мария Х.; Шмитц, Оливер Дж.; Броня, Амелия; Мейер, Мартин; Кляйн, Джулия; Меккельманн, Свен В. (2018). «Молекулярная эволюция в системе пептид-везикула» . Жизнь . 8 (2): 16. Бибкод : 2018Жизнь....8...16М . дои : 10.3390/life8020016 . ISSN 2075-1729 . ПМК 6027363 . ПМИД 29795023 .
- ^ Майер, Кристиан; Шрайбер, Ульрих; Давила, Мария Х. (1 июня 2015 г.). «Периодическое образование пузырьков в зонах тектонических разломов — идеальный сценарий молекулярной эволюции» . Происхождение жизни и эволюция биосфер . 45 (1): 139–148. Бибкод : 2015ОЛЕВ...45..139М . дои : 10.1007/s11084-015-9411-z . ISSN 1573-0875 . ПМЦ 4457167 . ПМИД 25716918 .
- ^ Давила, Мария Х.; Майер, Кристиан (2022). «Мембранная структура, полученная в ходе экспериментального процесса эволюции» . Жизнь . 12 (2): 145. Бибкод : 2022Life...12..145D . дои : 10.3390/life12020145 . ISSN 2075-1729 . ПМЦ 8875328 . ПМИД 35207433 .
- ^ Майер, Кристиан (2022). «Спонтанное формирование функциональных структур в беспорядочной среде» . Жизнь . 12 (5): 720. Бибкод : 2022Жизнь...12..720М . дои : 10.3390/life12050720 . ISSN 2075-1729 . ПМЦ 9148140 . ПМИД 35629387 .
- ^ Давила, Мария Х.; Майер, Кристиан (2023). «Структурные явления в мембране пузырька, полученные в ходе эволюционного эксперимента: исследование, основанное на моделировании МД» . Жизнь . 13 (8): 1735. Бибкод : 2023Life...13.1735D . дои : 10.3390/life13081735 . ISSN 2075-1729 . ПМЦ 10455627 . ПМИД 37629592 .
- ^ Майер, Кристиан (18 января 2020 г.). «Жизнь в контексте порядка и сложности» . Жизнь . 10 (1): 5. Бибкод : 2020Жизнь...10....5М . дои : 10.3390/life10010005 . ISSN 2075-1729 . ПМЦ 7175320 . ПМИД 31963637 .
- ^ Шаров, Алексей А.; Гордон, Ричард (2018). «Жизнь до Земли» . Обитаемость Вселенной до Земли: Жизнь до Земли . Астробиология, изучающая жизнь на Земле и за ее пределами. Академическая пресса . стр. 265–296. дои : 10.1016/B978-0-12-811940-2.00011-3 . ISBN 978-0-12-811940-2 . S2CID 117048600 . Архивировано из оригинала 30 апреля 2022 года . Проверено 30 апреля 2022 г.
- ^ Ледиман, Дж.; Ламберт, Дж.; Вайснер, КБ (2013). «Что такое сложная система?». Европейский журнал философии науки . 3 : 33–67. дои : 10.1007/s13194-012-0056-8 . S2CID 18787276 .
- ^ Эспозито, М.; Линденберг, Катя ; Ван ден Брук, К. (2010). «Производство энтропии как взаимосвязь между системой и резервуаром». Новый журнал физики . 12 (1): 013013. arXiv : 0908.1125 . Бибкод : 2010NJPh...12a3013E . дои : 10.1088/1367-2630/12/1/013013 . S2CID 26657293 .
- ^ Бар-Нун, А.; Бар-Нун, Н.; Бауэр, С.Х.; Саган, Карл (24 апреля 1970 г.). «Ударный синтез аминокислот в моделируемых примитивных средах». Наука . 168 (3930): 470–473. Бибкод : 1970Sci...168..470B . дои : 10.1126/science.168.3930.470 . ПМИД 5436082 . S2CID 42467812 .
- ^ Анбар, Майкл (27 сентября 1968 г.). «Кавитация при воздействии жидкой воды на воду: геохимические последствия». Наука . 161 (3848): 1343–1344. Бибкод : 1968Sci...161.1343A . дои : 10.1126/science.161.3848.1343 . ПМИД 17831346 .
- ^ Дхармаратне, Лина; Гризер, Франц (7 января 2016 г.). «Образование аминокислот при сонолизе водных растворов, содержащих уксусную кислоту, метан или углекислый газ, в присутствии газообразного азота». Журнал физической химии А. 120 (2): 191–199. Бибкод : 2016JPCA..120..191D . дои : 10.1021/acs.jpca.5b11858 . ПМИД 26695890 .
- ^ Патехебике, Йерсен; Чжао, Зе-Рун; Ван, Су; Сюй, Хао-Син; Чен, Цянь-Цянь; Ван, Сяо (2021). «Кавитация как возможная движущая сила пребиотического образования пуриновых нуклеозидов N9» . Отчеты о клетках Физические науки . 2 (3): 100375. Бибкод : 2021CRPS....200375P . дои : 10.1016/j.xcrp.2021.100375 . S2CID 233662126 .
- ^ Кальсон, Натан-Хаим; Фурман, Дэвид; Зейри, Иегуда (11 сентября 2017 г.). «Кавитационный синтез биогенных молекул на первичной Земле» . Центральная научная служба ACS . 3 (9): 1041–1049. дои : 10.1021/accentsci.7b00325 . ПМК 5620973 . ПМИД 28979946 . S2CID 21409351 .
- ^ Мюллер, Энтони WJ (1995). «Были ли первые организмы тепловыми двигателями? Новая модель биогенеза и ранняя эволюция преобразования биологической энергии» . Прогресс биофизики и молекулярной биологии . 63 (2): 193–231. дои : 10.1016/0079-6107(95)00004-7 . ПМИД 7542789 .
- ^ Мюллер, Энтони У.Дж.; Шульце-Макух, Дирк (2006). «Тепловая энергия и происхождение жизни». Происхождение жизни и эволюция биосфер . 36 (2): 77–189. Бибкод : 2006ОЛЕВ...36..177М . дои : 10.1007/s11084-005-9003-4 . ПМИД 16642267 . S2CID 22179552 .
- ^ Юнге, Вольфганг; Нельсон, Натан (2 июня 2015 г.). «АТФ-синтаза» . Ежегодный обзор биохимии . 84 (1): 631–657. doi : 10.1146/annurev-biochem-060614-034124 . ПМИД 25839341 .
- ^ Jump up to: а б с Дамер, Брюс; Димер, Дэвид (1 апреля 2020 г.). «Гипотеза происхождения жизни из горячих источников» . Астробиология . 20 (4): 429–452. Бибкод : 2020AsBio..20..429D . дои : 10.1089/ast.2019.2045 . ПМЦ 7133448 . ПМИД 31841362 .
- ^ Беннер, ЮАР; Белл, Э.А.; Бионди, Э.; Брассер, Р.; Карелл, Т.; Ким, Х.-Дж.; Мойжис, С.Дж.; Омран, А.; Пасек, Массачусетс; Трейл, Д. (2020). «Когда жизнь, вероятно, возникла на Земле в результате процесса, основанного на РНК?» . ХимСистемыХим . 2 (2). arXiv : 1908.11327 . дои : 10.1002/syst.201900035 .
- ^ * Копли, Шелли Д.; Смит, Эрик; Моровиц, Гарольд Дж. (декабрь 2007 г.). «Происхождение мира РНК: коэволюция генов и метаболизма» (PDF) . Биоорганическая химия . 35 (6): 430–443. дои : 10.1016/j.bioorg.2007.08.001 . ПМИД 17897696 . Архивировано (PDF) из оригинала 5 сентября 2013 года . Проверено 8 июня 2015 г.
Предположение о том, что жизнь на Земле возникла из мира РНК, широко распространено.
- Оргел, Лесли Э. (апрель 2003 г.). «Некоторые следствия гипотезы мира РНК». Происхождение жизни и эволюция биосфер . 33 (2): 211–218. Бибкод : 2003OLEB...33..211O . дои : 10.1023/A:1024616317965 . ПМИД 12967268 . S2CID 32779859 .
Теперь кажется весьма вероятным, что нашему знакомому миру ДНК/РНК/белков предшествовал мир РНК...
- Робертсон и Джойс 2012 : «В настоящее время имеются убедительные доказательства того, что мир РНК действительно существовал до того, как жизнь на основе ДНК и белков».
- Неве, Ким и Беннер, 2013 : «[Существование мира РНК] сегодня пользуется широкой поддержкой в обществе».
- Оргел, Лесли Э. (апрель 2003 г.). «Некоторые следствия гипотезы мира РНК». Происхождение жизни и эволюция биосфер . 33 (2): 211–218. Бибкод : 2003OLEB...33..211O . дои : 10.1023/A:1024616317965 . ПМИД 12967268 . S2CID 32779859 .
- ^ Jump up to: а б с Робертсон, Майкл П.; Джойс, Джеральд Ф. (май 2012 г.). «Происхождение мира РНК» . Перспективы Колд-Спринг-Харбор в биологии . 4 (5): а003608. doi : 10.1101/cshperspect.a003608 . ПМК 3331698 . ПМИД 20739415 .
- ^ Jump up to: а б с Чех, Томас Р. (июль 2012 г.). «Миры РНК в контексте» . Перспективы Колд-Спринг-Харбор в биологии . 4 (7): а006742. doi : 10.1101/cshperspect.a006742 . ПМЦ 3385955 . ПМИД 21441585 .
- ^ Пирс, Бен К.Д.; Пудриц, Ральф Э.; Семенов Дмитрий А.; Хеннинг, Томас К. (24 октября 2017 г.). «Происхождение мира РНК: судьба нуклеиновых оснований в теплых прудах» . ПНАС . 114 (43): 11327–11332. arXiv : 1710.00434 . Бибкод : 2017PNAS..11411327P . дои : 10.1073/pnas.1710339114 . ПМК 5664528 . ПМИД 28973920 .
- ^ Ярус, Майкл (апрель 2011 г.). «Пройдя через мир РНК: первоначальный дарвиновский предок» . Перспективы Колд-Спринг-Харбор в биологии . 3 (4): а003590. doi : 10.1101/cshperspect.a003590 . ПМК 3062219 . ПМИД 20719875 .
- ^ Foot & Foot 2004 , с. 29
- ^ Фокс, Джордж.Э. (9 июня 2010 г.). «Происхождение и эволюция рибосомы» . Перспективы Колд-Спринг-Харбор в биологии . 2 (9(а003483)): а003483. doi : 10.1101/cshperspect.a003483 . ПМЦ 2926754 . ПМИД 20534711 .
- ^ Невё, Марк; Ким, Хё Чжун; Беннер, Стивен А. (22 апреля 2013 г.). «Гипотеза о мире сильных РНК: пятьдесят лет». Астробиология . 13 (4): 391–403. Бибкод : 2013AsBio..13..391N . дои : 10.1089/ast.2012.0868 . ПМИД 23551238 .
- ^ Гилберт, Уолтер (20 февраля 1986 г.). «Происхождение жизни: мир РНК» . Природа . 319 (6055): 618. Бибкод : 1986Natur.319..618G . дои : 10.1038/319618a0 . S2CID 8026658 .
- ^ Оргел, Лесли Э. (октябрь 1994 г.). «Происхождение жизни на Земле». Научный американец . 271 (4): 76–83. Бибкод : 1994SciAm.271d..76O . doi : 10.1038/scientificamerican1094-76 . ПМИД 7524147 .
- ^ Линкольн, Трейси А.; Джойс, Джеральд Ф. (27 февраля 2009 г.). «Самоподдерживающаяся репликация фермента РНК» . Наука . 323 (5918): 1229–1232. Бибкод : 2009Sci...323.1229L . дои : 10.1126/science.1167856 . ПМК 2652413 . ПМИД 19131595 .
- ^ Джойс, Джеральд Ф. (2009). «Эволюция в мире РНК» . Перспективы Колд-Спринг-Харбор в биологии . 74 (Эволюция: Молекулярный ландшафт): 17–23. дои : 10.1101/sqb.2009.74.004 . ПМЦ 2891321 . ПМИД 19667013 .
- ^ Шостак, Джек В. (5 февраля 2015 г.). «Происхождение функций биологических нуклеиновых кислот, белков и мембран» . Чеви Чейз, Мэриленд: Медицинский институт Говарда Хьюза . Архивировано из оригинала 14 июля 2015 года . Проверено 16 июня 2015 г.
- ^ Бернштейн, Харрис; Байерли, Генри К.; Хопф, Фредерик А.; и др. (июнь 1983 г.). «Дарвиновская динамика». Ежеквартальный обзор биологии . 58 (2): 185–207. дои : 10.1086/413216 . JSTOR 2828805 . S2CID 83956410 .
- ^ Мишо 1999
- ^ Паласек, Стэн (23 мая 2013 г.). «Изначальная репликация РНК и применение в технологии ПЦР». arXiv : 1305.5581v1 [ q-bio.BM ].
- ^ Власов Александр Васильевич; Казаков Сергей А.; Джонстон, Брайан Х.; и др. (август 2005 г.). «Мир РНК на льду: новый сценарий появления информации о РНК». Журнал молекулярной эволюции . 61 (2): 264–273. Бибкод : 2005JMolE..61..264В . дои : 10.1007/s00239-004-0362-7 . ПМИД 16044244 . S2CID 21096886 .
- ^ Нусинов, Марк Д.; Отрощенко Владимир А.; Сантоли, Сальваторе (1997). «Зарождение неклеточной фазы жизни на мелкозернистых глинистых частицах раннего реголита Земли». БиоСистемы . 42 (2–3): 111–118. Бибкод : 1997BiSys..42..111N . дои : 10.1016/S0303-2647(96)01699-1 . ПМИД 9184757 .
- ^ Кюнляйн, Александра; Ланцмих, Саймон А.; Брун, Дитер (2 марта 2021 г.). «Последовательности тРНК могут собираться в репликатор» . электронная жизнь . 10 . doi : 10.7554/eLife.63431 . ПМЦ 7924937 . ПМИД 33648631 .
- ^ Ноллер, Гарри Ф. (апрель 2012 г.). «Эволюция синтеза белков из мира РНК» . Перспективы Колд-Спринг-Харбор в биологии . 4 (4): а003681. doi : 10.1101/cshperspect.a003681 . ПМЦ 3312679 . ПМИД 20610545 .
- ^ Кунин, Евгений В. (31 мая 2007 г.). «Космологическая модель вечной инфляции и переход от случайности к биологической эволюции в истории жизни» . Биология Директ . 2:15 . дои : 10.1186/1745-6150-2-15 . ЧВК 1892545 . ПМИД 17540027 .
- ^ Jump up to: а б Тамура, К.; Александр, RW (1 мая 2004 г.). «Синтез пептидов путем эволюции» . Клеточные и молекулярные науки о жизни . 61 (11): 1317–1330. дои : 10.1007/s00018-004-3449-9 . ISSN 1420-9071 . ПМЦ 11138682 . ПМИД 15170510 .
- ^ Тамура, Кодзи; Шиммель, Пол (22 июля 2003 г.). «Синтез пептидов с помощью матричной РНК-гида и аминоацилфосфатных адаптеров» . Труды Национальной академии наук . 100 (15): 8666–8669. Бибкод : 2003PNAS..100.8666T . дои : 10.1073/pnas.1432909100 . ISSN 0027-8424 . ПМК 166369 . ПМИД 12857953 .
- ^ Jump up to: а б Прессман, Эйб Д.; Лю, Цивэй; Янзен, Эван; Бланко, Селия; Мюллер, Ульрих Ф.; Джойс, Джеральд Ф.; Паскаль, Роберт; Чен, Ирен А. (17 апреля 2019 г.). «Картирование систематического ландшафта пригодности рибозимов показывает разочарованную эволюционную сеть самоаминоацилирующей РНК» . Журнал Американского химического общества . 141 (15): 6213–6223. дои : 10.1021/jacs.8b13298 . ISSN 0002-7863 . ПМК 6548421 . ПМИД 30912655 .
- ^ Тьяги, Санджай; Поннамперума, Сирил (1 мая 1990 г.). «Неслучайность в синтезе пребиотических пептидов» . Журнал молекулярной эволюции . 30 (5): 391–399. Бибкод : 1990JMolE..30..391T . дои : 10.1007/BF02101111 . ISSN 1432-1432 . ПМИД 2111852 .
- ^ Киф, Энтони Д.; Шостак, Джек В. (2001). «Функциональные белки из библиотеки случайных последовательностей» . Природа . 410 (6829): 715–718. Бибкод : 2001Natur.410..715K . дои : 10.1038/35070613 . ПМЦ 4476321 . ПМИД 11287961 .
- ^ Тонг, Шер Линг; Ли, Кун-Хва; Зеилиг, Буркхард (июнь 2021 г.). «Белки de novo из случайных последовательностей в результате эволюции in vitro» . Современное мнение в области структурной биологии . 68 : 129–134. дои : 10.1016/j.sbi.2020.12.014 . ПМЦ 8222087 . ПМИД 33517151 .
- ^ Бун, Дэвид Р.; Кастенхольц, Ричард В.; Гаррити, Джордж М., ред. (2001). Археи . и глубоко ветвящиеся и бактерии фототрофные Руководство Берджи по систематической бактериологии. Спрингер . ISBN 978-0-387-21609-6 . Архивировано из оригинала 25 декабря 2014 года. [ нужна страница ]
- ^ Вёзе, ЧР; Фокс, GE (1977). «Филогенетическая структура прокариотического домена: первичные царства» . ПНАС . 7 (11): 5088–5090. Бибкод : 1977PNAS...74.5088W . дои : 10.1073/pnas.74.11.5088 . ПМК 432104 . ПМИД 270744 .
- ^ Валас, Р.Э.; Борн, ЧП (2011). «Происхождение производного суперцарства: как грамположительная бактерия пересекла пустыню и стала археем» . Биология Директ . 6:16 . дои : 10.1186/1745-6150-6-16 . ПМК 3056875 . ПМИД 21356104 .
- ^ Кавальер-Смит, Томас (2006). «Укоренение древа жизни посредством анализа переходов» . Биология Директ . 1:19 . дои : 10.1186/1745-6150-1-19 . ПМК 1586193 . ПМИД 16834776 .
- ^ «В молодости нравилось погорячее» . Природа . 535 (7613): 468. 2016. doi : 10.1038/535468b . S2CID 49905802 .
- ^ Гогартен, Иоганн Петер; Димер, Дэвид (25 ноября 2016 г.). «Является ли LUCA термофильным предшественником?» . Природная микробиология . 1 (12): 16229. doi : 10.1038/nmicrobiol.2016.229 . ПМИД 27886195 . S2CID 205428194 . Архивировано из оригинала 3 апреля 2020 года . Проверено 21 сентября 2022 г.
- ^ Кэтчпол, Райан; Фортерре, Патрик (2019). «Эволюция обратной гиразы предполагает наличие негипертермофильного последнего универсального общего предка» . Молекулярная биология и эволюция . 36 (12): 2737–2747. дои : 10.1093/molbev/msz180 . ПМК 6878951 . ПМИД 31504731 . Архивировано из оригинала 27 января 2023 года . Проверено 18 сентября 2022 г.
- ^ Беркемер, Сара Дж.; МакГлинн, Шон Э (8 августа 2020 г.). «Новый анализ разделения доменов архей и бактерий: переменное филогенетическое расстояние и темп ранней эволюции» . Молекулярная биология и эволюция . 37 (8): 2332–2340. дои : 10.1093/molbev/msaa089 . ПМЦ 7403611 . ПМИД 32316034 . Архивировано из оригинала 27 января 2023 года . Проверено 21 сентября 2022 г.
- ^ Хоффманн, Джеффри В. (25 июня 1974 г.). «О происхождении генетического кода и стабильности аппарата трансляции». Журнал молекулярной биологии . 86 (2): 349–362. дои : 10.1016/0022-2836(74)90024-2 . ПМИД 4414916 .
- ^ Оргел, Лесли Э. (апрель 1963 г.). «Поддержание точности синтеза белка и его значение для старения» . ПНАС . 49 (4): 517–521. Бибкод : 1963PNAS...49..517O . дои : 10.1073/pnas.49.4.517 . ПМЦ 299893 . ПМИД 13940312 .
- ^ Хоффманн, Джеффри В. (октябрь 1975 г.). «Стохастическая теория происхождения генетического кода». Ежегодный обзор физической химии . 26 : 123–144. Бибкод : 1975ARPC...26..123H . дои : 10.1146/annurev.pc.26.100175.001011 .
- ^ Харрисон, Стюарт А.; Палмейра, Ракель Нуньес; Халперн, Аарон; Лейн, Ник (1 ноября 2022 г.). «Биофизическая основа возникновения генетического кода в протоклетках» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1863 (8): 148597. doi : 10.1016/j.bbabio.2022.148597 . PMID 35868450 . S2CID 250707510 .
- ^ Харрисон, Стюарт А.; Лейн, Ник (12 декабря 2018 г.). «Жизнь как руководство к синтезу пребиотических нуклеотидов» . Природные коммуникации . 9 (1): 5176. Бибкод : 2018NatCo...9.5176H . дои : 10.1038/s41467-018-07220-y . ПМК 6289992 . ПМИД 30538225 .
- ^ Кантин, Марджори Д.; Фурнье, Грегори П. (1 марта 2018 г.). «Адаптация окружающей среды от зарождения жизни до последнего вселенского общего предка» . Происхождение жизни и эволюция биосфер . 48 (1): 35–54. Бибкод : 2018OLEB...48...35C . дои : 10.1007/s11084-017-9542-5 . hdl : 1721.1/114219 . ПМИД 28685374 . S2CID 254888920 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Мэт, Вай-Кин (1 мая 2008 г.). «Геномика LUCA» (PDF) . Границы бионауки . 13 (14): 5605–5613. дои : 10.2741/3103 . ПМИД 18508609 . Архивировано (PDF) из оригинала 16 июня 2022 года . Проверено 8 декабря 2023 г.
- ^ Брейзер, доктор медицины (2012). Тайные камеры: внутренняя история клеток и сложной жизни . Издательство Оксфордского университета. п. 298.
- ^ Уорд, Питер и Киршвинк, Джо , цит., стр. 42
- ^ Jump up to: а б Колин-Гарсия, М.; Эредиа, А.; Кордеро, Г.; и др. (2016). «Гидротермальные источники и пребиотическая химия: обзор» . Бюллетень Мексиканского геологического общества . 68 (3): 599–620. дои : 10.18268/BSGM2016v68n3a13 . Архивировано из оригинала 18 августа 2017 года.
- ^ Ширбер, Майкл (24 июня 2014 г.). «Гидротермальные источники могут объяснить появление химических предшественников жизни» . Астробиология НАСА: Жизнь во Вселенной . НАСА. Архивировано из оригинала 29 ноября 2014 года . Проверено 19 июня 2015 г.
- ^ Jump up to: а б Мартин, Уильям ; Рассел, Майкл Дж. (29 января 2003 г.). «О происхождении клеток: гипотеза эволюционных переходов от абиотической геохимии к хемоавтотрофным прокариотам и от прокариотов к ядросодержащим клеткам» . Философские труды Королевского общества Б. 358 (1429): 59–83, обсуждение 83–85. дои : 10.1098/rstb.2002.1183 . ПМК 1693102 . ПМИД 12594918 .
- ^ Лейн 2009
- ^ Ашер, Оли (27 апреля 2015 г.). «Химия горячих источников морского дна может объяснить возникновение жизни» (Пресс-релиз). Университетский колледж Лондона . Архивировано из оригинала 20 июня 2015 года . Проверено 19 июня 2015 г.
- ^ Ролдан, Альберто; Холлингсворт, Натан; Роффи, Анна; и др. (май 2015 г.). «Биологическая конверсия CO2 с помощью катализаторов на основе сульфида железа в устойчивых условиях» . Химические коммуникации . 51 (35): 7501–7504. дои : 10.1039/C5CC02078F . ПМИД 25835242 . Архивировано из оригинала 20 июня 2015 года . Проверено 19 июня 2015 г.
- ^ Баросс, Дж.А.; Хоффман, SE (1985). «Подводные гидротермальные источники и связанная с ними градиентная среда как места зарождения и эволюции жизни». Происхождение жизни и эволюция биосфер . 15 (4): 327–345. Бибкод : 1985OrLi...15..327B . дои : 10.1007/bf01808177 . S2CID 4613918 .
- ^ Рассел, MJ; Холл, Эй Джей (1997). «Появление жизни из пузырьков моносульфида железа на подводном гидротермальном окислительно-восстановительном фронте и фронте pH». Журнал Геологического общества . 154 (3): 377–402. Бибкод : 1997JGSoc.154..377R . дои : 10.1144/gsjgs.154.3.0377 . ПМИД 11541234 . S2CID 24792282 .
- ^ Поправка, JP; ЛаРоу, Делавэр; Макколлом, ТМ; Шок, Э.Л. (2013). «Энергетика органического синтеза внутри и вне клетки» . Философские труды Королевского общества Б. 368 (1622): 20120255. doi : 10.1098/rstb.2012.0255 . ПМЦ 3685458 . ПМИД 23754809 .
- ^ Шок, EL; Бойд, ES (2015). «Геомикробиология и микробная геохимия: принципы геобиохимии». Элементы . 11 : 389–394. дои : 10.2113/gselements.11.6.395 .
- ^ Мартин, В.; Рассел, MJ (2007). «О происхождении биохимии на щелочно-гидротермальном источнике» . Философские труды Королевского общества Б. 362 (1486): 1887–1925. дои : 10.1098/rstb.2006.1881 . ПМЦ 2442388 . ПМИД 17255002 .
- ^ Лейн, Ник; Мартин, Уильям Ф. (21 декабря 2012 г.). «Происхождение мембранной биоэнергетики» . Клетка . 151 (7): 1406–1416. дои : 10.1016/j.cell.2012.11.050 . ПМИД 23260134 . S2CID 15028935 .
- ^ Бааске, Филипп; Вайнерт, Франц М.; Дур, Стефан; Лемке, Коно Х.; Рассел, Майкл Дж.; Браун, Дитер (29 мая 2007 г.). «Чрезвычайное накопление нуклеотидов в смоделированных гидротермальных поровых системах» . Труды Национальной академии наук . 104 (22): 9346–9351. дои : 10.1073/pnas.0609592104 . ПМК 1890497 . ПМИД 17494767 .
- ^ Нуньес Палмейра, Ракель; Колнаги, Марко; Харрисон, Стюарт А.; Помянковский, Эндрю; Лейн, Ник (9 ноября 2022 г.). «Границы метаболической наследственности в протоклетках» . Труды Королевского общества B: Биологические науки . 289 (1986). дои : 10.1098/rspb.2022.1469 . ПМЦ 9653231 . ПМИД 36350219 .
- ^ Вёзе, Карл Р. (1987). «Бактериальная эволюция» . Микробиологические обзоры . 51.2 (1987) (2): 221–271. дои : 10.1128/мр.51.2.221-271.1987 . ПМК 373105 . ПМИД 2439888 . S2CID 734579 .
- ^ Рассел, Майкл Дж.; Холл, Аллан Дж. (2006), «Зарождение и ранняя эволюция жизни» , Эволюция атмосферы, гидросферы и биосферы ранней Земли - ограничения, обусловленные рудными месторождениями , Геологическое общество Америки, номер документа : 10.1130/2006.1198(01) , ISBN 9780813711980 , заархивировано из оригинала 31 января 2024 года , получено 30 ноября 2023 года.
- ^ Буссау, Бастьен; Бланкар, Самуэль; Нексулея, Анамария; Лартильо, Николя; Гуи, Маноло (декабрь 2008 г.). «Параллельные адаптации к высоким температурам в архейском эоне» . Природа . 456 (7224): 942–945. Бибкод : 2008Natur.456..942B . дои : 10.1038/nature07393 . ПМИД 19037246 . S2CID 4348746 . Архивировано из оригинала 16 декабря 2023 года . Проверено 8 декабря 2023 г.
- ^ Jump up to: а б Галтье, Николя; Турасс, Николя; Гуи, Маноло (8 января 1999 г.). «Негипертермофильный общий предок существующих форм жизни» . Наука . 283 (5399): 220–221. дои : 10.1126/science.283.5399.220 . ПМИД 9880254 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Келли, Дебора С.; Карсон, Джеффри А.; Блэкман, Донна К.; Фрю-Грин, Гретхен Л.; Баттерфилд, Дэвид А.; Лилли, Марвин Д.; Олсон, Эрик Дж.; Шренк, Мэтью О.; Роу, Кевин К.; Лебон, Джефф Т.; Ривизиньо, Пит (июль 2001 г.). «Внеосевое гидротермальное поле возле Срединно-Атлантического хребта на 30 ° с.ш.» . Природа . 412 (6843): 145–149. Бибкод : 2001Natur.412..145K . дои : 10.1038/35084000 . ПМИД 11449263 . S2CID 4407013 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Jump up to: а б Эренфройнд, П.; Ирвин, В.; Беккер, Л.; Бланк, Дж.; Брукато-младший; и др. (август 2002 г.). «Астрофизические и астрохимические открытия о происхождении жизни» . Отчеты о прогрессе в физике . 65 (10): 1427. Бибкод : 2002РПФ...65.1427Е . дои : 10.1088/0034-4885/65/10/202 . S2CID 250904448 .
- ^ Чиба, CF; Чиба, CF; Хэнд, КП (2006). «Кометы и пребиотические органические молекулы на ранней Земле». Кометы и происхождение и эволюция жизни . Достижения астробиологии и биогеофизики. Шпрингер Берлин Гейдельберг. стр. 169–206. дои : 10.1007/3-540-33088-7_6 . ISBN 978-3-540-33086-8 . Архивировано из оригинала 31 января 2024 года . Проверено 30 ноября 2023 г.
- ^ Чаттерджи, Санкар (2023), Чаттерджи, Санкар (редактор), «Колыбель жизни» , От звездной пыли до первых клеток: происхождение и эволюция ранней жизни , Чам: Springer International Publishing, стр. 43–66, doi : 10.1007/978-3-031-23397-5_6 , ISBN 978-3-031-23397-5 , заархивировано из оригинала 31 января 2024 года , получено 5 декабря 2023 года.
- ^ Димер, Дэвид В. (7 февраля 2019 г.). «Перспективы жизни на других планетах». Сборка жизни . Издательство Оксфордского университета . дои : 10.1093/oso/9780190646387.003.0017 . ISBN 978-0-19-064638-7 . Архивировано из оригинала 31 января 2024 года . Проверено 6 декабря 2023 г.
- ^ Пирс, Бен К.Д.; Пудриц, Ральф Э.; Семенов Дмитрий А.; Хеннинг, Томас К. (2 октября 2017 г.). «Происхождение мира РНК: судьба нуклеиновых оснований в теплых прудах» . Труды Национальной академии наук . 114 (43): 11327–11332. arXiv : 1710.00434 . Бибкод : 2017PNAS..11411327P . дои : 10.1073/pnas.1710339114 . ПМК 5664528 . ПМИД 28973920 .
- ^ Jump up to: а б с Димер, Дэвид (10 февраля 2021 г.). «Где началась жизнь? Тестирование идей в условиях аналогов пребиотиков» . Жизнь . 11 (2): 134. Бибкод : 2021Life...11..134D . дои : 10.3390/life11020134 . ПМЦ 7916457 . ПМИД 33578711 .
- ^ Джордан, Шон Ф.; Рамму, Ханади; Желудев Иван Н.; Хартли, Эндрю М.; Марешаль, Амандин; Лейн, Ник (2019). «Стимулирование самосборки протоклеток из смешанных амфифилов при зарождении жизни» . Экология и эволюция природы . 3 (12): 1705–1714. Бибкод : 2019NatEE...3.1705J . дои : 10.1038/s41559-019-1015-y . ПМИД 31686020 .
- ^ Коренага, июнь (ноябрь 2021 г.). «Была ли земля на ранней Земле?» . Жизнь . 11 (11): 1142. Бибкод : 2021Жизнь...11.1142К . дои : 10.3390/life11111142 . ПМЦ 8623345 . ПМИД 34833018 .
- ^ Пауэр, Мэтью В.; Жерланд, Беатрис; Сазерленд, Джон Д. (май 2009 г.). «Синтез активированных пиримидинрибонуклеотидов в пребиотически приемлемых условиях» . Природа . 459 (7244): 239–242. Бибкод : 2009Natur.459..239P . дои : 10.1038/nature08013 . ПМИД 19444213 . S2CID 4412117 . Архивировано из оригинала 12 ноября 2023 года . Проверено 8 декабря 2023 г.
- ^ Занле, Кевин; Арндт, Ник; Кокелл, Чарльз; Холлидей, Алекс; Нисбет, Юан; Селсис, Франк; Сон, Норман Х. (1 марта 2007 г.). «Появление обитаемой планеты» . Обзоры космической науки . 129 (1): 35–78. Бибкод : 2007ССРв..129...35З . дои : 10.1007/s11214-007-9225-z . S2CID 12006144 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Вёзе, ЧР (июнь 1987 г.). «Бактериальная эволюция» . Микробиологические обзоры . 51 (2): 221–271. дои : 10.1128/мр.51.2.221-271.1987 . ПМК 373105 . ПМИД 2439888 .
- ^ Jump up to: а б с Мулкиджанян Армен Ю.; Бычков Андрей Ю.; Диброва Дарья Викторовна; Гальперин, Михаил Юрьевич; Кунин, Евгений В. (3 апреля 2012 г.). «Происхождение первых клеток на земных бескислородных геотермальных полях» . Труды Национальной академии наук . 109 (14): Е821-30. Бибкод : 2012PNAS..109E.821M . дои : 10.1073/pnas.1117774109 . ПМЦ 3325685 . ПМИД 22331915 .
- ^ Чандру, Кухан; Гуттенберг, Николас; Гири, Чайтанья; и др. (31 мая 2018 г.). «Простой пребиотический синтез динамических комбинаторных полиэфирных библиотек большого разнообразия» . Химия связи . 1 (1). дои : 10.1038/s42004-018-0031-1 .
- ^ Форсайт, Джей Г.; Ю, Шэн-Шэн; Мамаджанов, Ирена; и др. (17 августа 2015 г.). «Опосредованное эфиром образование амидной связи, вызванное циклами влажно-сухой: возможный путь к полипептидам на пребиотической Земле» . Angewandte Chemie International Edition на английском языке . 54 (34): 9871–9875. дои : 10.1002/anie.201503792 . ПМЦ 4678426 . ПМИД 26201989 .
- ^ Патель, Бхавеш Х.; Персиваль, Клаудия; Ритсон, Дугал Дж.; Даффи, Колм. Д.; Сазерленд, Джон Д. (16 марта 2015 г.). «Общее происхождение предшественников РНК, белков и липидов в цианосульфидном протометаболизме» . Природная химия . 7 (4): 301–307. Бибкод : 2015НатЧ...7..301П . дои : 10.1038/nchem.2202 . ПМЦ 4568310 . ПМИД 25803468 .
- ^ Кэтлинг, Дэвид К.; Занле, Кевин Дж. (28 февраля 2020 г.). «Архейская атмосфера» . Достижения науки . 6 (9): eaax1420. Бибкод : 2020SciA....6.1420C . дои : 10.1126/sciadv.aax1420 . ПМК 7043912 . ПМИД 32133393 .
- ^ Миллер, Стэнли Л.; Ласкано, Антонио (декабрь 1995 г.). «Происхождение жизни? Происходило ли оно при высоких температурах?» . Журнал молекулярной эволюции . 41 (6): 689–692. Бибкод : 1995JMolE..41..689M . дои : 10.1007/bf00173146 . hdl : 2060/19980211388 . ПМИД 11539558 . S2CID 25141419 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Фортерре, Патрик; Бержера, Агнес; Лопекс-Гарсия, Purificacion (май 1996 г.). «Уникальная топология ДНК и ДНК-топоизомеразы гипертермофильных архей» . Обзоры микробиологии FEMS . 18 (2–3): 237–248. дои : 10.1111/j.1574-6976.1996.tb00240.x . ПМИД 8639331 . S2CID 6001830 .
- ^ Брошье-Армане, Селин; Фортерре, Патрик (май 2006 г.). «Широко распространенное распространение архейной обратной гиразы в термофильных бактериях предполагает сложную историю вертикального наследования и латерального переноса генов» . Архея . 2 (2): 83–93. дои : 10.1155/2006/582916 . ПМК 2686386 . ПМИД 17350929 .
- ^ Фойлнер, Георг (июнь 2012 г.). «Проблема слабого молодого Солнца» . Обзоры геофизики . 50 (2). arXiv : 1204.4449 . Бибкод : 2012RvGeo..50.2006F . дои : 10.1029/2011RG000375 . S2CID 119248267 . Архивировано из оригинала 8 декабря 2023 года . Проверено 8 декабря 2023 г.
- ^ Бада, Дж.Л.; Бигэм, К.; Миллер, С.Л. (15 февраля 1994 г.). «Воздействие таяния замерзших океанов на раннюю Землю: последствия для происхождения жизни» . Труды Национальной академии наук . 91 (4): 1248–1250. Бибкод : 1994PNAS...91.1248B . дои : 10.1073/pnas.91.4.1248 . ПМЦ 43134 . ПМИД 11539550 .
- ^ Моннар, Пьер-Ален; Апель, Чарльз Л.; Канавариоти, Анастасия; Димер, Дэвид В. (июнь 2002 г.). «Влияние ионных неорганических растворов на процессы самосборки и полимеризации, связанные с ранними формами жизни: последствия для пребиотической водной среды» . Астробиология . 2 (2): 139–152. Бибкод : 2002AsBio...2..139M . дои : 10.1089/15311070260192237 . ПМИД 12469365 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Аттуотер, Джеймс; Вокнер, Аниела; Холлигер, Филипп (декабрь 2013 г.). «Эволюция рибозимной активности РНК-полимеразы во льду» . Природная химия . 5 (12): 1011–1018. Бибкод : 2013НатЧ...5.1011А . дои : 10.1038/nchem.1781 . ПМК 3920166 . ПМИД 24256864 .
- ^ Леви, Мэтью; Миллер, Стэнли Л.; Бринтон, Карен; Бада, Джеффри Л. (1 июня 2000 г.). «Пребиотический синтез аденина и аминокислот в условиях Европы» . Икар . 145 (2): 609–613. Бибкод : 2000Icar..145..609L . дои : 10.1006/icar.2000.6365 . ПМИД 11543508 .
- ^ Моултон, Винсент; Гарднер, Пол П.; Пойнтон, Роберт Ф.; Кример, Лоуренс К.; Джеймсон, Джеффри Б.; Пенни, Дэвид (1 октября 2000 г.). «Складывание РНК выступает против возникновения жизни с горячим стартом» . Журнал молекулярной эволюции . 51 (4): 416–421. Бибкод : 2000JMolE..51..416M . дои : 10.1007/s002390010104 . ПМИД 11040293 . S2CID 20787323 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Земора, Джорджета; Вальдсих, Кристина (ноябрь 2010 г.). «Сворачивание РНК в живых клетках» . Биология РНК . 7 (6): 634–641. дои : 10.4161/rna.7.6.13554 . ПМК 3073324 . ПМИД 21045541 . Архивировано из оригинала 31 января 2024 года . Проверено 8 декабря 2023 г.
- ^ Шрайбер, Ульрих; Локер-Грютен, Оливер; Майер, Кристиан (2012). «Гипотеза: происхождение жизни в глубоких тектонических разломах» . Происхождение жизни и эволюция биосфер . 42 (1): 47–54. Бибкод : 2012OLEB...42...47S . дои : 10.1007/s11084-012-9267-4 . ISSN 0169-6149 . ПМИД 22373604 .
- ^ Шрайбер, Ульрих; Майер, Кристиан; Шмитц, Оливер Дж.; Розендаль, Пиа; Бронха, Амела; Греле, Маркус; Кепплер, Фрэнк; Малдер, Инес; Саттлер, Тобиас; Шёлер, Хайнц Ф. (14 июня 2017 г.). Стюкен, Ева Элизабет (ред.). «Органические соединения во флюидных включениях архейского кварца — аналоги пребиотической химии на ранней Земле» . ПЛОС ОДИН . 12 (6): e0177570. Бибкод : 2017PLoSO..1277570S . дои : 10.1371/journal.pone.0177570 . ISSN 1932-6203 . ПМК 5470662 . ПМИД 28614348 .
- ^ Гроссманн, Йилдиз; Шрайбер, Ульрих; Майер, Кристиан; Шмитц, Оливер Дж. (21 июня 2022 г.). «Алифатические альдегиды в земной коре — остатки пребиотической химии?» . Жизнь . 12 (7): 925. Бибкод : 2022Life...12..925G . дои : 10.3390/life12070925 . ISSN 2075-1729 . ПМЦ 9319801 . ПМИД 35888015 .
- ^ Майер, Кристиан; Шрайбер, Ульрих; Давила, Мария (7 января 2017 г.). «Выбор пребиотических молекул в амфифильных средах» . Жизнь . 7 (1): 3. Бибкод : 2017Жизнь....7....3М . дои : 10.3390/life7010003 . ISSN 2075-1729 . ПМК 5370403 . ПМИД 28067845 .
- ^ Майер, Кристиан; Шрайбер, Ульрих; Давила, Мария Дж. (2015). «Периодическое образование пузырьков в зонах тектонических разломов — идеальный сценарий молекулярной эволюции» . Происхождение жизни и эволюция биосфер . 45 (1–2): 139–148. Бибкод : 2015ОЛЕВ...45..139М . дои : 10.1007/s11084-015-9411-z . ISSN 0169-6149 . ПМЦ 4457167 . ПМИД 25716918 .
- ^ Майер, Кристиан; Шрайбер, Ульрих; Давила, Мария; Шмитц, Оливер; Бронха, Амела; Мейер, Мартин; Кляйн, Джулия; Меккельманн, Свен (24 мая 2018 г.). «Молекулярная эволюция в системе пептид-везикула» . Жизнь . 8 (2): 16. Бибкод : 2018Жизнь....8...16М . дои : 10.3390/life8020016 . ISSN 2075-1729 . ПМК 6027363 . ПМИД 29795023 .
- ^ Jump up to: а б Плассон, Рафаэль; Кондепуди, Дилип К.; Берсини, Хьюз; и др. (август 2007 г.). «Появление гомохиральности в системах, далеких от равновесия: механизмы и роль в химии пребиотиков». Хиральность . 19 (8): 589–600. дои : 10.1002/чир.20440 . ПМИД 17559107 . «Специальный выпуск: материалы восемнадцатого международного симпозиума по хиральности (ISCD-18), Пусан, Корея, 2006 г.»
- ^ Чайчян, Рохас и Туреану, 2014 , стр. 353–364.
- ^ Джафарпур, Фаршид; Бьянкалани, Томмазо; Голденфельд, Найджел (2017). «Вызванное шумом нарушение симметрии вдали от равновесия и возникновение биологической гомохиральности» (PDF) . Физический обзор E . 95 (3): 032407. Бибкод : 2017PhRvE..95c2407J . дои : 10.1103/PhysRevE.95.032407 . ПМИД 28415353 . Архивировано из оригинала 2 апреля 2023 года . Проверено 29 августа 2019 г.
- ^ Джафарпур, Фаршид; Бьянкалани, Томмазо; Голденфельд, Найджел (2015). «Индуцированный шумом механизм биологической гомохиральности саморепликаторов раннего возраста». Письма о физических отзывах . 115 (15): 158101. arXiv : 1507.00044 . Бибкод : 2015PhRvL.115o8101J . doi : 10.1103/PhysRevLett.115.158101 . ПМИД 26550754 . S2CID 9775791 .
- ^ Франк, ФК (1953). «О спонтанном асимметричном синтезе». Биохимика и биофизика Acta . 11 (4): 459–463. дои : 10.1016/0006-3002(53)90082-1 . ПМИД 13105666 .
- ^ Кларк, Стюарт (июль – август 1999 г.). «Поляризованный звездный свет и направленность жизни». Американский учёный . 87 (4): 336. Бибкод : 1999AmSci..87..336C . дои : 10.1511/1999.4.336 . S2CID 221585816 .
- ^ Сибата, Таканори; Мориока, Хироши; Хаясе, Тадакацу; и др. (17 января 1996 г.). «Высокоэнантиоселективное каталитическое асимметричное автоумножение хирального пиримидилового спирта». Журнал Американского химического общества . 118 (2): 471–472. дои : 10.1021/ja953066g .
- ^ Соаи, Кенсо; Сато, Итару; Сибата, Таканори (2001). «Асимметричный автокатализ и происхождение хиральной гомогенности в органических соединениях». Химическая запись . 1 (4): 321–332. дои : 10.1002/tcr.1017 . ПМИД 11893072 .
- ^ Хазен 2005 , с. 184
- ^ Мейеренрих, Уве (2008). Аминокислоты и асимметрия жизни, пойманная в процессе формирования . Берлин: Шпрингер. стр. 76–79. ISBN 978-3-540-76886-9 .
- ^ Маллен, Лесли (5 сентября 2005 г.). «Строим жизнь из звездного материала» . Журнал астробиологии . Архивировано из оригинала 14 июля 2015 года.
{{cite journal}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Рут-Бернштейн, Роберт (23 июня 2010 г.). «Экспериментальный тест связывания L- и D-аминокислот с L- и D-кодонами позволяет предположить, что гомохиральность и направленность кодонов возникли вместе с генетическим кодом» . Симметрия . 2 (2): 1180–1200. Бибкод : 2010Symm....2.1180R . дои : 10.3390/sym2021180 .
Источники
[ редактировать ]- Альтерманн, Владислав (2009). «Введение: дорожная карта к Фата Моргане?». В Зекбахе, Йозеф; Уолш, Мод (ред.). От окаменелостей к астробиологии: записи жизни на Земле и поиск внеземных биосигнатур . Клеточное происхождение, жизнь в экстремальных средах обитания и астробиология. Том. 12. Дордрехт, Нидерланды; Лондон: Спрингер. ISBN 978-1-4020-8836-0 .
- Бада, Джеффри Л .; Ласкано, Антонио (2009). «Происхождение жизни» . В Русе, Майкл ; Трэвис, Джозеф (ред.). Эволюция: первые четыре миллиарда лет . Предисловие Эдварда О. Уилсона . Кембридж: Belknap Press издательства Гарвардского университета . ISBN 978-0-674-03175-3 . OCLC 225874308 .
- Бартон, Николас Х .; Бриггс, Дерек Э.Г .; Эйзен, Джонатан А .; и др. (2007). Эволюция . Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. ISBN 978-0-87969-684-9 . ОСЛК 86090399 .
- Бастиан, Х. Чарльтон (1871). Способы происхождения низших организмов . Лондон; Нью-Йорк: Макмиллан и компания . OCLC 42959303 . Проверено 6 июня 2015 г.
- Бернал, доктор юридических наук (1951). Физическая основа жизни . Лондон: Рутледж и Кеган Пол.
- Бернал, доктор юридических наук (1960). «Проблема стадий в биопоэзе» . Флоркин М. (ред.). Аспекты происхождения жизни . Международная серия монографий по чистой и прикладной биологии. Оксфорд, Великобритания; Нью-Йорк: Пергамон Пресс . ISBN 978-1-4831-3587-8 .
- Бернал, JD (1967) [Перепечатанная работа А. И. Опарина, первоначально опубликованная в 1924 году; Москва: Московский рабочий . Происхождение жизни . Естественная история Вайденфельда и Николсона. Перевод «Опарина» Энн Синг. Лондон: Вайденфельд и Николсон .
- Бок, Грегори Р.; Гуд, Джейми А., ред. (1996). Эволюция гидротермальных экосистем на Земле (и Марсе?) . Симпозиум Фонда Ciba. Том. 202. Чичестер, Великобритания; Нью-Йорк: Джон Уайли и сыновья . ISBN 978-0-471-96509-1 .
- Бондесон, январь (1999). Русалка Фиджи и другие очерки естественной и неестественной истории . Итака, Нью-Йорк: Издательство Корнельского университета . ISBN 978-0-8014-3609-3 .
- Брайсон, Билл (2004). Краткая история почти всего . Лондон: Черный лебедь . ISBN 978-0-552-99704-1 . OCLC 55589795 .
- Кальвин, Мелвин (1969). Химическая эволюция: молекулярная эволюция на пути к происхождению живых систем на Земле и в других местах . Оксфорд, Великобритания: Clarendon Press . ISBN 978-0-19-855342-7 . OCLC 25220 .
- Чайчян, Масуд; Рохас, Уго Перес; Туряну, Анка (2014). «Физика и жизнь». Основные понятия физики . Конспекты лекций бакалавриата по физике. Берлин; Гейдельберг: Springer Berlin Heidelberg . стр. 353–364. дои : 10.1007/978-3-642-19598-3_12 . ISBN 978-3-642-19597-6 . OCLC 900189038 . S2CID 115247432 .
- Чанг, Томас Минг Сви (2007). Искусственные клетки: биотехнология, наномедицина, регенеративная медицина, кровезаменители, биоинкапсуляция и терапия клетками/стволовыми клетками . Регенеративная медицина, искусственные клетки и наномедицина. Том. 1. Хакенсак, Нью-Джерси: World Scientific . ISBN 978-981-270-576-1 . OCLC 173522612 .
- Далримпл, Дж. Брент (2001). «Возраст Земли в двадцатом веке: проблема (в основном) решена». В Льюисе, CLE; Нелл, С.Дж. (ред.). Возраст Земли: с 4004 г. до н.э. по 2002 г. н.э. Специальное издание Геологического общества. Том. 190. Лондон: Лондонское геологическое общество . стр. 205–221. Бибкод : 2001GSLSP.190..205D . дои : 10.1144/gsl.sp.2001.190.01.14 . ISBN 978-1-86239-093-5 . ОСЛК 48570033 . S2CID 130092094 .
- Дарвин, Чарльз (1887). Дарвин, Фрэнсис (ред.). Жизнь и письма Чарльза Дарвина, включая автобиографическую главу . Том. 3 (3-е изд.). Лондон: Джон Мюррей . OCLC 834491774 .
- Дэвис, Пол (1999). Пятое чудо: поиски происхождения жизни . Лондон: Книги Пингвинов . ISBN 978-0-14-028226-9 .
- Добелл, Клиффорд (1960) [Первоначально опубликовано в 1932 году; Нью-Йорк: Harcourt, Brace & Company ]. Антоний ван Левенгук и его «Маленькие животные» . Нью-Йорк: Dover Publications .
- Дайсон, Фриман (1999). Происхождение жизни (пересмотренная ред.). Кембридж, Великобритания; Нью-Йорк: Издательство Кембриджского университета . ISBN 978-0-521-62668-2 .
- Фесенков, В.Г. (1959). «Некоторые соображения о первобытном состоянии Земли» . В Опарин А.И .; и др. (ред.). Происхождение жизни на Земле . Серия симпозиумов IUB. Том. для Международного союза биохимии 1. Под редакцией Фрэнка Кларка и RLM Synge (англо-французско-немецкое издание). Лондон; Нью-Йорк: Пергамон Пресс . ISBN 978-1-4832-2240-0 . Международный симпозиум по происхождению жизни на Земле (Москва, 19–24 августа 1957 г.)
- Хейзен, Роберт М. (2005). Бытие: научные поиски происхождения жизни . Вашингтон, округ Колумбия: Джозеф Генри Пресс . ISBN 978-0-309-09432-0 . ОСЛК 60321860 .
- Хаксли, Томас Генри (1968) [1897]. «VIII Биогенез и Абиогенез [1870]» . Дискурсы биологические и геологические . Сборник сочинений. Том. VIII (Переиздание). Нью-Йорк: Гринвуд Пресс . OCLC 476737627 . Архивировано из оригинала 7 мая 2015 года . Проверено 19 мая 2014 г.
- Клайце, Бриг (22 января 2001 г.). Кингсли, Стюарт А .; Бхатхал, Рагбир (ред.). Панспермия ставит новые вопросы . Поиски внеземного разума (SETI) в оптическом спектре III . Том. 4273. Беллингем, Вашингтон: SPIE . дои : 10.1117/12.435366 . ISBN 0-8194-3951-7 . Архивировано из оригинала 3 сентября 2013 года . Проверено 9 июня 2015 г. Материалы SPIE состоялись в Сан-Хосе, Калифорния, 22–24 января 2001 г.
- Лейн, Ник (2009). Жизнь по восходящей: 10 великих изобретений эволюции (1-е американское изд.). Нью-Йорк: WW Norton & Company. ISBN 978-0-393-06596-1 . OCLC 286488326 .
- Леннокс, Джеймс Г. (2001). Философия биологии Аристотеля: исследования истоков науки о жизни . Кембриджские исследования по философии и биологии. Кембридж, Великобритания; Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-65976-5 .
- Мишод, Ричард Э. (1999). Дарвиновская динамика: эволюционные переходы в приспособленности и индивидуальности . Принстон, Нью-Джерси: Издательство Принстонского университета . ISBN 978-0-691-02699-2 . OCLC 38948118 .
- Опарин, А.И. (1953) [Первоначально опубликовано в 1938 году; Нью-Йорк: Компания Macmillan ]. Происхождение жизни . Перевод и новое введение Сергия Моргулиса (2-е изд.). Минеола, Нью-Йорк: Dover Publications. ISBN 978-0-486-49522-4 .
- Росс, Александр (1652). Арканы Микрокосма . Том. II. Лондон. OCLC 614453394 . Архивировано из оригинала 24 февраля 2024 года . Проверено 14 июля 2015 г.
- Шелдон, Роберт Б. (22 сентября 2005 г.). «Историческое развитие различия между био- и абиогенезом» (PDF) . В Гувере, Ричард Б .; Левин, Гилберт В .; Розанов Алексей Юрьевич; Гладстон, Дж. Рэндалл (ред.). Астробиология и планетарные миссии . Астробиология и планетарные миссии . Том. 5906. Беллингем, Вашингтон: SPIE. стр. 59061И. дои : 10.1117/12.663480 . ISBN 978-0-8194-5911-4 . Архивировано (PDF) из оригинала 13 апреля 2015 г. Проверено 13 апреля 2015 г. Слушания SPIE состоялись в Сан-Диего, Калифорния, 31 июля – 2 августа 2005 г.
- Тиндаль, Джон (1905) [Первоначально опубликовано в 1871 году; Лондон; Нью-Йорк: Longmans , Green & Co.; Д. Эпплтон и компания ]. Фрагменты науки . Том. 2 (6-е изд.). Нью-Йорк: PF Collier & Sons . OCLC 726998155 .
- Воэт, Дональд ; Фут, Джудит Г. (2004). Биохимия . Том. 1 (3-е изд.). Нью-Йорк: Джон Уайли и сыновья. ISBN 978-0-471-19350-0 .
- Ярус, Майкл (2010). Жизнь из мира РНК: Внутренний предок . Кембридж, Массачусетс: Издательство Гарвардского университета . п. 287. ИСБН 978-0-674-05075-4 .
Внешние ссылки
[ редактировать ]- Прогресс в раскрытии тайн происхождения жизни – Адам Манн ( PNAS ; 14 апреля 2021 г.)
- Исследование истоков жизни. Архивировано 8 апреля 2023 года в Wayback Machine виртуальной выставке в Музее науки (Бостон).
- Как началась жизнь на Земле - Марсия Мэлори (Факты о Земле; 2015)
- Происхождение жизни - Ричард Докинз и др. ( Радио Би-би-си ; 2004)
- Жизнь во Вселенной - эссе Стивена Хокинга (1996)
Элементы природы |
|---|
