c-Раф
| РАФ1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | RAF1 , протоонкоген Raf-1, серин/треониновая киназа, CMD1NN, CRAF, NS5, Raf-1, c-Raf | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 164760 ; МГИ : 97847 ; Гомологен : 48145 ; GeneCards : RAF1 ; ОМА : РАФ1 – ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||



Протоонкогенная серин/треонин-протеинкиназа RAF , также известная как протоонкоген c-RAF или просто c-Raf или даже Raf-1 , представляет собой фермент [ 4 ] у людей кодируется геном RAF1 это . [ 5 ] [ 6 ] Белок c-Raf является частью пути ERK1/2 в виде киназы MAP (MAP3K), которая действует ниже подсемейства Ras мембранассоциированных ГТФаз. [ 7 ] C-Raf является членом семейства -киназ Raf серин/треонин-специфичных протеинкиназ из группы киназ TKL (тирозинкиназоподобных).
Открытие
[ редактировать ]Первый ген Raf, v-Raf, был обнаружен в 1983 году. Он был выделен из мышиного ретровируса под номером 3611. Вскоре было продемонстрировано, что он способен трансформировать фибробласты грызунов в раковые клеточные линии , поэтому этому гену было присвоено название Virus-. индуцированная быстро ускоряющаяся фибросаркома (V-RAF). [ 5 ] Год спустя в птичьем ретровирусе MH2 был обнаружен еще один трансформирующий ген, названный v-Mil, который оказался очень похожим на v-Raf. [ 8 ] Исследователям удалось продемонстрировать, что эти гены кодируют ферменты, обладающие серин-треониновой киназной активностью. [ 9 ] Нормальные клеточные гомологи v-Raf и v-Mil вскоре были обнаружены как в геноме мыши, так и в геноме курицы (отсюда и название c-Raf для нормального клеточного гена Raf), и стало ясно, что они тоже играют роль в регуляции роста и деление клеток. [ 10 ] [ 11 ]
c-Raf является основным компонентом пути митоген-активируемой протеинкиназы (MAPK): передача сигналов ERK1/2 . [ 12 ] Он действует как киназа MAP3, запуская весь киназный каскад. Последующие эксперименты показали, что нормальные клеточные гены Raf также могут мутировать и становиться онкогенами, «перегружая» активность MEK1/2 и ERK1/2. [ 13 ] Фактически, геномы позвоночных содержат множество генов Raf. Несколько лет спустя после открытия c-Raf были описаны еще две родственные киназы: A-Raf и B-Raf . Последнее стало предметом исследований в последние годы, поскольку большая часть опухолей человека несет онкогенные «драйверные» мутации в гене B-Raf. [ 14 ] Эти мутации вызывают неконтролируемую высокую активность ферментов Raf. Таким образом, в последние годы диагностический и терапевтический интерес к Raf-киназам достиг нового пика. [ 15 ]
Структура
[ редактировать ]Ген c-Raf человека расположен на хромосоме 3 . по крайней мере две изоформы мРНК Описаны (возникающие в результате включения или удаления альтернативного экзона ), которые имеют лишь незначительные различия. Более короткая основная изоформа, состоящая из 17 экзонов , кодирует протеинкиназу из 648 аминокислот. [ 16 ]

Подобно многим другим MAPKKK , c-Raf представляет собой многодоменный белок с несколькими дополнительными доменами , помогающими регулировать его каталитическую активность. На его N-концевом сегменте рядом друг с другом расположены Ras-связывающий домен (RBD) и гомологичный домен 1 C-киназы (домен C1). Структуры обоих консервативных доменов были раскрыты в последние десятилетия, что пролило свет на механизмы их регуляции.
Ras -связывающий домен имеет убиквитиноподобную складку (как и многие другие небольшие домены, ассоциированные с G-белками) и избирательно связывает только GTP-связанные белки Ras. [ 17 ] [ 18 ] [ 19 ] (Вы можете увидеть это взаимодействие более подробно в блоке PDB, прикрепленном к статье. Он показывает Rap1 в комплексе с RBD c-Raf.)
Домен C1 , расположенный сразу после связывающего домена Ras, представляет собой особый цинковый палец , богатый цистеинами и стабилизированный двумя ионами цинка. Он похож на диацилглицеринсвязывающие домены C1 ферментов протеинкиназы C (PKC). [ 20 ] [ 21 ] Но в отличие от PKC, домены C1 киназ семейства Raf не связывают диацилглицерин. [ 22 ] Вместо этого они взаимодействуют с другими липидами, такими как церамиды. [ 22 ] или фосфатидная кислота, [ 23 ] и даже помощь в распознавании активированного Ras (GTP-Ras). [ 21 ] [ 24 ]
Непосредственная близость этих двух доменов, а также несколько линий экспериментальных данных позволяют предположить, что они действуют как единое целое и отрицательно регулируют активность протеинкиназного домена путем прямого физического взаимодействия. [ 25 ] Исторически этот аутоингибирующий блок назывался регионом CR1 («Консервативный регион 1»), шарнирная область называлась CR2, а киназный домен CR3. К сожалению, точная структура аутоингибированной киназы остается неизвестной.
Между блоком аутоингибирующего домена и каталитическим киназным доменом можно обнаружить длинный сегмент, характерный для всех белков Raf. Он высоко обогащен сериновыми аминокислотами, но его точная последовательность плохо консервативна в родственных генах Raf. Этот регион кажется по своей сути неструктурированным и очень гибким. Его наиболее вероятная роль — действовать как естественный «шарнир» между жестко свернутыми аутоингибирующими и каталитическими доменами, обеспечивая сложные движения и глубокие конформационные перестройки внутри молекулы. [ 26 ] Эта шарнирная область содержит небольшой консервативный островок аминокислот, которые отвечают за распознавание белка 14-3-3 , но только тогда, когда критический серин (Ser259 в c-Raf человека) фосфорилируется. Второй аналогичный мотив обнаружен на крайнем С-конце (в центре вокруг фосфорилируемого Ser 621) всех ферментов Raf, но ниже киназного домена.
С-концевая половина c-Raf складывается в один белковый домен, отвечающий за каталитическую активность. Структура этого киназного домена хорошо известна как из c-Raf, так и из c-Raf. [ 27 ] и Б-Раф. [ 28 ] Он очень похож на другие киназы Raf и белки KSR, а также явно похож на некоторые другие киназы MAP3, такие как семейство киназ смешанного происхождения (MLK). Вместе они составляют группу протеинкиназ, подобных тирозинкиназе (TKL). Хотя некоторые особенности объединяют их каталитические домены с протеинтирозинкиназами, активность TKL ограничивается фосфорилированием остатков серина и треонина в белках-мишенях. Наиболее важным субстратом киназ Raf (кроме самих себя) являются киназы MKK1 и MKK2 , активность которых строго зависит от событий фосфорилирования, осуществляемых Rafs.
Эволюционные отношения
[ редактировать ]Человеческий c-Raf является членом более крупного семейства родственных протеинкиназ. Еще два члена, обнаруженные у большинства позвоночных, принадлежат к тому же семейству: B-Raf и A-Raf . Помимо разной длины их неконсервативных N- и C-концевых концов, все они имеют одинаковую доменную архитектуру, структуру и регуляцию. По сравнению с относительно известными c-Raf и B-Raf, о точной функции A-Raf известно очень мало, но также считается, что он похож на двух других членов семейства. Считается, что все эти гены являются продуктом полной дупликации генов или геномов на заре эволюции позвоночных из одного предкового гена Raf. Большинство других животных организмов обладают только одним геном Raf. У дрозофилы он называется Phl или Draf. [ 29 ] и Lin-45 у C. elegans. [ 30 ]

У многоклеточных животных также имеется тип киназы, тесно связанный с Raf: это киназный супрессор Ras (KSR). У позвоночных, таких как млекопитающие, есть два паралогичных гена KSR вместо одного: KSR1 и KSR2 . Их С-концевой киназный домен очень похож на Raf (первоначально называвшийся CA5 в KSR и CR3 в Raf), но N-концевая регуляторная область отличается. Хотя у них также есть гибкий шарнир (CA4 в KSR) и домен C1 (CA3 в KSR) перед ним, у KSR полностью отсутствует Ras-связывающий домен. Вместо этого у них есть уникальные регуляторные области на N-концах, первоначально названные CA1 («консервативная область 1») и CA2. Долгое время структура домена CA1 оставалась загадкой. Однако в 2012 году структура региона CA1 в KSR1 была решена: это оказался дивергентный домен SAM (стерильный альфа-мотив), дополненный спиральными спиралями (CC-SAM): это должно помочь KSR в мембране. привязка. [ 31 ] KSR, как и Rafs, также имеют двойные связывающие мотивы 14-3-3 (которые зависят от фосфорилирования), но также обладают новыми мотивами связывания MAPK в своих шарнирных областях. При типичной последовательности Phe-x-Phe-Pro (FxFP) эти мотивы важны для регуляции по принципу обратной связи Raf-киназ в пути ERK1/2 . Согласно нашим нынешним знаниям, KSR также участвуют в том же пути, что и Raf, хотя они играют лишь вспомогательную роль. Из-за очень низкой внутренней киназной активности они долгое время считались неактивными, пока в последние годы наконец не была продемонстрирована их каталитическая активность. [ 32 ] [ 33 ] Но даже в этом случае они вносят лишь незначительный вклад в фосфорилирование МКК1 и МКК2 . Основная роль KSR, по-видимому, заключается в обеспечении партнера по гетеродимеризации ферментов Raf, значительно облегчая их активацию посредством аллостерии. Подобные явления были описаны и для других киназ MAP3. ASK2, например, сам по себе является плохим ферментом, и его активность, по-видимому, связана с гетеродимеризацией ASK1/ASK2. [ 34 ]
Raf-подобные киназы полностью отсутствуют у грибов. Но недавнее секвенирование других опистоконтов (например, Capsaspora owczarzaki ) выявило присутствие подлинных киназ Raf у одноклеточных эукариот. Следовательно, возможно, что белки Raf являются древним наследием и предки грибов вторично утратили Raf-зависимую передачу сигналов. грибов Пути киназы MAP , которые гомологичны пути ERK1/2 млекопитающих (Fus3 и Kss1 у дрожжей), активируются родственными MEKK киназами (например, Ste11 у дрожжей) вместо ферментов Raf.
Киназы Raf, обнаруженные в ретровирусах (таких как мышиный v-Raf), вторично происходят из соответствующих генов позвоночных их хозяев. Эти гены Raf кодируют сильно усеченные белки, в которых отсутствует весь N-концевой аутоингибирующий домен и мотивы связывания 14-3-3. Известно, что такие серьезные сокращения вызывают неконтролируемую активность Raf-киназ: это именно то, что может понадобиться вирусу для эффективного размножения.
Регулирование деятельности
[ редактировать ]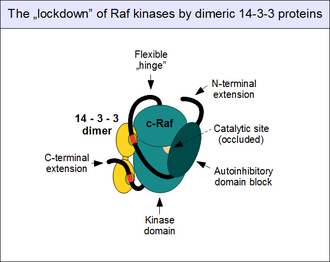
Как упоминалось выше, регуляция активности c-Raf является сложной. Будучи «привратником» пути ERK1/2 , он контролируется множеством тормозных механизмов и обычно не может быть активирован за один этап. Наиболее важный механизм регуляции включает прямую физическую ассоциацию N-концевого аутоингибирующего блока с киназным доменом c-Raf. Это приводит к окклюзии каталитического сайта и полному прекращению активности киназы. [ 25 ] Это «закрытое» состояние можно облегчить только в том случае, если аутоингибирующий домен Raf вовлекает партнера, конкурирующего с его собственным киназным доменом, что наиболее важно с GTP-связанным Ras. Таким образом, активированные малые G-белки могут разрушать внутримолекулярные взаимодействия: это приводит к конформационному изменению («открытию») c-Raf. [ 37 ] необходим для активации киназы и связывания субстрата.
Белки 14-3-3 также способствуют аутоингибированию. Поскольку известно, что все белки 14-3-3 образуют конститутивные димеры, их сборки имеют два сайта связывания. [ 38 ] Таким образом, димер действует как «молекулярные наручники», удерживая своих партнеров по связыванию на фиксированном расстоянии и в фиксированной ориентации. Когда точно расположенные двойные связывающие мотивы 14-3-3 взаимодействуют с одним димером белка 14-3-3 (например, 14-3-3-дзета), они замыкаются в конформацию, которая способствует аутоингибированию и не позволяет расцепиться. автоингибирующих и каталитических доменов. [ 39 ] Эта «блокировка» c-Raf (и других Rafs, а также KSR) контролируется фосфорилированием мотива. Нефосфорилированные мотивы, связывающие 14-3-3, не связывают своих партнеров: им необходимо сначала фосфорилироваться по консервативным серинам (Ser 259 и Ser 621) с помощью других протеинкиназ. Наиболее важной киназой, участвующей в этом событии, является киназа 1, активируемая TGF-бета (TAK1), а ферментами, предназначенными для удаления этих фосфатов, являются комплексы протеинфосфатазы 1 (PP1) и протеинфосфатазы 2A (PP2A). [ 40 ] [ 41 ]
Обратите внимание, что связывание 14-3-3 ферментов Raf не обязательно является ингибирующим: как только Raf открывается и димеризуется, 14-3-3 также могут связываться в транс , соединяя две киназы и «связывая» их вместе, чтобы укрепить димер, а не держать их подальше друг от друга. [ 42 ] Существуют и другие способы взаимодействия 14-3-3 с c-Raf, но их роль малоизвестна. [ 43 ]
Димеризация является еще одним важным механизмом регуляции активности c-Raf, необходимым для фосфорилирования петли активации Raf . Обычно в димеризации участвуют только «открытые» киназные домены. В отличие от B-Raf, который легко образует гомодимеры сам с собой, c-Raf предпочитает гетеродимеризацию либо с B-Raf, либо с KSR1. [ нужна ссылка ] Гомодимеры и гетеродимеры ведут себя одинаково. [ 33 ] Структура домена гомодимеркиназы B-Raf ясно показывает, что петли активации (которые контролируют каталитическую активность всех известных протеинкиназ) расположены в димере в активно-подобной конформации. Это происходит из-за аллостерического эффекта связывания другой молекулы с «обратной» стороной киназы; такие димеры симметричны и имеют два частично активных каталитических центра. На этом этапе активность Raf-киназ низкая и нестабильная.

Для достижения полной активности и стабилизации активного состояния петлю активации c-Raf необходимо фосфорилировать. Единственные киназы, которые, как известно, выполняют это действие, — это сами киназы семейства Raf. Но некоторые другие киназы, такие как PAK1, могут фосфорилировать другие остатки вблизи киназного домена c-Raf: точная роль этих вспомогательных киназ неизвестна. В контексте c-Raf и c-Raf, и KSR1 необходимы для стадии «трансфосфорилирования». Из-за строения димеров это фосфорилирование может происходить только в транс-режиме (т.е. один димер фосфорилирует другой в четырехчленном переходном комплексе). [ 44 ] Взаимодействуя с консервативными остатками Arg и Lys в киназном домене, фосфорилированные петли активации меняют конформацию и становятся упорядоченными, навсегда фиксируя киназный домен в полностью активном состоянии до тех пор, пока он не будет дефосфорилирован. Фосфорилированные петли активации также делают киназу нечувствительной к присутствию ее аутоингибирующего домена. [ 45 ] KSR не могут пройти этот последний этап, поскольку в их петлях активации отсутствуют какие-либо фосфорилируемые остатки. Но как только c-Raf полностью активирован, в этом больше нет необходимости: активные ферменты Raf теперь могут взаимодействовать со своими субстратами. [ 46 ] Как и большинство протеинкиназ, c-Raf имеет несколько субстратов. БАД (Bcl2-атагонист клеточной гибели) непосредственно фосфорилируется c-Raf, [ 47 ] наряду с несколькими типами аденилатциклаз , [ 48 ] миозинфосфатаза (MYPT), [ 49 ] тропонин Т сердечной мышцы (TnTc), [ 50 ] и т. д. белок ретинобластомы (pRb) и фосфатаза Cdc25 . В качестве возможных субстратов были также предложены [ 51 ]
Наиболее важными мишенями всех ферментов Raf являются MKK1(MEK1) и MKK2(MEK2) . Хотя структура фермент-субстратного комплекса c-Raf:MKK1 неизвестна, ее можно точно смоделировать по комплексу KSR2:MKK1. [ 33 ] Здесь никакого настоящего катализа не происходит, но считается, что он очень похож на то, как Raf связывается со своими субстратами. Основной интерфейс взаимодействия обеспечивается С-концевыми долями обоих киназных доменов; большая, неупорядоченная, богатая пролином петля, уникальная для MKK1 и MKK2, также играет важную роль в ее позиционировании по отношению к Raf (и KSR). [ 52 ] Эти МКК фосфорилируются по крайней мере в двух сайтах своих петель активации при связывании с Raf: это также активирует их. Мишенью киназного каскада являются ERK1 и ERK2, которые избирательно активируются МКК1 или МКК2. ERK имеют многочисленные субстраты в клетках; они также способны перемещаться в ядро для активации ядерных факторов транскрипции. Активированные ERK являются плейотропными эффекторами клеточной физиологии и играют важную роль в контроле экспрессии генов, участвующих в цикле деления клеток, миграции клеток, ингибировании апоптоза и дифференцировке клеток.
Сопутствующие заболевания человека
[ редактировать ]Наследственные мутации c-Raf, приводящие к увеличению функции, участвуют в некоторых редких, но тяжелых синдромах. Большинство этих мутаций включают изменения отдельных аминокислот в одном из двух мотивов связывания 14-3-3. [ 53 ] [ 54 ] Мутация c-Raf является одной из возможных причин синдрома Нунан : у больных наблюдаются врожденные пороки сердца, низкий и дисморфный рост и ряд других уродств. Подобные мутации в c-Raf также могут вызывать родственное состояние, называемое синдромом LEOPARD (лентиго, электрокардиографические нарушения, глазной гипертелоризм, легочный стеноз, аномалии гениталий, задержка роста, глухота) со сложной ассоциацией дефектов.
Роль в раке
[ редактировать ]Хотя c-Raf явно способен мутировать в онкоген в экспериментальных условиях и даже в некоторых опухолях человека, [ 55 ] [ 56 ] его сестринская киназа B-Raf действительно играет важную роль в канцерогенезе у человека. [ 57 ]
Мутации B-Raf
[ редактировать ]Примерно 20% всех исследованных образцов опухолей человека содержат мутированный ген B-Raf. [ 58 ] Подавляющее большинство этих мутаций связано с заменой одной аминокислоты: Val 600 на Glu, и этот аберрантный генный продукт (BRAF-V600E) можно визуализировать с помощью иммуногистохимии для клинической молекулярной диагностики. [ 59 ] [ 60 ] Аберрация может имитировать фосфорилирование петли активации и, пропуская все этапы управления при нормальной активации, немедленно сделать киназный домен полностью активным. [ 61 ] Поскольку B-Raf также может активироваться путем гомодимеризации, а c-Raf - путем гетеродимеризации, эта мутация имеет катастрофический эффект, делая путь ERK1/2 конститутивно активным и запуская неконтролируемый процесс клеточного деления. [ 62 ]
В качестве терапевтической цели
[ редактировать ]Из-за важности мутаций Ras и B-Raf в онкогенезе было разработано несколько ингибиторов Raf для борьбы с раком, особенно против B-Raf, проявляющего мутацию V600E. Сорафениб стал первым клинически полезным препаратом, который представляет собой фармакологическую альтернативу для лечения ранее практически неизлечимых злокачественных новообразований, таких как почечно-клеточный рак и меланома. [ 63 ] Далее последовали несколько других молекул, таких как вемурафениб , регорафениб , дабрафениб и т. д.
К сожалению, АТФ-конкурентные ингибиторы B-Raf могут оказывать нежелательное воздействие на K-Ras-зависимые виды рака: они просто слишком селективны в отношении B-Raf. Хотя они прекрасно ингибируют активность B-Raf в случае, если мутантный B-Raf является основным виновником, они также способствуют гомо- и гетеродимеризации B-Raf с самим собой и c-Raf. Это фактически усилит активацию c-Raf, а не ингибирует ее в случае, если в каких-либо генах Raf нет мутаций, но их общий вышестоящий белок-активатор K-Ras является мутированным. [ 27 ] Эта «парадоксальная» активация c-Raf обуславливает необходимость скрининга мутаций B-Raf у пациентов (путем генетической диагностики) перед началом терапии ингибиторами B-Raf. [ 64 ]
Список взаимодействующих белков
[ редактировать ]Было показано, что C-Raf взаимодействует с:
- АКТ1 , [ 65 ]
- СПРОС1 , [ 66 ]
- СУМКА1 , [ 67 ]
- БРАФ , [ 68 ]
- Бкл-2 , [ 69 ]
- CDC25A , [ 70 ] [ 71 ]
- ЦФЛАР , [ 72 ]
- ФИН , [ 73 ]
- ГРБ10 , [ 74 ] [ 75 ]
- ХРАС , [ 76 ] [ 77 ] [ 78 ] [ 79 ] [ 80 ] [ 81 ] [ 82 ] [ 83 ] [ 84 ] [ 85 ] [ 86 ] [ 87 ] [ 88 ] [ 89 ] [ 90 ] [ 91 ] [ 92 ]
- ХСП90АА1 , [ 93 ] [ 94 ]
- КРАС , [ 81 ] [ 82 ]
- МАП2К1 , [ 95 ]
- МАП3К1 , [ 96 ]
- MAPK7 , [ 97 ]
- МАПК8IP3 , [ 98 ] [ 99 ]
- ПАК1 , [ 100 ]
- ПЭБП1 , [ 95 ]
- ПОБ , [ 101 ]
- ПРКЦЗ , [ 102 ]
- РАП1А , [ 17 ] [ 86 ] [ 103 ] [ 104 ]
- РЕБ , [ 105 ] [ 106 ] [ 107 ]
- RRAS2 [ 81 ] [ 108 ]
- РБ1 , [ 101 ] [ 109 ]
- РБЛ2 , [ 109 ]
- ШОК2 , [ 81 ]
- ШТУЦ1 , [ 93 ]
- СРК , [ 73 ]
- ТСЦ22Д3 , [ 110 ]
- Иегова , [ 80 ] [ 102 ] [ 111 ] [ 112 ] [ 113 ] [ 114 ]
- ЯХВЕ , [ 113 ] [ 114 ]
- ЮВАГ , [ 102 ] [ 115 ] [ 116 ]
- ЯХА , [ 102 ] [ 113 ] [ 117 ]
- ЮВАК , [ 95 ] [ 102 ] [ 115 ] [ 118 ] и
- ЯВАЗ . [ 102 ] [ 119 ] [ 120 ] [ 121 ] [ 122 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000000441 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Ли П., Вуд К., Мамон Х., Хазер В., Робертс Т. (февраль 1991 г.). «Raf-1: киназа, в настоящее время не имеющая причины, но не лишенная последствий». Клетка . 64 (3): 479–82. дои : 10.1016/0092-8674(91)90228-Q . ПМИД 1846778 . S2CID 37427156 .
- ^ Перейти обратно: а б Рэпп У.Р., Голдсборо, доктор медицинских наук, Марк Дж.Е., Боннер Т.И., Гроффен Дж., Рейнольдс Ф.Х., Стивенсон-младший (июль 1983 г.). «Структура и биологическая активность v-raf, уникального онкогена, трансдуцируемого ретровирусом» . Учеб. Натл. акад. наук. США . 80 (14): 4218–22. Бибкод : 1983PNAS...80.4218R . дои : 10.1073/pnas.80.14.4218 . ПМК 384008 . ПМИД 6308607 .
- ^ Боннер Т., О'Брайен С.Дж., Нэш В.Г., Рэпп У.Р., Мортон К.С., Ледер П. (январь 1984 г.). «Человеческие гомологи онкогена raf (mil) расположены на хромосомах 3 и 4 человека». Наука . 223 (4631): 71–4. Бибкод : 1984Sci...223...71B . дои : 10.1126/science.6691137 . ПМИД 6691137 .
- ^ «Ген Энтреза: гомолог 1 вирусного онкогена мышиного лейкоза RAF1 v-raf-1» .
- ^ Сутраве П., Боннер Т.И., Рапп У.Р., Янсен Х.В., Пачинский Т., Бистер К. (1984). «Нуклеотидная последовательность птичьего ретровирусного онкогена v-mil: гомолог мышиного ретровирусного онкогена v-raf». Природа . 309 (5963): 85–8. Бибкод : 1984Natur.309...85S . дои : 10.1038/309085a0 . ПМИД 6325930 . S2CID 4357047 .
- ^ Моеллинг К., Хейманн Б., Беймлинг П., Рапп У.Р., Сандер Т. (1984). «Серин- и треонин-специфическая протеинкиназная активность очищенных белков gag-mil и gag-raf». Природа . 312 (5994): 558–61. Бибкод : 1984Natur.312..558M . дои : 10.1038/312558a0 . ПМИД 6438534 . S2CID 4269749 .
- ^ Колх В., Хайдекер Г., Ллойд П., Рапп У.Р. (январь 1991 г.). «Протеинкиназа Raf-1 необходима для роста индуцированных клеток NIH/3T3» . Природа . 349 (6308): 426–8. Бибкод : 1991Natur.349..426K . дои : 10.1038/349426a0 . ПМИД 1992343 . S2CID 4368936 .
- ^ Марк Г.Е., Рапп УР (апрель 1984 г.). «Первичная структура v-raf: родство с семейством онкогенов src». Наука . 224 (4646): 285–9. Бибкод : 1984Sci...224..285M . дои : 10.1126/science.6324342 . ПМИД 6324342 .
- ^ Кириакис Дж. М., Апп Х., Чжан К. Ф., Банерджи П., Бротиган Д. Л., Рапп У. Р., Авруч Дж. (июль 1992 г.). «Raf-1 активирует MAP-киназу-киназу». Природа . 358 (6385): 417–21. Бибкод : 1992Natur.358..417K . дои : 10.1038/358417a0 . ПМИД 1322500 . S2CID 4335307 .
- ^ Симидзу К., Накацу Ю., Номото С., Секигути М. (1986). «Структура активированного гена c-raf-1 рака желудка человека». Межд. Симп. Принцесса Такамацу Рак Рез. Фонд . 17 : 85–91. ПМИД 2843497 .
- ^ Дэвис Х, Бигнелл Г.Р., Кокс С., Стивенс П., Эдкинс С., Клегг С., Тиг Дж., Воффендин Х., Гарнетт М.Дж., Боттомли В., Дэвис Н., Дикс Е., Юинг Р., Флойд Ю., Грей К., Холл С., Хоуз Р. , Хьюз Дж., Космиду В., Мензис А., Молд С., Паркер А., Стивенс С., Уотт С., Хупер С., Уилсон Р., Джаятилаке Х., Гастерсон Б.А., Купер С., Шипли Дж., Харгрейв Д., Притчард-Джонс К., Мейтленд Н., Ченевикс-Тренч Дж., Риггинс Г.Дж., Бигнер Д.Д., Палмиери Дж., Коссу А., Фланаган А., Николсон А., Хо Дж.В. , Люнг С.Ю., Юэнь С.Т., Вебер Б.Л., Зейглер Х.Ф., Дэрроу Т.Л., Патерсон Х., Марэ Р., Маршалл С.Дж., Вустер Р., Страттон М.Р., Futreal PA (июнь 2002 г.). «Мутации гена BRAF при раке человека» (PDF) . Природа . 417 (6892): 949–54. Бибкод : 2002Natur.417..949D . дои : 10.1038/nature00766 . ПМИД 12068308 . S2CID 3071547 .
- ^ Шридхар С.С., Хедли Д., Сиу Л.Л. (апрель 2005 г.). «Киназа Raf как мишень для противораковой терапии» . Мол. Рак Там . 4 (4): 677–85. doi : 10.1158/1535-7163.MCT-04-0297 . ПМИД 15827342 .
- ^ Дозье С., Ансьо С., Феррейра Э., Колл Дж., Стехелин Д. (август 1991 г.). «Альтернативно сплайсированная мРНК c-mil/raf преимущественно экспрессируется в мышечных тканях кур и консервативна среди видов позвоночных». Онкоген . 6 (8): 1307–11. ПМИД 1886707 .
- ^ Перейти обратно: а б Нассар Н., Хорн Г., Херрманн С., Шерер А., Маккормик Ф., Виттингхофер А. (июнь 1995 г.). «Кристаллическая структура 2,2 A Ras-связывающего домена серин/треониновой киназы c-Raf1 в комплексе с Rap1A и аналогом GTP». Природа . 375 (6532): 554–60. Бибкод : 1995Natur.375..554N . дои : 10.1038/375554a0 . ПМИД 7791872 . S2CID 4347807 .
- ^ Эмерсон С.Д., Мэдисон В.С., Палермо Р.Э., Во Д.С., Шеффлер Дж.Э., Цао К.Л., Кифер С.Е., Лю С.П., Фрай, округ Колумбия (май 1995 г.). «Структура раствора Ras-связывающего домена c-Raf-1 и идентификация его поверхности взаимодействия Ras». Биохимия . 34 (21): 6911–8. дои : 10.1021/bi00021a001 . ПМИД 7766599 .
- ^ Муди С.А., Виллумсен Б.М., Вебер М.Дж., Человек-волк А. (июнь 1993 г.). «Комплексы Ras.GTP с Raf-1 и митоген-активируемой протеинкиназной киназой». Наука . 260 (5114): 1658–61. Бибкод : 1993Sci...260.1658M . дои : 10.1126/science.8503013 . ПМИД 8503013 .
- ^ Мотт Х.Р., Карпентер Дж.В., Чжун С., Гош С., Белл Р.М., Кэмпбелл С.Л. (август 1996 г.). «Структура раствора богатого цистеином домена Raf-1: новый сайт связывания ras и фосфолипидов» . Учеб. Натл. акад. наук. США . 93 (16): 8312–7. Бибкод : 1996PNAS...93.8312M . дои : 10.1073/pnas.93.16.8312 . ПМК 38667 . ПМИД 8710867 .
- ^ Перейти обратно: а б Дауб М., Йокель Дж., Кряк Т., Вебер К.К., Шмитц Ф., Рапп У.Р., Виттингхофер А., Блок C (ноябрь 1998 г.). «Богатый цистеином домен RafC1 содержит множество различных регуляторных эпитопов, которые контролируют Ras-зависимую активацию Raf» . Мол. Клетка. Биол . 18 (11): 6698–710. дои : 10.1128/mcb.18.11.6698 . ПМК 109253 . ПМИД 9774683 .
- ^ Перейти обратно: а б Инь X, Зафрулла М, Ли Х, Хаймовиц-Фридман А, Фукс З, Колесник Р (2009). «Церамид-связывающий домен C1 опосредует киназный супрессор мембранной транслокации ras» . Клетка. Физиол. Биохим . 24 (3–4): 219–30. дои : 10.1159/000233248 . ПМЦ 2978518 . ПМИД 19710537 .
- ^ Крафт К.А., Гарридо Дж.Л., Флухарти Э., Лейва-Вега Л., Ромеро Дж. (декабрь 2008 г.). «Роль фосфатидной кислоты в соединении каскада ERK» . Ж. Биол. Хим . 283 (52): 36636–45. дои : 10.1074/jbc.M804633200 . ПМК 2606017 . ПМИД 18952605 .
- ^ Бртва Т.Р., Друган Дж.К., Гош С., Террелл Р.С., Кэмпбелл-Берк С., Белл Р.М., Дер СиДжей (апрель 1995 г.). «Два отдельных домена Raf опосредуют взаимодействие с Ras» . Ж. Биол. Хим . 270 (17): 9809–12. дои : 10.1074/jbc.270.17.9809 . ПМИД 7730360 .
- ^ Перейти обратно: а б Катлер Р.Э., Стивенс Р.М., Сарасино М.Р., Моррисон Д.К. (август 1998 г.). «Ауторегуляция серин/треонин киназы Raf-1» . Учеб. Натл. акад. наук. США . 95 (16): 9214–9. Бибкод : 1998PNAS...95.9214C . дои : 10.1073/pnas.95.16.9214 . ПМК 21318 . ПМИД 9689060 .
- ^ Хмиту И., Дрюилленек С., Валлюэ А., Пейссонно К., Эйшен А. (январь 2007 г.). «Дифференциальная регуляция изоформ B-raf посредством механизмов фосфорилирования и аутоингибирования» . Мол. Клетка. Биол . 27 (1): 31–43. дои : 10.1128/MCB.01265-06 . ПМК 1800654 . ПМИД 17074813 .
- ^ Перейти обратно: а б Хацивассилиу Г, Сонг К, Йен И, Брандхубер Б.Дж., Андерсон DJ, Альварадо Р., Лудлам М.Дж., Стоко Д., Глур С.Л., Вигерс Г., Моралес Т., Алиагас И., Лю Б., Сидерис С., Хефлих К.П., Джайсвал Б.С., Сешагири С. , Кеппен Х., Белвин М., Фридман Л.С., Малек С. (март 2010 г.). «Ингибиторы RAF стимулируют RAF дикого типа для активации пути MAPK и усиления роста» . Природа . 464 (7287): 431–5. Бибкод : 2010Natur.464..431H . дои : 10.1038/nature08833 . ПМИД 20130576 .
- ^ Ван П.Т., Гарнетт М.Дж., Роу С.М., Ли С., Никулеску-Дуваз Д., Гуд В.М., Джонс К.М., Маршалл С.Дж., Спрингер С.Дж., Барфорд Д., Марэ Р. (март 2004 г.). «Механизм активации сигнального пути RAF-ERK онкогенными мутациями B-RAF» . Клетка . 116 (6): 855–67. дои : 10.1016/S0092-8674(04)00215-6 . ПМИД 15035987 . S2CID 126161 .
- ^ Марк Дж.Э., Макинтайр Р.Дж., Диган М.Е., Амбросио Л., Перримон Н. (июнь 1987 г.). «Гомологи онкогена raf Drosophila melanogaster» . Мол. Клетка. Биол . 7 (6): 2134–40. дои : 10.1128/mcb.7.6.2134 . ПМЦ 365335 . ПМИД 3037346 .
- ^ Чонг Х., Викис Х.Г., Гуань К.Л. (май 2003 г.). «Механизмы регуляции семейства Raf-киназ». Клетка. Сигнал . 15 (5): 463–9. дои : 10.1016/S0898-6568(02)00139-0 . ПМИД 12639709 .
- ^ Ковеал Д., Шу-Нуфер Н., Ритт Д., Пейдж Р., Моррисон Д.К., Пети В. (декабрь 2012 г.). «CC-SAM, для спирально-стерильного α-мотива, нацеливает каркас KSR-1 на определенные участки плазматической мембраны» . Научный сигнал . 5 (255): ра94. дои : 10.1126/scisignal.2003289 . ПМЦ 3740349 . ПМИД 23250398 .
- ^ Ху Дж., Ю Х., Корнев А.П., Чжао Дж., Филберт Э.Л., Тейлор С.С., Шоу А.С. (апрель 2011 г.). «Мутация, которая блокирует связывание АТФ, создает псевдокиназу, стабилизирующую каркасную функцию супрессора киназы Ras, CRAF и BRAF» . Учеб. Натл. акад. наук. США . 108 (15): 6067–72. Бибкод : 2011PNAS..108.6067H . дои : 10.1073/pnas.1102554108 . ПМК 3076888 . ПМИД 21441104 .
- ^ Перейти обратно: а б с Бреннан Д.Ф., Дар А.С., Герц НТ, Чао В.К., Берлингейм А.Л., Шокат К.М., Барфорд Д. (апрель 2011 г.). «Аллостерический переход KSR, индуцированный Raf, стимулирует фосфорилирование МЕК». Природа . 472 (7343): 366–9. Бибкод : 2011Natur.472..366B . дои : 10.1038/nature09860 . ПМИД 21441910 . S2CID 18818 .
- ^ Ортнер Э., Мёллинг К. (октябрь 2007 г.). «Образование гетеромерного комплекса ASK2 и ASK1 регулирует передачу сигналов, индуцированную стрессом». Биохим. Биофиз. Рез. Коммун . 362 (2): 454–9. дои : 10.1016/j.bbrc.2007.08.006 . ПМИД 17714688 .
- ^ Перейти обратно: а б Маталланас Д., Биртвистл М., Романо Д., Зебиш А., Раух Дж., фон Кригсхайм А., Колх В. (2011). «Киназы семейства Раф: старые собаки научились новым трюкам» . Гены рака . 2 (3): 232–60. дои : 10.1177/1947601911407323 . ПМК 3128629 . ПМИД 21779496 .
- ^ Перейти обратно: а б Алекса А., Варга Дж., Ремени А. (2010). «Скаффолды являются «активными» регуляторами сигнальных модулей» . ФЕБС Дж . 277 (21): 4376–82. дои : 10.1111/j.1742-4658.2010.07867.x . ПМИД 20883493 . S2CID 43848866 .
- ^ Тераи К., Мацуда М. (март 2005 г.). «Связывание Ras открывает c-Raf, открывая место стыковки для митоген-активируемой протеинкиназы» . Представитель ЭМБО . 6 (3): 251–5. дои : 10.1038/sj.embor.7400349 . ПМЦ 1299259 . ПМИД 15711535 .
- ^ Лю Д., Бьенковска Дж., Петоса С., Коллиер Р.Дж., Фу Х., Лиддингтон Р. (июль 1995 г.). «Кристаллическая структура зета-изоформы белка 14-3-3». Природа . 376 (6536): 191–4. Бибкод : 1995Natur.376..191L . дои : 10.1038/376191a0 . ПМИД 7603574 . S2CID 4366970 .
- ^ Фишер А., Бальюлс А., Рейндерс Дж., Нехорошкова Е., Сибилски С., Мец Р., Альберт С., Раджалингам К., Хекман М., Рапп У.Р. (январь 2009 г.). «Регуляция активности RAF белками 14-3-3: киназы RAF функционально связаны как с гомо-, так и с гетеродимерными формами белков 14-3-3» . Ж. Биол. Хим . 284 (5): 3183–94. дои : 10.1074/jbc.M804795200 . ПМИД 19049963 .
- ^ Родригес-Вициана П., Осес-Прието Дж., Берлингейм А., Фрид М., МакКормик Ф. (апрель 2006 г.). «Голофермент фосфатазы, состоящий из Shoc2/Sur8 и каталитической субъединицы PP1, действует как эффектор M-Ras для модуляции активности Raf» . Мол. Клетка . 22 (2): 217–30. doi : 10.1016/j.molcel.2006.03.027 . ПМИД 16630891 .
- ^ Жаумот М., Хэнкок Дж. Ф. (июль 2001 г.). «Протеинфосфатазы 1 и 2А способствуют активации Raf-1, регулируя взаимодействия 14-3-3» . Онкоген . 20 (30): 3949–58. дои : 10.1038/sj.onc.1204526 . ПМИД 11494123 . S2CID 8800975 .
- ^ Цивион Г., Луо З., Авруч Дж. (июль 1998 г.). «Димерный белок 14-3-3 является важным кофактором активности киназы Raf». Природа . 394 (6688): 88–92. Бибкод : 1998Natur.394...88T . дои : 10.1038/27938 . ПМИД 9665134 . S2CID 204998340 .
- ^ Молзан М., Оттманн С. (ноябрь 2012 г.). «Синергическое связывание фосфорилированных сайтов связывания S233 и S259 C-RAF с одним димером 14-3-3ζ». Дж. Мол. Биол . 423 (4): 486–95. дои : 10.1016/j.jmb.2012.08.009 . ПМИД 22922483 .
- ^ Маккей М.М., Фриман А.К., Моррисон Д.К. (2011). «Сложность функции KSR, выявленная с помощью исследования ингибитора Raf и структуры KSR» . Малые ГТФазы . 2 (5): 276–281. дои : 10.4161/sgtp.2.5.17740 . ПМЦ 3265819 . ПМИД 22292131 .
- ^ Чонг Х, Гуань К.Л. (сентябрь 2003 г.). «Регуляция Raf посредством фосфорилирования и взаимодействия N-конца и C-конца» . Ж. Биол. Хим . 278 (38): 36269–76. дои : 10.1074/jbc.M212803200 . ПМИД 12865432 .
- ^ Ши Ф., Леммон М.А. (май 2011 г.). «Биохимия. КСР играет в CRAF-ти». Наука . 332 (6033): 1043–4. Бибкод : 2011Sci...332.1043S . дои : 10.1126/science.1208063 . ПМИД 21617065 . S2CID 38639290 .
- ^ Е Д.З., Цзинь С., Чжо Ю., Филд Дж. (2011). Бауэр Дж.А. (ред.). «p21-активированная киназа 1 (Pak1) фосфорилирует BAD непосредственно по серину 111 in vitro и опосредованно через Raf-1 по серину 112» . ПЛОС ОДИН . 6 (11): e27637. Бибкод : 2011PLoSO...627637Y . дои : 10.1371/journal.pone.0027637 . ПМК 3214075 . ПМИД 22096607 .
- ^ Дин Кью, Грос Р., Грей И.Д., Тауссиг Р., Фергюсон С.С., Фельдман Р.Д. (октябрь 2004 г.). «Активация Raf-киназы аденилатциклаз: изоформ-селективная регуляция». Мол. Фармакол . 66 (4): 921–8. дои : 10.1124/моль.66.4.921 . ПМИД 15385642 . S2CID 9876605 .
- ^ Брустас К.Г., Грамматикакис Н., Это М., Дент П., Бротиган Д.Л., Касид У. (январь 2002 г.). «Фосфорилирование миозинсвязывающей субъединицы миозинфосфатазы с помощью Raf-1 и ингибирование активности фосфатазы» . Ж. Биол. Хим . 277 (4): 3053–9. дои : 10.1074/jbc.M106343200 . ПМИД 11719507 .
- ^ Пфлейдерер П., Сумандея М.П., Рыбин В.О., Ван С., Стейнберг С.Ф. (2009). «Raf-1: новая киназа сердечного тропонина Т» . Дж. Мышцы Res. Клетка. Мобильная . 30 (1–2): 67–72. дои : 10.1007/s10974-009-9176-y . ПМЦ 2893395 . ПМИД 19381846 .
- ^ Хиндли А., Колх В. (апрель 2002 г.). «Киназа, регулируемая внеклеточным сигналом (ERK) / митоген-активируемая протеинкиназа (MAPK), независимые функции Raf-киназ». Дж. Клеточная наука . 115 (Часть 8): 1575–81. дои : 10.1242/jcs.115.8.1575 . ПМИД 11950876 .
- ^ Кэтлинг А.Д., Шеффер Х.Дж., Рейтер К.В., Редди Г.Р., Вебер М.Дж. (октябрь 1995 г.). «Богатая пролином последовательность, уникальная для MEK1 и MEK2, необходима для связывания Raf и регулирует функцию MEK» . Мол. Клетка. Биол . 15 (10): 5214–25. дои : 10.1128/mcb.15.10.5214 . ПМК 230769 . ПМИД 7565670 .
- ^ Пандит Б, Саркози А, Пеннаккио Л.А., Карта С, Оиси К., Мартинелли С., Понья Э.А., Шаквиц В., Усташевска А, Ландстрем А., Бос Дж.М., Оммен С.Р., Эспозито Г., Лепри Ф, Фаул С., Мундел П., Лопес Сигеро ДжейПи, Тенкони Р, Селикорни А, Росси С, Маццанти Л, Торренте И, Марино Б, Диджилио MC, Зампино Дж., Акерман М.Дж., Даллапиккола Б., Тарталья М., Гельб Б.Д. (август 2007 г.). «Мутации усиления функции RAF1 вызывают синдромы Нунан и LEOPARD с гипертрофической кардиомиопатией». Нат. Жене . 39 (8): 1007–12. дои : 10.1038/ng2073 . ПМИД 17603483 . S2CID 19335210 .
- ^ Молзан М., Шумахер Б., Оттманн С., Бальюлс А., Полцин Л., Вейанд М., Тиль П., Роуз Р., Роуз М., Куэнне П., Кайзер М., Рапп У.Р., Кульманн Дж., Оттманн С. (октябрь 2010 г.). «Нарушение связывания 14-3-3 с C-RAF при синдроме Нунан предполагает новые подходы к заболеваниям с повышенной передачей сигналов Ras» . Мол. Клетка. Биол . 30 (19): 4698–711. дои : 10.1128/MCB.01636-09 . ПМЦ 2950525 . ПМИД 20679480 .
- ^ Шторм С.М., Рапп УР (апрель 1993 г.). «Активация онкогена: мутации гена c-raf-1 в экспериментальных и естественных опухолях» . Токсикол. Летт . 67 (1–3): 201–10. дои : 10.1016/0378-4274(93)90056-4 . ПМИД 8451761 .
- ^ Зебиш А., Стабер П.Б., Делавар А., Боднер С., Хиден К., Фишередер К., Джанакираман М., Линкеш В., Аунер Х.В., Эмбергер В., Виндпассингер С., Шимек М.Г., Хёфлер Г., Троппмайр Дж., Силл Х. (апрель 2006 г.). «Две трансформирующие мутации зародышевой линии C-RAF, выявленные у пациентов с острым миелоидным лейкозом, связанным с терапией» . Рак Рез . 66 (7): 3401–8. дои : 10.1158/0008-5472.CAN-05-0115 . ПМИД 16585161 .
- ^ Эмусс В., Гарнетт М., Мейсон С., Марэ Р. (ноябрь 2005 г.). «Мутации C-RAF редки при раке человека, поскольку C-RAF имеет низкую базальную киназную активность по сравнению с B-RAF» . Рак Рез . 65 (21): 9719–26. дои : 10.1158/0008-5472.CAN-05-1683 . ПМИД 16266992 .
- ^ Форбс С.А., Биндал Н., Бэмфорд С., Коул С., Кок С.И., Беар Д., Цзя М., Шепард Р., Люнг К., Мензис А., Тиг Дж.В., Кэмпбелл П.Дж., Страттон М.Р., Futreal PA (январь 2011 г.). «COSMIC: исследование полных раковых геномов в Каталоге соматических мутаций рака» . Нуклеиновые кислоты Рез . 39 (Проблема с базой данных): D945–50. дои : 10.1093/nar/gkq929 . ПМК 3013785 . ПМИД 20952405 .
- ^ Каппер Д., Бергхофф А.С., Магерле М., Ильхан А., Верер А., Хакль М., Пихлер Дж., Пуш С., Мейер Дж., Хабель А., Петцельбауэр П., Бирнер П., фон Даймлинг А., Пройссер М. (2012). «Иммуногистохимическое тестирование статуса BRAF V600E в 1120 образцах опухолевой ткани пациентов с метастазами в головной мозг». Акта Нейропатол . 123 (2): 223–33. дои : 10.1007/s00401-011-0887-y . ПМИД 22012135 . S2CID 35623183 .
- ^ Каппер Д., Пройссер М., Хабель А., Сам Ф., Аккерман У., Шиндлер Г., Пуш С., Мехтершаймер Г., Зентграф Х., фон Даймлинг А. (2011). «Оценка статуса мутации BRAF V600E методом иммуногистохимии с использованием моноклонального антитела, специфичного для мутации». Акта Нейропатол . 122 (1): 11–9. дои : 10.1007/s00401-011-0841-z . ПМИД 21638088 . S2CID 25647782 .
- ^ Тран Н.Х., Ву X, Фрост Дж.А. (апрель 2005 г.). «B-Raf и Raf-1 регулируются разными механизмами авторегуляции» . Ж. Биол. Хим . 280 (16): 16244–53. дои : 10.1074/jbc.M501185200 . ПМИД 15710605 .
- ^ Гарнетт М.Дж., Рана С., Патерсон Х., Барфорд Д., Марэ Р. (декабрь 2005 г.). «Дикий тип и мутантный B-RAF активируют C-RAF посредством различных механизмов, включающих гетеродимеризацию» . Мол. Клетка . 20 (6): 963–9. doi : 10.1016/j.molcel.2005.10.022 . ПМИД 16364920 .
- ^ Маурер Г., Тарковски Б., Баккарини М. (август 2011 г.). «Роль киназ Raf в раке и терапевтические возможности» . Онкоген . 30 (32): 3477–88. дои : 10.1038/onc.2011.160 . ПМИД 21577205 .
- ^ Ким Д.Х., Сим Т. (март 2012 г.). «Новые низкомолекулярные ингибиторы киназы Raf для таргетной терапии рака». Арх. Фарм. Рез . 35 (4): 605–15. дои : 10.1007/s12272-012-0403-5 . ПМИД 22553052 . S2CID 26714490 .
- ^ Циммерманн С., Мёллинг К. (ноябрь 1999 г.). «Фосфорилирование и регуляция Raf с помощью Akt (протеинкиназа B)». Наука . 286 (5445): 1741–4. дои : 10.1126/science.286.5445.1741 . ПМИД 10576742 .
- ^ Чен Дж., Фуджи К., Чжан Л., Робертс Т., Фу Х. (июль 2001 г.). «Raf-1 способствует выживанию клеток путем противодействия киназы 1, регулирующей сигнал апоптоза, посредством независимого от MEK-ERK механизма» . Учеб. Натл. акад. наук. США . 98 (14): 7783–8. Бибкод : 2001PNAS...98.7783C . дои : 10.1073/pnas.141224398 . ПМЦ 35419 . ПМИД 11427728 .
- ^ Ван Х.Г., Такаяма С., Рапп У.Р., Рид Дж.К. (июль 1996 г.). «Белок, взаимодействующий с Bcl-2, BAG-1, связывается с киназой Raf-1 и активирует ее» . Учеб. Натл. акад. наук. США . 93 (14): 7063–8. Бибкод : 1996PNAS...93.7063W . дои : 10.1073/pnas.93.14.7063 . ПМК 38936 . ПМИД 8692945 .
- ^ Вебер С.К., Слупски-младший, Калмес Х.А., Рапп УР (май 2001 г.). «Активный Ras индуцирует гетеродимеризацию cRaf и BRaf». Рак Рез . 61 (9): 3595–8. ПМИД 11325826 .
- ^ Ван Х.Г., Рапп У.Р., Рид Дж.К. (ноябрь 1996 г.). «Bcl-2 нацеливает протеинкиназу Raf-1 на митохондрии» . Клетка . 87 (4): 629–38. дои : 10.1016/s0092-8674(00)81383-5 . ПМИД 8929532 . S2CID 16559750 .
- ^ Галактионов К., Джессус С., Бич Д. (май 1995 г.). «Взаимодействие Raf1 с фосфатазой Cdc25 связывает передачу митогенного сигнала с активацией клеточного цикла» . Генс Дев . 9 (9): 1046–58. дои : 10.1101/gad.9.9.1046 . ПМИД 7744247 .
- ^ Хуан Т.С., Шу Ч., Ян В.К., Ван-Пэн Дж. (июль 1997 г.). «Активация фосфатазы CDC 25 и киназы CDC 2, участвующих в GL331-индуцированном апоптозе». Рак Рез . 57 (14): 2974–8. ПМИД 9230211 .
- ^ Катаока Т., Бадд Р.К., Холлер Н., Том М., Мартинон Ф., Ирмлер М., Бернс К., Хане М., Кеннеди Н., Ковачович М., Чопп Дж. (июнь 2000 г.). «Ингибитор каспазы-8 FLIP способствует активации сигнальных путей NF-kappaB и Erk» . Курс. Биол . 10 (11): 640–8. Бибкод : 2000CBio...10..640K . дои : 10.1016/s0960-9822(00)00512-1 . ПМИД 10837247 . S2CID 14819939 .
- ^ Перейти обратно: а б Клегон В., Моррисон Д.К. (июль 1994 г.). «Raf-1 взаимодействует с Fyn и Src не фосфотирозин-зависимым образом» . Ж. Биол. Хим . 269 (26): 17749–55. дои : 10.1016/S0021-9258(17)32504-8 . ПМИД 7517401 .
- ^ Нантель А., Хубер М., Томас Д.Ю. (декабрь 1999 г.). «Локализация эндогенного Grb10 в митохондриях и его взаимодействие с митохондриально-ассоциированным пулом Raf-1» . Ж. Биол. Хим . 274 (50): 35719–24. дои : 10.1074/jbc.274.50.35719 . ПМИД 10585452 .
- ^ Нантель А., Мохаммад-Али К., Шерк Дж., Познер Б.И., Томас Д.Ю. (апрель 1998 г.). «Взаимодействие адаптерного белка Grb10 с киназами Raf1 и MEK1» . Ж. Биол. Хим . 273 (17): 10475–84. дои : 10.1074/jbc.273.17.10475 . ПМИД 9553107 .
- ^ Стэнг С., Ботторфф Д., Стоун Дж.К. (июнь 1997 г.). «Взаимодействие активированного Ras только с Raf-1 может быть достаточным для трансформации клеток rat2» . Мол. Клетка. Биол . 17 (6): 3047–55. дои : 10.1128/MCB.17.6.3047 . ПМК 232157 . ПМИД 9154803 .
- ^ Джермани А., Прабэль А., Мура С., Подгорняк М.П., Ди Карло А., Эрлих Р., Гиссельбрехт С., Варин-Бланк Н., Кальво Ф., Брузцони-Джованелли Х. (декабрь 2003 г.). «SIAH-1 взаимодействует с CtIP и способствует его деградации по протеасомному пути» . Онкоген . 22 (55): 8845–51. дои : 10.1038/sj.onc.1206994 . ПМИД 14654780 .
- ^ Митин Н.Ю., Рамоцкий М.Б., Зулло А.Дж., Дер С.Дж., Конечный С.Ф., Тапаровский Э.Дж. (май 2004 г.). «Идентификация и характеристика дождя, нового Ras-взаимодействующего белка с уникальной субклеточной локализацией» . Ж. Биол. Хим . 279 (21): 22353–61. дои : 10.1074/jbc.M312867200 . ПМИД 15031288 .
- ^ Варджиу П., Де Абахо Р., Гарсия-Ранеа Х.А., Валенсия А., Сантистебан П., Креспо П., Берналь Дж. (январь 2004 г.). «Небольшой GTP-связывающий белок, Rhes, регулирует передачу сигнала от рецепторов, связанных с G-белком» . Онкоген . 23 (2): 559–68. дои : 10.1038/sj.onc.1207161 . ПМИД 14724584 .
- ^ Перейти обратно: а б Юрьев А., Венногле Л.П. (февраль 2003 г.). «Новые белок-белковые взаимодействия киназы raf, обнаруженные с помощью исчерпывающего двухгибридного анализа дрожжей». Геномика . 81 (2): 112–25. дои : 10.1016/s0888-7543(02)00008-3 . ПМИД 12620389 .
- ^ Перейти обратно: а б с д Ли В., Хан М., Гуань К.Л. (апрель 2000 г.). «Богатый лейцином повторяющийся белок SUR-8 усиливает активацию киназы MAP и образует комплекс с Ras и Raf» . Генс Дев . 14 (8): 895–900. дои : 10.1101/gad.14.8.895 . ПМК 316541 . ПМИД 10783161 .
- ^ Перейти обратно: а б Киёно М., Като Дж., Катаока Т., Казиро Ю., Сато Т. (сентябрь 2000 г.). «Стимуляция обменной активности гуаниновых нуклеотидов Ras-GRF1/CDC25(Mm) при фосфорилировании тирозина с помощью Cdc42-регулируемой киназы ACK1» . Ж. Биол. Хим . 275 (38): 29788–93. дои : 10.1074/jbc.M001378200 . ПМИД 10882715 .
- ^ Жануэ-Лерози I, Пашева Е, де Танд М.Ф., Тавитян А, де Гинцбург Ж (март 1998 г.). «Идентификация специфического эффектора малого ГТФ-связывающего белка Rap2» . Евро. Дж. Биохим . 252 (2): 290–8. дои : 10.1046/j.1432-1327.1998.2520290.x . ПМИД 9523700 .
- ^ Беттнер Б., Говек Э.Э., Кросс Дж., Ван Алст Л. (август 2000 г.). «Соединительный мультидоменный белок AF-6 является партнером по связыванию Rap1A GTPase и связывается с актиновым регулятором цитоскелета профилином» . Учеб. Натл. акад. наук. США . 97 (16): 9064–9. Бибкод : 2000PNAS...97.9064B . дои : 10.1073/pnas.97.16.9064 . ПМК 16822 . ПМИД 10922060 .
- ^ Карбовничек М., Робертсон Г.П., Хенске Е.П. (сентябрь 2006 г.). «Rheb ингибирует активность C-raf и гетеродимеризацию B-raf/C-raf» . Ж. Биол. Хим . 281 (35): 25447–56. дои : 10.1074/jbc.M605273200 . ПМИД 16803888 .
- ^ Перейти обратно: а б Хан Л., Количелли Дж. (март 1995 г.). «Человеческий белок, выбранный для вмешательства в функцию Ras, напрямую взаимодействует с Ras и конкурирует с Raf1» . Мол. Клетка. Биол . 15 (3): 1318–23. дои : 10.1128/mcb.15.3.1318 . ПМК 230355 . ПМИД 7862125 .
- ^ Елинек Т., Кэтлинг А.Д., Рейтер К.В., Муди С.А., Вольфман А., Вебер М.Дж. (декабрь 1994 г.). «РАС и РАФ-1 образуют сигнальный комплекс с МЕК-1, но не с МЕК-2» . Мол. Клетка. Биол . 14 (12): 8212–8. дои : 10.1128/mcb.14.12.8212 . ПМК 359360 . ПМИД 7969158 .
- ^ Ромеро Ф, Мартинес-А С, Камонис Х, Реболло А (июнь 1999 г.). «Фактор транскрипции Aiolos контролирует гибель клеток в Т-клетках, регулируя экспрессию Bcl-2 и его клеточную локализацию» . ЭМБО Дж . 18 (12): 3419–30. дои : 10.1093/emboj/18.12.3419 . ПМЦ 1171421 . ПМИД 10369681 .
- ^ Моркос П., Тапар Н., Тасним Н., Стейси Д., Таманой Ф. (май 1996 г.). «Идентификация мутантов нейрофибромина, которые проявляют аллельную специфичность или повышенное сродство к Ras, что приводит к подавлению активированных аллелей ras» . Мол. Клетка. Биол . 16 (5): 2496–503. дои : 10.1128/mcb.16.5.2496 . ПМК 231238 . ПМИД 8628317 .
- ^ Ху С.Д., Кария К., Тамада М., Акасака К., Ширузу М., Ёкояма С., Катаока Т. (декабрь 1995 г.). «Богатая цистеином область Raf-1 взаимодействует с активаторным доменом посттрансляционно модифицированного Ha-Ras» . Ж. Биол. Хим . 270 (51): 30274–7. дои : 10.1074/jbc.270.51.30274 . ПМИД 8530446 .
- ^ Родригес-Вициана П., Уорн П.Х., Ходжа А., Марте Б.М., Паппин Д., Дас П., Уотерфилд, доктор медицинских наук, Ридли А., Даунвард Дж. (май 1997 г.). «Роль фосфоинозитид-3-ОН-киназы в трансформации клеток и контроле актинового цитоскелета с помощью Ras» . Клетка . 89 (3): 457–67. дои : 10.1016/s0092-8674(00)80226-3 . ПМИД 9150145 . S2CID 14459536 .
- ^ Хуан Ю.З., Занг М., Сюн В.К., Луо З., Мэй Л. (январь 2003 г.). «Эрбин подавляет путь киназы MAP» . Ж. Биол. Хим . 278 (2): 1108–14. дои : 10.1074/jbc.M205413200 . ПМИД 12379659 .
- ^ Перейти обратно: а б Доган Т., Хармс Г.С., Хекман М., Карреман С., Оберой Т.К., Алнемри Э.С., Рапп У.Р., Раджалингам К. (декабрь 2008 г.). «Х-связанные и клеточные IAP модулируют стабильность киназы C-RAF и подвижность клеток». Нат. Клеточная Биол . 10 (12): 1447–55. дои : 10.1038/ncb1804 . ПМИД 19011619 . S2CID 6553549 .
- ^ Станкато Л.Ф., Чоу Ю.Х., Хатчисон К.А., Пердью Г.Х., Джоув Р., Пратт В.Б. (октябрь 1993 г.). «Raf существует в нативном гетерокомплексе с hsp90 и p50, который может быть восстановлен в бесклеточной системе» . Ж. Биол. Хим . 268 (29): 21711–6. дои : 10.1016/S0021-9258(20)80600-0 . ПМИД 8408024 .
- ^ Перейти обратно: а б с Юнг К., Янош П., Макферран Б., Роуз Д.В., Мишак Х., Седиви Дж.М., Колх В. (май 2000 г.). «Механизм подавления киназного пути, регулируемого Raf/MEK/внеклеточным сигналом, с помощью белка-ингибитора киназы Raf» . Мол. Клетка. Биол . 20 (9): 3079–85. дои : 10.1128/mcb.20.9.3079-3085.2000 . ПМК 85596 . ПМИД 10757792 .
- ^ Карандикар М., Сюй С., Кобб М.Х. (декабрь 2000 г.). «MEKK1 связывает raf-1 и компоненты каскада ERK2» . Ж. Биол. Хим . 275 (51): 40120–7. дои : 10.1074/jbc.M005926200 . ПМИД 10969079 .
- ^ Инглиш Дж. М., Пирсон Г., Хокенберри Т., Шивакумар Л., Уайт М.А., Кобб М.Х. (октябрь 1999 г.). «Вклад пути ERK5/MEK5 в передачу сигналов Ras/Raf и контроль роста» . Ж. Биол. Хим . 274 (44): 31588–92. дои : 10.1074/jbc.274.44.31588 . ПМИД 10531364 .
- ^ Кубоки Ю, Ито М, Такамацу Н, Ямамото К.И., Шиба Т., Ёсиока К. (декабрь 2000 г.). «Каркасный белок в NH2-концевых сигнальных путях киназы c-Jun подавляет сигнальные пути киназы, регулируемые внеклеточными сигналами» . Ж. Биол. Хим . 275 (51): 39815–8. дои : 10.1074/jbc.C000403200 . ПМИД 11044439 .
- ^ Ито М., Ёсиока К., Акэти М., Ямашита С., Такамацу Н., Сугияма К., Хиби М., Накабеппу Ю., Шиба Т., Ямамото К.И. (ноябрь 1999 г.). «JSAP1, новый jun N-концевой протеинкиназа (JNK), связывающий белок, который действует как каркасный фактор в сигнальном пути JNK» . Мол. Клетка. Биол . 19 (11): 7539–48. дои : 10.1128/mcb.19.11.7539 . ПМЦ 84763 . ПМИД 10523642 .
- ^ Занг М., Хейн С., Луо З. (февраль 2002 г.). «Взаимодействие между активным Pak1 и Raf-1 необходимо для фосфорилирования и активации Raf-1» . Ж. Биол. Хим . 277 (6): 4395–405. дои : 10.1074/jbc.M110000200 . ПМИД 11733498 .
- ^ Перейти обратно: а б Ван С., Нат Н., Фусаро Дж., Челлаппан С. (ноябрь 1999 г.). «Rb и запретин нацелены на отдельные области E2F1 для репрессии и отвечают на разные восходящие сигналы» . Мол. Клетка. Биол . 19 (11): 7447–60. дои : 10.1128/mcb.19.11.7447 . ПМЦ 84738 . ПМИД 10523633 .
- ^ Перейти обратно: а б с д и ж Ван дер Хувен PC, Ван дер Вал JC, Руурс П., Ван Дейк MC, Ван Блиттерсвейк Дж (январь 2000 г.). «Изотипы 14-3-3 облегчают связывание протеинкиназы C-дзета с Raf-1: негативная регуляция посредством фосфорилирования 14-3-3» . Биохим. Дж . 345 (2): 297–306. дои : 10.1042/0264-6021:3450297 . ПМК 1220759 . ПМИД 10620507 .
- ^ Ху С.Д., Кария К., Окада Т., Ци Икс, Сонг С., Катаока Т. (январь 1999 г.). «Влияние фосфорилирования на активность Rap1A по взаимодействию с Raf-1 и подавлению Ras-зависимой активации Raf-1» . Ж. Биол. Хим . 274 (1): 48–51. дои : 10.1074/jbc.274.1.48 . ПМИД 9867809 .
- ^ Окада Т., Ху С.Д., Джин Т.Г., Кария К., Ямаваки-Катаока Ю., Катаока Т. (сентябрь 1999 г.). «Важнейший фактор, определяющий реакцию Рафа на малые ГТФазы семейства Ras» . Мол. Клетка. Биол . 19 (9): 6057–64. дои : 10.1128/mcb.19.9.6057 . ПМЦ 84512 . ПМИД 10454553 .
- ^ Лонг Икс, Лин Ю, Ортис-Вега С, Ёнезава К, Авруч Дж (апрель 2005 г.). «Rheb связывает и регулирует киназу mTOR» . Курс. Биол . 15 (8): 702–13. Бибкод : 2005CBio...15..702L . дои : 10.1016/j.cub.2005.02.053 . ПМИД 15854902 . S2CID 3078706 .
- ^ Карбовничек М., Кэш Т., Чунг М., Робертсон Г.П., Астринидис А., Хенске Е.П. (июль 2004 г.). «Регуляция активности киназы B-Raf с помощью туберина и Rheb является независимой от рапамицина (mTOR) мишенью млекопитающих» . Ж. Биол. Хим . 279 (29): 29930–7. дои : 10.1074/jbc.M402591200 . ПМИД 15150271 .
- ^ Йи В.М., Уорли П.Ф. (февраль 1997 г.). «Rheb взаимодействует с киназой Raf-1 и может функционировать для интеграции сигналов, зависящих от фактора роста и протеинкиназы А» . Мол. Клетка. Биол . 17 (2): 921–33. дои : 10.1128/mcb.17.2.921 . ПМК 231818 . ПМИД 9001246 .
- ^ Мовилла Н., Креспо П., Бустело XR (октябрь 1999 г.). «Элементы сигнальной трансдукции TC21, онкогенного члена подсемейства R-Ras GTP-связывающих белков» . Онкоген . 18 (43): 5860–9. дои : 10.1038/sj.onc.1202968 . ПМИД 10557073 .
- ^ Перейти обратно: а б Ван С., Гош Р.Н., Челлаппан С.П. (декабрь 1998 г.). «Raf-1 физически взаимодействует с Rb и регулирует его функцию: связь между митогенной передачей сигналов и регуляцией клеточного цикла» . Мол. Клетка. Биол . 18 (12): 7487–98. дои : 10.1128/mcb.18.12.7487 . ПМК 109329 . ПМИД 9819434 .
- ^ Айрольди Э, Золло О, Маккиаруло А, Ди Марко Б, Маркетти С, Риккарди С (ноябрь 2002 г.). «Лейциновая застежка-молния, индуцированная глюкокортикоидами, ингибирует киназный путь, регулируемый внеклеточным сигналом Raf, путем связывания с Raf-1» . Мол. Клетка. Биол . 22 (22): 7929–41. дои : 10.1128/mcb.22.22.7929-7941.2002 . ПМК 134721 . ПМИД 12391160 .
- ^ Труонг А.Б., Masters SC, Ян Х, Фу Х (ноябрь 2002 г.). «Роль С-концевой петли 14-3-3 во взаимодействии лигандов». Белки . 49 (3): 321–5. дои : 10.1002/прот.10210 . ПМИД 12360521 . S2CID 31480274 .
- ^ Юрьев А, Оно М, Гофф С.А., Макалузо Ф, Венногле Л.П. (июль 2000 г.). «Изоформ-специфическая локализация A-RAF в митохондриях» . Мол. Клетка. Биол . 20 (13): 4870–8. дои : 10.1128/mcb.20.13.4870-4878.2000 . ПМК 85938 . ПМИД 10848612 .
- ^ Перейти обратно: а б с Винченц С., Диксит В.М. (август 1996 г.). «Белки 14-3-3 связываются с А20 специфичным для изоформ образом и функционируют как молекулы-шапероны и адаптеры» . Ж. Биол. Хим . 271 (33): 20029–34. дои : 10.1074/jbc.271.33.20029 . ПМИД 8702721 .
- ^ Перейти обратно: а б Конклин Д.С., Галактионов К., Бич Д. (август 1995 г.). «Белки 14-3-3 связываются с фосфатазами cdc25» . Учеб. Натл. акад. наук. США . 92 (17): 7892–6. Бибкод : 1995PNAS...92.7892C . дои : 10.1073/pnas.92.17.7892 . ПМК 41252 . ПМИД 7644510 .
- ^ Перейти обратно: а б Юинг Р.М., Чу П., Элизма Ф., Ли Х., Тейлор П., Клими С., МакБрум-Серажевски Л., Робинсон М.Д., О'Коннор Л., Ли М., Тейлор Р., Дарси М., Хо Ю, Хейлбут А., Мур Л., Чжан С., Орнацкий О, Бухман Ю.В., Этьер М., Шэн Ю., Василеску Дж., Абу-Фарха М., Ламберт Дж.П., Дьюэл Х.С., Стюарт II, Кюль Б., Хог К., Колвилл К., Гладвиш К., Маскат Б., Кинач Р., Адамс С.Л., Моран М.Ф., Морин ГБ, Топалоглу Т., Фигейс Д. (2007). «Крупномасштабное картирование белково-белковых взаимодействий человека методом масс-спектрометрии» . Мол. Сист. Биол . 3 (1): 89. дои : 10.1038/msb4100134 . ПМЦ 1847948 . ПМИД 17353931 .
- ^ Аутьери М.В., Карбоне CJ (июль 1999 г.). «14-3-3Гамма взаимодействует и фосфорилируется с помощью нескольких изоформ протеинкиназы C в стимулированных PDGF клетках сосудистых гладких мышц человека». ДНК Клеточная Биол . 18 (7): 555–64. дои : 10.1089/104454999315105 . ПМИД 10433554 .
- ^ Ичимура Т., Вакамия-Цурута А., Итагаки С., Таока М., Хаяно Т., Нацумэ Т., Исобе Т. (апрель 2002 г.). «Зависимое от фосфорилирования взаимодействие легкой цепи 2 кинезина и белка 14-3-3». Биохимия . 41 (17): 5566–72. дои : 10.1021/bi015946f . ПМИД 11969417 .
- ^ Лю Ю.К., Элли С., Йошида Х., Боннефой-Берар Н., Альтман А. (июнь 1996 г.). «Модулируемая активацией ассоциация белков 14-3-3 с Cbl в Т-клетках» . Ж. Биол. Хим . 271 (24): 14591–5. дои : 10.1074/jbc.271.24.14591 . ПМИД 8663231 .
- ^ Кларк Г.Дж., Друган Дж.К., Россман К.Л., Карпентер Дж.В., Роджерс-Грэм К., Фу Х., Дер СиДжей, Кэмпбелл С.Л. (август 1997 г.). «14-3-3-дзета отрицательно регулирует активность Raf-1 путем взаимодействия с богатым цистеином доменом Raf-1» . Ж. Биол. Хим . 272 (34): 20990–3. дои : 10.1074/jbc.272.34.20990 . ПМИД 9261098 .
- ^ Цивион Г., Луо З.Дж., Авруч Дж. (сентябрь 2000 г.). «Каликулин А-индуцированное фосфорилирование виментина секвестрирует 14-3-3 и вытесняет других партнеров 14-3-3 in vivo» . Ж. Биол. Хим . 275 (38): 29772–8. дои : 10.1074/jbc.M001207200 . ПМИД 10887173 .
- ^ Кояма С., Уильямс Л.Т., Кикучи А. (июль 1995 г.). «Характеристика взаимодействия Raf-1 с белком ras p21 или 14-3-3 в интактных клетках» . ФЭБС Летт . 368 (2): 321–5. дои : 10.1016/0014-5793(95)00686-4 . ПМИД 7628630 . S2CID 29625141 .
- ^ Чоу CW, Дэвис Р.Дж. (январь 2000 г.). «Интеграция сигнальных путей кальция и циклического АМФ с помощью 14-3-3» . Мол. Клетка. Биол . 20 (2): 702–12. дои : 10.1128/MCB.20.2.702-712.2000 . ПМЦ 85175 . ПМИД 10611249 .
Дальнейшее чтение
[ редактировать ]- Рид Дж.К., Чжа Х., Эме-Семпе С., Такаяма С., Ван Х.Г. (1996). «Структурно-функциональный анализ белков семейства BCL-2». Механизмы активации лимфоцитов и иммунной регуляции VI . Адв. Эксп. Мед. Биол. Том. 406. стр. 99–112. дои : 10.1007/978-1-4899-0274-0_10 . ISBN 978-1-4899-0276-4 . ПМИД 8910675 .
- Гейер М., Факлер О.Т., Петерлин Б.М. (2001). «Структурно-функциональные взаимоотношения при ВИЧ-1 Nef» . Представитель ЭМБО . 2 (7): 580–5. doi : 10.1093/embo-reports/kve141 . ПМЦ 1083955 . ПМИД 11463741 .
- Диллон А.С., Колх В. (2002). «Отмена регулирования киназы Raf-1». Арх. Биохим. Биофиз . 404 (1): 3–9. дои : 10.1016/S0003-9861(02)00244-8 . ПМИД 12127063 .
- Гринуэй А.Л., Холлоуэй Г., Макфи Д.А., Эллис П., Корналл А., Лидман М. (2004). «Контроль Nef ВИЧ-1 клеточных сигнальных молекул: несколько стратегий содействия репликации вируса». Дж. Биоски . 28 (3): 323–35. дои : 10.1007/BF02970151 . ПМИД 12734410 . S2CID 33749514 .
- Чен Х., Куннималайян М., Ван Гомпел Дж.Дж. (2006). «Медуллярный рак щитовидной железы: функции raf-1 и гомолога ахета-скута-1 человека». Щитовидная железа . 15 (6): 511–21. дои : 10.1089/thy.2005.15.511 . ПМИД 16029117 .
Внешние ссылки
[ редактировать ]- Запись GeneReviews/NCBI/NIH/UW о синдроме Нунан
- доменной структуры Диаграммы Raf-1, A-Raf и B-Raf.
- дрозофилы Отверстие для полюса - The Interactive Fly
- c-raf + Proteins в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Местоположение генома человека RAF1 и страница сведений о гене RAF1 в браузере генома UCSC .
- Обзор всей структурной информации, доступной в PDB для UniProt : P04049 (протоонкогенная серин/треонин-протеинкиназа RAF) в PDBe-KB .
галерея PDB |
|---|




