Эволюция
| Часть серии по биологии . |
| Эволюция |
|---|
 |
| Механизмы и процессы |
| Исследования и история |
| эволюционной биологии Области |
Эволюция — это изменение наследственных характеристик биологических популяций на протяжении последующих поколений. [1] [2] Это происходит, когда эволюционные процессы, такие как естественный отбор и генетический дрейф, воздействуют на генетическую изменчивость, в результате чего определенные характеристики становятся более или менее распространенными в популяции на протяжении последующих поколений. [3] Процесс эволюции привел к появлению биоразнообразия на всех уровнях биологической организации . [4] [5]
Научная теория эволюции путем естественного отбора была независимо разработана двумя британскими натуралистами, Чарльзом Дарвином и Альфредом Расселом Уоллесом , в середине XIX века как объяснение того, почему организмы адаптируются к своей физической и биологической среде. Впервые теория была подробно изложена в книге Дарвина «Происхождение видов» . [6] Эволюция путем естественного отбора подтверждается наблюдаемыми фактами о живых организмах: (1) часто рождается больше потомства, чем может выжить; (2) черты различаются у разных людей в зависимости от их морфологии , физиологии и поведения; (3) разные признаки обеспечивают разную степень выживаемости и воспроизводства (дифференцированная приспособленность ); и (4) черты могут передаваться из поколения в поколение ( наследственность приспособленности). [7] Поэтому в последующих поколениях члены популяции с большей вероятностью будут заменены потомками родителей с благоприятными характеристиками для этой среды.
В начале 20-го века конкурирующие идеи эволюции были опровергнуты , и эволюция была объединена с менделевской наследственностью и популяционной генетикой , что дало начало современной эволюционной теории. [8] В этом синтезе основой наследственности являются молекулы ДНК , передающие информацию из поколения в поколение. Процессы, которые изменяют ДНК в популяции, включают естественный отбор, генетический дрейф, мутации и поток генов . [3]
Вся жизнь на Земле, включая человечество , имеет общего последнего универсального предка (LUCA). [9] [10] [11] которые жили примерно 3,5–3,8 миллиарда лет назад. [12] Летопись окаменелостей включает в себя развитие раннего биогенного графита. [13] в микробного мата окаменелости [14] [15] [16] окаменевшим многоклеточным организмам . Существующие модели биоразнообразия формировались в результате повторяющихся образований новых видов ( видообразование ), изменений внутри видов ( анагенез ) и утраты видов ( вымирание ) на протяжении всей эволюционной истории жизни на Земле. [17] Морфологические и биохимические черты, как правило, более схожи между видами, имеющими более недавнего общего предка , который исторически использовался для реконструкции филогенетических деревьев , хотя прямое сравнение генетических последовательностей сегодня является более распространенным методом. [18] [19]
Эволюционные биологи продолжали изучать различные аспекты эволюции, формируя и проверяя гипотезы , а также создавая теории, основанные на полевых или лабораторных данных , а также на данных, полученных методами математической и теоретической биологии . Их открытия повлияли не только на развитие биологии , но и на другие области, включая сельское хозяйство, медицину и информатику . [20]
Наследственность

Эволюция организмов происходит посредством изменения наследственных характеристик — унаследованных характеристик организма. У людей, например, цвет глаз является наследственной характеристикой, и человек может унаследовать «черту карих глаз» от одного из своих родителей. [21] организма Наследственные признаки контролируются генами, а полный набор генов в геноме (генетический материал) называется его генотипом . [22]
Полный набор наблюдаемых признаков, составляющих строение и поведение организма, называется его фенотипом . Некоторые из этих черт возникают в результате взаимодействия его генотипа с окружающей средой, тогда как другие нейтральны. [23] Некоторые наблюдаемые характеристики не наследуются. Например, загорелая кожа возникает в результате взаимодействия генотипа человека и солнечного света; таким образом, загар не передается детям. Фенотип – это способность кожи загорать под воздействием солнечного света. Однако некоторые люди загорают легче, чем другие, из-за различий в генотипических вариациях; Ярким примером являются люди с наследственной чертой альбинизма , которые вообще не загорают и очень чувствительны к солнечным ожогам . [24]
Наследственные характеристики передаются от одного поколения к другому через ДНК , — молекулу которая кодирует генетическую информацию. [22] ДНК представляет собой длинный биополимер, состоящий из четырех типов оснований. Последовательность оснований в конкретной молекуле ДНК определяет генетическую информацию подобно последовательности букв, составляющих предложение. Прежде чем клетка делится, ДНК копируется, так что каждая из двух образовавшихся клеток унаследует последовательность ДНК. Части молекулы ДНК, определяющие одну функциональную единицу, называются генами; разные гены имеют разные последовательности оснований. Внутри клеток каждая длинная цепь ДНК называется хромосомой . Конкретное расположение последовательности ДНК внутри хромосомы известно как локус . Если последовательность ДНК в локусе различается у разных людей, разные формы этой последовательности называются аллелями. Последовательности ДНК могут меняться в результате мутаций, создавая новые аллели. Если внутри гена происходит мутация, новая аллель может повлиять на признак, который контролирует ген, изменяя фенотип организма. [25] Однако, хотя это простое соответствие между аллелем и признаком в некоторых случаях работает, на большинство признаков влияют многочисленные гены количественным или эпистатическим образом . [26] [27]
Источники вариаций


Эволюция может произойти, если внутри популяции существуют генетические вариации. Вариации возникают в результате мутаций в геноме, перетасовки генов посредством полового размножения и миграции между популяциями ( поток генов ). Несмотря на постоянное появление новых вариаций посредством мутаций и потока генов, большая часть генома вида очень похожа у всех особей этого вида. [28] Однако открытия в области эволюционной биологии развития показали, что даже относительно небольшие различия в генотипе могут привести к резким различиям в фенотипе как внутри вида, так и между видами.
Фенотип отдельного организма зависит как от его генотипа, так и от влияния среды, в которой он жил. [27] Современный эволюционный синтез определяет эволюцию как изменение во времени этой генетической вариации. Частота одного конкретного аллеля станет более или менее преобладающей по сравнению с другими формами этого гена. Вариация исчезает, когда новый аллель достигает точки фиксации — когда он либо исчезает из популяции, либо полностью заменяет предковый аллель. [29]
Мутация
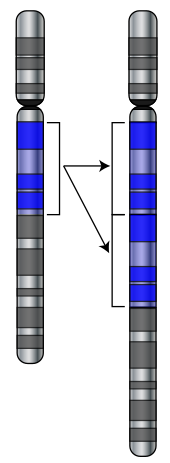
Мутации — это изменения в последовательности ДНК клеточного генома, которые являются основным источником генетических вариаций во всех организмах. [30] Когда происходят мутации, они могут изменить продукт гена , помешать функционированию гена или не оказать никакого эффекта.
Около половины мутаций в кодирующих областях генов, кодирующих белки, являются вредными, а другая половина — нейтральными. Небольшой процент всех мутаций в этом регионе дает преимущества в фитнесе. [31] Некоторые мутации в других частях генома вредны, но подавляющее большинство нейтральны. Некоторые из них полезны.
больших участков хромосомы Мутации могут включать в себя дублирование (обычно путем генетической рекомбинации ), что может привести к появлению дополнительных копий гена в геноме. [32] Дополнительные копии генов являются основным источником сырья, необходимого для эволюции новых генов. [33] Это важно, поскольку большинство новых генов развиваются внутри семейств генов из ранее существовавших генов, имеющих общих предков. [34] Например, человеческий глаз использует четыре гена для создания структур, воспринимающих свет: три для цветового зрения и один для ночного зрения ; все четверо произошли от одного предкового гена. [35]
Новые гены могут быть созданы из предкового гена, когда дубликат мутирует и приобретает новую функцию. Этот процесс становится проще, если ген дублируется, поскольку это увеличивает избыточность системы; один ген в паре может приобрести новую функцию, в то время как другая копия продолжает выполнять свою первоначальную функцию. [36] [37] Другие типы мутаций могут даже генерировать совершенно новые гены из ранее некодирующей ДНК — явление, называемое de novo рождением генов . [38] [39]
Генерация новых генов также может включать дублирование небольших частей нескольких генов, при этом эти фрагменты затем рекомбинируются с образованием новых комбинаций с новыми функциями ( перетасовка экзонов ). [40] [41] Когда новые гены собираются из перетасованных ранее существовавших частей, домены действуют как модули с простыми независимыми функциями, которые можно смешивать вместе для создания новых комбинаций с новыми и сложными функциями. [42] Например, поликетидсинтазы — это крупные ферменты , вырабатывающие антибиотики ; они содержат до 100 независимых доменов, каждый из которых катализирует один этап общего процесса, как этап на сборочной линии. [43]
Одним из примеров мутации являются поросята диких кабанов . Они имеют камуфляжную окраску и имеют характерный рисунок из темных и светлых продольных полос. Однако мутации в рецепторе меланокортина 1 ( MC1R ) нарушают эту закономерность. Большинство пород свиней несут мутации MC1R, нарушающие окраску дикого типа, а также различные мутации, вызывающие доминирующую черную окраску. [44]
Секс и рекомбинация
У бесполых организмов гены наследуются вместе или сцеплены , поскольку они не могут смешиваться с генами других организмов во время размножения. Напротив, потомство половых организмов содержит случайные смеси хромосом своих родителей, которые производятся посредством независимого ассортимента. В родственном процессе, называемом гомологичной рекомбинацией , половые организмы обмениваются ДНК между двумя совпадающими хромосомами. [45] Рекомбинация и реассортация не изменяют частоты аллелей, а вместо этого изменяют то, какие аллели связаны друг с другом, производя потомство с новыми комбинациями аллелей. [46] Секс обычно увеличивает генетическую изменчивость и может увеличить скорость эволюции. [47] [48]

Двойная стоимость секса была впервые описана Джоном Мейнардом Смитом . [49] Первая цена заключается в том, что у видов с половым диморфизмом только один из двух полов может вынашивать потомство. Эта стоимость не распространяется на гермафродитные виды, такие как большинство растений и многих беспозвоночных . Вторая цена заключается в том, что любая особь, размножающаяся половым путем, может передать только 50% своих генов любому отдельному потомку, а с каждым новым поколением передается еще меньше. [50] Однако половое размножение является более распространенным способом размножения среди эукариот и многоклеточных организмов. Гипотеза Красной Королевы использовалась для объяснения значения полового размножения как средства, обеспечивающего непрерывную эволюцию и адаптацию в ответ на коэволюцию с другими видами в постоянно меняющейся среде. [50] [51] [52] [53] Другая гипотеза состоит в том, что половое размножение — это, прежде всего, адаптация, способствующая точному рекомбинационному восстановлению повреждений в ДНК зародышевой линии, и что увеличение разнообразия является побочным продуктом этого процесса, который иногда может быть адаптивно полезным. [54] [55]
Поток генов
Поток генов — это обмен генами между популяциями и между видами. [56] Следовательно, он может быть источником вариаций, новых для популяции или вида. Поток генов может быть вызван перемещением особей между отдельными популяциями организмов, как это может быть вызвано перемещением мышей между внутренними и прибрежными популяциями или перемещением пыльцы между толерантными к тяжелым металлам и чувствительными к тяжелым металлам популяциями. травы.
Перенос генов между видами включает образование гибридных организмов и горизонтальный перенос генов . Горизонтальный перенос генов — это передача генетического материала от одного организма к другому организму, не являющемуся его потомком; это наиболее распространено среди бактерий. [57] В медицине это способствует распространению устойчивости к антибиотикам , поскольку, когда одна бактерия приобретает гены устойчивости, она может быстро передать их другим видам. [58] горизонтальный перенос генов от бактерий к эукариотам, таким как дрожжи Saccharomyces cerevisiae и долгоносик адзуки Callosobruchus chinensis . Произошел [59] [60] Примером крупномасштабного переноса являются эукариотические бделлоидные коловратки , которые получили ряд генов от бактерий, грибов и растений. [61] Вирусы также могут переносить ДНК между организмами, что позволяет передавать гены даже между биологическими доменами . [62]
Крупномасштабный перенос генов произошел также между предками эукариотических клеток и бактерий во время приобретения хлоропластов и митохондрий . Возможно, что сами эукариоты произошли в результате горизонтального переноса генов между бактериями и архей . [63]
Эпигенетика
Некоторые наследственные изменения нельзя объяснить изменениями последовательности нуклеотидов в ДНК. Эти явления классифицируются как эпигенетические системы наследования. [64] Метилирование ДНК маркирующее хроматин , самоподдерживающиеся метаболические петли, подавление генов посредством интерференции РНК и трехмерная конформация белков , (таких как прионы ) — это области, в которых эпигенетические системы наследования были обнаружены на организменном уровне. [65] Биологи развития предполагают, что сложные взаимодействия в генетических сетях и коммуникации между клетками могут привести к наследственным вариациям, которые могут лежать в основе некоторых механизмов развития пластичности и канализации . [66] Наследственность может также проявляться в еще более крупных масштабах. Например, экологическое наследование в процессе построения ниш определяется регулярной и повторяющейся деятельностью организмов в окружающей среде. Это порождает наследие эффектов, которые модифицируют и возвращаются в режим отбора последующих поколений. [67] Другие примеры наследственности в эволюции, не находящиеся под прямым контролем генов, включают наследование культурных признаков и симбиогенез . [68] [69]
Эволюционные силы

С неодарвинистской точки зрения эволюция происходит, когда происходят изменения в частотах аллелей внутри популяции скрещивающихся организмов. [70] например, аллель черного цвета в популяции бабочек становится все более распространенным. Механизмы, которые могут привести к изменениям частот аллелей, включают естественный отбор, генетический дрейф и предвзятость мутаций.
Естественный отбор
Эволюция путем естественного отбора — это процесс, в ходе которого черты, способствующие выживанию и воспроизводству, становятся более распространенными в последующих поколениях популяции. Он воплощает в себе три принципа: [7]
- В популяциях организмов существуют вариации в отношении морфологии, физиологии и поведения (фенотипические вариации).
- Разные черты определяют разную степень выживаемости и воспроизводства (дифференциальная приспособленность).
- Эти черты могут передаваться из поколения в поколение (наследственность приспособленности).
Производится больше потомства, чем может выжить, и эти условия вызывают конкуренцию между организмами за выживание и размножение. Следовательно, организмы с признаками, которые дают им преимущество перед конкурентами, с большей вероятностью передадут свои черты следующему поколению, чем организмы с признаками, которые не дают преимущества. [71] Эта телеономия — это качество, благодаря которому процесс естественного отбора создает и сохраняет черты, которые, по-видимому, подходят для выполнения функциональных ролей, которые они выполняют. [72] Последствия отбора включают неслучайное спаривание . [73] и генетический автостоп .
Центральная концепция естественного отбора — эволюционная приспособленность организма. [74] Пригодность измеряется способностью организма выживать и размножаться, что определяет размер его генетического вклада в следующее поколение. [74] Однако приспособленность — это не то же самое, что общее число потомков: вместо этого приспособленность определяется долей последующих поколений, несущих гены организма. [75] Например, если бы организм мог хорошо выживать и быстро размножаться, но его потомство было бы слишком маленьким и слабым, чтобы выжить, этот организм внес бы небольшой генетический вклад в будущие поколения и, следовательно, имел бы низкую приспособленность. [74]
Если аллель увеличивает приспособленность больше, чем другие аллели этого гена, то с каждым поколением этот аллель имеет более высокую вероятность стать распространенным в популяции. Говорят, что эти черты «отбираются для ». Примерами черт, которые могут повысить приспособленность, являются повышенная выживаемость и повышенная плодовитость . И наоборот, более низкая приспособленность, вызванная наличием менее полезного или вредного аллеля, приводит к тому, что этот аллель, вероятно, становится более редким — они «отбираются против ». [76]
Важно отметить, что приспособленность аллели не является фиксированной характеристикой; если окружающая среда изменится, ранее нейтральные или вредные черты могут стать полезными, а ранее полезные черты станут вредными. [25] Однако даже если направление отбора таким образом изменится, черты, утраченные в прошлом, не смогут вновь развиться в идентичной форме. [77] [78] Однако повторная активация спящих генов, пока они не были удалены из генома, а лишь подавлялись, возможно, на протяжении сотен поколений, может привести к повторному появлению признаков, которые, как считается, были утеряны, например, задние ноги у дельфинов, зубы у кур, крылья у бескрылых палочников, хвосты и дополнительные соски у людей и т. д. Подобные «возвраты» известны как атавизмы . [79]

· График 1 показывает направленный отбор одному крайнему фенотипу . , при котором предпочтение отдается
· График 2 изображает стабилизирующий отбор , при котором промежуточный фенотип имеет преимущество над крайними признаками.
· На графике 3 показан разрушительный отбор , при котором крайние фенотипы имеют преимущество перед промежуточными.
Естественный отбор внутри популяции по признаку, который может варьироваться в зависимости от диапазона значений, например, по росту, можно разделить на три разных типа. Первый — направленный отбор , который представляет собой сдвиг среднего значения признака с течением времени — например, организмы медленно становятся выше. [80] Во-вторых, разрушительный отбор — это отбор по крайним значениям признаков, который часто приводит к тому, что наиболее распространенными становятся два разных значения , при этом отбор осуществляется против среднего значения. Это было бы тогда, когда преимущество имели либо низкие, либо высокие организмы, но не организмы среднего роста. Наконец, стабилизирующий отбор осуществляется против крайних значений признаков с обеих сторон, что приводит к уменьшению дисперсии вокруг среднего значения и меньшему разнообразию. [71] [81] Это, например, привело бы к тому, что организмы в конечном итоге стали бы иметь одинаковую высоту.
Естественный отбор в большинстве случаев делает природу мерилом, при котором индивидуумы и отдельные черты с большей или меньшей вероятностью выживут. «Природа» в этом смысле относится к экосистеме , то есть системе, в которой организмы взаимодействуют со всеми другими элементами, как физическими , так и биологическими , в своей местной среде обитания. Юджин Одум , основатель экологии, определил экосистему как: «Любая единица, включающая в себя все организмы... на данной территории, взаимодействующая с физической средой так, что поток энергии приводит к четко определенной трофической структуре, биотическому разнообразию, и материальные циклы (т.е. обмен веществами между живыми и неживыми частями) внутри системы...». [82] Каждая популяция в экосистеме занимает отдельную нишу или положение с разными отношениями с другими частями системы. Эти отношения включают историю жизни организма, его положение в пищевой цепи и его географический ареал. Такое широкое понимание природы позволяет ученым определить конкретные силы, которые в совокупности составляют естественный отбор.
Естественный отбор может действовать на разных уровнях организации , таких как гены, клетки, отдельные организмы, группы организмов и виды. [83] [84] [85] Отбор может действовать на нескольких уровнях одновременно. [86] Примером отбора, происходящего ниже уровня отдельного организма, являются гены, называемые транспозонами , которые могут реплицироваться и распространяться по всему геному. [87] Отбор на уровне выше индивидуального, например групповой отбор , может способствовать развитию сотрудничества. [88]
Генетический дрейф

Генетический дрейф — это случайное колебание частот аллелей внутри популяции от одного поколения к другому. [89] Когда силы отбора отсутствуют или относительно слабы, частоты аллелей с одинаковой вероятностью смещаются вверх или вниз. [ нужны разъяснения ] в каждом последующем поколении, поскольку аллели подвержены ошибкам выборки . [90] Этот дрейф прекращается, когда аллель в конечном итоге становится фиксированной, либо исчезая из популяции, либо полностью заменяя другие аллели. Таким образом, генетический дрейф может исключить некоторые аллели из популяции только по случайности. Даже в отсутствие сил отбора генетический дрейф может привести к тому, что две отдельные популяции, изначально имеющие одну и ту же генетическую структуру, разделятся на две расходящиеся популяции с разными наборами аллелей. [91]
Согласно нейтральной теории молекулярной эволюции, большинство эволюционных изменений являются результатом фиксации нейтральных мутаций в результате генетического дрейфа. [92] Таким образом, в этой модели большинство генетических изменений в популяции являются результатом постоянного мутационного давления и генетического дрейфа. [93] Эта форма нейтральной теории обсуждается, поскольку она, похоже, не соответствует каким-либо генетическим вариациям, наблюдаемым в природе. [94] [95] Более обоснованной версией этой модели является почти нейтральная теория , согласно которой мутация, которая была бы фактически нейтральной в небольшой популяции, не обязательно является нейтральной в большой популяции. [71] Другие теории предполагают, что генетический дрейф затмевается другими стохастическими силами эволюции, такими как генетический автостоп, также известный как генетический драфт. [90] [96] [97] Другая концепция — конструктивная нейтральная эволюция (CNE), которая объясняет, что сложные системы могут возникать и распространяться в популяции посредством нейтральных переходов благодаря принципам избыточной мощности, пресупрессии и храповика. [98] [99] [100] и он применялся в самых разных областях: от происхождения сплайсосом до сложной взаимозависимости микробных сообществ . [101] [102] [103]
Время, необходимое нейтральному аллелю для закрепления в результате генетического дрейфа, зависит от размера популяции; фиксация происходит быстрее в небольших популяциях. [104] Число особей в популяции не имеет решающего значения, а является мерой, известной как эффективный размер популяции. [105] Эффективная популяция обычно меньше общей популяции, поскольку она учитывает такие факторы, как уровень инбридинга и стадию жизненного цикла, на которой популяция наименьшая. [105] Эффективный размер популяции может не быть одинаковым для каждого гена в одной и той же популяции. [106]
Обычно трудно измерить относительную важность отбора и нейтральных процессов, включая дрейф. [107] Сравнительная важность адаптивных и неадаптивных сил в движущих силах эволюционных изменений является областью текущих исследований . [108]
Предвзятость мутации
Смещение мутации обычно понимается как разница в ожидаемых скоростях для двух разных типов мутаций, например, смещение переход-трансверсия, смещение GC-AT, смещение делеции-вставки. Это связано с идеей отклонения в развитии . Холдейн [109] и Фишер [110] утверждал, что, поскольку мутация представляет собой слабое давление, которое легко преодолевается отбором, тенденции к мутациям будут неэффективными, за исключением условий нейтральной эволюции или чрезвычайно высоких темпов мутаций. Этот аргумент противоположного давления долгое время использовался, чтобы отвергнуть возможность внутренних тенденций в эволюции. [111] пока молекулярная эра не вызвала возобновление интереса к нейтральной эволюции.
Нобору Суэока [112] и Эрнст Фриз [113] предположил, что систематические ошибки в мутациях могут быть ответственны за систематические различия в геномном составе GC между видами. Идентификация мутаторного штамма E. coli , ориентированного на GC , в 1967 году. [114] наряду с предложением нейтральной теории установил правдоподобие мутационных объяснений молекулярных закономерностей, которые сейчас распространены в литературе по молекулярной эволюции.
Например, мутационные ошибки часто используются в моделях использования кодонов. [115] Такие модели также включают эффекты отбора в соответствии с моделью мутации-отбора-дрейфа. [116] что допускает как мутационные ошибки, так и дифференциальный отбор, основанный на влиянии на трансляцию. Гипотезы мутационной предвзятости сыграли важную роль в развитии представлений об эволюции состава генома, включая изохоры. [117] Различная систематическая ошибка вставки или делеции в разных таксонах может привести к эволюции геномов разного размера. [118] [119] Гипотеза Линча относительно размера генома основана на мутационной склонности к увеличению или уменьшению размера генома.
Однако мутационные гипотезы эволюции состава уменьшились в своих масштабах, когда было обнаружено, что (1) конверсия генов, обусловленная GC, вносит важный вклад в состав диплоидных организмов, таких как млекопитающие. [120] и (2) бактериальные геномы часто имеют мутации, связанные с АТ. [121]
Современные размышления о роли мутационных предубеждений отражают теорию, отличную от теории Холдейна и Фишера. Более поздние работы [111] показали, что первоначальная теория «давления» предполагает, что эволюция основана на постоянной изменчивости: когда эволюция зависит от событий мутации, которые приводят к появлению новых аллелей, мутационные и онтогенетические отклонения при введении изменчивости (предвзятости прибытия) могут накладывать предвзятости на эволюцию, не требуя нейтральности. эволюция или высокая скорость мутаций. [111] [122] В нескольких исследованиях сообщается, что мутации, участвующие в адаптации, отражают распространенные мутационные предубеждения. [123] [124] [125] хотя другие оспаривают эту интерпретацию. [126]
Генетический автостоп
Рекомбинация позволяет аллелям на одной и той же цепи ДНК разделиться. Однако скорость рекомбинации низкая (примерно два события на хромосому за поколение). В результате гены, расположенные близко друг к другу на хромосоме, не всегда могут быть отодвинуты друг от друга, а гены, расположенные близко друг к другу, имеют тенденцию наследоваться вместе — явление, известное как сцепление . [127] Эта тенденция измеряется путем определения того, как часто два аллеля встречаются вместе на одной хромосоме по сравнению с ожидаемым , что называется их неравновесием по сцеплению . Набор аллелей, который обычно наследуется в группе, называется гаплотипом . Это может быть важно, когда одна аллель в определенном гаплотипе очень полезна: естественный отбор может привести к избирательному охвату , который также приведет к тому, что другие аллели в гаплотипе станут более распространенными в популяции; этот эффект называется генетическим автостопом или генетическим проектом. [128] Генетический выброс, вызванный тем фактом, что некоторые нейтральные гены генетически связаны с другими, находящимися в стадии селекции, может быть частично учтен соответствующим эффективным размером популяции. [96]
Половой отбор

Особым случаем естественного отбора является половой отбор, который представляет собой отбор по любому признаку, который увеличивает успех спаривания за счет увеличения привлекательности организма для потенциальных партнеров. [130] Признаки, развившиеся в результате полового отбора, особенно заметны у самцов некоторых видов животных. Несмотря на сексуальное предпочтение, такие черты, как громоздкие рога, брачные крики, большой размер тела и яркая окраска, часто привлекают хищников, что ставит под угрозу выживание отдельных самцов. [131] [132] Этот недостаток выживания уравновешивается более высоким репродуктивным успехом у самцов, которые демонстрируют эти трудно поддельные , отобранные половым путем черты. [133]
Естественные результаты
Эволюция влияет на каждый аспект формы и поведения организмов. Наиболее заметными являются специфические поведенческие и физические адаптации, являющиеся результатом естественного отбора. Эти адаптации повышают физическую форму, помогая таким действиям, как поиск еды, избегание хищников или привлечение партнеров. Организмы также могут реагировать на отбор, сотрудничая друг с другом, обычно помогая своим родственникам или вступая во взаимовыгодный симбиоз . В долгосрочной перспективе эволюция производит новые виды путем разделения предковых популяций организмов на новые группы, которые не могут или не будут скрещиваться. Эти результаты эволюции различаются по временной шкале: макроэволюция и микроэволюция. Макроэволюция относится к эволюции, которая происходит на уровне вида или выше, в частности видообразованию и вымиранию, тогда как микроэволюция относится к меньшим эволюционным изменениям внутри вида или популяции, в частности, к сдвигам частоты аллелей и адаптации. [135] Макроэволюция – это результат длительных периодов микроэволюции. [136] Таким образом, различие между микро- и макроэволюцией не является фундаментальным — разница лишь во времени. [137] Однако в макроэволюции могут иметь значение черты всего вида. Например, большое количество вариаций среди особей позволяет виду быстро адаптироваться к новым местам обитания , уменьшая вероятность его вымирания, в то время как широкий географический диапазон увеличивает вероятность видообразования, повышая вероятность того, что часть популяции вымрет. оказаться в изоляции. В этом смысле микроэволюция и макроэволюция могут включать отбор на разных уровнях: микроэволюция воздействует на гены и организмы, а не макроэволюционные процессы, такие как отбор видов , воздействующие на целые виды и влияющие на скорость их видообразования и вымирания. [138] [139] [140]
Распространенным заблуждением является то, что у эволюции есть цели, долгосрочные планы или врожденная тенденция к «прогрессу», что выражается в таких убеждениях, как ортогенез и эволюционизм; однако на самом деле эволюция не имеет долгосрочной цели и не обязательно приводит к увеличению сложности. [141] [142] [143] Хотя сложные виды эволюционировали, они возникают как побочный эффект увеличения общего числа организмов, а простые формы жизни по-прежнему остаются более распространенными в биосфере. [144] Например, подавляющее большинство видов — это микроскопические прокариоты формируют около половины мировой биомассы . , которые, несмотря на свои небольшие размеры, [145] и составляют подавляющее большинство биоразнообразия Земли. [146] Таким образом, простые организмы были доминирующей формой жизни на Земле на протяжении всей ее истории и продолжают оставаться основной формой жизни до наших дней, а сложная жизнь кажется более разнообразной только потому, что она более заметна . [147] Действительно, эволюция микроорганизмов особенно важна для эволюционных исследований, поскольку их быстрое размножение позволяет изучать экспериментальную эволюцию и наблюдать за эволюцией и адаптацией в реальном времени. [148] [149]
Приспособление

Адаптация – это процесс, который делает организмы более приспособленными к среде их обитания. [150] [151] Кроме того, термин «адаптация» может относиться к черте, важной для выживания организма. Например, приспособление зубов лошадей к скрежетанию травы. Используя термин «адаптация» для обозначения эволюционного процесса и «адаптивная черта» для продукта (части или функции тела), можно различить два значения этого слова. Адаптации производятся естественным отбором. [152] Следующие определения принадлежат Феодосию Добжанскому:
- Адаптация – это эволюционный процесс, в ходе которого организм становится более способным жить в своей среде обитания или средах обитания. [153]
- Адаптированность — это состояние адаптированности: степень, в которой организм способен жить и размножаться в заданном наборе сред обитания. [154]
- Адаптивная черта — это аспект модели развития организма, который обеспечивает или повышает вероятность выживания и размножения этого организма. [155]
Адаптация может привести либо к приобретению нового признака, либо к потере наследственного признака. Примером, показывающим оба типа изменений, является адаптация бактерий к отбору антибиотиков, при этом генетические изменения вызывают устойчивость к антибиотикам, изменяя мишень лекарства или увеличивая активность транспортеров, которые выкачивают лекарство из клетки. [156] Другими яркими примерами являются бактерии Escherichia coli, развили способность использовать лимонную кислоту в качестве питательного вещества которые в ходе длительного лабораторного эксперимента . [157] Flavobacterium вырабатывает новый фермент, который позволяет этим бактериям расти на побочных продуктах производства нейлона. [158] [159] и почвенная бактерия Sphingobium развивает совершенно новый метаболический путь , который разлагает синтетический пестицид пентахлорфенол . [160] [161] Интересная, но все еще спорная идея заключается в том, что некоторые адаптации могут повысить способность организмов генерировать генетическое разнообразие и адаптироваться посредством естественного отбора (увеличивая способность организмов к эволюции). [162] [163] [164] [165]
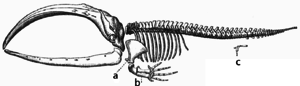
Адаптация происходит посредством постепенной модификации существующих структур. Следовательно, структуры со сходной внутренней организацией могут иметь разные функции у родственных организмов. Это результат того, что единая наследственная структура была адаптирована для функционирования по-разному. Кости крыльев летучей мыши, например, очень похожи на кости ног мышей и рук приматов , поскольку все эти структуры произошли от общего предка млекопитающих. [167] Однако, поскольку все живые организмы в той или иной степени родственны, [168] даже органы, которые, по-видимому, имеют небольшое структурное сходство или вообще не имеют никакого структурного сходства, такие как глаза членистоногих , кальмаров и позвоночных или конечности и крылья членистоногих и позвоночных, могут зависеть от общего набора гомологичных генов, которые контролируют их сборку и функцию; это называется глубокой гомологией . [169] [170]
В ходе эволюции некоторые структуры могут утратить свою первоначальную функцию и стать рудиментарными структурами. [171] Такие структуры могут иметь небольшую функцию или вообще не выполнять ее у современных видов, но иметь четкую функцию у предковых видов или других близкородственных видов. Примеры включают псевдогены , [172] нефункциональные остатки глаз у слепых пещерных рыб, [173] крылья у нелетающих птиц, [174] наличие бедренных костей у китов и змей, [166] и половые признаки у организмов, размножающихся бесполым путем. [175] Примеры рудиментарных структур у человека включают зубы мудрости , [176] копчик , [171] отросток червеобразный , [171] и другие поведенческие проявления, такие как мурашки по коже [177] [178] и примитивные рефлексы . [179] [180] [181]
Однако многие черты, которые кажутся простыми адаптациями, на самом деле являются экзаптациями : структуры, изначально приспособленные для одной функции, но по совпадению в процессе стали в некоторой степени полезными для какой-то другой функции. [182] Одним из примеров является африканская ящерица Holaspis guentheri , у которой развилась чрезвычайно плоская голова, позволяющая прятаться в расщелинах, в чем можно убедиться, посмотрев на ее ближайших родственников. Однако у этого вида голова настолько уплощена, что помогает скользить с дерева на дерево — экзаптация. [182] Внутри клеток существуют молекулярные механизмы, такие как бактериальные жгутики . [183] и машины для сортировки белков [184] эволюционировал за счет привлечения нескольких ранее существовавших белков, ранее выполнявших разные функции. [135] Другим примером является использование ферментов гликолиза и метаболизма ксенобиотиков в качестве структурных белков, называемых кристаллинами, в хрусталике глаз организмов. [185] [186]
Областью текущих исследований эволюционной биологии развития являются основы развития адаптаций и экзаптаций. [187] Это исследование посвящено происхождению и эволюции эмбрионального развития , а также тому, как модификации развития и процессов развития приводят к возникновению новых особенностей. [188] Эти исследования показали, что эволюция может изменить развитие, создав новые структуры, такие как эмбриональные костные структуры, которые развиваются в челюсть у других животных, а не образуют часть среднего уха у млекопитающих . [189] Также возможно, что структуры, утраченные в ходе эволюции, вновь появятся из-за изменений в генах развития, например, мутации у кур, в результате которой у эмбрионов вырастают зубы, похожие на зубы крокодилов. [190] Сейчас становится ясно, что большинство изменений в форме организмов происходят из-за изменений в небольшом наборе консервативных генов. [191]
Коэволюция

Взаимодействия между организмами могут приводить как к конфликту, так и к сотрудничеству. Когда взаимодействие происходит между парами видов, например, патогеном и хозяином или хищником и его добычей, эти виды могут развивать согласованные наборы адаптаций. Здесь эволюция одного вида вызывает адаптацию второго вида. Эти изменения у второго вида, в свою очередь, вызывают новые адаптации у первого вида. Этот цикл отбора и реагирования называется коэволюцией. [192] Примером может служить выработка тетродотоксина у грубокожего тритона и эволюция устойчивости к тетродотоксину у его хищника, обыкновенной подвязочной змеи . В этой паре хищник-жертва эволюционная гонка вооружений привела к высоким уровням токсинов у тритонов и, соответственно, к высоким уровням устойчивости к токсинам у змей. [193]
Сотрудничество
Не все коэволюционирующие взаимодействия между видами связаны с конфликтом. [194] Возникло множество случаев взаимовыгодного взаимодействия. Например, между растениями и микоризными грибами, которые растут на их корнях и помогают растению поглощать питательные вещества из почвы, существует тесная связь. [195] Это взаимные отношения, поскольку растения снабжают грибы сахарами в результате фотосинтеза . Здесь грибы фактически растут внутри растительных клеток, позволяя им обмениваться питательными веществами со своими хозяевами, одновременно посылая сигналы , подавляющие иммунную систему растений . [196]
Возникли также коалиции между организмами одного вида. Крайним случаем является эусоциальность , обнаруженная у социальных насекомых, таких как пчелы , термиты и муравьи , когда стерильные насекомые кормят и охраняют небольшое количество организмов в колонии , способных к размножению. В еще меньшем масштабе соматические клетки, составляющие тело животного, ограничивают его размножение, чтобы они могли поддерживать стабильный организм, который затем поддерживает небольшое количество зародышевых клеток животного для производства потомства. Здесь соматические клетки реагируют на определенные сигналы, которые указывают им, расти ли, оставаться такими, какие они есть, или умереть. Если клетки игнорируют эти сигналы и размножаются ненадлежащим образом, их неконтролируемый рост вызывает рак . [197]
Такое сотрудничество внутри видов, возможно, развилось в процессе родственного отбора , при котором один организм действует, помогая вырастить потомство родственника. [198] Эта деятельность выбрана потому, что если помогающий человек содержит аллели, которые способствуют помогающей деятельности, вполне вероятно, что его родственники также будут содержать эти аллели и, таким образом, эти аллели будут передаваться дальше. [199] Другие процессы, которые могут способствовать сотрудничеству, включают групповой отбор, когда сотрудничество приносит пользу группе организмов. [200]
Видообразование

Видообразование — это процесс, при котором вид разделяется на два или более вида-потомка. [201]
Существует несколько способов определения понятия «вид». Выбор определения зависит от особенностей рассматриваемого вида. [202] Например, концепции некоторых видов с большей готовностью применимы к организмам, размножающимся половым путем, тогда как другие лучше подходят к бесполым организмам. Несмотря на разнообразие концепций различных видов, эти различные концепции могут быть помещены в один из трех широких философских подходов: скрещивания, экологического и филогенетического. [203] Концепция биологических видов (BSC) является классическим примером подхода скрещивания. Согласно определению биолога-эволюциониста Эрнста Майра в 1942 году, BSC утверждает, что «виды — это группы фактически или потенциально скрещивающихся природных популяций, которые репродуктивно изолированы от других таких групп». [204] Несмотря на широкое и долгосрочное использование, BSC, как и концепции других видов, не лишен противоречий, например, потому, что генетическая рекомбинация среди прокариот не является неотъемлемым аспектом воспроизводства; [205] это называется проблемой видов . [202] Некоторые исследователи пытались дать единое монистическое определение вида, в то время как другие придерживаются плюралистического подхода и предполагают, что могут быть разные способы логической интерпретации определения вида. [202] [203]
Барьеры для воспроизводства между двумя популяциями разного пола необходимы для того, чтобы популяции стали новыми видами. Поток генов может замедлить этот процесс, распространяя новые генетические варианты и на другие популяции. В зависимости от того, насколько далеко разошлись два вида с момента их последнего общего предка , они все еще могут производить потомство, как это происходит в случае спаривания лошадей и ослов для получения мулов . [206] Такие гибриды, как правило, бесплодны . В этом случае близкородственные виды могут регулярно скрещиваться, но гибриды будут отбираться, а виды останутся отдельными. Однако время от времени образуются жизнеспособные гибриды, и эти новые виды могут либо иметь промежуточные свойства между родительскими видами, либо обладать совершенно новым фенотипом. [207] Важность гибридизации для создания новых видов животных неясна, хотя случаи наблюдались у многих типов животных. [208] серая древесная лягушка . особенно хорошо изученным примером является [209]
Видообразование наблюдалось неоднократно как в контролируемых лабораторных условиях , так и в природе. [210] У организмов, размножающихся половым путем, видообразование является результатом репродуктивной изоляции, за которой следует генеалогическая дивергенция. Существует четыре основных географических способа видообразования. Наиболее распространенным у животных является аллопатрическое видообразование , которое происходит в популяциях, изначально изолированных географически, например, в результате фрагментации среды обитания или миграции. Отбор в этих условиях может привести к очень быстрым изменениям внешнего вида и поведения организмов. [211] [212] Поскольку отбор и дрейф действуют независимо на популяции, изолированные от остальных видов, разделение может в конечном итоге привести к появлению организмов, которые не могут скрещиваться. [213]
Второй способ видообразования — перипатрическое видообразование , которое происходит, когда небольшие популяции организмов изолируются в новой среде. Это отличается от аллопатрического видообразования тем, что изолированные популяции численно намного меньше родительской популяции. Здесь эффект основателя вызывает быстрое видообразование после того, как увеличение инбридинга увеличивает отбор гомозигот, что приводит к быстрым генетическим изменениям. [214]
Третий способ — парапатрическое видообразование . Это похоже на перипатрическое видообразование тем, что небольшая популяция попадает в новую среду обитания, но отличается тем, что между этими двумя популяциями нет физического разделения. Вместо этого видообразование является результатом эволюции механизмов, которые уменьшают поток генов между двумя популяциями. [201] Обычно это происходит, когда произошло резкое изменение окружающей среды в среде обитания родительского вида. Одним из примеров является трава Anthoxanthum odoratum , которая может подвергаться парапатрическому видообразованию в ответ на локальное загрязнение металлами из шахт. [215] Здесь развиваются растения, обладающие устойчивостью к высоким уровням металлов в почве. Отбор против скрещивания с металлочувствительной родительской популяцией привел к постепенному изменению времени цветения металлоустойчивых растений, что в конечном итоге привело к полной репродуктивной изоляции. Отбор против гибридов между двумя популяциями может вызвать усиление , то есть эволюцию признаков, способствующих спариванию внутри вида, а также смещение признаков , когда два вида становятся более разными по внешнему виду. [216]

Наконец, при симпатрическом видообразовании виды расходятся без географической изоляции или изменения среды обитания. Эта форма встречается редко, поскольку даже небольшой поток генов может устранить генетические различия между частями популяции. [217] Как правило, симпатрическое видообразование у животных требует эволюции как генетических различий , так и неслучайного спаривания, чтобы обеспечить развитие репродуктивной изоляции. [218]
Один тип симпатрического видообразования включает скрещивание двух родственных видов для получения нового гибридного вида. Это не характерно для животных, поскольку гибриды животных обычно стерильны. Это связано с тем, что во время мейоза каждого гомологичные хромосомы родителя принадлежат разным видам и не могут успешно спариваться. Однако это чаще встречается у растений, поскольку растения часто удваивают количество хромосом, образуя полиплоиды . [219] Это позволяет хромосомам каждого родительского вида образовывать совпадающие пары во время мейоза, поскольку хромосомы каждого родителя уже представлены парой. [220] Примером такого видообразования является скрещивание видов растений Arabidopsis thaliana и Arabidopsis arenosa с образованием нового вида Arabidopsis suecica . [221] Это произошло около 20 000 лет назад. [222] и процесс видообразования был повторен в лаборатории, что позволяет изучить генетические механизмы, участвующие в этом процессе. [223] Действительно, удвоение хромосом внутри вида может быть распространенной причиной репродуктивной изоляции, поскольку половина удвоенных хромосом не будет иметь совпадений при скрещивании с неудвоенными организмами. [224]
События видообразования играют важную роль в теории прерывистого равновесия , которая объясняет в летописи окаменелостей короткие «всплески» эволюции, перемежающиеся относительно длительными периодами застоя, когда виды остаются относительно неизменными. [225] В этой теории видообразование и быстрая эволюция связаны, причем естественный отбор и генетический дрейф наиболее сильно действуют на организмы, претерпевающие видообразование в новых средах обитания или небольших популяциях. В результате периоды застоя в летописи окаменелостей соответствуют родительской популяции, а организмы, претерпевающие видообразование и быструю эволюцию, обнаруживаются в небольших популяциях или географически ограниченных средах обитания и поэтому редко сохраняются в виде окаменелостей. [139]
Вымирание

Вымирание – это исчезновение целого вида. Вымирание не является чем-то необычным, поскольку виды регулярно появляются в результате видообразования и исчезают в результате вымирания. [226] Почти все виды животных и растений, жившие на Земле, в настоящее время вымерли. [227] и вымирание, похоже, является окончательной судьбой всех видов. [228] Эти вымирания происходили постоянно на протяжении всей истории жизни, хотя время от времени темпы вымирания резко возрастают . [229] Событие мел-палеогенового вымирания , во время которого вымерли нептичьи динозавры, является наиболее известным, но более раннее пермско-триасовое вымирание было еще более серьезным: около 96% всех морских видов были уничтожены. [229] Голоценовое вымирание — это продолжающееся массовое вымирание, связанное с распространением человечества по всему земному шару в течение последних нескольких тысяч лет. Современные темпы вымирания в 100–1000 раз превышают фоновые темпы, и до 30% нынешних видов могут исчезнуть к середине 21 века. [230] Деятельность человека в настоящее время является основной причиной продолжающегося вымирания; [231] [232] глобальное потепление может еще больше ускорить его в будущем. [233] Несмотря на предполагаемое вымирание более 99% всех видов, когда-либо живших на Земле, [234] [235] По оценкам, в настоящее время на Земле обитает около 1 триллиона видов, из которых описана лишь одна тысячная 1%. [236]
Роль вымирания в эволюции не очень хорошо изучена и может зависеть от того, какой тип вымирания рассматривается. [229] Причины непрерывных событий «низкого уровня» вымирания, которые составляют большинство вымираний, могут быть результатом конкуренции между видами за ограниченные ресурсы ( принцип конкурентного исключения ). [237] Если один вид сможет превзойти другой, это может привести к отбору видов, при котором более приспособленные виды выживут, а другие виды будут вымирать. [84] Периодические массовые вымирания также важны, но вместо того, чтобы действовать как селективная сила, они резко сокращают разнообразие неспецифическим образом и способствуют всплескам быстрой эволюции и видообразования у выживших. [238]
Приложения
Концепции и модели, используемые в эволюционной биологии, такие как естественный отбор, имеют множество применений. [239]
Искусственный отбор — это преднамеренный отбор признаков в популяции организмов. Это использовалось в течение тысяч лет при одомашнивании растений и животных. [240] Совсем недавно такой отбор стал жизненно важной частью генной инженерии , при этом селектируемые маркеры, такие как гены устойчивости к антибиотикам, используются для манипулирования ДНК. Белки с ценными свойствами возникли в результате повторяющихся раундов мутаций и отбора (например, модифицированные ферменты и новые антитела ) в процессе, называемом направленной эволюцией . [241]
Понимание изменений, произошедших в ходе эволюции организма, может выявить гены, необходимые для построения частей тела, гены, которые могут быть вовлечены в генетические нарушения человека . [242] Например, мексиканская тетра — пещерная рыба- альбинос , потерявшая зрение в ходе эволюции. Скрещивание разных популяций этой слепой рыбы дало потомство с функциональными глазами, поскольку в изолированных популяциях, которые развивались в разных пещерах, произошли разные мутации. [243] Это помогло идентифицировать гены, необходимые для зрения и пигментации. [244]
Эволюционная теория имеет множество применений в медицине . Многие болезни человека не являются статическими явлениями, а способны к эволюции. Вирусы, бактерии, грибки и раковые клетки развиваются и становятся устойчивыми к иммунной защите хозяина, а также к фармацевтическим препаратам . [245] [246] [247] Те же самые проблемы возникают в сельском хозяйстве, где используются пестициды. [248] и гербицид [249] сопротивление. Вполне возможно, что мы приближаемся к концу эффективного срока службы большинства доступных антибиотиков. [250] и предсказание эволюции и эволюционируемости [251] наших патогенов и разработка стратегий, чтобы замедлить или обойти их, требуют более глубокого знания сложных сил, движущих эволюцию на молекулярном уровне. [252]
В информатике моделирование эволюции с использованием эволюционных алгоритмов и искусственной жизни началось в 1960-х годах и было расширено моделированием искусственного отбора. [253] Искусственная эволюция стала широко признанным методом оптимизации благодаря работе Инго Рехенберга в 1960-х годах. Он использовал стратегии эволюции для решения сложных инженерных задач. [254] Генетические алгоритмы , в частности, стали популярными благодаря трудам Джона Генри Холланда . [255] Практическое применение также включает автоматическую эволюцию компьютерных программ . [256] Эволюционные алгоритмы теперь используются для более эффективного решения многомерных проблем, чем программное обеспечение, созданное людьми-разработчиками, а также для оптимизации проектирования систем. [257]
Эволюционная история жизни
−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||||||||||||||||||||
Происхождение жизни
Земле около 4,54 миллиарда лет . [258] [259] [260] Самые ранние неоспоримые свидетельства существования жизни на Земле датируются по меньшей мере 3,5 миллиарда лет назад. [12] [261] во время Эоархейской эры после того, как геологическая кора начала затвердевать после более раннего расплавленного Гадейского эона. Окаменелости микробного мата были обнаружены в песчанике возрастом 3,48 миллиарда лет в Западной Австралии. [14] [15] [16] возрастом 3,7 миллиарда лет, Другим ранним физическим свидетельством биогенного вещества является графит в метаосадочных породах обнаруженных в Западной Гренландии. [13] а также «остатки биотической жизни », обнаруженные в скалах возрастом 4,1 миллиарда лет в Западной Австралии. [262] [263] Комментируя австралийские находки, Стивен Блер Хеджес написал: «Если бы жизнь возникла на Земле сравнительно быстро, то она могла бы быть обычным явлением во Вселенной». [262] [264] В июле 2016 года ученые сообщили об идентификации набора из 355 генов последнего универсального общего предка (LUCA) всех организмов, живущих на Земле. [265]
Более 99% всех видов, а это более пяти миллиардов видов, [266] виды, когда-либо жившие на Земле, считаются вымершими. [234] [235] По оценкам, численность современных видов на Земле колеблется от 10 до 14 миллионов. [267] [268] из которых, по оценкам, около 1,9 миллиона были названы [269] и 1,6 миллиона зарегистрировано на сегодняшний день в центральной базе данных, [270] оставляя по крайней мере 80% еще не описанными.
Считается, что высокоэнергетическая химия создала самовоспроизводящуюся молекулу около 4 миллиардов лет назад, а полмиллиарда лет спустя существовал последний общий предок всей жизни. [10] В настоящее время научный консенсус заключается в том, что сложная биохимия, из которой состоит жизнь, возникла в результате более простых химических реакций. [271] [272] Начало жизни могло включать в себя самовоспроизводящиеся молекулы, такие как РНК. [273] и сборка простых клеток. [274]
Общее происхождение
Все организмы на Земле произошли от общего предка или генофонда предков . [168] [275] Современные виды представляют собой этап процесса эволюции, а их разнообразие является продуктом длинной серии событий видообразования и вымирания. [276] Общее происхождение организмов было впервые выведено из четырех простых фактов об организмах: во-первых, они имеют географическое распространение, которое нельзя объяснить местной адаптацией. Во-вторых, разнообразие жизни – это не набор совершенно уникальных организмов, а организмы, имеющие морфологическое сходство. В-третьих, рудиментарные черты без четкой цели напоминают функциональные наследственные черты. В-четвертых, на основе этих сходств организмы можно классифицировать в иерархию вложенных групп, подобную генеалогическому древу. [277]
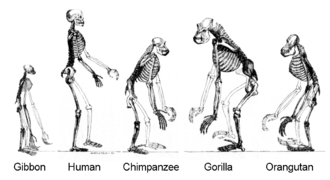
Из-за горизонтального переноса генов это «дерево жизни» может быть более сложным, чем простое ветвящееся дерево, поскольку некоторые гены распространились независимо между отдаленно родственными видами. [278] [279] Чтобы решить эту и другие проблемы, некоторые авторы предпочитают использовать « Коралл жизни » как метафору или математическую модель, иллюстрирующую эволюцию жизни. Эта точка зрения восходит к идее, кратко упомянутой Дарвином, но позже оставленной. [280]
Предыдущие виды также оставили записи своей эволюционной истории. Окаменелости, наряду со сравнительной анатомией современных организмов, составляют морфологическую или анатомическую летопись. [281] Сравнивая анатомию современных и вымерших видов, палеонтологи могут сделать вывод о происхождении этих видов. Однако этот подход наиболее успешен для организмов, у которых были твердые части тела, такие как раковины, кости или зубы. Кроме того, поскольку прокариоты, такие как бактерии и археи, имеют ограниченный набор общих морфологий, их окаменелости не предоставляют информации об их происхождении.
Совсем недавно доказательства общего происхождения были получены в результате изучения биохимического сходства между организмами. Например, все живые клетки используют один и тот же базовый набор нуклеотидов и аминокислот . [282] Развитие молекулярной генетики выявило записи эволюции, оставшиеся в геномах организмов: датирование момента, когда виды разошлись по молекулярным часам , вызванным мутациями. [283] Например, эти сравнения последовательностей ДНК показали, что люди и шимпанзе имеют общие геномы на 98%, а анализ тех немногих областей, где они различаются, помогает пролить свет на то, когда существовал общий предок этих видов. [284]
Эволюция жизни

Прокариоты населяли Землю примерно 3–4 миллиарда лет назад. [286] [287] Никаких очевидных изменений в морфологии или клеточной организации у этих организмов в течение следующих нескольких миллиардов лет не произошло. [288] Эукариотические клетки возникли между 1,6 и 2,7 миллиарда лет назад. Следующее серьезное изменение в клеточной структуре произошло, когда бактерии были поглощены эукариотическими клетками, образовав кооперативную ассоциацию, называемую эндосимбиозом . [289] [290] Поглощенные бактерии и клетка-хозяин затем подверглись совместной эволюции, при этом бактерии эволюционировали либо в митохондрии, либо в гидрогеносомы . [291] Очередное поглощение цианобактериоподобных организмов привело к образованию хлоропластов у водорослей и растений. [292]
История жизни была историей одноклеточных период в океанах начали появляться многоклеточные организмы эукариот, прокариотов и архей примерно до 610 миллионов лет назад, когда в эдиакарский . [286] [293] Эволюция многоклеточности происходила в ходе множества независимых событий в таких разнообразных организмах, как губки , бурые водоросли , цианобактерии, слизевики и миксобактерии . [294] В январе 2016 года ученые сообщили, что около 800 миллионов лет назад незначительное генетическое изменение в одной молекуле под названием GK-PID, возможно, позволило организмам перейти от одноклеточного организма к одной из многих клеток. [295]
Вскоре после появления этих первых многоклеточных организмов в течение примерно 10 миллионов лет появилось значительное биологическое разнообразие в результате события, названного Кембрийским взрывом . Здесь в палеонтологической летописи появилось большинство типов современных животных, а также уникальные линии, впоследствии вымершие. [296] Были предложены различные причины кембрийского взрыва, в том числе накопление кислорода в атмосфере в результате фотосинтеза. [297]
Около 500 миллионов лет назад растения и грибы колонизировали землю, а вскоре за ними последовали членистоногие и другие животные. [298] Насекомые добились особенного успеха и даже сегодня составляют большинство видов животных. [299] Земноводные впервые появились около 364 миллионов лет назад, за ними последовали ранние амниоты и птицы около 155 миллионов лет назад (обе из линий, подобных «рептилиям»), млекопитающие около 129 миллионов лет назад, гоминины около 10 миллионов лет назад и современные люди около 250 000 лет назад. назад. [300] [301] [302] Однако, несмотря на эволюцию этих крупных животных, более мелкие организмы, подобные тем типам, которые развились на ранних этапах этого процесса, продолжают быть весьма успешными и доминировать на Земле, причем большая часть как биомассы, так и видов является прокариотами. [146]
История эволюционной мысли




Классическая античность
Предположение о том, что один тип организма может произойти от другого типа, восходит к некоторым из первых греческих философов -досократиков , таких как Анаксимандр и Эмпедокл . [304] Подобные предложения сохранились и во времена Римской империи. Поэт и философ Лукреций последовал за Эмпедоклом в своем шедевре De rerum natura ( букв. « О природе вещей » ). [305] [306]
Средний возраст
В отличие от этих материалистических взглядов, аристотелизм рассматривал все естественные вещи как реализации фиксированных природных возможностей, известных как формы . [307] [308] Это стало частью средневекового телеологического понимания природы , в котором все вещи играют определенную роль в божественном космическом порядке. Вариации этой идеи стали стандартным пониманием средневековья и были интегрированы в христианское учение, но Аристотель не требовал, чтобы реальные типы организмов всегда соответствовали один к одному точным метафизическим формам, а конкретно приводил примеры того, как новые типы живых существ все может случиться. [309]
Ряд арабских мусульманских ученых писали об эволюции, в первую очередь Ибн Халдун , написавший книгу «Мукаддима» в 1377 году нашей эры, в которой он утверждал, что люди развились из «мира обезьян» в процессе, посредством которого «виды становятся более многочисленными». ". [310]
Додарвинистский
« Новая наука» 17 века отвергла аристотелевский подход. Он стремился объяснить явления природы с помощью физических законов , которые были одинаковыми для всех видимых вещей и не требовали существования каких-либо фиксированных природных категорий или божественного космического порядка. Однако этот новый подход медленно укоренялся в биологических науках: последнем бастионе концепции фиксированных природных типов. Джон Рэй применил один из ранее использовавшихся более общих терминов для фиксированных природных типов, «видов», к типам растений и животных, но он строго определил каждый тип живого существа как вид и предположил, что каждый вид можно определить по признакам, которые увековечили его. себя поколение за поколением. [311] введенная Биологическая классификация, Карлом Линнеем в 1735 году, явно признавала иерархическую природу взаимоотношений видов, но все же рассматривала виды как фиксированные в соответствии с божественным замыслом. [312]
Другие натуралисты того времени размышляли об эволюционном изменении видов с течением времени в соответствии с законами природы. В 1751 году Пьер Луи Мопертюи писал о естественных изменениях, происходящих во время размножения и накапливающихся на протяжении многих поколений, образуя новые виды. [313] Жорж-Луи Леклерк, граф де Бюффон , предположил, что виды могут деградировать в разные организмы, а Эразм Дарвин предположил, что все теплокровные животные могли произойти от одного микроорганизма (или «нити»). [314] Первой полноценной эволюционной схемой была теория «трансмутации» Жана-Батиста Ламарка 1809 года. [315] который предполагал спонтанное зарождение , постоянно производя простые формы жизни, которые развивались с большей сложностью в параллельных линиях с присущей им прогрессивной тенденцией, и постулировал, что на локальном уровне эти линии адаптируются к окружающей среде, наследуя изменения, вызванные их использованием или неиспользованием у родителей. [316] (Последний процесс позже был назван ламаркизмом .) [316] [317] [318] Эти идеи были осуждены авторитетными натуралистами как предположения, не имеющие эмпирической поддержки. В частности, Жорж Кювье настаивал на том, что виды не связаны между собой и фиксированы, а их сходство отражает божественный замысел функциональных потребностей. Тем временем идеи Рэя о доброжелательном замысле были развиты Уильямом Пейли в книге « Естественная теология или доказательства существования и атрибутов божества» (1802 г.), в которой предлагались сложные адаптации в качестве доказательства божественного замысла и которой восхищался Чарльз Дарвин. [319] [320]
Дарвиновская революция
Решающий отход от концепции постоянных типологических классов или типов в биологии произошел с теорией эволюции посредством естественного отбора, которая была сформулирована Чарльзом Дарвином и Альфредом Уоллесом в терминах переменных популяций. Дарвин использовал выражение « происхождение с модификациями », а не «эволюция». [321] Частично под влиянием «Опыта о принципе народонаселения » (1798) Томаса Роберта Мальтуса , Дарвин отмечал, что рост населения приведет к «борьбе за существование», в которой преобладают благоприятные вариации, а другие погибают. В каждом поколении многим потомкам не удается дожить до репродуктивного возраста из-за ограниченности ресурсов. Это могло бы объяснить разнообразие растений и животных, имеющих общее происхождение, благодаря действию законов природы одинаково для всех типов организмов. [322] [323] [324] [325] Дарвин разработал свою теорию «естественного отбора», начиная с 1838 года, и писал свою «большую книгу» на эту тему, когда Альфред Рассел Уоллес прислал ему версию практически той же теории в 1858 году. Их отдельные статьи были представлены вместе на встрече 1858 года. Линнеевского общества в Лондоне . [326] В конце 1859 года Дарвин опубликовал свой «реферат» под названием «Происхождение видов » , объясняющий естественный отбор подробно и таким образом, что это привело к все более широкому признанию дарвиновских концепций эволюции за счет альтернативных теорий . Томас Генри Хаксли применил идеи Дарвина к людям, используя палеонтологию и сравнительную анатомию, чтобы предоставить убедительные доказательства того, что люди и обезьяны имели общее происхождение. Некоторых это встревожило, поскольку это подразумевало, что у людей нет особого места во Вселенной . [327]
Пангенезис и наследственность
Механизмы репродуктивной наследственности и происхождение новых признаков оставались загадкой. С этой целью Дарвин разработал свою предварительную теорию пангенезиса . [328] В 1865 году Грегор Мендель сообщил, что черты наследуются предсказуемым образом посредством независимого ассортимента и разделения элементов (позже известных как гены). Законы наследственности Менделя в конечном итоге вытеснили большую часть теории пангенезиса Дарвина. [329] Август Вейсман провел важное различие между половыми клетками , дающими начало гаметам (таким как сперматозоиды и яйцеклетки ), и соматическими клетками организма, продемонстрировав, что наследственность передается только через зародышевую линию. Хьюго де Врис соединил теорию пангенезиса Дарвина с различием зародышевых и соматических клеток Вейсмана и предположил, что дарвиновские пангены концентрируются в ядре клетки и при экспрессии могут перемещаться в цитоплазму, изменяя структуру клетки . Де Врис также был одним из исследователей, которые сделали работу Менделя широко известной, полагая, что менделевские черты соответствуют передаче наследственных вариаций по зародышевой линии. [330] Чтобы объяснить, как возникают новые варианты, де Врис разработал теорию мутаций , которая привела к временному расколу между теми, кто признавал дарвиновскую эволюцию, и биометристами, которые объединились с де Фрисом. [331] [332] В 1930-х годах пионеры в области популяционной генетики , такие как Рональд Фишер , Сьюэлл Райт и Дж.Б.С. Холдейн, заложили основы эволюции на надежную статистическую философию. Таким образом, было устранено ложное противоречие между теорией Дарвина, генетическими мутациями и менделевской наследственностью . [333]
«Современный синтез»
В 1920-х и 1930-х годах современный синтез объединил естественный отбор и популяционную генетику, основанную на менделевском наследовании, в единую теорию, которая включала случайный генетический дрейф, мутации и поток генов. Эта новая версия эволюционной теории сосредоточена на изменениях частот аллелей в популяции. Это объяснило закономерности, наблюдаемые у разных видов в популяциях, через переходы окаменелостей в палеонтологии. [333]
Дальнейшие синтезы
С тех пор дальнейшие синтезы расширили объяснительную силу эволюции в свете многочисленных открытий, охватив биологические явления по всей биологической иерархии от генов до популяций. [334]
Публикация структуры ДНК Джеймсом Уотсоном и Фрэнсисом Криком при участии Розалинды Франклин в 1953 году продемонстрировала физический механизм наследования. [335] Молекулярная биология улучшила понимание взаимосвязи между генотипом и фенотипом . Достижения были также достигнуты в филогенетической систематике , отображающей переход признаков в сравнительную и проверяемую структуру посредством публикации и использования эволюционных деревьев . [336] В 1973 году биолог-эволюционист Теодосий Добжанский написал, что « ничто в биологии не имеет смысла, кроме как в свете эволюции », поскольку она вывела на свет отношения того, что сначала казалось разрозненными фактами естественной истории, в связный объяснительный массив знаний, который описывает и предсказывает множество наблюдаемых фактов о жизни на этой планете. [337]
Одно из расширений, известное как эволюционная биология развития и неофициально называемое «эво-дево», подчеркивает, как изменения между поколениями (эволюция) влияют на закономерности изменений внутри отдельных организмов ( развитие ). [237] [338] С начала 21-го века некоторые биологи выступали за расширенный эволюционный синтез , который бы учитывал эффекты негенетических способов наследования, таких как эпигенетика , родительские эффекты , экологическое наследование и культурное наследование , а также эволюционность . [339] [340]
Социальные и культурные реакции

В 19 веке, особенно после публикации « Происхождения видов» в 1859 году, идея о том, что жизнь возникла, была активным источником академических дебатов, сосредоточенных на философских, социальных и религиозных последствиях эволюции. Сегодня современный эволюционный синтез принимается подавляющим большинством ученых. [237] Однако эволюция остается спорной концепцией для некоторых теистов . [342]
В то время как различные религии и конфессии примирили свои убеждения с эволюцией посредством таких концепций, как теистическая эволюция , есть креационисты , которые считают, что эволюция противоречит мифам о сотворении мира, присутствующим в их религиях, и которые выдвигают различные возражения против эволюции . [135] [343] [344] Как было продемонстрировано ответами на публикацию « Остатков естественной истории творения» биологии является вывод о том в 1844 году, наиболее спорным аспектом эволюционной , что люди имеют общее происхождение с обезьянами и что умственные и моральные способности человечества изменились. те же самые типы естественных причин, что и другие наследственные признаки у животных. [345] В некоторых странах, особенно в Соединенных Штатах, эта напряженность между наукой и религией подпитывает нынешние противоречия между сотворением и эволюцией, религиозный конфликт, сосредоточенный на политике и государственном образовании . [346] В то время как другие научные области, такие как космология [347] и наука о Земле [348] Также вступая в противоречие с буквальными интерпретациями многих религиозных текстов , эволюционная биология испытывает значительно большее сопротивление со стороны религиозных буквалистов.
Преподавание эволюции на уроках биологии в американской средней школе было редкостью на протяжении большей части первой половины 20-го века. Решение по делу Скоупса 1925 года привело к тому, что этот предмет стал очень редким в американских учебниках средней школы по биологии на протяжении целого поколения, но позже он был постепенно вновь введен и стал юридически защищен решением 1968 года по делу Эпперсон против Арканзаса . С тех пор конкурирующая религиозная вера креационизма была юридически запрещена в учебных программах средней школы различными решениями в 1970-х и 1980-х годах, но она вернулась в псевдонаучной форме как разумный замысел (ID), чтобы быть снова исключенной в деле Китцмиллер против Дувра 2005 года. Дело школьного округа . [349] Дебаты по поводу идей Дарвина не вызвали серьезных разногласий в Китае. [350]
См. также
- Деволюция (биология) - представление о том, что виды могут вернуться к примитивным формам.
Ссылки
- ^ Холл и Халлгримссон 2008 , стр. 4–6
- ^ «Ресурсы эволюции» . Вашингтон, округ Колумбия: Национальные академии наук, техники и медицины . 2016. Архивировано из оригинала 3 июня 2016 года.
- ^ Перейти обратно: а б Скотт-Филлипс, Томас К.; Лаланд, Кевин Н .; Шукер, Дэвид М.; и др. (май 2014 г.). «Перспектива строительства ниши: критическая оценка» . Эволюция . 68 (5): 1231–1243. дои : 10.1111/evo.12332 . ISSN 0014-3820 . ПМК 4261998 . ПМИД 24325256 .
Эволюционные процессы обычно рассматриваются как процессы, посредством которых происходят эти изменения. Широко признаны четыре таких процесса: естественный отбор (в широком смысле, включая половой отбор), генетический дрейф, мутация и миграция (Fisher 1930; Haldane 1932). Последние два порождают вариации; первые два сортируют его.
- ^ Холл и Халлгримссон 2008 , стр. 3–5
- ^ Voet, Voet & Pratt 2016 , стр. 1–22, Глава 1: Введение в химию жизни.
- ^ Дарвин 1859 г.
- ^ Перейти обратно: а б Левонтин, Ричард К. (ноябрь 1970 г.). «Единицы выбора» (PDF) . Ежегодный обзор экологии и систематики . 1 :1–18. doi : 10.1146/annurev.es.01.110170.000245 . ISSN 0066-4162 . JSTOR 2096764 . S2CID 84684420 . Архивировано (PDF) из оригинала 6 февраля 2015 года.
- ^ Futuyma & Kirkpatrick 2017 , стр. 3–26, Глава 1: Эволюционная биология
- ^ Кампуракис 2014 , стр. 127–129
- ^ Перейти обратно: а б Дулитл, В. Форд (февраль 2000 г.). «Искоренение Древа Жизни» (PDF) . Научный американец . 282 (2): 90–95. Бибкод : 2000SciAm.282b..90D . doi : 10.1038/scientificamerican0200-90 . ISSN 0036-8733 . ПМИД 10710791 . Архивировано из оригинала (PDF) 7 сентября 2006 года . Проверено 5 апреля 2015 г.
- ^ Глансдорф, Николас; Ин Сюй; Лабедан, Бернар (9 июля 2008 г.). «Последний универсальный общий предок: появление, конституция и генетическое наследие неуловимого предшественника» . Биология Директ . 3:29 . дои : 10.1186/1745-6150-3-29 . ISSN 1745-6150 . ПМЦ 2478661 . ПМИД 18613974 .
- ^ Перейти обратно: а б Шопф, Дж. Уильям ; Кудрявцев Анатолий Б.; Чая, Эндрю Д.; Трипати, Абхишек Б. (5 октября 2007 г.). «Свидетельства архейской жизни: строматолиты и микроокаменелости». Докембрийские исследования . 158 (3–4): 141–155. Бибкод : 2007PreR..158..141S . doi : 10.1016/j.precamres.2007.04.009 . ISSN 0301-9268 .
- ^ Перейти обратно: а б Отомо, Йоко; Какегава, Такеши; Исида, Акизуми; и др. (январь 2014 г.). «Доказательства наличия биогенного графита в метаосадочных породах раннего архейского периода Иисуса». Природа Геонауки . 7 (1): 25–28. Бибкод : 2014NatGe...7... 25O дои : 10.1038/ngeo2025 . ISSN 1752-0894 .
- ^ Перейти обратно: а б Боренштейн, Сет (13 ноября 2013 г.). «Найдена самая старая окаменелость: познакомьтесь со своей микробной мамой» . Возбуждайте . Йонкерс, Нью-Йорк: Интерактивная сеть Mindspark . Ассошиэйтед Пресс . Архивировано из оригинала 29 июня 2015 года . Проверено 31 мая 2015 г.
- ^ Перейти обратно: а б Перлман, Джонатан (13 ноября 2013 г.). «Обнаружены древнейшие признаки жизни на Земле» . «Дейли телеграф» . Лондон. Архивировано из оригинала 16 декабря 2014 года . Проверено 15 декабря 2014 г.
- ^ Перейти обратно: а б Ноффке, Нора ; Кристиан, Дэниел; Уэйси, Дэвид; Хейзен, Роберт М. (16 ноября 2013 г.). «Микробно-индуцированные осадочные структуры, фиксирующие древнюю экосистему формации Дрессер возрастом около 3,48 миллиардов лет, Пилбара, Западная Австралия» . Астробиология . 13 (12): 1103–1124. Бибкод : 2013AsBio..13.1103N . дои : 10.1089/ast.2013.1030 . ISSN 1531-1074 . ПМК 3870916 . ПМИД 24205812 .
- ^ Футуйма 2004 , с. 33
- ^ Ткань 2005 , стр. хв-16
- ^ НАН 2008 , с. 17. Архивировано 30 июня 2015 г. в Wayback Machine.
- ^ Футуйма, Дуглас Дж. , изд. (1999). «Эволюция, наука и общество: эволюционная биология и национальная программа исследований» (PDF) (резюме). Нью-Брансуик, Нью-Джерси: Отдел университетских публикаций, Рутгерс, Государственный университет Нью-Джерси . OCLC 43422991 . Архивировано из оригинала (PDF) 31 января 2012 года . Проверено 24 ноября 2014 г.
- ^ Штурм, Ричард А.; Фрудакис, Тони Н. (август 2004 г.). «Цвет глаз: порталы в гены пигментации и происхождение». Тенденции в генетике . 20 (8): 327–332. дои : 10.1016/j.tig.2004.06.010 . ISSN 0168-9525 . ПМИД 15262401 .
- ^ Перейти обратно: а б Пирсон, Хелен (25 мая 2006 г.). «Генетика: что такое ген?» . Природа . 441 (7092): 398–401. Бибкод : 2006Natur.441..398P . дои : 10.1038/441398a . ISSN 0028-0836 . ПМИД 16724031 . S2CID 4420674 .
- ^ Вишер, Питер М.; Хилл, Уильям Г .; Рэй, Наоми Р. (апрель 2008 г.). «Наследственность в эпоху геномики — концепции и заблуждения». Обзоры природы Генетика . 9 (4): 255–266. дои : 10.1038/nrg2322 . ISSN 1471-0056 . ПМИД 18319743 . S2CID 690431 .
- ^ Эттинг, Уильям С.; Бриллиант, Мюррей Х.; Кинг, Ричард А. (август 1996 г.). «Клинический спектр альбинизма у человека». Молекулярная медицина сегодня . 2 (8): 330–335. дои : 10.1016/1357-4310(96)81798-9 . ISSN 1357-4310 . ПМИД 8796918 .
- ^ Перейти обратно: а б Футуйма 2005 г. [ нужна страница ]
- ^ Филлипс, Патрик К. (ноябрь 2008 г.). «Эпистаз — существенная роль взаимодействий генов в структуре и эволюции генетических систем» . Обзоры природы Генетика . 9 (11): 855–867. дои : 10.1038/nrg2452 . ISSN 1471-0056 . ПМК 2689140 . ПМИД 18852697 .
- ^ Перейти обратно: а б Жунлин Ву; Мин Линь (март 2006 г.). «Функциональное картирование - как составить карту и изучить генетическую архитектуру динамических сложных признаков». Обзоры природы Генетика . 7 (3): 229–237. дои : 10.1038/nrg1804 . ISSN 1471-0056 . ПМИД 16485021 . S2CID 24301815 .
- ^ Батлин, Роджер К.; Трегенза, Том (28 февраля 1998 г.). «Уровни генетического полиморфизма: маркерные локусы и количественные признаки» . Философские труды Королевского общества Б. 353 (1366): 187–198. дои : 10.1098/rstb.1998.0201 . ISSN 0962-8436 . ПМК 1692210 . ПМИД 9533123 .
- Батлин, Роджер К.; Трегенза, Том (29 декабря 2000 г.). «Поправка на Бутлина и Трегензу, Уровни генетического полиморфизма: маркерные локусы и количественные признаки» . Философские труды Королевского общества Б. 355 (1404): 1865. doi : 10.1098/rstb.2000.2000 . ISSN 0962-8436 .
Некоторые значения в таблице 1 на с. 193 были даны неверно. Ошибки не влияют на выводы, сделанные в статье. Исправленная таблица воспроизведена ниже.
- Батлин, Роджер К.; Трегенза, Том (29 декабря 2000 г.). «Поправка на Бутлина и Трегензу, Уровни генетического полиморфизма: маркерные локусы и количественные признаки» . Философские труды Королевского общества Б. 355 (1404): 1865. doi : 10.1098/rstb.2000.2000 . ISSN 0962-8436 .
- ^ Амос, Уильям; Харвуд, Джон (28 февраля 1998 г.). «Факторы, влияющие на уровень генетического разнообразия природных популяций» . Философские труды Королевского общества Б. 353 (1366): 177–186. дои : 10.1098/rstb.1998.0200 . ISSN 0962-8436 . ПМК 1692205 . ПМИД 9533122 .
- ^ Futuyma & Kirkpatrick 2017 , стр. 79–102, Глава 4: Мутация и вариация.
- ^ Кейтли, PD (2012). «Скорость и последствия новых мутаций у человека» . Генетика . 190 (2): 295–304. дои : 10.1534/genetics.111.134668 . ПМК 3276617 . ПМИД 22345605 .
- ^ Гастингс, П.Дж.; Лупски, Джеймс Р .; Розенберг, Сьюзен М.; Ира, Гжегож (август 2009 г.). «Механизмы изменения числа копий гена» . Обзоры природы Генетика . 10 (8): 551–564. дои : 10.1038/nrg2593 . ISSN 1471-0056 . ПМК 2864001 . ПМИД 19597530 .
- ^ Кэрролл, Гренье и Уэзерби, 2005 г. [ нужна страница ]
- ^ Харрисон, Пол М.; Герштейн, Марк (17 мая 2002 г.). «Изучение геномов на протяжении веков: семейства белков, псевдогены и эволюция протеомов». Журнал молекулярной биологии . 318 (5): 1155–1174. дои : 10.1016/S0022-2836(02)00109-2 . ISSN 0022-2836 . ПМИД 12083509 .
- ^ Боумейкер, Джеймс К. (май 1998 г.). «Эволюция цветового зрения у позвоночных» . Глаз . 12 (3б): 541–547. дои : 10.1038/eye.1998.143 . ISSN 0950-222X . ПМИД 9775215 . S2CID 12851209 .
- ^ Грегори, Т. Райан ; Хеберт, Пол Д.Н. (апрель 1999 г.). «Модуляция содержания ДНК: непосредственные причины и конечные последствия» . Геномные исследования . 9 (4): 317–324. дои : 10.1101/гр.9.4.317 . ISSN 1088-9051 . ПМИД 10207154 . S2CID 16791399 . Архивировано из оригинала 23 августа 2014 года . Проверено 11 декабря 2014 г.
- ^ Херлс, Мэтью (13 июля 2004 г.). «Дупликация генов: геномная торговля запасными частями» . ПЛОС Биология . 2 (7): е206. дои : 10.1371/journal.pbio.0020206 . ISSN 1545-7885 . ПМК 449868 . ПМИД 15252449 .
- ^ Лю, На; Окамура, Кацутомо; Тайлер, Дэвид М.; и др. (октябрь 2008 г.). «Эволюция и функциональное разнообразие генов микроРНК животных» . Клеточные исследования . 18 (10): 985–996. дои : 10.1038/cr.2008.278 . ISSN 1001-0602 . ПМК 2712117 . ПМИД 18711447 .
- ^ Сипель, Адам (октябрь 2009 г.). «Дарвиновская алхимия: гены человека из некодирующей ДНК» . Геномные исследования . 19 (10): 1693–1695. дои : 10.1101/гр.098376.109 . ISSN 1088-9051 . ПМЦ 2765273 . ПМИД 19797681 .
- ^ Оренго, Кристина А.; Торнтон, Джанет М. (июль 2005 г.). «Белковые семейства и их эволюция - структурная перспектива». Ежегодный обзор биохимии . 74 . Годовые обзоры : 867–900. doi : 10.1146/annurev.biochem.74.082803.133029 . ISSN 0066-4154 . ПМИД 15954844 . S2CID 7483470 .
- ^ Лонг, Манюань; Бетран, Эстер; Торнтон, Кевин; Ван, Вэнь (ноябрь 2003 г.). «Происхождение новых генов: взгляды молодых и старых». Обзоры природы Генетика . 4 (11): 865–875. дои : 10.1038/nrg1204 . ISSN 1471-0056 . ПМИД 14634634 . S2CID 33999892 .
- ^ Ван, Минглей; Каэтано-Аноллес, Густаво (14 января 2009 г.). «Эволюционная механика организации доменов в протеомах и рост модульности в белковом мире» . Структура . 17 (1): 66–78. дои : 10.1016/j.str.2008.11.008 . ISSN 1357-4310 . ПМИД 19141283 .
- ^ Вайсман, Кира Дж.; Мюллер, Рольф (14 апреля 2008 г.). «Белко-белковые взаимодействия в мультиферментных мегасинтетазах». ХимБиоХим . 9 (6): 826–848. дои : 10.1002/cbic.200700751 . ISSN 1439-4227 . ПМИД 18357594 . S2CID 205552778 .
- ^ Андерссон, Лейф (2020). «Мутации домашних животных, нарушающие или создающие структуру пигментации» . Границы экологии и эволюции . 8 . дои : 10.3389/fevo.2020.00116 . ISSN 2296-701X .
- ^ Раддинг, Чарльз М. (декабрь 1982 г.). «Гомологическое спаривание и обмен цепями при генетической рекомбинации». Ежегодный обзор генетики . 16 : 405–437. дои : 10.1146/annurev.ge.16.120182.002201 . ISSN 0066-4197 . ПМИД 6297377 .
- ^ Агравал, Анил Ф. (5 сентября 2006 г.). «Эволюция пола: почему организмы меняют свои генотипы?». Современная биология . 16 (17): Р696–Р704. Бибкод : 2006CBio...16.R696A . CiteSeerX 10.1.1.475.9645 . дои : 10.1016/j.cub.2006.07.063 . ISSN 0960-9822 . ПМИД 16950096 . S2CID 14739487 .
- ^ Питерс, Эндрю Д.; Отто, Сара П. (июнь 2003 г.). «Освобождение генетической изменчивости посредством пола». Биоэссе . 25 (6): 533–537. дои : 10.1002/bies.10291 . ISSN 0265-9247 . ПМИД 12766942 .
- ^ Годдард, Мэтью Р.; Годфрей, Х. Чарльз Дж .; Берт, Остин (31 марта 2005 г.). «Секс повышает эффективность естественного отбора в экспериментальных популяциях дрожжей» . Природа . 434 (7033): 636–640. Бибкод : 2005Natur.434..636G . дои : 10.1038/nature03405 . ISSN 0028-0836 . ПМИД 15800622 . S2CID 4397491 .
- ^ Мейнард Смит 1978 [ нужна страница ]
- ^ Перейти обратно: а б Ридли 2004 , с. 314
- ^ Ван Вален, Ли (1973). «Новый эволюционный закон» (PDF) . Эволюционная теория . 1 :1–30. ISSN 0093-4755 . Архивировано из оригинала (PDF) 22 декабря 2014 года . Проверено 24 декабря 2014 г.
- ^ Гамильтон, Вашингтон ; Аксельрод, Роберт ; Танезе, Рэйко (1 мая 1990 г.). «Половое размножение как приспособление к сопротивлению паразитам (обзор)» . ПНАС . 87 (9): 3566–3573. Бибкод : 1990PNAS...87.3566H . дои : 10.1073/pnas.87.9.3566 . ISSN 0027-8424 . ПМЦ 53943 . ПМИД 2185476 .
- ^ Бердселл и Уиллс 2003 , стр. 113–117.
- ^ Бернштейн Х., Байерли Х.К., Хопф Ф.А., Мишод Р.Э. Генетические повреждения, мутации и эволюция пола. Наука. 1985, 20 сентября; 229 (4719): 1277–81. дои : 10.1126/science.3898363 . ПМИД 3898363
- ^ Бернштейн Х., Хопф Ф.А., Мишод Р.Э. Молекулярные основы эволюции пола. Адв Генет. 1987;24:323-70. дои : 10.1016/s0065-2660(08)60012-7 . ПМИД 3324702
- ^ Морджан, Кэрри Л.; Ризеберг, Лорен Х. (июнь 2004 г.). «Как виды эволюционируют коллективно: последствия потока генов и отбора для распространения выгодных аллелей» . Молекулярная экология . 13 (6): 1341–1356. Бибкод : 2004MolEc..13.1341M . дои : 10.1111/j.1365-294X.2004.02164.x . ISSN 0962-1083 . ПМК 2600545 . ПМИД 15140081 .
- ^ Баучер, Ян; Дуади, Кристоф Дж.; Папке, Р. Тейн; и др. (декабрь 2003 г.). «Боковой перенос генов и происхождение прокариотических групп». Ежегодный обзор генетики . 37 : 283–328. дои : 10.1146/annurev.genet.37.050503.084247 . ISSN 0066-4197 . ПМИД 14616063 .
- ^ Уолш, Тимоти Р. (октябрь 2006 г.). «Комбинаторная генетическая эволюция мультирезистентности». Современное мнение в микробиологии . 9 (5): 476–482. дои : 10.1016/j.mib.2006.08.009 . ISSN 1369-5274 . ПМИД 16942901 .
- ^ Кондо, Нацуко; Нико, Наруо; Иджичи, Нобуюки; и др. (29 октября 2002 г.). «Фрагмент генома эндосимбионта Wolbachia , перенесенный на Х-хромосому насекомого-хозяина» . ПНАС . 99 (22): 14280–14285. Бибкод : 2002PNAS...9914280K . дои : 10.1073/pnas.222228199 . ISSN 0027-8424 . ПМЦ 137875 . ПМИД 12386340 .
- ^ Спраг, Джордж Ф. младший (декабрь 1991 г.). «Генетический обмен между королевствами». Текущее мнение в области генетики и развития . 1 (4): 530–533. дои : 10.1016/S0959-437X(05)80203-5 . ISSN 0959-437X . ПМИД 1822285 .
- ^ Гладышев Евгений А.; Мезельсон, Мэтью ; Архипова Ирина Р. (30 мая 2008 г.). «Массовый горизонтальный перенос генов у бделлоидных коловраток» . Наука . 320 (5880): 1210–1213. Бибкод : 2008Sci...320.1210G . дои : 10.1126/science.1156407 . ISSN 0036-8075 . ПМИД 18511688 . S2CID 11862013 . Архивировано из оригинала 30 июля 2022 года . Проверено 30 июля 2022 г.
- ^ Бальдо, Анджела М.; МакКлюр, Марселла А. (сентябрь 1999 г.). «Эволюция и горизонтальный перенос генов, кодирующих дУТФазу, в вирусах и их хозяевах» . Журнал вирусологии . 73 (9): 7710–7721. doi : 10.1128/JVI.73.9.7710-7721.1999 . ISSN 0022-538X . ПМЦ 104298 . ПМИД 10438861 .
- ^ Ривера, Мария К.; Лейк, Джеймс А. (9 сентября 2004 г.). «Кольцо жизни является доказательством происхождения эукариот в результате слияния геномов» . Природа . 431 (7005): 152–155. Бибкод : 2004Natur.431..152R . дои : 10.1038/nature02848 . ISSN 0028-0836 . ПМИД 15356622 . S2CID 4349149 .
- ^ Яблонка, Ева; Раз, Гал (июнь 2009 г.). «Трансгенерационная эпигенетическая наследственность: распространенность, механизмы и последствия для изучения наследственности и эволюции» (PDF) . Ежеквартальный обзор биологии . 84 (2): 131–176. CiteSeerX 10.1.1.617.6333 . дои : 10.1086/598822 . ISSN 0033-5770 . ПМИД 19606595 . S2CID 7233550 . Архивировано (PDF) из оригинала 15 июля 2011 года . Проверено 30 июля 2022 г.
- ^ Боссдорф, Оливер; Аркури, Давиде; Ричардс, Кристина Л.; Пильуччи, Массимо (май 2010 г.). «Экспериментальное изменение метилирования ДНК влияет на фенотипическую пластичность экологически значимых признаков Arabidopsis thaliana » (PDF) . Эволюционная экология . 24 (3): 541–553. Бибкод : 2010EvEco..24..541B . дои : 10.1007/s10682-010-9372-7 . ISSN 0269-7653 . S2CID 15763479 . Архивировано (PDF) из оригинала 5 июня 2022 года . Проверено 30 июля 2022 г.
- ^ Яблонка, Ева; Лэмб, Мэрион Дж. (декабрь 2002 г.). «Изменяющаяся концепция эпигенетики». Анналы Нью-Йоркской академии наук . 981 (1): 82–96. Бибкод : 2002NYASA.981...82J . дои : 10.1111/j.1749-6632.2002.tb04913.x . ISSN 0077-8923 . ПМИД 12547675 . S2CID 12561900 .
- ^ Лаланд, Кевин Н.; Стерельный, Ким (сентябрь 2006 г.). «Перспектива: семь причин (не) пренебрегать строительством ниши» . Эволюция . 60 (9): 1751–1762. дои : 10.1111/j.0014-3820.2006.tb00520.x . ISSN 0014-3820 . ПМИД 17089961 . S2CID 22997236 .
- ^ Чепмен, Майкл Дж.; Маргулис, Линн (декабрь 1998 г.). «Морфогенез посредством симбиогенеза» (PDF) . Международная микробиология . 1 (4): 319–326. ISSN 1139-6709 . ПМИД 10943381 . Архивировано из оригинала (PDF) 23 августа 2014 года . Проверено 9 декабря 2014 г.
- ^ Уилсон, Дэвид Слоан ; Уилсон, Эдвард О. (декабрь 2007 г.). «Переосмысление теоретической основы социобиологии» (PDF) . Ежеквартальный обзор биологии . 82 (4): 327–348. дои : 10.1086/522809 . ISSN 0033-5770 . ПМИД 18217526 . S2CID 37774648 . Архивировано из оригинала (PDF) 11 мая 2011 года.
- ^ Юэнс 2004 [ нужна страница ]
- ^ Перейти обратно: а б с Херст, Лоуренс Д. (февраль 2009 г.). «Основные понятия генетики: генетика и понимание отбора». Обзоры природы Генетика . 10 (2): 83–93. дои : 10.1038/nrg2506 . ПМИД 19119264 . S2CID 1670587 .
- ^ Дарвин 1859 г. , Глава XIV
- ^ Отто, Сара П .; Серведио, Мария Р .; Нуисмер, Скотт Л. (август 2008 г.). «Частотно-зависимый отбор и эволюция ассортативного спаривания» . Генетика . 179 (4): 2091–2112. doi : 10.1534/genetics.107.084418 . ПМК 2516082 . ПМИД 18660541 .
- ^ Перейти обратно: а б с Орр, Х. Аллен (август 2009 г.). «Приспособленность и ее роль в эволюционной генетике» . Обзоры природы Генетика . 10 (8): 531–539. дои : 10.1038/nrg2603 . ISSN 1471-0056 . ПМЦ 2753274 . ПМИД 19546856 .
- ^ Холдейн, JBS (14 марта 1959 г.). «Теория естественного отбора сегодня» . Природа . 183 (4663): 710–713. Бибкод : 1959Natur.183..710H . дои : 10.1038/183710a0 . ПМИД 13644170 . S2CID 4185793 .
- ^ Ланде, Рассел ; Арнольд, Стеван Дж. (ноябрь 1983 г.). «Измерение отбора коррелирующих символов». Эволюция . 37 (6): 1210–1226. дои : 10.1111/j.1558-5646.1983.tb00236.x . ISSN 0014-3820 . JSTOR 2408842 . ПМИД 28556011 . S2CID 36544045 .
- ^ Гольдберг, Эмма Э.; Игич, Борис (ноябрь 2008 г.). «О филогенетических тестах необратимой эволюции». Эволюция . 62 (11): 2727–2741. дои : 10.1111/j.1558-5646.2008.00505.x . ISSN 0014-3820 . ПМИД 18764918 . S2CID 30703407 .
- ^ Коллин, Рэйчел; Миглиетта, Мария Пиа (ноябрь 2008 г.). «Изменение мнений о законе Долло». Тенденции в экологии и эволюции . 23 (11): 602–609. Бибкод : 2008TEcoE..23..602C . дои : 10.1016/j.tree.2008.06.013 . ПМИД 18814933 .
- ^ Томич, Ненад; Мейер-Рохов, Виктор Бенно (2011). «Атавизмы: медицинские, генетические и эволюционные последствия» . Перспективы биологии и медицины . 54 (3): 332–353. дои : 10.1353/pbm.2011.0034 . ПМИД 21857125 . S2CID 40851098 .
- ^ Хоекстра, Хопи Э.; Хекстра, Джонатан М.; Берриган, Дэвид; и др. (31 июля 2001 г.). «Сила и темп направленного выбора в дикой природе» . ПНАС . 98 (16): 9157–9160. Бибкод : 2001PNAS...98.9157H . дои : 10.1073/pnas.161281098 . ПМК 55389 . ПМИД 11470913 .
- ^ Фельзенштейн, Джозеф (ноябрь 1979 г.). «Экскурсии по границе между подрывным и стабилизирующим отбором» . Генетика . 93 (3): 773–795. дои : 10.1093/генетика/93.3.773 . ПМЦ 1214112 . ПМИД 17248980 .
- ^ Одум 1971 , с. 8
- ^ Окаша 2006 г.
- ^ Перейти обратно: а б Гулд, Стивен Джей (28 февраля 1998 г.). «Дальнейшие путешествия Гулливера: необходимость и трудность иерархической теории отбора» . Философские труды Королевского общества Б. 353 (1366): 307–314. дои : 10.1098/rstb.1998.0211 . ISSN 0962-8436 . ПМЦ 1692213 . ПМИД 9533127 .
- ^ Майр, Эрнст (18 марта 1997 г.). «Объекты выбора» . ПНАС . 94 (6): 2091–2094. Бибкод : 1997ПНАС...94.2091М . дои : 10.1073/pnas.94.6.2091 . ISSN 0027-8424 . ПМК 33654 . ПМИД 9122151 .
- ^ Мейнард Смит 1998 , стр. 203–211, обсуждение 211–217.
- ^ Хики, Донал А. (1992). «Эволюционная динамика мобильных элементов у прокариот и эукариот». Генетика . 86 (1–3): 269–274. дои : 10.1007/BF00133725 . ISSN 0016-6707 . ПМИД 1334911 . S2CID 6583945 .
- ^ Гулд, Стивен Джей; Ллойд, Элизабет А. (12 октября 1999 г.). «Индивидуальность и адаптация на разных уровнях отбора: как нам назвать и обобщить единицу дарвинизма?» . ПНАС . 96 (21): 11904–11909. Бибкод : 1999PNAS...9611904G . дои : 10.1073/pnas.96.21.11904 . ISSN 0027-8424 . ЧВК 18385 . ПМИД 10518549 .
- ^ Футуйма и Киркпатрик, 2017 , стр. 55–66, Глава 3: Естественный отбор и адаптация.
- ^ Перейти обратно: а б Мазель, Джоанна (25 октября 2011 г.). «Генетический дрейф» . Современная биология . 21 (20): Р837–Р838. Бибкод : 2011CBio...21.R837M . дои : 10.1016/j.cub.2011.08.007 . ISSN 0960-9822 . ПМИД 22032182 . S2CID 17619958 .
- ^ Ланде, Рассел (1989). «Теории видообразования Фишера и Райта» . Геном . 31 (1): 221–227. дои : 10.1139/g89-037 . ISSN 0831-2796 . ПМИД 2687093 .
- ^ Кимура, Мотоо (1991). «Нейтральная теория молекулярной эволюции: обзор последних данных» . Японский журнал генетики человека . 66 (4): 367–386. дои : 10.1266/jjg.66.367 . ПМИД 1954033 . Архивировано из оригинала 5 июня 2022 года.
- ^ Кимура, Мотоо (1989). «Нейтральная теория молекулярной эволюции и мировоззрение нейтралистов». Геном . 31 (1): 24–31. дои : 10.1139/g89-009 . ISSN 0831-2796 . ПМИД 2687096 .
- ^ Крейтман, Мартин (август 1996 г.). «Нейтральная теория мертва. Да здравствует нейтральная теория» . Биоэссе . 18 (8): 678–683, обсуждение 683. doi : 10.1002/bies.950180812 . ISSN 0265-9247 . ПМИД 8760341 .
- ^ Ли, Э.Г. младший (ноябрь 2007 г.). «Нейтральная теория: историческая перспектива» . Журнал эволюционной биологии . 20 (6): 2075–2091. дои : 10.1111/j.1420-9101.2007.01410.x . ISSN 1010-061X . ПМИД 17956380 . S2CID 2081042 .
- ^ Перейти обратно: а б Гиллеспи, Джон Х. (ноябрь 2001 г.). «Влияет ли размер популяции вида на его эволюцию?» . Эволюция . 55 (11): 2161–2169. дои : 10.1111/j.0014-3820.2001.tb00732.x . ISSN 0014-3820 . ПМИД 11794777 . S2CID 221735887 .
- ^ Неер, Ричард А.; Шрайман, Борис И. (август 2011 г.). «Генетический проект и квазинейтральность в больших факультативно-сексуальных популяциях» . Генетика . 188 (4): 975–996. arXiv : 1108.1635 . Бибкод : 2011arXiv1108.1635N . дои : 10.1534/genetics.111.128876 . ПМК 3176096 . ПМИД 21625002 .
- ^ Штольцфус, Арлин (1999). «О возможности конструктивной нейтральной эволюции» . Журнал молекулярной эволюции . 49 (2): 169–181. Бибкод : 1999JMolE..49..169S . дои : 10.1007/PL00006540 . ПМИД 10441669 . S2CID 1743092 . Архивировано из оригинала 30 июля 2022 года . Проверено 30 июля 2022 г.
- ^ Штольцфус, Арлин (13 октября 2012 г.). «Конструктивная нейтральная эволюция: исследование любопытного противоречия эволюционной теории» . Биология Директ . 7 (1): 35. дои : 10.1186/1745-6150-7-35 . ПМЦ 3534586 . ПМИД 23062217 .
- ^ Муньос-Гомес, Серхио А.; Билоликар, Гаурав; Уайдман, Джереми Г.; и др. (1 апреля 2021 г.). «Конструктивная нейтральная эволюция 20 лет спустя» . Журнал молекулярной эволюции . 89 (3): 172–182. Бибкод : 2021JMolE..89..172M . дои : 10.1007/s00239-021-09996-y . ПМЦ 7982386 . ПМИД 33604782 .
- ^ Лукеш, Юлиус; Арчибальд, Джон М.; Килинг, Патрик Дж.; и др. (2011). «Как нейтральный эволюционный храповик может усложнить клетку» . ИУБМБ Жизнь . 63 (7): 528–537. дои : 10.1002/iub.489 . ПМИД 21698757 . S2CID 7306575 .
- ^ Воссеберг, Джулиан; Снел, Беренд (1 декабря 2017 г.). «Одомашнивание самосплайсинговых интронов во время эукариогенеза: возникновение сложного сплайсосомного аппарата» . Биология Директ . 12 (1): 30. дои : 10.1186/s13062-017-0201-6 . ПМК 5709842 . ПМИД 29191215 .
- ^ Брюне, TDP; Дулитл, У. Форд (19 марта 2018 г.). «Общность конструктивной нейтральной эволюции». Биология и философия . 33 (1): 2. дои : 10.1007/s10539-018-9614-6 . S2CID 90290787 .
- ^ Отто, Сара П.; Уитлок, Майкл К. (июнь 1997 г.). «Вероятность фиксации в популяциях меняющегося размера» (PDF) . Генетика . 146 (2): 723–733. дои : 10.1093/генетика/146.2.723 . ПМК 1208011 . ПМИД 9178020 . Архивировано (PDF) из оригинала 19 марта 2015 года . Проверено 18 декабря 2014 г.
- ^ Перейти обратно: а б Чарльзуорт, Брайан (март 2009 г.). «Фундаментальные концепции генетики: эффективный размер популяции и закономерности молекулярной эволюции и изменчивости». Обзоры природы Генетика . 10 (3): 195–205. дои : 10.1038/nrg2526 . ПМИД 19204717 . S2CID 205484393 .
- ^ Каттер, Ашер Д.; Чхве, Джэ Ён (август 2010 г.). «Естественный отбор формирует полиморфизм нуклеотидов в геноме нематоды Caenorhabditis briggsae » . Геномные исследования . 20 (8): 1103–1111. дои : 10.1101/гр.104331.109 . ПМК 2909573 . ПМИД 20508143 .
- ^ Митчелл-Олдс, Томас; Уиллис, Джон Х.; Гольдштейн, Дэвид Б. (ноябрь 2007 г.). «Какие эволюционные процессы влияют на естественные генетические вариации фенотипических признаков?». Обзоры природы Генетика . 8 (11): 845–856. дои : 10.1038/nrg2207 . ISSN 1471-0056 . ПМИД 17943192 . S2CID 14914998 .
- ^ Ней, Масатоши (декабрь 2005 г.). «Селекционизм и нейтрализм в молекулярной эволюции» . Молекулярная биология и эволюция . 22 (12): 2318–2342. дои : 10.1093/molbev/msi242 . ISSN 0737-4038 . ПМЦ 1513187 . ПМИД 16120807 .
- Ней, Масатоши (май 2006 г.). «Селекционизм и нейтрализм в молекулярной эволюции» . Молекулярная биология и эволюция (ошибка). 23 (5): 2318–42. дои : 10.1093/molbev/msk009 . ISSN 0737-4038 . ПМЦ 1513187 . ПМИД 16120807 .
- ^ Холдейн, JBS (июль 1927 г.). «Математическая теория естественного и искусственного отбора, Часть V: Отбор и мутация». Труды Кембриджского философского общества . 26 (7): 838–844. Бибкод : 1927PCPS...23..838H . дои : 10.1017/S0305004100015644 . S2CID 86716613 .
- ^ Фишер 1930 г.
- ^ Перейти обратно: а б с Ямпольский Лев Юрьевич; Штольцфус, Арлин (20 декабря 2001 г.). «Предвзятость во введении вариаций как ориентирующий фактор эволюции». Эволюция и развитие . 3 (2): 73–83. дои : 10.1046/j.1525-142x.2001.003002073.x . ПМИД 11341676 . S2CID 26956345 .
- ^ Суэока, Нобору (1 апреля 1962 г.). «О генетических основах изменчивости и гетерогенности базового состава ДНК» . ПНАС . 48 (4): 582–592. Бибкод : 1962ПНАС...48..582С . дои : 10.1073/pnas.48.4.582 . ПМК 220819 . ПМИД 13918161 .
- ^ Фриз, Эрнст (июль 1962 г.). «Об эволюции основного состава ДНК». Журнал теоретической биологии . 3 (1): 82–101. Бибкод : 1962JThBi...3...82F . дои : 10.1016/S0022-5193(62)80005-8 .
- ^ Кокс, Эдвард К.; Янофски, Чарльз (1 ноября 1967 г.). «Измененное соотношение оснований в ДНК штамма-мутатора Escherichia coli» . Учеб. Натл. акад. наук. США . 58 (5): 1895–1902. Бибкод : 1967PNAS...58.1895C . дои : 10.1073/pnas.58.5.1895 . ПМК 223881 . ПМИД 4866980 .
- ^ Шах, Премал; Гилкрист, Майкл А. (21 июня 2011 г.). «Объяснение сложных моделей использования кодонов с учетом эффективности трансляции, предвзятости мутаций и генетического дрейфа» . ПНАС . 108 (25): 10231–10236. Бибкод : 2011PNAS..10810231S . дои : 10.1073/pnas.1016719108 . ПМК 3121864 . ПМИД 21646514 .
- ^ Балмер, Майкл Г. (ноябрь 1991 г.). «Теория отбора-мутации-дрейфа использования синонимичных кодонов» . Генетика . 129 (3): 897–907. дои : 10.1093/генетика/129.3.897 . ПМК 1204756 . ПМИД 1752426 .
- ^ Фрикселл, Карл Дж.; Цукеркандль, Эмиль (сентябрь 2000 г.). «Дезаминирование цитозина играет основную роль в эволюции изохор млекопитающих» . Молекулярная биология и эволюция . 17 (9): 1371–1383. doi : 10.1093/oxfordjournals.molbev.a026420 . ПМИД 10958853 .
- ^ Петров Дмитрий А.; Сангстер, Тодд А.; Джонстон, Дж. Спенсер; и др. (11 февраля 2000 г.). «Доказательства потери ДНК как определяющего фактора размера генома». Наука . 287 (5455): 1060–1062. Бибкод : 2000Sci...287.1060P . дои : 10.1126/science.287.5455.1060 . ISSN 0036-8075 . ПМИД 10669421 . S2CID 12021662 .
- ^ Петров, Дмитрий А. (май 2002 г.). «Потеря ДНК и эволюция размера генома у дрозофилы » . Генетика . 115 (1): 81–91. дои : 10.1023/А:1016076215168 . ISSN 0016-6707 . ПМИД 12188050 . S2CID 5314242 .
- ^ Дюре, Лоран; Галтье, Николя (сентябрь 2009 г.). «Смещенная конверсия генов и эволюция геномных ландшафтов млекопитающих». Ежегодный обзор геномики и генетики человека . 10 . Годовые обзоры: 285–311. doi : 10.1146/annurev-genom-082908-150001 . ПМИД 19630562 . S2CID 9126286 .
- ^ Хершберг, Рут; Петров, Дмитрий А. (9 сентября 2010 г.). «Доказательства того, что мутация у бактерий повсеместно смещена в сторону АТ» . ПЛОС Генетика . 6 (9): e1001115. дои : 10.1371/journal.pgen.1001115 . ПМЦ 2936535 . ПМИД 20838599 .
- ^ А. Штольцфус (2019). «Понимание предвзятости при введении вариаций как эволюционной причины». В Уллере, Т.; Лаланд, КНЦ (ред.). Эволюционная причинность: биологические и философские размышления . Кембридж, Массачусетс: MIT Press.
- ^ Штольцфус, Арлин; МакКэндлиш, Дэвид М. (сентябрь 2017 г.). «Мутационные предубеждения влияют на параллельную адаптацию» . Молекулярная биология и эволюция . 34 (9): 2163–2172. дои : 10.1093/molbev/msx180 . ПМК 5850294 . ПМИД 28645195 .
- ^ Пейн, Джошуа Л.; Менардо, Фабрицио; Траунер, Андрей; и др. (13 мая 2019 г.). «Предвзятость перехода влияет на эволюцию устойчивости к антибиотикам у микобактерий туберкулеза » . ПЛОС Биология . 17 (5): e3000265. дои : 10.1371/journal.pbio.3000265 . ПМК 6532934 . ПМИД 31083647 .
- ^ Сторц, Джей Ф.; Натараджан, Чандрасекхар; Синьор, Энтони В.; и др. (22 июля 2019 г.). «Роль мутационной предвзятости в адаптивной молекулярной эволюции: выводы из конвергентных изменений в функции белка» . Философские труды Королевского общества Б. 374 : 20180238.doi : (1777 ) 10.1098/rstb.2018.0238 . ПМК 6560279 . ПМИД 31154983 .
- ^ Свенссон, Эрик И.; Бергер, Дэвид (1 мая 2019 г.). «Роль мутационной предвзятости в адаптивной эволюции». Тенденции в экологии и эволюции . 34 (5): 422–434. Бибкод : 2019TEcoE..34..422S . дои : 10.1016/j.tree.2019.01.015 . ПМИД 31003616 . S2CID 125066709 .
- ^ Лиен, Сигбьёрн; Шида, Джоанна; Шечингер, Биргит; и др. (февраль 2000 г.). «Доказательства гетерогенности рекомбинации в псевдоаутосомной области человека: анализ высокого разрешения с помощью типирования сперматозоидов и радиационно-гибридного картирования» . Американский журнал генетики человека . 66 (2): 557–566. дои : 10.1086/302754 . ISSN 0002-9297 . ПМЦ 1288109 . ПМИД 10677316 .
- ^ Бартон, Николас Х. (29 ноября 2000 г.). «Генетический автостоп» . Философские труды Королевского общества Б. 355 (1403): 1553–1562. дои : 10.1098/rstb.2000.0716 . ISSN 0962-8436 . ПМК 1692896 . ПМИД 11127900 .
- ^ Райс, К; Спете, Дж; Штатечный, М; Строндль, К; Хёдль, Вт (20 октября 2008 г.). «Синий и ультрафиолет: изменение цвета в зависимости от пола во время брачного сезона у балканской болотной лягушки» . Журнал зоологии . 276 (3): 229–236. doi : 10.1111/j.1469-7998.2008.00456.x – через Академию Google.
- ^ Андерссон, Мальте; Симмонс, Ли В. (июнь 2006 г.). «Половой отбор и выбор партнера» (PDF) . Тенденции в экологии и эволюции . 21 (6): 296–302. CiteSeerX 10.1.1.595.4050 . дои : 10.1016/j.tree.2006.03.015 . ISSN 0169-5347 . ПМИД 16769428 . Архивировано (PDF) из оригинала 9 марта 2013 года.
- ^ Кокко, Ханна ; Брукс, Роберт; Макнамара, Джон М.; Хьюстон, Аласдер И. (7 июля 2002 г.). «Континуум полового отбора» . Труды Королевского общества Б. 269 (1498): 1331–1340. дои : 10.1098/рспб.2002.2020 . ISSN 0962-8452 . ПМК 1691039 . ПМИД 12079655 .
- ^ Куинн, Томас П.; Хендри, Эндрю П.; Бак, Грегори Б. (2001). «Баланс естественного и полового отбора нерки: взаимодействие между размером тела, репродуктивными возможностями и уязвимостью к хищникам со стороны медведей» (PDF) . Исследования в области эволюционной экологии . 3 : 917–937. ISSN 1522-0613 . Архивировано (PDF) из оригинала 5 марта 2016 года . Проверено 15 декабря 2014 г.
- ^ Хант, Джон; Брукс, Роберт; Дженнионс, Майкл Д.; и др. (23 декабря 2004 г.). «Высококачественные самцы полевых сверчков вкладывают значительные средства в сексуальные проявления, но умирают молодыми». Природа . 432 (7020): 1024–1027. Бибкод : 2004Natur.432.1024H . дои : 10.1038/nature03084 . ISSN 0028-0836 . ПМИД 15616562 . S2CID 4417867 .
- ^ Байм, Майкл; Либерман, Тами Д.; Келсик, Эрик Д.; и др. (9 сентября 2016 г.). «Пространственно-временная эволюция микробов на ландшафтах антибиотиков» . Наука . 353 (6304): 1147–1151. Бибкод : 2016Sci...353.1147B . дои : 10.1126/science.aag0822 . ISSN 0036-8075 . ПМЦ 5534434 . ПМИД 27609891 .
- ^ Перейти обратно: а б с Скотт, Юджини С .; Мацке, Николас Дж. (15 мая 2007 г.). «Биологический дизайн в научных классах» . ПНАС . 104 (Приложение 1): 8669–8676. Бибкод : 2007PNAS..104.8669S . дои : 10.1073/pnas.0701505104 . ПМК 1876445 . ПМИД 17494747 .
- ^ Хендри, Эндрю Пол; Киннисон, Майкл Т. (ноябрь 2001 г.). «Введение в микроэволюцию: скорость, закономерности, процесс». Генетика . 112–113 (1): 1–8. дои : 10.1023/A:1013368628607 . ISSN 0016-6707 . ПМИД 11838760 . S2CID 24485535 .
- ^ Леруа, Арманд М. (март – апрель 2000 г.). «Масштабная независимость эволюции». Эволюция и развитие . 2 (2): 67–77. CiteSeerX 10.1.1.120.1020 . дои : 10.1046/j.1525-142x.2000.00044.x . ISSN 1520-541X . ПМИД 11258392 . S2CID 17289010 .
- ^ Гулд 2002 , стр. 657–658.
- ^ Перейти обратно: а б Гулд, Стивен Джей (19 июля 1994 г.). «Темп и модус в макроэволюционной реконструкции дарвинизма» . ПНАС . 91 (15): 6764–6771. Бибкод : 1994PNAS...91.6764G . дои : 10.1073/pnas.91.15.6764 . ПМК 44281 . ПМИД 8041695 .
- ^ Яблонски, Дэвид (2000). «Микро- и макроэволюция: масштаб и иерархия в эволюционной биологии и палеобиологии». Палеобиология . 26 (сп4): 15–52. doi : 10.1666/0094-8373(2000)26[15:MAMSAH]2.0.CO;2 . S2CID 53451360 .
- ^ Догерти, Майкл Дж. (20 июля 1998 г.). «Человеческая раса развивается или деградирует?» . Научный американец . ISSN 0036-8733 . Архивировано из оригинала 6 мая 2014 года . Проверено 11 сентября 2015 г.
- ^ Исаак, Марк, изд. (22 июля 2003 г.). «Заявление CB932: Эволюция вырожденных форм» . Архив TalkOrigins . Хьюстон, Техас: Фонд TalkOrigins. Архивировано из оригинала 23 августа 2014 года . Проверено 19 декабря 2014 г.
- ^ Лейн 1996 , с. 61
- ^ Кэрролл, Шон Б. (22 февраля 2001 г.). «Случайность и необходимость: эволюция морфологической сложности и разнообразия» . Природа . 409 (6823): 1102–1109. Бибкод : 2001Natur.409.1102C . дои : 10.1038/35059227 . ПМИД 11234024 . S2CID 4319886 .
- ^ Уитмен, Уильям Б.; Коулман, Дэвид С.; Вибе, Уильям Дж. (9 июня 1998 г.). «Прокариоты: невидимое большинство» . ПНАС . 95 (12): 6578–6583. Бибкод : 1998PNAS...95.6578W . дои : 10.1073/pnas.95.12.6578 . ISSN 0027-8424 . ПМК 33863 . ПМИД 9618454 .
- ^ Перейти обратно: а б Шлосс, Патрик Д.; Хандельсман, Джо (декабрь 2004 г.). «Состояние микробной переписи» . Обзоры микробиологии и молекулярной биологии . 68 (4): 686–691. дои : 10.1128/MMBR.68.4.686-691.2004 . ПМК 539005 . ПМИД 15590780 .
- ^ Нилсон, Кеннет Х. (январь 1999 г.). «Микробиология после викингов: новые подходы, новые данные, новые идеи» . Происхождение жизни и эволюция биосфер . 29 (1): 73–93. Бибкод : 1999ОЛЕВ...29...73Н . дои : 10.1023/А:1006515817767 . ISSN 0169-6149 . ПМИД 11536899 . S2CID 12289639 .
- ^ Баклинг, Ангус; Маклин, Р. Крейг; Брокхерст, Майкл А.; Коулгрейв, Ник (12 февраля 2009 г.). «Бигль в бутылке» . Природа . 457 (7231): 824–829. Бибкод : 2009Natur.457..824B . дои : 10.1038/nature07892 . ISSN 0028-0836 . ПМИД 19212400 . S2CID 205216404 .
- ^ Елена, Сантьяго Ф.; Ленски, Ричард Э. (июнь 2003 г.). «Эволюционные эксперименты с микроорганизмами: динамика и генетические основы адаптации». Обзоры природы Генетика . 4 (6): 457–469. дои : 10.1038/nrg1088 . ISSN 1471-0056 . ПМИД 12776215 . S2CID 209727 .
- ^ Майр 1982 , с. 483: «Адаптация... больше не могла считаться статическим состоянием, продуктом творческого прошлого, а вместо этого стала продолжающимся динамическим процессом».
- ^ Шестое издание Оксфордского научного словаря (2010 г.) определяет адаптацию как «любое изменение в структуре или функционировании последующих поколений популяции, которое делает ее более приспособленной к окружающей среде».
- ^ Орр, Х. Аллен (февраль 2005 г.). «Генетическая теория адаптации: краткая история». Обзоры природы Генетика . 6 (2): 119–127. дои : 10.1038/nrg1523 . ISSN 1471-0056 . ПМИД 15716908 . S2CID 17772950 .
- ^ Добжанский 1968 , стр. 1–34.
- ^ Добжанский 1970 , стр. 4–6, 79–82, 84–87.
- ^ Добжанский, Феодосий (март 1956 г.). «Генетика природных популяций. XXV. Генетические изменения в популяциях Drosophila pseudoobscura и Drosophila persimilis в некоторых местностях Калифорнии» . Эволюция . 10 (1): 82–92. дои : 10.2307/2406099 . ISSN 0014-3820 . JSTOR 2406099 .
- ^ Накадзима, Акира; Сугимото, Ёко; Ёнеяма, Хироши; и др. (июнь 2002 г.). «Высокий уровень устойчивости к фторхинолонам у Pseudomonas aeruginosa из-за взаимодействия эффлюксного насоса MexAB-OprM и мутации ДНК-гиразы» . Микробиология и иммунология . 46 (6): 391–395. дои : 10.1111/j.1348-0421.2002.tb02711.x . ISSN 1348-0421 . ПМИД 12153116 . S2CID 22593331 .
- ^ Блаунт, Закари Д.; Борланд, Кристина З.; Ленски, Ричард Э. (10 июня 2008 г.). «Первая статья: Историческая случайность и эволюция ключевой инновации в экспериментальной популяции Escherichia coli » . ПНАС . 105 (23): 7899–7906. Бибкод : 2008PNAS..105.7899B . дои : 10.1073/pnas.0803151105 . ISSN 0027-8424 . ПМЦ 2430337 . ПМИД 18524956 .
- ^ Окада, Хиросукэ; Негоро, Сейджи; Кимура, Хироюки; и др. (10 ноября 1983 г.). «Эволюционная адаптация кодируемых плазмидами ферментов для разложения олигомеров нейлона». Природа . 306 (5939): 203–206. Бибкод : 1983Natur.306..203O . дои : 10.1038/306203a0 . ISSN 0028-0836 . ПМИД 6646204 . S2CID 4364682 .
- ^ Оно, Сусуму (апрель 1984 г.). «Рождение уникального фермента из альтернативной рамки считывания ранее существовавшей, внутренне повторяющейся кодирующей последовательности» . ПНАС . 81 (8): 2421–2425. Бибкод : 1984PNAS...81.2421O . дои : 10.1073/pnas.81.8.2421 . ISSN 0027-8424 . ПМК 345072 . ПМИД 6585807 .
- ^ Копли, Шелли Д. (июнь 2000 г.). «Эволюция метаболического пути разложения токсичного ксенобиотика: лоскутный подход». Тенденции биохимических наук . 25 (6): 261–265. дои : 10.1016/S0968-0004(00)01562-0 . ISSN 0968-0004 . ПМИД 10838562 .
- ^ Кроуфорд, Рональд Л.; Юнг, Карина М.; Ремень, Дженис Л. (октябрь 2007 г.). «Недавняя эволюция пентахлорфенол (PCP)-4-монооксигеназы (PcpB) и связанные с ней пути бактериальной деградации PCP». Биодеградация . 18 (5): 525–539. дои : 10.1007/s10532-006-9090-6 . ISSN 0923-9820 . ПМИД 17123025 . S2CID 8174462 .
- ^ Альтенберг 1995 , стр. 205–259.
- ^ Мазель, Джоанна ; Бергман, Авив (июль 2003 г.). «Эволюция свойств эволюционируемости дрожжевого приона [PSI +]» . Эволюция . 57 (7): 1498–1512. дои : 10.1111/j.0014-3820.2003.tb00358.x . ПМИД 12940355 . S2CID 30954684 .
- ^ Ланкастер, Алекс К.; Бардилл, Дж. Патрик; Верно, Хизер Л.; Мазель, Джоанна (февраль 2010 г.). «Скорость спонтанного появления дрожжевого приона [ PSI +] и ее влияние на эволюцию свойств эволюционности системы [ PSI +]» . Генетика . 184 (2): 393–400. дои : 10.1534/genetics.109.110213 . ISSN 0016-6731 . ПМЦ 2828720 . ПМИД 19917766 .
- ^ Драги, Джереми; Вагнер, Гюнтер П. (февраль 2008 г.). «Эволюция эволюционности в модели развития». Эволюция . 62 (2): 301–315. дои : 10.1111/j.1558-5646.2007.00303.x . ПМИД 18031304 . S2CID 11560256 .
- ^ Перейти обратно: а б Бейдер, Ларс; Холл, Брайан К. (ноябрь 2002 г.). «Конечности у китов и отсутствие конечностей у других позвоночных: механизмы эволюционной и развивающей трансформации и утраты». Эволюция и развитие . 4 (6): 445–458. дои : 10.1046/j.1525-142X.2002.02033.x . ПМИД 12492145 . S2CID 8448387 .
- ^ Янг, Натан М.; Халлгримссон, Бенедикт (декабрь 2005 г.). «Серийная гомология и эволюция ковариационной структуры конечностей млекопитающих» . Эволюция . 59 (12): 2691–2704. дои : 10.1554/05-233.1 . ISSN 0014-3820 . ПМИД 16526515 . S2CID 198156135 .
- ^ Перейти обратно: а б Пенни, Дэвид; Пул, Энтони (декабрь 1999 г.). «Природа последнего всеобщего общего предка». Текущее мнение в области генетики и развития . 9 (6): 672–677. дои : 10.1016/S0959-437X(99)00020-9 . ПМИД 10607605 .
- ^ Холл, Брайан К. (август 2003 г.). «Происхождение с модификацией: единство, лежащее в основе гомологии и гомоплазии, как видно из анализа развития и эволюции» . Биологические обзоры . 78 (3): 409–433. дои : 10.1017/S1464793102006097 . ISSN 1464-7931 . ПМИД 14558591 . S2CID 22142786 .
- ^ Шубин, Нил ; Табин, Клиффорд Дж .; Кэрролл, Шон Б. (12 февраля 2009 г.). «Глубокая гомология и истоки эволюционной новизны» . Природа . 457 (7231): 818–823. Бибкод : 2009Natur.457..818S . дои : 10.1038/nature07891 . ПМИД 19212399 . S2CID 205216390 .
- ^ Перейти обратно: а б с Фонг, Дэниел Ф.; Кейн, Томас С.; Калвер, Дэвид К. (ноябрь 1995 г.). «Рудистигиализация и потеря нефункциональных персонажей». Ежегодный обзор экологии и систематики . 26 : 249–268. doi : 10.1146/annurev.es.26.110195.001341 .
- ^ ЧжаоЛэй Чжан; Герштейн, Марк (август 2004 г.). «Масштабный анализ псевдогенов в геноме человека». Текущее мнение в области генетики и развития . 14 (4): 328–335. дои : 10.1016/j.где.2004.06.003 . ISSN 0959-437X . ПМИД 15261647 .
- ^ Джеффри, Уильям Р. (май – июнь 2005 г.). «Адаптивная эволюция дегенерации глаз у мексиканской слепой пещерной рыбы». Журнал наследственности . 96 (3): 185–196. CiteSeerX 10.1.1.572.6605 . doi : 10.1093/jhered/esi028 . ПМИД 15653557 .
- ^ Максвелл, Эрин Э.; Ларссон, Ханс CE (май 2007 г.). «Остеология и миология крыла эму ( Dromaius novaehollandiae ) и ее влияние на эволюцию рудиментарных структур». Журнал морфологии . 268 (5): 423–441. дои : 10.1002/jmor.10527 . ISSN 0362-2525 . ПМИД 17390336 . S2CID 12494187 .
- ^ ван дер Кои, Каспер Дж.; Швандер, Таня (ноябрь 2014 г.). «О судьбе сексуальных признаков при асексуальности» (PDF) . Биологические обзоры . 89 (4): 805–819. дои : 10.1111/brv.12078 . ISSN 1464-7931 . ПМИД 24443922 . S2CID 33644494 . Архивировано (PDF) из оригинала 23 июля 2015 года . Проверено 5 августа 2015 г.
- ^ Сильвестри, Энтони Р. младший; Сингх, Икбал (апрель 2003 г.). «Нерешенная проблема третьего моляра: будет ли людям лучше без него?» . Журнал Американской стоматологической ассоциации . 134 (4): 450–455. дои : 10.14219/jada.archive.2003.0194 . ПМИД 12733778 . Архивировано из оригинала 23 августа 2014 года.
- ^ Койн 2009 , с. 62
- ^ Дарвин 1872 , стр. 101, 103.
- ^ Грей 2007 , с. 66
- ^ Койн 2009 , стр. 85–86.
- ^ Стивенс 1982 , с. 87
- ^ Перейти обратно: а б Гулд 2002 , стр. 1235–1236.
- ^ Паллен, Марк Дж.; Мацке, Николас Дж. (октябрь 2006 г.). «От происхождения видов к происхождению бактериальных жгутиков» (PDF) . Обзоры природы по микробиологии (PDF) . 4 (10): 784–790. дои : 10.1038/nrmicro1493 . ISSN 1740-1526 . ПМИД 16953248 . S2CID 24057949 . Архивировано из оригинала (PDF) 26 декабря 2014 года . Проверено 25 декабря 2014 г.
- ^ Клементс, Эбигейл; Бурсак, Деян; Гацос, Ксения; и др. (15 сентября 2009 г.). «Уменьшаемая сложность митохондриальной молекулярной машины» . ПНАС . 106 (37): 15791–15795. Бибкод : 2009PNAS..10615791C . дои : 10.1073/pnas.0908264106 . ПМК 2747197 . ПМИД 19717453 .
- ^ Пятигорский и др. 1994 , стр. 241–250.
- ^ Вистоу, Грэм (август 1993 г.). «Кристаллины хрусталика: рекрутирование генов и эволюционный динамизм» . Тенденции биохимических наук . 18 (8): 301–306. дои : 10.1016/0968-0004(93)90041-К . ISSN 0968-0004 . ПМИД 8236445 .
- ^ Джонсон, Норман А.; Портер, Адам Х. (ноябрь 2001 г.). «К новому синтезу: популяционная генетика и эволюционная биология развития». Генетика . 112–113 (1): 45–58. дои : 10.1023/A:1013371201773 . ISSN 0016-6707 . ПМИД 11838782 . S2CID 1651351 .
- ^ Багуна, Хауме; Гарсия-Фернандес, Хорди (2003). «Эво-Дево: долгий и извилистый путь» . Международный журнал биологии развития . 47 (7–8): 705–713. ISSN 0214-6282 . ПМИД 14756346 . Архивировано из оригинала 28 ноября 2014 года.
- С любовью, Алан К. (март 2003 г.). «Эволюционная морфология, инновации и синтез эволюционной биологии и биологии развития» . Биология и философия . 18 (2): 309–345. дои : 10.1023/A:1023940220348 . S2CID 82307503 .
- ^ Аллин, Эдгар Ф. (декабрь 1975 г.). «Эволюция среднего уха млекопитающих». Журнал морфологии . 147 (4): 403–437. дои : 10.1002/jmor.1051470404 . ISSN 0362-2525 . ПМИД 1202224 . S2CID 25886311 .
- ^ Харрис, Мэтью П.; Хассо, Шон М.; Фергюсон, Марк У.Дж.; Фэллон, Джон Ф. (21 февраля 2006 г.). «Развитие зубов первого поколения архозавров у куриного мутанта» . Современная биология . 16 (4): 371–377. Бибкод : 2006CBio...16..371H . дои : 10.1016/j.cub.2005.12.047 . PMID 16488870 . S2CID 15733491 .
- ^ Кэрролл, Шон Б. (11 июля 2008 г.). «Эво-Дево и расширяющийся эволюционный синтез: генетическая теория морфологической эволюции» . Клетка . 134 (1): 25–36. дои : 10.1016/j.cell.2008.06.030 . ПМИД 18614008 . S2CID 2513041 .
- ^ Уэйд, Майкл Дж. (март 2007 г.). «Коэволюционная генетика экологических сообществ». Обзоры природы Генетика . 8 (3): 185–195. дои : 10.1038/nrg2031 . ПМИД 17279094 . S2CID 36705246 .
- ^ Геффини, Шана; Броди, Эдмунд Д. младший; Рубен, Питер К.; Броди, Эдмунд Д. III (23 августа 2002 г.). «Механизмы адаптации в гонке вооружений хищник-жертва: устойчивые к ТТХ натриевые каналы». Наука . 297 (5585): 1336–1339. Бибкод : 2002Sci...297.1336G . дои : 10.1126/science.1074310 . ПМИД 12193784 . S2CID 8816337 .
- Броди, Эдмунд Д. младший; Риденхур, Бенджамин Дж.; Броди, Эдмунд Д. III (октябрь 2002 г.). «Эволюционная реакция хищников на опасную добычу: горячие и холодные точки в географической мозаике коэволюции подвязочных змей и тритонов» . Эволюция . 56 (10): 2067–2082. doi : 10.1554/0014-3820(2002)056[2067:teropt]2.0.co;2 . ПМИД 12449493 . S2CID 8251443 .
- Кэрролл, Шон Б. (21 декабря 2009 г.). «Все, что не убивает некоторых животных, может сделать их смертельными» . Нью-Йорк Таймс . Нью-Йорк. Архивировано из оригинала 23 апреля 2015 года . Проверено 26 декабря 2014 г.
- ^ Сакс, Джоэл Л. (сентябрь 2006 г.). «Сотрудничество внутри и между видами». Журнал эволюционной биологии . 19 (5): 1415–1418, обсуждение 1426–1436. дои : 10.1111/j.1420-9101.2006.01152.x . ПМИД 16910971 . S2CID 4828678 .
- Новак, Мартин А. (8 декабря 2006 г.). «Пять правил эволюции сотрудничества» . Наука . 314 (5805): 1560–1563. Бибкод : 2006Sci...314.1560N . дои : 10.1126/science.1133755 . ПМК 3279745 . ПМИД 17158317 .
- ^ Пашковский, Ута (август 2006 г.). «Мутуализм и паразитизм: инь и янь растительных симбиозов». Современное мнение в области биологии растений . 9 (4): 364–370. Бибкод : 2006COPB....9..364P . дои : 10.1016/j.pbi.2006.05.008 . ISSN 1369-5266 . ПМИД 16713732 .
- ^ Хаус, Беттина; Фестер, Томас (май 2005 г.). «Молекулярная и клеточная биология арбускулярного микоризного симбиоза». Планта . 221 (2): 184–196. Бибкод : 2005Plant.221..184H . дои : 10.1007/s00425-004-1436-x . ПМИД 15871030 . S2CID 20082902 .
- ^ Бертрам, Джон С. (декабрь 2000 г.). «Молекулярная биология рака». Молекулярные аспекты медицины . 21 (6): 167–223. дои : 10.1016/S0098-2997(00)00007-8 . ПМИД 11173079 . S2CID 24155688 .
- ^ Рив, Х. Керн; Хёльдоблер, Берт (5 июня 2007 г.). «Появление суперорганизма посредством межгрупповой конкуренции» . ПНАС . 104 (23): 9736–9740. Бибкод : 2007PNAS..104.9736R . дои : 10.1073/pnas.0703466104 . ISSN 0027-8424 . ПМК 1887545 . ПМИД 17517608 .
- ^ Аксельрод, Роберт; Гамильтон, штат Вашингтон (27 марта 1981 г.). «Эволюция сотрудничества» . Наука . 211 (4489): 1390–1396. Бибкод : 1981Sci...211.1390A . дои : 10.1126/science.7466396 . ПМИД 7466396 .
- ^ Уилсон, Эдвард О.; Хёльдоблер, Берт (20 сентября 2005 г.). «Эусоциальность: происхождение и последствия» . ПНАС . 102 (38): 13367–1371. Бибкод : 2005PNAS..10213367W . дои : 10.1073/pnas.0505858102 . ПМЦ 1224642 . ПМИД 16157878 .
- ^ Перейти обратно: а б Гаврилец, Сергей (октябрь 2003 г.). «Перспектива: модели видообразования: чему мы научились за 40 лет?». Эволюция . 57 (10): 2197–2215. дои : 10.1554/02-727 . ПМИД 14628909 . S2CID 198158082 .
- ^ Перейти обратно: а б с де Кейрос, Кевин (3 мая 2005 г.). «Эрнст Майр и современная концепция вида» . ПНАС . 102 (Приложение 1): 6600–6607. Бибкод : 2005PNAS..102.6600D . дои : 10.1073/pnas.0502030102 . ПМЦ 1131873 . ПМИД 15851674 .
- ^ Перейти обратно: а б Эрешефски, Марк (декабрь 1992 г.). «Элиминативный плюрализм» . Философия науки . 59 (4): 671–690. дои : 10.1086/289701 . JSTOR 188136 . S2CID 224829314 .
- ^ Майр 1942 , с. 120
- ^ Фрейзер, Кристоф; Альм, Эрик Дж.; Польц, Мартин Ф.; и др. (6 февраля 2009 г.). «Проблема бактериальных видов: понимание генетического и экологического разнообразия». Наука . 323 (5915): 741–746. Бибкод : 2009Sci...323..741F . дои : 10.1126/science.1159388 . ПМИД 19197054 . S2CID 15763831 .
- ^ Коротко, Роджер Валентайн (октябрь 1975 г.). «Вклад мула в научную мысль». Журнал репродукции и фертильности. Приложение (23): 359–364. OCLC 1639439 . ПМИД 1107543 .
- ^ Гросс, Бриана Л.; Ризеберг, Лорен Х. (май – июнь 2005 г.). «Экологическая генетика гомоплоидного гибридного видообразования» . Журнал наследственности . 96 (3): 241–252. doi : 10.1093/jhered/esi026 . ISSN 0022-1503 . ПМК 2517139 . ПМИД 15618301 .
- ^ Берк, Джон М.; Арнольд, Майкл Л. (декабрь 2001 г.). «Генетика и приспособленность гибридов». Ежегодный обзор генетики . 35 : 31–52. дои : 10.1146/annurev.genet.35.102401.085719 . ISSN 0066-4197 . ПМИД 11700276 . S2CID 26683922 .
- ^ Вриенгук, Роберт К. (4 апреля 2006 г.). «Полиплоидные гибриды: множественное происхождение видов квакш» . Современная биология . 16 (7): Р245–Р247. Бибкод : 2006CBio...16.R245V . дои : 10.1016/j.cub.2006.03.005 . ISSN 0960-9822 . ПМИД 16581499 . S2CID 11657663 .
- ^ Райс, Уильям Р.; Хостерт, Эллен Э. (декабрь 1993 г.). «Лабораторные эксперименты по видообразованию: чему мы научились за 40 лет?». Эволюция . 47 (6): 1637–1653. дои : 10.1111/j.1558-5646.1993.tb01257.x . ISSN 0014-3820 . JSTOR 2410209 . ПМИД 28568007 . S2CID 42100751 .
- Джиггинс, Крис Д.; Уздечка, Джон Р. (март 2004 г.). «Видообразование яблочной личинки: смесь винтажей?». Тенденции в экологии и эволюции . 19 (3): 111–114. дои : 10.1016/j.tree.2003.12.008 . ISSN 0169-5347 . ПМИД 16701238 .
- Боксхорн, Джозеф (1 сентября 1995 г.). «Наблюдаемые случаи видообразования» . Архив TalkOrigins . Хьюстон, Техас: The TalkOrigins Foundation, Inc. Архивировано из оригинала 22 января 2009 года . Проверено 26 декабря 2008 г.
- Вайнберг, Джеймс Р.; Старчак, Виктория Р.; Йорг, Даниэле (август 1992 г.). «Доказательства быстрого видообразования после события основателя в лаборатории» . Эволюция . 46 (4): 1214–1220. дои : 10.2307/2409766 . ISSN 0014-3820 . JSTOR 2409766 . ПМИД 28564398 .
- ^ Херрел, Энтони; Хьюг, Кэтлин; Ванхойдонк, Бике; и др. (25 марта 2008 г.). «Быстрое крупномасштабное эволюционное расхождение в морфологии и продуктивности, связанное с использованием различных пищевых ресурсов» . ПНАС . 105 (12): 4792–4795. Бибкод : 2008PNAS..105.4792H . дои : 10.1073/pnas.0711998105 . ISSN 0027-8424 . ПМК 2290806 . ПМИД 18344323 .
- ^ Лосос, Джонатан Б.; Уорхельт, Кеннет И.; Шонер, Томас В. (1 мая 1997 г.). «Адаптивная дифференциация после экспериментальной колонизации островов у Anolis ящериц » . Природа . 387 (6628): 70–73. Бибкод : 1997Natur.387...70L . дои : 10.1038/387070a0 . ISSN 0028-0836 . S2CID 4242248 .
- ^ Хоскин, Конрад Дж.; Хиггл, Меган; Макдональд, Кейт Р.; Мориц, Крейг (27 октября 2005 г.). «Подкрепление приводит к быстрому аллопатрическому видообразованию» . Природа . 437 (7063): 1353–1356. Бибкод : 2005Natur.437.1353H . дои : 10.1038/nature04004 . ПМИД 16251964 . S2CID 4417281 .
- ^ Темплтон, Алан Р. (апрель 1980 г.). «Теория видообразования через принцип основателя» . Генетика . 94 (4): 1011–1038. дои : 10.1093/генетика/94.4.1011 . ПМЦ 1214177 . ПМИД 6777243 . Архивировано из оригинала 23 августа 2014 года . Проверено 29 декабря 2014 г.
- ^ Антонович, Янис (июль 2006 г.). «Эволюция в близко расположенных популяциях растений X: долгосрочное сохранение предрепродуктивной изоляции на границе шахты». Наследственность . 97 (1): 33–37. дои : 10.1038/sj.hdy.6800835 . ISSN 0018-067X . ПМИД 16639420 . S2CID 12291411 .
- ^ Носил, Патрик; Креспи, Бернард Дж.; Грис, Регина; Грис, Герхард (март 2007 г.). «Естественный отбор и расхождение в предпочтениях партнера во время видообразования» . Генетика . 129 (3): 309–327. дои : 10.1007/s10709-006-0013-6 . ISSN 0016-6707 . ПМИД 16900317 . S2CID 10808041 .
- ^ Саволайнен, Винсент ; Анстетт, Мари-Шарлотта; Лексер, Кристиан; и др. (11 мая 2006 г.). «Симпатрическое видообразование у пальм на океаническом острове» . Природа . 441 (7090): 210–213. Бибкод : 2006Natur.441..210S . дои : 10.1038/nature04566 . ISSN 0028-0836 . ПМИД 16467788 . S2CID 867216 .
- Барлуэнга, Марта; Стольтинг, Кай Н.; Зальцбургер, Вальтер; и др. (9 февраля 2006 г.). «Симпатрическое видообразование цихлид кратерного озера Никарагуа» . Природа . 439 (7077): 719–23. Бибкод : 2006Natur.439..719B . дои : 10.1038/nature04325 . ISSN 0028-0836 . ПМИД 16467837 . S2CID 3165729 . Архивировано из оригинала 30 июля 2022 года . Проверено 30 июля 2022 г.
- ^ Гаврилец, Сергей (21 марта 2006 г.). «Модель симпатрического видообразования Мейнарда Смита». Журнал теоретической биологии . 239 (2): 172–182. Бибкод : 2006JThBi.239..172G . дои : 10.1016/j.jtbi.2005.08.041 . ISSN 0022-5193 . ПМИД 16242727 .
- ^ Вуд, Трой Э.; Такебаяси, Наоки; Баркер, Майкл С.; и др. (18 августа 2009 г.). «Частота полиплоидного видообразования у сосудистых растений» . ПНАС . 106 (33): 13875–13879. Бибкод : 2009PNAS..10613875W . дои : 10.1073/pnas.0811575106 . ISSN 0027-8424 . ПМЦ 2728988 . ПМИД 19667210 .
- ^ Хегарти, Мэтью Дж.; Хискок, Саймон Дж. (20 мая 2008 г.). «Геномные ключи к эволюционному успеху полиплоидных растений» . Современная биология . 18 (10): Р435–Р444. Бибкод : 2008CBio...18.R435H . дои : 10.1016/j.cub.2008.03.043 . ISSN 0960-9822 . ПМИД 18492478 . S2CID 1584282 .
- ^ Якобссон, Маттиас; Хагенблад, Дженни; Таваре, Симон ; и др. (июнь 2006 г.). «Уникальное недавнее происхождение аллотетраплоидных видов Arabidopsis suecica : данные по маркерам ядерной ДНК» . Молекулярная биология и эволюция . 23 (6): 1217–1231. дои : 10.1093/molbev/msk006 . ПМИД 16549398 . Архивировано из оригинала 15 февраля 2022 года . Проверено 30 июля 2022 г.
- ^ Селл, Торбьёрн; Якобссон, Маттиас; Линд-Халден, Кристина; Халлден, Кристер (сентябрь 2003 г.). «ДНК хлоропластов указывает на единое происхождение аллотетраплоида Arabidopsis suecica » . Журнал эволюционной биологии . 16 (5): 1019–1029. дои : 10.1046/j.1420-9101.2003.00554.x . ПМИД 14635917 . S2CID 29281998 .
- ^ Бомблис, Кирстен ; Вайгель, Детлеф (декабрь 2007 г.). « Арабидопсис — модельный род для видообразования». Текущее мнение в области генетики и развития . 17 (6): 500–504. дои : 10.1016/j.где.2007.09.006 . ПМИД 18006296 .
- ^ Семон, Мари; Вулф, Кеннет Х. (декабрь 2007 г.). «Последствия дупликации генома». Текущее мнение в области генетики и развития . 17 (6): 505–512. дои : 10.1016/j.где.2007.09.007 . ПМИД 18006297 .
- ^ Элдридж и Гулд 1972 , стр. 82–115.
- ^ Бентон, Майкл Дж. (7 апреля 1995 г.). «Диверсификация и вымирание в истории жизни» . Наука . 268 (5207): 52–58. Бибкод : 1995Sci...268...52B . дои : 10.1126/science.7701342 . ISSN 0036-8075 . ПМИД 7701342 .
- ^ Рауп, Дэвид М. (28 марта 1986 г.). «Биологическое вымирание в истории Земли». Наука . 231 (4745): 1528–1533. Бибкод : 1986Sci...231.1528R . дои : 10.1126/science.11542058 . ПМИД 11542058 . S2CID 23012011 .
- ^ Авис, Джон К.; Хаббелл, Стивен П .; Аяла, Франсиско Дж. (12 августа 2008 г.). «В свете эволюции II: Биоразнообразие и вымирание» . ПНАС . 105 (Приложение 1): 11453–11457. Бибкод : 2008PNAS..10511453A . дои : 10.1073/pnas.0802504105 . ПМК 2556414 . ПМИД 18695213 .
- ^ Перейти обратно: а б с Рауп, Дэвид М. (19 июля 1994 г.). «Роль вымирания в эволюции» . ПНАС . 91 (15): 6758–6763. Бибкод : 1994PNAS...91.6758R . дои : 10.1073/pnas.91.15.6758 . ПМК 44280 . ПМИД 8041694 .
- ^ Новачек, Майкл Дж.; Клеланд, Эльза Э. (8 мая 2001 г.). «Текущее исчезновение биоразнообразия: сценарии смягчения последствий и восстановления» . ПНАС . 98 (10): 5466–5470. Бибкод : 2001PNAS...98.5466N . дои : 10.1073/pnas.091093698 . ISSN 0027-8424 . ПМК 33235 . ПМИД 11344295 .
- ^ Пимм, Стюарт ; Рэйвен, Питер ; Петерсон, Алан; и др. (18 июля 2006 г.). «Воздействие человека на темпы недавнего, настоящего и будущего вымирания птиц» . ПНАС . 103 (29): 10941–10946. Бибкод : 2006PNAS..10310941P . дои : 10.1073/pnas.0604181103 . ISSN 0027-8424 . ПМЦ 1544153 . ПМИД 16829570 .
- ^ Барноски, Энтони Д .; Кох, Пол Л.; Феранец, Роберт С.; и др. (1 октября 2004 г.). «Оценка причин позднеплейстоценовых вымираний на континентах». Наука . 306 (5693): 70–75. Бибкод : 2004Sci...306...70B . CiteSeerX 10.1.1.574.332 . дои : 10.1126/science.1101476 . ISSN 0036-8075 . ПМИД 15459379 . S2CID 36156087 .
- ^ Льюис, Оуэн Т. (29 января 2006 г.). «Изменение климата, кривые зависимости вида от площади и кризис вымирания» . Философские труды Королевского общества Б. 361 (1465): 163–171. дои : 10.1098/rstb.2005.1712 . ISSN 0962-8436 . ПМЦ 1831839 . ПМИД 16553315 .
- ^ Перейти обратно: а б Стернс и Стернс 1999 , с. Х
- ^ Перейти обратно: а б Новачек, Майкл Дж. (8 ноября 2014 г.). «Блестящее будущее предыстории» . Нью-Йорк Таймс . Нью-Йорк. ISSN 0362-4331 . Архивировано из оригинала 29 декабря 2014 года . Проверено 25 декабря 2014 г.
- ^ «Исследователи обнаружили, что на Земле может обитать 1 триллион видов» . Национальный научный фонд . Округ Арлингтон, Вирджиния. 2 мая 2016 г. Архивировано из оригинала 4 мая 2016 г. . Проверено 6 мая 2016 г.
- ^ Перейти обратно: а б с Кучера, Ульрих ; Никлас, Карл Дж. (июнь 2004 г.). «Современная теория биологической эволюции: расширенный синтез». Naturwissenschaften . 91 (6): 255–276. Бибкод : 2004NW.....91..255K . дои : 10.1007/s00114-004-0515-y . ISSN 1432-1904 . ПМИД 15241603 . S2CID 10731711 .
- ^ Яблонски, Дэвид (8 мая 2001 г.). «Уроки прошлого: эволюционные последствия массовых вымираний» . ПНАС . 98 (10): 5393–5398. Бибкод : 2001PNAS...98.5393J . дои : 10.1073/pnas.101092598 . ПМК 33224 . ПМИД 11344284 .
- ^ Булл, Джеймс Дж .; Вичман, Холли А. (ноябрь 2001 г.). «Прикладная эволюция». Ежегодный обзор экологии и систематики . 32 : 183–217. doi : 10.1146/annurev.ecolsys.32.081501.114020 . ISSN 1545-2069 .
- ^ Добли, Джон Ф.; Гаут, Брэндон С.; Смит, Брюс Д. (29 декабря 2006 г.). «Молекулярная генетика одомашнивания сельскохозяйственных культур» . Клетка . 127 (7): 1309–1321. дои : 10.1016/j.cell.2006.12.006 . ISSN 0092-8674 . ПМИД 17190597 . S2CID 278993 .
- ^ Йекель, Кристиан; Каст, Питер; Хилверт, Дональд (июнь 2008 г.). «Протеиновый дизайн путем направленной эволюции». Ежегодный обзор биофизики . 37 : 153–173. doi : 10.1146/annurev.biophys.37.032807.125832 . ISSN 1936-122Х . ПМИД 18573077 .
- ^ Махер, Брендан (8 апреля 2009 г.). «Эволюция: следующая топ-модель биологии?» . Природа . 458 (7239): 695–698. дои : 10.1038/458695а . ISSN 0028-0836 . ПМИД 19360058 . S2CID 41648315 .
- ^ Боровски, Ричард (8 января 2008 г.). «Восстановление зрения у слепых пещерных рыб» . Современная биология . 18 (1): С23–С24. Бибкод : 2008CBio...18..R23B . дои : 10.1016/j.cub.2007.11.023 . ISSN 0960-9822 . ПМИД 18177707 . S2CID 16967690 .
- ^ Гросс, Джошуа Б.; Боровски, Ричард; Табин, Клиффорд Дж. (2 января 2009 г.). Барш, Грегори С. (ред.). «Новая роль Mc1r в параллельной эволюции депигментации в независимых популяциях пещерной рыбы Astyanax mexicanus » . ПЛОС Генетика . 5 (1): e1000326. дои : 10.1371/journal.pgen.1000326 . ISSN 1553-7390 . ПМК 2603666 . ПМИД 19119422 .
- ^ Мерло, Лорен М.Ф.; Пеппер, Джон В.; Рид, Брайан Дж.; Мэйли, Карло К. (декабрь 2006 г.). «Рак как эволюционный и экологический процесс». Обзоры природы Рак . 6 (12): 924–935. дои : 10.1038/nrc2013 . ISSN 1474-175Х . ПМИД 17109012 . S2CID 8040576 .
- ^ Хлеб, Дабо; Вэйвэй Сюэ; Вэньци Чжан; и др. (октябрь 2012 г.). «Понимание механизма лекарственной устойчивости вируса гепатита С от NS3/4A до ITMN-191 из-за мутаций R155K, A156V, D168A/E: компьютерное исследование». Acta биохимии и биофизики (BBA) – общие предметы 1820 (10): 1526–1534. дои : 10.1016/j.bbagen.2012.06.001 . ISSN 0304-4165 . ПМИД 22698669 .
- ^ Вудфорд, Нил; Эллингтон, Мэтью Дж. (январь 2007 г.). «Появление устойчивости к антибиотикам путем мутации» . Клиническая микробиология и инфекции . 13 (1): 5–18. дои : 10.1111/j.1469-0691.2006.01492.x . ISSN 1198-743X . ПМИД 17184282 .
- ^ Лаббе, Пьеррик; Бертикат, Клэр; Бертомье, Арно; и др. (16 ноября 2007 г.). «Сорок лет беспорядочной эволюции устойчивости к инсектицидам у комаров Culex pipiens » . ПЛОС Генетика . 3 (11): е205. дои : 10.1371/journal.pgen.0030205 . ISSN 1553-7390 . ПМК 2077897 . ПМИД 18020711 .
- ^ Нив, Пол (октябрь 2007 г.). «Проблемы развития и управления устойчивостью к гербицидам: 50 лет после Харпера». Исследования сорняков . 47 (5): 365–369. Бибкод : 2007WeedR..47..365N . дои : 10.1111/j.1365-3180.2007.00581.x . ISSN 0043-1737 .
- ^ Родригес-Рохас, Александро; Родригес-Бельтран, Херонимо; Кос, Алехандро; Бласкес, Хесус (август 2013 г.). «Антибиотики и устойчивость к антибиотикам: ожесточенная борьба с эволюцией». Международный журнал медицинской микробиологии . 303 (6–7): 293–297. дои : 10.1016/j.ijmm.2013.02.004 . ISSN 1438-4221 . ПМИД 23517688 .
- ^ Шенк, Мартин Ф.; Сзендро, Иван Г.; Круг, Иоахим; де Виссер, генеральный директор Дж. Арьян (28 июня 2012 г.). «Количественная оценка адаптивного потенциала фермента устойчивости к антибиотикам» . ПЛОС Генетика . 8 (6): e1002783. дои : 10.1371/journal.pgen.1002783 . ISSN 1553-7390 . ПМЦ 3386231 . ПМИД 22761587 .
- ^ Прочтите, Эндрю Ф.; Линч, Пенелопа А.; Томас, Мэтью Б. (7 апреля 2009 г.). «Как создать устойчивые к эволюции инсектициды для борьбы с малярией» . ПЛОС Биология . 7 (4): e1000058. дои : 10.1371/journal.pbio.1000058 . ПМК 3279047 . ПМИД 19355786 .
- ^ Фрейзер, Алекс С. (18 января 1958 г.). «Анализ генетических моделей методом Монте-Карло» . Природа . 181 (4603): 208–209. Бибкод : 1958Natur.181..208F . дои : 10.1038/181208a0 . ISSN 0028-0836 . ПМИД 13504138 . S2CID 4211563 .
- ^ Рехенберг 1973 г.
- ^ Голландия 1975 г.
- ^ Коза 1992
- ^ Джамшиди, Миссури (15 августа 2003 г.). «Инструменты интеллектуального управления: нечеткие контроллеры, нейронные сети и генетические алгоритмы». Философские труды Королевского общества А. 361 (1809): 1781–1808. Бибкод : 2003RSPTA.361.1781J . дои : 10.1098/rsta.2003.1225 . ПМИД 12952685 . S2CID 34259612 .
- ^ «Возраст Земли» . Геологическая служба США . 9 июля 2007 года. Архивировано из оригинала 23 декабря 2005 года . Проверено 31 мая 2015 г.
- ^ Далримпл 2001 , стр. 205–221.
- ^ Манхеса, Жерар; Аллегре, Клод Ж .; Дюпреа, Бернар; Хамельн, Бруно (май 1980 г.). «Изотопное исследование свинца основных-ультраосновных слоистых комплексов: размышления о возрасте Земли и характеристиках примитивной мантии» . Письма о Земле и планетологии . 47 (3): 370–382. Бибкод : 1980E&PSL..47..370M . дои : 10.1016/0012-821X(80)90024-2 . ISSN 0012-821X .
- ^ Рэйвен и Джонсон 2002 , с. 68
- ^ Перейти обратно: а б Боренштейн, Сет (19 октября 2015 г.). «Намеки на жизнь на ранней Земле, которая считалась пустынной» . Возбуждайте . Йонкерс, Нью-Йорк: Интерактивная сеть Mindspark . Ассошиэйтед Пресс . Архивировано из оригинала 23 октября 2015 года . Проверено 8 октября 2018 г.
- ^ Белл, Элизабет А.; Бенике, Патрик; Харрисон, Т. Марк; Мао, Венди Л. (24 ноября 2015 г.). «Потенциально биогенный углерод сохранился в цирконе возрастом 4,1 миллиарда лет» (PDF) . ПНАС . 112 (47): 14518–14521. Бибкод : 2015PNAS..11214518B . дои : 10.1073/pnas.1517557112 . ISSN 0027-8424 . ПМЦ 4664351 . ПМИД 26483481 . Архивировано (PDF) из оригинала 6 ноября 2015 года . Проверено 30 декабря 2015 г.
- ^ Схоутен, Люси (20 октября 2015 г.). «Когда на Земле впервые появилась жизнь? Может быть, намного раньше, чем мы думали» . Христианский научный монитор . Бостон, Массачусетс: Издательское общество христианской науки . ISSN 0882-7729 . Архивировано из оригинала 22 марта 2016 года . Проверено 11 июля 2018 г.
- ^ Уэйд, Николас (25 июля 2016 г.). «Знакомьтесь, Лука, предок всего живого» . Нью-Йорк Таймс . Нью-Йорк. ISSN 0362-4331 . Архивировано из оригинала 28 июля 2016 года . Проверено 25 июля 2016 г.
- ^ МакКинни 1997 , с. 110
- ^ Мора, Камило; Титтенсор, Дерек П.; Адл, Сина; и др. (23 августа 2011 г.). «Сколько видов существует на Земле и в океане?» . ПЛОС Биология . 9 (8): e1001127. дои : 10.1371/journal.pbio.1001127 . ISSN 1545-7885 . ПМК 3160336 . ПМИД 21886479 .
- ^ Миллер и Спулман 2012 , с. 62
- ^ Чепмен 2009
- ^ Росков Ю.; Абукай, Л.; Оррелл, Т.; Николсон, Д.; и др., ред. (2016). «Каталог жизни видов 2000 и ITIS, ежегодный контрольный список за 2016 год» . Вид 2000 . Лейден, Нидерланды: Центр биоразнообразия Naturalis . ISSN 2405-884X . Архивировано из оригинала 12 ноября 2016 года . Проверено 6 ноября 2016 г.
- ^ Перето, Хули (март 2005 г.). «Споры о происхождении жизни» (PDF) . Международная микробиология . 8 (1): 23–31. ISSN 1139-6709 . ПМИД 15906258 . Архивировано из оригинала (PDF) 24 августа 2015 года.
- ^ Маршалл, Майкл (11 ноября 2020 г.). «Предположение Чарльза Дарвина о раннем периоде жизни, вероятно, было верным – в нескольких нацарапанных записках своему другу биолог Чарльз Дарвин выдвинул теорию о том, как зародилась жизнь. Мало того, что это, вероятно, было верным, его теория на столетие опередила свое время» . Новости Би-би-си . Архивировано из оригинала 11 ноября 2020 года . Проверено 11 ноября 2020 г.
- ^ Джойс, Джеральд Ф. (11 июля 2002 г.). «Древность эволюции на основе РНК». Природа . 418 (6894): 214–221. Бибкод : 2002Natur.418..214J . дои : 10.1038/418214a . ПМИД 12110897 . S2CID 4331004 .
- ^ Треворс, Джек Т.; Псеннер, Роланд (декабрь 2001 г.). «От самосборки жизни до современных бактерий: возможная роль наноклеток» . Обзоры микробиологии FEMS . 25 (5): 573–582. дои : 10.1111/j.1574-6976.2001.tb00592.x . ISSN 1574-6976 . ПМИД 11742692 .
- ^ Теобальд, Дуглас Л. (13 мая 2010 г.). «Формальная проверка теории универсального общего происхождения» . Природа . 465 (7295): 219–222. Бибкод : 2010Natur.465..219T . дои : 10.1038/nature09014 . ISSN 0028-0836 . ПМИД 20463738 . S2CID 4422345 .
- ^ Баптест, Эрик; Уолш, Дэвид А. (июнь 2005 г.). «Звучит ли «Кольцо жизни» правдиво?». Тенденции в микробиологии . 13 (6): 256–261. дои : 10.1016/j.tim.2005.03.012 . ISSN 0966-842X . ПМИД 15936656 .
- ^ Дарвин 1859 , с. 1
- ^ Дулиттл, У. Форд; Баптесте, Эрик (13 февраля 2007 г.). «Плюрализм моделей и гипотеза Древа жизни» . ПНАС . 104 (7): 2043–2049. Бибкод : 2007PNAS..104.2043D . дои : 10.1073/pnas.0610699104 . ISSN 0027-8424 . ЧВК 1892968 . ПМИД 17261804 .
- ^ Кунин, Виктор; Голдовский, Леон; Дарзентас, Никос; Узунис, Христос А. (июль 2005 г.). «Сеть жизни: Реконструкция микробной филогенетической сети» . Геномные исследования . 15 (7): 954–959. дои : 10.1101/гр.3666505 . ISSN 1088-9051 . ПМЦ 1172039 . ПМИД 15965028 .
- ^ Дарвин 1837 , с. 25
- ^ Яблонски, Дэвид (25 июня 1999 г.). «Будущее ископаемых пластинок». Наука . 284 (5423): 2114–2116. дои : 10.1126/science.284.5423.2114 . ISSN 0036-8075 . ПМИД 10381868 . S2CID 43388925 .
- ^ Мейсон, Стивен Ф. (6 сентября 1984 г.). «Истоки биомолекулярной рукости» . Природа . 311 (5981): 19–23. Бибкод : 1984Natur.311...19M . дои : 10.1038/311019a0 . ISSN 0028-0836 . ПМИД 6472461 . S2CID 103653 .
- ^ Вольф, Юрий И.; Рогозин Игорь Б.; Гришин Ник В.; Кунин, Евгений В. (1 сентября 2002 г.). «Геномные деревья и древо жизни» . Тенденции в генетике . 18 (9): 472–479. дои : 10.1016/S0168-9525(02)02744-0 . ISSN 0168-9525 . ПМИД 12175808 .
- ^ Варки, Аджит ; Альтейде, Таша К. (декабрь 2005 г.). «Сравнение геномов человека и шимпанзе: поиск иголок в стоге сена». Геномные исследования . 15 (12): 1746–1758. CiteSeerX 10.1.1.673.9212 . дои : 10.1101/гр.3737405 . ISSN 1088-9051 . ПМИД 16339373 .
- ^ Сиссарелли, Франческа Д.; Доркс, Тобиас; фон Меринг, Кристиан; и др. (3 марта 2006 г.). «На пути к автоматической реконструкции древа жизни с высоким разрешением» (PDF) . Наука . 311 (5765): 1283–1287. Бибкод : 2006Sci...311.1283C . CiteSeerX 10.1.1.381.9514 . дои : 10.1126/science.1123061 . ISSN 0036-8075 . ПМИД 16513982 . S2CID 1615592 . Архивировано (PDF) из оригинала 4 марта 2016 г.
- ^ Перейти обратно: а б Кавальер-Смит, Томас (29 июня 2006 г.). «Эволюция клеток и история Земли: застой и революция» . Философские труды Королевского общества Б. 361 (1470): 969–1006. дои : 10.1098/rstb.2006.1842 . ISSN 0962-8436 . ПМЦ 1578732 . ПМИД 16754610 .
- ^ Шопф, Дж. Уильям (29 июня 2006 г.). «Ископаемые свидетельства архейской жизни» . Философские труды Королевского общества Б. 361 (1470): 869–885. дои : 10.1098/rstb.2006.1834 . ПМЦ 1578735 . ПМИД 16754604 .
- Альтерманн, Владислав; Казмерчак, Юзеф (ноябрь 2003 г.). «Архейские микроокаменелости: переоценка ранней жизни на Земле» . Исследования в области микробиологии . 154 (9): 611–617. дои : 10.1016/j.resmic.2003.08.006 . ПМИД 14596897 .
- ^ Шопф, Дж. Уильям (19 июля 1994 г.). «Разные темпы, разные судьбы: темп и способ эволюции менялись от докембрия к фанерозою» . ПНАС . 91 (15): 6735–6742. Бибкод : 1994PNAS...91.6735S . дои : 10.1073/pnas.91.15.6735 . ПМЦ 44277 . ПМИД 8041691 .
- ^ Пул, Энтони М.; Пенни, Дэвид (январь 2007 г.). «Оценка гипотез происхождения эукариот». Биоэссе . 29 (1): 74–84. doi : 10.1002/bies.20516 . ISSN 0265-9247 . ПМИД 17187354 .
- ^ Дьялл, Сабрина Д.; Браун, Марк Т.; Джонсон, Патрисия Дж. (9 апреля 2004 г.). «Древние вторжения: от эндосимбионтов к органеллам». Наука . 304 (5668): 253–257. Бибкод : 2004Sci...304..253D . дои : 10.1126/science.1094884 . ПМИД 15073369 . S2CID 19424594 .
- ^ Мартин, Уильям (октябрь 2005 г.). «Недостающее звено между гидрогеносомами и митохондриями». Тенденции в микробиологии . 13 (10): 457–459. дои : 10.1016/j.tim.2005.08.005 . ПМИД 16109488 .
- ^ Ланг, Б. Франц; Грей, Майкл В.; Бургер, Гертрауд (декабрь 1999 г.). «Эволюция митохондриального генома и происхождение эукариот». Ежегодный обзор генетики . 33 : 351–397. дои : 10.1146/annurev.genet.33.1.351 . ISSN 0066-4197 . ПМИД 10690412 .
- Макфадден, Джеффри Ян (1 декабря 1999 г.). «Эндосимбиоз и эволюция растительной клетки». Современное мнение в области биологии растений . 2 (6): 513–519. Бибкод : 1999COPB....2..513M . дои : 10.1016/S1369-5266(99)00025-4 . ПМИД 10607659 .
- ^ Делонг, Эдвард Ф .; Пейс, Норман Р. (1 августа 2001 г.). «Экологическое разнообразие бактерий и архей» . Систематическая биология . 50 (4): 470–478. CiteSeerX 10.1.1.321.8828 . дои : 10.1080/106351501750435040 . ISSN 1063-5157 . ПМИД 12116647 .
- ^ Кайзер, Дейл (декабрь 2001 г.). «Построение многоклеточного организма». Ежегодный обзор генетики . 35 : 103–123. дои : 10.1146/annurev.genet.35.102401.090145 . ISSN 0066-4197 . ПМИД 11700279 . S2CID 18276422 .
- ^ Циммер, Карл (7 января 2016 г.). «Генетический переворот помог организмам перейти от одной клетки к множеству» . Нью-Йорк Таймс . Нью-Йорк. ISSN 0362-4331 . Архивировано из оригинала 7 января 2016 года . Проверено 7 января 2016 г.
- ^ Валентайн, Джеймс В .; Яблонски, Дэвид; Эрвин, Дуглас Х. (1 марта 1999 г.). «Ископаемые, молекулы и эмбрионы: новые взгляды на кембрийский взрыв» . Разработка . 126 (5): 851–859. дои : 10.1242/dev.126.5.851 . ISSN 0950-1991 . ПМИД 9927587 . Архивировано из оригинала 1 марта 2015 года . Проверено 30 декабря 2014 г.
- ^ Оно, Сусуму (январь 1997 г.). «Причина, а также последствия кембрийского взрыва в эволюции животных». Журнал молекулярной эволюции . 44 (Приложение 1): С23–С27. Бибкод : 1997JMolE..44S..23O . дои : 10.1007/PL00000055 . ISSN 0022-2844 . ПМИД 9071008 . S2CID 21879320 .
- Валентин, Джеймс В.; Яблонски, Дэвид (2003). «Морфологическая макроэволюция и развитие: палеонтологическая перспектива» . Международный журнал биологии развития . 47 (7–8): 517–522. ISSN 0214-6282 . ПМИД 14756327 . Архивировано из оригинала 24 октября 2014 года . Проверено 30 декабря 2014 г.
- ^ Уотерс, Элизабет Р. (декабрь 2003 г.). «Молекулярная адаптация и происхождение наземных растений». Молекулярная филогенетика и эволюция . 29 (3): 456–463. Бибкод : 2003MolPE..29..456W . дои : 10.1016/j.ympev.2003.07.018 . ISSN 1055-7903 . ПМИД 14615186 .
- ^ Мэйхью, Питер Дж. (август 2007 г.). «Почему существует так много видов насекомых? Перспективы окаменелостей и филогении» . Биологические обзоры . 82 (3): 425–454. дои : 10.1111/j.1469-185X.2007.00018.x . ISSN 1464-7931 . ПМИД 17624962 . S2CID 9356614 .
- ^ Кэрролл, Роберт Л. (май 2007 г.). «Палеозойское происхождение саламандр, лягушек и червяг» . Зоологический журнал Линнеевского общества . 150 (Приложение с1): 1–140. дои : 10.1111/j.1096-3642.2007.00246.x . ISSN 1096-3642 .
- ^ Уибл, Джон Р.; Ружье, Гильермо В.; Новачек, Майкл Дж.; Ашер, Роберт Дж. (21 июня 2007 г.). «Меловые эвтерии и лавразийское происхождение плацентарных млекопитающих вблизи границы K/T» . Природа . 447 (7147): 1003–1006. Бибкод : 2007Natur.447.1003W . дои : 10.1038/nature05854 . ISSN 0028-0836 . ПМИД 17581585 . S2CID 4334424 .
- ^ Уитмер, Лоуренс М. (28 июля 2011 г.). «Палеонтология: икона, сбитая с насеста». Природа . 475 (7357): 458–459. дои : 10.1038/475458a . ISSN 0028-0836 . ПМИД 21796198 . S2CID 205066360 .
- ^ Дарвин 1909 , с. 53
- ^ Кирк, Рэйвен и Шофилд 1983 , стр. 100–142, 280–321.
- ^ Лукреций
- ^ Седли, Дэвид (2003). «Лукреций и Новый Эмпедокл» (PDF) . Лидс Международные классические исследования . 2 (4). ISSN 1477-3643 . Архивировано из оригинала (PDF) 23 августа 2014 года . Проверено 25 ноября 2014 г.
- ^ Торри, Гарри Бил; Фелин, Фрэнсис (март 1937 г.). «Был ли Аристотель эволюционистом?» . Ежеквартальный обзор биологии . 12 (1): 1–18. дои : 10.1086/394520 . ISSN 0033-5770 . JSTOR 2808399 . S2CID 170831302 .
- ^ Халл, Дэвид Л. (декабрь 1967 г.). «Метафизика эволюции». Британский журнал истории науки . 3 (4). Кембридж : Издательство Кембриджского университета от имени Британского общества истории науки : 309–337. дои : 10.1017/S0007087400002892 . JSTOR 4024958 . S2CID 170328394 .
- ^ Мейсон 1962 , стр. 43–44.
- ^ Кирос, Теодрос. Исследования африканской политической мысли . 2001, стр. 55
- ^ Майр 1982 , стр. 256–257.
- ^ Ваггонер, Бен (7 июля 2000 г.). «Карл Линней (1707–1778)» . Эволюция (Онлайн-выставка). Беркли, Калифорния: Музей палеонтологии Калифорнийского университета . Архивировано из оригинала 30 апреля 2011 года . Проверено 11 февраля 2012 г.
- ^ Боулер 2003 , стр. 73–75.
- ^ «Эразм Дарвин (1731–1802)» . Эволюция (Онлайн-выставка). Беркли, Калифорния: Музей палеонтологии Калифорнийского университета. 4 октября 1995 года. Архивировано из оригинала 19 января 2012 года . Проверено 11 февраля 2012 г.
- ^ Ламарк 1809 г.
- ^ Перейти обратно: а б Нардон и Гренье 1991 , с. 162
- ^ Гизелин, Майкл Т. (сентябрь – октябрь 1994 г.). «Воображаемый Ламарк: взгляд на фальшивую «историю» в школьных учебниках» . Учебное письмо . OCLC 23228649 . Архивировано из оригинала 12 февраля 2008 года . Проверено 23 января 2008 г.
- ^ Яблонка, Ева ; Лэмб, Мэрион Дж. (август 2007 г.). «Краткая информация об эволюции в четырех измерениях». Поведенческие и мозговые науки . 30 (4): 353–365. дои : 10.1017/S0140525X07002221 . ISSN 0140-525X . ПМИД 18081952 . S2CID 15879804 .
- ^ Буркхардт и Смит, 1991 г.
- «Дарвин, ЧР, Лаббоку, Джону» . Дарвиновский заочный проект . Кембридж: Кембриджский университет . Архивировано из оригинала 15 декабря 2014 года . Проверено 1 декабря 2014 г. Письмо 2532 от 22 ноября 1859 г.
- ^ Саллоуэй, Фрэнк Дж. (июнь 2009 г.). «Почему Дарвин отверг разумный замысел». Журнал биологических наук . 34 (2): 173–183. дои : 10.1007/s12038-009-0020-8 . ISSN 0250-5991 . ПМИД 19550032 . S2CID 12289290 .
- ^ «Результаты поиска по запросу «происхождение с модификацией» - Полное собрание сочинений Чарльза Дарвина в Интернете» . Архивировано из оригинала 5 июня 2022 года . Проверено 30 июля 2022 г.
- ^ Трезвый, Эллиотт (16 июня 2009 г.). «Дарвин написал « Происхождение» задом наперед?» . ПНАС . 106 (Приложение 1): 10048–10055. Бибкод : 2009PNAS..10610048S . дои : 10.1073/pnas.0901109106 . ISSN 0027-8424 . ПМК 2702806 . ПМИД 19528655 .
- ^ Майр 2002 , с. 165
- ^ Боулер 2003 , стр. 145–146.
- ^ Сокал, Роберт Р .; Кровелло, Теодор Дж. (март – апрель 1970 г.). «Концепция биологических видов: критическая оценка». Американский натуралист . 104 (936): 127–153. дои : 10.1086/282646 . ISSN 0003-0147 . JSTOR 2459191 . S2CID 83528114 .
- ^ Дарвин, Чарльз ; Уоллес, Альфред (20 августа 1858 г.). «О склонности видов к образованию разновидностей и о сохранении разновидностей и видов естественными средствами отбора» . Журнал трудов Лондонского Линнеевского общества. Зоология . 3 (9): 45–62. дои : 10.1111/j.1096-3642.1858.tb02500.x . ISSN 1096-3642 . Архивировано из оригинала 14 июля 2007 года . Проверено 13 мая 2007 г.
- ^ Десмонд, Адриан Дж. (17 июля 2014 г.). «Томас Генри Хаксли» . Британская онлайн-энциклопедия . Чикаго, Иллинойс: Британская энциклопедия, Inc. Архивировано из оригинала 19 января 2015 года . Проверено 2 декабря 2014 г.
- ^ Ю. -С. Лю; ХМ Чжоу; МХ Чжи; XJ Ли; К.Л. Ван (сентябрь 2009 г.). «Вклад Дарвина в генетику». Журнал прикладной генетики . 50 (3): 177–184. дои : 10.1007/BF03195671 . ISSN 1234-1983 . ПМИД 19638672 . S2CID 19919317 .
- ^ Вейлинг, Франц (июль 1991 г.). «Историческое исследование: Иоганн Грегор Мендель 1822–1884». Американский журнал медицинской генетики . 40 (1): 1–25, обсуждение 26. doi : 10.1002/ajmg.1320400103 . ПМИД 1887835 .
- ^ Райт 1984 , с. 480
- ^ Источник 1971 г.
- ^ Стамхейс, Ида Х.; Мейер, Онно Г.; Зевенхейзен, Эрик Дж. А. (июнь 1999 г.). «Гюго де Врис о наследственности, 1889–1903: статистика, менделевские законы, пангены, мутации» . Исида . 90 (2): 238–267. дои : 10.1086/384323 . JSTOR 237050 . ПМИД 10439561 . S2CID 20200394 .
- ^ Перейти обратно: а б Боулер 1989 , стр. 307–318.
- ^ Левинсон 2019 .
- ^ Уотсон, Джей Ди ; Крик, FHC (25 апреля 1953 г.). «Молекулярная структура нуклеиновых кислот: структура нуклеиновой кислоты дезоксирибозы» (PDF) . Природа . 171 (4356): 737–738. Бибкод : 1953Natur.171..737W . дои : 10.1038/171737a0 . ISSN 0028-0836 . ПМИД 13054692 . S2CID 4253007 . Архивировано (PDF) из оригинала 23 августа 2014 года . Проверено 4 декабря 2014 г.
От нашего внимания не ускользнул тот факт, что специфическое спаривание, которое мы постулировали, сразу предполагает возможный механизм копирования генетического материала.
- ^ Хенниг 1999 , стр. 280.
- ^ Добжанский, Феодосий (март 1973 г.). «Ничто в биологии не имеет смысла, кроме как в свете эволюции» (PDF) . Американский учитель биологии . 35 (3): 125–129. CiteSeerX 10.1.1.324.2891 . дои : 10.2307/4444260 . JSTOR 4444260 . S2CID 207358177 . Архивировано из оригинала (PDF) 23 октября 2015 года.
- ^ Авис, Джон К .; Аяла, Франсиско Дж. (11 мая 2010 г.). «В свете эволюции IV: Состояние человека» (PDF) . ПНАС . 107 (Приложение 2): 8897–8901. дои : 10.1073/pnas.1003214107 . ISSN 0027-8424 . ПМК 3024015 . ПМИД 20460311 . Архивировано (PDF) из оригинала 23 августа 2014 года . Проверено 29 декабря 2014 г.
- ^ Даншен, Этьен; Шармантье, Энн; Шампанское, Фрэнсис А .; Месуди, Алекс; Пужоль, Бенуа; Бланше, Саймон (июнь 2011 г.). «За пределами ДНК: интеграция инклюзивного наследования в расширенную теорию эволюции». Обзоры природы Генетика . 12 (7): 475–486. дои : 10.1038/nrg3028 . ISSN 1471-0056 . ПМИД 21681209 . S2CID 8837202 .
- ^ Пильуччи и Мюллер, 2010 г.
- ^ Браун 2003 , стр. 376–379.
- ^ Обзор философских, религиозных и космологических противоречий см.: О научном и социальном восприятии эволюции в XIX и начале XX веков см.:
- Джонстон, Ян К. (1999). «Раздел третий: Истоки эволюционной теории» . ... И все же мы развиваемся: Справочник по ранней истории современной науки (3-е исправленное изд.). Нанаймо, Британская Колумбия: Факультет гуманитарных исследований Университетского колледжа Маласпины . Архивировано из оригинала 16 апреля 2016 года . Проверено 1 января 2015 г.
- Боулер 2003
- Цукеркандль, Эмиль (30 декабря 2006 г.). «Разумный замысел и биологическая сложность». Джин . 385 : 2–18. дои : 10.1016/j.gene.2006.03.025 . ISSN 0378-1119 . ПМИД 17011142 .
- ^ Росс, Маркус Р. (май 2005 г.). «Кто во что верит? Прояснение путаницы в отношении разумного замысла и креационизма молодой Земли» (PDF) . Журнал геонаучного образования . 53 (3): 319–323. Бибкод : 2005JGeEd..53..319R . CiteSeerX 10.1.1.404.1340 . дои : 10.5408/1089-9995-53.3.319 . ISSN 1089-9995 . S2CID 14208021 . Архивировано (PDF) из оригинала 11 мая 2008 г. Проверено 28 апреля 2008 г.
- ^ Хамид, Салман (12 декабря 2008 г.). «Наготове к исламскому креационизму» (PDF) . Наука . 322 (5908): 1637–1638. дои : 10.1126/science.1163672 . ISSN 0036-8075 . ПМИД 19074331 . S2CID 206515329 . Архивировано из оригинала (PDF) 10 ноября 2014 года.
- ^ Боулер 2003
- ^ Миллер, Джон Д.; Скотт, Юджини К.; Окамото, Синдзи (11 августа 2006 г.). «Общественное признание эволюции». Наука . 313 (5788): 765–766. дои : 10.1126/science.1126746 . ISSN 0036-8075 . ПМИД 16902112 . S2CID 152990938 .
- ^ Спергель, Дэвид Натаниэль ; Верде, Лисия; Пейрис, Хиранья В.; и др. (2003). «Наблюдения первокурсника Уилкинсона с помощью микроволнового зонда анизотропии (WMAP): определение космологических параметров». Серия дополнений к астрофизическому журналу . 148 (1): 175–194. arXiv : astro-ph/0302209 . Бибкод : 2003ApJS..148..175S . дои : 10.1086/377226 . S2CID 10794058 .
- ^ Уайльд, Саймон А.; Вэлли, Джон В.; Пек, Уильям Х.; Грэм, Колин М. (11 января 2001 г.). «Свидетельства обломочных цирконов о существовании континентальной коры и океанов на Земле 4,4 миллиарда лет назад» . Природа . 409 (6817): 175–178. Бибкод : 2001Natur.409..175W . дои : 10.1038/35051550 . ISSN 0028-0836 . ПМИД 11196637 . S2CID 4319774 .
- ^ Бранч, Гленн (март 2007 г.). «Понимание креационизма после Китцмиллера » . Бионаука . 57 (3): 278–284. дои : 10.1641/B570313 . ISSN 0006-3568 . S2CID 86665329 .
- ^ Сяосин Цзинь (март 2019 г.). «Перевод и трансмутация: происхождение видов в Китае». Британский журнал истории науки . 52 (1). Кембридж: Издательство Кембриджского университета от имени Британского общества истории науки: 117–141. дои : 10.1017/S0007087418000808 . ПМИД 30587253 . S2CID 58605626 .
Библиография
- Альтенберг, Ли (1995). «Рост генома и эволюция карты генотип-фенотип». В Банцхафе, Вольфганг; Экман, Фрэнк Х. (ред.). Эволюция и биовычисления: вычислительные модели эволюции . Конспекты лекций по информатике. Том. 899. Берлин; Нью-Йорк: Springer-Verlag Berlin Heidelberg . стр. 205–259. CiteSeerX 10.1.1.493.6534 . дои : 10.1007/3-540-59046-3_11 . ISBN 978-3-540-59046-0 . ISSN 0302-9743 . LCCN 95005970 . ОСЛК 32049812 .
- Бердселл, Джон А.; Уиллс, Кристофер (2003). «Эволюционное происхождение и поддержание половой рекомбинации: обзор современных моделей». В Макинтайре, Росс Дж.; Клегг, Майкл Т. (ред.). Эволюционная биология . Том. 33. Нью-Йорк: Springer Science+Business Media . ISBN 978-1-4419-3385-0 . ISSN 0071-3260 . OCLC 751583918 .
- Боулер, Питер Дж. (1989). Менделевская революция: появление наследственных концепций в современной науке и обществе . Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-3888-0 . LCCN 89030914 . OCLC 19322402 .
- Боулер, Питер Дж. (2003). Эволюция: История идеи (3-е полностью переработанное и расширенное изд.). Беркли, Калифорния: Издательство Калифорнийского университета . ISBN 978-0-520-23693-6 . LCCN 2002007569 . OCLC 49824702 .
- Браун, Джанет (2003). Чарльз Дарвин: Сила места . Том. 2. Лондон: Пимлико . ISBN 978-0-7126-6837-8 . LCCN 94006598 . OCLC 52327000 .
- Буркхардт, Фредерик ; Смит, Сидней, ред. (1991). Переписка Чарльза Дарвина . Том. 7 : 1858–1859. Кембридж: Издательство Кембриджского университета . ISBN 978-0-521-38564-0 . LCCN 84045347 . OCLC 185662993 .
- Кэрролл, Шон Б .; Гренье, Дженнифер К.; Уэзерби, Скотт Д. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных (2-е изд.). Молден, Массачусетс: Blackwell Publishing . ISBN 978-1-4051-1950-4 . LCCN 2003027991 . OCLC 53972564 .
- Чепмен, Артур Д. (2009). Численность живых видов в Австралии и мире (2-е изд.). Канберра: Департамент окружающей среды, водных ресурсов, наследия и искусства : Исследование австралийских биологических ресурсов . ISBN 978-0-642-56860-1 . OCLC 780539206 . Архивировано из оригинала 25 декабря 2016 года . Проверено 6 ноября 2016 г.
- Койн, Джерри А. (2009). Почему эволюция верна . Нью-Йорк: Викинг . ISBN 978-0-670-02053-9 . LCCN 2008033973 . OCLC 233549529 .
- Далримпл, Дж. Брент (2001). «Возраст Земли в двадцатом веке: проблема (в основном) решена». В Льюисе, CLE; Нелл, С.Дж. (ред.). Возраст Земли: с 4004 г. до н.э. по 2002 г. н.э. Специальное издание Геологического общества. Том. 190. стр. 205–221. Бибкод : 2001GSLSP.190..205D . дои : 10.1144/gsl.sp.2001.190.01.14 . ISBN 978-1-86239-093-5 . LCCN 2003464816 . ОСЛК 48570033 . S2CID 130092094 .
- Дарвин, Чарльз (1837). Блокнот «Б» . Записную книжку можно приобрести в онлайн-каталоге «Полное собрание сочинений Чарльза Дарвина», заархивированном 18 марта 2022 года на сайте Wayback Machine . Проверено 9 октября 2019 г.
- Дарвин, Чарльз (1859). О происхождении видов путем естественного отбора, или о сохранении избранных рас в борьбе за жизнь (1-е изд.). Лондон: Джон Мюррей . LCCN 06017473 . OCLC 741260650 . Книга доступна в онлайн-каталоге «Полное собрание сочинений Чарльза Дарвина», заархивированном 27 января 2015 года на сайте Wayback Machine . Проверено 21 ноября 2014 г.
- Дарвин, Чарльз (1872). Выражение эмоций у человека и животных . Лондон: Джон Мюррей. LCCN 04002793 . ОСЛК 1102785 .
- Дарвин, Фрэнсис , изд. (1909). Основы «Происхождения видов», очерк, написанный в 1842 году (PDF) . Кембридж: напечатано в University Press. LCCN 61057537 . OCLC 1184581 . Архивировано (PDF) из оригинала 4 марта 2016 года . Проверено 27 ноября 2014 г.
- Деннетт, Дэниел (1995). Опасная идея Дарвина: эволюция и смысл жизни . Нью-Йорк: Саймон и Шустер . ISBN 978-0-684-80290-9 . LCCN 94049158 . OCLC 31867409 .
- Добжанский, Феодосий (1968). «О некоторых фундаментальных понятиях дарвиновской биологии». У Добжанского Феодосия; Хехт, Макс К.; Стир, Уильям К. (ред.). Эволюционная биология (1-е изд.). Нью-Йорк: Appleton-Century-Crofts . стр. 1–34. дои : 10.1007/978-1-4684-8094-8_1 . ISBN 978-1-4684-8096-2 . OCLC 24875357 .
- Добжанский, Феодосий (1970). Генетика эволюционного процесса . Нью-Йорк: Издательство Колумбийского университета . ISBN 978-0-231-02837-0 . LCCN 72127363 . OCLC 97663 .
- Элдридж, Найлс ; Гулд, Стивен Джей (1972). «Периодическое равновесие: альтернатива филетическому градуализму». В Шопфе, Томас Дж. М. (ред.). Модели в палеобиологии . Сан-Франциско, Калифорния: Фриман, Купер. ISBN 978-0-87735-325-6 . LCCN 72078387 . OCLC 572084 .
- Элдридж, Найлз (1985). Временные рамки: переосмысление дарвиновской эволюции и теории прерывистого равновесия . Нью-Йорк: Саймон и Шустер . ISBN 978-0-671-49555-8 . LCCN 84023632 . OCLC 11443805 .
- Юэнс, Уоррен Дж. (2004). Математическая популяционная генетика . Междисциплинарная прикладная математика. Том. Я. Теоретическое введение (2-е изд.). Нью-Йорк: Springer-Verlag Нью-Йорк . ISBN 978-0-387-20191-7 . LCCN 2003065728 . OCLC 53231891 .
- Фишер, Рональд А. (1930). Генетическая теория естественного отбора . Оксфорд: Кларендон Пресс . ISBN 978-0-19-850440-5 . LCCN 30029177 . ОСЛК 18500548 .
- Футуйма, Дуглас Дж. (2004). «Плод Древа Жизни: взгляд на эволюцию и экологию». В Cracraft, Джоэл; Донохью, Майкл Дж. (ред.). Сборка Древа Жизни . Оксфорд; Нью-Йорк: Издательство Оксфордского университета . ISBN 978-0-19-517234-8 . LCCN 2003058012 . OCLC 61342697 . «Материалы симпозиума, проведенного в Американском музее естественной истории в Нью-Йорке, 2002 г.».
- Футуйма, Дуглас Дж. (2005). Эволюция . Сандерленд, Массачусетс: Sinauer Associates . ISBN 978-0-87893-187-3 . LCCN 2004029808 . OCLC 57311264 .
- Футуйма, Дуглас Дж.; Киркпатрик, Марк (2017). Эволюция (Четвертое изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-1-60535-605-1 . LCCN 2017000562 . OCLC 969439375 .
- Гулд, Стивен Джей (2002). Структура эволюционной теории . Кембридж, Массачусетс : Belknap Press издательства Гарвардского университета . ISBN 978-0-674-00613-3 . LCCN 2001043556 . OCLC 47869352 .
- Грей, Питер (2007). Психология (5-е изд.). Нью-Йорк: Worth Publishers . ISBN 978-0-7167-0617-5 . LCCN 2006921149 . OCLC 76872504 .
- Холл, Брайан К .; Халлгримссон, Бенедикт (2008). Эволюция Стрикбергера (4-е изд.). Садбери, Массачусетс: Издательство Jones and Bartlett. ISBN 978-0-7637-0066-9 . LCCN 2007008981 . OCLC 85814089 .
- Хенниг, Вилли (1999) [Первоначально опубликовано в 1966 г. (переиздано в 1979 г.); перевод из неопубликованной авторской редакции Grundzüge einer Theorie der phylogenetischen Systematik , опубликованной в 1950 году]. Филогенетическая систематика . Перевод Д. Дуайта Дэвиса и Райнера Зангерла; предисловие Донна Э. Розена, Гарета Нельсона и Колина Паттерсона (переиздание). Урбана, Иллинойс: Издательство Университета Иллинойса . ISBN 978-0-252-06814-0 . LCCN 78031969 . OCLC 722701473 .
- Холланд, Джон Х. (1975). Адаптация в природных и искусственных системах: вводный анализ с приложениями к биологии, управлению и искусственному интеллекту . Анн-Арбор, Мичиган: Издательство Мичиганского университета . ISBN 978-0-472-08460-9 . LCCN 74078988 . ОСЛК 1531617 .
- Кампуракис, Костас (2014). Понимание эволюции . Кембридж; Нью-Йорк: Издательство Кембриджского университета. ISBN 978-1-107-03491-4 . LCCN 2013034917 . OCLC 855585457 .
- Кирк, Джеффри ; Рэйвен, Джон ; Шофилд, Малькольм (1983). Досократические философы: критическая история с подборкой текстов (2-е изд.). Кембридж; Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-27455-5 . LCCN 82023505 . ОСЛК 9081712 .
- Коза, Джон Р. (1992). Генетическое программирование: о программировании компьютеров посредством естественного отбора . Сложные адаптивные системы. Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-11170-6 . LCCN 92025785 . OCLC 26263956 .
- Ламарк, Жан-Батист (1809). Зоологическая философия . Париж: Денту и автор. ОСЛК 2210044 . Зоологическая философия (1809 г.) в Интернет-архиве . Проверено 29 ноября 2014 г.
- Лейн, Дэвид Х. (1996). Феномен Тейяра: Пророк нового века (1-е изд.). Мейкон, Джорджия: Издательство Университета Мерсер . ISBN 978-0-86554-498-7 . LCCN 96008777 . ОСЛК 34710780 .
- Левинсон, Джин (2019). Переосмысление эволюции: революция, которая скрывается на виду . Хакенсак, Нью-Джерси: World Scientific . ISBN 978-1-78634-726-8 . LCCN 2019013762 . OCLC 1138095098 . Архивировано из оригинала 21 мая 2022 года . Проверено 30 июля 2022 г.
- Лукреций . «Книга V, строки 855–877» . Де Рерум Натура . Отредактировано и переведено Уильямом Эллери Леонардом (1916). Медфорд/Сомервилл, Массачусетс: Университет Тафтса . OCLC 33233743 . Архивировано из оригинала 4 сентября 2014 года . Проверено 25 ноября 2014 г. - из цифровой библиотеки Персея .
- Мейсон, Стивен Ф. (1962). История наук . Книги Кольера. Научная библиотека, CS9 (новая ред.). Нью-Йорк: Collier Books . LCCN 62003378 . OCLC 568032626 .
- Мейнард Смит, Джон (1978). Эволюция секса . Кембридж; Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-29302-0 . LCCN 77085689 . ОСЛК 3413793 .
- Мейнард Смит, Джон (1998). «Единицы отбора». В Боке, Грегори Р.; Гуд, Джейми А. (ред.). Пределы редукционизма в биологии . Симпозиумы Фонда Новартис. Том. 213. Чичестер ; Нью-Йорк: Джон Уайли и сыновья . стр. 203–221. дои : 10.1002/9780470515488.ch15 . ISBN 978-0-471-97770-4 . LCCN 98002779 . ОСЛК 38311600 . ПМИД 9653725 . «Материалы симпозиума по пределам редукционизма в биологии, состоявшегося в Фонде Novartis, Лондон, 13–15 мая 1997 г.».
- Майр, Эрнст (1942). Систематика и происхождение видов с точки зрения зоолога . Колумбийская биологическая серия. Том. 13. Нью-Йорк: Издательство Колумбийского университета. LCCN 43001098 . OCLC 766053 .
- Майр, Эрнст (1982). Рост биологической мысли: разнообразие, эволюция и наследование . Перевод Джона Рэя Э. Силка. Кембридж, Массачусетс: Belknap Press . ISBN 978-0-674-36445-5 . LCCN 81013204 . OCLC 7875904 .
- Майр, Эрнст (2002) [Первоначально опубликовано в 2001 году; Нью-Йорк: Основные книги ]. Что такое эволюция . Магистр наук. Лондон: Вайденфельд и Николсон . ISBN 978-0-297-60741-0 . LCCN 2001036562 . ОСЛК 248107061 .
- МакКинни, Майкл Л. (1997). «Как редким видам удается избежать вымирания? Палеонтологический взгляд». В Кунине, Уильям Э.; Гастон, Кевин Дж. (ред.). Биология редкости: причины и последствия редких и общих различий (1-е изд.). Лондон; Нью-Йорк: Чепмен и Холл . ISBN 978-0-412-63380-5 . LCCN 96071014 . OCLC 36442106 .
- Миллер, Дж. Тайлер; Спулман, Скотт Э. (2012). Наука об окружающей среде (14-е изд.). Бельмонт, Калифорния: Брукс/Коул . ISBN 978-1-111-98893-7 . LCCN 2011934330 . OCLC 741539226 . Архивировано из оригинала 2 мая 2019 года . Проверено 27 декабря 2014 г.
- Нардон, Пол; Гренье, Анн-Мари (1991). «Теория серийного эндосимбиоза и эволюция долгоносика: роль симбиоза». В Маргулисе, Линн; Фестер, Рене (ред.). Симбиоз как источник эволюционных инноваций: видообразование и морфогенез . Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-13269-5 . LCCN 90020439 . OCLC 22597587 . «На основе конференции, состоявшейся в Белладжио, Италия, 25–30 июня 1989 г.»
- Национальная академия наук ; Медицинский институт (2008). Наука, эволюция и креационизм . Вашингтон, округ Колумбия: Издательство Национальной академии. ISBN 978-0-309-10586-6 . LCCN 2007015904 . OCLC 123539346 . Проверено 22 ноября 2014 г.
- Одум, Юджин П. (1971). Основы экологии (3-е изд.). Филадельфия, Пенсильвания: Сондерс . ISBN 978-0-7216-6941-0 . LCCN 76081826 . OCLC 154846 .
- Окаша, Самир (2006). Эволюция и уровни отбора . Оксфорд; Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-926797-2 . LCCN 2006039679 . OCLC 70985413 .
- Панно, Джозеф (2005). Клетка: эволюция первого организма . Факты о файловой библиотеке. Нью-Йорк: факты в архиве . ISBN 978-0-8160-4946-2 . LCCN 2003025841 . OCLC 53901436 .
- Пятигорский, Иорам; Канторов, Марк; Гопал-Шривастава, Рашми; Томарев, Станислав И. (1994). «Привлечение ферментов и стрессовых белков в виде кристаллинов хрусталика». Ин Янссон, Бенгт; Йорнвалль, Ганс; Ридберг, Ульф; и др. (ред.). К молекулярной основе употребления и злоупотребления алкоголем . Эксперименты. Том. 71. Базель; Бостон: Birkhäuser Verlag . стр. 241–50. дои : 10.1007/978-3-0348-7330-7_24 . ISBN 978-3-7643-2940-2 . LCCN 94010167 . ОСЛК 30030941 . ПМИД 8032155 .
- Пильуччи, Массимо ; Мюллер, Герд Б. , ред. (2010). Эволюция, расширенный синтез . Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-51367-8 . LCCN 2009024587 . OCLC 804875316 . Архивировано из оригинала 18 сентября 2015 года.
- Провайн, Уильям Б. (1971). Истоки теоретической популяционной генетики . Чикагская история науки и медицины (2-е изд.). Чикаго, Иллинойс: Издательство Чикагского университета . ISBN 978-0-226-68464-2 . LCCN 2001027561 . OCLC 46660910 .
- Рэйвен, Питер Х .; Джонсон, Джордж Б. (2002). Биология (6-е изд.). Бостон, Массачусетс: МакГроу-Хилл . ISBN 978-0-07-112261-0 . LCCN 2001030052 . OCLC 45806501 .
- Рэй, Джон (1686). История растений [ История растений ]. Том. I. Лондон: Напечатано Мэри Кларк. LCCN agr11000774 . ОСЛК 2126030 .
- Речехберг, Инго (1973). эволюционная стратегия; Оптимизация технических систем согласно принципам биологической эволюции (кандидатская диссертация). Проблемата (на немецком языке). Том 15. Послесловие Манфреда Эйгена . Штутгарт-Бад-Каннштатт: Фромманн-Хольцбуг. ISBN 978-3-7728-0373-4 . LCCN 74320689 . ОСЛК 9020616 .
- Ридли, Марк (2004). Эволюция . Оксфорд: Блэквелл. ISBN 978-1-4051-0345-9 .
- Стернс, Беверли Петерсон; Стернс, Стивен К. (1999). Смотрю с края вымирания . Нью-Хейвен, Коннектикут: Издательство Йельского университета . ISBN 978-0-300-08469-6 . LCCN 98034087 . OCLC 803522914 .
- Стивенс, Энтони (1982). Архетип: естественная история личности . Лондон: Рутледж и Кеган Пол . ISBN 978-0-7100-0980-7 . LCCN 84672250 . OCLC 10458367 .
- Воэт, Дональд ; Воэт, Джудит Г .; Пратт, Шарлотта В. (2016). Основы биохимии: жизнь на молекулярном уровне (Пятое изд.). Хобокен, Нью-Джерси : John Wiley & Sons . ISBN 978-1-118-91840-1 . LCCN 2016002847 . OCLC 939245154 .
- Райт, Сьюэлл (1984). Генетические и биометрические основы . Эволюция и генетика популяций. Том. 1. Чикаго, Иллинойс: Издательство Чикагского университета. ISBN 978-0-226-91038-3 . LCCN 67025533 . OCLC 246124737 .
Дальнейшее чтение
- Вводное чтение
- Барретт, Пол Х.; Вайншанк, Дональд Дж.; Готтлбер, Тимоти Т., ред. (1981). Согласие с дарвиновским происхождением видов, первое издание . Итака, Нью-Йорк: Издательство Корнельского университета . ISBN 978-0-8014-1319-3 . LCCN 80066893 . OCLC 610057960 .
- Кэрролл, Шон Б. (2005). Бесконечные самые красивые формы: новая наука Эво Дево и создание царства животных . иллюстрации Джейми В. Кэрролла, Джоша П. Клэйсса, Линн М. Олдс (1-е изд.). Нью-Йорк: WW Norton & Company. ISBN 978-0-393-06016-4 . LCCN 2004029388 . OCLC 57316841 .
- Чарльзуорт, Брайан ; Чарльзуорт, Дебора (2003). Эволюция: очень краткое введение . Очень короткие вступления. Оксфорд; Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-280251-4 . LCCN 2003272247 . OCLC 51668497 .
- Гулд, Стивен Джей (1989). Замечательная жизнь: сланцы Берджесс и природа истории (1-е изд.). Нью-Йорк: WW Norton & Company. ISBN 978-0-393-02705-1 . LCCN 88037469 . OCLC 18983518 .
- Джонс, Стив (1999). Почти как кит: обновлено происхождение видов . Лондон; Нью-Йорк: Даблдей . ISBN 978-0-385-40985-8 . LCCN 2002391059 . OCLC 41420544 .
- —— (2000). Призрак Дарвина: Обновленное происхождение видов (1-е изд.). Нью-Йорк: Рэндом Хаус . ISBN 978-0-375-50103-6 . LCCN 99053246 . OCLC 42690131 . Американская версия.
- Мадер, Сильвия С. (2007). Биология . Значительный вклад Мюррея П. Пендарвиса (9-е изд.). Бостон, Массачусетс: Высшее образование Макгроу-Хилла . ISBN 978-0-07-246463-4 . LCCN 2005027781 . OCLC 61748307 .
- Мейнард Смит, Джон (1993). Теория эволюции (изд. Canto). Кембридж; Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-45128-4 . LCCN 93020358 . OCLC 27676642 .
- Паллен, Марк Дж. (2009). Примерное руководство по эволюции . Грубые руководства Справочные руководства. Лондон; Нью-Йорк: Грубые гиды . ISBN 978-1-85828-946-5 . LCCN 2009288090 . OCLC 233547316 .
- Продвинутое чтение
- Бартон, Николас Х .; Бриггс, Дерек Э.Г .; Эйзен, Джонатан А .; и др. (2007). Эволюция . Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. ISBN 978-0-87969-684-9 . LCCN 2007010767 . ОСЛК 86090399 .
- Койн, Джерри А.; Орр, Х. Аллен (2004). Видообразование . Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-089-0 . LCCN 2004009505 . OCLC 55078441 .
- Бергстрем, Карл Т .; Дугаткин, Ли Алан (2012). Эволюция (1-е изд.). Нью-Йорк: WW Norton & Company. ISBN 978-0-393-91341-5 . LCCN 2011036572 . OCLC 729341924 .
- Холл, Брайан К.; Олсон, Венди, ред. (2003). Ключевые слова и концепции эволюционной биологии развития . Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-00904-2 . LCCN 2002192201 . OCLC 50761342 .
- Кауфман, Стюарт А. (1993). Истоки порядка: самоорганизация и отбор в эволюции . Нью-Йорк; Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-507951-7 . LCCN 91011148 . OCLC 895048122 .
- Мейнард Смит, Джон; Сатмари, Эорс (1995). Основные переходы в эволюции . Оксфорд; Нью-Йорк: WH Freeman Spektrum. ISBN 978-0-7167-4525-9 . LCCN 94026965 . OCLC 30894392 .
- Майр, Эрнст (2001). Что такое эволюция . Нью-Йорк: Основные книги. ISBN 978-0-465-04426-9 . LCCN 2001036562 . OCLC 47443814 .
- Минелли, Алессандро (2009). Формы становления: эволюционная биология развития . Перевод Марка Эпштейна. Принстон, Нью-Джерси; Оксфорд: Издательство Принстонского университета . ISBN 978-0-691-13568-7 . LCCN 2008028825 . ОСЛК 233030259 .
Внешние ссылки
- Общая информация
- «Эволюция» в программе «В наше время » на BBC
- «Эволюционные ресурсы национальных академий» . Вашингтон, округ Колумбия: Национальная академия наук . Проверено 30 мая 2011 г.
- «Понимание эволюции: ваш универсальный ресурс для информации об эволюции» . Беркли, Калифорния: Калифорнийский университет, Беркли . Проверено 30 мая 2011 г.
- «Эволюция эволюции – 150 лет Дарвина «О происхождении видов» » . Округ Арлингтон, Вирджиния: Национальный научный фонд . Архивировано из оригинала 19 мая 2011 года . Проверено 30 мая 2011 г.
- «Интерактивная временная шкала эволюции человека» . Смитсоновский институт , Национальный музей естественной истории . 28 января 2010 года . Проверено 14 июля 2018 г. Требуется Adobe Flash.
- « История эволюции в Соединенных Штатах ». Салон . Проверено 24 августа 2021 г.
- Видео (1980; анимация «Космос»; 8:01): «Эволюция» - Карл Саган на YouTube
- Эксперименты
- Ленски, Ричард Э. «Экспериментальная эволюция» . Ист-Лансинг, Мичиган: Университет штата Мичиган . Проверено 31 июля 2013 г.
- Честейн, Эрик; Ливнат, Ади; Пападимитриу, Христос ; Вазирани, Умеш (22 июля 2014 г.). «Алгоритмы, игры и эволюция» . ПНАС . 111 (29): 10620–10623. Бибкод : 2014PNAS..11110620C . дои : 10.1073/pnas.1406556111 . ISSN 0027-8424 . ПМЦ 4115542 . ПМИД 24979793 .
- Онлайн лекции
- «Серия лекций «Эволюция имеет значение» . Портал онлайн-обучения Гарварда . Кембридж, Массачусетс: Гарвардский университет . Архивировано из оригинала 18 декабря 2017 года . Проверено 15 июля 2018 г.
- Стернс, Стивен К. «EEB 122: Принципы эволюции, экологии и поведения» . Открытые курсы Йельского университета . Нью-Хейвен, Коннектикут: Йельский университет . Архивировано из оригинала 1 декабря 2017 года . Проверено 14 июля 2018 г.