Цианобактерии
| Цианобактерии Временной диапазон: (Возможные палеоархейские записи) | |
|---|---|
 | |
| Изображение под микроскопом Cylindrospermum , нитчатого рода цианобактерий. | |
| Научная классификация | |
| Домен: | Бактерии |
| Клэйд : | Террабактерии |
| Клэйд : | Группа цианобактерий-мелайнабактерий |
| Тип: | Цианобактерии Станьер , 1973 год. |
| Сорт: | Цианофицеи |
| Заказы [3] | |
По состоянию на 2014 год [update] таксономия находилась на пересмотре [1] [2] | |
| Синонимы | |
Список | |
Цианобактерии ( / s aɪ ˌ æ n oʊ b æ k ˈ t ɪər i . ə / ), также называемые Cyanobacteriota или Cyanophyta , представляют собой автотрофных тип грамотрицательных бактерий. [4] которые могут получать биологическую энергию посредством кислородного фотосинтеза . Название «цианобактерии» (от древнегреческого κύανος ( куанос ) «синий») относится к их голубовато-зеленому ( циановому ) цвету. [5] [6] что лежит в основе неофициального общего названия цианобактерий — сине-зеленых водорослей . [7] [8] [9] хотя, будучи прокариотами, они с научной точки зрения не классифицируются как водоросли . [примечание 1]
Цианобактерии, вероятно, являются самым многочисленным таксоном , когда-либо существовавшим на Земле, и первыми известными организмами, производящими кислород . [10] появившись в среднем архее и зародившись, по-видимому, в пресноводной или земной среде . [11] Их фотопигменты могут поглощать частоты красного и синего спектра солнечного света (таким образом, отражая зеленоватый цвет), расщепляя молекулы воды на ионы водорода и кислорода. Ионы водорода используются для реакции с диоксидом углерода с образованием сложных органических соединений, таких как углеводы (процесс, известный как фиксация углерода ), а кислород выделяется в качестве побочного продукта . Считается, что непрерывно производя и выделяя кислород в течение миллиардов лет, цианобактерии превратили ранней Земли бескислородную, слабовосстанавливающую пребиотическую атмосферу в окислительную атмосферу со свободным газообразным кислородом (который ранее был немедленно удален различными поверхностными восстановителями ). что привело к Великому Событию окисления и « ржавлению Земли » в раннем протерозое . [12] резко изменил состав форм жизни на Земле. [13] Последующая адаптация ранних одноклеточных организмов к выживанию в кислородной среде, вероятно, привела к эндосимбиозу между анаэробами и аэробами и, следовательно, к эволюции эукариот в палеопротерозое .
Цианобактерии используют фотосинтетические пигменты, такие как различные формы хлорофилла , каротиноиды , фикобилины, для преобразования фотонной энергии солнечного света в химическую энергию . В отличие от гетеротрофных прокариот, цианобактерии имеют внутренние мембраны . Это уплощенные мешочки, называемые тилакоидами , в которых осуществляется фотосинтез. [14] [15] Фотоавтотрофные эукариоты, такие как красные , зеленые водоросли и растения, осуществляют фотосинтез в хлорофиллических органеллах , которые, как полагают, произошли от цианобактерий, приобретенных давным-давно посредством эндосимбиоза. Эти эндосимбионтные цианобактерии в эукариотах затем развились и дифференцировались в специализированные органеллы, такие как хлоропласты , хромопласты , этиопласты и лейкопласты , известные под общим названием пластиды .
Серицитохроматия, предложенное название парафилетической и самой базальной группы, является предком как нефотосинтетической группы мелаинабактерий , так и фотосинтезирующих цианобактерий, также называемых оксифотобактериями. [16]
Цианобактерии Synechocystis и Cyanothece являются важными модельными организмами с потенциальным применением в биотехнологии для производства биоэтанола , пищевых красителей, в качестве источника пищи для людей и животных, пищевых добавок и сырья. [17] Цианобактерии производят ряд токсинов, известных как цианотоксины , которые могут вызывать вредные последствия для здоровья людей и животных.
Обзор
[ редактировать ]
Цианобактерии — очень большой и разнообразный тип фотосинтезирующих прокариот . [19] Они характеризуются уникальной комбинацией пигментов и способностью осуществлять кислородный фотосинтез . Они часто живут колониальными скоплениями , которые могут принимать самые разные формы. [20] Особый интерес представляют нитчатые виды , которые часто доминируют в верхних слоях микробных матов, встречающихся в экстремальных условиях, таких как горячие источники , гиперсоленая вода , пустыни и полярные регионы. [21] но также широко распространены и в более обыденной среде. [22] Они эволюционно оптимизированы для условий окружающей среды с низким содержанием кислорода. [23] Некоторые виды являются азотфиксаторами и обитают в самых разных влажных почвах и воде либо свободно, либо в симбиотических отношениях с растениями или грибами лишайниковообразующими ( как, например, лишайники рода Peltigera ). [24]

Цианобактерии являются широко распространенными фотосинтезирующими прокариотами во всем мире и вносят основной вклад в глобальные биогеохимические циклы . [25] Они являются единственными оксигенными фотосинтезирующими прокариотами и процветают в разнообразных и экстремальных средах обитания. [26] Они являются одними из старейших организмов на Земле, окаменелости которых датируются не менее 2,1 миллиарда лет назад. [27] С тех пор цианобактерии стали важными игроками в экосистемах Земли. Планктонные цианобактерии являются фундаментальным компонентом морских пищевых сетей и вносят основной вклад в глобальные потоки углерода и азота . [28] [29] Некоторые цианобактерии образуют вредоносное цветение водорослей, вызывающее нарушение функций водных экосистем и интоксикацию дикой природы и человека за счет производства мощных токсинов ( цианотоксинов ), таких как микроцистины , сакситоксин и цилиндроспермопсин . [30] [31] В настоящее время цветение цианобактерий представляет серьезную угрозу для водной среды и здоровья населения, а его частота и масштабы растут во всем мире. [32] [25]
Цианобактерии повсеместно распространены в морской среде и играют важную роль в качестве первичных продуцентов . Они являются частью морского фитопланктона , который в настоящее время обеспечивает почти половину общей первичной продукции Земли. [33] Около 25% мировой первичной продукции морской среды приходится на цианобактерии. [34]
Среди цианобактерий лишь несколько линий колонизировали открытый океан: Crocosphaera и родственные ей, цианобактерии UCYN-A , Trichodesmium , а также Prochromococcus и Synechococcus . [35] [36] [37] [38] Из этих линий особенно важны азотфиксирующие цианобактерии, поскольку они контролируют первичную продуктивность и экспорт органического углерода в глубины океана. [35] путем преобразования газообразного азота в аммоний, который позже используется для производства аминокислот и белков. Морские пикоцианобактерии ( Prochromococcus и Synechococcus ) численно доминируют в большинстве сообществ фитопланктона в современных океанах, внося важный вклад в первичную продуктивность. [37] [38] [39] Хотя некоторые планктонные цианобактерии представляют собой одноклеточные и свободноживущие клетки (например, Crocosphaera , Prochromococcus , Synechococcus ); другие установили симбиотические отношения с гаптофитными водорослями , такими как кокколитофоры . [36] Среди нитчатых форм Trichodesmium являются свободноживущими и образуют агрегаты. Однако нитчатые гетероцистообразующие цианобактерии (например, Richelia , Calothrix ) встречаются в сочетании с такими диатомовыми водорослями , как Hemiaulus , Rhizosolenia и Chaetoceros . [40] [41] [42] [43]
Морские цианобактерии включают самые маленькие известные фотосинтезирующие организмы. Самый маленький из них, прохлорококк , имеет диаметр всего от 0,5 до 0,8 микрометра. [44] С точки зрения численности особей, Prochromococcus, возможно, является самым многочисленным родом на Земле: один миллилитр поверхностной морской воды может содержать 100 000 клеток этого рода и более. По оценкам, во всем мире насчитывается несколько октиллионов (10 27 , миллиард миллиардов миллиардов) особей. [45] Прохлорококк распространен повсеместно между 40° с.ш. и 40° ю.ш. и доминирует в олиготрофных (бедных питательными веществами) регионах океанов. [46] На долю бактерии приходится около 20% кислорода в атмосфере Земли. [47]
Морфология
[ редактировать ]Цианобактерии разнообразны по морфологии: от одноклеточных и нитчатых до колониальных форм . Нитчатые формы демонстрируют функциональную дифференцировку клеток, такую как гетероцисты (для фиксации азота), акинеты (клетки на стадии покоя) и гормогонии (репродуктивные, подвижные нити). Вместе с имеющимися у них межклеточными связями они считаются первыми признаками многоклеточности. [48] [49] [50] [25]
Многие цианобактерии образуют подвижные клеточные нити, называемые гормогониями , которые удаляются от основной биомассы, чтобы отпочковаться и сформировать новые колонии в другом месте. [51] [52] Клетки гормогония часто тоньше, чем в вегетативном состоянии, а клетки на обоих концах подвижной цепи могут сужаться. Чтобы оторваться от родительской колонии, гормогонию часто приходится разрывать более слабую клетку нити, называемую некридием.

масштабные линейки около 10 мкм


• Негетероцитарные : (c) Arthrospira maxima ,


Некоторые нитчатые виды могут дифференцироваться в несколько различных клеток типов :
- Вегетативные клетки – нормальные фотосинтезирующие клетки, образующиеся при благоприятных условиях выращивания.
- Акинеты - устойчивые к климату споры, которые могут образовываться, когда условия окружающей среды становятся суровыми.
- Толстостенные гетероцисты , содержащие фермент нитрогеназу, необходимый для фиксации азота. [54] [55] [56] в анаэробной среде из-за чувствительности к кислороду. [56]
Каждая отдельная клетка (каждая отдельная цианобактерия) обычно имеет толстую студенистую клеточную стенку . [57] У них отсутствуют жгутики , но гормогонии некоторых видов могут передвигаться, скользя по поверхности. [58] Многие многоклеточные нитчатые формы Oscillatoria способны к волнообразным движениям; нить колеблется взад и вперед. В толщах воды некоторые цианобактерии плавают, образуя газовые пузырьки , как у архей . [59] Эти везикулы не являются органеллами как таковыми. Они ограничены не липидными мембранами , а белковой оболочкой.
Фиксация азота
[ редактировать ]
Некоторые цианобактерии способны фиксировать атмосферный азот в анаэробных условиях с помощью специализированных клеток — гетероцист . [55] [56] Гетероцисты также могут образовываться в соответствующих условиях окружающей среды (бескислородных), когда фиксированного азота недостаточно. Виды, образующие гетероцисты, специализируются на фиксации азота и способны превращать газообразный азот в аммиак . NH 3 ), нитриты ( NO - 2 ) или нитраты ( NO - 3 ), которые могут усваиваться растениями и превращаться в белок и нуклеиновые кислоты (атмосферный азот не биодоступен для растений, за исключением имеющих эндосимбиотические азотфиксирующие бактерии , особенно семейства Fabaceae , среди другие).
Свободноживущие цианобактерии присутствуют в воде рисовых полей , а цианобактерии можно обнаружить в виде эпифитов на поверхности зеленой водоросли Чара , где они могут фиксировать азот. [60] Цианобактерии, такие как Anabaena (симбионт водного папоротника Azolla ), могут обеспечить рисовые плантации биоудобрением . [61]
Фотосинтез
[ редактировать ]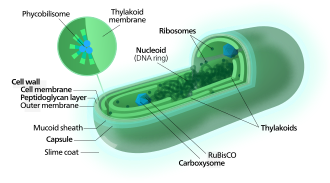

Фиксация углерода
[ редактировать ]Цианобактерии используют энергию солнечного света для фотосинтеза — процесса, при котором энергия света используется для синтеза органических соединений из углекислого газа. Поскольку они являются водными организмами, они обычно используют несколько стратегий, которые вместе известны как «механизм концентрации CO 2 », чтобы помочь в приобретении неорганического углерода (CO 2 или бикарбоната ). Среди более конкретных стратегий — широкое распространение бактериальных микрокомпартментов, известных как карбоксисомы . [63] которые взаимодействуют с активными переносчиками CO 2 и бикарбоната, с целью накопления бикарбоната в цитоплазме клетки. [64] Карбоксисомы представляют собой икосаэдрические структуры, состоящие из белков гексамерной оболочки, которые собираются в клеточные структуры, диаметр которых может достигать нескольких сотен нанометров. Считается, что эти структуры привязывают CO 2 фермент, фиксирующий , RuBisCO , к внутренней части оболочки, а также фермент карбоангидразу , используя метаболические каналы для увеличения локальных концентраций CO 2 и, таким образом, повышая эффективность фермента RuBisCO. . [65]
Электронный транспорт
[ редактировать ]В отличие от пурпурных бактерий и других бактерий, осуществляющих аноксигенный фотосинтез , тилакоидные мембраны цианобактерий не являются сплошными с плазматической мембраной, а представляют собой отдельные компартменты. [66] Фотосинтетический аппарат встроен в тилакоидные мембраны, при этом фикобилисомы действуют как светособирающие антенны, прикрепленные к мембране, что дает зеленую пигментацию, наблюдаемую (с длинами волн от 450 до 660 нм) у большинства цианобактерий. [67]
Хотя большая часть высокоэнергетических электронов, полученных из воды, используется клетками цианобактерий для собственных нужд, часть этих электронов может быть передана во внешнюю среду посредством электрогенной активности. [68]
Дыхание
[ редактировать ]Дыхание у цианобактерий может происходить в тилакоидной мембране наряду с фотосинтезом. [69] при этом их фотосинтетический транспорт электронов находится в том же отсеке, что и компоненты дыхательного транспорта электронов. В то время как цель фотосинтеза состоит в сохранении энергии путем создания углеводов из CO 2 , дыхание является обратным: углеводы снова превращаются в CO 2 , сопровождаясь высвобождением энергии.
Цианобактерии, по-видимому, разделяют эти два процесса, поскольку их плазматическая мембрана содержит только компоненты дыхательной цепи, в то время как в тилакоидной мембране находится взаимосвязанная дыхательная и фотосинтетическая цепь транспорта электронов. [69] Цианобактерии используют для дыхания электроны от сукцинатдегидрогеназы, а не от НАДФН . [69]
Цианобактерии дышат только ночью (или в темноте), поскольку на свету средства, используемые для транспорта электронов, используются наоборот для фотосинтеза. [70]
Электронно-транспортная цепь
[ редактировать ]Многие цианобактерии способны восстанавливать азот и углекислый газ в аэробных условиях, и этот факт может быть ответственен за их эволюционный и экологический успех. Водно-окисляющий фотосинтез осуществляется за счет сопряжения активности фотосистем (ФС) II и I ( Z-схема ). В отличие от зеленых серных бактерий , которые используют только одну фотосистему, использование воды в качестве донора электронов требует энергии и требует наличия двух фотосистем. [71]
Прикрепленные к тилакоидной мембране фикобилисомы действуют как светособирающие антенны для фотосистем. [72] Компоненты фикобилисом ( фикобилипротеины ) ответственны за сине-зеленую пигментацию большинства цианобактерий. [73] Вариации на эту тему обусловлены главным образом каротиноидами и фикоэритринами , которые придают клеткам красно-коричневатую окраску. У некоторых цианобактерий цвет света влияет на состав фикобилисом. [74] [75] При зеленом свете клетки накапливают больше фикоэритрина, который поглощает зеленый свет, тогда как при красном свете они производят больше фикоцианина , который поглощает красный свет. Таким образом, эти бактерии могут менять цвет от кирпично-красного до ярко-сине-зеленого в зависимости от того, подвергаются ли они воздействию зеленого или красного света. [76] Этот процесс «дополнительной хроматической адаптации» позволяет клеткам максимально использовать доступный свет для фотосинтеза.
У некоторых родов фикобилисомы отсутствуют, а вместо них есть хлорофилл b ( Prochromon , Prochromococcus , Prochromothrix ). Первоначально они были сгруппированы как прохлорофиты или хлороксибактерии, но, по-видимому, развились в несколько различных линий цианобактерий. По этой причине их теперь рассматривают как часть группы цианобактерий. [77] [78]
Метаболизм
[ редактировать ]В общем, фотосинтез у цианобактерий использует воду в качестве донора электронов и производит кислород в качестве побочного продукта, хотя некоторые могут также использовать сероводород. [79] процесс, который происходит среди других фотосинтезирующих бактерий, таких как пурпурные серобактерии .
Углекислый газ восстанавливается с образованием углеводов посредством цикла Кальвина . [80] Считается, что большое количество кислорода в атмосфере впервые появилось в результате деятельности древних цианобактерий. [81] Они часто встречаются в качестве симбионтов с рядом других групп организмов, таких как грибы (лишайники), кораллы , птеридофиты ( Азолла ), покрытосеменные ( Гуннера ) и др. [82] Углеродный обмен цианобактерий включает неполный цикл Кребса , [83] пентозофосфатный путь и гликолиз . [84]
Есть группы, способные к гетеротрофному росту. [85] в то время как другие являются паразитическими , вызывая заболевания у беспозвоночных или водорослей (например, болезнь черной полосы ). [86] [87] [88]
Экология
[ редактировать ]
Цианобактерии можно найти практически во всех наземных и водных средах обитания — океанах , пресной воде , влажной почве, временно увлажненных камнях в пустынях , голых камнях и почве и даже антарктических скалах. Они могут встречаться в виде планктонных клеток или образовывать фототрофные биопленки . Они встречаются внутри камней и ракушек (в эндолитных экосистемах ). [90] Некоторые из них являются лишайников , простейших растений, различных эндосимбионтами или губок и обеспечивают хозяина энергией . Некоторые живут в мехе ленивцев , обеспечивая им маскировку . [91]
Водные цианобактерии известны своим обширным и хорошо заметным цветением , которое может образовываться как в пресноводной , так и в морской среде. Цветки могут иметь вид сине-зеленой краски или накипи. Эти цветы могут быть токсичными и часто приводят к закрытию рекреационных водоёмов при их обнаружении. Морские бактериофаги являются важными паразитами одноклеточных морских цианобактерий. [92]
Росту цианобактерий благоприятствуют пруды и озера, где вода спокойна и не имеет бурного перемешивания. [93] Их жизненный цикл нарушается, когда вода естественным или искусственным образом смешивается из-за бурлящих течений, вызванных текущей водой ручьев или пенящейся водой фонтанов. По этой причине цветение цианобактерий в реках редко происходит, если вода не течет медленно. Росту также благоприятствуют более высокие температуры, которые позволяют видам Microcystis конкурировать с диатомовыми водорослями и зелеными водорослями и потенциально способствуют образованию токсинов. [93]
Основываясь на экологических тенденциях, модели и наблюдения предполагают, что цианобактерии, вероятно, будут увеличивать свое доминирование в водной среде. Это может привести к серьезным последствиям, в частности к загрязнению источников питьевой воды . Исследователи, в том числе Линда Лоутон из Университета Роберта Гордона , разработали методы их изучения. [94] Цианобактерии могут мешать очистке воды различными способами, в первую очередь засоряя фильтры (часто большие пласты песка и аналогичных сред) и производя цианотоксины , которые могут вызвать серьезные заболевания при употреблении в пищу. Последствия могут также касаться рыболовства и практики обращения с отходами. Антропогенная эвтрофикация , повышение температуры, вертикальная стратификация и увеличение содержания углекислого газа в атмосфере способствуют увеличению доминирования цианобактерий в водных экосистемах. [95]

Было обнаружено, что цианобактерии играют важную роль в наземной среде обитания и сообществах организмов. Широко сообщалось, что почвенные корки цианобактерий помогают стабилизировать почву, предотвращая эрозию и удерживая воду. [96] Примером вида цианобактерий, который делает это, является Microcoleus vaginatus . M. vaginatus стабилизирует почву, используя полисахаридную оболочку, которая связывается с частицами песка и поглощает воду. [97] M. vaginatus также вносит значительный вклад в сплоченность биологической почвенной корки . [98]
Некоторые из этих организмов вносят значительный вклад в глобальную экологию и кислородный цикл . Крошечная морская цианобактерия Prochromococcus была открыта в 1986 году и обеспечивает более половины фотосинтеза открытого океана. [99] циркадные ритмы Когда-то считалось, что существуют только в эукариотических клетках, но многие цианобактерии обладают бактериальным циркадным ритмом .
«Цианобактерии, возможно, являются самой успешной группой микроорганизмов на Земле. Они наиболее генетически разнообразны; они занимают широкий спектр сред обитания во всех широтах, широко распространены в пресноводных, морских и наземных экосистемах и встречаются в самых экстремальных нишах. такие как горячие источники, соляные заводы и гиперсоленые заливы. Фотоавтотрофные цианобактерии, производящие кислород, создали условия в ранней атмосфере планеты, которые направили эволюцию аэробного метаболизма и эукариотического фотосинтеза. Цианобактерии выполняют жизненно важные экологические функции в мировых океанах, внося важный вклад. к глобальным бюджетам углерода и азота». - Стюарт и Фальконер [100]
Цианобионты
[ редактировать ]
Заселение листьев и корней цианобактериями
(2) На поверхности корня цианобактерии демонстрируют два типа колонизации; в корневых волосках нити видов Anabaena и Nostoc образуют рыхлые колонии, а в ограниченной зоне на поверхности корня специфические виды Nostoc образуют колонии цианобактерий.
(3) Совместная инокуляция 2,4-D и Nostoc spp. увеличивает образование параклубеньков и фиксацию азота. Большое количество Nostoc spp. изоляты колонизируют корневую эндосферу и образуют параклубеньки. [101]
Некоторые цианобактерии, так называемые цианобионты (цианобактериальные симбионты), вступают в симбиотические отношения с другими организмами, как одноклеточными, так и многоклеточными. [102] Как показано справа, существует множество примеров симбиотического взаимодействия цианобактерий с наземными растениями . [103] [104] [105] [106] Цианобактерии могут проникать в растение через устьица и колонизировать межклеточное пространство, образуя петли и внутриклеточные клубки. [107] Анабаена виды. колонизировать корни растений пшеницы и хлопчатника. [108] [109] [110] Калотрикс сп. также был обнаружен в корневой системе пшеницы. [109] [110] Однодольные растения , такие как пшеница и рис, были заселены видами Nostoc . [111] [112] [113] [114] В 1991 году Гантер и другие выделили из корней растений и почвы разнообразные гетероцистные азотфиксирующие цианобактерии, в том числе Nostoc , Anabaena и Cylindrospermum . Оценка корней проростков пшеницы выявила два типа паттернов ассоциации: свободную колонизацию корневых волосков Anabaena и плотную колонизацию поверхности корня в ограниченной зоне Nostoc . [111] [101]

(а) O. Magnificus с многочисленными цианобионтами, присутствующими в верхних и нижних поясных списках (черные наконечники стрел) цингулума, называемого симбиотической камерой.
(б) O. steinii с многочисленными цианобионтами, обитающими в симбиотической камере.
(c) Увеличенная область на (b), показывающая двух цианобионтов, которые разделяются в результате бинарного поперечного деления (белые стрелки).

Особого внимания заслуживают взаимоотношения между цианобионтами (цианобактериальными симбионтами) и протистанами-хозяевами, поскольку некоторые азотфиксирующие цианобактерии ( диазотрофы ) играют важную роль в первичной продукции , особенно в азото-лимитированных олиготрофных океанах. [115] [116] [117] Цианобактерии, в основном размером с пико Synechococcus и Prochromococcus , распространены повсеместно и являются наиболее многочисленными фотосинтезирующими организмами на Земле, на их долю приходится четверть всего углерода, зафиксированного в морских экосистемах. [39] [118] [46] Известно, что в отличие от свободноживущих морских цианобактерий некоторые цианобионты ответственны за фиксацию азота, а не за фиксацию углерода в организме хозяина. [119] [120] Однако физиологические функции большинства цианобионтов остаются неизвестными. Цианобионты были обнаружены в многочисленных группах протистов, включая динофлагелляты , тинтинниды , радиолярии , амебы , диатомовые водоросли и гаптофиты . [121] [122] Среди этих цианобионтов мало что известно о природе (например, генетическом разнообразии, специфичности хозяина или цианобионта и сезонности цианобионта) вовлеченного симбиоза, особенно в отношении хозяина-динофлагелляты. [102]
Коллективное поведение
[ редактировать ]
Некоторые цианобактерии, даже одноклеточные, демонстрируют поразительное коллективное поведение и образуют колонии (или цветки ), которые могут плавать в воде и играть важную экологическую роль. Например, миллиарды лет назад сообщества морских палеопротерозойских цианобактерий могли помочь создать биосферу , какой мы ее знаем, похоронив соединения углерода и позволив первоначальному накоплению кислорода в атмосфере. [124] С другой стороны, ядовитое цветение цианобактерий становится все более серьезной проблемой для общества, поскольку их токсины могут быть вредными для животных. [32] Экстремальное цветение также может привести к истощению воды кислородом и уменьшению проникновения солнечного света и видимости, тем самым ставя под угрозу пищевое и спаривающееся поведение светозависимых видов. [123]
Как показано на диаграмме справа, бактерии могут оставаться во взвешенном состоянии в виде отдельных клеток, коллективно прилипать к поверхностям, образуя биопленки, пассивно оседать или флокулировать, образуя взвешенные агрегаты. Цианобактерии способны производить сульфатированные полисахариды (желтая дымка, окружающая скопления клеток), которые позволяют им образовывать плавающие агрегаты. В 2021 году Маэда и др. обнаружили, что кислород, вырабатываемый цианобактериями, захватывается сетью полисахаридов и клеток, позволяя микроорганизмам образовывать плавучие цветки. [125] Считается, что специфические белковые волокна, известные как пили (представленные в виде линий, расходящихся от клеток), могут действовать как дополнительный способ связывания клеток друг с другом или с поверхностями. Некоторые цианобактерии также используют сложные внутриклеточные газовые везикулы в качестве средства плавучести. [123]


На диаграмме слева выше показана предлагаемая модель микробного распределения, пространственной организации, круговорота углерода и O 2 в скоплениях и прилегающих территориях. (а) Комки содержат более плотные нити цианобактерий и гетеротрофные микробы. Первоначальные различия в плотности зависят от подвижности цианобактерий и могут быть установлены за короткие промежутки времени. Более темный синий цвет за пределами комка указывает на более высокую концентрацию кислорода в областях, прилегающих к комкам. Кислородная среда увеличивает частоту разворота любых нитей, которые начинают покидать комки, тем самым уменьшая чистую миграцию от комков. Это обеспечивает сохранение первоначальных группировок в течение коротких периодов времени; (б) Пространственная связь между фотосинтезом и дыханием в сгустках. Кислород, вырабатываемый цианобактериями, диффундирует в вышележащую среду или используется для аэробного дыхания. Растворенный неорганический углерод (DIC) диффундирует в комок из вышележащей среды, а также образуется внутри комка при дыхании. В кислородных растворах высокое содержание O Концентрации 2 снижают эффективность фиксации СО 2 и приводят к выведению гликолата. В этих условиях слипание может быть полезным для цианобактерий, если оно стимулирует удержание углерода и ассимиляцию неорганического углерода цианобактериями внутри сгустков. Этот эффект, по-видимому, способствует накоплению частиц органического углерода (клеток, оболочек и гетеротрофных организмов) в сгустках. [126]
Пока неясно, почему и как цианобактерии образуют сообщества. Агрегация должна отвлекать ресурсы от основного бизнеса по созданию большего количества цианобактерий, поскольку обычно она предполагает производство обильных количеств внеклеточного материала. Кроме того, клетки в центре плотных агрегатов также могут страдать как от затенения, так и от нехватки питательных веществ. [127] [128] Так какую же пользу цианобактериям приносит такая совместная жизнь? [123]

Новое понимание того, как цианобактерии формируют цветение, было получено в результате исследования цианобактерии Synechocystis , проведенного в 2021 году . Они используют набор генов, которые регулируют производство и экспорт сульфатированных полисахаридов — цепочек молекул сахара, модифицированных сульфатными группами, которые часто можно найти в морских водорослях и тканях животных. Многие бактерии производят внеклеточные полисахариды, но сульфатированные обнаружены только у цианобактерий. У Synechocystis эти сульфатированные полисахариды помогают цианобактериям образовывать плавучие агрегаты, улавливая пузырьки кислорода в слизистой сети клеток и полисахаридов. [125] [123]
Предыдущие исследования Synechocystis показали, что пили IV типа , украшающие поверхность цианобактерий, также играют роль в формировании цветков. [130] [127] Эти втягивающиеся и клейкие белковые волокна важны для подвижности, адгезии к субстратам и поглощения ДНК. [131] Для формирования цветков могут потребоваться как пили IV типа, так и синехан – например, пили могут способствовать экспорту полисахарида за пределы клетки. Действительно, активность этих белковых волокон может быть связана с продукцией внеклеточных полисахаридов у нитчатых цианобактерий. [132] Более очевидным ответом было бы то, что пили помогают строить агрегаты, связывая клетки друг с другом или с внеклеточным полисахаридом. Как и другие виды бактерий, [133] определенные компоненты пилей могут позволить цианобактериям одного и того же вида узнавать друг друга и устанавливать первоначальные контакты, которые затем стабилизируются за счет создания массы внеклеточного полисахарида. [123]
Механизм пузырьковой флотации, установленный Maeda et al. объединяет ряд известных стратегий, позволяющих цианобактериям контролировать свою плавучесть, таких как использование газовых везикул или накопление углеводных балластов. [134] Пили типа IV сами по себе также могут контролировать положение морских цианобактерий в толще воды, регулируя вязкое сопротивление. [135] Внеклеточный полисахарид, по-видимому, является многоцелевым активом для цианобактерий: от плавучего устройства до хранения пищи, защитного механизма и средства передвижения. [132] [123]
Клеточная смерть
[ редактировать ]
Одним из наиболее важных процессов, определяющих экофизиологию цианобактерий, является клеточная смерть . Имеющиеся данные подтверждают существование контролируемой гибели клеток у цианобактерий, а различные формы гибели клеток были описаны как реакция на биотические и абиотические стрессы. Однако исследования клеточной гибели цианобактерий — относительно молодая область, и понимание основных механизмов и молекулярных механизмов, лежащих в основе этого фундаментального процесса, остается во многом неуловимым. [25] Однако сообщения о гибели клеток морских и пресноводных цианобактерий указывают на то, что этот процесс имеет серьезные последствия для экологии микробных сообществ. [137] [138] [139] [140] Различные формы гибели клеток наблюдались у цианобактерий при нескольких стрессовых условиях. [141] [142] и смерть клеток, как предполагается, играет ключевую роль в процессах развития, таких как дифференцировка акинет и гетероцист, а также в стратегии выживания популяции. [136] [143] [144] [48] [25]
Цианофаги
[ редактировать ]

Цианофаги – это вирусы, поражающие цианобактерии. Цианофаги можно найти как в пресноводной, так и в морской среде. [145] Морские и пресноводные цианофаги имеют икосаэдрические головки, содержащие двухцепочечную ДНК, прикрепленную к хвосту соединительными белками. [146] Размер головы и хвоста различается у разных видов цианофагов. Цианофаги, как и другие бактериофаги полагаются на броуновское движение , при столкновении с бактериями , а затем используют белки, связывающие рецепторы, для распознавания белков клеточной поверхности, что приводит к прилипанию. Вирусы с сократительными хвостами затем полагаются на рецепторы, обнаруженные на их хвостах, для распознавания высококонсервативных белков на поверхности клетки-хозяина. [147]
Цианофаги заражают широкий спектр цианобактерий и являются ключевыми регуляторами популяций цианобактерий в водной среде, а также могут способствовать предотвращению цветения цианобактерий в пресноводных и морских экосистемах. Это цветение может представлять опасность для людей и других животных, особенно в эвтрофных пресноводных озерах. Инфицирование этими вирусами широко распространено в клетках, принадлежащих Synechococcus spp. в морской среде, где, как сообщается, до 5% клеток, принадлежащих морским цианобактериям, содержат зрелые фаговые частицы. [148]
Первый цианофаг ЛПП-1 был открыт в 1963 году. [149] Цианофаги классифицируются внутри бактериофагов семейств Myoviridae (например, AS-1 , N-1 ), Podoviridae (например, LPP-1) и Siphoviridae (например, S-1 ). [149]
Движение
[ редактировать ]
Давно известно, что нитчатые цианобактерии совершают поверхностные движения и что эти движения являются результатом пилей IV типа . [150] [132] [151] Кроме того, известно, что Synechococcus , морская цианобактерия, плавает со скоростью 25 мкм/с по механизму, отличному от механизма бактериальных жгутиков. [152] Считается, что образование волн на поверхности цианобактерий отталкивает окружающую воду назад. [153] [154] Известно, что клетки подвижны методом скольжения. [155] и новый нехарактерный нефототаксический метод плавания. [156] это не связано с движением жгутиков.
Многие виды цианобактерий способны планировать. Скольжение — это форма движения клеток, которая отличается от ползания или плавания тем, что не зависит от какого-либо очевидного внешнего органа или изменения формы клетки и происходит только в присутствии субстрата . [157] [158] Скольжение нитчатых цианобактерий, по-видимому, осуществляется за счет механизма «струи слизи», при котором клетки выдавливают гель, который быстро расширяется по мере гидратации, создавая движущую силу. [159] [160] хотя некоторые одноклеточные цианобактерии используют пили IV типа для скольжения. [161] [22]
Цианобактерии предъявляют строгие требования к свету. Слишком мало света может привести к недостаточному производству энергии, а у некоторых видов может привести к тому, что клетки прибегнут к гетеротрофному дыханию. [21] Слишком много света может подавлять клетки, снижать эффективность фотосинтеза и вызывать повреждение в результате обесцвечивания. УФ-излучение особенно смертельно опасно для цианобактерий, при этом нормальный уровень солнечной радиации в некоторых случаях оказывается для этих микроорганизмов значительно вредным. [20] [162] [22]
Нитчатые цианобактерии, живущие в микробных матах, часто мигрируют внутри мата вертикально и горизонтально, чтобы найти оптимальную нишу, которая уравновешивает их потребности в свете для фотосинтеза и их чувствительность к фотоповреждениям. Например, нитчатые цианобактерии Oscillatoria sp. и Spirulina subsalsa, обнаруженные в гиперсоленых донных матах Герреро-Негро , Мексика, в течение дня мигрируют вниз, в нижние слои, чтобы избежать интенсивного солнечного света, а затем поднимаются на поверхность в сумерках. [163] Напротив, популяция Microcoleus chthonoplastes, обнаруженная в гиперсоленых матах в Камарге , Франция, мигрирует в верхний слой мата в течение дня и равномерно распределяется по мату ночью. [164] Эксперимент in vitro с использованием Phormidium uncinatum также продемонстрировал склонность этого вида мигрировать, чтобы избежать вредного воздействия радиации. [20] [162] Эти миграции обычно являются результатом какого-то фотодвижения, хотя свою роль могут сыграть и другие формы такси. [165] [22]
Фотодвижение – модуляция движения клеток в зависимости от падающего света – используется цианобактериями как средство поиска оптимальных условий освещения в окружающей среде. Существует три типа фотодвижения: фотокинез, фототаксис и фотофобные реакции. [166] [167] [168] [22]
Фотокинетические микроорганизмы модулируют скорость своего скольжения в зависимости от интенсивности падающего света. Например, скорость, с которой скользит Phormidium Autumnale, линейно увеличивается с интенсивностью падающего света. [169] [22]
Фототаксические микроорганизмы движутся в соответствии с направлением света в окружающей среде, так что положительно фототаксические виды будут иметь тенденцию двигаться примерно параллельно свету и к источнику света. Такие виды, как Phormidium uncinatum, не могут двигаться прямо к свету, а полагаются на случайные столкновения, чтобы сориентироваться в правильном направлении, после чего они стремятся больше двигаться к источнику света. Другие, такие как Anabaena variabilis , могут управлять путем сгибания трихомы. [170] [22]
Наконец, фотофобные микроорганизмы реагируют на пространственные и временные градиенты света. Усиливающаяся фотофобная реакция возникает, когда организм входит в более яркую область поля из более темной, а затем меняет направление, избегая таким образом яркого света. Противоположная реакция, называемая реакцией понижения, возникает, когда организм входит в темную область из яркой области, а затем меняет направление, оставаясь, таким образом, на свету. [22]
Эволюция
[ редактировать ]История Земли
[ редактировать ]−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||||||||||||||||||||
Строматолиты — слоистые биохимические аккреционные структуры, образующиеся на мелководье путем захвата, связывания и цементации осадочных зерен биопленками ( микробными матами ) микроорганизмов , особенно цианобактерий. [171]
В докембрии строматолитовые сообщества микроорганизмов росли в большинстве морских и неморских сред фотической зоны . После кембрийского взрыва морских животных выпас травоядных животных на строматолитовых матах значительно сократил распространенность строматолитов в морской среде. С тех пор они встречаются в основном в гиперсоленых условиях, где не могут жить пасущиеся беспозвоночные (например, залив Шарк , Западная Австралия). Строматолиты предоставляют древние записи о жизни на Земле в виде ископаемых останков, датируемых 3,5 млрд лет назад. [172] Самые старые неоспоримые свидетельства существования цианобактерий датируются 2,1 млрд лет назад, но есть некоторые свидетельства их существования еще 2,7 млрд лет назад. [27] Цианобактерии также могли возникнуть 3,5 млрд лет назад. [173] Концентрация кислорода в атмосфере оставалась около 0,001% от сегодняшнего уровня или ниже до 2,4 млрд лет назад ( Великое событие оксигенации ). [174] Повышение содержания кислорода могло вызвать падение концентрации метана в атмосфере и спровоцировать гуронское оледенение примерно 2,4–2,1 млрд лет назад. Таким образом, цианобактерии, возможно, уничтожили большинство других бактерий того времени. [175]
Онколиты — это осадочные структуры , состоящие из онкоидов, которые представляют собой слоистые структуры, образованные в результате роста цианобактерий. Онколиты подобны строматолитам, но вместо того, чтобы образовывать колонны, они образуют примерно сферические структуры, не прикрепившиеся к подстилающему субстрату при своем формировании. [176] Онкоиды часто формируются вокруг центрального ядра, например фрагмента скорлупы. [177] а структура карбоната кальция образуется в результате инкрустации микробов . Онколиты являются индикаторами теплых вод в фотической зоне , но известны и в современной пресноводной среде. [178] Эти структуры редко превышают 10 см в диаметре.
Одна из прежних классификационных схем окаменелостей цианобактерий делила их на поростроматы и спонгиостромы . В настоящее время они признаны формальными таксонами и считаются таксономически устаревшими; однако некоторые авторы выступают за то, чтобы термины оставались неформальными для описания формы и структуры бактериальных окаменелостей. [179]
 Строматолиты, оставленные цианобактериями, являются старейшими известными окаменелостями жизни на Земле. Этому окаменелому один миллиард лет.
Строматолиты, оставленные цианобактериями, являются старейшими известными окаменелостями жизни на Земле. Этому окаменелому один миллиард лет.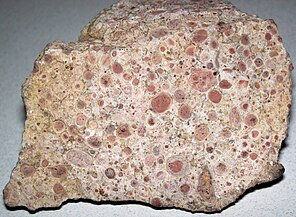 Онколитический известняк, образовавшийся из последовательных слоев карбоната кальция, осажденного цианобактериями.
Онколитический известняк, образовавшийся из последовательных слоев карбоната кальция, осажденного цианобактериями.
![Цианобактериальные остатки кольчатого трубчатого микрофоссилия Oscillatoriopsis longa [180] Масштабная линейка: 100 мкм.](//upload.wikimedia.org/wikipedia/commons/thumb/3/38/Oscillatoriopsis_longa_fossil.jpg/217px-Oscillatoriopsis_longa_fossil.jpg) Цианобактериальные остатки кольчатого трубчатого микрофоссилия Oscillatoriopsis longa [180]
Цианобактериальные остатки кольчатого трубчатого микрофоссилия Oscillatoriopsis longa [180]
Масштабная линейка: 100 мкм



![Цианобактериальные остатки кольчатого трубчатого микрофоссилия Oscillatoriopsis longa [180] Масштабная линейка: 100 мкм.](http://upload.wikimedia.org/wikipedia/commons/thumb/3/38/Oscillatoriopsis_longa_fossil.jpg/217px-Oscillatoriopsis_longa_fossil.jpg)
Происхождение фотосинтеза
[ редактировать ]Кислородный фотосинтез возник только один раз (у прокариотических цианобактерий), и все фотосинтезирующие эукариоты (включая все растения и водоросли ) приобрели эту способность в результате эндосимбиоза с цианобактериями или их эндосимбионтными хозяевами. Другими словами, весь кислород, который делает атмосферу пригодной для дыхания аэробных организмов, изначально поступает от цианобактерий или их пластидных потомков. [181]
Цианобактерии оставались основными первичными продуцентами на протяжении второй половины архея и , отчасти потому большей части протерозоя , что окислительно-восстановительная структура океанов благоприятствовала фотоавтотрофам, способным к фиксации азота . Однако утверждается, что их популяция значительно менялась на протяжении этого эона. [10] [182] [183] Археопластиды , такие как зеленые и красные водоросли, в конце концов превзошли цианобактерии в качестве основных первичных продуцентов на континентальных шельфах ближе к концу неопротерозоя , но только с мезозойской (251–65 млн лет назад) радиацией вторичных фотоавтотрофов, таких как динофлагелляты , кокколитофориды и диатомовые водоросли , началось первичное производство в морские шельфовые воды принимают современный вид. Цианобактерии по-прежнему имеют решающее значение для морских экосистем как первичные продуценты в океанических круговоротах, как агенты биологической фиксации азота и, в модифицированной форме, как пластиды морских водорослей . [184]
Происхождение хлоропластов
[ редактировать ]Первичные хлоропласты — это клеточные органеллы, обнаруженные у некоторых эукариотических линий, где они специализируются на фотосинтезе. Считается, что они произошли от эндосимбиотических цианобактерий. [185] [186] После нескольких лет дебатов, [187] В настоящее время общепринято, что три основные группы первичных эндосимбиотических эукариот (т.е. зеленые растения , красные водоросли и глаукофиты ) образуют одну большую монофилетическую группу , называемую Archaeplastida , которая развилась после одного уникального эндосимбиотического события. [188] [189] [190] [191]
О морфологическом сходстве хлоропластов и цианобактерий впервые сообщил немецкий ботаник Андреас Франц Вильгельм Шимпер в XIX веке. [192] Хлоропласты встречаются только у растений и водорослей . [193] таким образом, российский биолог Константин Мерешковский в 1905 году предположил симбиогенное происхождение пластид. [194] Линн Маргулис снова привлекла внимание к этой гипотезе более 60 лет спустя. [195] но эта идея не получила полного признания до тех пор, пока не начали накапливаться дополнительные данные. Цианобактериальное происхождение пластид в настоящее время подтверждается различными данными филогенетики . [196] [188] [191] геномный , [197] биохимический [198] [199] и структурные доказательства. [200] Описание другого независимого и более недавнего события первичного эндосимбиоза между цианобактерией и отдельной линией эукариот ( ризариев Paulinella chromatophora ) также подтверждает эндосимбиотическое происхождение пластид. [201]


В дополнение к этому первичному эндосимбиозу, многие эукариотические линии были подвержены вторичным или даже третичным эндосимбиотическим событиям , то есть « матрешкоподобному » поглощению эукариотом другого эукариота, несущего пластиды. [203] [185]
Хлоропласты имеют много общего с цианобактериями, включая кольцевую хромосому прокариотического типа , рибосомы и сходные белки в фотосинтетическом реакционном центре. [204] [205] Эндосимбиотическая теория предполагает, что фотосинтезирующие бактерии были приобретены (путем эндоцитоза ) ранними эукариотическими клетками и образовали первые растительные клетки. Следовательно, хлоропласты могут быть фотосинтезирующими бактериями, приспособившимися к жизни внутри растительных клеток. Как и митохондрии , хлоропласты по-прежнему обладают собственной ДНК, отдельной от ядерной ДНК клеток-хозяев растений, и гены в этой ДНК хлоропластов напоминают гены цианобактерий. [206] ДНК в хлоропластах кодирует окислительно-восстановительные белки, такие как фотосинтетические реакционные центры. Гипотеза CoRR предполагает, что такое совместное расположение необходимо для окислительно-восстановительной регуляции.
Морское происхождение
[ редактировать ]
| Часть серии о |
| Планктон |
|---|
 |
Цианобактерии коренным образом изменили геохимию планеты. [210] [207] Многочисленные геохимические данные подтверждают возникновение интервалов глубоких глобальных изменений окружающей среды в начале и конце протерозоя ( 2500–542 млн лет назад). [211] [212] [213] Хотя широко признано, что присутствие молекулярного кислорода в ранней летописи окаменелостей было результатом деятельности цианобактерий, мало что известно о том, как эволюция цианобактерий (например, предпочтение среды обитания) могла способствовать изменениям в биогеохимических циклах на протяжении истории Земли. произошел первый этап увеличения насыщения кислородом поверхности Земли, известный как Великое событие окисления (GOE). Геохимические данные показали, что в раннем палеопротерозое (2500–1600 млн лет назад) [210] [207] Второе, но гораздо более резкое повышение уровня кислорода, известное как неопротерозойское событие оксигенации (NOE), [212] [81] [214] произошло примерно от 800 до 500 млн лет назад. [213] [215] Последние данные по изотопам хрома указывают на низкие уровни атмосферного кислорода на поверхности Земли в середине протерозоя. [211] что согласуется с поздней эволюцией морских планктонных цианобактерий во время криогенного периода ; [216] оба типа доказательств помогают объяснить позднее появление и диверсификацию животных. [217] [43]
Понимание эволюции планктонных цианобактерий важно, поскольку их происхождение фундаментально изменило циклы азота и углерода к концу докембрия . [215] Однако остается неясным, какие эволюционные события привели к появлению планктонных форм открытого океана среди цианобактерий и как эти события связаны с геохимическими данными в докембрийском периоде. [212] На данный момент кажется, что геохимия океана (например, эвксинные условия в период раннего и среднего протерозоя) [212] [214] [218] и доступность питательных веществ [219] вероятно, способствовали очевидной задержке диверсификации и повсеместной колонизации среды открытого океана планктонными цианобактериями в неопротерозое . [215] [43]
Генетика
[ редактировать ]Цианобактерии способны к естественной генетической трансформации . [220] [221] [222] Естественная генетическая трансформация — это генетическое изменение клетки, возникающее в результате прямого поглощения и включения экзогенной ДНК из ее окружения. Чтобы произошла бактериальная трансформация, бактерии-реципиенты должны находиться в состоянии компетентности , которое может возникать в природе в ответ на такие условия, как голод, высокая плотность клеток или воздействие агентов, повреждающих ДНК. При хромосомной трансформации гомологичная трансформирующая ДНК может быть интегрирована в геном реципиента путем гомологичной рекомбинации , и этот процесс, по-видимому, является адаптацией для восстановления повреждений ДНК . [223]
восстановление ДНК
[ редактировать ]Цианобактерии подвергаются воздействию стрессов окружающей среды и образующихся внутри них активных форм кислорода , которые вызывают повреждение ДНК . Цианобактерии обладают многочисленными E. coli подобными репарации ДНК, генами . [224] Некоторые гены репарации ДНК высококонсервативны у цианобактерий, даже в небольших геномах , что позволяет предположить, что основные процессы репарации ДНК, такие как рекомбинационная репарация , эксцизионная репарация нуклеотидов и метил-направленная репарация несовпадений ДНК, распространены среди цианобактерий. [224]
Классификация
[ редактировать ]Филогения
[ редактировать ]| на основе 16S рРНК ДП _12_2021 г. [225] [226] [227] | GTDB 08-RS214 по базе данных таксономии генома [228] [229] [230] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
Таксономия
[ редактировать ]
Исторически бактерии были впервые отнесены к растениям, составляющим класс Schizomycetes, которые вместе с Schizophyceae (сине-зеленые водоросли/цианобактерии) образовали тип Schizophyta. [231] затем в типе Monera в царстве Protista Геккеля Protogens в 1866 году, включающем , Protamaeba, Vampyrella, Protomonae и Vibrio , но не Nostoc и другие цианобактерии, которые были отнесены к водорослям, [232] реклассифицирован как прокариоты Чаттоном позже . [233]
Цианобактерии традиционно разделяли по морфологии на пять отделов, обозначенных цифрами I – V. Первые три — Chroococcales , Pleurocapsales и Oscillatoriales — не подтверждены филогенетическими исследованиями. Последние два – Nostocales и Stigonematales – монофилетичны как единое целое и составляют гетероцистные цианобактерии. [234] [235]
Представители Chroococales одноклеточные и обычно собираются в колонии. Классическим таксономическим критерием была морфология клеток и плоскость клеточного деления. Клетки Pleurocapsales обладают способностью образовывать внутренние споры (баоциты). Остальные секции включают нитчатые виды. У Oscillatoriales клетки расположены однорядно и не образуют специализированных клеток (акинет и гетероцист). [236] У Nostocales и Stigonematales клетки обладают способностью в определенных условиях образовывать гетероцисты. Stigonematales, в отличие от Nostocales, включает виды с действительно разветвленными трихомами. [234]
Большинство таксонов, включенных в тип или подраздел Cyanobacteria, еще не были официально опубликованы в соответствии с Международным кодексом номенклатуры прокариот (ICNP), за исключением:
- Классы Chroobacteria , Hormogoneae и Gloeobacteria.
- Порядки Chroococcales , Gloeobacterales , Nostocales , Oscillatoriales , Pleurocapsales и Stigonematales.
- Семейства Прохлоровые и Прохлортриховые.
- Роды Halospirulina , Planktothricoides , Prochromococcus , Prochromon и Prochromothrix.
Остальные действительно опубликованы в соответствии с Международным кодексом номенклатуры водорослей, грибов и растений .
Раньше считалось, что некоторые бактерии, такие как Beggiatoa , являются бесцветными цианобактериями. [237]
Принятая в настоящее время таксономия основана на Списке названий прокариот, имеющих номенклатуру (LPSN). [238] и Национальный центр биотехнологической информации (NCBI). [239] Класс «Цианобактерии»
- Подкласс « Глоеобактерии » Кавальер-Смит, 2002 г.
- Gloeobacterales Кавальер-Смит 2002 г.
- Подкласс «Фикобактерии» Кавальер-Смит, 2002 г.
- Акариохлориды Мияшита и др. 2003 г. ex Струнецкий и Мареш 2022 г. [вкл. Термосинехококки]
- Эгеококки Струнецкий и Мареш 2022
- " Элайнеллалес "
- « Эврикоккал »
- Geitlerinematales Струнецкий и Мареш 2022
- Gloeomargaritales Moreira et al. 2016 год
- " Leptolyngbyales " Струнецкий и Мареш 2022
- Nodosilineales Струнецкий и Мареш 2022
- Окулателлалес Струнецкий и Мареш 2022
- « Формедесмиалы ».
- Proхлорококковые Комарек и Струнецкий 2020 {"PCC-6307"}
- Pseudanabaenales Хоффманн, Комарек и Кастовски 2005 г.
- « Псевдоформиалес ».
- Термостихалес Комарек и Струнецкий 2020
- Synechococcophycidae Хоффманн, Комарек и Кастовски 2005 г.
- « Лимнотрихалес »
- Прохлортрихалес Струнецкий и Мареш 2022 (PCC-9006)
- Synechococcales Хоффманн, Комарек и Кастовски, 2005 г.
- Nostocophycidae Хоффманн, Комарек и Кастовски 2005 г.
- Цианобактерии Риппка и Коэн-Базир 1983 ( Chamaesiphonales , Chroococcales , Chroococcidiopsidales , Nostocales , Oscillatoriales , Pleurocapsales , Spirulinales , Stigonematales )
Отношение к людям
[ редактировать ]Биотехнология
[ редактировать ]
Одноклеточная цианобактерия Synechocystis sp. PCC6803 был третьим прокариотом и первым фотосинтезирующим организмом, геном которого был полностью секвенирован . [240] Он продолжает оставаться важным модельным организмом. [241] Cyanothece ATCC 51142 — важный модельный диазотрофный организм. Самые маленькие геномы обнаружены у Prochromococcus spp. (1,7 Мб ) [242] [243] и самый большой — Nostoc punctiforme (9 Мб). [144] Представители Calothrix spp. оцениваются в 12–15 Мб, [244] размером с дрожжи .
Недавние исследования показали потенциальное применение цианобактерий для производства возобновляемой энергии путем прямого преобразования солнечного света в электричество. Внутренние пути фотосинтеза могут быть связаны с химическими медиаторами, которые переносят электроны на внешние электроды . [245] [246] В краткосрочной перспективе предпринимаются усилия по коммерциализации топлива на основе водорослей, такого как дизельное топливо , бензин и авиакеросин . [68] [247] [248] Цианобактерии также были созданы для производства этанола. [249] и эксперименты показали, что когда один или два гена CBB чрезмерно экспрессируются, выход может быть еще выше. [250] [251]
Цианобактерии могут обладать способностью производить вещества, которые однажды смогут служить противовоспалительными средствами и бороться с бактериальными инфекциями у людей. [252] Было продемонстрировано, что фотосинтетическая продукция сахара и кислорода цианобактериями имеет терапевтическую ценность у крыс с сердечными приступами. [253] Хотя цианобактерии могут естественным образом продуцировать различные вторичные метаболиты, они могут служить выгодными хозяевами для производства метаболитов растительного происхождения благодаря биотехнологическим достижениям в области системной биологии и синтетической биологии. [254]
Экстрагированный синий цвет спирулины используется в качестве натурального пищевого красителя. [255]
Исследователи из нескольких космических агентств утверждают, что цианобактерии могут быть использованы для производства товаров для потребления человеком в будущих пилотируемых аванпостах на Марсе путем преобразования материалов, доступных на этой планете. [256]
Питание человека
[ редактировать ]
Некоторые цианобактерии продаются в пищу, особенно Arthrospira Platensis ( Spirulina ) и другие ( Aphanizomenon flos-aquae ). [257]
Некоторые микроводоросли содержат вещества высокой биологической ценности, такие как полиненасыщенные жирные кислоты, аминокислоты, белки, пигменты, антиоксиданты, витамины и минералы. [258] Съедобные сине-зеленые водоросли снижают выработку провоспалительных цитокинов путем ингибирования пути NF-κB в макрофагах и спленоцитах. [259] Сульфатные полисахариды проявляют иммуномодулирующую, противоопухолевую, антитромботическую, антикоагулянтную, антимутагенную, противовоспалительную, противомикробную и даже противовирусную активность в отношении ВИЧ, герпеса и гепатита. [260]
Риски для здоровья
[ редактировать ]Некоторые цианобактерии могут продуцировать нейротоксины , цитотоксины , эндотоксины и гепатотоксины (например, продуцирующие микроцистин бактерии рода microcystis ), которые вместе известны как цианотоксины .
Конкретные токсины включают анатоксин-а , гуанитоксин , аплизиатоксин , цианопептолин, цилиндроспермопсин , домоевую кислоту , нодуларин R (из Nodularia ), неосакситоксин и сакситоксин . Цианобактерии при определенных условиях бурно размножаются. Это приводит к цветению водорослей , которое может нанести вред другим видам и представлять опасность для людей и животных, если вовлеченные цианобактерии производят токсины. Задокументировано несколько случаев отравления людей, но недостаток знаний не позволяет точно оценить риски. [261] [262] [263] [264] и исследование Линды Лоутон , FRSE в Университете Роберта Гордона , Абердин, и его соавторов на протяжении 30 лет изучает это явление и методы повышения безопасности воды. [265]
Недавние исследования показывают, что значительное воздействие высоких уровней цианобактерий, продуцирующих токсины, такие как BMAA, может вызвать боковой амиотрофический склероз (БАС). У людей, живущих в радиусе полумили от озер, загрязненных цианобактериями, риск развития БАС в 2,3 раза выше, чем у остального населения; в Нью-Гемпшире у жителей озера Маскома риск развития БАС был в 25 раз выше ожидаемого. [266] BMAA из корок пустынь, обнаруженных по всему Катару, возможно, способствовало более высокому уровню заболеваемости БАС у ветеранов войны в Персидском заливе . [262] [267]
Химический контроль
[ редактировать ]Некоторые химические вещества могут устранить цветение цианобактерий в небольших системах на водной основе, таких как плавательные бассейны. К ним относятся гипохлорит кальция , сульфат меди , куприцид (хелатная медь) и симазин . [268] Необходимое количество гипохлорита кальция варьируется в зависимости от цветения цианобактерий, поэтому обработка необходима периодически. По данным Министерства сельского хозяйства Австралии, для лечения цветения часто бывает достаточно 12 г 70% материала на 1000 л воды. [268] Сульфат меди также широко используется, но Министерство сельского хозяйства Австралии больше не рекомендует его, поскольку он убивает домашний скот, ракообразных и рыбу. [268] Куприцид — это хелатный продукт меди, который устраняет поседение с меньшим риском токсичности, чем сульфат меди. Рекомендации по дозировке варьируются от 190 мл до 4,8 л на 1000 м2. 2 . [268] Обработка железоалюминиевыми квасцами в дозе 50 мг/л уменьшит цветение водорослей. [268] [269] Симазин, который также является гербицидом, будет продолжать убивать цветы в течение нескольких дней после применения. Симазин продается в различной концентрации (25, 50 и 90%), рекомендуемое количество, необходимое на один кубометр воды на продукт, составляет 25% продукта на 8 мл; 50% продукт 4 мл; или 90% продукта 2,2 мл. [268]
Изменение климата
[ редактировать ]Изменение климата , вероятно, приведет к увеличению частоты, интенсивности и продолжительности цветения цианобактерий во многих эвтрофных озерах, водохранилищах и эстуариях. [270] [32] Цианобактерии, образующие цветение, производят различные нейротоксины , гепатотоксины и дерматоксины , которые могут быть смертельными для птиц и млекопитающих (включая водоплавающих птиц, крупный рогатый скот и собак) и угрожать использованию вод для отдыха, производства питьевой воды, сельскохозяйственного орошения и рыболовства. [32] Токсичные цианобактерии вызвали серьезные проблемы с качеством воды, например, в озере Тайху (Китай), озере Эри (США), озере Окичоби (США), озере Виктория (Африка) и Балтийском море . [32] [271] [272] [273]
Изменение климата прямо и косвенно благоприятствует цветению цианобактерий. [32] Многие цветущие цианобактерии могут расти при относительно высоких температурах. [274] Повышенная термическая стратификация озер и водохранилищ позволяет плавучим цианобактериям всплывать вверх и образовывать плотные поверхностные цветения, что дает им лучший доступ к свету и, следовательно, селективное преимущество перед неплавучими организмами фитопланктона. [275] [93] Длительные засухи летом увеличивают время пребывания воды в водохранилищах, реках и эстуариях, и эти застойные теплые воды могут обеспечить идеальные условия для развития цветения цианобактерий. [276] [273]
Способность вредных цианобактерий рода Microcystis адаптироваться к повышенному уровню CO 2 была продемонстрирована как в лабораторных, так и в полевых экспериментах. [277] виды Microcystis . поглощать CO 2 и HCO −
3 и накапливают неорганический углерод в карбоксисомах , а конкурентоспособность штамма зависит от концентрации неорганического углерода. В результате , что изменение климата и повышение уровня CO 2 повлияют на штаммовый состав цветения цианобактерий. ожидается [277] [273]
Галерея
[ редактировать ] Активность цианобактерий окрасила озеро кальдеры Коатепеке в бирюзовый цвет
Активность цианобактерий окрасила озеро кальдеры Коатепеке в бирюзовый цвет Цветение цианобактерий возле Фиджи
Цветение цианобактерий возле Фиджи Цианобактерии в озере Кёйлиё .
Цианобактерии в озере Кёйлиё .- Видео — Oscillatoria и Gleocapsa — с колебательным движением, когда нити Oscillatoria ориентируются на свет.



См. также
[ редактировать ]- Архейский Эон
- Бактериальные типы , другие основные линии бактерий.
- Биодизель
- Цианобионт
- Эндосимбиотическая теория
- Геологическая история кислорода
- Гиполит
Примечания
[ редактировать ]- ^ Ботаники ограничивают название водоросли» протистскими « эукариотами , что не распространяется на цианобактерии, которые являются прокариотами . Однако общее название «сине-зеленые водоросли» продолжает использоваться как синоним цианобактерий за пределами биологических наук.
Ссылки
[ редактировать ]- ^ Сильва ПК, Мо РЛ (декабрь 2019 г.). «Цианофицеи» . ДоступНаука . Макгроу Хилл Образование . дои : 10.1036/1097-8542.175300 . Проверено 21 апреля 2011 г.
- ^ Орен А. (сентябрь 2004 г.). «Предложение по дальнейшей интеграции цианобактерий в Бактериологический кодекс» . Международный журнал систематической и эволюционной микробиологии . 54 (Часть 5): 1895–1902. дои : 10.1099/ijs.0.03008-0 . ПМИД 15388760 .
- ^ Комарек Ю., Каштовский Ю., Мареш Ю., Йохансен Ю.Р. (2014). «Таксономическая классификация цианопрокариотов (родов цианобактерий) 2014 г. с использованием полифазного подхода» (PDF) . Преслия . 86 : 295–335.
- ^ Синха Р.П., Хадер Д.П. (2008). «УФ-защита в цианобактериях». Наука о растениях . 174 (3): 278–289. Бибкод : 2008PlnSc.174..278S . doi : 10.1016/j.plantsci.2007.12.004 .
- ^ Харпер, Дуглас. «голубой» . Интернет-словарь этимологии . Проверено 21 января 2018 г.
- ^ κύανος . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей» .
- ^ «История жизни и экология цианобактерий» . Музей палеонтологии Калифорнийского университета . Архивировано из оригинала 19 сентября 2012 года . Проверено 17 июля 2012 г.
- ^ «Браузер таксономии – цианобактерии» . Национальный центр биотехнологической информации . NCBI:txid1117 . Проверено 12 апреля 2018 г.
- ^ Аллаби М., изд. (1992). «Водоросли». Краткий словарь ботаники . Оксфорд: Издательство Оксфордского университета .
- ^ Перейти обратно: а б Крокфорд П.В., Бар Он Ю.М., Уорд Л.М., Майло Р., Халеви И. (ноябрь 2023 г.). «Геологическая история первичной продуктивности». Современная биология . 33 (21): 4741–4750.e5. Бибкод : 2023CBio...33E4741C . дои : 10.1016/j.cub.2023.09.040 . ПМИД 37827153 . S2CID 263839383 .
- ^ Сталь ЖЖ, Кретойу М.С. (2016). Морской микробиом: неиспользованный источник биоразнообразия и биотехнологического потенциала . Springer Science+Business Media . ISBN 978-3319330006 .
- ^ Уиттон Б.А., изд. (2012). «Ископаемая летопись цианобактерий» . Экология цианобактерий II: их разнообразие в пространстве и времени . Springer Science+Business Media . п. 17. ISBN 978-94-007-3855-3 .
- ^ «Бактерии» . Базовая биология. 18 марта 2016 г.
- ^ Либертон М., Пакраси Х.Б. (2008). «Глава 10. Мембранные системы цианобактерий». В Herrero A, Flore E (ред.). Цианобактерии: молекулярная биология, геномика и эволюция . Норидж, Великобритания: Horizon Scientific Press . стр. 217–287. ISBN 978-1-904455-15-8 .
- ^ Либертон М., Пейдж Л.Е., О'Делл В.Б., О'Нил Х., Мамонтов Э., Урбан В.С., Пакраси Х.Б. (февраль 2013 г.). «Организация и гибкость тилакоидных мембран цианобактерий, исследованная методом рассеяния нейтронов» . Журнал биологической химии . 288 (5): 3632–3640. дои : 10.1074/jbc.M112.416933 . ПМЦ 3561581 . ПМИД 23255600 .
- ^ Моншам М.Э., Спаак П., Помати Ф. (27 июля 2019 г.). «Долгосрочное разнообразие и распространение нефотосинтезирующих цианобактерий в приальпийских озерах» . Границы микробиологии . 9 : 3344. doi : 10.3389/fmicb.2018.03344 . ПМК 6340189 . ПМИД 30692982 .
- ^ Патхак Дж., Раджниш, Маурья П.К., Сингх С.П., Хадер Д.П., Синха Р.П. (2018). «Цианобактериальное сельское хозяйство для экологически чистых и устойчивых методов ведения сельского хозяйства: инновации и перспективы» . Границы в науке об окружающей среде . 6 . дои : 10.3389/fenvs.2018.00007 . ISSN 2296-665X .
- ^ Моррисон Дж. (11 января 2016 г.). «Живые бактерии движутся в воздушных потоках Земли» . Смитсоновский журнал . Проверено 10 августа 2022 г.
- ^ Уиттон Б.А., Поттс М. (2012). «Знакомство с цианобактериями». В Уиттоне, BA (ред.). Экология цианобактерий II . стр. 1–13. дои : 10.1007/978-94-007-3855-3_1 . ISBN 978-94-007-3854-6 .
- ^ Перейти обратно: а б с Тамулонис С., Постма М., Каандорп Дж. (2011). «Моделирование нитчатых цианобактерий раскрывает преимущества длинных и быстрых трихом для оптимизации светового воздействия» . ПЛОС ОДИН . 6 (7): e22084. Бибкод : 2011PLoSO...622084T . дои : 10.1371/journal.pone.0022084 . ПМЦ 3138769 . ПМИД 21789215 .
- ^ Перейти обратно: а б Оставайся в ЖЖ (5 июля 2012). «Цианобактериальные маты и строматолиты» . В Уиттоне, BA (ред.). Экология цианобактерий II: их разнообразие в пространстве и времени . Springer Science & Business Media. ISBN 9789400738553 . Проверено 15 февраля 2022 г. - через Google Книги.
- ^ Перейти обратно: а б с д и ж г час Тамулонис С., Постма М., Каандорп Дж. (2011). «Моделирование нитчатых цианобактерий раскрывает преимущества длинных и быстрых трихом для оптимизации светового воздействия» . ПЛОС ОДИН . 6 (7): e22084. Бибкод : 2011PLoSO...622084T . дои : 10.1371/journal.pone.0022084 . ПМЦ 3138769 . ПМИД 21789215 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Вайс КР (30 июля 2006 г.). «Первобытный прилив токсинов» . Лос-Анджелес Таймс . Архивировано из оригинала 14 августа 2006 года.
- ^ Доддс В.К., Гаддер Д.А., Молленхауэр Д. (1995). «Экология Ностока» . Журнал психологии . 31 (1): 2–18. Бибкод : 1995JPcgy..31....2D . дои : 10.1111/j.0022-3646.1995.00002.x . S2CID 85011483 .
- ^ Перейти обратно: а б с д и ж Агилера А., Клеменчич М., Суэльдо Дж., Ржимски П., Джаннуцци Л., Мартин М.В. (2021). «Клеточная смерть цианобактерий: современное понимание и рекомендации по достижению консенсуса по ее номенклатуре» . Границы микробиологии . 12 : 631654. doi : 10.3389/fmicb.2021.631654 . ПМЦ 7965980 . ПМИД 33746925 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Рэйвен РА (5 июля 2012 г.). «Физиологическая экология: Углерод». В Уиттоне, BA (ред.). Экология цианобактерий II: их разнообразие в пространстве и времени . Спрингер. п. 442. ИСБН 9789400738553 .
- ^ Перейти обратно: а б Ширмейстер Б.Е., де Вос Ж.М., Антонелли А., Багери Х.К. (январь 2013 г.). «Эволюция многоклеточности совпала с увеличением разнообразия цианобактерий и Великим событием окисления» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (5): 1791–1796. Бибкод : 2013PNAS..110.1791S . дои : 10.1073/pnas.1209927110 . ПМЦ 3562814 . ПМИД 23319632 .
- ^ Буллерьян Г.С., Post AF (2014). «Физиология и молекулярная биология водных цианобактерий» . Границы микробиологии . 5 : 359. дои : 10.3389/fmicb.2014.00359 . ПМК 4099938 . ПМИД 25076944 .
- ^ Тан В., Ван С., Фонсека-Батиста Д., Дехайрс Ф., Гиффорд С., Гонсалес А.Г. и др. (февраль 2019 г.). «Возврат к распределению океанической фиксации N 2 и оценка вклада диазотрофов в морскую продукцию» . Природные коммуникации . 10 (1): 831. doi : 10.1038/s41467-019-08640-0 . ПМК 6381160 . ПМИД 30783106 .
- ^ Блаха Л., Бабица П., Маршалек Б. (июнь 2009 г.). «Токсины, образующиеся при цветении цианобактерий в воде – токсичность и риски» . Междисциплинарная токсикология . 2 (2): 36–41. дои : 10.2478/v10102-009-0006-2 . ПМК 2984099 . ПМИД 21217843 .
- ^ Паерл Х.В., Оттен Т.Г. (май 2013 г.). «Вредное цветение цианобактерий: причины, последствия и меры борьбы». Микробная экология . 65 (4): 995–1010. Бибкод : 2013MicEc..65..995P . дои : 10.1007/s00248-012-0159-y . ПМИД 23314096 . S2CID 5718333 .
- ^ Перейти обратно: а б с д и ж Хейсман Дж., Кодд Г.А., Паерл Х.В., Ибелингс Б.В., Верспаген Дж.М., Виссер П.М. (август 2018 г.). «Цианобактерии цветут». Обзоры природы. Микробиология . 16 (8): 471–483. дои : 10.1038/s41579-018-0040-1 . ПМИД 29946124 . S2CID 49427202 .
- ^ Филд CB, Беренфельд М.Дж., Рандерсон Дж.Т., Фальковски П. (июль 1998 г.). «Первичная продукция биосферы: интеграция наземных и океанических компонентов» . Наука . 281 (5374): 237–240. Бибкод : 1998Sci...281..237F . дои : 10.1126/science.281.5374.237 . ПМИД 9657713 .
- ^ Кабельо-Йевес П.Дж., Сканлан Дж., Каллиери С., Пикасо А., Шалленберг Л., Хубер П. и др. (октябрь 2022 г.). «α-цианобактерии, обладающие формой IA RuBisCO, глобально доминируют в водных средах обитания» . Журнал ISME . 16 (10). ООО «Спрингер Сайенс энд Бизнес Медиа»: 2421–2432. Бибкод : 2022ISMEJ..16.2421C . дои : 10.1038/s41396-022-01282-z . ПМЦ 9477826 . ПМИД 35851323 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б Зер Дж. П. (апрель 2011 г.). «Азотфиксация морскими цианобактериями». Тенденции в микробиологии . 19 (4): 162–173. дои : 10.1016/j.tim.2010.12.004 . ПМИД 21227699 .
- ^ Перейти обратно: а б Томпсон А.В., Фостер Р.А., Крупке А., Картер Б.Дж., Мусат Н., Валот Д. и др. (сентябрь 2012 г.). «Одноклеточная цианобактерия, симбиотическая с одноклеточной эукариотической водорослью». Наука . 337 (6101): 1546–1550. Бибкод : 2012Sci...337.1546T . дои : 10.1126/science.1222700 . ПМИД 22997339 . S2CID 7071725 .
- ^ Перейти обратно: а б Джонсон З.И., Зинсер Э.Р., Коу А., МакНалти Н.П., Вудворд Э.М., Чисхолм С.В. (март 2006 г.). «Разделение ниш между экотипами Prochromococcus по градиентам окружающей среды в масштабе океана». Наука . 311 (5768): 1737–1740. Бибкод : 2006Sci...311.1737J . дои : 10.1126/science.1118052 . ПМИД 16556835 . S2CID 3549275 .
- ^ Перейти обратно: а б Сканлан Д.Д., Островски М., Мазар С., Дюфрен А., Гарчарек Л., Хесс В.Р. и др. (июнь 2009 г.). «Экологическая геномика морских пикоцианобактерий» . Обзоры микробиологии и молекулярной биологии . 73 (2): 249–299. дои : 10.1128/MMBR.00035-08 . ПМЦ 2698417 . ПМИД 19487728 .
- ^ Перейти обратно: а б Фломбаум П., Гальегос Дж.Л., Гордилло Р.А., Корнер Дж., Шафран Л.Л., Цзяо Н. и др. (июнь 2013 г.). «Настоящее и будущее глобальное распространение морских цианобактерий Prochromococcus и Synechococcus» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (24): 9824–9 Бибкод : 2013PNAS..110.9824F . дои : 10.1073/pnas.1307701110 . ПМЦ 3683724 . ПМИД 23703908 .
- ^ Фостер Р.А., Кайперс М.М., Вагнер Т., Паерл Р.В., Мусат Н., Зер Дж.П. (сентябрь 2011 г.). «Фиксация и перенос азота в диатомево-цианобактериальных симбиозах открытого океана» . Журнал ISME . 5 (9): 1484–1493. Бибкод : 2011ISMEJ...5.1484F . дои : 10.1038/ismej.2011.26 . ПМК 3160684 . ПМИД 21451586 .
- ^ Вильярреал ТА (1990). «Лабораторная культура и предварительная характеристика азотфиксирующего симбиоза ризосолении и рихелии». Морская экология . 11 (2): 117–132. Бибкод : 1990Март..11..117В . дои : 10.1111/j.1439-0485.1990.tb00233.x .
- ^ Янсон С., Воутерс Дж., Бергман Б., Карпентер Э.Дж. (октябрь 1999 г.). «Специфичность хозяина в симбиозе Richelia-диатомовых водорослей, выявленная с помощью анализа последовательности гена hetR». Экологическая микробиология . 1 (5): 431–438. Бибкод : 1999EnvMi...1..431J . дои : 10.1046/j.1462-2920.1999.00053.x . ПМИД 11207763 .
- ^ Перейти обратно: а б с д и Санчес-Баракальдо П. (декабрь 2015 г.). «Происхождение морских планктонных цианобактерий» . Научные отчеты . 5 : 17418. Бибкод : 2015NatSR...517418S . дои : 10.1038/srep17418 . ПМК 4665016 . ПМИД 26621203 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Кеттлер Г.К., Мартини А.С., Хуанг К., Цукер Дж., Коулман М.Л., Родриг С. и др. (декабрь 2007 г.). «Закономерности и последствия приобретения и потери генов в эволюции прохлорококка» . ПЛОС Генетика . 3 (12): е231. дои : 10.1371/journal.pgen.0030231 . ПМК 2151091 . ПМИД 18159947 .
- ^ Немирофф Р., Боннелл Дж., ред. (27 сентября 2006 г.). «Земля от Сатурна» . Астрономическая картина дня . НАСА .
- ^ Перейти обратно: а б Партенский Ф., Гесс В.Р., Вало Д. (март 1999 г.). «Прохлорококк, морской фотосинтезирующий прокариот мирового значения» . Обзоры микробиологии и молекулярной биологии . 63 (1): 106–127. дои : 10.1128/ММБР.63.1.106-127.1999 . ПМК 98958 . ПМИД 10066832 .
- ^ «Самый важный микроб, о котором вы никогда не слышали» . npr.org .
- ^ Перейти обратно: а б Классен Д., Розен Д.Е., Койперс О.П., Согаард-Андерсен Л., ван Везель Г.П. (февраль 2014 г.). «Бактериальные решения проблемы многоклеточности: рассказ о биопленках, нитях и плодовых телах» (PDF) . Обзоры природы. Микробиология . 12 (2): 115–124. дои : 10.1038/nrmicro3178 . hdl : 11370/0db66a9c-72ef-4e11-a75d-9d1e5827573d . ПМИД 24384602 . S2CID 20154495 .
- ^ Нюрнбергский DJ, Марискаль В., Паркер Дж., Мастроянни Дж., Флорес Э., Муллино CW (март 2014 г.). «Ветвление и межклеточная коммуникация у цианобактерии Раздела V Mastigocladus laminosus, сложного многоклеточного прокариота». Молекулярная микробиология . 91 (5): 935–949. дои : 10.1111/mmi.12506 . hdl : 10261/99110 . ПМИД 24383541 . S2CID 25479970 .
- ^ Эрреро А., Ставанс Дж., Флорес Э. (ноябрь 2016 г.). «Многоклеточная природа нитчатых гетероцистообразующих цианобактерий». Обзоры микробиологии FEMS . 40 (6): 831–854. дои : 10.1093/femsre/fuw029 . hdl : 10261/140753 . ПМИД 28204529 .
- ^ Риссер Д.Д., Чу В.Г., Микс Дж.К. (апрель 2014 г.). «Генетическая характеристика локуса hmp, кластера генов, подобного хемотаксису, который регулирует развитие и подвижность гормогоний у Nostoc punctiforme» . Молекулярная микробиология . 92 (2): 222–233. дои : 10.1111/mmi.12552 . ПМИД 24533832 . S2CID 37479716 .
- ^ Хайатан Б., Бэйнс Д.К., Ченг М.Х., Чо Ю.В., Хьюнь Дж., Ким Р. и др. (май 2017 г.). «Предполагаемая O-связанная β- N -ацетилглюкозаминтрансфераза необходима для развития и подвижности гормонов в нитчатых цианобактериях Nostoc punctiforme» . Журнал бактериологии . 199 (9): e00075–17. дои : 10.1128/JB.00075-17 . ПМК 5388816 . ПМИД 28242721 .
- ^ Эстевес-Феррейра А.А., Кавальканти Дж.Х., Ваз М.Г., Альваренга Л.В., Нуньес-Неси А., Араужо В.Л. (2017). «Цианобактериальные нитрогеназы: филогенетическое разнообразие, регуляция и функциональные прогнозы» . Генетика и молекулярная биология . 40 (1 прил. 1): 261–275. дои : 10.1590/1678-4685-GMB-2016-0050 . ПМЦ 5452144 . ПМИД 28323299 .
- ^ Микс Дж.К., Элхай Дж., Тиль Т., Поттс М., Лаример Ф., Ламердин Дж. и др. (2001). «Обзор генома Nostoc punctiforme, многоклеточной симбиотической цианобактерии». Исследования фотосинтеза . 70 (1): 85–106. дои : 10.1023/А:1013840025518 . ПМИД 16228364 . S2CID 8752382 .
- ^ Перейти обратно: а б Golden JW, Юн Х.С. (декабрь 1998 г.). «Формирование гетероцист в Анабаене». Современное мнение в микробиологии . 1 (6): 623–629. дои : 10.1016/s1369-5274(98)80106-9 . ПМИД 10066546 .
- ^ Перейти обратно: а б с Фэй П. (июнь 1992 г.). «Кислородные отношения азотфиксации у цианобактерий» . Микробиологические обзоры . 56 (2): 340–373. дои : 10.1128/MMBR.56.2.340-373.1992 . ПМК 372871 . ПМИД 1620069 .
- ^ Сингх В., Панде ПК, Джайн Д.К. (ред.). «Цианобактерии, актиномицеты, микоплазмы и риккетсии» . Учебник по ботанике «Разнообразие микробов и криптогам» . Публикации Растоги. п. 72. ИСБН 978-8171338894 .
- ^ «Различия между бактериями и цианобактериями» . Заметки по микробиологии . 29 октября 2015 года . Проверено 21 января 2018 г.
- ^ Уолсби А.Е. (март 1994 г.). «Газовые пузырьки» . Микробиологические обзоры . 58 (1): 94–144. дои : 10.1128/ММБР.58.1.94-144.1994 . ПМК 372955 . ПМИД 8177173 .
- ^ Симс ГК, Дуниган EP (1984). «Суточные и сезонные изменения активности нитрогеназы C
22Ч
2 сокращение) корней риса». Биология и биохимия почвы . 16 : 15–18. doi : 10.1016/0038-0717(84)90118-4 . - ^ Бокки С., Мальджиольо А (2010). «Азолла-Анабаена как биоудобрение для рисовых полей в долине реки По, рисовой зоне с умеренным климатом в Северной Италии» . Международный журнал агрономии . 2010 : 1–5. дои : 10.1155/2010/152158 . hdl : 2434/149583 .
- ^ Хуокко Т., Ни Т., Дайкс Г.Ф., Симпсон Д.М., Браунридж П., Конради Ф.Д. и др. (июнь 2021 г.). «Изучение пути биогенеза и динамики тилакоидных мембран» . Природные коммуникации . 12 (1): 3475. Бибкод : 2021NatCo..12.3475H . дои : 10.1038/s41467-021-23680-1 . ПМЦ 8190092 . ПМИД 34108457 .
- ^ Керфельд Калифорния, Хайнхорст С, Кэннон GC (2010). «Бактериальные микрокомпарты» . Ежегодный обзор микробиологии . 64 (1): 391–408. дои : 10.1146/annurev.micro.112408.134211 . ПМК 6022854 . ПМИД 20825353 .
- ^ Рэй Б.Д., Лонг Б.М., Бэджер М.Р., Прайс Г.Д. (сентябрь 2013 г.). «Функции, состав и эволюция двух типов карбоксисом: многогранных микрокомпартментов, которые облегчают фиксацию CO2 у цианобактерий и некоторых протеобактерий» . Обзоры микробиологии и молекулярной биологии . 77 (3): 357–379. дои : 10.1128/MMBR.00061-12 . ПМЦ 3811607 . ПМИД 24006469 .
- ^ Лонг Б.М., Бэджер М.Р., Уитни С.М., Прайс Г.Д. (октябрь 2007 г.). «Анализ карбоксисом Synechococcus PCC7942 выявил множественные комплексы Рубиско с карбоксисомными белками CcmM и CcaA» . Журнал биологической химии . 282 (40): 29323–29335. дои : 10.1074/jbc.M703896200 . ПМИД 17675289 .
- ^ Воткнехт Калифорнийский университет, Вестхофф П. (декабрь 2001 г.). «Биогенез и происхождение тилакоидных мембран» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1541 (1–2): 91–101. дои : 10.1016/S0167-4889(01)00153-7 . ПМИД 11750665 .
- ^ Собеховска-Сасим М, Стон-Эгерт Дж, Косаковска А (февраль 2014 г.). «Количественный анализ экстрагированных фикобилиновых пигментов в цианобактериях – оценка спектрофотометрическими и спектрофлуориметрическими методами» . Журнал прикладной психологии . 26 (5): 2065–2074. Бибкод : 2014JAPco..26.2065S . дои : 10.1007/s10811-014-0244-3 . ПМК 4200375 . ПМИД 25346572 .
- ^ Перейти обратно: а б Пишотта Дж.М., Зоу Ю., Баскаков И.В. (май 2010 г.). Ян Ч. (ред.). «Светозависимая электрогенная активность цианобактерий» . ПЛОС ОДИН . 5 (5): е10821. Бибкод : 2010PLoSO...510821P . дои : 10.1371/journal.pone.0010821 . ПМК 2876029 . ПМИД 20520829 .
- ^ Перейти обратно: а б с Вермаас В.Ф. (2001). «Фотосинтез и дыхание цианобактерий». Фотосинтез и дыхание цианобактерий. ЭЛС . John Wiley & Sons , Ltd. doi : 10.1038/npg.els.0001670 . ISBN 978-0-470-01590-2 . S2CID 19016706 .
- ^ Армстронф Дж. Э. (2015). Как Земля стала зеленой: краткая история растений за 3,8 миллиарда лет . университета Издательство Чикагского . ISBN 978-0-226-06977-7 .
- ^ Клатт Дж. М., де Бир Д., Хойслер С., Полерецкий Л. (2016). «Цианобактерии в микробных матах сульфидного источника могут осуществлять кислородный и аноксигенный фотосинтез одновременно в течение всего суточного периода» . Границы микробиологии . 7 : 1973. doi : 10.3389/fmicb.2016.01973 . ПМК 5156726 . ПМИД 28018309 .
- ^ Гроссман А.Р., Шефер М.Р., Чан Г.Г., Коллиер Дж.Л. (сентябрь 1993 г.). «Фикобилисома, светособирающий комплекс, реагирующий на условия окружающей среды» . Микробиологические обзоры . 57 (3): 725–749. дои : 10.1128/MMBR.57.3.725-749.1993 . ПМК 372933 . PMID 8246846 .
- ^ «Цвета от бактерий | Причины цвета» . www.webexhibits.org . Проверено 22 января 2018 г.
- ^ Гарсия-Пишель Ф (2009). «Цианобактерии». В Шехтере М. (ред.). Энциклопедия микробиологии (третье изд.). стр. 107–24. дои : 10.1016/B978-012373944-5.00250-9 . ISBN 978-0-12-373944-5 .
- ^ Кехо Д.М. (май 2010 г.). «Хроматическая адаптация и эволюция светочувствительности у цианобактерий» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (20): 9029–9030. Бибкод : 2010PNAS..107.9029K . дои : 10.1073/pnas.1004510107 . ПМЦ 2889117 . ПМИД 20457899 .
- ^ Кехо Д.М., Гуту А. (2006). «Реакция на цвет: регуляция дополнительной хроматической адаптации». Ежегодный обзор биологии растений . 57 : 127–150. doi : 10.1146/annurev.arplant.57.032905.105215 . ПМИД 16669758 .
- ^ Паленик Б., Хазелькорн Р. (январь 1992 г.). «Множественное эволюционное происхождение прохлорофитов, прокариот, содержащих хлорофилл b». Природа . 355 (6357): 265–267. Бибкод : 1992Natur.355..265P . дои : 10.1038/355265a0 . ПМИД 1731224 . S2CID 4244829 .
- ^ Урбах Э., Робертсон Д.Л., Чисхолм С.В. (январь 1992 г.). «Множественное эволюционное происхождение прохлорофитов в цианобактериальной радиации». Природа . 355 (6357): 267–270. Бибкод : 1992Natur.355..267U . дои : 10.1038/355267a0 . ПМИД 1731225 . S2CID 2011379 .
- ^ Коэн Ю., Йоргенсен Б.Б., Ревсбех Н.П., Поплавски Р. (февраль 1986 г.). «Адаптация к сероводороду кислородного и аноксигенного фотосинтеза цианобактерий» . Прикладная и экологическая микробиология . 51 (2): 398–407. Бибкод : 1986ApEnM..51..398C . дои : 10.1128/АЕМ.51.2.398-407.1986 . ПМК 238881 . ПМИД 16346996 .
- ^ Бланкеншип RE (2014). Молекулярные механизмы фотосинтеза . Уайли-Блэквелл . стр. 147–73. ISBN 978-1-4051-8975-0 .
- ^ Перейти обратно: а б Оч Л.М., Шилдс-Чжоу Г.А. (январь 2012 г.). «Неопротерозойское событие оксигенации: экологические возмущения и биогеохимический цикл». Обзоры наук о Земле . 110 (1–4): 26–57. Бибкод : 2012ESRv..110...26O . doi : 10.1016/j.earscirev.2011.09.004 .
- ^ Адамс Д.Г., Бергман Б., Нирцвицки-Бауэр С.А., Дагган П.С., Рай А.Н., Шюсслер А (2013). «Цианобактериально-растительные симбиозы». Розенберг Э., Делонг Э.Ф., Лори С., Стакебрандт Э., Томпсон Ф. (ред.). Прокариоты . Шпрингер, Берлин, Гейдельберг. стр. 359–400. дои : 10.1007/978-3-642-30194-0_17 . ISBN 978-3-642-30193-3 .
- ^ Чжан С., Брайант Д.А. (декабрь 2011 г.). «Цикл трикарбоновых кислот у цианобактерий». Наука . 334 (6062): 1551–1553. Бибкод : 2011Sci...334.1551Z . дои : 10.1126/science.1210858 . ПМИД 22174252 . S2CID 206536295 .
- ^ Сюн В., Ли Т.К., Роммельфангер С., Йерсинг Э., Кано М., Манесс ПК. и др. (декабрь 2015 г.). «Путь фосфокетолазы способствует метаболизму углерода у цианобактерий». Природные растения . 2 (1): 15187. doi : 10.1038/nplants.2015.187 . ПМИД 27250745 . S2CID 40094360 .
- ^ Смит А (1973). «Синтез продуктов обмена веществ» . В Карр Н.Г., Уиттон Б.А. (ред.). Биология сине-зеленых водорослей . Издательство Калифорнийского университета. стр. 30– . ISBN 978-0-520-02344-4 .
- ^ Жангу М (1987). «Болезни иглокожих. I. Возбудители микроорганизмов и протистанов» . Болезни водных организмов . 2 : 147–62. дои : 10.3354/dao002147 .
- ^ Кинне О, изд. (1980). Болезни морских животных (PDF) . Том. 1. Чичестер, Великобритания: John Wiley & Sons. ISBN 978-0-471-99584-5 .
- ^ Кристиансен А (1964). « Sarcinastrum urosporae , бесцветная паразитическая сине-зеленая водоросль» (PDF) . Психология . 4 (1): 19–22. Бибкод : 1964Phyco...4...19K . дои : 10.2216/i0031-8884-4-1-19.1 . Архивировано из оригинала (PDF) 6 января 2015 года.
- ^ Мазард С., Пенесян А., Островски М., Полсен И.Т., Иган С. (май 2016 г.). «Маленькие микробы с большим влиянием: роль цианобактерий и их метаболитов в формировании нашего будущего» . Морские наркотики . 14 (5): 97. дои : 10.3390/md14050097 . ПМЦ 4882571 . ПМИД 27196915 .
- ^ де лос Риос А., Грубе М., Санчо Л.Г., Аскасо С. (февраль 2007 г.). «Ультраструктурные и генетические характеристики эндолитных цианобактериальных биопленок, колонизирующих гранитные породы Антарктики» . ФЭМС Микробиология Экология . 59 (2): 386–395. Бибкод : 2007FEMME..59..386D . дои : 10.1111/j.1574-6941.2006.00256.x . ПМИД 17328119 .
- ^ Воган Т. (2011). Маммология . Джонс и Барлетт. п. 21. ISBN 978-0763762995 .
- ^ Шульц Н. (30 августа 2009 г.). «Фотосинтетические вирусы поддерживают повышенный уровень кислорода в мире» . Новый учёный .
- ^ Перейти обратно: а б с Джонк К.Д., Хейсман Дж., Шарплс Дж., Соммейер Б., Виссер П.М., Струм Дж.М. (1 марта 2008 г.). «Летняя жара способствует цветению вредных цианобактерий» . Биология глобальных изменений . 14 (3): 495–512. Бибкод : 2008GCBio..14..495J . дои : 10.1111/j.1365-2486.2007.01510.x . S2CID 54079634 .
- ^ «Линда Лоутон – 11-я Международная конференция по токсичным цианобактериям» . Проверено 25 июня 2021 г.
- ^ Паерл Х.В., Пол В.Дж. (апрель 2012 г.). «Изменение климата: связь с глобальной экспансией вредных цианобактерий». Исследования воды . 46 (5): 1349–1363. Бибкод : 2012WatRe..46.1349P . дои : 10.1016/j.watres.2011.08.002 . ПМИД 21893330 .
- ^ Томас А.Д., Дугилл А.Дж. (15 марта 2007 г.). «Пространственное и временное распределение цианобактериальных почвенных корок в Калахари: влияние на свойства поверхности почвы». Геоморфология . 85 (1): 17–29. Бибкод : 2007Geomo..85...17T . дои : 10.1016/j.geomorph.2006.03.029 .
- ^ Белнап Дж., Гарднер Дж.С. (1993). «Микроструктура почвы в почвах плато Колорадо: роль цианобактерии Microcoleus Vaginatus». Натуралист Великого Бассейна . 53 (1): 40–47. JSTOR 41712756 .
- ^ Нельсон, Кори; Хиральдо-Сильва, Ана; Варсоп Томас, Финли; Гарсия-Пишель, Ферран (16 ноября 2022 г.). «Пространственная самосегрегация пионерских видов цианобактерий стимулирует организацию микробиома в биокорках» . ИСМЕ Коммуникации . 2 (1): 114. дои : 10.1038/s43705-022-00199-0 . ПМЦ 9723579 . ПМИД 37938289 .
- ^ Надис С. (декабрь 2003 г.). «Клетки, управляющие морями» (PDF) . Научный американец . 289 (6): 52–53. Бибкод : 2003SciAm.289f..52N . doi : 10.1038/scientificamerican1203-52 . ПМИД 14631732 . Архивировано из оригинала (PDF) 19 апреля 2014 года . Проверено 19 апреля 2014 г.
- ^ Стюарт I, Falconer IR (2 сентября 2011 г.). «Цианобактерии и цианобактериальные токсины» . В Уолше П.Дж., Смите С., Флеминге Л., Соло-Габриэле Х., Гервике У.Х. (ред.). Океаны и здоровье человека: морские риски и средства правовой защиты . Академическая пресса . стр. 271–296. ISBN 978-0-08-087782-2 .
- ^ Перейти обратно: а б с Ли С.М., Рю С.М. (4 февраля 2021 г.). «Водоросли как новенькие в полезном микробиоме растений» . Границы в науке о растениях . 12 . Frontiers Media SA: 599742. doi : 10.3389/fpls.2021.599742 . ПМЦ 7889962 . ПМИД 33613596 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б с Ким М., Чой Д.Х., Пак М.Г. (май 2021 г.). «Генетическое разнообразие цианобионтов и специфичность хозяина динофлагелляты Ornithocercus, несущей цианобионты, в прибрежных водах умеренного климата» . Научные отчеты . 11 (1): 9458. Бибкод : 2021НатСР..11.9458К . дои : 10.1038/s41598-021-89072-z . ПМК 8097063 . ПМИД 33947914 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Гантар М., Эльхай Дж. (1999). «Колонизация параклубеньков пшеницы N2-фиксирующей цианобактерией Nostocsp . Штамм 2S9B» . Новый фитолог . 141 (3): 373–379. дои : 10.1046/j.1469-8137.1999.00352.x .
- ^ Гантар М (2000). «Механическое повреждение корней обеспечивает усиленную колонизацию эндоризосферы пшеницы диазотфиксирующей цианобактерией Nostoc sp. Штамм 2S9B». Биология и плодородие почв . 32 (3): 250–255. Бибкод : 2000BioFS..32..250G . дои : 10.1007/s003740000243 . S2CID 7590731 .
- ^ Тревес Х., Раанан Х., Кедем И., Мурик О., Керен Н., Зер Х. и др. (июнь 2016 г.). «Механизмы, благодаря которым зеленая водоросль Chlorella ohadii, выделенная из корки пустынной почвы, демонстрирует беспрецедентную устойчивость к фотоповреждениям» . Новый фитолог . 210 (4): 1229–1243. дои : 10.1111/nph.13870 . ПМИД 26853530 .
- ^ Чжу Х, Ли С, Ху З, Лю Г (декабрь 2018 г.). «Молекулярная характеристика эукариотических сообществ водорослей тропической филлосферы на основе секвенирования гена 18S рДНК в реальном времени» . Биология растений BMC . 18 (1): 365. дои : 10.1186/s12870-018-1588-7 . ПМК 6299628 . ПМИД 30563464 .
- ^ Крингс М., Хасс Х., Керп Х., Тейлор Т.Н., Агерер Р., Доцлер Н. (2009). «Эндофитные цианобактерии в наземном растении возрастом 400 миллионов лет: сценарий возникновения симбиоза?». Обзор палеоботаники и палинологии . 153 (1–2): 62–69. Бибкод : 2009RPaPa.153...62K . дои : 10.1016/j.revpalbo.2008.06.006 .
- ^ Картикеян Н., Прасанна Р., Суд А., Джайсвал П., Наяк С., Кошик Б.Д. (2009). «Физиологическая характеристика и электронно-микроскопическое исследование цианобактерий, связанных с ризосферой пшеницы». Фолиа микробиологическая . 54 (1): 43–51. дои : 10.1007/s12223-009-0007-8 . ПМИД 19330544 . S2CID 23420342 .
- ^ Перейти обратно: а б Бабу С., Прасанна Р., Бидьярани Н., Сингх Р. (2015). «Анализ колонизации инокулированных цианобактерий в растениях пшеницы с использованием биохимических и молекулярных инструментов». Журнал прикладной психологии . 27 (1): 327–338. Бибкод : 2015JAPco..27..327B . дои : 10.1007/s10811-014-0322-6 . S2CID 17353123 .
- ^ Перейти обратно: а б Бидьярани Н., Прасанна Р., Чавла Г., Бабу С., Сингх Р. (апрель 2015 г.). «Расшифровка факторов, связанных с заселением растений риса цианобактериями». Журнал фундаментальной микробиологии . 55 (4): 407–419. дои : 10.1002/jobm.201400591 . ПМИД 25515189 . S2CID 5401526 .
- ^ Перейти обратно: а б Гантар М., Керби Н.В., Роуэлл П. (1991). «Колонизация пшеницы (Triticum vulgare L.) N2-фиксирующими цианобактериями: II. Ультраструктурное исследование» . Новый фитолог . 118 (3): 485–492. дои : 10.1111/j.1469-8137.1991.tb00031.x .
- ^ Ахмед М., Сталь Ж.Дж., Хаснайн С. (2010). «Ассоциация негетероцистных цианобактерий с сельскохозяйственными растениями» . Растение и почва . 336 (1–2): 363–375. Бибкод : 2010ПлСой.336..363А . дои : 10.1007/s11104-010-0488-x . S2CID 21309970 .
- ^ Хусейн А., Хамаюн М., Шах С.Т. (ноябрь 2013 г.). «Корневая колонизация и фитостимуляция фитогормонами, продуцирующими энтофитный Nostoc sp. AH-12». Современная микробиология . 67 (5): 624–630. дои : 10.1007/s00284-013-0408-4 . ПМИД 23794014 . S2CID 14704537 .
- ^ Хусейн А., Шах С.Т., Рахман Х., Иршад М., Икбал А. (2015). «Влияние IAA на рост in vitro и колонизацию Nostoc в корнях растений» . Границы в науке о растениях . 6 : 46. дои : 10.3389/fpls.2015.00046 . ПМЦ 4318279 . ПМИД 25699072 .
- ^ Капоне Д.Г. (1997). «Триходезмий, морская цианобактерия глобального значения». Наука . 276 (5316): 1221–1229. дои : 10.1126/science.276.5316.1221 .
- ^ Фальковский П.Г., Барбер Р.Т., Сметачек В. (июль 1998 г.). «Биогеохимический контроль и обратная связь на первичное производство океана». Наука . 281 (5374): 200–207. дои : 10.1126/science.281.5374.200 . ПМИД 9660741 .
- ^ Хатчинс Д.А., Фу FX, Чжан Ю., Уорнер М.Э., Фэн Ю., Портун К., Бернхардт П.В., Малхолланд М.Р. (2007). «Контроль CO2 за фиксацией триходезмия N2, фотосинтезом, скоростью роста и соотношением элементов: последствия для биогеохимии океана в прошлом, настоящем и будущем» . Лимнология и океанография . 52 (4): 1293–1304. Бибкод : 2007LimOc..52.1293H . дои : 10.4319/lo.2007.52.4.1293 . S2CID 55606811 .
- ^ Хуан С., Вильгельм С.В., Харви Х.Р., Тейлор К., Цзяо Н., Чен Ф. (февраль 2012 г.). «Новые линии прохлорококков и синехококков в мировом океане» . Журнал ISME . 6 (2): 285–297. Бибкод : 2012ISMEJ...6..285H . дои : 10.1038/ismej.2011.106 . ПМК 3260499 . ПМИД 21955990 .
- ^ Шривастава А.К., Рай А.М., Нейлан Б.А. (март 2013 г.). Стрессовая биология цианобактерий: молекулярные механизмы клеточных реакций . ЦРК Пресс. ISBN 9781466504783 – через Google Книги .
- ^ Зер Дж.П., Бенч С.Р., Картер Б.Дж., Хьюсон И., Ниязи Ф., Ши Т. и др. (ноябрь 2008 г.). «Глобально распространенные некультивируемые океанические N2-фиксирующие цианобактерии лишены кислородной фотосистемы II». Наука . 322 (5904): 1110–1112. Бибкод : 2008Sci...322.1110Z . дои : 10.1126/science.1165340 . ПМИД 19008448 . S2CID 206516012 .
- ^ Десель Дж., Колин С., Фостер Р.А. (2015). «Фотосимбиоз морских планктонных протистов». Морские протисты . стр. 465–500. дои : 10.1007/978-4-431-55130-0_19 . ISBN 978-4-431-55129-4 .
- ^ Фостер Р.А., Зер Дж.П. (сентябрь 2019 г.). «Разнообразие, геномика и распространение одноклеточных симбиотических ассоциаций фитопланктона и цианобактерий». Ежегодный обзор микробиологии . 73 : 435–456. doi : 10.1146/annurev-micro-090817-062650 . ПМИД 31500535 . S2CID 202414294 .
- ^ Перейти обратно: а б с д и ж г Муллино CW, Уайльд А (июнь 2021 г.). «Социальная жизнь цианобактерий» . электронная жизнь . 10 . doi : 10.7554/eLife.70327 . ПМК 8208810 . ПМИД 34132636 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Каменная Н.А., Земля М., Махони Л., Чен Л., Холман Э., Холман Х.И. и др. (май 2018 г.). «Высокое pCO 2 -индуцированное богатые экзополисахаридами балластные агрегаты планктонных цианобактерий могут объяснить палеопротерозойское захоронение углерода» . Природные коммуникации . 9 (1): 2116. Бибкод : 2018NatCo...9.2116K . дои : 10.1038/s41467-018-04588-9 . ПМК 5974010 . ПМИД 29844378 .
- ^ Перейти обратно: а б Маэда К., Окуда Й., Эномото Г., Ватанабэ С., Икеучи М. (июнь 2021 г.). «Биосинтез сульфатированного экзополисахарида, синехана и образование цветения в модельной цианобактерии Synechocystis sp., штамм PCC 6803» . электронная жизнь . 10 . doi : 10.7554/eLife.66538 . ПМК 8205485 . ПМИД 34127188 .
- ^ Перейти обратно: а б Сим М.С., Лян Б., Петрофф А.П., Эванс А., Клепак-Серадж В., Фланнери Д.Т. и др. (2012). «Кислородозависимый морфогенез современных слипшихся фотосинтетических матов и последствия для архейских строматолитовых летописей» . Геонауки . 2 (4): 235–259. Бибкод : 2012Geosc...2..235S . doi : 10.3390/geosciences2040235 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0 .
- ^ Перейти обратно: а б Конради Ф.Д., Чжоу Р.К., Озер С., Шугерс Н., Уайлд А., Муллино CW (октябрь 2019 г.). «Факторы, контролирующие образование и структуру хлопьев в штамме Cyanobacterium Synechocystis sp. PCC 6803» . Журнал бактериологии . 201 (19). дои : 10.1128/JB.00344-19 . ПМЦ 6755745 . ПМИД 31262837 .
- ^ Эномото Дж., Икеучи М. (март 2020 г.). «Цианобактериохромы, чувствительные к синему/зеленому свету, являются датчиками затенения клеток в нишах, насыщенных красным светом» . iScience . 23 (3): 100936. Бибкод : 2020iSci...23j0936E . дои : 10.1016/j.isci.2020.100936 . ПМК 7063230 . ПМИД 32146329 .
- ^ Галлуцци Л., Браво-Сан-Педро Дж.М., Витале И., Ааронсон С.А., Абрамс Дж.М., Адам Д. и др. (январь 2015 г.). «Основные и дополнительные аспекты гибели клеток: рекомендации NCCD 2015» . Смерть клеток и дифференцировка . 22 (1): 58–73. дои : 10.1038/cdd.2014.137 . ПМЦ 4262782 . ПМИД 25236395 .
- ^ Аллен Р., Риттманн Б.Е., Кертисс Р. (апрель 2019 г.). «Формирование и агрегация аксенической биопленки штаммом Synechocystis sp. PCC 6803 индуцируются изменениями концентрации питательных веществ и требуют структур клеточной поверхности» . Прикладная и экологическая микробиология . 85 (7). Бибкод : 2019ApEnM..85E2192A . дои : 10.1128/АЕМ.02192-18 . ПМК 6585507 . ПМИД 30709828 .
- ^ Шугерс Н., Уайльд А. (март 2015 г.). «Придатки цианобактериальной клетки» . Жизнь . 5 (1): 700–715. Бибкод : 2015Жизнь....5..700С . дои : 10.3390/life5010700 . ПМК 4390875 . ПМИД 25749611 .
- ^ Перейти обратно: а б с Хаятан Б., Микс Дж.К., Риссер Д.Д. (декабрь 2015 г.). «Доказательства того, что модифицированная пилоподобная система IV типа обеспечивает скользящую подвижность и секрецию полисахаридов у нитчатых цианобактерий» . Молекулярная микробиология . 98 (6): 1021–1036. дои : 10.1111/mmi.13205 . ПМИД 26331359 . S2CID 8749419 .
- ^ Адамс Д.В., Штуцманн С., Штуцманн С., Блокеш М. (сентябрь 2019 г.). «Пили Vibrio cholerae, поглощающие ДНК, необходимы для колонизации хитина и способны к распознаванию родственников посредством специфичного для последовательности самодействия» . Природная микробиология . 4 (9): 1545–1557. дои : 10.1038/s41564-019-0479-5 . ПМК 6708440 . ПМИД 31182799 .
- ^ Кромкамп Дж., Уолсби А.Е. (1990). «Компьютерная модель плавучести и вертикальной миграции цианобактерий». Журнал исследований планктона . 12 : 161–183. дои : 10.1093/планкт/12.1.161 .
- ^ Агило-Ферретьянс, М.Д., Бош Р., Пуксти Р.Дж., Латва М., Заджелович В., Чхун А. и др. (март 2021 г.). «Пили позволяют доминирующим морским цианобактериям избегать погружения и уклоняться от хищников» . Природные коммуникации . 12 (1): 1857. Бибкод : 2021NatCo..12.1857A . дои : 10.1038/s41467-021-22152-w . ПМЦ 7994388 . ПМИД 33767153 .
- ^ Перейти обратно: а б Ху С., Ржимский П. (декабрь 2019 г.). «Запрограммированное клеточное смерть и сопутствующее высвобождение микроцистина в пресноводных цветущих цианобактериях Microcystis : от идентификации к экологической значимости» . Токсины . 11 (12): 706. doi : 10.3390/toxins11120706 . ПМК 6950475 . ПМИД 31817272 .
- ^ Агусти С. (июнь 2004 г.). «Жизнеспособность и сегрегация ниш клеток прохлорококка и синехококка в центральной части Атлантического океана» . Водная микробная экология . 36 (1): 53–59. дои : 10.3354/ame036053 . hdl : 10261/86957 .
- ^ Агусти С., Алоу Э.В., Хойер М.В., Фрейзер Т.К. , Кэнфилд Д.Э. (2006). «Гибель клеток в сообществах фитопланктона озер». Пресноводная биология . 51 (8): 1496–1506. Бибкод : 2006FrBio..51.1496A . дои : 10.1111/j.1365-2427.2006.01584.x .
- ^ Франклин DJ, Brussaard CP, Berges JA (2006). «Какова роль и природа запрограммированной гибели клеток в экологии фитопланктона?» . Европейский журнал психологии . 41 (1): 1–14. Бибкод : 2006EJPhy..41....1F . дои : 10.1080/09670260500505433 . S2CID 53599616 .
- ^ Сиджи, округ Колумбия, Селвин А., Галлуа П., Дин А.П. (2007). «Закономерности гибели клеток пресноводных колониальных цианобактерий во время цветения в конце лета». Психология . 46 (3): 284–292. Бибкод : 2007Phyco..46..284S . дои : 10.2216/06-69.1 . S2CID 86268392 .
- ^ Берман-Франк И., Бидле К.Д., Харамати Л., Фальковски П.Г. (2004). «Гибель морской цианобактерии Trichodesmium SPP. через автокатализируемый путь гибели клеток» . Лимнология и океанография . 49 (4): 997–1005. Бибкод : 2004LimOc..49..997B . дои : 10.4319/lo.2004.49.4.0997 .
- ^ Ху С., Ржимский П. (декабрь 2019 г.). «Запрограммированное клеточное смерть и сопутствующее высвобождение микроцистина в пресноводных цветущих цианобактериях Microcystis : от идентификации к экологической значимости» . Токсины . 11 (12): 706. doi : 10.3390/toxins11120706 . ПМК 6950475 . ПМИД 31817272 .
- ^ Ржимский П., Климашик П., Юрчак Т., Понедзялек Б. (2020). «Окислительный стресс, запрограммированная гибель клеток и высвобождение микроцистина у Microcystis aeruginosa в ответ на травоядных дафний » . Границы микробиологии . 11 : 1201. дои : 10.3389/fmicb.2020.01201 . ПМК 7311652 . ПМИД 32625177 .
- ^ Перейти обратно: а б Микс Дж.К., Элхай Дж., Тиль Т., Поттс М., Лаример Ф., Ламердин Дж. и др. (2001). «Обзор генома Nostoc punctiforme, многоклеточной симбиотической цианобактерии». Исследования фотосинтеза . 70 (1): 85–106. дои : 10.1023/А:1013840025518 . ПМИД 16228364 . S2CID 8752382 .
- ^ Саттл, Калифорния (1 января 2000 г.). «Цианофаги и их роль в экологии цианобактерий». В Whitton BA, Potts M (ред.). Экология цианобактерий . Спрингер Нидерланды. стр. 563–589. дои : 10.1007/0-306-46855-7_20 . ISBN 9780792347354 .
- ^ Саттл, Калифорния, Чан А.М. (1993). «Морские цианофаги, заражающие океанические и прибрежные штаммы Synechococcus: численность, морфология, перекрестная инфекционность и характеристики роста» . Серия «Прогресс в области морской экологии» . 92 : 99–109. Бибкод : 1993MEPS...92...99S . дои : 10.3354/meps092099 .
- ^ Фокин А., Россманн М.Г. (январь 2014 г.). «Молекулярная архитектура хвостатых двухцепочечных ДНК-фагов» . Бактериофаг . 4 (1): e28281. дои : 10.4161/bact.28281 . ПМЦ 3940491 . ПМИД 24616838 .
- ^ Проктор Л.М., Фурман Дж.А. (1990). «Вирусная смертность морских бактерий и цианобактерий». Природа . 343 (6253): 60–62. Бибкод : 1990Natur.343...60P . дои : 10.1038/343060a0 . S2CID 4336344 .
- ^ Перейти обратно: а б Сарма ТА. «Цианофаги» в Справочнике цианобактерий ( CRC Press ; 2012) ( ISBN 1466559411 )
- ^ Дагган П.С., Готтарделло П., Адамс Д.Г. (июнь 2007 г.). «Молекулярный анализ генов Nostoc punctiforme, участвующих в биогенезе пилуса и заражении растений» . Журнал бактериологии . 189 (12): 4547–4551. дои : 10.1128/JB.01927-06 . ЧВК 1913353 . ПМИД 17416648 .
- ^ Уайльд А., Муллино CW (декабрь 2015 г.). «Подвижность цианобактерий: полисахаридные треки и моторы пилуса IV типа» . Молекулярная микробиология . 98 (6): 998–1001. дои : 10.1111/mmi.13242 . ПМИД 26447922 . S2CID 22585994 .
- ^ Уотербери Дж. Б., Уилли Дж. М., Фрэнкс Д. Г., Валуа Ф. В., Уотсон С. В. (октябрь 1985 г.). «Цианобактерия, способная к плавательной подвижности». Наука . 230 (4721): 74–76. Бибкод : 1985Sci...230...74W . дои : 10.1126/science.230.4721.74 . ПМИД 17817167 . S2CID 29180516 .
- ^ Элерс К., Остер Г. (2012). «О загадочном движении синехококка» . ПЛОС ОДИН . 7 (5): e36081. Бибкод : 2012PLoSO...736081E . дои : 10.1371/journal.pone.0036081 . ПМЦ 3342319 . ПМИД 22567124 .
- ^ Мията М., Робинсон Р.К., Уеда Т.К., Фукумори Ю., Фукусима С.И., Харута С. и др. (январь 2020 г.). «Древо подвижности. Предлагаемая история систем подвижности на древе жизни» . Гены в клетки . 25 (1): 6–21. дои : 10.1111/gtc.12737 . ПМК 7004002 . ПМИД 31957229 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Кастенхольц Р.В. (1982). «Подвижность и налоги». В Карр Н.Г., Уиттон Б.А. (ред.). Биология цианобактерий . Калифорнийский университет Press, Беркли и Лос-Анджелес. стр. 413–439. ISBN 978-0-520-04717-4 .
- ^ Уотербери Дж. Б., Уилли Дж. М., Фрэнкс Д. Г., Валуа Ф. В., Уотсон С. В. (октябрь 1985 г.). «Цианобактерия, способная к плавательной подвижности». Наука . 230 (4721): 74–76. Бибкод : 1985Sci...230...74W . дои : 10.1126/science.230.4721.74 . ПМИД 17817167 . S2CID 29180516 .
- ^ Макбрайд, MJ (2001). «Скользящая подвижность бактерий: множественные механизмы движения клеток по поверхностям». Ежегодный обзор микробиологии . 55 : 49–75. дои : 10.1146/annurev.micro.55.1.49 . ПМИД 11544349 .
- ^ Райхенбах Х (1981). «Таксономия планирующих бактерий». Ежегодный обзор микробиологии . 35 : 339–364. дои : 10.1146/annurev.mi.35.100181.002011 . ПМИД 6794424 .
- ^ Хойчик Э., Баумайстер В. (октябрь 1998 г.). «Комплекс соединительных пор, прокариотическая органелла секреции, является молекулярным мотором, лежащим в основе скользящей подвижности у цианобактерий» . Современная биология . 8 (21): 1161–1168. Бибкод : 1998CBio....8.1161H . дои : 10.1016/S0960-9822(07)00487-3 . ПМИД 9799733 . S2CID 14384308 .
- ^ Хойчик Э (2000). «Скользящая подвижность цианобактерий: наблюдения и возможные объяснения». Архив микробиологии . 174 (1–2): 11–17. Бибкод : 2000ArMic.174...11H . дои : 10.1007/s002030000187 . ПМИД 10985737 . S2CID 9927312 .
- ^ Бхая Д., Ватанабэ Н., Огава Т., Гроссман А.Р. (март 1999 г.). «Роль альтернативного сигма-фактора в подвижности и формировании ворсинок у цианобактерии Synechocystis sp., штамма PCC6803» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (6): 3188–3193. Бибкод : 1999PNAS...96.3188B . дои : 10.1073/pnas.96.6.3188 . ЧВК 15917 . ПМИД 10077659 .
- ^ Перейти обратно: а б Донкор В.А., Амеовор Д.Х., Хадер Д.П. (1993). «Влияние тропической солнечной радиации на подвижность нитчатых цианобактерий» . ФЭМС Микробиология Экология . 12 (2): 143–147. Бибкод : 1993ЖЕНЩИНА..12..143D . дои : 10.1111/j.1574-6941.1993.tb00026.x .
- ^ Гарсиа-Пишель Ф., Мехлинг М., Кастенхольц Р.В. (май 1994 г.). «Суточная миграция микроорганизмов в бентосном сообществе гиперсоленого мата» . Прикладная и экологическая микробиология . 60 (5): 1500–1511. Бибкод : 1994ApEnM..60.1500G . doi : 10.1128/aem.60.5.1500-1511.1994 . ПМК 201509 . ПМИД 16349251 .
- ^ Фурсан А, Соле А, Диестра Е, Раншу-Пейрус А, Эстев И, Кометт П, Дюран Р (сентябрь 2006 г.). «Вертикальная миграция фототрофных бактериальных популяций в гиперсоленом микробном мате из Сален-де-Жиро (Камарг, Франция)» . ФЭМС Микробиология Экология . 57 (3): 367–377. Бибкод : 2006FEMME..57..367F . дои : 10.1111/j.1574-6941.2006.00124.x . ПМИД 16907751 .
- ^ Ричардсон Л.Л., Кастенхольц Р.В. (сентябрь 1987 г.). «Вертикальные движения цианобактерии Oscillatoria terebriformis в богатом сульфидами микробном мате горячих источников» . Прикладная и экологическая микробиология . 53 (9): 2142–2150. Бибкод : 1987ApEnM..53.2142R . doi : 10.1128/aem.53.9.2142-2150.1987 . ПМК 204072 . ПМИД 16347435 .
- ^ Хадер Д.П. (1987). «ВЛИЯНИЕ УФ-Б-ОБЛУЧЕНИЯ НА ФОТОДВИЖЕНИЕ В ДЕСМИДЕ Cosmarium cucumis». Фотохимия и фотобиология . 46 : 121–126. дои : 10.1111/j.1751-1097.1987.tb04745.x . S2CID 97100233 .
- ^ Нульч В., Хедер Д.П. (июнь 1988 г.). «Фотодвижение подвижных микроорганизмов-II» . Фотохимия и фотобиология . 47 (6): 837–869. дои : 10.1111/j.1751-1097.1988.tb01668.x . ПМИД 3064112 . S2CID 26445775 .
- ^ Чекуччи Г., Сгарбосса А. и Ленчи Ф. (2004) «Фотодвижения микроорганизмов: Введение». Справочник CRC по органической фотохимии и фотобиологии , 2-е изд., CRC Press.
- ^ Нульч, Вильгельм (1962) «ВЛИЯНИЕ СВЕТА НА ДВИЖЕНИЕ ЦИАНОФИЦОВ: III. Коммуникация: ФОТОФОБОТАКСИС PHORMIDIUM UNCINATUM». Планта , 58 (6): 647–63.
- ^ Нульч В., Шухарт Х., Хёль М. (1979). «Исследования фототаксической ориентации Anabaena variabilis». Архив микробиологии . 122 (1): 85–91. Бибкод : 1979ArMic.122...85N . дои : 10.1007/BF00408050 . S2CID 12242837 .
- ^ Езда на Р (2007). «Термин строматолит: к существенному определению». Летайя . 32 (4): 321–30. дои : 10.1111/j.1502-3931.1999.tb00550.x .
- ^ Баумгартнер Р.Дж., Ван Кранендонк М.Дж., Уэйси Д., Фиорентини М.Л., Сондерс М., Карузо С. и др. (2019). «Нанопористый пирит и органическое вещество в строматолитах возрастом 3,5 миллиарда лет свидетельствуют о первозданной жизни» (PDF) . Геология . 47 (11): 1039–43. Бибкод : 2019Geo....47.1039B . дои : 10.1130/G46365.1 . S2CID 204258554 .
- ^ Шопф Дж.В., Пакер Б.М. (июль 1987 г.). «Раннеархейские (возрастом от 3,3 до 3,5 миллиардов лет) микроокаменелости из группы Варравуна, Австралия». Наука . 237 (4810): 70–73. Бибкод : 1987Sci...237...70S . дои : 10.1126/science.11539686 . ПМИД 11539686 .
- ^ Фаркуар Дж., Бао Х., Тименс М. (август 2000 г.). «Атмосферное влияние самого раннего цикла серы на Земле». Наука . 289 (5480): 756–759. Бибкод : 2000Sci...289..756F . дои : 10.1126/science.289.5480.756 . ПМИД 10926533 .
- ^ Переулок N (6 февраля 2010 г.). «Первое дыхание: миллиарды лет борьбы Земли за кислород» . Новый учёный . стр. 36–39. См. также прилагаемый график.
- ^ Корсетти Ф.А., Авраамик С.М., Пирс Д. (апрель 2003 г.). «Сложная микробиота времен снежного кома Земли: микроокаменелости из неопротерозойской формации Кингстон-Пик, Долина Смерти, США» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (8): 4399–4404. Бибкод : 2003PNAS..100.4399C . дои : 10.1073/pnas.0730560100 . ПМК 153566 . ПМИД 12682298 .
- ^ Гутчик Р.К., Перри Т.Г. (1 ноября 1959 г.). «Губки Саппингтона (Киндерхука) и их окружающая среда [Монтана]» . Журнал палеонтологии . 33 (6): 977–85 . Проверено 28 июня 2007 г.
- ^ Езда Р (1991). Известковые водоросли и строматолиты . Спрингер-Верлаг Пресс. п. 32.
- ^ Монти КЛ (1981). «Спонгиостромат против поростроматных строматолитов и онколитов» . В Монти CL (ред.). Фанерозойские строматолиты . Берлин, Гейдельберг: Springer. стр. 1–4. дои : 10.1007/978-3-642-67913-1_1 . ISBN 978-3-642-67913-1 .
- ^ Кастеллани С., Маас А., Эрикссон М.Е., Хауг Дж.Т., Хауг С., Валошек Д. (2018). «Первая запись цианобактерий в кембрийских отложениях Орстен в Швеции». Палеонтология . 61 (6): 855–880. Бибкод : 2018Palgy..61..855C . дои : 10.1111/пала.12374 . S2CID 134049042 .
- ^ «Как растения производят кислород? Спросите у цианобактерий» . Физика.орг . Наука X. 30 марта 2017 г. Проверено 26 октября 2017 г.
- ^ Крокфорд П.В., Кунцманн М., Беккер А., Хейлс Дж., Бао Х., Халверсон Г.П. и др. (20 мая 2019 г.). «Клейпул продолжил: Расширение изотопной записи осадочных сульфатов» . Химическая геология . 513 : 200–225. Бибкод : 2019ЧГео.513..200С . doi : 10.1016/j.chemgeo.2019.02.030 . ISSN 0009-2541 . S2CID 134406359 .
- ^ Ходжскисс М.С., Крокфорд П.В., Пэн Ю., Винг Б.А., Хорнер Т.Дж. (август 2019 г.). «Падение производительности положит конец Великому окислению Земли» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (35): 17207–17212. Бибкод : 2019PNAS..11617207H . дои : 10.1073/pnas.1900325116 . ПМК 6717284 . ПМИД 31405980 .
- ^ Эрреро А (2008). Флорес Э (ред.). Цианобактерии: молекулярная биология, геномика и эволюция (1-е изд.). Кайстер Академик Пресс. ISBN 978-1-904455-15-8 .
- ^ Перейти обратно: а б Килинг Пи Джей (2013). «Число, скорость и влияние пластидных эндосимбиозов на эволюцию эукариот». Ежегодный обзор биологии растений . 64 : 583–607. doi : 10.1146/annurev-arplant-050312-120144 . ПМИД 23451781 . S2CID 207679266 .
- ^ Мур К.Р., Магнабоско С., Момпер Л., Голд Д.А., Босак Т., Фурнье ГП (2019). «Расширенная рибосомальная филогения цианобактерий поддерживает глубокое размещение пластид» . Границы микробиологии . 10 : 1612. дои : 10.3389/fmicb.2019.01612 . ПМК 6640209 . ПМИД 31354692 .
- ^ Хоу С.Дж., Барбрук А.С., Нисбет Р.Э., Локхарт П.Дж., Ларкум А.В. (август 2008 г.). «Происхождение пластид» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 363 (1504): 2675–2685. дои : 10.1098/rstb.2008.0050 . ПМК 2606771 . ПМИД 18468982 .
- ^ Перейти обратно: а б Родригес-Эспелета Н., Бринкманн Х., Берей С.С., Рур Б., Бургер Г., Лёффельхардт В. и др. (июль 2005 г.). «Монофилия первичных фотосинтезирующих эукариот: зеленых растений, красных водорослей и глаукофитов» . Современная биология . 15 (14): 1325–1330. Бибкод : 2005CBio...15.1325R . дои : 10.1016/j.cub.2005.06.040 . ПМИД 16051178 .
- ^ Адл С.М., Симпсон А.Г., Лейн С.Э., Лукеш Дж., Басс Д., Баузер С.С. и др. (сентябрь 2012 г.). «Пересмотренная классификация эукариот» . Журнал эукариотической микробиологии . 59 (5): 429–493. дои : 10.1111/j.1550-7408.2012.00644.x . ПМЦ 3483872 . ПМИД 23020233 .
- ^ Прайс Д.С., Чан С.Х., Юн Х.С., Ян Э.К., Цю Х., Вебер А.П. и др. (февраль 2012 г.). «Геном Cyanophora paradoxa объясняет происхождение фотосинтеза у водорослей и растений». Наука . 335 (6070): 843–847. Бибкод : 2012Sci...335..843P . дои : 10.1126/science.1213561 . ПМИД 22344442 . S2CID 17190180 .
- ^ Перейти обратно: а б Понсе-Толедо Р.И., Дешам П., Лопес-Гарсия П., Живанович Ю., Бензерара К., Морейра Д. (февраль 2017 г.). «Ранневетвящиеся пресноводные цианобактерии у происхождения пластид» . Современная биология . 27 (3): 386–391. Бибкод : 2017CBio...27..386P . дои : 10.1016/j.cub.2016.11.056 . ПМК 5650054 . ПМИД 28132810 .
- ^ Шимпер А.Ф. (1883 г.). «О развитии хлорофилловых зерен и пятен» . Газета (на немецком языке). 41 : 105–14, 121–31, 137–46, 153–62. Архивировано из оригинала 19 октября 2013 года.
- ^ Альбертс Б (2002). «Хлоропласты и фотосинтез». Молекулярная биология клетки (4-е изд.). Нью-Йорк [ua]: Гирлянда. ISBN 978-0-8153-4072-0 .
- ^ Мерешковский С (1905). «О природе и происхождении хроматофоров в растительном мире». Biol Centralbl (на немецком языке). 25 :593-604.
- ^ Саган Л. (март 1967 г.). «О происхождении митозирующих клеток». Журнал теоретической биологии . 14 (3): 255–274. Бибкод : 1967JThBi..14..225S . дои : 10.1016/0022-5193(67)90079-3 . ПМИД 11541392 .
- ^ Шварц Р.М., Дайхофф Миссури (январь 1978 г.). «Происхождение прокариотов, эукариотов, митохондрий и хлоропластов». Наука . 199 (4327): 395–403. Бибкод : 1978Sci...199..395S . дои : 10.1126/science.202030 . ПМИД 202030 .
- ^ Арчибальд Дж. М. (август 2015 г.). «Геномные взгляды на рождение и распространение пластид» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (33): 10147–10153. Бибкод : 2015PNAS..11210147A . дои : 10.1073/pnas.1421374112 . ПМЦ 4547232 . ПМИД 25902528 .
- ^ Бланкеншип RE (октябрь 2010 г.). «Ранняя эволюция фотосинтеза» . Физиология растений . 154 (2): 434–438. дои : 10.1104/стр.110.161687 . ПМК 2949000 . ПМИД 20921158 .
- ^ Роквелл, Северная Каролина, Лагариас Дж.К., Бхаттачарья Д. (2014). «Первичный эндосимбиоз и эволюция восприятия света и кислорода у фотосинтезирующих эукариот» . Границы экологии и эволюции . 2 (66). дои : 10.3389/fevo.2014.00066 . ПМЦ 4343542 . ПМИД 25729749 .
- ^ Краткое изложение в Кавалер-Смит Т. (апрель 2000 г.). «Мембранная наследственность и ранняя эволюция хлоропластов». Тенденции в науке о растениях . 5 (4): 174–182. дои : 10.1016/S1360-1385(00)01598-3 . ПМИД 10740299 .
- ^ Новак ЕС, Мелконян М, Глекнер Г (март 2008 г.). «Последовательность хроматофорного генома Паулинеллы проливает свет на приобретение фотосинтеза эукариотами» . Современная биология . 18 (6): 410–418. Бибкод : 2008CBio...18..410N . дои : 10.1016/j.cub.2008.02.051 . ПМИД 18356055 .
- ^ Килинг П.Дж. (октябрь 2004 г.). «Разнообразие и история эволюции пластид и их хозяев» . Американский журнал ботаники . 91 (10): 1481–1493. дои : 10.3732/ajb.91.10.1481 . ПМИД 21652304 .
- ^ Арчибальд Дж. М. (январь 2009 г.). «Загадка эволюции пластид» . Современная биология . 19 (2): Р81–Р88. Бибкод : 2009CBio...19..R81A . дои : 10.1016/j.cub.2008.11.067 . ПМИД 19174147 .
- ^ Дуглас С.Е. (декабрь 1998 г.). «Эволюция пластид: происхождение, разнообразие, тенденции». Текущее мнение в области генетики и развития . 8 (6): 655–661. дои : 10.1016/S0959-437X(98)80033-6 . ПМИД 9914199 .
- ^ Рейес-Прието А., Вебер А.П., Бхаттачарья Д. (2007). «Происхождение и формирование пластид у водорослей и растений». Ежегодный обзор генетики . 41 : 147–168. дои : 10.1146/annurev.genet.41.110306.130134 . ПМИД 17600460 .
- ^ Рэйвен Дж.А., Аллен Дж.Ф. (2003). «Геномика и эволюция хлоропластов: что цианобактерии сделали для растений?» . Геномная биология . 4 (3): 209. doi : 10.1186/gb-2003-4-3-209 . ПМК 153454 . ПМИД 12620099 .
- ^ Перейти обратно: а б с Беккер А., Холланд Х.Д., Ван П.Л., Рамбл Д., Стейн Х.Дж., Ханна Дж.Л. и др. (январь 2004 г.). «Датировка повышения содержания кислорода в атмосфере». Природа . 427 (6970): 117–120. Бибкод : 2004Natur.427..117B . дои : 10.1038/nature02260 . ПМИД 14712267 . S2CID 4329872 .
- ^ Кумп Л.Р., Джуниум С., Артур М.А., Брайер А., Фаллик А., Мележик В. и др. (декабрь 2011 г.). «Изотопные доказательства массового окисления органического вещества после великого события окисления» . Наука . 334 (6063): 1694–1696. Бибкод : 2011Sci...334.1694K . дои : 10.1126/science.1213999 . ПМИД 22144465 . S2CID 27680273 .
- ^ Фралик П., Дэвис Д.В., Киссин С.А. (2002). «Возраст формации Ганфлинт, Онтарио, Канада: определение возраста U – Pb отдельных цирконов по переработанному вулканическому пеплу». Канадский журнал наук о Земле . 39 (7): 1085–1091. Бибкод : 2002CaJES..39.1085F . дои : 10.1139/e02-028 .
- ^ Перейти обратно: а б Голландия HD (июнь 2006 г.). «Окисление атмосферы и океанов» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 361 (1470): 903–915. дои : 10.1098/rstb.2006.1838 . ПМЦ 1578726 . ПМИД 16754606 .
- ^ Перейти обратно: а б с д Лайонс Т.В., Рейнхард К.Т., Планавски, Нью-Джерси (февраль 2014 г.). «Повышение содержания кислорода в раннем океане и атмосфере Земли». Природа . 506 (7488): 307–315. Бибкод : 2014Natur.506..307L . дои : 10.1038/nature13068 . ПМИД 24553238 . S2CID 4443958 .
- ^ Перейти обратно: а б Саху С.К., Планавски, Нью-Джерси, Кендалл Б., Ван Х, Ши Х, Скотт С. и др. (сентябрь 2012 г.). «Окисление океана после Мариноского оледенения». Природа . 489 (7417): 546–549. Бибкод : 2012Natur.489..546S . дои : 10.1038/nature11445 . ПМИД 23018964 . S2CID 4391882 .
- ^ Перейти обратно: а б Кэнфилд, Д.Э., Поултон, SW, генеральный директор Нарбонны (январь 2007 г.). «Поздненеопротерозойская глубоководная оксигенация океана и возникновение животной жизни» . Наука . 315 (5808): 92–95. Бибкод : 2007Sci...315...92C . дои : 10.1126/science.1135013 . ПМИД 17158290 . S2CID 24761414 .
- ^ Перейти обратно: а б с Файк Д.А., Гротцингер Дж.П., Пратт Л.М., Summons RE (декабрь 2006 г.). «Окисление Эдиакарского океана» . Природа . 444 (7120): 744–747. Бибкод : 2006Natur.444..744F . дои : 10.1038/nature05345 . ПМИД 17151665 . S2CID 4337003 .
- ^ Санчес-Баракальдо П., Риджвелл А., Рэйвен Х.А. (март 2014 г.). «Неопротерозойский переход в морском азотном цикле» . Современная биология . 24 (6): 652–657. Бибкод : 2014CBio...24..652S . дои : 10.1016/j.cub.2014.01.041 . ПМИД 24583016 . S2CID 16756351 .
- ^ Эрвин Д.Х., Лафламм М., Твидт С.М., Сперлинг Э.А., Пизани Д., Петерсон К.Дж. (ноябрь 2011 г.). «Кембрийская загадка: раннее расхождение, а затем экологический успех в ранней истории животных». Наука . 334 (6059): 1091–1097. Бибкод : 2011Sci...334.1091E . дои : 10.1126/science.1206375 . ПМИД 22116879 . S2CID 7737847 .
- ^ Поултон SW, Кэнфилд DE (2011). «Железистые условия: доминирующая особенность океана на протяжении всей истории Земли». Элементы . 7 (2): 107–112. Бибкод : 2011Элеме...7..107П . дои : 10.2113/gselements.7.2.107 . S2CID 131561186 .
- ^ Анбар А.Д., Нолл А.Х. (август 2002 г.). «Химия и эволюция протерозойского океана: бионеорганический мост?». Наука . 297 (5584): 1137–1142. Бибкод : 2002Sci...297.1137A . дои : 10.1126/science.1069651 . ПМИД 12183619 . S2CID 5578019 .
- ^ Орквишевский К.Г., Кейни А.Р. (июнь 1974 г.). «Генетическая трансформация сине-зеленой бактерии Anacystis nidulans». Архив микробиологии . 98 (1): 31–37. Бибкод : 1974ArMic..98...31O . дои : 10.1007/BF00425265 . ПМИД 4209657 . S2CID 5635245 .
- ^ Стивенс С.Е., Портер Р.Д. (октябрь 1980 г.). «Превращение в Agmenellum Quadruplicatum» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (10): 6052–6056. Бибкод : 1980PNAS...77.6052S . дои : 10.1073/pnas.77.10.6052 . ПМК 350211 . ПМИД 16592896 .
- ^ Григорьева Г, Шестаков С (1982). «Трансформация цианобактерии Synechocystis sp. 6803» . Письма FEMS по микробиологии . 13 (4): 367–70. дои : 10.1111/j.1574-6968.1982.tb08289.x .
- ^ Бернштейн Х., Бернштейн К., Мишод Р.Э. (январь 2018 г.). «Пол у микробных патогенов» . Инфекция, генетика и эволюция . 57 : 8–25. Бибкод : 2018InfGE..57....8B . дои : 10.1016/j.meegid.2017.10.024 . ПМИД 29111273 .
- ^ Перейти обратно: а б Касье-Шова С, Водор Т, Шова Ф (2016). «Сравнительная геномика рекомбинации и репарации ДНК цианобактерий: биотехнологические последствия» . Границы микробиологии . 7 : 1809. дои : 10.3389/fmicb.2016.01809 . ПМК 5101192 . ПМИД 27881980 .
- ^ «ЛТП» . Проверено 23 февраля 2021 г.
- ^ «Дерево LTP_all в формате Ньюика» . Проверено 23 февраля 2021 г.
- ^ «Примечания к выпуску LTP_12_2021» (PDF) . Проверено 23 февраля 2021 г.
- ^ «Выпуск GTDB 08-RS214» . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ "bac120_r214.sp_label" . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ «История таксонов» . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ Нэгели С. (1857 г.). Каспари Р. (ред.). «Отчет о материалах 33-го совещания немецких естествоиспытателей и врачей, проходившего в Бонне с 18 по 24 сентября 1857 года». Ботаническая газета . 15 :749-776.
- ^ Геккель Э (1867). Общая морфология организмов . Раймер, Берлин.
- ^ Чаттон Э (1925). « perplexa : амеб с защищенными спорами, паразит дафний. Размышления о биологии и филогении простейших» Pansporella . Энн. наук. Нат. Зоол . 10 (на французском языке). VII : 1–84.
- ^ Перейти обратно: а б Гуггер М.Ф., Хоффманн Л. (март 2004 г.). «Полифилия истинно ветвящихся цианобактерий (Stigonematales)» . Международный журнал систематической и эволюционной микробиологии . 54 (Часть 2): 349–357. дои : 10.1099/ijs.0.02744-0 . ПМИД 15023942 .
- ^ Ховард-Аззе М., Шамсир Л., Шеллхорн Х.Э., Гупта Р.С. (ноябрь 2014 г.). «Филогенетический анализ и молекулярные подписи, определяющие монофилетическую кладу гетероцистных цианобактерий и идентифицирующие ее ближайших родственников». Исследования фотосинтеза . 122 (2): 171–185. Бибкод : 2014PhoRe.122..171H . дои : 10.1007/s11120-014-0020-x . ПМИД 24917519 . S2CID 17745718 .
- ^ Комарек Ю., Каштовский Ю., Мареш Ю., Йохансен Ю.Р. (2014). «Таксономическая классификация цианопрокариотов (родов цианобактерий) 2014 г. с использованием полифазного подхода» (PDF) . Преслия . 86 : 295–335.
- ^ Прингсхайм ЕС (1963). Бесцветные водоросли: вклад в ( эволюционные исследования на немецком языке). Густав Фишер Верлаг.
- ^ Эузеби Ж.П. « Цианобактерии » . Список названий прокариот, имеющих номенклатуру (LPSN) . Проверено 22 января 2022 г.
- ^ «Цианобактерии» . База данных таксономии Национального центра биотехнологической информации (NCBI) . Проверено 20 марта 2021 г.
- ^ Канеко Т., Сато С., Котани Х., Танака А., Асамидзу Э., Накамура Ю. и др. (июнь 1996 г.). «Анализ последовательности генома одноклеточной цианобактерии Synechocystis sp. штамма PCC6803. II. Определение последовательности всего генома и определение потенциальных белок-кодирующих областей» . Исследование ДНК . 3 (3): 109–136. дои : 10.1093/dnares/3.3.109 . ПМИД 8905231 .
- ^ Табей Ю, Окада К, Цузуки М (апрель 2007 г.). «Sll1330 контролирует экспрессию гликолитических генов у Synechocystis sp. PCC 6803». Связь с биохимическими и биофизическими исследованиями . 355 (4): 1045–1050. дои : 10.1016/j.bbrc.2007.02.065 . ПМИД 17331473 .
- ^ Рокап Г., Лаример Ф.В., Ламердин Дж., Малфатти С., Чейн П., Альгрен Н.А. и др. (август 2003 г.). «Дивергенция генома двух экотипов Prochromococcus отражает дифференциацию океанических ниш» . Природа . 424 (6952): 1042–1047. Бибкод : 2003Natur.424.1042R . дои : 10.1038/nature01947 . ПМИД 12917642 . S2CID 4344597 .
- ^ Дюфрен А., Саланубат М., Партенский Ф., Артигенав Ф., Аксманн И.М., Барбе В. и др. (август 2003 г.). «Последовательность генома цианобактерии Prochromococcus marinus SS120, почти минимального оксифототрофного генома» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (17): 10020–10025. Бибкод : 2003PNAS..10010020D . дои : 10.1073/pnas.1733211100 . ЧВК 187748 . ПМИД 12917486 .
- ^ Хердман М., Жанвье М., Риппка Р., Станье Р.Ю. (1979). «Размер генома цианобактерий» . Журнал общей микробиологии . 111 : 73–85. дои : 10.1099/00221287-111-1-73 .
- ^ Кинтана Н., Ван дер Кой Ф., Ван де Ри, доктор медицинских наук, Вошол Г.П., Верпоорт Р. (август 2011 г.). «Возобновляемая энергия из цианобактерий: оптимизация производства энергии путем разработки метаболических путей» . Прикладная микробиология и биотехнология . 91 (3): 471–490. дои : 10.1007/s00253-011-3394-0 . ПМК 3136707 . ПМИД 21691792 .
- ^ Чен X, Лоуренс Дж.М., Вей Л.Т., Шертель Л., Цзин К., Виньолини С. и др. (июль 2022 г.). «3D-печатные иерархические столбчатые электроды для высокопроизводительного полуискусственного фотосинтеза» . Природные материалы . 21 (7): 811–818. Бибкод : 2022NatMa..21..811C . дои : 10.1038/s41563-022-01205-5 . PMID 35256790 . S2CID 247255146 .
- ^ «Сине-зеленые бактерии могут помочь генерировать «зеленое» электричество» , The Hindu , 21 июня 2010 г.
- ^ «Джоуль выигрывает ключевой патент на ГМО-цианобактерии, которые создают топливо из солнечного света, CO2 и воды: Biofuels Digest» . 14 сентября 2010 года . Проверено 10 августа 2022 г.
- ^ Дэн, доктор медицинских наук, Коулман-младший (февраль 1999 г.). «Синтез этанола методом генной инженерии в цианобактериях» . Прикладная и экологическая микробиология . 65 (2): 523–528. Бибкод : 1999ApEnM..65..523D . дои : 10.1128/АЕМ.65.2.523-528.1999 . ПМК 91056 . ПМИД 9925577 .
- ^ Лян Ф., Линдберг П., Линдблад П. (20 ноября 2018 г.). «Инженерия фотоавтотрофной фиксации углерода для улучшения роста и продуктивности» . Устойчивая энергетика и топливо . 2 (12): 2583–2600. дои : 10.1039/C8SE00281A .
- ^ Руссу С., Альбергати А., Лян Ф., Линдблад П. (июнь 2021 г.). «Инженерные цианобактерии с дополнительной сверхэкспрессией выбранных ферментов Кальвина-Бенсона-Башама демонстрируют дальнейшее увеличение производства этанола» . Метаболические инженерные коммуникации . 12 : e00161. дои : 10.1016/j.mec.2021.e00161 . ПМЦ 7820548 . ПМИД 33520653 . S2CID 231727024 .
- ^ Чой Х., Маскух С.Дж., Вилла Ф.А., Байрум Т., Тисдейл М.Э., Смит Дж.Э. и др. (май 2012 г.). «Хонауцины AC, мощные ингибиторы воспаления и чувства бактериального кворума: синтетические производные и взаимосвязь структура-активность» . Химия и биология . 19 (5): 589–598. doi : 10.1016/j.chembiol.2012.03.014 . ПМК 3361693 . ПМИД 22633410 .
- ^ Фришкорн К. (19 июня 2017 г.). «Нужно вылечить сердечный приступ? Попробуйте фотосинтез» . Смитсоновский институт . Проверено 20 мая 2021 г.
- ^ Чон Й, Чо С.Х., Ли Х., Чхве Х.К., Ким Д.М., Ли К.Г. и др. (ноябрь 2020 г.). «Текущее состояние и будущие стратегии по увеличению производства вторичных метаболитов цианобактерий» . Микроорганизмы . 8 (12): 1849. doi : 10.3390/microorganisms8121849 . ПМЦ 7761380 . ПМИД 33255283 .
- ^ Ньюсом АГ, Калвер, Калифорния, ван Бримен РБ (июль 2014 г.). «Палитра природы: поиск натуральных синих красителей». Журнал сельскохозяйственной и пищевой химии . 62 (28): 6498–6511. дои : 10.1021/jf501419q . ПМИД 24930897 .
- ^ Версо С., Баке М., Лехто К., де Вера Дж.П., Ротшильд Л.Дж. , Билли Д. (2016). «Устойчивое поддержание жизни на Марсе – потенциальная роль цианобактерий» . Международный журнал астробиологии . 15 (1): 65–92. Бибкод : 2016IJAsB..15...65В . дои : 10.1017/S147355041500021X .
- ^ Сполаоре П., Жоаннис-Кассан С., Дюран Э., Исамберт А. (февраль 2006 г.). «Коммерческое применение микроводорослей». Журнал бионауки и биоинженерии . 101 (2): 87–96. дои : 10.1263/jbb.101.87 . ПМИД 16569602 . S2CID 16896655 .
- ^ Кристаки Э., Флору-Панери П., Бонос Э. (декабрь 2011 г.). «Микроводоросли: новый ингредиент в питании». Международный журнал пищевых наук и питания . 62 (8): 794–799. дои : 10.3109/09637486.2011.582460 . ПМИД 21574818 . S2CID 45956239 .
- ^ Ку CS, Фам Техас, Пак Ю, Ким Б, Шин М.С., Кан И, Ли Дж (апрель 2013 г.). «Съедобные сине-зеленые водоросли снижают выработку провоспалительных цитокинов путем ингибирования пути NF-κB в макрофагах и спленоцитах» . Biochimica et Biophysical Acta (BBA) – Общие предметы . 1830 (4): 2981–2988. дои : 10.1016/j.bbagen.2013.01.018 . ПМЦ 3594481 . ПМИД 23357040 .
- ^ Мишурцова Л, Шкрованкова С, Самек Д, Амброжова Ю, Мачу Л (2012). «Польза для здоровья полисахаридов водорослей в питании человека». Достижения в области исследований продуктов питания и питания . 66 :75–145. дои : 10.1016/B978-0-12-394597-6.00003-3 . ISBN 978-0-12-394597-6 . ПМИД 22909979 .
- ^ Тебо Л., Лесн Дж., Бутен Дж.П. (1995). «[Цианобактерии, их токсины и риски для здоровья]». Тропическая медицина . 55 (4): 375–380. ПМИД 8830224 .
- ^ Перейти обратно: а б «Сине-зеленые водоросли (цианобактерии) и их токсины. Питьевая вода» . 30 мая 2008 г. Архивировано из оригинала 30 мая 2008 г. . Проверено 10 августа 2022 г.
- ^ «Вредное цветение озера Атитлан, Гватемала» . Earthobservatory.nasa.gov . 3 декабря 2009 года . Проверено 10 августа 2022 г. из Земной обсерватории НАСА ,
- ^ Вейрих К.А., Миллер Т.Р. (январь 2014 г.). «Вредное цветение пресноводных водорослей: токсины и здоровье детей». Современные проблемы детского и подросткового здравоохранения . 44 (1): 2–24. дои : 10.1016/j.cppeds.2013.10.007 . ПМИД 24439026 .
- ^ «Профессор Линда Лоутон» . rgu-repository.worktribe.com . Проверено 25 июня 2021 г.
- ^ Коллер Т.А., Дулин Дж.В., Хейни Дж.Ф., Мерби А.Дж., Вест К.Г., Фаррар Х.Э. и др. (2009). «Кластер бокового амиотрофического склероза в Нью-Гэмпшире: возможная роль цветения токсичных цианобактерий». Боковой амиотрофический склероз . 10 (Приложение 2): 101–108. дои : 10.3109/17482960903278485 . ПМИД 19929741 . S2CID 35250897 .
- ^ Кокс П.А., Ричер Р., Меткалф Дж.С., Банак С.А., Кодд Г.А., Брэдли В.Г. (2009). «Воздействие цианобактерий и BMAA из пыли пустыни: возможная связь со спорадическим БАС среди ветеранов войны в Персидском заливе». Боковой амиотрофический склероз . 10 (Приложение 2): 109–117. дои : 10.3109/17482960903286066 . ПМИД 19929742 . S2CID 1748627 .
- ^ Перейти обратно: а б с д и ж Главный ДК (2006). «Цветение токсичных водорослей» (PDF) . Ветеринарный патологоанатом, Южный Перт . agric.wa.gov.au. Архивировано из оригинала (PDF) 1 декабря 2014 года . Проверено 18 ноября 2014 г.
- ^ Мэй В., Бейкер Х. (1978). «Уменьшение количества токсичных водорослей на фермерских плотинах с помощью железных квасцов». Техн. Бык . 19 : 1–16.
- ^ Паерл Х.В., Хьюсман Дж. (апрель 2008 г.). «Климат. Цветение любит жару». Наука . 320 (5872): 57–58. дои : 10.1126/science.1155398 . ПМИД 18388279 . S2CID 142881074 .
- ^ Ситоки Л., Курмайер Р., Ротт Э. (июль 2012 г.). «Пространственные вариации состава фитопланктона, биообъема и результирующих концентраций микроцистина в заливе Ньянза (озеро Виктория, Кения)» . Гидробиология . 691 (1): 109–122. дои : 10.1007/s10750-012-1062-8 . ПМЦ 3968937 . ПМИД 24683268 .
- ^ Меткалф Дж.С., Банак С.А., Пауэлл Дж.Т., Тимм Ф.Дж., Марч С.Дж., Брэнд Л.Е., Кокс Пенсильвания (2018). «Реакция общественного здравоохранения на цветение токсичных цианобактерий: перспективы событий во Флориде в 2016 году» . Водная политика . 20 (5): 919–932. дои : 10.2166/wp.2018.012 . S2CID 88502303 .
- ^ Перейти обратно: а б с Кавиччиоли Р., Риппл В.Дж., Тиммис К.Н., Азам Ф., Баккен Л.Р., Бэйлис М. и др. (сентябрь 2019 г.). «Предупреждение ученых человечеству: микроорганизмы и изменение климата» . Обзоры природы. Микробиология . 17 (9): 569–586. дои : 10.1038/s41579-019-0222-5 . ПМЦ 7136171 . ПМИД 31213707 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Виссер П.М., Верспаген Дж.М., Сандрини Дж., Сталь Л.Дж., Маттейс Х.К., Дэвис Т.В. и др. (апрель 2016 г.). «Как рост выбросов CO 2 и глобальное потепление могут стимулировать вредоносное цветение цианобактерий» . Вредные водоросли . 54 : 145–159. дои : 10.1016/j.hal.2015.12.006 . ПМИД 28073473 .
- ^ Уолсби А.Э., Хейс П.К., Бойе Р., Сталь Л.Дж. (июль 1997 г.). «Селективное преимущество плавучести, обеспечиваемое газовыми пузырьками для планктонных цианобактерий в Балтийском море». Новый фитолог . 136 (3): 407–417. дои : 10.1046/j.1469-8137.1997.00754.x . ПМИД 33863010 .
- ^ Леман П.В., Куробе Т., Лесмейстер С., Бакса Д., Тунг А., Тех С.Дж. (март 2017 г.). «Воздействие сильной засухи 2014 года на цветение Microcystis в устье Сан-Франциско» . Вредные водоросли . 63 : 94–108. Бибкод : 2017HAlga..63...94L . дои : 10.1016/j.hal.2017.01.011 . ПМИД 28366405 .
- ^ Перейти обратно: а б Сандрини Дж., Джи Икс, Верспаген Дж.М., Танн Р.П., Slot PC, Luimstra VM и др. (август 2016 г.). «Быстрая адаптация вредных цианобактерий к повышению уровня CO2» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (33): 9315–9320. Бибкод : 2016PNAS..113.9315S . дои : 10.1073/pnas.1602435113 . ПМЦ 4995953 . ПМИД 27482094 .
Атрибуция
![]() В эту статью включен текст , доступный по лицензии CC BY 2.5 .
В эту статью включен текст , доступный по лицензии CC BY 2.5 .
Дальнейшее чтение
[ редактировать ]- Джиллиан С. (1997). Природный суперпродукт: революция сине-зеленых водорослей (первое изд.). Ньюлиф. ISBN 978-0-7522-0569-4 .
- Сэвидж М (1994). Проект «Миллениал»: колонизация Галактики за восемь простых шагов . Литтл Браун и Ко ISBN 978-0-316-77163-4 .
- Фогг Дж.Э., Стюарт В.Д., Фэй П., Уолсби А.Е. (1973). Сине-зеленые водоросли . Лондон и Нью-Йорк: Академическая пресса . ISBN 978-0-12-261650-1 .
- «Архитекторы земной атмосферы. Знакомство с цианобактериями» . Калифорнийский университет в Беркли . 3 февраля 2006 г.
- Уиттон, Б.А. (2002). «Тип Cyanophyta (Цианобактерии)». Пресноводная водорослевая флора Британских островов . Кембридж: Издательство Кембриджского университета . ISBN 978-0-521-77051-4 .
- Пятидесятница А, Франке У (2010). «Фотосинтез и кальцификация строматолитовой пресноводной цианобактерии Rivularia » . Европейский журнал психологии . 45 (4): 345–53. Бибкод : 2010EJPhy..45..345P . дои : 10.1080/09670262.2010.492914 .
- Уиттон Б.А., Поттс М., ред. (2000). Экология цианобактерий: их разнообразие во времени и пространстве . Спрингер. ISBN 978-0-7923-4735-4 .
- «От микроводорослей до голубой нефти» . Обзор ПарижТех . Декабрь 2011. Архивировано из оригинала 17 апреля 2016 года . Проверено 2 марта 2012 г.

