Белок

Белки представляют собой большие биомолекулы и макромолекулы , которые составляют одну или несколько длинных цепочек аминокислотных остатков . Белки выполняют широкий спектр функций внутри организмов, включая катализ метаболических реакций , репликацию ДНК , реагирование на стимулы , обеспечивая структуру клеткам и организмам , а также транспортирующие молекулы из одного места в другое. Белки отличаются друг от друга, главным образом, по их последовательности аминокислот, что продиктовано нуклеотидной последовательности их генов , и которая обычно приводит к складыванию белков в определенную трехмерную структуру , которая определяет его активность.
Линейная цепь аминокислотных остатков называется полипептидом . Белок содержит по крайней мере один длинный полипептид. Короткие полипептиды, содержащие менее 20–30 остатков, редко считаются белками и обычно называют пептидами . Индивидуальные аминокислотные остатки связываются вместе пептидными связями и прилегающими аминокислотными остатками. Последовательность генетическом аминокислотных остатков в белке определяется последовательности гена , которая кодируется в коде . В целом, генетический код определяет 20 стандартных аминокислот; Но в некоторых организмах генетический код может включать селеноцистеин и - в определенных археи - пирролизин . Вскоре после или даже во время синтеза остатки в белке часто химически модифицируются посттрансляционной модификацией , которая изменяет физические и химические свойства, складывание, стабильность, активность и, в конечном счете, функцию белков. Некоторые белки имеют прикрепленные непептидные группы, которые можно назвать протезными группами или кофакторами . Белки также могут работать вместе для достижения определенной функции, и они часто связывают, чтобы сформировать стабильный белковые комплексы .
После образования белки существуют только в течение определенного периода, а затем деградируют и переработаны механизмом клеток в процессе текучести кадров . Срок службы белка измеряется с точки зрения его полураспада и охватывает широкий диапазон. Они могут существовать в течение нескольких минут или лет со средней продолжительностью жизни 1–2 дня в клетках млекопитающих. Аномальные или неправильно свернутые белки деградируются быстрее либо из -за того, что они нацелены на разрушение, либо из -за нестабильных.
Как и другие биологические макромолекулы, такие как полисахариды и нуклеиновые кислоты , белки являются важными частями организмов и участвуют практически во всех процессах внутри клеток . Многие белки - это ферменты , которые катализируют биохимические реакции и имеют жизненно важное значение для метаболизма . Белки также имеют структурные или механические функции, такие как актин и миозин в мышцах и белки в цитоскелете , которые образуют систему каркасов , которая поддерживает форму клеток. Другие белки важны для передачи сигналов клеток, иммунных ответов , клеточной адгезии и клеточного цикла . У животных белки необходимы в рационе , чтобы обеспечить незаменимые аминокислоты , которые нельзя синтезировать . Пищеварение разбивает белки для метаболического использования.
История и этимология
Открытие и ранние исследования
Белки были изучены и признаны с 1700 -х годов Антуаном Фуркрой и другими, [ 1 ] [ 2 ] который часто коллективно называл их « альбуминами », или «альбоминовыми материалами» ( Eiweisskörper , на немецком языке). [ 2 ] глютен был впервые отделен от пшеницы в опубликованных исследованиях около 1747 года, а затем решил существовать во многих растениях. Например, [ 1 ] В 1789 году Антуан Фуркрой признал три различных разновидности животных белков: альбумин , фибрин и желатин . [ 3 ] Овощные (растительные) белки, изученные в конце 1700 -х и начале 1800 -х годов, включали глютен , альбумин растений , глиадин и богобомин . [ 1 ]
Белки были впервые описаны голландским химиком Джерардом Йоханнесом Малдером и назвали шведским химиком Йеном Джейкобом Берзелиусом в 1838 году. [ 4 ] [ 5 ] [ Лучший источник необходим ] Малдер провел элементарный анализ общих белков и обнаружил, что почти все белки имели одинаковую эмпирическую формулу , C 400 H 620 N 100 O 120 P 1 S 1 . [ 6 ] Он пришел к ошибочному выводу, что они могут состоять из одного типа (очень большой) молекулы. Термин «белок» для описания этих молекул был предложен ассоциированным Берзелиусом Малдера; белок получен из греческого слова πρώτειος ( протеис ), что означает «первичный», [ 7 ] "В лидере", или "Стоя впереди", [ 2 ] + -in . Малдер продолжал идентифицировать продукты разложения белка, такие как аминокислотный лейцин, для которого он обнаружил (почти правильную) молекулярную массу 131 Да . [ 6 ]
Ранние ученые по питанию, такие как немецкий Карл фон Войт, считали, что белок является наиболее важным питательным веществом для поддержания структуры организма, потому что обычно считалось, что «плоть делает плоть». [ 8 ] Около 1862 года Карл Генрих Риттхаузен выделял аминокислотную глутаминовую кислоту . [ 9 ] Томас Берр Осборн собрал подробный обзор растительных белков на сельскохозяйственной экспериментальной станции Коннектикута . Затем работа с [ нужно разъяснения ] Lafayette Mendel и применение закона Либига минимума , который утверждает, что рост ограничен самым скудным ресурсом, для кормления лабораторных крыс, эссенционные аминокислоты с питанием были установлены . Работа была продолжена и сообщена Уильямом Каммингом Роуз .
Трудность очищающих белков в больших количествах затрудняла изучение ранних биохимиков белка. Следовательно, ранние исследования были сосредоточены на белках, которые можно очистить в больших количествах, в том числе из крови, яичных белков и различных токсинов, а также пищеварительных и метаболических ферментов, полученных из бойни. [ нужно разъяснения ] В 1950 -х годах компания Armor Hot Dog Company очистила 1 кг чистой бычьей поджелудочной железы, рибонуклеаза А и сделала ее свободно доступной для ученых; Этот жест помог рибонуклеазу стать основной целью для биохимического исследования в течение следующих десятилетий. [ 6 ]
Полипептиды

Понимание белков в качестве полипептидов или цепочек аминокислот, получило работу Франца Хофмайстера и Германа Эмиля Фишера в 1902 году. [ 10 ] [ 11 ] Центральная роль белков как ферментов в живых организмах, которые катализировали реакции, не была полностью оценена до 1926 года, когда Джеймс Б. Самнер показал, что ферментативная уреаза на самом деле является белком. [ 12 ]
Линусу Полинг приписывают успешное прогнозирование регулярных белковых вторичных структур на основе водородной связи , идея, впервые выдвинутая Уильямом Астбери в 1933 году. [ 13 ] Позже работа Уолтера Каузмана по денатурации , [ 14 ] [ 15 ] Основано частично на предыдущих исследованиях Кадж Линдерстром-Ланг , [ 16 ] Внесли понимание складывания белка и структуры, опосредованного гидрофобными взаимодействиями .
аминокислотную цепь, Первым белком, имеющим секвенированную был инсулин Фредериком Сэнгером , в 1949 году. Сэнгер правильно определил аминокислотную последовательность инсулина, что убедительно демонстрируя, что белки состояли из линейных полимеров аминокислот, а не разветвленных цепей, коллоиды или Циклолы . [ 17 ] Он получил Нобелевскую премию за это достижение в 1958 году. [ 18 ] Христианского Анфинсена Исследования о окислительном процессе складывания рибонуклеазы А, за что он выиграл Нобелевскую премию в 1972 году, укрепил термодинамическую гипотезу складывания белка, согласно которой сложенная форма белка представляет минимум свободной энергии . [ 19 ] [ 20 ]
Структура

С развитием рентгеновской кристаллографии стало возможным определять как белковые структуры, а также их последовательности. [ 21 ] Первыми белковыми структурами, которые должны быть решены, были гемоглобин от Макса Перутца и миоглобина от Джона Кендрю в 1958 году. [ 22 ] [ 23 ] Использование компьютеров и увеличение вычислительной мощности также поддерживало секвенирование сложных белков. В 1999 году Роджер Корнберг преуспел в секвенировании очень сложной структуры РНК-полимеразы с использованием рентгеновских лучей высокой интенсивности от синхротронов . [ 21 ]
С тех пор криоэлектронная микроскопия (крио-ЭМ) крупных макромолекулярных сборок [ 24 ] был разработан. Cryo-EM использует образцы белка, которые заморожены, а не кристаллы, и балки электронов , а не рентген. Это наносит меньший ущерб выборке, позволяя ученым получать больше информации и анализировать более крупные структуры. [ 21 ] Вычислительная структура белка прогнозирование малых белков структурных доменов [ 25 ] также помог исследователям подойти к разрешению белковых структур на уровне атомного уровня. По состоянию на апрель 2024 года [update], Банк данных белка содержит 181 018 рентгеновских лучей, 19 809 EM и 12 697 ЯМР -структур. [ 26 ]
Классификация
Белки в основном классифицируются по последовательности и структуре, хотя обычно используются другие классификации. Специально для ферментов Система номеров ЕС обеспечивает функциональную схему классификации. Точно так же генная онтология классифицирует как гены, так и белки по их биологической и биохимической функции, а также по их внутриклеточному расположению.
Сходство последовательности используется для классификации белков как с точки зрения эволюционного, так и функционального сходства. Это может использовать либо целые белки, либо белковые домены , особенно в многодоменных белках . Белковые домены позволяют классифицировать белок комбинацией последовательности, структуры и функции, и их можно объединить по -разному. В раннем исследовании 170 000 белков было назначено около двух третей, по крайней мере, один домен, при этом более крупные белки содержали больше доменов (например, белки более 600 аминокислот, имеющие в среднем более 5 доменов). [ 27 ]
Биохимия

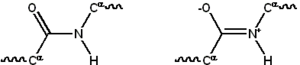
Большинство белков состоят из линейных полимеров , построенных из серии до 20 различных L -аминокислот . Все протеиногенные аминокислоты обладают общими структурными особенностями, в том числе α-углерод, которым аминогрузка , карбоксильная группа и переменная боковая цепь связаны с . Только пролин отличается от этой основной структуры, поскольку он содержит необычное кольцо для группы аминов N, которая заставляет ко-н-амидную часть в фиксированную конформацию. [ 28 ] Боковые цепи стандартных аминокислот, подробно описанные в списке стандартных аминокислот , имеют большое разнообразие химических структур и свойств; Это комбинированный эффект всех аминокислотных боковых цепей в белке, который в конечном итоге определяет его трехмерную структуру и его химическую реактивность. [ 29 ] Аминокислоты в полипептидной цепи связаны пептидными связями . После связи в белковой цепи, отдельная аминокислота называется остатком, а связанный серия углерода, азота и атомов кислорода известна как основная цепь или ливня белка. [ 30 ] : 19
Пептидная связь имеет две резонансные формы, которые вносят вклад некоторый характер с двойным связью и ингибируют вращение вокруг его оси, так что альфа-углероды являются примерно копланаром . Два других двугральных угла в пептидной связи определяют локальную форму, принятую белковой основой. [ 30 ] : 31 Конец со свободной аминогрузкой известен как N-конце или амино-концерн, тогда как конец белка со свободной карбоксильной группой известен как C-конце или карбокси (последовательность белка написана из N- Термин в C-конце, слева направо).
Слова белок , полипептид и пептид являются немного неоднозначными и могут перекрываться по значению. Белок обычно используется для обозначения полной биологической молекулы в стабильной конформации , тогда как пептид обычно зарезервирован для коротких аминокислотных олигомеров, которые часто не хватает стабильной трехмерной структуры. Но граница между ними недостаточно определена и обычно лежит около 20–30 остатков. [ 31 ] Полипептид может относиться к любой отдельной линейной цепи аминокислот, обычно независимо от длины, но часто подразумевает отсутствие определенной конформации .
Взаимодействия
Белки могут взаимодействовать со многими типами молекул, в том числе с другими белками , с липидами , с углеводами и с ДНК . [ 32 ] [ 33 ] [ 30 ] [ 34 ]
Изобилие в клетках
Было подсчитано, что бактерии среднего размера содержат около 2 миллионов белков на клетку (например, E. coli и staphylococcus aureus ). Меньшие бактерии, такие как микоплазма или спирохеты , содержат меньше молекул по порядку от 50 000 до 1 миллиона. Напротив, эукариотические клетки больше и, таким образом, содержат гораздо больше белка. Например, , дрожжевые по оценкам клетки содержат около 50 миллионов белков и клеток человека по порядку от 1 до 3 миллиардов. [ 35 ] Концентрация отдельных белковых копий варьируется от нескольких молекул на клетку до 20 миллионов. [ 36 ] Не все гены, кодирующие белки, экспрессируются в большинстве клеток, и их число зависит от типа клеток и внешних стимулов. Например, из около 20 000 белков, кодируемых геномом человека, только 6000 обнаруживаются в лимфобластоидных клетках. [ 37 ]
Синтез
Биосинтез

Белки собираются из аминокислот с использованием информации, кодируемой в генах. Каждый белок имеет свою уникальную аминокислотную последовательность, которая указывает нуклеотидной последовательности гена, кодирующего этот белок. Генетический код представляет собой набор трехнуклеотидных наборов, называемых кодонами , и каждая трехнуклеотидная комбинация обозначает аминокислоту, например, AUG ( аденин - урацил - гуанин ) является кодом метионина . Поскольку ДНК содержит четыре нуклеотида, общее количество возможных кодонов составляет 64; Следовательно, в генетическом коде есть некоторая избыточность, причем некоторые аминокислоты указаны более чем одним кодоном. [ 34 ] : 1002–42 Гены, кодируемые в ДНК, сначала транскрибируются в РНК-досенджер (мРНК) белками, такими как РНК-полимераза . Большинство организмов затем обрабатывают пре-мРНК (также известную как первичную транскрипцию ), используя различные формы посттранскрипционной модификации, образуя зрелую мРНК, которая затем используется в качестве шаблона для синтеза белка рибосомой . В прокариотах мРНК может быть либо использована, как только она будет получена, либо была связана рибосомой после того, как отошла от нуклеоида . Напротив, эукариоты делают мРНК в клеточном ядре , а затем транслируют ее через ядерную мембрану в цитоплазму , где синтез белка затем происходит . Скорость синтеза белка выше у прокариот, чем у эукариот, и может достигать до 20 аминокислот в секунду. [ 38 ]
Процесс синтеза белка из матрицы мРНК известен как трансляция . МРНК загружается на рибосому и читается три нуклеотида одновременно, сопоставляя каждый кодон с его , антикодоном, расположенным на молекуле РНК переноса которая несет аминокислоту, соответствующую кодону, который он распознает. Фермент аминоациль -синтетаза «заряжает» молекулы тРНК с правильными аминокислотами. Растущий полипептид часто называют зарождающейся цепью . Белки всегда биосинтезируются от N-конца до C-конца . [ 34 ] : 1002–42
Размер синтезированного белка может быть измерен по количеству аминокислот, которые он содержит, и его общей молекулярной массой , которая обычно сообщается в единицах далтонов (синонимично с атомными массовыми единицами ) или производной килодалтон (KDA). Средний размер белка увеличивается от археи до бактерий к эукариоте (283, 311, 438 остатков и 31, 34, 49 кДа соответственно) из -за большего числа белковых доменов, составляющих белки в более высоких организмах. [ 39 ] Например, дрожжевые белки в среднем имеют длину 466 аминокислот и 53 кДа в массе. [ 31 ] Самыми крупными известными белками являются Titins , компонент мышечной саркории , с молекулярной массой почти 3000 кДа и общей длиной почти 27 000 аминокислот. [ 40 ]
Химический синтез

Короткие белки также могут быть синтезированы химически с помощью семейства методов, известных как синтез пептидов , которые зависят от методов органического синтеза , таких как химическое лигирование, для производства пептидов с высоким выходом. [ 41 ] Химический синтез позволяет внедрить не натуральные аминокислоты в полипептидные цепи, такие как прикрепление флуоресцентных зондов к аминокислотным цепи. [ 42 ] Эти методы полезны при лабораторной биохимии и клеточной биологии , хотя, как правило, не для коммерческих применений. Химический синтез неэффективен для полипептидов длиннее, чем около 300 аминокислот, и синтезированные белки могут не с готовностью предполагать свою собственную третичную структуру . Большинство методов химического синтеза переходят от С-конца к N-концу, противоположной биологической реакции. [ 43 ]
Структура

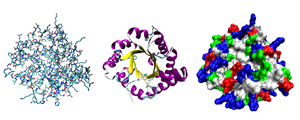
Большинство белков складываются в уникальные 3D -структуры. Форма, в которой белок естественным образом складывается, известна как его нативная конформация . [ 30 ] : 36 Хотя многие белки могут складываться без посторонней помощи, просто с помощью химических свойств их аминокислот, другие требуют помощи молекулярных шаперонов , чтобы складываться в свои родные состояния. [ 30 ] : 37 Биохимики часто относятся к четырем различным аспектам структуры белка: [ 30 ] : 30–34
- Первичная структура : аминокислотная последовательность . Белок - это полиамид .
- Вторичная структура : регулярно повторяя локальные структуры, стабилизированные водородными связями . Наиболее распространенными примерами являются α-спираль , β-лист и повороты . Поскольку вторичные структуры являются локальными, в одной и той же молекуле белка могут присутствовать многие области разной вторичной структуры.
- Третичная структура : общая форма молекулы одного белка; Пространственная связь вторичных структур друг к другу. Третичная структура, как правило, стабилизируется нелокальными взаимодействиями, чаще всего образование гидрофобного ядра , а также посредством солевых мостов , водородных связей, дисульфидных связей и даже посттрансляционных модификаций . Термин «третичная структура» часто используется в качестве синонима с термином сгиба . Третичная структура - это то, что контролирует основную функцию белка.
- Четвертичная структура : структура, образованная несколькими молекулами белка (полипептидные цепи), обычно называемую белковыми субъединицами в этом контексте, которые функционируют как единый белковый комплекс .
- КВИНАРНАЯ СТРУКТУРА : сигнатуры поверхности белка, которые организуют переполненный клеточный интерьер. Квиарная структура зависит от переходных, но существенных макромолекулярных взаимодействий, которые происходят внутри живых клеток.
Белки не являются полностью жесткими молекулами. В дополнение к этим уровням структуры белки могут смещаться между несколькими родственными структурами, пока они выполняют свои функции. В контексте этих функциональных перестановки эти третичные или четвертичные структуры обычно называют « конформациями », а переходы между ними называются конформационными изменениями. Такие изменения часто вызывают связывание молекулы субстрата фермента с активным сайтом или физической областью белка, которая участвует в химическом катализе. В растворе белки также подвергаются изменению структуры посредством термической вибрации и столкновения с другими молекулами. [ 34 ] : 368–75

Белки могут быть неофициально разделены на три основных класса, которые коррелируют с типичными третичными структурами: глобулярные белки , волокнистые белки и мембранные белки . Почти все глобулярные белки растворимы , а многие - ферменты. Фиброзные белки часто являются структурными, такими как коллаген , основной компонент соединительной ткани или кератина , белковой компонент волос и ногтей. Мембранные белки часто служат рецепторами или предоставляют каналы для полярных или заряженных молекул для прохождения через клеточную мембрану . [ 34 ] : 165–85
Особый случай внутримолекулярных водородных связей в белках, плохо защищенных от водной атаки и, следовательно, способствуя их собственному обезвоживанию , называются дегидронами . [ 44 ]
Белковые домены
Многие белки состоят из нескольких белковых доменов , т.е. сегменты белка, которые складываются в различные структурные единицы. Домены обычно также имеют специфические функции, такие как ферментативная активность (например, киназа ) или они служат связывающими модулями (например, домен SH3 связывается с пролиновыми последовательностями в других белках).
Мотив последовательности
Короткие аминокислотные последовательности в белках часто действуют как сайты распознавания для других белков. [ 45 ] Например, домены SH3 обычно связываются с короткими мотивами PXXP (т.е. 2 пролины [P], разделенные двумя неопределенными аминокислотами [x], хотя окружающие аминокислоты могут определять точную специфичность связывания). Многие такие мотивы были собраны в базе данных эукариотических линейных мотивов (ELM).
Белковая топология
Топология белка описывает запутанность основы и расположение контактов в сложенной цепи. [ 46 ] Две теоретические основы теории узлов и топологии схемы были применены для характеристики топологии белка. Способность описать топологию белка открывает новые пути для инженерии белков и фармацевтического развития и добавляет к нашему пониманию заболеваний, неправильно сводящих белок, таких как нервно -мышечные расстройства и рак.
Клеточные функции
Белки являются главными субъектами в клетке, которые, как говорят, выполняют обязанности, указанные информацией, кодируемой в генах. [ 31 ] За исключением определенных типов РНК , большинство других биологических молекул являются относительно инертными элементами, на которых действуют белки. Белки составляют половину сухого веса клеток Coli Escherichia , тогда как другие макромолекулы, такие как ДНК и РНК, составляют только 3% и 20% соответственно. [ 47 ] Набор белков, экспрессируемых в конкретном типе клеток или клеток, известен как его протеом .

Главной характеристикой белков, которые также позволяют их разнообразному набору функций, является их способность связывать другие молекулы конкретно и тесно и тесно. Область белка, ответственная за связывание другой молекулы, известна как сайт связывания и часто является депрессией или «карманом» на молекулярной поверхности. Эта способность связывания опосредована третичной структурой белка, которая определяет карман сайта связывания и химическими свойствами окружающих аминокислотных боковых цепей. Связывание белка может быть чрезвычайно напряженным и специфичным; Например, белок ингибитора рибонуклеазы связывается с ангиогенином человека с субфемтомолярной постоянной диссоциации (<10 −15 M) но вообще не связывается с его амфибийской гомологом онназы (> 1 м). Чрезвычайно незначительные химические изменения, такие как добавление одной метильной группы к партнеру по связыванию, иногда могут быть достаточно для почти удаления связывания; Например, аминоациль -синтетаза, специфичная для аминокислотного валина , дискриминирует от очень похожей боковой цепи аминокислотного изолецина . [ 48 ]
Белки могут связываться с другими белками, а также с малой молекулярными субстратами. Когда белки связываются конкретно с другими копиями той же молекулы, они могут олигомеризировать с образованием фибриллов; Этот процесс часто встречается в структурных белках, которые состоят из шаровых мономеров, которые самимника с образованием жестких волокон. Белковые взаимодействия также регулируют ферментативную активность, контрольную прогрессию через клеточный цикл и позволяют сборке крупных белковых комплексов , которые выполняют много тесно связанных реакций с общей биологической функцией. Белки также могут связываться или даже интегрироваться в клеточные мембраны. Способность связывать партнеров индуцировать конформационные изменения в белках позволяет создавать чрезвычайно сложные сигнальные сети. [ 34 ] : 830–49 Поскольку взаимодействия между белками обратимы и в значительной степени зависят от наличия различных групп партнерских белков с образованием агрегатов, которые способны выполнять дискретные наборы функции, изучение взаимодействия между конкретными белками является ключом к пониманию важных аспектов клеточной функции и, в конечном счете, свойства, которые различают конкретные типы клеток. [ 49 ] [ 50 ]
Ферменты
Самая известная роль белков в клетке-это как ферменты , которые катализируют химические реакции. Ферменты обычно очень специфичны и ускоряют только одну или несколько химических реакций. Ферменты выполняют большинство реакций, связанных с метаболизмом , а также манипулируют ДНК в таких процессах, как репликация ДНК , репарация ДНК и транскрипция . Некоторые ферменты действуют на другие белки, чтобы добавить или удалять химические группы в процессе, известном как посттрансляционная модификация. Известно, что около 4000 реакций катализируются ферментами. [ 51 ] Ускорение скорости, предоставляемое ферментативным катализом, часто является огромным - как 10 17 -Краткое увеличение скорости по сравнению с неэтализированной реакцией в случае оротата -декарбоксилазы (78 миллионов лет без фермента, 18 миллисекунд с ферментом). [ 52 ]
Молекулы, связанные и действующие ферментами, называются субстратами . Хотя ферменты могут состоять из сотен аминокислот, обычно это лишь небольшая часть остатков, которые вступают в контакт с субстратом, и еще меньшая фракция - от три до четырех остатков - которые непосредственно участвуют в катализе. [ 53 ] Область фермента, который связывает субстрат и содержит каталитические остатки, известна как активный сайт .
Диригентные белки являются членами класса белков, которые диктуют стереохимию соединения, синтезированного другими ферментами. [ 54 ]
Сигнализация клеток и связывание лиганда
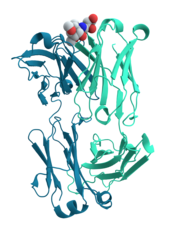
Многие белки участвуют в процессе передачи сигналов и передачи сигналов . Некоторые белки, такие как инсулин , являются внеклеточными белками, которые передают сигнал из клетки, в которой они были синтезированы с другими клетками в отдаленных тканях . Другие - это мембранные белки , которые действуют как рецепторы , основной функцией которого является связывание сигнальной молекулы и индуцировать биохимический ответ в клетке. Многие рецепторы имеют сайт связывания, подвергающийся воздействию на клеточную поверхность, и эффекторный домен внутри клетки, который может обладать ферментативной активностью или может подвергаться конформационному изменению, обнаруженному другими белками в клетке. [ 33 ] : 251–81
Антитела являются белковыми компонентами адаптивной иммунной системы, основной функцией которого является связывание антигенов или посторонних веществ в организме и нацеливаться на них для разрушения. Антитела могут быть секретированы во внеклеточную среду или закреплены на мембранах специализированных В -клеток, известных как плазматические клетки . Принимая во внимание, что ферменты ограничены в своем сродстве к связыванию к своим субстратам, необходимой провести их реакцию, антитела не имеют таких ограничений. Связывание антител к ее цели чрезвычайно высока. [ 34 ] : 275–50
Многие белки транспорта лигандов связывают определенные небольшие биомолекулы и транспортируют их в другие места в организме многоклеточного организма. Эти белки должны иметь высокую аффинность связывания, когда их лиганд присутствует в высоких концентрациях, но также должен высвобождать лиганд, когда он присутствует в низких концентрациях в тканях -мишенях. Каноническим примером лиганд-связывающего белка является гемоглобин , который переносит кислород из легких в другие органы и ткани у всех позвоночных и имеет близкие гомологи во всех биологических королевствах . [ 34 ] : 222–29 Лектины представляют собой белки, связывающие сахар , которые очень специфичны для их сахарных фрагментов. Лектины обычно играют роль в явлениях биологического распознавания с участием клеток и белков. [ 55 ] Рецепторы и гормоны являются очень специфическими связывающими белками.
Трансмембранные белки также могут служить белками транспорта лигандов, которые изменяют проницаемость клеточной мембраны на мелкие молекулы и ионы. Только мембрана имеет гидрофобное ядро, посредством которого полярные или заряженные молекулы не могут диффундировать . Мембранные белки содержат внутренние каналы, которые позволяют таким молекулам вводить и выходить из клетки. Многие белки ионных каналов специализируются на выборе только для определенного иона; Например, калий и натриевые каналы часто различают только один из двух ионов. [ 33 ] : 232–34
Структурные белки

Структурные белки придают жесткости и жесткости биологическим компонентам иначе. Большинство структурных белков являются фиброзными белками ; Например, коллаген и эластин являются критическими компонентами соединительной ткани , такой как хрящ , а кератин находится в твердых или нитевидных структурах, таких как волосы , ногти , перья , копыта и некоторые раковины животных . [ 34 ] : 178–81 Некоторые глобулярные белки также могут играть в структурные функции, например, актин и тубулин являются шаровидными и растворимыми в качестве мономеров, но полимеризуются с образованием длинных, жестких волокон, которые составляют цитоскелет , который позволяет клетке сохранять свою форму и размер.
Другими белками, которые выполняют структурные функции, являются моторные белки, такие как миозин , кинезин и динеин , которые способны генерировать механические силы. Эти белки имеют решающее значение для клеточной подвижности одноклеточных организмов и сперматозоидов многих многоклеточных организмов, которые размножаются сексуально . Они также генерируют силы, оказываемые сжимающими мышцами [ 34 ] : 258–64, 272 и играть важную роль в внутриклеточном транспорте.
Эволюция белка
Ключевым вопросом в молекулярной биологии является то, как развиваются белки, то есть как мутации (или, скорее, изменения в аминокислотной последовательности) привести к новым структурам и функциям? Большинство аминокислот в белке могут быть изменены без нарушения активности или функции, как видно из многочисленных гомологичных белков разных видов (как собрано в специализированных базах данных для семейств белков , например, PFAM ). [ 56 ] Чтобы предотвратить драматические последствия мутаций, ген может быть дублирован, прежде чем он сможет свободно мутировать. Однако это также может привести к полной потере функции генов и, следовательно, псевдогенам . [ 57 ] Чаще всего изменения аминокислот имеют ограниченные последствия, хотя некоторые могут существенно изменять функцию белка, особенно в ферментах . Например, многие ферменты могут изменить свою субстратную специфичность на одну или несколько мутаций. [ 58 ] Изменения в специфичности субстрата облегчаются распущенностью субстрата , т.е. способность многих ферментов связывать и обрабатывать несколько субстратов . Когда возникают мутации, специфичность фермента может увеличивать (или уменьшать) и, следовательно, его ферментативную активность. [ 58 ] Таким образом, бактерии (или другие организмы) могут адаптироваться к различным источникам пищи, включая неестественные субстраты, такие как пластик. [ 59 ]
Методы обучения
Методы, обычно используемые для изучения структуры и функции белка, включают иммуногистохимию , сайт-направленный мутагенез , рентгеновскую кристаллографию , ядерный магнитный резонанс и масс-спектрометрию .
Активность и структуры белков могут быть исследованы in vitro , in vivo и in silico . Исследования очищенных белков в контролируемых средах в контролируемых средах полезны для изучения того, как белок выполняет его функцию: например, исследования кинетики фермента исследуют химический механизм каталитической активности фермента и его относительная сродство к различным возможным молекулам субстрата. Напротив, эксперименты in vivo могут предоставить информацию о физиологической роли белка в контексте клетки или даже целого организма . В исследованиях Silico используются вычислительные методы для изучения белков.
Очистка белка
Белки могут быть очищены от других клеточных компонентов, используя различные методы, такие как ультрацентрифугирование , осаждение , электрофорез и хроматография ; Появление генетической инженерии стало возможным ряд методов для облегчения очистки.
Для проведения анализа in vitro белок должен быть очищен от других клеточных компонентов. Этот процесс обычно начинается с лизиса клеток , в котором нарушается мембрана клетки, а ее внутреннее содержимое выделяется в раствор, известный как грубое лизат . Полученная смесь может быть очищена с использованием ультрацентрифугирования , которая фракционирует различные клеточные компоненты в фракции, содержащие растворимые белки; мембранные липиды и белки; клеточные органеллы и нуклеиновые кислоты . Осадки методом, известным как солирование, может сконцентрировать белки из этого лизата. Затем используются различные типы хроматографии для выделения белка или белка, представляющих интерес на основе таких свойств, как молекулярный вес, чистый заряд и аффинность связывания. [ 30 ] : 21–24 Уровень очистки может контролироваться с использованием различных типов гелевого электрофореза, желаемого белка молекулярная масса и изоэлектрическую точку если известны с помощью спектроскопии , если белок обладает различимыми спектроскопическими признаками или с помощью ферментов, если белок обладает ферментатической активностью. Кроме того, белки могут быть выделены в соответствии с их зарядом с использованием электрофокусировки . [ 60 ]
Для природных белков может потребоваться серия стадий очистки для получения белка, достаточно чистого для лабораторных применений. Чтобы упростить этот процесс, генетическая инженерия часто используется для добавления химических особенностей к белкам, которые облегчают их очистку, не влияя на их структуру или активность. Здесь «метка», состоящая из специфической аминокислотной последовательности, часто серии остатков гистидина (« His-tag »), прикреплен к одному конце белка. В результате, когда лизат передается по колонке хроматографии, содержащей никель , остатки гистидина лигают никель и прикрепляются к колонке, в то время как негрированные компоненты лизата проходят беспрепятственно. Был разработан ряд различных тегов, чтобы помочь исследователям очистить определенные белки из сложных смесей. [ 61 ]
Клеточная локализация
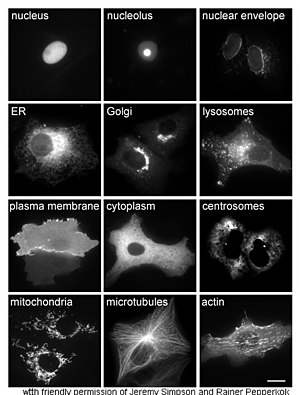
Изучение белков in vivo часто связано с синтезом и локализацией белка в клетке. Хотя многие внутриклеточные белки синтезируются в цитоплазме и мембрановой или секретируемых белках в эндоплазматическом ретикулуме , специфика того, как белки нацелены на специфические органеллы или клеточные структуры, часто неясны. Полезный метод оценки клеточной локализации использует генетическую инженерию для экспрессии в клетке слитого белка или химеры , состоящего из природного белка, представляющего интерес, связанный с « репортером », таким как зеленый флуоресцентный белок (GFP). [ 62 ] Положение слитого белка в клетке может быть затем чисто и эффективно визуализировано с помощью микроскопии , [ 63 ] Как показано на рисунке напротив.
Другие методы выяснения клеточного расположения белков требуют использования известных компартментальных маркеров для таких областей, как ER, гольги, лизосомы или вакуоли, митохондрии, хлоропласты, плазматические мембраны и т. Д. Антител к известным маркерам становится намного проще идентифицировать локализацию интересующего белка. Например, косвенная иммунофлуоресценция обеспечит флуоресцентную колокализацию и демонстрацию местоположения. Флуоресцентные красители используются для маркировки клеточных компартментов для аналогичной цели. [ 64 ]
Существуют и другие возможности. Например, иммуногистохимия обычно использует антитело к одному или нескольким белкам, представляющим интерес, которые конъюгированы с ферментами, дающими либо люминесцентные, либо хромогенные сигналы, которые можно сравнить между образцами, что позволяет получить информацию о локализации. Другим применимым методикой является COFRactionation в сахарозе (или других материалах) градиентах с использованием изопикнического центрифугирования . [ 65 ] Хотя этот метод не доказывает колокализацию компартмента известной плотности и интересующего белка, он увеличивает вероятность и более поддается масштабным исследованиям.
Наконец, золотым стандартом метода клеточной локализации является иммуноэлектронная микроскопия . Этот метод также использует антитело к интересующему белку, а также классическую электронную микроскопию. Образец готовит для нормального электронного микроскопического исследования, а затем обрабатывает антителом к интересующему белу, который конъюгируется с чрезвычайно электрическим материалом, обычно золотым. Это допускает локализацию как ультраструктурных деталей, так и интересующего белка. [ 66 ]
Благодаря другому генетическому инженерному приложению, известному как сайт-направленный мутагенез , исследователи могут изменить белковую последовательность и, следовательно, ее структуру, клеточную локализацию и восприимчивость к регуляции. Этот метод даже позволяет включать неестественные аминокислоты в белки, используя модифицированные тРНК, [ 67 ] и может позволить рациональный дизайн новых белков с новыми свойствами. [ 68 ]
Протеомика
Общий комплемент белков, присутствующих за раз в клеточном или клеточном типе, известен как его протеом , и изучение таких крупномасштабных наборов данных определяет область протеомики , названную аналогией связанной области геномики . Ключевые экспериментальные методы в протеомике включают 2D -электрофорез , [ 69 ] что позволяет разделять многие белки, масс -спектрометрию , [ 70 ] что позволяет быстро идентифицировать высокопроизводительность белков и секвенирование пептидов (чаще всего после расщепления в геле ), белковых микрочипов , которые позволяют обнаружить относительные уровни различных белков, присутствующих в клетке, и двухгибридный скрининг , который, который, который, которые, которые, которые, которые, которые, которые могут Позволяет систематическому изучению белковых взаимодействий . [ 71 ] Общий дополнение биологически возможных таких взаимодействий известно как взаимодействие . [ 72 ] Систематическая попытка определить структуры белков, представляющих каждую возможную складку, известна как структурная геномика . [ 73 ]
Определение структуры
Обнаружение третичной структуры белка или четвертичной структуры его комплексов может дать важные подсказки о том, как белок выполняет свою функцию и как его можно влиять, т.е. в дизайне лекарств . Поскольку белки слишком малы, чтобы их можно было увидеть под световым микроскопом , другие методы должны быть использованы для определения их структуры. Общие экспериментальные методы включают рентгеновскую кристаллографию и ЯМР-спектроскопию , которые могут производить структурную информацию при атомном разрешении. Однако эксперименты ЯМР способны предоставить информацию, из которой можно оценить подмножество расстояний между парами атомов, и окончательные возможные конформации для белка определяются путем решения проблемы геометрии расстояния . Двойная поляризационная интерферометрия является количественным аналитическим методом для измерения общей конформации белка и конформационных изменений из -за взаимодействия или другого стимула. Круговой дихроизм является еще одним лабораторным методом для определения внутреннего β-листа / α-спирального состава белков. Криоэлектронная микроскопия используется для получения структурной информации с более низким разрешением об очень больших белковых комплексах, включая собранные вирусы ; [ 33 ] : 340–41 В некоторых случаях вариант, известный как кристаллография электронов , также может дать информацию с высоким разрешением, особенно для двухмерных кристаллов мембранных белков. [ 74 ] Решенные структуры обычно депонируются в банке данных белка (PDB), свободно доступном ресурсе, из которого можно получить структурные данные о тысячах белков в форме декартовых координат для каждого атома в белке. [ 75 ]
Гораздо больше генных последовательностей известно, чем белковые структуры. Кроме того, набор решаемых структур смещен в сторону белков, которые могут быть легко подвергнуты условиям, необходимым для рентгеновской кристаллографии , одного из основных методов определения структуры. В частности, глобулярные белки сравнительно легко кристаллизовать при подготовке к рентгеновской кристаллографии. Мембранные белки и крупные белковые комплексы, напротив, трудно кристаллизовать и недопредставлены в PDB. [ 76 ] Структурные инициативы геномики пытались исправить эти недостатки путем систематического решения репрезентативных структур основных классов сгиба. Методы прогнозирования структуры белка пытаются обеспечить средства генерации правдоподобной структуры для белков, структуры которых не были экспериментально определены. [ 77 ]
Прогноз структуры

Дополнительная к области структурной геномики, прогнозирование структуры белка развивает эффективные математические модели белков для вычисления, предсказания молекулярных образов в теории, вместо того, чтобы обнаружить структуры с лабораторным наблюдением. [ 78 ] Наиболее успешный тип прогнозирования структуры, известный как моделирование гомологии , опирается на существование структуры «шаблона» с сходством последовательностей с моделируемым белком; Цель структурной геномики состоит в том, чтобы обеспечить достаточное представление в решительных структурах для моделирования большинства из тех, кто остается. [ 79 ] Хотя создание точных моделей остается проблемой, когда доступны только отдаленно связанные структуры шаблонов, было высказано предположение, что выравнивание последовательностей является узким местом в этом процессе, так как можно создать довольно точные модели, если известно «идеальная» выравнивание последовательности. [ 80 ] Многие методы прогнозирования структуры послужили для информирования о появлении области инженерии белков , в которой уже были разработаны новые белковые складки. [ 81 ] Также белки (у эукариот ~ 33%) содержат большие неструктурированные, но биологически функциональные сегменты и могут быть классифицированы как внутренние белки . [ 82 ] Следовательно, прогнозирование и анализ белкового расстройства является важной частью характеристики структуры белка. [ 83 ]
Биоинформатика
Был разработан огромный спектр вычислительных методов для анализа структуры, функции и эволюции белков. Разработка таких инструментов была обусловлена большим и быстрорастущим количеством геномных и протеомных данных, доступных для различных организмов, включая геном человека . Ресурсы не существуют для изучения всех белков экспериментально, поэтому лишь немногие подвергаются лабораторным экспериментам, в то время как вычислительные инструменты используются для экстраполяции в подобные белки. Такие гомологичные белки могут быть эффективно идентифицированы в отдаленно связанных организмах путем выравнивания последовательностей . Последовательности генома и генов можно искать по множеству инструментов для определенных свойств. Инструменты профилирования последовательностей могут найти рестрикционные ферментные сайты, открытые кадры считывания в нуклеотидных последовательностях и прогнозировать вторичные структуры . Филогенетические деревья могут быть сконструированы, а эволюционные гипотезы, разработанные с использованием специального программного обеспечения, такого как Clustal W, в отношении происхождения современных организмов и генов, которые они выражают. Поле Биоинформатика в настоящее время необходима для анализа генов и белков.
При силико моделировании динамических процессов
Более сложной вычислительной проблемой является прогноз межмолекулярных взаимодействий, таких как молекулярная стыковка , [ 84 ] Складывание белка , взаимодействие белка -белка и химическая реакционная способность. Математические модели для моделирования этих динамических процессов включают молекулярную механику , в частности, молекулярную динамику . В связи с этим при силико моделировании обнаружили складывание небольших α-спиральных белковых доменов, таких как головной убор виллина , [ 85 ] ВИЧ -аксессуарный белок [ 86 ] и гибридные методы, объединяющие стандартную молекулярную динамику с квантовой механической математикой, исследовали электронные состояния родопсинов . [ 87 ]
Помимо классической молекулярной динамики, методы квантовой динамики позволяют моделировать белки в атомистических деталях с точным описанием квантовых механических эффектов. Примеры включают многослойный метод мультиконфигурации, зависящий от времени Hartree (MCTDH), и иерархические уравнения движения (HEOM), которые были применены к криптохромам растений [ 88 ] и бактерий легких комплексов, [ 89 ] соответственно. Как квантовое, так и классическое механическое моделирование систем биологического масштаба чрезвычайно требовательно вычислительно, поэтому распределенные вычислительные инициативы (например, Folding@Home проект [ 90 ] ) Облегчить молекулярное моделирование путем использования достижений в графических процессоров параллельной обработке Монте -Карло и методов .
Химический анализ
Общее содержание азота в органическом веществе в основном образуется аминогрупп в белках. Общий азот Kjeldahl ( TKN ) является мерой азота, широко используемого при анализе (отходов) воды, почвы, пищи, кормления и органического вещества в целом. Как следует из названия, метод Kjeldahl применяется . Доступны более чувствительные методы. [ 91 ] [ 92 ]
Питание
Большинство микроорганизмов и растений могут биосинтезировать все 20 стандартных аминокислот , в то время как животные (включая людей) должны получить некоторые из аминокислот из рациона . [ 47 ] Аминокислоты, которые организм не может синтезировать самостоятельно, называются незаменимыми аминокислотами . Ключевые ферменты, которые синтезируют определенные аминокислоты, не присутствуют у животных, таких как аспартакиназа , которая катализирует первый этап в синтезе лизина , метионина и треонина из аспартата . Если в окружающей среде присутствуют аминокислоты, микроорганизмы могут сохранять энергию, взяв аминокислоты из их окружения и подавляя их биосинтетические пути.
У животных аминокислоты получают за счет потребления продуктов, содержащих белок. Затем проглатываемые белки разбиваются на аминокислоты посредством расщепления , которое обычно включает в себя денатурацию белка посредством воздействия кислоты и гидролиза ферментами, называемыми протеазами . Некоторые проглатываемые аминокислоты используются для биосинтеза белка, в то время как другие превращаются в глюкозу посредством глюконеогенеза или подаются в цикл лимонной кислоты . Такое использование белка в качестве топлива особенно важно в условиях голода , поскольку он позволяет использовать собственные белки организма для поддержки жизни, особенно в мышцах . [ 93 ]
У животных, таких как собаки и кошки , белок поддерживает здоровье и качество кожи, стимулируя рост волосяных фолликулов и кератинизацию, и, таким образом, снижая вероятность возникновения проблем с кожей. [ 94 ] Некачественные белки также играют роль в отношении здоровья желудочно-кишечного тракта, увеличивая потенциал метеоризма и пахотных соединений у собак, потому что, когда белки достигают толстой кишки в непереваренном состоянии, они ферментируются, продуцируя газовый газ, индол и скатол. [ 95 ] Собаки и кошки переваривают животные белки лучше, чем из растений, но продукты низкокачественного животного происхождения плохо перевариваются, включая кожу, перья и соединительную ткань. [ 95 ]
Механические свойства
Механические свойства белков очень разнообразны и часто являются центральными для их биологической функции, как в случае белков, таких как кератин и коллаген . [ 96 ] Например, способность мышечной ткани постоянно расширяться и сокращаться напрямую связана с упругими свойствами их основного белкового макияжа. [ 97 ] [ 98 ] Помимо фиброзных белков, конформационная динамика ферментов [ 99 ] и структура биологических мембран , среди других биологических функций, определяется механическими свойствами белков. Помимо их биологического контекста, уникальные механические свойства многих белков, наряду с их относительной устойчивостью по сравнению с синтетическими полимерами , сделали их желательными мишенями для дизайна материалов следующего поколения. [ 100 ] [ 101 ]
Модуль Янга
Модуль Янга , E, рассчитывается как осевое напряжение σ над результирующей деформацией ε. Это мера относительной жесткости материала. В контексте белков эта жесткость часто напрямую коррелирует с биологической функцией. Например, коллаген , обнаруженный в соединительной ткани , костях и хряще , и кератин , обнаруженный в ногтях, когтях и волосах, наблюдали жесткости, которые на несколько порядков выше, чем у эластина , [ 102 ] Что состоит в том, чтобы придать эластичность таким структурам, как кровеносные сосуды , легочная ткань и ткань мочевого пузыря , среди прочих. [ 103 ] [ 104 ] По сравнению с этим глобулярные белки , такие как бычий сывороточный альбумин , который относительно свободно плавает в цитозоле и часто функционирует как ферменты (и, следовательно, переносится частыми конформационными изменениями), имеют сравнительно более низкие модули Янга. [ 105 ] [ 106 ]
Модуль Янга одного белка можно найти с помощью молекулярной динамики. Использование либо атомистических силовых полей, таких как Charmm или Gromos , или крупнозернистые силовые поля, такие как Martini, [ 107 ] Одна молекула белка может быть растянута одноосной силой, в то время как результирующее разгибание регистрируется для расчета штамма. [ 108 ] [ 109 ] Экспериментально, такие методы, как микроскопия атомной силы, могут использоваться для получения аналогичных данных. [ 110 ]
На макроскопическом уровне модуль молодых сетей белковых сетей может быть получен с помощью более традиционных механических испытаний . Экспериментально наблюдаемые значения для нескольких белков можно увидеть ниже.
| Белок | Класс белка | Модуль Янга |
|---|---|---|
| Кератин (сшитый) | Волокнистый | 1,5-10 ГПа [ 111 ] |
| Эластин (сшитый) | Волокнистый | 1 МПа [ 102 ] |
| Фибрин (сшитый) | Волокнистый | 1-10 МПа [ 102 ] |
| Коллаген (сшитый) | Волокнистый | 5-7,5 ГПа [ 102 ] [ 112 ] |
| Resilin (сшитый) | Волокнистый | 1-2 МПа [ 102 ] |
| Бычий сывороточный альбумин (сшитый) | Глобулярный | 2,5-15 кПа [ 105 ] |
| β-боловые белки из внешней мембраны | Мембрана | 20-45 ГПа [ 113 ] |
Вязкость
В дополнение к служащим в качестве ферментов в клетке, глобулярные белки часто действуют как ключевые транспортные молекулы. Например, сывороточные альбумины , ключевой компонент крови , необходимы для транспортировки множества малых молекул по всему телу. [ 114 ] Из -за этого, зависимое от концентрации поведение этих белков в растворе, непосредственно связано с функцией системы кровообращения . В качестве количественной оценки этого поведения происходит вязкость решения.
Вязкость, η, обычно приводится в меру сопротивления жидкости деформации. Его можно рассчитать как отношение между приложенным напряжением и скоростью изменения результирующего деформации сдвига, то есть скорости деформации. Вязкость сложных жидких смесей, таких как кровь, часто сильно зависит от температуры и концентрации растворенного вещества. [ 115 ] Для сывороточного альбумина, в частности бычьего сывороточного альбумина следующую связь между вязкостью и температурой и концентрацией . , можно использовать [ 116 ]
![{\ displaystyle \ eta = \ exp \ left [{\ frac {c} {\ alpha -\ beta \ c}} \ left (-b+dt+{\ frac {\ delta e} {rt}} \ right) \ верно]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/c875efcb377556dd7908fc43705d64ab6b749489)
Где C -концентрация, t -температура, R - постоянная газа , а α, β, B , D и Δ E -все константы свойства на основе материала. Это уравнение имеет форму уравнения Аррениуса , назначая вязкость, экспоненциальную зависимость от температуры и концентрации.
Смотрите также
Ссылки
- ^ Подпрыгнуть до: а беременный в Осборн Т.Б. (1909). «История» . Овощные белки . С. 1–6.
- ^ Подпрыгнуть до: а беременный в Рейнольдс Дж.А., Танфорд С. (2003). Роботы природы: история белков (Оксфордская книга в мягкой обложке) . Нью -Йорк, Нью -Йорк: издательство Оксфордского университета. п. 15. ISBN 978-0-19-860694-9 .
- ^ Танфорд С. (2001). Роботы природы: история белков . Интернет -архив. Оксфорд; Торонто: издательство Оксфордского университета. ISBN 978-0-19-850466-5 .
- ^ Малдер Г.Дж. (1838). «О композиции некоторых животных веществ» . Бюллетень физических и естественных наук на голландском языке : 104.
- ^ Хартли Х (август 1951 г.). «Происхождение слова« белок » . Природа . 168 (4267): 244. Bibcode : 1951natur.168..244H . doi : 10.1038/168244a0 . PMID 14875059 . S2CID 4271525 .
- ^ Подпрыгнуть до: а беременный в Perrett D (август 2007 г.). «От« белка »до начала клинической протеомики». Протеомика. Клиническое применение . 1 (8): 720–738. doi : 10.1002/prca.200700525 . PMID 21136729 . S2CID 32843102 .
- ^ «Белок (п.)». Оксфордский английский словарь . Июль 2023 года. DOI : 10.1093/OED/5657543824 .
- ^ Bischoff TL, VOIT C (1860). Законы травяной церемонии травоворитера, обнаруженные в новых исследованиях (на немецком языке). Лейпциг, Гейдельберг.
{{cite book}}: CS1 Maint: местоположение отсутствует издатель ( ссылка ) - ^ Осборн Т.Б. (апрель 1913). «В память о Генрихе Риттхаузен» . Биохимический бюллетень . II (7). Колумбийского университета Биохимическая ассоциация : 338 . Получено 1 января 2016 года . , заархивировано в библиотеке наследия биоразнообразия
- ^ "Хофмайстер, Франц" . Encyclopedia.com. Архивировано из оригинала 5 апреля 2017 года . Получено 4 апреля 2017 года .
- ^ Koshland DE, Haurowitz F. «Белок, раздел: классификация белка» . Britannica.com. Архивировано с оригинала 4 апреля 2017 года . Получено 4 апреля 2017 года .
- ^ Самнер Дж.Б. (август 1926). «Изоляция и кристаллизация ферментной уреазы» . Журнал биологической химии . 69 (2): 435–441. doi : 10.1016/s0021-9258 (18) 84560-4 .
- ^ Полинг Л, Кори Р.Б. (май 1951 г.). «Атомные координаты и структурные факторы для двух спиральных конфигураций полипептидных цепей» . Труды Национальной академии наук Соединенных Штатов Америки . 37 (5): 235–240. Bibcode : 1951pnas ... 37..235p . doi : 10.1073/pnas.37.5.235 . PMC 1063348 . PMID 14834145 .
- ^ Kauzmann W (май 1956 г.). «Структурные факторы в денатурации белка». Журнал клеточной физиологии. Добавка . 47 (Suppl 1): 113–131. doi : 10.1002/jcp.1030470410 . PMID 13332017 .
- ^ Kauzmann W (1959). «Некоторые факторы в интерпретации белковой денатурации». Достижения в химии белка, том 14 . Тол. 14. С. 1–63. doi : 10.1016/s0065-3233 (08) 60608-7 . ISBN 978-0-12-034214-3 Полем PMID 14404936 .
- ^ Калман С.М., Линдерстром-Ланг К., Оттесен М., Ричардс Ф.М. (февраль 1955 г.). «Разложение рибонуклеазы субтилизином». Biochimica et Biophysica Acta . 16 (2): 297–299. doi : 10.1016/0006-3002 (55) 90224-9 . PMID 14363272 .
- ^ Сэнгер F (1949). «Терминальные пептиды инсулина» . Биохимический журнал . 45 (5): 563–574. doi : 10.1042/bj0450563 . PMC 1275055 . PMID 15396627 .
- ^ Сэнгер Ф. (1958), Нобелевская лекция: Химия инсулина (PDF) , Nobelprize.org, архивировал (PDF) из оригинала 2013-03-19 , извлечен 2016-02-09
- ^ Ричардс Ф.М. (1972). «Нобелевская премия 1972 года за химию». Наука . 178 (4060): 492–3. Bibcode : 1972sci ... 178..492r . doi : 10.1126/science.178.4060.492 . PMID 17754377 .
- ^ Marshall GR, Feng JA, Kuster DJ (2008). «Вернуться в будущее: рибонуклеаза А» . Биополимеры . 90 (3): 259–77. doi : 10.1002/bip.20845 . PMID 17868092 .
- ^ Подпрыгнуть до: а беременный в Stoddart C (1 марта 2022 г.). «Структурная биология: как белки получили свой крупный план» . Познаваемый журнал . doi : 10.1146/Познание-022822-1 .
- ^ Muirhead H, Perutz MF (август 1963 г.). «Структура гемоглобина. Трехмерный синтез Фурье сниженного человеческого гемоглобина при разрешении 5,5 Å». Природа . 199 (4894): 633–638. Bibcode : 1963natur.199..633m . doi : 10.1038/1996333a0 . PMID 14074546 . S2CID 4257461 .
- ^ Kendrew JC, Bodo G, Dintzis HM, Parrish RG, Wyckoff H, Phillips DC (март 1958 г.). «Трехмерная модель молекулы миоглобина, полученная с помощью рентгеновского анализа». Природа . 181 (4610): 662–666. Bibcode : 1958natur.181..662K . doi : 10.1038/181662a0 . PMID 13517261 . S2CID 4162786 .
- ^ Чжоу Чж (апрель 2008 г.). «На пути к структурному определению атомного разрешения с помощью одночастической криоэлектронной микроскопии» . Современное мнение в структурной биологии . 18 (2): 218–228. doi : 10.1016/j.sbi.2008.03.004 . PMC 2714865 . PMID 18403197 .
- ^ Кескин О., Тункбаг Н., Гурсой А (апрель 2008 г.). «Характеристика и прогнозирование белковых интерфейсов, чтобы вывести сети взаимодействия белка-белка». Текущая фармацевтическая биотехнология . 9 (2): 67–76. doi : 10.2174/138920108783955191 . HDL : 11511/32640 . PMID 18393863 .
- ^ «Сводная статистика» . RCSB PDB . Получено 2024-04-20 .
- ^ Ekman D, Björklund AK, Frey-Skött J, Elofsson A (апрель 2005 г.). «Многодоменные белки в трех королевствах жизни: сирот домены и другие незнашиваемые регионы». Журнал молекулярной биологии . 348 (1): 231–243. doi : 10.1016/j.jmb.2005.02.007 . PMID 15808866 .
- ^ Нельсон Д.Л., Кокс М.М. (2005). Принципы биохимии Лехнингера (4 -е изд.). Нью -Йорк, Нью -Йорк: WH Freeman and Company.
- ^ Гайтитер А., Торнтон Дж. М. (ноябрь 2005 г.). «Понимание каталитического инструментария природы». Тенденции в биохимических науках . 30 (11): 622–629. doi : 10.1016/j.tibs.2005.09.006 . PMID 16214343 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин Murray RF, Harper HW, Granner DK, Mayes PA, Rodwell VW (2006). Иллюстрированная биохимия Харпера . Нью-Йорк: Lange Medical Books/McGraw-Hill. ISBN 978-0-07-146197-9 .
- ^ Подпрыгнуть до: а беременный в Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, et al. (2004). Молекулярная клеточная биология (5 -е изд.). Нью -Йорк, Нью -Йорк: WH Freeman and Company.
- ^ Ardejani MS, Powers ET, Kelly JW (август 2017 г.). «Использование кооперативно сложенных пептидов для измерения энергий взаимодействия и конформационных склонностей» . Счета химических исследований . 50 (8): 1875–1882. doi : 10.1021/acs.accounts.7b00195 . PMC 5584629 . PMID 28723063 .
- ^ Подпрыгнуть до: а беременный в дюймовый Бранден С., Туз Дж. (1999). Введение в структуру белка . Нью -Йорк: Garland Pub. ISBN 978-0-8153-2305-1 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж Ван Холде К.Е., Мэтьюз К.К. (1996). Биохимия . Менло Парк, Калифорния: Бенджамин/Каммингс Паб. Co., Inc. ISBN 978-0-8053-3931-4 .
- ^ Мило Р (декабрь 2013 г.). «Какое общее количество белковых молекул на объем клеток? Призыв переосмыслить некоторые опубликованные значения» . Биологии . 35 (12): 1050–1055. doi : 10.1002/bies.201300066 . PMC 3910158 . PMID 24114984 .
- ^ Beck M, Schmidt A, Malmstroem J, Claassen M, Ori A, Szymborska A, et al. (Ноябрь 2011). «Количественный протеом человеческой клеточной линии» . Биология молекулярных систем . 7 : 549. doi : 10.1038/msb.2011.82 . PMC 3261713 . PMID 22068332 .
- ^ Ву Л., Кандилль С.И., Чой Ю., Се Д., Цзян Л., Ли-Пок-Тан Дж. И др. (Июль 2013). «Вариация и генетический контроль изобилия белка у людей» . Природа . 499 (7456): 79–82. Bibcode : 2013natur.499 ... 79w . doi : 10.1038/nature12223 . PMC 3789121 . PMID 23676674 .
- ^ Добсон К.М. (2000). «Природа и значение складывания белка». В боли RH (ред.). Механизмы складывания белка . Оксфорд, Оксфордшир: издательство Оксфордского университета. С. 1–28. ISBN 978-0-19-963789-8 .
- ^ Kozlowski LP (январь 2017 г.). «Протеом-PI: Протеоемная изоэлектрическая база данных точек» . Исследование нуклеиновых кислот . 45 (D1): D1112 - D1116. doi : 10.1093/nar/gkw978 . PMC 5210655 . PMID 27789699 .
- ^ Fulton AB, Isaacs WB (апрель 1991 г.). «Титин, огромный, эластичный саркомерный белок с вероятной ролью в морфогенезе». Биологии . 13 (4): 157–161. doi : 10.1002/bies.950130403 . PMID 1859393 . S2CID 20237314 .
- ^ Bruckdorfer T, Marder O, Albericio F (февраль 2004 г.). «От производства пептидов в количестве миллиграммов для исследований до множества величин для лекарств в будущем». Текущая фармацевтическая биотехнология . 5 (1): 29–43. doi : 10.2174/1389201043489620 . PMID 14965208 .
- ^ Schwarzer D, Cole PA (декабрь 2005 г.). «Полусинтез белка и экспрессированный перевязку белка: погоня за хвостом белка». Современное мнение о химической биологии . 9 (6): 561–569. doi : 10.1016/j.cbpa.2005.09.018 . PMID 16226484 .
- ^ Кент С.Б. (февраль 2009 г.). «Общий химический синтез белков». Обзоры химического общества . 38 (2): 338–351. doi : 10.1039/b700141j . PMID 19169452 . S2CID 5432012 .
- ^ Фернандес А., Скотт Р (сентябрь 2003 г.). «Дегидрон: структурно кодируемый сигнал для взаимодействия белка» . Биофизический журнал . 85 (3): 1914–1928. Bibcode : 2003bpj .... 85.1914f . doi : 10.1016/s0006-3495 (03) 74619-0 . PMC 1303363 . PMID 12944304 .
- ^ Дэйви Н.Е., Ван Рои К., Уэтеритт Р.Дж., Тодт Г., Уяр Б., Альтенберг Б. и др. (Январь 2012 г.). «Атрибуты коротких линейных мотивов». Молекулярные биосистемы . 8 (1): 268-281. Doi : 10.1039 / c1mb05231d . PMID 21909575 .
- ^ Скальвини Б., Шейххассани В., Вудард Дж., Аупич Дж, Дам Р.Т., Джерала Р. и др. (Июль 2020 г.). «Топология сложенных молекулярных цепей: от отдельных биомолекул до инженерных оригами». Тенденции в химии . 2 (7): 609–622. doi : 10.1016/j.trechm.2020.04.009 . HDL : 1887/3245505 . S2CID 218957613 .
- ^ Подпрыгнуть до: а беременный Foot D, Foot Jg. (2004). Биохимия полная 1 3 -е изд.
- ^ Sannararayanan R, Moras D (2001). "Точность перевода генетического кода " Acta Biochimica Polonica 48 (2): 323–3 Doi : 10.18388/ abp.2001_3 11732604PMID
- ^ Копленд Дж. А., Шеффилд-Мур М., Колдич-Зиванович Н., Джентри С., Лампро Г., Цорцату-Статопулу Ф. и др. (Июнь 2009 г.). «Половые стероидные рецепторы при дифференцировке скелета и эпителиальной неоплазии: возможно ли тканевое вмешательство?». Биологии . 31 (6): 629–641. doi : 10.1002/bies.200800138 . PMID 19382224 . S2CID 205469320 .
- ^ Самарин С., Нусрат А (январь 2009 г.). «Регуляция эпителиального апикального соединительного комплекса с помощью GTPase Family Family» . Границы в биологии . 14 (3): 1129–1142. doi : 10.2741/3298 . PMID 19273120 .
- ^ Байрох А (январь 2000 г.). «База данных ферментов в 2000 году» . Исследование нуклеиновых кислот . 28 (1): 304–305. doi : 10.1093/nar/28.1.304 . PMC 102465 . PMID 10592255 .
- ^ Радзицка А., Вольфенден Р. (январь 1995 г.). «Опытный фермент». Наука . 267 (5194): 90–93. Bibcode : 1995sci ... 267 ... 90r . doi : 10.1126/science.7809611 . PMID 7809611 .
- ^ EBI Внешние службы (2010-01-20). «Каталитические атлас в Европейском институте биоинформатики» . Ebi.ac.uk. Архивировано из оригинала 2013-08-03 . Получено 2011-01-16 .
- ^ Пикель Б., Шаллер А (октябрь 2013 г.). «Диригентные белки: молекулярные характеристики и потенциальные биотехнологические применения». Прикладная микробиология и биотехнология . 97 (19): 8427–8438. doi : 10.1007/s00253-013-5167-4 . PMID 23989917 . S2CID 1896003 .
- ^ Rüdiger H, Siebert HC, Solís D, Jiménez-Barbero J, Romero A, Von Der Lieth CW, et al. (Апрель 2000). «Лекарственная химия, основанная на сахарном коде: основы лектинологии и экспериментальных стратегий с лектинами в качестве целей». Текущая лекарственная химия . 7 (4): 389–416. doi : 10.2174/0929867003375164 . PMID 10702616 .
- ^ Малдер Н.Дж. (2007-09-28). «Базы данных семейства белков». эл . Чичестер, Великобритания: John Wiley & Sons, Ltd. с. A0003058.Pub2. doi : 10.1002/9780470015902.a0003058.pub2 . ISBN 978-0-470-01617-6 .
- ^ Sisu C, Pei B, Leng J, Frankish A, Zhang Y, Balasubramanian S, et al. (Сентябрь 2014). «Сравнительный анализ псевдогенов в трех филах» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (37): 13361–13366. Bibcode : 2014pnas..11113361S . doi : 10.1073/pnas.1407293111 . PMC 4169933 . PMID 25157146 .
- ^ Подпрыгнуть до: а беременный Guzmán GI, Sandberg TE, Lacroix RA, Nyerges A, Papp H, De Raad M, et al. (Апрель 2019). «Адаптация распределения ферментов формирует к новым субстратам роста» . Биология молекулярных систем . 15 (4): E8462. doi : 10.15252/msb.20188462 . PMC 6452873 . PMID 30962359 .
- ^ Бано К., Куддус М., Захир М.Р., Зия К., Хан М.Ф., Ашраф Г.М. и др. (2017). «Микробная ферментативная деградация биоразлагаемых пластиков». Текущая фармацевтическая биотехнология . 18 (5): 429–440. doi : 10.2174/1389201018666170523165742 . PMID 28545359 .
- ^ Привет, J, Posch A, Cohen A, Liu N, Harbers A (2008). «Фракционирование сложных белковых смесей с помощью изоэлектрической фокусировки жидкофазной фазы». 2D -страница: образец подготовка и фракционирование . Методы в молекулярной биологии. Тол. 424. С. 225–239. doi : 10.1007/978-1-60327-064-9_19 . ISBN 978-1-58829-722-8 Полем PMID 18369866 .
- ^ Терпе К (январь 2003 г.). «Обзор слияния белка TAG: от молекулярных и биохимических основ в коммерческие системы». Прикладная микробиология и биотехнология . 60 (5): 523–533. doi : 10.1007/s00253-002-1158-6 . PMID 12536251 . S2CID 206934268 .
- ^ Stepanenko OV, Verkhusha VV, Kuznetsova IM, Uversky VN, Turoverov KK (август 2008 г.). «Флуоресцентные белки как биомаркеры и биосенсоры: бросание цветовых огней на молекулярные и клеточные процессы» . Текущая белковая и пептидная наука . 9 (4): 338–369. doi : 10.2174/138920308785132668 . PMC 2904242 . PMID 18691124 .
- ^ Yuste R (декабрь 2005 г.). «Флуоресцентная микроскопия сегодня». Природные методы . 2 (12): 902–904. doi : 10.1038/nmeth1205-902 . PMID 16299474 . S2CID 205418407 .
- ^ Марголин W (январь 2000 г.). «Зеленый флуоресцентный белок в качестве репортера для макромолекулярной локализации в бактериальных клетках». Методы 20 (1): 62–72. doi : 10.1006/Meth.1999.0906 . PMID 10610805 .
- ^ Уокер Дж. Х., Уилсон К. (2000). Принципы и методы практической биохимии . Кембридж, Великобритания: издательство Кембриджского университета. С. 287–89. ISBN 978-0-521-65873-7 .
- ^ Mayhew TM, Lucocq JM (август 2008 г.). «Разработка в клеточной биологии для количественной иммуноэлектронной микроскопии на основе тонких срезов: обзор» . Гистохимия и клеточная биология . 130 (2): 299–313. doi : 10.1007/s00418-008-0451-6 . PMC 2491712 . PMID 18553098 .
- ^ Hohsaka T, Sisido M (декабрь 2002 г.). «Включение не натуральных аминокислот в белки». Современное мнение о химической биологии . 6 (6): 809–815. doi : 10.1016/s1367-5931 (02) 00376-9 . PMID 12470735 .
- ^ Cedrone F, Ménez A, Quéméneur E (август 2000 г.). «Адаптирование новых ферментных функций с помощью рационального редизайна». Современное мнение в структурной биологии . 10 (4): 405–410. doi : 10.1016/s0959-440x (00) 00106-8 . PMID 10981626 .
- ^ Görg A, Weiss W, Dunn MJ (декабрь 2004 г.). «Современная двумерная технология электрофореза для протеомики». Протеомика . 4 (12): 3665–3685. doi : 10.1002/pmic.200401031 . PMID 15543535 . S2CID 28594824 .
- ^ Conrotto P, Souchelnytskyi S (сентябрь 2008 г.). «Протеомные подходы в биологических и медицинских науках: принципы и применение». Экспериментальная онкология . 30 (3): 171–180. PMID 18806738 .
- ^ Koegl M, Uetz P (декабрь 2007 г.). «Улучшение дрожжевых двухгибридных систем скрининга». Брифинги в функциональной геномике и протеомике . 6 (4): 302–312. doi : 10.1093/bfgp/elm035 . PMID 18218650 .
- ^ Plewczyński D, Ginalski K (2009). «Interactome: прогнозирование взаимодействия белкового белка в клетках» . Клеточная и молекулярная биологическая буква . 14 (1): 1–22. doi : 10.2478/s11658-008-0024-7 . PMC 6275871 . PMID 18839074 .
- ^ Чжан С., Ким Ш. (Февраль 2003 г.). «Обзор структурной геномики: от структуры к функции» . Современное мнение о химической биологии . 7 (1): 28–32. doi : 10.1016/s1367-5931 (02) 00015-7 . PMID 12547423 .
- ^ Gonen T, Cheng Y, Sliz P, Hiroaki Y, Fujiyoshi Y, Harrison SC, et al. (Декабрь 2005 г.). «Взаимодействие липид-белков в двухслойных двухмерных кристаллах AQP0» . Природа . 438 (7068): 633–638. Bibcode : 2005natur.438..633G . doi : 10.1038/nature04321 . PMC 1350984 . PMID 16319884 .
- ^ Стэндли Д.М., Кинджо А.Р., Киношита К., Накамура Х (июль 2008 г.). «Базы данных структуры белка с новыми веб -сервисами для структурной биологии и биомедицинских исследований» . Брифинги в биоинформатике . 9 (4): 276–285. doi : 10.1093/bib/bbn015 . PMID 18430752 .
- ^ Walian P, Cross TA, Jap BK (2004). «Структурная геномика мембранных белков» . Биология генома . 5 (4): 215. DOI : 10.1186/GB-2004-5-4-215 . PMC 395774 . PMID 15059248 .
- ^ Sleator Rd (2012). «Прогнозирование функций белка». Функциональная геномика . Методы в молекулярной биологии. Тол. 815. С. 15–24. doi : 10.1007/978-1-61779-424-7_2 . ISBN 978-1-61779-423-0 Полем PMID 22130980 .
- ^ Чжан Y (июнь 2008 г.). «Прогресс и проблемы в прогнозировании структуры белка» . Современное мнение в структурной биологии . 18 (3): 342–348. doi : 10.1016/j.sbi.2008.02.004 . PMC 2680823 . PMID 18436442 .
- ^ Xiang Z (июнь 2006 г.). «Достижения в моделировании структуры белка гомологии» . Текущая белковая и пептидная наука . 7 (3): 217–227. doi : 10.2174/13892030306777452312 . PMC 1839925 . PMID 16787261 .
- ^ Zhang Y, Skolnick J (январь 2005 г.). «Проблема прогнозирования структуры белка может быть решена с использованием текущей библиотеки PDB» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (4): 1029–1034. Bibcode : 2005pnas..102.1029Z . doi : 10.1073/pnas.0407152101 . PMC 545829 . PMID 15653774 .
- ^ Kuhlman B, Dantas G, Ireton GC, Varani G, Stoddard BL, Baker D (ноябрь 2003 г.). «Дизайн новой глобулярной белковой складки с точностью атомного уровня». Наука . 302 (5649): 1364–1368. Bibcode : 2003sci ... 302.1364K . doi : 10.1126/science.1089427 . PMID 14631033 . S2CID 1939390 .
- ^ Ward JJ, Sodhi JS, McGuffin LJ, Buxton BF, Jones DT (март 2004 г.). «Прогнозирование и функциональный анализ нативного расстройства в белках из трех королевств жизни». Журнал молекулярной биологии . 337 (3): 635–645. Citeseerx 10.1.1.120.5605 . doi : 10.1016/j.jmb.2004.02.002 . PMID 15019783 .
- ^ Tompa P, Fersht A (2009). Структура и функция внутренне неупорядоченных белков . CRC Press. ISBN 978-1-4200-7893-0 . [ страница необходима ]
- ^ Ritchie DW (февраль 2008 г.). «Недавний прогресс и будущие направления в стыковке белка белка». Текущая белковая и пептидная наука . 9 (1): 1–15. Citeseerx 10.1.1.211.4946 . doi : 10.2174/138920308783565741 . PMID 18336319 .
- ^ Zagrovic B, Snow CD, рубашки MR, Pande vs (ноябрь 2002 г.). «Моделирование складывания небольшого альфа-спирального белка в атомистических деталях с использованием вычислений по всему миру». Журнал молекулярной биологии . 323 (5): 927–937. Citeseerx 10.1.1.142.8664 . doi : 10.1016/s0022-2836 (02) 00997-x . PMID 12417204 .
- ^ Herges T, Wenzel W (январь 2005 г.). «В Silico складывание белка трех спиралей и характеристика его свободного ландшафта в полевом полевом поле». Письма о физическом обзоре . 94 (1): 018101. ARXIV : Физика/0310146 . Bibcode : 2005phrvl..94a8101h . doi : 10.1103/physrevlett.94.018101 . PMID 15698135 . S2CID 1477100 .
- ^ Хоффманн М., Ванко М., Стродель П., Кениг П.П., Фрауэнхайм Т., Шультен К. и др. (Август 2006 г.). «Настройка цвета у родопсинов: механизм спектрального сдвига между бактериодопсином и сенсорным родопсином II». Журнал Американского химического общества . 128 (33): 10808–10818. doi : 10.1021/ja062082i . PMID 16910676 .
- ^ Mendive-Tapia D, Mangaud E, Firmino T, De La Lande A, Desouter-Lecomte M, Meyer HD, et al. (Январь 2018). «Многомерное квантовое механическое моделирование переноса электронов и электронную когерентность в растениях криптохроме: роль начальных условий ванны». Журнал физической химии б . 122 (1): 126–136. doi : 10.1021/acs.jpcb.7b10412 . PMID 29216421 .
- ^ Strümpfer J, Schulten K (август 2012 г.). «Открытые расчеты квантовой динамики с уравнениями движения иерархии на параллельных компьютерах» . Журнал химической теории и вычислений . 8 (8): 2808–2816. doi : 10.1021/ct3003833 . PMC 3480185 . PMID 23105920 .
- ^ Scheraga HA, Khalili M, Liwo A (2007). «Динамика белка: обзор методов молекулярного моделирования». Ежегодный обзор физической химии . 58 : 57–83. Bibcode : 2007Arpc ... 58 ... 57 с . doi : 10.1146/annurev.physchem.58.032806.104614 . PMID 17034338 .
- ^ Муньос-Хуэрта Р.Ф., Гевара-Гонсалес Р.Г., Контрерас-Медина Л.М., Торрес-Пачеко I, Прадо-Оливарес Дж., Окамп-Веласкес Р.В. (август 2013 г.). «Обзор методов восприятия статуса азота у растений: преимущества, недостатки и последние достижения» . Датчики . 13 (8): 10823–10843. Bibcode : 2013senso..1310823M . doi : 10.3390/s130810823 . PMC 3812630 . PMID 23959242 .
- ^ Martin PD, Malley DF, Manning G, Fuller L (ноябрь 2002 г.). «Определение органического углерода и азота в почве на уровне поля с использованием спектроскопии в ближней инфракрасной линии». Канадский журнал по почвенной науке . 82 (4): 413–422. doi : 10.4141/s01-054 .
- ^ Brosnan JT (июнь 2003 г.). «Межорганский транспорт аминокислот и его регулирование» . Журнал питания . 133 (6 Suppl 1): 2068s - 2072s. doi : 10.1093/jn/133.6.2068s . PMID 12771367 .
- ^ Watson TD (декабрь 1998 г.). «Диета и кожные заболевания у собак и кошек» . Журнал питания . 128 (12 Suppl): 2783S - 2789S. doi : 10.1093/jn/128.12.2783s . PMID 9868266 .
- ^ Подпрыгнуть до: а беременный Case LP, Daristotle L, Hayek MG, Raasch MF (2010). Клык и кошачьи питание-е-книга: ресурс для профессионалов животных-компаньонов . Elsevier Health Sciences.
- ^ Gosline J, Lillie M, Carrington E, Guerette P, Ortlepp C, Savage K (февраль 2002 г.). Бейли А.Дж., Макмиллан Дж., Трут -Пар, Татхам А.С. (ред.). «Упругие белки: биологические роли и механические свойства» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 357 (1418): 121–132. doi : 10.1098/rstb.2001.1022 . PMC 1692928 . PMID 11911769 .
- ^ Maruyama K, Natori R, Unomura Y (июль 1976 г.). «Новый эластичный белок из мышц». Природа . 262 (5563): 58–60. Bibcode : 1976natur.262 ... 58m . doi : 10.1038/262058A0 . PMID 934326 .
- ^ Tskhovrebova L, Trinick J (февраль 2012 г.). «Создание мышечной упругих: структурная основа растяжения миомезина» . PLOS Биология . 10 (2): E1001264. doi : 10.1371/journal.pbio.1001264 . PMC 3279349 . PMID 22347814 .
- ^ Мизраджи Э., Аренденза Л, Лин Дж (ноябрь 1987). «Вязкоупругие модели для ферментов с несколькими конформационными состояниями». Журнал теоретической биологии . 129 (2): 163–175. Bibcode : 1987jthbi.129..163m . doi : 10.1016/s0022-5193 (87) 80010-3 . PMID 3455460 .
- ^ Schiller T, Scheibel T (2024-04-18). «Сколовые и биомиметические волокна на основе биомиметического белка и их применение» . Коммуникационные материалы . 5 (1): 56. Bibcode : 2024comat ... 5 ... 56 с . doi : 10.1038/s43246-024-00488-2 . ISSN 2662-4443 .
- ^ Sun J, He H, Zhao K, Cheng W, Li Y, Zhang P, et al. (Сентябрь 2023 г.). «Протеиновые волокна с самостоятельными механическими свойствами посредством динамической химии имин» . Природная связь . 14 (1): 5348. Bibcode : 2023natco..14.5348S . doi : 10.1038/s41467-023-41084-1 . PMC 10475138 . PMID 37660126 .
- ^ Подпрыгнуть до: а беременный в дюймовый и Guthold M, Liu W, Sparks EA, Jawerth LM, Peng L, Falvo M, et al. (2007-10-02). «Сравнение механических и структурных свойств фибриновых волокон с другими белковыми волокнами» . Клеточная биохимия и биофизика . 49 (3): 165–181. doi : 10.1007/s12013-007-9001-4 . PMC 3010386 . PMID 17952642 .
- ^ Wang K, Meng X, Guo Z (2021). «Структура эластина, синтез, регуляторный механизм и связь с сердечно -сосудистыми заболеваниями» . Границы в клеточной биологии и развитии . 9 : 596702. DOI : 10.3389/fcell.2021.596702 . PMC 8670233 . PMID 34917605 .
- ^ Девель Л., Тамбурро А.М. (февраль 1999 г.). «Эластин: молекулярное описание и функция». Международный журнал биохимии и клеточной биологии . 31 (2): 261–272. doi : 10.1016/s1357-2725 (98) 00098-3 . PMID 10216959 .
- ^ Подпрыгнуть до: а беременный Хури Л.Р., Попа I (ноябрь 2019). «Химическое развертывание белковых доменов индуцирует изменение формы в запрограммированных белковых гидрогелях» . Природная связь . 10 (1): 5439. Bibcode : 2019natco..10.5439K . doi : 10.1038/s41467-019-13312-0 . PMC 6884551 . PMID 31784506 .
- ^ Тан Р., Шин Дж., Хео Дж., Коул Б.Д., Хонг Дж, Джанг Й (октябрь 2020 г.). «Настройка структурной целостности и механических свойств глобулярных белковых пузырьков путем смешивания сшиваемых и не связанных с ними строительных блоков». Биомакромолекулы . 21 (10): 4336–4344. doi : 10.1021/acs.biomac.0c01147 . PMID 32955862 .
- ^ Souza PC, Alessandri R, Barnoud J, Thallmair S, Faustino I, Grünewald F, et al. (Апрель 2021 г.). «Мартини 3: Силовое поле общего назначения для крупнозернистой молекулярной динамики» (PDF) . Природные методы . 18 (4): 382–388. doi : 10.1038/s41592-021-01098-3 . PMID 33782607 .
- ^ «Домашняя страница Петра Шимчака» . www.fuw.edu.pl. Получено 2024-05-13 .
- ^ Mapplebeck S, Booth J, Shalashilin D (август 2021 г.). «Моделирование динамики тяги белка во втором времени с молекулярной динамикой в штучной упаковке» . Журнал химической физики . 155 (8): 085101. BIBCODE : 2021JCHPH.155H5101M . doi : 10.1063/5.0059321 . PMID 34470356 .
- ^ Carrion-Vazquez M, Marszalek PE, Oberhauser AF, Fernandez JM (сентябрь 1999 г.). «Атомная силовая микроскопия отражает фенотипы длины в отдельных белках» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (20): 11288–11292. Bibcode : 1999pnas ... 9611288c . doi : 10.1073/pnas.96.20.11288 . PMC 18026 . PMID 10500169 .
- ^ McKittrick J, Chen Py, Bodde SG, Yang W, Novitskaya EE, Meyers MA (2012-04-03). «Структура, функции и механические свойства кератина» . Jom . 64 (4): 449–468. Bibcode : 2012Jom .... 64D.449M . doi : 10.1007/s11837-012-0302-8 . ISSN 1047-4838 .
- ^ Ян Л., Ван дер Верф К.О., Фитие К.Ф., Беннинк М.Л., Дейкстра П.Дж., Фейджен Дж. (Март 2008 г.). «Механические свойства нативных и сшитых коллагеновых фибриллов типа I» . Биофизический журнал . 94 (6): 2204–2211. Bibcode : 2008bpj .... 94 2204y . Doi : 10.1529/biophysj.107.111013 . PMC 2257912 . PMID 18032556 .
- ^ Усветите HJ, Fleming PJ, Fleming KG, Sodt AJ (август 2018 г.). «Строительные блоки внешней мембраны: расчет общей модели эластичной энергии для белков мембраны β-баррель» . Журнал химической теории и вычислений . 14 (8): 4487–4497. doi : 10.1021/acs.jctc.8b00377 . PMC 6191857 . PMID 29979594 .
- ^ Мишра В., Хит Р.Дж. (август 2021 г.). «Структурные и биохимические особенности человеческого сывороточного альбумина, необходимый для эукариотической клеточной культуры» . Международный журнал молекулярных наук . 22 (16): 8411. DOI : 10.3390/IJMS22168411 . PMC 8395139 . PMID 34445120 .
- ^ Спенсер С.Дж., Ранганатан В.Т., Йетирадж А., Эндрюс Г.Т. (март 2024 г.). «Концентрационная зависимость упругих и вязкоупругих свойств водных растворов фиколла и бычьего сывороточного альбумина с помощью спектроскопии легкого рассеяния Brillouin». Langmuir: ACS Journal of Turfaces и Colloids . 40 (9): 4615–4622. Arxiv : 2309.10967 . doi : 10.1021/acs.langmuir.3c02967 . PMID 38387073 .
- ^ Monkos K (февраль 1996 г.). «Вязкость водных растворов бычьего сывороточного альбумина как функция температуры и концентрации». Международный журнал биологических макромолекул . 18 (1–2): 61–68. doi : 10.1016/0141-8130 (95) 01057-2 . PMID 8852754 .
Дальнейшее чтение
- Учебники
- Бранден С., Туз Дж. (1999). Введение в структуру белка . Нью -Йорк: Garland Pub. ISBN 978-0-8153-2305-1 .
- Murray RF, Harper HW, Granner DK, Mayes PA, Rodwell VW (2006). Иллюстрированная биохимия Харпера . Нью-Йорк: Lange Medical Books/McGraw-Hill. ISBN 978-0-07-146197-9 .
- Ван Холде К.Е., Мэтьюз К.К. (1996). Биохимия . Менло Парк, Калифорния: Бенджамин/Каммингс Паб. Co., Inc. ISBN 978-0-8053-3931-4 .
Внешние ссылки
 Определения из Wiktionary
Определения из Wiktionary  СМИ из Commons
СМИ из Commons
Базы данных и проекты
- База данных белков NCBI Entrez
- База данных структуры белка NCBI
- Справочная база данных белка человека
- Человеческая протеинпедия
- Фолдинга@Home (Стэнфордский университет) Архивировал 2012-09-08 на The Wayback Machine
- Databank Protein в Европе (см. Также Pdbequips , короткие статьи и учебные пособия по интересным структурам PDB)
- Исследования сотрудничают по структурной биоинформатике (см. Также молекулу месяца, архивировав 2020-07-24 на машине Wayback , представляя короткие отчеты о выбранных белках из PDB)
- Протеопедия - жизнь в 3D : вращающаяся, увеличиваемая 3D -модель с вики -аннотациями для каждой известной молекулярной структуры белка.
- Uniprot Универсальный белковый ресурс
Учебные пособия и образовательные сайты
- «Введение в белки» из надежд (Проект по информационным технологиям в Хантингтоне для образования в Стэнфорде)
- Белки: биогенез деградации - виртуальная библиотека биохимии и клеточной биологии