В пределах
| |||
| |||
| Имена | |||
|---|---|---|---|
| Название ИЮПАК
D -рибоза
| |||
| Систематическое название ИЮПАК
( 2R ,3R , 4S , 5R ) -5-(гидроксиметил)оксолан-2,3,4-триол | |||
| Другие имена
г -рибоза
| |||
| Идентификаторы | |||
3D model ( JSmol )
|
| ||
| ХЭМБЛ | |||
| ХимическийПаук |
| ||
| Лекарственный Банк | |||
| Номер ЕС |
| ||
ПабХим CID
|
|||
| НЕКОТОРЫЙ | |||
| Характеристики [ 1 ] [ 2 ] | |||
| С 5 Н 10 О 5 | |||
| Молярная масса | 150.13 | ||
| Появление | Белый твердый | ||
| Температура плавления | 95 ° C (203 ° F; 368 К) | ||
| 100 г/л (25 °C, 77 °F) | |||
Хиральное вращение ([α] D )
|
−21,5° (Н 2 О) | ||
| Родственные соединения | |||
Родственные альдопентозы
|
арабиноза Ксилоза Ликсоза | ||
Родственные соединения
|
Дезоксирибоза | ||
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа).
| |||

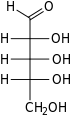

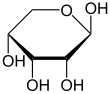

Рибоза представляет собой простой сахар и углевод с молекулярной формулой C 5 H 10 O 5 и линейной формой состава H-(C=O)-(CHOH) 4 -H. Встречающаяся в природе форма, d -рибоза — компонент рибонуклеотидов, из построена РНК , поэтому это соединение необходимо для кодирования , декодирования , регуляции и экспрессии генов которых . Он имеет структурный аналог дезоксирибозу , которая является столь же важным компонентом ДНК . L -рибоза — это ненатуральный сахар, который впервые был получен Эмилем Фишером и Оскаром Пилоти в 1891 году. [ 3 ] Лишь в 1909 году Фебус Левен и Уолтер Джейкобс осознали, что d -рибоза была природным продуктом , энантиомером продукта Фишера и Пилоти и важным компонентом нуклеиновых кислот . [ 4 ] [ 5 ] [ 6 ] Фишер выбрал название «рибоза», поскольку оно представляет собой частичную перегруппировку названия другого сахара, арабинозы которого является , эпимером 2'-углерод; оба названия также относятся к гуммиарабику , из которого впервые была выделена арабиноза и из которой были приготовлены л- рибоза . [ 6 ] [ 7 ]

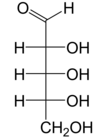

Справа: проекция Фишера форм открытой цепи д - и л - рибоза
Как и большинство сахаров, рибоза существует в виде смеси циклических форм, находящихся в равновесии с ее линейной формой, и они легко взаимопревращаются, особенно в водном растворе . [ 8 ] Название «рибоза» используется в биохимии и биологии для обозначения всех этих форм, хотя при необходимости для каждой используются более конкретные названия. В своей линейной форме рибозу можно рассматривать как пентозный сахар со всеми гидроксильными функциональными группами, расположенными на одной стороне в проекции Фишера . d -Рибоза имеет эти гидроксильные группы с правой стороны и связана с систематическим названием (2R , 3R , 4R ) -2,3,4,5-тетрагидроксипентаналь, [ 9 ] пока У l -рибозы гидроксильные группы расположены слева в проекции Фишера. Циклизация рибозы происходит посредством образования полуацеталя из-за атаки на альдегид гидроксильной группы C4' с образованием фуранозной формы или гидроксильной группы C5' с образованием формы пиранозы . В каждом случае есть два возможных геометрических результата, названных α- и β- и известных как аномеры , в зависимости от стереохимии полуацетального атома углерода («аномерный углерод»). При комнатной температуре около 76% d -рибоза присутствует в формах пиранозы. [ 8 ] : 228 (а:б = 1:2) [ 10 ] и 24% в фуранозных формах [ 8 ] : 228 (а:б = 1:3), [ 10 ] присутствует только около 0,1% линейной формы. [ 11 ] [ 12 ]
Рибонуклеозиды аденозин , цитидин , гуанозин и уридин являются производными β- д -рибофураноза. Метаболически важные виды, которые включают фосфорилированную рибозу, включают АДФ , АТФ , кофермент А , [ 8 ] : 228–229 и НАДН . цАМФ и цГМФ служат вторичными мессенджерами в некоторых сигнальных путях, а также являются производными рибозы. Рибозный фрагмент присутствует в некоторых фармацевтических агентах, включая антибиотики неомицин и паромомицин . [ 10 ]
Синтез и источники
[ редактировать ]Рибоза в виде ее 5-фосфатного эфира обычно образуется из глюкозы пентозофосфатным путем . По крайней мере, у некоторых архей были идентифицированы альтернативные пути. [ 13 ]
Рибозу можно синтезировать химическим путем, но коммерческое производство основано на ферментации глюкозы. Используя генетически модифицированные штаммы B. subtilis , из 200 г глюкозы можно получить 90 г/л рибозы. Преобразование влечет за собой посредничество глюконата и рибулозы. [ 14 ]
Рибоза была обнаружена в метеоритах . [ 15 ] [ 16 ]
Структура
[ редактировать ]Рибоза представляет собой альдопентозу (моносахарид, содержащий пять атомов углерода , который в форме открытой цепи имеет альдегидную функциональную группу на одном конце). В общепринятой схеме нумерации моносахаридов атомы углерода нумеруются от C1' (в альдегидной группе) до C5'. Производное дезоксирибозы , обнаруженное в ДНК, отличается от рибозы наличием атома водорода вместо гидроксильной группы при С2'. Эта гидроксильная группа выполняет функцию сплайсинга РНК .
" д -" в названии d -рибоза относится к стереохимии хирального атома углерода , наиболее удаленного от альдегидной группы (C4'). В d -рибоза, как и все d -сахара, этот атом углерода имеет ту же конфигурацию, что и в д -глицеральдегид .
-
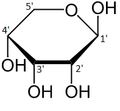
 а- d -рибопираноза
а- d -рибопираноза -
 б- d -рибопираноза
б- d -рибопираноза -
 а- d -рибофураноза
а- d -рибофураноза -
 б- d -рибофураноза
б- d -рибофураноза




Относительное содержание форм рибозы в растворе: β- d- рибопираноза (59%), α- d -рибопираноза (20%), β- d -рибофураноза (13%), α- d -рибофураноза (7%) и открытая цепь (0,1%). [ 11 ]
Для остатков рибозы в нуклеозидах и нуклеотидах торсионные углы вращения, охватывающие связи, влияют на конфигурацию соответствующего нуклеозида и нуклеотида. Вторичная структура нуклеиновой кислоты определяется поворотом ее 7 торсионных углов . [ 17 ] Наличие большого количества углов скручивания обеспечивает большую гибкость.
В рибозах с закрытыми кольцами наблюдаемая гибкость, упомянутая выше, не наблюдается, поскольку кольцевой цикл накладывает ограничение на количество возможных углов скручивания в структуре. [ 17 ] Конформеры рибоз закрытой формы различаются тем, как одиночный кислород в молекуле расположен относительно азотистого основания (также известного как азотистое основание или просто основание), присоединенного к рибозе. Если углерод обращен к основанию, то рибоза обозначается как эндо. Если углерод обращен от основания, то рибоза обозначается как экзо. Если к 2'-углероду рибозы замкнутого цикла присоединена молекула кислорода, то экзоподтверждение более стабильно, поскольку оно уменьшает взаимодействие кислорода с основанием. [ 17 ] Разница сама по себе весьма невелика, но если рассматривать всю цепочку РНК, то небольшая разница будет иметь существенное значение.
- Некоторые складчатые конфигурации рибозы
-
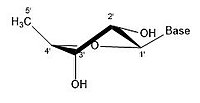 2' эндо
2' эндо -
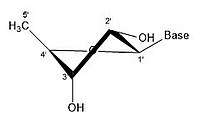 2' эндо 3' экзо
2' эндо 3' экзо -
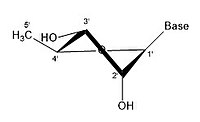 3' эндо 2' экзо
3' эндо 2' экзо -
 3' эндо
3' эндо




Молекулу рибозы обычно изображают на бумаге как плоскую молекулу. Несмотря на это, он обычно неплоский по своей природе. Даже между атомами водорода множество компонентов молекулы рибозы вызывают стерические препятствия и напряжения между ними. Чтобы уменьшить это скопление и напряжение кольца , кольцо сморщивается, то есть становится неплоским. [ 18 ] Это сморщивание достигается за счет смещения атома из плоскости, снятия напряжения и получения более стабильной конформации. [ 17 ] Сморщивание, иначе известное как конформация сахарного кольца (в частности, сахара рибозы), можно описать амплитудой складки, а также углом псевдовращения . Угол псевдоповорота можно описать как диапазон «север (N)» или «юг (S)». Хотя оба диапазона встречаются в виде двойных спиралей, северный диапазон обычно связан с РНК и А-формой ДНК . Напротив, южный диапазон связан с ДНК формы B. Z-ДНК содержит сахара как в северном, так и в южном диапазоне. [ 19 ] Когда смещается только один атом, это называется складкой «конверта». Когда два атома смещаются, это называется «закрученной» складкой в связи с зигзагообразной ориентацией. [ 20 ] В «эндо» складке основное смещение атомов происходит на β-грани, той же стороне, что и связь C4’-C5’ и основание. В «экзо»-складке основное смещение атомов происходит на α-грани, на противоположной стороне кольца. Основными формами рибозы являются 3'-эндо-складка (обычно используемая в РНК и А-форме ДНК) и 2'-эндо-складка (обычно используемая в B-форме ДНК). [ 21 ] Эти кольцевые складки образуются в результате изменения углов скручивания колец; существует бесконечное количество комбинаций углов, поэтому существует бесконечное количество мобильных конформаций складок, каждая из которых разделена несопоставимыми энергиями активации.
Функции
[ редактировать ]АТФ является производным рибозы; он содержит одну рибозу, три фосфатные группы и адениновое основание. АТФ создается во время клеточного дыхания из аденозиндифосфата (АТФ с одной фосфатной группой меньше).
Сигнальные пути
[ редактировать ]Рибоза является строительным блоком вторичных сигнальных молекул, таких как циклический аденозинмонофосфат (цАМФ), который образуется из АТФ. Одним из конкретных случаев использования цАМФ является цАМФ-зависимые сигнальные пути . активирует либо стимулирующий, либо ингибирующий гормональный рецептор В сигнальных путях цАМФ сигнальная молекула . Эти рецепторы связаны со стимулирующим или тормозным регулятивным G-белком . Когда активируется стимулирующий G-белок, аденилатциклаза катализирует АТФ в цАМФ с помощью магния. 2+ или Мн 2+ . цАМФ, вторичный мессенджер, затем активирует протеинкиназу А , фермент, регулирующий клеточный метаболизм . Протеинкиназа А регулирует метаболические ферменты путем фосфорилирования , которое вызывает изменения в клетке в зависимости от исходной сигнальной молекулы. Обратное происходит, когда активируется ингибирующий G-белок; G-белок ингибирует аденилатциклазу, и АТФ не превращается в цАМФ.

Метаболизм
[ редактировать ]Рибозу называют «молекулярной валютой» из-за ее участия во внутриклеточной передаче энергии. [ нужна ссылка ] Например, никотинамидадениндинуклеотид (НАД), флавинадениндинуклеотид (ФАД) и никотинамидадениндинуклеотидфосфат (НАДФ) содержат d -рибофуранозный фрагмент . Каждый из них может быть получен из d -рибоза после ее превращения в d- рибозо-5-фосфат под действием фермента рибокиназы . [ 22 ] [ 23 ] НАД, ФАД и НАДФ действуют как акцепторы электронов в биохимических окислительно-восстановительных реакциях в основных метаболических путях, включая гликолиз , цикл лимонной кислоты , ферментацию и цепь транспорта электронов .

Биосинтез нуклеотидов
[ редактировать ]Нуклеотиды синтезируются путем утилизации или синтеза de novo . [ 24 ] Для спасения нуклеотидов используются фрагменты ранее созданных нуклеотидов и повторно синтезируются их для использования в будущем. аминокислоты, диоксид углерода, производные фолиевой кислоты и фосфорибозилпирофосфат (PRPP). In de novo для синтеза нуклеотидов используются [ 24 ] Как de novo, так и для спасения требуется PRPP, который синтезируется из АТФ и рибозо-5-фосфата ферментом, называемым PRPP-синтетазой . [ 24 ]
Модификации
[ редактировать ]Модификации в природе
[ редактировать ]Рибокиназа катализирует превращение d -рибоза d- рибозо-5-фосфат . После преобразования, d- рибозо-5-фосфат доступен для производства аминокислот триптофана и гистидина или для использования в пентозофосфатном пути . Поглощение d -рибоза составляет 88–100% в тонком кишечнике (до 200 мг/кг·ч). [ 25 ]
Одна важная модификация происходит в положении C2' молекулы рибозы. При добавлении О-алкильной группы ядерная устойчивость РНК увеличивается из-за дополнительных стабилизирующих сил. Эти силы стабилизируются за счет увеличения внутримолекулярной водородной связи и увеличения стабильности гликозидной связи . [ 26 ] результате повышение резистентности приводит к увеличению периода полураспада миРНК Возникающее в и потенциального терапевтического потенциала в клетках и животных. [ 27 ] Метилирование рибозы в определенных участках коррелирует со снижением иммунной стимуляции. [ 28 ]
Синтетические модификации
[ редактировать ]Наряду с фосфорилированием молекулы рибофуранозы могут обменивать кислород на селен и серу с образованием аналогичных сахаров, которые различаются только в 4'-положении. Эти производные более липофильны, чем исходная молекула. Повышенная липофильность делает эти виды более подходящими для использования в таких методах, как ПЦР , РНК-аптамеров постмодификация , антисмысловая технология и для фазирования рентгеновских кристаллографических данных. [ 27 ]
Подобно природным 2'-модификациям, синтетическая модификация рибозы включает добавление фтора в 2'-положение. Эта фторированная рибоза действует аналогично метилированной рибозе, поскольку она способна подавлять иммунную стимуляцию в зависимости от местоположения рибозы в цепи ДНК. [ 26 ] Большая разница между метилированием и фторированием заключается в том, что последнее происходит только посредством синтетических модификаций. Добавление фтора приводит к усилению стабилизации гликозидной связи и увеличению внутримолекулярных водородных связей. [ 26 ]
Медицинское использование
[ редактировать ]d -рибозу было предложено использовать при лечении застойной сердечной недостаточности. [ 29 ] (а также других форм заболеваний сердца) и синдрома хронической усталости (СХУ), также называемого миалгическим энцефаломиелитом (МЭ), в открытом неслепом, нерандомизированном и неперекрестном субъективном исследовании. [ 30 ]
Дополнительный d -рибоза может обходить часть пентозофосфатного пути , пути производства энергии, для производства d- рибозо-5-фосфат. Фермента глюкозо-6-фосфатдегидрогеназы (G-6-PDH) часто не хватает в клетках, но в большей степени в пораженных тканях, например, в клетках миокарда у пациентов с сердечными заболеваниями. Поставка d -рибоза в митохондриях напрямую коррелирует с выработкой АТФ; уменьшился Поставка d -рибозы снижает количество вырабатываемого АТФ. Исследования показывают, что добавление d -рибоза после тканевой ишемии (например, ишемии миокарда) увеличивает выработку АТФ в миокарде и, следовательно, функцию митохондрий. По сути, введение дополнительных d -рибоза обходит ферментативный этап пентозофосфатного пути, обеспечивая альтернативный источник 5-фосфо- d -рибозо-1- пирофосфат для производства АТФ. Дополнительный d -рибоза улучшает восстановление уровня АТФ, а также уменьшает повреждение клеток у людей и других животных. Одно исследование показало, что использование дополнительных D -рибоза снижает частоту приступов стенокардии у мужчин с диагностированной ишемической болезнью сердца . [ 31 ] d -Рибоза использовалась для лечения многих патологических состояний, таких как синдром хронической усталости, фибромиалгия и дисфункция миокарда. Он также используется для уменьшения симптомов спазмов, боли, скованности и т. д. после тренировки и для улучшения спортивных результатов. [ нужна ссылка ] .
Ссылки
[ редактировать ]- ^ Индекс Merck: Энциклопедия химических веществ, лекарств и биологических препаратов (11-е изд.), Merck, 1989, ISBN. 091191028X , 8205
- ^ Уэст, Роберт С., изд. (1981). Справочник CRC по химии и физике (62-е изд.). Бока-Ратон, Флорида: CRC Press. п. С-506. ISBN 0-8493-0462-8 .
- ^ Фишер, Эмиль ; Пилоти, Оскар (1891). «О новой пентоновой кислоте и второй неактивной триоксиглутаровой кислоте » . Отчеты Немецкого химического общества (на немецком языке). 24 (2): 4214–4225. дои : 10.1002/cber.189102402322 . Архивировано из оригинала 4 июня 2020 года . Проверено 12 марта 2020 г.
- ^ Левен, Пенсильвания ; Джейкобс, Вашингтон (1909). «Об инозиновой кислоте». Отчеты Немецкого химического общества (на немецком языке). 42 (1): 1198–1203. дои : 10.1002/cber.190904201196 .
- ^ Левен, Пенсильвания ; Джейкобс, Вашингтон (1909). «О пентозе в нуклеиновых кислотах». Отчеты Немецкого химического общества (на немецком языке). 42 (3): 3247–3251. дои : 10.1002/cber.19090420351 .
- ^ Jump up to: а б Жанлоз, Роджер В .; Флетчер, Хьюитт Г. (1951). «Химия рибозы» . В Гудзоне, Клод С .; Кантор, Сидни М. (ред.). Достижения в химии углеводов . Том. 6. Академическая пресса . стр. 135–174. дои : 10.1016/S0096-5332(08)60066-1 . ISBN 9780080562650 . ПМИД 14894350 . Архивировано из оригинала 26 октября 2023 года . Проверено 15 декабря 2019 г.
- ^ Нечамкин, Ховард (1958). «Некоторые интересные этимологические производные химической терминологии». Научное образование . 42 (5): 463–474. Бибкод : 1958SciEd..42..463N . дои : 10.1002/sce.3730420523 .
- ^ Jump up to: а б с д Дьюик, Пол М. (2013). «Кислород как нуклеофил: гемицетали, гемикетали, ацетали и кетали» . Основы органической химии: для студентов фармации, медицинской химии и биологической химии . Джон Уайли и сыновья . стр. 224–234. ISBN 9781118681961 . Архивировано из оригинала 26 октября 2023 года . Проверено 15 декабря 2019 г.
- ^ Ли, Джеффри (июль – август 2012 г.). «Номенклатурные системы, не входящие в ИЮПАК» . Химия Интернэшнл . 34 (4). Международный союз теоретической и прикладной химии . Архивировано из оригинала 5 декабря 2019 года . Проверено 15 декабря 2019 г.
- ^ Jump up to: а б с Бутани, СП (2019). «Альдопентозы — сахара нуклеиновых кислот» . Химия биомолекул (2-е изд.). ЦРК Пресс . стр. 63–65. ISBN 9781000650907 . Архивировано из оригинала 26 октября 2023 года . Проверено 15 декабря 2019 г.
- ^ Jump up to: а б Дрю, Кеннет Н.; Заичек, Ярослав; Бондо, Гейл; Бозе, Бидиша; Серианни, Энтони С. (февраль 1998 г.). " 13 C-меченные альдопентозы: обнаружение и количественное определение циклических и ациклических форм с помощью гетероядерной 1D и 2D ЯМР-спектроскопии» . Carbodrive Research . 307 (3–4): 199–209. doi : 10.1016/S0008-6215(98)00040-8 .
- ^ де Вульф, П.; Вандам, Э.Дж. (1997). «Микробный синтез ᴅ-рибозы: метаболическая дерегуляция и процесс ферментации». Достижения прикладной микробиологии . 44 : 167–214. дои : 10.1016/S0065-2164(08)70462-3 . ISBN 9780120026449 .
- ^ Тамбула, ДЛ; Тенг, К.; Бартлетт, Миннесота; Уитмен, ВБ (1997). «Биосинтез рибозы и доказательства альтернативного первого этапа общего пути ароматических аминокислот у Methanococcus maripaludis» . Журнал бактериологии . 179 (19): 6010–6013. дои : 10.1128/jb.179.19.6010-6013.1997 . ПМК 179501 . ПМИД 9324245 .
- ^ Вульф, П. Де; Вандам, Э.Дж. (1997). «Производство d-рибозы путем ферментации». Прикладная микробиология и биотехнология . 48 (2): 141–148. дои : 10.1007/s002530051029 . hdl : 11572/262019 . ПМИД 9299771 . S2CID 34340369 .
- ^ Штайгервальд, Билл; Джонс, Нэнси; Фурукава, Ёсихиро (18 ноября 2019 г.). «Первое обнаружение сахаров в метеоритах дает ключ к разгадке происхождения жизни» . НАСА . Архивировано из оригинала 15 января 2021 года . Проверено 18 ноября 2019 г.
- ^ Фурукава, Ёсихиро; Чикараиси, Ёсито; Окоучи, Наахико; Огава, Нанако О.; Главин, Дэниел П.; Дворкин, Джейсон П.; Абэ, Чиаки; Накамура, Томоки (2019). «Внеземные рибоза и другие сахара в примитивных метеоритах» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (49): 24440–24445. Бибкод : 2019PNAS..11624440F . дои : 10.1073/pnas.1907169116 . ПМК 6900709 . ПМИД 31740594 .
- ^ Jump up to: а б с д Блумфилд, Виктор; Кротерс, Дональд; Тиноко, Игнасио (2000). Нуклеиновые кислоты: структура, свойства и функции . Университетские научные книги. стр. 19–25 . ISBN 9780935702491 .
- ^ Фут, Дональд; Фут, Джудит (2011). Биохимия . Джон Уайли и сыновья, Inc. стр. 1152 , 1153. ISBN 978-0470570951 .
- ^ Фолоппе, Николя; МакКерелл, Александр Д. (август 1998 г.). «Конформационные свойства дезоксирибозных и рибозных фрагментов нуклеиновых кислот: квантово-механическое исследование». Журнал физической химии Б. 102 (34): 6669–6678. дои : 10.1021/jp9818683 . ISSN 1520-6106 .
- ^ «Архитектура нуклеиновой кислоты» . fbio.uh.cu . Архивировано из оригинала 17 мая 2018 года . Проверено 8 октября 2019 г.
- ^ Нидл, Стивен (2008). «Строительные блоки ДНК и РНК». В Нидле, Стивен (ред.). Принципы структуры нуклеиновых кислот . Академическая пресса . стр. 20–37 . дои : 10.1016/B978-012369507-9.50003-0 . ISBN 9780123695079 .
- ^ Борк, Пер ; Сандер, Крис ; Валенсия, Альфонсо (1993). «Конвергентная эволюция сходных ферментативных функций в разных белковых складках: семейства сахаркиназ гексокиназы, рибокиназы и галактокиназы» . Белковая наука . 2 (1): 31–40. дои : 10.1002/pro.5560020104 . ПМК 2142297 . ПМИД 8382990 .
- ^ Пак, Джэ; Гупта, Рэдхи С. (2008). «Аденозинкиназа и рибокиназа – семейство белков RK» . Клеточные и молекулярные науки о жизни . 65 (18): 2875–2896. дои : 10.1007/s00018-008-8123-1 . ПМЦ 11131688 . ПМИД 18560757 . S2CID 11439854 .
- ^ Jump up to: а б с Пучсервер, Пере (2018). «Сигнальная трансдукция и метаболомика». В Хоффмане, Рональде; Бенц, Эдвард Дж.; Зильберштейн, Лесли Э.; Хеслоп, Хелен Э. (ред.). Гематология (7-е изд.). Эльзевир. стр. 68–78. дои : 10.1016/B978-0-323-35762-3.00007-X . ISBN 9780323357623 .
- ^ «Лекарственные травы, добавки, индекс AZ» . PDRHealth.com . ПДР, ООО. Архивировано из оригинала 11 октября 2008 года.
- ^ Jump up to: а б с Хэмлоу, Лукас; Он, Ченчен; Фан, Лин; Ву, Ранран; Ян, Бо; Роджерс, Монтана; Берден, Гиль; Ооменс, Дж. (июнь 2015 г.). Структурные [sic] эффекты модификаций цитидин-2'-рибозы, определенные с помощью спектроскопии действия Irmpd . 70-й Международный симпозиум по молекулярной спектроскопии. Университет Иллинойса Урбана-Шампейн . Бибкод : 2015isms.confEMI13H . дои : 10.15278/isms.2015.MI13 .
- ^ Jump up to: а б Эвич, Марина; Спринг-Коннелл, Александр М.; Германн, Маркус В. (27 января 2017 г.). «Влияние модифицированных сахаров рибозы на конформацию и функцию нуклеиновой кислоты» . Гетероциклические связи . 23 (3): 155–165. дои : 10.1515/hc-2017-0056 . ISSN 2191-0197 . S2CID 91052034 .
- ^ Пикок, Хайден; Фучини, Раймонд В.; Джаялат, Прасанна; Ибарра-Соса, Хосе М.; Харингсма, Генри Дж.; Фланаган, В. Майкл; Уиллингем, Ааррон; Бил, Питер А. (2011). «Модификации нуклеиновых оснований и рибозы контролируют иммуностимуляцию с помощью РНК-миметика микроРНК-122» . Журнал Американского химического общества . 133 (24): 9200–9203. дои : 10.1021/ja202492e . ПМК 3116021 . ПМИД 21612237 .
- ^ Омран, Хейдер; Маккартер, Дин; Сен-Сир, Джон; Людериц, Берндт (2004). «ᴅ-Рибоза помогает пациентам с застойной сердечной недостаточностью» . Экспериментальная и клиническая кардиология . Лето (9 (2)): 117–118. ПМК 2716264 . ПМИД 19641697 .
- ^ Тейтельбаум, Джейкоб Э.; Джонсон, Кларенс; Сен-Сир, Джон (26 ноября 2006 г.). «Использование ᴅ-рибозы при синдроме хронической усталости и фибромиалгии: пилотное исследование». Журнал альтернативной и дополнительной медицины . 12 (9): 857–862. CiteSeerX 10.1.1.582.4800 . дои : 10.1089/acm.2006.12.857 . ПМИД 17109576 .
- ^ «Рибоза» . wa.kaiserpermanente.org . Архивировано из оригинала 3 марта 2021 года . Проверено 7 октября 2019 г.
Виды углеводов |
|---|
| Базы данных органов управления : Национальные |
|---|