Разновидность
Вид или ( мн. :вид) часто определяется как самая большая группа организмов в которой любые две особи соответствующего пола , типа спаривания могут производить плодовитое потомство , как правило, путем полового размножения . [ 1 ] Это основная единица классификации и таксономического ранга организма, а также единица биоразнообразия . Другие способы определения видов включают их кариотип , последовательность ДНК , морфологию , поведение или экологическую нишу . Кроме того, палеонтологи используют концепцию хроновида, поскольку воспроизводство ископаемых невозможно изучить. По последним строгим оценкам общее количество видов эукариот составляет от 8 до 8,7 миллионов. [ 2 ] [ 3 ] [ 4 ] Около 14% из них были описаны к 2011 году. [ 4 ] Всем видам (кроме вирусов ) присвоено название, состоящее из двух частей , «биномиальное». Первая часть бинома — это род , к которому принадлежит вид. Вторая часть называется видовым названием или видовым эпитетом (в ботанической номенклатуре , иногда также в зоологической номенклатуре ). Например, удав — один из видов рода Boa , удав эпитет вида — .
Хотя приведенные выше определения на первый взгляд могут показаться адекватными, при более внимательном рассмотрении они представляют собой проблемные концепции видов. Например, границы между близкородственными видами становятся нечеткими при гибридизации , в видовом комплексе из сотен сходных микровидов и в кольцевых видах . Кроме того, среди организмов, которые размножаются только бесполым путем , концепция репродуктивного вида разрушается, и каждый клон потенциально является микровидом. Хотя ни одно из этих определений не является полностью удовлетворительным, и хотя концепция вида, возможно, не является идеальной моделью жизни, она по-прежнему остается полезным инструментом для ученых и защитников природы для изучения жизни на Земле, независимо от теоретических трудностей. Если бы виды были фиксированными и четко отличались друг от друга, проблем не было бы, но эволюционные процессы заставляют виды изменяться. Это обязывает систематиков решать, например, когда произошло достаточно изменений, чтобы объявить, что линия должна быть разделена на несколько хроновиды , или когда популяции разошлись, чтобы иметь достаточно различные состояния признаков, чтобы их можно было описать как кладистические виды.
Виды и высшие таксоны рассматривались со времен Аристотеля до XVIII века как категории, которые можно было выстроить в иерархию, великую цепь бытия . В 19 веке биологи поняли, что виды могут эволюционировать, если им будет достаточно времени. Книга Чарльза Дарвина 1859 года «Происхождение видов» объяснила, как виды могли возникнуть в результате естественного отбора . Это понимание было значительно расширено в 20 веке благодаря генетике и популяционной экологии . Генетическая изменчивость возникает в результате мутаций и рекомбинаций , в то время как сами организмы мобильны, что приводит к географической изоляции и генетическому дрейфу с различным давлением отбора . Иногда обмен генами между видами может осуществляться путем горизонтального переноса генов ; новые виды могут быстро возникать за счет гибридизации и полиплоидии ; и виды могут вымереть по разным причинам. Вирусы представляют собой особый случай, обусловленный балансом мутаций и отбора , и их можно рассматривать как квазивиды .
Определение
[ редактировать ]Биологи и систематики предприняли множество попыток определить виды, начиная с морфологии и заканчивая генетикой . У ранних систематиков, таких как Линней, не было другого выбора, кроме как описывать то, что они видели: позже это было формализовано как типологическая или морфологическая концепция вида. Эрнст Майр подчеркивал репродуктивную изоляцию, но это, как и концепции других видов, трудно или даже невозможно проверить. [ 5 ] [ 6 ] Позже биологи попытались уточнить определение Майра, в частности, используя концепции распознавания и сплоченности. [ 7 ] Многие понятия весьма схожи или пересекаются, поэтому их нелегко сосчитать: биолог Р. Л. Мэйден зафиксировал около 24 концепций, [ 8 ] а философ науки Джон Уилкинс насчитал 26. [ 5 ] Уилкинс далее сгруппировал концепции видов в семь основных видов концепций: (1) агамовиды для бесполых организмов (2) биовиды для репродуктивно изолированных половых организмов (3) эковиды, основанные на экологических нишах (4) эволюционные виды, основанные на происхождении (5) генетические виды на основе генофонда (6) морфовиды, основанные на форме или фенотипе, и (7) таксономические виды, виды, определенные систематиком. [ 9 ]
Типологический или морфологический вид
[ редактировать ]
Типологический вид — это группа организмов, особи которой соответствуют определенным фиксированным свойствам (типу), так что даже неграмотные люди часто признают тот же таксон, что и современные систематики. [ 11 ] [ 12 ] Группы вариаций или фенотипов внутри особей (например, более длинные или короткие хвосты) будут различать виды. Этот метод использовался как «классический» метод определения видов, например, у Линнея, на ранних этапах эволюционной теории. Однако разные фенотипы не обязательно относятся к разным видам (например, четырехкрылая дрозофила, рожденная от двукрылой матери, не является отдельным видом). Виды, названные таким образом, называются морфовидами . [ 13 ] [ 14 ]
В 1970-х годах Роберт Р. Сокал , Теодор Дж. Кровелло и Питер Снит предложили вариацию концепции морфологического вида, фенетический вид, определяемый как набор организмов со схожим фенотипом друг с другом, но отличающимся фенотипом от других наборов. организмов. [ 15 ] Он отличается от концепции морфологического вида тем, что включает числовую меру расстояния или сходства с объектами кластера, основанную на многомерных сравнениях достаточно большого количества фенотипических признаков. [ 16 ]
Распознавание и сплоченность видов
[ редактировать ]Виды, распознающие партнеров, — это группа организмов, размножающихся половым путем, которые распознают друг друга как потенциальных партнеров. [ 17 ] [ 18 ] Если расширить это, чтобы обеспечить изоляцию после спаривания, сплоченный вид представляет собой наиболее инклюзивную популяцию особей, обладающую потенциалом фенотипической сплоченности посредством внутренних механизмов сплоченности; независимо от того, могут ли популяции успешно гибридизоваться, они по-прежнему остаются отдельными сплоченными видами, если степень гибридизации недостаточна для полного смешивания их соответствующих генофондов . [ 19 ] Дальнейшее развитие концепции узнавания дает биосемиотическая концепция вида. [ 20 ]
Генетическое сходство и виды штрих-кода
[ редактировать ]
В микробиологии гены могут свободно перемещаться даже между отдаленно родственными бактериями, возможно, распространяясь на весь бактериальный домен. Как правило, микробиологи предполагают, что представителей бактерий или архей с последовательностями генов 16S рибосомальной РНК, более похожими друг на друга более чем на 97%, необходимо проверять с помощью гибридизации ДНК-ДНК , чтобы решить, принадлежат ли они к одному и тому же виду. [ 21 ] Это понятие было сужено в 2006 году до сходства 98,7%. [ 22 ]
Метод средней идентичности нуклеотидов (ANI) позволяет количественно оценить генетическое расстояние между целыми геномами , используя области длиной около 10 000 пар оснований . При наличии достаточного количества данных из геномов одного рода можно использовать алгоритмы для классификации видов, как для Pseudomonas avellanae в 2013 г., [ 23 ] и для всех секвенированных бактерий и архей с 2020 года. [ 24 ] Наблюдаемые значения ANI среди последовательностей, по-видимому, имеют «разрыв ANI» на уровне 85–95%, что позволяет предположить наличие генетической границы, подходящей для определения концепции вида. [ 25 ]
Штрих-кодирование ДНК было предложено как способ различения видов, который может использовать даже неспециалист. [ 26 ] Один из штрих-кодов представляет собой участок митохондриальной ДНК внутри гена цитохром-с-оксидазы . База данных Barcode of Life Data System содержит последовательности штрих-кодов ДНК более чем 190 000 видов. [ 27 ] [ 28 ] Однако такие ученые, как Роб ДеСалль, выразили обеспокоенность тем, что классическую таксономию и штрих-кодирование ДНК, которые они считают неправильным, необходимо согласовать, поскольку они по-разному разграничивают виды. [ 29 ] Генетическая интрогрессия , опосредованная эндосимбионтами и другими векторами, может еще больше сделать штрих-коды неэффективными для идентификации видов. [ 30 ]
Филогенетические или кладистические виды
[ редактировать ]
Филогенетический или кладистический вид — это «наименьшая совокупность популяций (половых) или линий (бесполых), которую можно диагностировать по уникальной комбинации состояний признаков у сопоставимых особей (семафоронтов)». [ 31 ] Эмпирическая основа – наблюдаемые состояния характера – предоставляет доказательства в поддержку гипотез об эволюционно расходящихся линиях, которые сохранили свою наследственную целостность во времени и пространстве. [ 32 ] [ 33 ] [ 34 ] [ 35 ] Молекулярные маркеры могут быть использованы для определения диагностических генетических различий в ядерной или митохондриальной ДНК различных видов. [ 36 ] [ 31 ] [ 37 ] Например, в исследовании, проведенном на грибах , изучение нуклеотидных признаков с использованием кладистических видов дало наиболее точные результаты в распознавании многочисленных видов грибов из всех изученных концепций. [ 37 ] [ 38 ] Версии филогенетической концепции вида, подчеркивающие монофилию или диагностируемость. [ 39 ] может привести к разделению существующих видов, например, у Bovidae , путем признания старых подвидов видами, несмотря на то, что не существует репродуктивных барьеров, и популяции могут интерградировать морфологически. [ 40 ] Другие назвали этот подход таксономической инфляцией , размывая концепцию вида и делая таксономию нестабильной. [ 41 ] Третьи защищают этот подход, считая «таксономическую инфляцию» уничижительной и называя противоположную точку зрения «таксономическим консерватизмом»; утверждая, что политически целесообразно разделить виды и признать меньшие популяции на уровне видов, потому что это означает, что их легче включить как находящихся под угрозой исчезновения в МСОП Красный список , и они могут привлечь природоохранное законодательство и финансирование. [ 42 ]
В отличие от концепции биологического вида, кладистический вид не опирается на репродуктивную изоляцию - его критерии не зависят от процессов, которые являются неотъемлемой частью других концепций. [ 31 ] Следовательно, это применимо к бесполым линиям. [ 36 ] [ 37 ] Однако он не всегда обеспечивает четкие и интуитивно удовлетворительные границы между таксонами и может потребовать нескольких источников доказательств, таких как более одного полиморфного локуса, чтобы дать правдоподобные результаты. [ 37 ]
Эволюционные виды
[ редактировать ]Эволюционный вид, предложенный Джорджем Гейлордом Симпсоном в 1951 году, представляет собой «субъектность, состоящую из организмов, которая сохраняет свою идентичность от других таких сущностей во времени и в пространстве и которая имеет свою собственную независимую эволюционную судьбу и исторические тенденции». [ 8 ] [ 43 ] Это отличается от концепции биологических видов тем, что воплощает устойчивость во времени. Уайли и Мэйден заявили, что они считают концепцию эволюционного вида «идентичной» концепции вида как родословной Вилли Хеннига , и утверждали, что концепция биологического вида, «несколько версий» концепции филогенетического вида и идея о том, что виды относятся к тому же виду, что и высшие таксоны, и не подходят для изучения биоразнообразия (с целью точной оценки количества видов). Они также предположили, что эта концепция работает как для бесполых, так и для видов, размножающихся половым путем. [ 44 ] Вариантом этой концепции является Кевина де Кейроза . «Общая концепция происхождения видов» [ 45 ]
Экологические виды
[ редактировать ]Экологический вид — это совокупность организмов, приспособленных к определенному набору ресурсов, называемому нишей в окружающей среде. Согласно этой концепции, популяции образуют дискретные фенетические кластеры, которые мы признаем как виды, потому что экологические и эволюционные процессы, контролирующие распределение ресурсов, имеют тенденцию создавать эти кластеры. [ 46 ]
Генетические виды
[ редактировать ]Генетический вид, по определению Роберта Бейкера и Роберта Брэдли, представляет собой набор генетически изолированных скрещивающихся популяций. Это похоже на концепцию биологических видов Майра, но подчеркивает генетическую, а не репродуктивную изоляцию. [ 47 ] В XXI веке генетический вид можно будет установить путем сравнения последовательностей ДНК. Ранее были доступны другие методы, такие как сравнение кариотипов (наборов хромосом ) и аллозимов ( вариантов ферментов ). [ 48 ]
Эволюционно значимая единица
[ редактировать ]( Эволюционно значимая единица ESU) или «виды дикой природы». [ 49 ] популяция организмов, считающихся обособленными в целях сохранения. [ 50 ]
Хроновиды
[ редактировать ]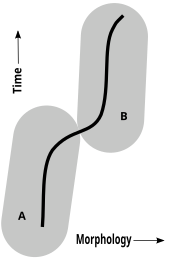
В палеонтологии только сравнительная анатомия (морфология) и гистология. [ 51 ] на основе окаменелостей концепцию хроновида . в качестве доказательства можно применить Во время анагенеза (эволюции, не обязательно связанной с ветвлением) некоторые палеонтологи стремятся идентифицировать последовательность видов, каждый из которых произошел от филетически вымершего ранее вида посредством непрерывных, медленных и более или менее однородных изменений. В такой временной последовательности некоторые палеонтологи оценивают, сколько изменений необходимо, чтобы морфологически отличная форма считалась видом, отличным от ее предков. [ 52 ] [ 53 ] [ 54 ] [ 55 ]
Вирусные квазивиды
[ редактировать ]Вирусы имеют огромные популяции, и их жизнь сомнительна, поскольку они состоят всего лишь из цепочки ДНК или РНК в белковой оболочке, и быстро мутируют. Все эти факторы делают традиционные концепции видов практически неприменимыми. [ 56 ] Вирусный квазивид — это группа генотипов, связанных сходными мутациями, конкурирующих в высокомутагенной среде и, следовательно, регулируемых балансом мутаций и отбора . Прогнозируется, что вирусный квазивид, находящийся на низком, но эволюционно нейтральном и высокосвязанном (то есть плоском) участке ландшафта приспособленности, вытеснит квазивид, находящийся на более высоком, но более узком пике приспособленности, к которому окружающие мутанты непригодны. эффект» или «выживание самого плоского». Нет никаких предположений о том, что вирусный квазивид напоминает традиционный биологический вид. [ 57 ] [ 58 ] [ 59 ] Международный комитет по таксономии вирусов с 1962 года разработал универсальную таксономическую схему вирусов; это стабилизировало вирусную таксономию. [ 60 ] [ 61 ] [ 62 ]
Концепция биологических видов Майра
[ редактировать ]
В большинстве современных учебников используется определение Эрнста Майра , данное в 1942 году: [ 63 ] [ 64 ] известная как «Концепция биологических видов», как основа для дальнейшего обсуждения определения вида. Ее также называют репродуктивной или изоляционной концепцией. Это определяет вид как [ 65 ]
группы фактически или потенциально скрещивающихся природных популяций, репродуктивно изолированных от других таких групп. [ 65 ]
Утверждалось, что это определение является естественным следствием влияния полового размножения на динамику естественного отбора. [ 66 ] [ 67 ] [ 68 ] [ 69 ] Использование Майром прилагательного «потенциально» стало предметом споров; некоторые интерпретации исключают необычные или искусственные спаривания, которые происходят только в неволе или с участием животных, способных к спариванию, но обычно не делающих этого в дикой природе. [ 65 ]
Проблема видов
[ редактировать ]Трудно определить вид таким образом, чтобы это применимо ко всем организмам. [ 70 ] Спор о концепциях видов называется проблемой видов. [ 65 ] [ 71 ] [ 72 ] [ 73 ] Проблема была осознана еще в 1859 году, когда Дарвин написал в «Происхождении видов» :
Ни одно определение не удовлетворило всех натуралистов; однако каждый натуралист смутно знает, что он имеет в виду, когда говорит о виде. Обычно этот термин включает неизвестный элемент отдельного акта творения. [ 74 ]
Когда концепция Майра терпит крах
[ редактировать ]
Многие авторы утверждают, что простое определение из учебника, соответствующее концепции Майра, хорошо работает для большинства многоклеточных организмов , но в некоторых ситуациях не работает:
- Когда организмы размножаются бесполым путем , как у одноклеточных организмов, таких как бактерии и другие прокариоты , [ 75 ] и партеногенетические или апомиктические многоклеточные организмы. штрих-кодирование ДНК и филогенетика . В этих случаях обычно используются [ 76 ] [ 77 ] [ 78 ] Термин «квазивид» иногда используется для обозначения быстро мутирующих объектов, таких как вирусы. [ 79 ] [ 80 ]
- Когда ученые не знают, способны ли две морфологически сходные группы организмов к скрещиванию; это относится ко всем вымершим формам жизни в палеонтологии , поскольку эксперименты по разведению невозможны. [ 81 ]
- Когда гибридизация обеспечивает значительный поток генов между видами. [ 82 ]
- У кольцевых видов , когда представители соседних популяций в широко непрерывном ареале успешно скрещиваются, а представители более отдаленных популяций - нет. [ 83 ]


Идентификация видов затруднена из-за несоответствия между молекулярными и морфологическими исследованиями; их можно разделить на два типа: (i) одна морфология, множественные линии (например, морфологическая конвергенция , загадочные виды ) и (ii) одна линия, множественные морфологии (например, фенотипическая пластичность , несколько стадий жизненного цикла ). [ 84 ] Кроме того, горизонтальный перенос генов (HGT) затрудняет определение вида. [ 85 ] Все определения видов предполагают, что организм получает свои гены от одного или двух родителей, очень похоже на «дочерний» организм, но это не то, что происходит при HGT. [ 86 ] Имеются убедительные доказательства существования ГПГ между очень несходными группами прокариот и, по крайней мере, иногда между несходными группами эукариот . [ 85 ] включая некоторых ракообразных и иглокожих . [ 87 ]
Эволюционный биолог Джеймс Маллет заключает, что
нет простого способа определить, принадлежат ли родственные географические или временные формы одному и тому же виду или разным видам. Пробелы в видах можно проверить только локально и в определенный момент времени. Приходится признать, что догадка Дарвина верна: любая локальная реальность или целостность видов значительно уменьшаются в больших географических диапазонах и периодах времени. [ 19 ]
The botanist Brent Mishler [ 88 ] утверждал, что концепция вида недействительна, особенно потому, что поток генов уменьшается постепенно, а не дискретными шагами, что затрудняет объективное разграничение видов. [ 89 ] наблюдались сложные и нестабильные закономерности потока генов цихлид Действительно, у костистых Великих восточноафриканских озер . [ 90 ] Уилкинс утверждал, что «если бы мы были верны теории эволюции и вытекающему из нее филогенетическому подходу к таксонам, нам следует заменить ее идеей «наименьшей клады»» (концепцией филогенетических видов). [ 91 ] Мишлер и Уилкинс [ 92 ] и другие [ 93 ] согласны с этим подходом, хотя это и вызовет трудности в биологической номенклатуре. Уилкинс процитировал замечание ихтиолога Чарльза Тейта Ригана в начале 20-го века о том, что «вид — это то, что биолог с достаточной квалификацией решит назвать видом». [ 91 ] Уилкинс отметил, что философ Филип Китчер назвал это «концепцией циничного вида». [ 94 ] и утверждая, что это далеко не цинично, а с пользой приводит к эмпирической таксономии для любой данной группы, основанной на опыте систематиков. [ 91 ] Другие биологи пошли еще дальше и заявили, что мы должны полностью отказаться от видов и обратиться к «наименее инклюзивным таксономическим единицам» (LITU). [ 95 ] точка зрения, которая соответствовала бы современной эволюционной теории. [ 93 ]
Агрегаты микровидов
[ редактировать ]Концепция вида еще более ослабляется существованием микровидов , групп организмов, включающих множество растений, с очень небольшой генетической изменчивостью, обычно образующих видовые агрегаты . [ 96 ] Например, одуванчик Taraxacum officinale и ежевика Rubus fruticosus представляют собой агрегаты со многими микровидами — около 400 у ежевики и более 200 у одуванчика. [ 97 ] осложняются гибридизацией , апомиксисом и полиплоидией , из-за чего поток генов между популяциями трудно определить, а их таксономия является спорной. [ 98 ] [ 99 ] [ 100 ] Комплексы видов встречаются у насекомых, таких как Heliconius , бабочки [ 101 ] позвоночные, такие как древесные лягушки Hypsiboas , [ 102 ] и грибы, такие как мухомор . [ 103 ]
-
 Ежевика принадлежит к любому из сотен микровидов Rubus fruticosus совокупности видов .
Ежевика принадлежит к любому из сотен микровидов Rubus fruticosus совокупности видов . -
 Род бабочек Heliconius включает множество схожих видов.
Род бабочек Heliconius включает множество схожих видов. -
 Комплекс видов Hypsiboascalcaratus - fasciatus включает не менее шести видов древесных лягушек.
Комплекс видов Hypsiboascalcaratus - fasciatus включает не менее шести видов древесных лягушек.



Гибридизация
[ редактировать ]Естественная гибридизация представляет собой проблему для концепции репродуктивно изолированного вида, поскольку плодовитые гибриды допускают поток генов между двумя популяциями. Например, ворона-падальщик Corvus corone и ворона Corvus cornix появляются и классифицируются как отдельные виды, однако они могут гибридизироваться там, где их географические ареалы пересекаются. [ 104 ]
- Гибридизация падали и вороны обеспечивает поток генов между «видами»
-

-
 Гибрид с темным брюшком.
Гибрид с темным брюшком. -




Виды колец
[ редактировать ]Кольцевой вид — это связный ряд соседних популяций, каждая из которых может скрещиваться половым путем с соседними родственными популяциями, но для которых существуют по крайней мере две «конечные» популяции в серии, слишком отдаленно связанные с скрещиванием, хотя существует потенциальный поток генов между каждой «связанной» популяцией. [ 105 ] Такие неразмножающиеся, хотя и генетически связанные, «конечные» популяции могут сосуществовать в одном и том же регионе, замыкая кольцо. Таким образом, кольцевые виды представляют собой трудность для любой концепции вида, основанной на репродуктивной изоляции. [ 106 ] Однако кольцевые виды в лучшем случае редки. Предлагаемые примеры включают сельдевую чайку – комплекс малых черных чаек вокруг Северного полюса, группу Ensatina eschscholtzii из 19 популяций саламандр в Америке, [ 107 ] и зеленоватая славка в Азии, [ 108 ] но многие так называемые кольцевые виды оказались результатом неправильной классификации, что привело к вопросу о том, действительно ли существуют какие-либо кольцевые виды. [ 109 ] [ 110 ] [ 111 ] [ 112 ]
-
 Семь «видов» чаек Larus скрещиваются в кольце вокруг Арктики.
Семь «видов» чаек Larus скрещиваются в кольце вокруг Арктики. -
 Противоположные концы кольца: серебристая чайка ( Larus argentatus ) (спереди) и малая чайка ( Larus fuscus ) в Норвегии.
Противоположные концы кольца: серебристая чайка ( Larus argentatus ) (спереди) и малая чайка ( Larus fuscus ) в Норвегии. -
 Зеленоватая славка . Phylloscopus trochiloides
Зеленоватая славка . Phylloscopus trochiloides -





Таксономия и именование
[ редактировать ]
Распространенные и научные названия
[ редактировать ]Обычно используемые названия видов организмов часто неоднозначны: «кошка» может означать домашнюю кошку Felis catus или семейство кошачьих Felidae . Другая проблема с общими именами заключается в том, что они часто различаются от места к месту, так что пума, пума, катамаунт, пантера, художник и горный лев означают Puma concolor в разных частях Америки , в то время как «пантера» может также означать ягуара ( Panthera onca ) Латинской Америки или леопард ( Panthera pardus ) Африки и Азии. Напротив, научные названия видов выбираются уникальными и универсальными (за исключением некоторых межкодовых омонимов ); они состоят из двух частей, используемых вместе : род , как у Puma , и видовой эпитет, как у concolor . [ 113 ] [ 114 ]
Описание вида
[ редактировать ]
Виду дается таксономическое название, когда типовой экземпляр официально описан в публикации, которая присваивает ему уникальное научное название. Описание обычно предоставляет средства для идентификации нового вида, которые не могут быть основаны исключительно на морфологии. [ 115 ] (см. загадочные виды ), отличая его от других ранее описанных и родственных или запутанных видов, а также предоставляя действительно опубликованное название (в ботанике) или доступное название (в зоологии), когда статья принимается к публикации. Типовой материал обычно хранится в постоянном хранилище, часто в исследовательской коллекции крупного музея или университета, что позволяет проводить независимую проверку и позволяет сравнивать образцы. [ 116 ] [ 117 ] [ 118 ] Описателям новых видов предлагается выбирать названия, которые, по словам Международного кодекса зоологической номенклатуры , «уместны, компактны, благозвучны, запоминаются и не вызывают обид». [ 119 ]
Сокращения
[ редактировать ]В книгах и статьях иногда намеренно не идентифицируются виды полностью, используя аббревиатуру «sp.». в единственном числе или «спп». (расшифровывается как «виды множественного числа », что по латыни означает «несколько видов») во множественном числе вместо конкретного названия или эпитета (например, Canis sp.). Обычно это происходит, когда авторы уверены, что некоторые особи принадлежат к определенному роду, но не уверены, к какому конкретному виду они принадлежат, как это часто бывает в палеонтологии . [ 120 ]
Авторы также могут использовать «spp». как короткий способ сказать, что что-то применимо ко многим видам внутри рода, но не ко всем. Если ученые имеют в виду, что что-то применимо ко всем видам внутри рода, они используют название рода без конкретного названия или эпитета. Названия родов и видов обычно печатаются курсивом . Однако такие сокращения, как «sp.» не следует выделять курсивом. [ 120 ]
Если идентичность вида не ясна, специалист может использовать «ср.». перед эпитетом, чтобы указать, что требуется подтверждение. Сокращения «№». (рядом) или «афф». (аффинный) может использоваться, когда идентичность неясна, но когда вид кажется похожим на вид, упомянутый ниже. [ 120 ]
Идентификационные коды
[ редактировать ]С появлением онлайн-баз данных были разработаны коды для идентификации уже определенных видов, в том числе:
- Национальный центр биотехнологической информации (NCBI) использует числовой «таксид» или идентификатор таксономии , «стабильный уникальный идентификатор», например, таксид Homo sapiens — 9606. [ 121 ]
- Киотская энциклопедия генов и геномов (KEGG) использует трех- или четырехбуквенный код для ограниченного числа организмов; в этом коде, например, H. sapiens — это просто hsa . [ 122 ]
- UniProt использует «мнемонику организма», состоящую не более чем из пяти буквенно-цифровых символов, например, ЧЕЛОВЕК для H. sapiens . [ 123 ]
- Интегрированная система таксономической информации (ITIS) предоставляет уникальный номер каждому виду. LSID для Homo sapiens : urn :lsid:catalogueoflife.org:taxon:4da6736d-d35f-11e6-9d3f-bc764e092680:col20170225 . [ 124 ]
Комкование и расщепление
[ редактировать ]Наименование конкретного вида, включая то, к какому роду (и более высоким таксонам) он отнесен, является гипотезой об эволюционных связях и различимости этой группы организмов. По мере поступления дополнительной информации гипотеза может быть подтверждена или опровергнута. Иногда, особенно в прошлом, когда общение было более трудным, систематики, работающие изолированно, давали два разных названия отдельным организмам, которые позже были идентифицированы как один и тот же вид. Когда обнаруживается, что два названия вида применимы к одному и тому же виду, более старое название вида имеет приоритет и обычно сохраняется, а более новое название рассматривается как младший синоним, этот процесс называется синонимией . Разделение таксона на несколько, часто новых, таксонов называется расщеплением . Коллеги часто называют систематиков «сборщиками» или «расщепителями», в зависимости от их личного подхода к признанию различий или сходств между организмами. [ 125 ] [ 126 ] [ 120 ] Ограничение таксонов, считающееся таксономическим решением по усмотрению компетентных специалистов, не регулируется Кодексами зоологической или ботанической номенклатуры, в отличие от Филокода , и противоречит тому, что делается в ряде других областей, в которых определения технические термины, такие как геохронологические единицы и геополитические объекты, четко разграничены. [ 127 ] [ 93 ]
Широкие и узкие смыслы
[ редактировать ]Номенклатурные коды , регулирующие наименование видов, включая МКЗН для животных и МКП для растений, не устанавливают правил для определения границ вида. Исследования могут изменить границы, также известные как ограничения, на основе новых данных. Тогда, возможно, потребуется различать виды с помощью используемых граничных определений, и в таких случаях названия могут быть уточнены с помощью sensu stricto («в узком смысле»), чтобы обозначить использование в точном значении, данном автором, например человеком, назвавшим вид, тогда как антоним sensu lato («в широком смысле») обозначает более широкое использование, например, включая другие подвиды. Другие сокращения, такие как «аук». («автор»), а такие уточнения, как «не» («не»), могут использоваться для дальнейшего уточнения смысла, в котором указанные авторы определили или описали этот вид. [ 120 ] [ 128 ] [ 129 ]
Изменять
[ редактировать ]Виды подвержены изменениям, будь то путем эволюции в новые виды, [ 130 ] обмен генами с другими видами, [ 131 ] сливаясь с другими видами или вымирая. [ 132 ]
Видообразование
[ редактировать ]Эволюционный процесс , посредством которого биологические популяции организмов, размножающихся половым путем, превращаются в отдельные или репродуктивно изолированные виды, называется видообразованием . [ 133 ] [ 134 ] Чарльз Дарвин был первым, кто описал роль естественного отбора в видообразовании в своей книге 1859 года «Происхождение видов» . [ 135 ] Видообразование зависит от степени репродуктивной изоляции , уменьшенного потока генов. Легче всего это происходит при аллопатрическом видообразовании, когда популяции разделены географически и могут постепенно расходиться по мере накопления мутаций. Репродуктивная изоляция находится под угрозой из-за гибридизации, но ее можно отобрать, если пара популяций имеет несовместимые аллели одного и того же гена, как описано в модели Бейтсона-Добжанского-Мюллера . [ 130 ] Другой механизм, филетическое видообразование, предполагает постепенное изменение одной линии с течением времени в новую и отдельную форму ( хроновид ) без увеличения числа образующихся видов. [ 136 ]
Обмен генами между видами
[ редактировать ]Горизонтальный перенос генов между организмами разных видов посредством гибридизации , антигенного сдвига или реассортации иногда является важным источником генетических вариаций. Вирусы могут передавать гены между видами. Бактерии могут обмениваться плазмидами с бактериями других видов, в том числе с некоторыми явно отдаленно родственными в разных филогенетических доменах , что затрудняет анализ их взаимоотношений и ослабляет концепцию бактериального вида. [ 137 ] [ 85 ] [ 138 ] [ 131 ]
Луи-Мари Бобей и Говард Охман предполагают, основываясь на анализе геномов многих типов бактерий, что их часто можно сгруппировать «в сообщества, которые регулярно обмениваются генами», почти так же, как растения и животные могут быть сгруппированы в репродуктивные группы. изолированные гнездящиеся популяции. Таким образом, бактерии могут образовывать виды, аналогичные концепции биологических видов Майра, состоящие из популяций, размножающихся бесполым путем, которые обмениваются генами путем гомологичной рекомбинации. [ 139 ] [ 140 ]
Вымирание
[ редактировать ]Вид вымирает, когда умирает последняя особь он может этого вида, но функционально вымереть задолго до этого момента. По оценкам, более 99 процентов всех видов, когда-либо живших на Земле, а это около пяти миллиардов видов, сейчас вымерли. Некоторые из них находились в состоянии массового вымирания , например, в конце ордовикского , девонского , пермского , триасового и мелового периодов. Массовые вымирания имели множество причин, включая вулканическую активность , изменение климата и изменения в химии океана и атмосферы, а они, в свою очередь, оказали серьезное воздействие на экологию Земли, атмосферу, поверхность суши и воды. [ 141 ] [ 142 ] Другая форма вымирания — ассимиляция одного вида другим путем гибридизации. Получившийся единственный вид был назван « компиловидом ». [ 143 ]
Практические последствия
[ редактировать ]Биологам и защитникам природы необходимо классифицировать и идентифицировать организмы в ходе своей работы. Сложность надежного отнесения организмов к виду представляет собой угрозу достоверности результатов исследований, например, проведение измерений того, насколько распространен тот или иной вид в экосистеме, является спорным. Исследования, использующие концепцию филогенетических видов, выявили на 48% больше видов и, соответственно, меньшие популяции и ареалы, чем исследования, использующие нефилогенетические концепции; это было названо «таксономической инфляцией», [ 144 ] что может вызвать ложное впечатление об изменении количества исчезающих видов и, как следствие, политические и практические трудности. [ 145 ] [ 146 ] Некоторые наблюдатели утверждают, что существует внутренний конфликт между желанием понять процессы видообразования и необходимостью идентифицировать и классифицировать. [ 146 ]
Законы о сохранении во многих странах предусматривают специальные положения, предотвращающие вымирание видов. Зоны гибридизации между двумя видами, одним из которых является охраняемый, а другой нет, иногда приводили к конфликтам между законодателями, землевладельцами и защитниками природы. Одним из классических случаев в Северной Америке является случай охраняемой северной пятнистой совы , которая гибридизируется с незащищенной калифорнийской пятнистой совой и полосатой совой ; это привело к юридическим дебатам. [ 147 ] Утверждалось, что проблема видов создается в результате разнообразного использования концепции вида, и что решение состоит в том, чтобы отказаться от нее и всех других таксономических рангов и вместо этого использовать монофилетические группы без ранжирования - подход, которому способствует ФилоКодекс . Утверждалось также, что, поскольку виды несопоставимы, их подсчет не является достоверной мерой биоразнообразия ; были предложены альтернативные меры филогенетического биоразнообразия. [ 148 ] [ 89 ] [ 149 ]
История
[ редактировать ]Классические формы
[ редактировать ]В своей биологии Аристотель использовал термин γένος (генос) для обозначения вида, например птицы или рыбы , и εἶδος (эйдос) для обозначения конкретной формы внутри вида, например (внутри птиц) журавля , орла , ворона или воробей . Эти термины были переведены на латынь как «род» и «вид», хотя они не соответствуют названным таким образом терминам Линнея ; сегодня птицы — это класс , журавли — семейство , а вороны — род . Вид отличался своими атрибутами ; например, у птицы есть перья, клюв, крылья, яйцо с твердой скорлупой и теплая кровь. Форма отличалась тем, что была общей для всех ее членов: молодые люди наследовали любые вариации, которые они могли иметь от своих родителей. Аристотель считал, что все виды и формы различны и неизменны. Что еще более важно, в работах Аристотеля термины γένος (генос) и εἶδος (эйдос) относительны; таксон, который считается эйдосом в одном контексте, может считаться геносом в другом и далее подразделяться на эйдос (множественное число от эйдос). [ 150 ] [ 151 ] Его подход оставался влиятельным до эпохи Возрождения . [ 152 ] и до сих пор, в меньшей степени, сегодня. [ 153 ]
Фиксированные виды
[ редактировать ]

Когда наблюдатели раннего Нового времени начали разрабатывать системы организации живых существ, они поместили каждый вид животных или растений в определенный контекст. Многие из этих ранних схем разграничения теперь считались бы причудливыми: схемы включали кровное родство, основанное на цвете (все растения с желтыми цветами) или поведении (змеи, скорпионы и некоторые кусающие муравьи). Джон Рэй , английский натуралист, первым попытался дать биологическое определение вида в 1686 году следующим образом:
Мне не приходило в голову более надежного критерия определения вида, чем отличительные признаки, сохраняющиеся при размножении из семян. Таким образом, какие бы вариации ни происходили у особей или видов, если они происходят из семени одного и того же растения, это случайные вариации, а не такие, которые отличают вид... виды постоянно; один вид никогда не возникает из семени другого, и наоборот. [ 154 ]
В 18 веке шведский учёный Карл Линней классифицировал организмы по общим физическим характеристикам, а не просто по различиям. [ 155 ] Как и многие современные систематики, [ 156 ] [ 157 ] [ 158 ] он установил идею таксономической иерархии классификации, основанной на наблюдаемых характеристиках и призванной отражать естественные отношения. [ 159 ] [ 160 ] Однако в то время еще широко распространено мнение, что между видами (за исключением, возможно, представителей данного рода) не существует никакой органической связи. [ 93 ] какими бы похожими они ни казались. На эту точку зрения повлияло европейское научное и религиозное образование, которое считало, что таксоны были созданы Богом, образуя аристотелевскую иерархию, scala naturae или великую цепь бытия . Однако вне зависимости от того, предполагалось ли ее закрепить, лестница (лестница) по своей сути подразумевала возможность подъема. [ 161 ]
Мутабельность
[ редактировать ]Рассматривая свидетельства гибридизации, Линней признал, что виды не фиксированы и могут меняться; он не считал, что могут возникнуть новые виды, и придерживался точки зрения на божественно закрепленные виды, которые могут измениться в результате процессов гибридизации или акклиматизации. [ 162 ] К 19 веку натуралисты поняли, что виды могут со временем менять форму и что история планеты предоставила достаточно времени для серьезных изменений. Жан-Батист Ламарк в своей «Зоологической философии» 1809 года описал трансмутацию видов , предположив, что вид может меняться с течением времени, что радикально отличается от аристотелевского мышления. [ 163 ]
В 1859 году Чарльз Дарвин и Альфред Рассел Уоллес предоставили убедительное объяснение эволюции и образования новых видов. Дарвин утверждал, что именно популяции, а не отдельные особи, эволюционировали в результате естественного отбора из естественных вариаций среди особей. [ 164 ] Это потребовало нового определения вида. Дарвин пришел к выводу, что виды — это то, чем они кажутся: идеи, временно полезные для обозначения групп взаимодействующих особей, написав:
Я смотрю на термин «вид» как на термин, произвольно данный для удобства множеству особей, очень похожих друг на друга… Он по существу не отличается от слова «разнообразие», которое присваивается менее отчетливым и более изменчивым формам. Термин разнообразие, опять-таки, по сравнению с простыми индивидуальными различиями также применяется произвольно и ради удобства. [ 165 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Циммер, Карл (19 февраля 2024 г.). «Что такое вид? - Некоторые из самых известных видов на Земле могут быть не такими, какими кажутся» . Нью-Йорк Таймс . Архивировано из оригинала 19 февраля 2024 года . Проверено 19 февраля 2024 г.
- ^ Уилсон, Эдвард О. (3 марта 2018 г.). «Мнение: 8 миллионов видов, которых мы не знаем» . Нью-Йорк Таймс . ISSN 0362-4331 . Проверено 25 января 2020 г.
- ^ Боренштейн, С. (2019). «Отчет ООН: Люди ускоряют вымирание других видов» . Ассошиэйтед Пресс .
- ^ Перейти обратно: а б Мора, Камило; Титтенсор, Дерек П.; Адл, Сина; Симпсон, Аластер ГБ; Червь, Борис (23 августа 2011 г.). «Сколько видов существует на Земле и в океане?» . ПЛОС Биология . 9 (8): e1001127. дои : 10.1371/journal.pbio.1001127 . ПМК 3160336 . ПМИД 21886479 .
- ^ Перейти обратно: а б «Концепции видов» . Научный американец . 20 апреля 2012 года. Архивировано из оригинала 14 марта 2017 года . Проверено 14 марта 2017 г.
- ^ Маллет, Джеймс (1995). «Определение вида для современного синтеза». Тенденции в экологии и эволюции . 10 (7): 294–299. Бибкод : 1995TEcoE..10..294M . дои : 10.1016/0169-5347(95)90031-4 . ПМИД 21237047 .
- ^ Мастерс, JC; Спенсер, Х.Г. (1989). «Почему нам нужна новая концепция генетических видов». Систематическая зоология . 38 (3): 270–279. дои : 10.2307/2992287 . JSTOR 2992287 .
- ^ Перейти обратно: а б Мэйден, РЛ (1997). «Иерархия концепций видов: развязка проблемы видов». В Кларидже, штат Миссури; Дава, штат Ха; Уилсон, MR (ред.). Единицы биоразнообразия – виды на практике, специальный том 54 . Ассоциация систематиков.
- ^ Захос 2016 , с. 79.
- ^ Гудерс, Джон (1986). Полевой справочник Зимородка по птицам Британии и Ирландии . Книги Зимородка. п. 246. ИСБН 978-0-86272-139-8 .
- ^ Гулд, Стивен Джей (1980). «Куахог есть Куахог» . В: Большой палец панды: больше размышлений о естествознании . Нью-Йорк: WW Norton & Company. стр. 204–213 . ISBN 978-0-393-30023-9 .
- ^ Мейнард Смит, Джон (1989). Эволюционная генетика . Издательство Оксфордского университета. стр. 273–274. ISBN 978-0-19-854215-5 .
- ^ Русе, Майкл (1969). «Определения видов в биологии». Британский журнал философии науки . 20 (2): 97–119. дои : 10.1093/bjps/20.2.97 . JSTOR 686173 . S2CID 121580839 .
- ^ Левин, Ральф А. (1981). «Концепция трех видов». Таксон . 30 (3): 609–613. дои : 10.2307/1219942 . JSTOR 1219942 .
- ^ Кларидж, Дава и Уилсон 1997 , стр. 404
- ^ Гизелин, Майкл Т. (1974). «Радикальное решение проблемы видов» . Систематическая биология . 23 (4): 536–544. дои : 10.1093/sysbio/23.4.536 .
- ^ Кларидж и др.: 408–409.
- ^ Патерсон, HEH (1985). «Виды и видообразование». Врба, Е.С. (ред.). Монография № 4: Концепция распознавания видов . Претория: Музей Трансвааля.
- ^ Перейти обратно: а б Маллет, Джеймс (28 сентября 1999 г.). «Виды, понятия» (PDF) . В Калоу, П. (ред.). Энциклопедия экологии и природопользования . Блэквелл. стр. 709–711. ISBN 978-0-632-05546-3 . Архивировано (PDF) из оригинала 19 октября 2016 г.
- ^ Кулл, Калеви (2016). «Биосемиотическая концепция вида» . Биосемиотика . 9 : 61–71. дои : 10.1007/s12304-016-9259-2 . S2CID 18470078 . Архивировано из оригинала 7 февраля 2018 года.
- ^ Стакебрандт, Э.; Гебель, Б.М. (1994). «Таксономическое примечание: место для реассоциации ДНК-ДНК и анализа последовательности 16S рРНК в современном определении вида в бактериологии» . Международный журнал систематической бактериологии . 44 (4): 846–849. дои : 10.1099/00207713-44-4-846 .
- ^ Стакебрандт, Э.; Эберс, Дж. (2006). «Возвращение к таксономическим параметрам: потускневшие золотые стандарты» (PDF) . Микробиология сегодня . 33 (4): 152–155. Архивировано (PDF) из оригинала 25 марта 2018 г.
- ^ Земля, Мириам; Хаузер, Лорен; Джун, Се-Ран; Нукаев, Интават; Лёз, Майкл Р.; Ан, Тэ Хёк; Карпинец Татьяна; Лунд, Оле; Кора, Гурупрасед; Вассенаар, Труди; Пудель, Суреш; Ассери, Дэвид В. (2015). «Итоги секвенирования бактериального генома за 20 лет» . Функциональная и интегративная геномика . 15 (2): 141–161. дои : 10.1007/s10142-015-0433-4 . ПМЦ 4361730 . ПМИД 25722247 .
 Эта статья содержит цитаты из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0). Архивировано 16 октября 2017 г. по лицензии Wayback Machine .
Эта статья содержит цитаты из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0). Архивировано 16 октября 2017 г. по лицензии Wayback Machine .
- ^ Паркс, ДХ; Чувочина, М.; Шомей, Пенсильвания; Ринке, К.; Массиг, Эй Джей; Гугенгольц, П. (сентябрь 2020 г.). «Полная межвидовая таксономия бактерий и архей» . Природная биотехнология . 38 (9): 1079–1086. bioRxiv 10.1101/771964 . дои : 10.1038/s41587-020-0501-8 . ПМИД 32341564 . S2CID 216560589 .
- ^ Родригес-Р, Луис М.; Джайн, Чираг; Конрад, Рот Э.; Алуру, Шринивас; Константинидис, Константинос Т. (7 июля 2021 г.). «Ответ на: «Переоценка доказательств универсальной генетической границы между видами микробов» » . Природные коммуникации . 12 (1): 4060. Бибкод : 2021NatCo..12.4060R . дои : 10.1038/s41467-021-24129-1 . ПМЦ 8263725 . ПМИД 34234115 .
- ^ «Что такое штрих-кодирование ДНК?» . Штрих-код жизни. Архивировано из оригинала 1 июля 2017 года . Проверено 11 октября 2017 г.
- ^ Ратнасингем, Судживан; Хеберт, Пол Д.Н. (2007). «ЖИРНЫЙ: Система данных «Штрих-код жизни» (http://www.barcodinglife.org)» . Заметки по молекулярной экологии . 7 (3): 355–364. дои : 10.1111/j.1471-8286.2007.01678.x . ПМК 1890991 . ПМИД 18784790 .
- ^ Стокль, Марк (ноябрь – декабрь 2013 г.). «Штрих-кодирование ДНК готово к прорыву» . ДжинВотч . 26 (5). Архивировано из оригинала 3 апреля 2014 года . Проверено 25 марта 2018 г.
- ^ ДеСалле, Р.; Иган, Миннесота; Сиддалл, М. (2005). «Несвятая троица: таксономия, разграничение видов и штрих-кодирование ДНК» . Философские труды Королевского общества B: Биологические науки . 360 (1462): 1905–1916. дои : 10.1098/rstb.2005.1722 . ПМК 1609226 . ПМИД 16214748 .
- ^ Уитворт, ТЛ; Доусон, РД; Магалон, Х.; Бодри, Э. (2007). «Штрих-кодирование ДНК не может надежно идентифицировать виды рода мух Protocalliphora (Diptera: Calliphoridae)» . Труды Королевского общества B: Биологические науки . 274 (1619): 1731–1739. дои : 10.1098/rspb.2007.0062 . ПМЦ 2493573 . ПМИД 17472911 .
- ^ Перейти обратно: а б с Никсон, КЦ; Уиллер, QD (1990). «Расширение концепции филогенетического вида» . Кладистика . 6 (3): 211–223. дои : 10.1111/j.1096-0031.1990.tb00541.x . S2CID 84095773 .
- ^ Уилер, Квентин Д.; Платник, Норман И. 2000. Концепция филогенетического вида (в смысле Уилера и Платника). В: Wheeler & Meier, 2000 , стр. 55–69.
- ^ Жиро, Т.; Рефрегье, Г.; Ле Гак, М.; де Вьен, DM; Худ, Мэн (2008). «Видообразование грибов». Грибковая генетика и биология . 45 (6): 791–802. дои : 10.1016/j.fgb.2008.02.001 . ПМИД 18346919 .
- ^ Бернардо, Дж. (2011). «Критическая оценка значения и диагностируемости загадочного эволюционного разнообразия, а также его последствий для сохранения природы перед лицом изменения климата». В Ходкинсоне, Т.; Джонс, М.; Уолдрен, С.; Парнелл, Дж. (ред.). Изменение климата, экология и систематика. Специальная серия Ассоциации Систематики . Издательство Кембриджского университета. стр. 380–438. ISBN 978-0-521-76609-8 . .
- ^ Брауэр, Эндрю В.З. и Рэндалл Т. Шу. (2021). Биологическая систематика: принципы и приложения. Издательство Корнельского университета, Итака, Нью-Йорк.
- ^ Перейти обратно: а б Жиро, Т.; Рефрегье, Г.; Ле Гак, М.; де Вьен, DM; Худ, Мэн (2008). «Видообразование грибов». Грибковая генетика и биология . 45 (6): 791–802. дои : 10.1016/j.fgb.2008.02.001 . ПМИД 18346919 .
- ^ Перейти обратно: а б с д Тейлор, Дж.В.; Джейкобсон, диджей; Крокен, С.; Касуга, Т.; Гейзер, Д.М.; Хиббетт, Д.С.; Фишер, MC (2000). «Филогенетическое распознавание видов и концепции видов грибов». Грибковая генетика и биология . 31 (1): 21–32. дои : 10.1006/fgbi.2000.1228 . ПМИД 11118132 . S2CID 2551424 .
- ^ Тейлор, Дж.В.; Тернер, Э.; Таунсенд, JP; Деттман, младший; Джейкобсон, Д. (2006). «Эукариотические микробы, распознавание видов и географические пределы видов: примеры из царства грибов» . Философские труды Королевского общества B: Биологические науки . 361 (1475): 1947–1963. дои : 10.1098/rstb.2006.1923 . ПМК 1764934 . ПМИД 17062413 .
- ^ Захос 2016 , стр. 91–92.
- ^ Гроувс, К.; Грабб, П. 2011. Таксономия копытных. Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса.
- ^ Хеллер, Р.; Франдсен, П.; Лоренцен, Эд; Зигизмунд, HR (2013). «Неужели видов быков в два раза больше, чем мы думали?» . Систематическая биология . 62 (3): 490–493. дои : 10.1093/sysbio/syt004 . hdl : 10400.7/566 . ПМИД 23362112 .
- ^ Коттерилл, Ф.; Тейлор, П.; Гипполити, С.; и др. (2014). «Почему одного столетия фенетики достаточно: ответ на вопрос: «Неужели видов коров в два раза больше, чем мы думали?» " . Систематическая биология . 63 (5): 819–832. дои : 10.1093/sysbio/syu003 . ПМИД 24415680 .
- ^ Лапорт, LOF (1994). «Симпсон о видах». Журнал истории биологии . 27 (1): 141–159. дои : 10.1007/BF01058629 . ПМИД 11639257 . S2CID 34975382 .
- ^ Wheeler & Meier 2000 , стр. 70–92, 146–160, 198–208.
- ^ де Кейрос, Кевин (1998). «Общая концепция происхождения видов, видовые критерии и процесс видообразования». В DJ Howard; С.Х. Берлохер (ред.). Бесконечные формы: виды и видообразование . Издательство Оксфордского университета. стр. 57–75.
- ^ Ридли, Марк . «Идея вида». Эволюция (2-е изд.). Блэквелл Наука. п. 719. ИСБН 978-0-86542-495-1 .
- ^ Бейкер, Роберт Дж.; Брэдли, Роберт Д. (2006). «Видообразование млекопитающих и концепция генетического вида» . Журнал маммологии . 87 (4): 643–662. doi : 10.1644/06-MAMM-F-038R2.1 . ПМЦ 2771874 . ПМИД 19890476 .
- ^ Бейкер, Роберт Дж.; Брэдли, Роберт Д. (2006). «Видообразование млекопитающих и концепция генетического вида» . Журнал маммологии . 87 (4): 643–662. doi : 10.1644/06-MAMM-F-038R2.1 . ПМЦ 2771874 . ПМИД 19890476 .
- ^ Правительство Канады. Комитет по статусу дикой природы, находящейся под угрозой исчезновения, в Канаде. «Процесс и критерии оценки COSEWIC» . Cosepac.gc.ca. Архивировано из оригинала 12 апреля 2015 года . Проверено 7 апреля 2015 г.
- ^ ДеВеердт, Сара (29 июля 2002 г.). «Что на самом деле является эволюционно значимой единицей?» . Университет Вашингтона. Архивировано из оригинала 5 февраля 2017 года . Проверено 1 декабря 2016 г.
- ^ де Буффрениль, Вивиан; де Риклес, Арманд Дж; Зильберберг, Луиза; Падиан, Кевин; Лорен, Мишель; Кильяк, Александра (2021). Гистология скелета позвоночных и палеогистология . Бока-Ратон, Флорида: CRC Press. стр. xii + 825. ISBN 978-1351189576 .
- ^ «Хроновиды» . Оксфордский справочник. Архивировано из оригинала 1 октября 2014 года . Проверено 12 января 2018 г.
- ^ Карр, Стивен М. (2005). «Эволюционные виды и хроновиды» . Мемориальный университет Ньюфаундленда и Лабрадора. Архивировано из оригинала 10 марта 2016 года . Проверено 12 января 2018 г.
- ^ Дзик, Дж. (1985). «Типологические и популяционные концепции хроновидов: значение для биостратиграфии аммонитов» (PDF) . Acta Palaeontologica Polonica . 30 (1–2): 71–92. Архивировано (PDF) из оригинала 13 марта 2017 года.
- ^ О'Брайен, Майкл Дж.; Лайман, Р. Ли (2007). Применение эволюционной археологии: систематический подход . Спрингер. стр. 146–149. ISBN 978-0-306-47468-2 . Архивировано из оригинала 7 февраля 2018 года.
- ^ Ван Регенмортель, Марк Х.В. (2010). «Логические загадки и научные противоречия: природа видов, вирусов и живых организмов». Систематическая и прикладная микробиология . 33 (1): 1–6. Бибкод : 2010СиАпМ..33....1В . дои : 10.1016/j.syapm.2009.11.001 . ПМИД 20005655 .
- ^ ван Нимвеген, Эрик; Кратчфилд, Джеймс П.; Хюйнен, Мартейн (август 1999 г.). «Нейтральная эволюция мутационной устойчивости» . ПНАС . 96 (17): 9716–9720. arXiv : adap-org/9903006 . Бибкод : 1999PNAS...96.9716V . дои : 10.1073/pnas.96.17.9716 . ПМК 22276 . ПМИД 10449760 .
- ^ Вилке, Клаус О.; Ван, Цзя Лань; Офрия, Чарльз; Ленски, Ричард Э.; Адами, Кристоф (2001). «Эволюция цифровых организмов с высокой скоростью мутаций приводит к выживанию самых плоских» (PDF) . Природа . 412 (6844): 331–333. Бибкод : 2001Natur.412..331W . дои : 10.1038/35085569 . ПМИД 11460163 . S2CID 1482925 .
- ^ Елена, Сан-Франциско; Агудело-Ромеро, П.; Карраско, П.; и др. (2008). «Экспериментальная эволюция растительных РНК-вирусов» . Наследственность . 100 (5): 478–483. дои : 10.1038/sj.hdy.6801088 . ПМК 7094686 . ПМИД 18253158 .
- ^ Кинг, Эндрю MQ; Лефковиц, Э.; Адамс, MJ; Карстенс, Э.Б. (2012). Классификация и номенклатура вирусов по таксономии вирусов: девятый отчет Международного комитета по таксономии вирусов . Эльзевир. ISBN 978-0-12-384684-6 . Архивировано из оригинала 10 декабря 2017 года . Проверено 13 апреля 2017 г. .
- ^ Фоке, СМ; Фаргетт, Д. (2005). «Международный комитет по таксономии вирусов и 3142 неустановленных видов» . Вирусологический журнал . 2:64 . дои : 10.1186/1743-422X-2-64 . ПМК 1208960 . ПМИД 16105179 .
- ^ Гиббс, Эй Джей (2013). «Вирусная систематика нуждается в генеральной чистке; эра ее исследований закончилась» . Вирусологический журнал . 10 :254. дои : 10.1186/1743-422X-10-254 . ПМЦ 3751428 . ПМИД 23938184 .
- ^ Майр, Эрнст (1942). Систематика и происхождение видов . Нью-Йорк: Издательство Колумбийского университета.
- ^ Уиллер, стр. 17–29.
- ^ Перейти обратно: а б с д де Кейроз, К. (2005). «Эрнст Майр и современная концепция вида» . ПНАС . 102 (Приложение 1): 6600–6607. Бибкод : 2005PNAS..102.6600D . дои : 10.1073/pnas.0502030102 . ПМЦ 1131873 . ПМИД 15851674 .
- ^ Хопф, ФА; Хопф, ФРВ (1985). «Роль эффекта Аллее в упаковке видов». Теоретическая популяционная биология . 27 (1): 27–50. Бибкод : 1985TPBio..27...27H . дои : 10.1016/0040-5809(85)90014-0 .
- ^ Бернштейн, Х.; Байерли, ХК; Хопф, ФА; Мишо, RE (1985). «Секс и возникновение видов». Журнал теоретической биологии . 117 (4): 665–690. Бибкод : 1985JThBi.117..665B . дои : 10.1016/S0022-5193(85)80246-0 . ПМИД 4094459 .
- ^ Бернштейн, Кэрол; Бернштейн, Харрис (1991). Старение, пол и восстановление ДНК . Бостон: Академическая пресса. ISBN 978-0-12-092860-6 .
- ^ Мишод, Ричард Э. (1995). Эрос и эволюция: естественная философия секса . Аддисон-Уэсли. ISBN 978-0-201-44232-8 .
- ^ Ханаге, Уильям П. (2013), «Возвращение к нечетким видам», BMC Biology , 11 (41): 41, doi : 10.1186/1741-7007-11-41 , PMC 3626887 , PMID 23587266
- ^ Кох, Х. (2010). «Сочетание морфологии и штрих-кодирования ДНК решает таксономию западно-малагасийской Liotrigona Moure, 1961» (PDF) . Африканские беспозвоночные . 51 (2): 413–421. дои : 10.5733/afin.051.0210 . Архивировано (PDF) из оригинала 1 декабря 2016 года.
- ^ Де Кейрос, К. (2007). «Концепции видов и разграничение видов» . Систематическая биология . 56 (6): 879–886. дои : 10.1080/10635150701701083 . ПМИД 18027281 .
- ^ Фрейзер, К.; Альм, Э.Дж.; Польц, МФ; Спратт, Б.Г.; Ханаге, WP (2009). «Проблема видов бактерий: понимание генетического и экологического разнообразия». Наука . 323 (5915): 741–746. Бибкод : 2009Sci...323..741F . дои : 10.1126/science.1159388 . ПМИД 19197054 . S2CID 15763831 .

- ^ «Дарвин 1859 г., глава II, стр. 59» . Дарвин-онлайн.org.uk. Архивировано из оригинала 21 октября 2012 года . Проверено 25 ноября 2012 г.
- ^ Геверс, Дирк; Коэн, Фредерик М.; Лоуренс, Джеффри Г.; Спратт, Брайан Г.; Коэнье, Том; Фейл, Эдвард Дж.; Штакебрандт, Эрко; Де Пер, Ив Ван; Вандам, Питер; Томпсон, Фабиан Л.; Качели, Жан (2005). «Мнение: переоценка видов прокариот» . Обзоры природы Микробиология . 3 (9): 733–9. дои : 10.1038/nrmicro1236 . ПМИД 16138101 . S2CID 41706247 .
- ^ Темплтон, Арканзас (1989). «Значение видов и видообразований: генетическая перспектива». Ин Отте, Д.; Эндлер, Дж. А. (ред.). Видообразование и его последствия . Синауэр Ассошиэйтс. стр. 3–27.
- ^ Эдвард Г. Рики; Фахри А. Баззаз (2005). Репродуктивное распределение у растений . Академическая пресса. п. 99. ИСБН 978-0-12-088386-8 . Архивировано из оригинала 17 июня 2013 года.
- ^ Росселло-Мора, Рамон; Аманн, Рудольф (январь 2001 г.). «Видовая концепция прокариот» . Обзоры микробиологии FEMS . 25 (1): 39–67. дои : 10.1111/j.1574-6976.2001.tb00571.x . ПМИД 11152940 .
- ^ Андино, Рауль; Доминго, Эстебан (2015). «Вирусные квазивиды» . Вирусология . 479–480: 46–51. дои : 10.1016/j.virol.2015.03.022 . ПМЦ 4826558 . ПМИД 25824477 .
- ^ Бибрихер, КК; Эйген, М. (2006). «Что такое квазивид?». Квазивиды: концепция и значение для вирусологии . Актуальные темы микробиологии и иммунологии. Том. 299. Спрингер. стр. 1–31. дои : 10.1007/3-540-26397-7_1 . ISBN 978-3-540-26397-5 . ПМИД 16568894 .
- ^ Тойеман, А.Е. (2009). «Концепция вида в палеонтологии» . Геологический журнал . 61 (8): 355–360. Бибкод : 1924ГеоМ...61..355Т . дои : 10.1017/S001675680008660X . S2CID 84339122 . Архивировано из оригинала 14 марта 2017 года.
- ^ Захос 2016 , с. 101.
- ^ Захос 2016 , стр. 156–157.
- ^ Лар, диджей; Дом смеха, HD; Оливерио, AM; Гао, Ф.; Кац, Луизиана (2014). «Как противоречивая морфологическая и молекулярная эволюция микроорганизмов может пересмотреть наши представления о биоразнообразии на Земле» . Биоэссе . 36 (10): 950–959. doi : 10.1002/bies.201400056 . ПМЦ 4288574 . ПМИД 25156897 .
- ^ Перейти обратно: а б с Мельчер, Ульрих (2001). «Молекулярная генетика: Горизонтальный перенос генов» . Государственный университет Оклахомы. Архивировано из оригинала 4 марта 2016 года.
- ^ Баптест, Э.; и др. (май 2005 г.). «Действительно ли филогения ортологичных генов поддерживает древовидное мышление?» . Эволюционная биология BMC . 5 (33): 33. Бибкод : 2005BMCEE...5...33B . дои : 10.1186/1471-2148-5-33 . ПМЦ 1156881 . ПМИД 15913459 .
- ^ Уильямсон, Дэвид И. (2003). Происхождение личинок . Клювер. ISBN 978-1-4020-1514-4 .
- ^ Мишлер, Брент Д. (2022). «Экология, эволюция и систематика в поствидовом мире». В Уилкинсе, Джон С.; Захос, Фрэнк Э.; Павлинов, Игорь (ред.). Проблемы видов и за их пределами: современные проблемы философии и практики . Бока-Ратон: CRC Press, Taylor & Francisco Group. ISBN 978-0-367-85560-4 . OCLC 1273727987 .
- ^ Перейти обратно: а б Мишлер, Брент Д. (1999). «Избавление от видов?». В Уилсоне, Р. (ред.). Виды: Новые междисциплинарные очерки (PDF) . МТИ Пресс. стр. 307–315. ISBN 978-0262731232 . Архивировано (PDF) из оригинала 10 января 2018 г.
- ^ Зальцбургер, Вальтер (ноябрь 2018 г.). «Понимание взрывной диверсификации посредством геномики цихлидных рыб» . Обзоры природы Генетика . 19 (11): 705–717. дои : 10.1038/s41576-018-0043-9 . ISSN 1471-0064 . ПМИД 30111830 . S2CID 52008986 .
- ^ Перейти обратно: а б с Уилкинс, Джон С. (2022). «Добрый вид». В Уилкинсе, Джон С.; Захос, Фрэнк Э.; Павлинов, Игорь (ред.). Проблемы видов и за их пределами: современные проблемы философии и практики . Бока-Ратон: CRC Press, Taylor & Francisco Group. ISBN 978-0-367-85560-4 . OCLC 1273727987 .
- ^ Мишлер, Брент Д .; Уилкинс, Джон С. (2018). «Охота на SNARC: странное решение проблемы видов» . Философия, теория и практика биологии . 10 (20220112). Библиотека Мичиганского университета. дои : 10.3998/ptpbio.16039257.0010.001 . hdl : 2027/spo.16039257.0010.001 .
- ^ Перейти обратно: а б с д Лорен, Мишель (3 августа 2023 г.). Появление PhyloCode: продолжающаяся эволюция биологической номенклатуры . ЦРК Пресс. п. xv + 209. doi : 10.1201/9781003092827 . ISBN 978-1-003-09282-7 .
- ^ Китчер, Филип (июнь 1984 г.). "Разновидность". Философия науки . 51 (2): 308–333. дои : 10.1086/289182 . S2CID 224836299 .
Я защищаю точку зрения на видовую категорию, плюралистический реализм, который призван отдать должное взглядам многих различных групп систематиков.
- ^ Плейжель, Ф.; Роуз, GW (22 марта 2000 г.). «Наименее инклюзивная таксономическая единица: новая таксономическая концепция биологии» . Труды Лондонского королевского общества. Серия Б: Биологические науки . 267 (1443): 627–630. дои : 10.1098/rspb.2000.1048 . ISSN 0962-8452 . ПМК 1690571 . ПМИД 10787169 .
- ^ Хейвуд, В.Х. (1962). «Совокупность видов» в теории и практике». В Хейвуде, штат Вирджиния; Лёве, А. (ред.). Симпозиум по биосистематике, Монреаль, октябрь 1962 г. стр. 26–36.
- ^ Пиментел, Дэвид (2014). Биологические инвазии: экономические и экологические издержки чужеродных растений, животных и микробов . ЦРК Пресс. п. 92. ИСБН 978-1-4200-4166-8 . Архивировано из оригинала 7 февраля 2018 года.
- ^ Джарвис, CE (1992). «Семьдесят два предложения по сохранению типов избранных родовых названий Линнея, отчет подкомитета 3C по лектотипизации родовых названий Линнея». Таксон . 41 (3): 552–583. дои : 10.2307/1222833 . JSTOR 1222833 .
- ^ Витцель, Хакан (1999). «Вариации ДНК хлоропластов и эволюция сетчатой структуры в половых и апомиктических секциях одуванчиков». Молекулярная экология . 8 (12): 2023–2035. Бибкод : 1999MolEc...8.2023W . дои : 10.1046/j.1365-294x.1999.00807.x . ПМИД 10632854 . S2CID 25180463 .
- ^ Дейк, Питер Дж. Ван (2003). Экологические и эволюционные возможности апомиксиса: идеи и Chondrilla . « Taraxacum Философские труды Королевского общества Б. 358 (1434): 1113–1121. дои : 10.1098/rstb.2003.1302 . ПМК 1693208 . ПМИД 12831477 .
- ^ Маллет, Джеймс ; Бельтран, М.; Нойкирхен, В.; Линарес, М. (2007). «Естественная гибридизация бабочек-геликониин: граница видов как континуум» . Эволюционная биология BMC . 7 (1): 28. Бибкод : 2007BMCEE...7...28M . дои : 10.1186/1471-2148-7-28 . ПМК 1821009 . ПМИД 17319954 .
- ^ Рон, Сантьяго; Каминер, Марсель (2014). «Систематика древесных лягушек комплекса видов Hypsiboas calcaratus и Hypsiboas fasciatus (Anura, Hylidae) с описанием четырех новых видов» . ZooKeys (370): 1–68. Бибкод : 2014ЗооК..370....1С . дои : 10.3897/zookeys.370.6291 . ПМК 3904076 . ПМИД 24478591 .
- ^ Гемл, Дж.; Таллосс, RE; Лаурсен, Джорджия; Сасанова Н.А.; Тейлор, Д.Л. (2008). «Доказательства сильной меж- и внутриконтинентальной филогеографической структуры у Amanita muscaria , распространяемого ветром эктомикоризного базидиомицета». Молекулярная филогенетика и эволюция . 48 (2): 694–701. Бибкод : 2008МОЛПЭ..48..694Г . дои : 10.1016/j.ympev.2008.04.029 . ПМИД 18547823 . S2CID 619242 .
- ^ «Определение вида» . Калифорнийский университет в Беркли. Архивировано из оригинала 13 марта 2017 года . Проверено 12 марта 2017 г.
- ^ Захос 2016 , с. 188.
- ^ Стамос, Дэвид Н. (2003). Проблема видов: биологические виды, онтология и метафизика биологии . Лексингтонские книги. п. 330. ИСБН 978-0-7391-6118-0 . Архивировано из оригинала 12 марта 2017 года.
- ^ Мориц, К.; Шнайдер, CJ; Уэйк, Д.Б. (1992). «Эволюционные взаимоотношения внутри комплекса Ensatina eschscholtzii подтверждают интерпретацию кольцевых видов» (PDF) . Систематическая биология . 41 (3): 273–291. дои : 10.1093/sysbio/41.3.273 . Архивировано (PDF) из оригинала 12 января 2018 г.
- ^ Ирвин, Делавэр; Бенш, Стаффан; Ирвин, Джессика Х.; Прайс, Тревор Д. (2005). «Видообразование по расстоянию у кольцевых видов». Наука . 307 (5708): 414–6. Бибкод : 2005Sci...307..414I . дои : 10.1126/science.1105201 . ПМИД 15662011 . S2CID 18347146 .
- ^ Мартенс, Йохен; Пакерт, Мартин (2007). «Кольцевые виды - существуют ли они у птиц?». Зоологический вестник . 246 (4): 315–324. Бибкод : 2007ЗооАн.246..315М . дои : 10.1016/j.jcz.2007.07.004 .
- ^ Алькайд, М.; Скордато, ESC; Цена, ТД; Ирвин, Делавэр (2014). «Геномная дивергенция в кольцевом комплексе видов». Природа . 511 (7507): 83–85. Бибкод : 2014Natur.511...83A . дои : 10.1038/nature13285 . hdl : 10261/101651 . ПМИД 24870239 . S2CID 4458956 .
- ^ Либерс, Дорит; Книфф, Питер де; Хельбиг, Андреас Дж. (2004). «Комплекс серебристых чаек не является кольцевым видом» . Труды Лондонского королевского общества. Серия Б: Биологические науки . 271 (1542): 893–901. дои : 10.1098/rspb.2004.2679 . ПМК 1691675 . ПМИД 15255043 .
- ^ Хайтон, Р. (1998). «Является ли Ensatina eschscholtzii кольцевым видом?». Герпетологика . 54 (2): 254–278. JSTOR 3893431 .
- ^ «Слово о названиях видов…» Смитсоновская морская станция в Форт-Пирсе. Архивировано из оригинала 24 марта 2017 года . Проверено 11 марта 2017 г.
- ^ Хоун, Дэйв (19 июня 2013 г.). «Что в имени? Почему важны научные названия» . Хранитель . Архивировано из оригинала 15 февраля 2017 года . Проверено 11 марта 2017 г.
- ^ Лоули, Джонатан В.; Гамеро-Мора, Эдгар; Маронна, Максимилиано М.; Кьяверано, Лучано М.; Стампар, Сержио Н.; Хопкрофт, Рассел Р.; Коллинз, Аллен Г.; Морандини, Андре К. (29 сентября 2022 г.). «Морфология не всегда полезна для диагностики, и это нормально: гипотезы о видах не должны быть привязаны к классу данных. Ответ Брауну и Гиббонсу (S Afr J Sci. 2022; 118 (9/10), Art. # 12590). " . Южноафриканский научный журнал . 118 (9/10). дои : 10.17159/sajs.2022/14495 . S2CID 252562185 .
- ^ Один из примеров аннотации статьи, в которой назван новый вид, можно найти по адресу. Веллнер, С.; Лоддерс, Н.; Кемпфер, П. (2012). « Mmethylobacterium cerastii sp. nov., новый вид, выделенный с поверхности листьев Cerastium holosteoides » . Международный журнал систематической и эволюционной микробиологии . 62 (Часть 4): 917–924. дои : 10.1099/ijs.0.030767-0 . ПМИД 21669927 .
- ^ Хичкок, А.С. (1921), «Понятие типа в систематической ботанике» , Американский журнал ботаники , 8 (5): 251–255, doi : 10.2307/2434993 , JSTOR 2434993
- ^ Николсон, Дэн Х. «Ботаническая номенклатура, типы и стандартные справочные материалы» . Смитсоновский национальный музей естественной истории, отдел ботаники. Архивировано из оригинала 16 сентября 2015 года . Проверено 17 ноября 2015 г.
- ^ «Международный кодекс зоологической номенклатуры, рекомендация 25C» . Архивировано из оригинала 8 июня 2011 года . Проверено 18 июня 2011 г.
- ^ Перейти обратно: а б с д и Уинстон, Джудит Э. (1999). Описание видов. Практическая таксономическая процедура для биологов . Нью-Йорк: Издательство Колумбийского университета. стр. 141–144.
- ^ «Главная – Таксономия – NCBI» . Ncbi.nlm.nih.gov. 19 октября 2012 года. Архивировано из оригинала 7 февраля 2018 года . Проверено 25 ноября 2012 г.
- ^ «Организмы KEGG: полные геномы» . Геном.jp. Архивировано из оригинала 14 октября 2012 года . Проверено 25 ноября 2012 г.
- ^ «Таксономия» . Uniprot.org . Архивировано из оригинала 27 ноября 2012 года . Проверено 25 ноября 2012 г.
- ^ «ИТИС: Человек разумный» . Каталог жизни. Архивировано из оригинала 12 марта 2017 года . Проверено 11 марта 2017 г.
- ^ Симпсон, Джордж Гейлорд (1945). «Принципы классификации и классификация млекопитающих». Бюллетень Американского музея естественной истории . 85:23 .
- ^ Чейз, Боб (2005). «Выскочка Антихрист». Журнал исторической мастерской . 60 (1): 202–206. дои : 10.1093/hwj/dbi042 . S2CID 201790420 .
- ^ Лорен, Мишель (23 июля 2023 г.). «Филокод: логический результат тысячелетней эволюции биологической номенклатуры?» . Зоологика Скрипта . 52 (6): 543–555. дои : 10.1111/zsc.12625 . ISSN 0300-3256 . S2CID 260224728 .
- ^ Уилсон, Филип (2016). «sensu stricto, sensu lato» . AZ дерева терминов . Архивировано из оригинала 10 января 2018 года . Проверено 9 января 2018 г.
- ^ «Глоссарий: чувственность» . Международная комиссия по зоологической номенклатуре . Архивировано из оригинала 2 августа 2017 года . Проверено 9 января 2018 г.
- ^ Перейти обратно: а б Бартон, Нью-Хэмпшир (июнь 2010 г.). «Какую роль играет естественный отбор в видообразовании?» . Философские труды Королевского общества Б. 365 (1547): 1825–1840. дои : 10.1098/rstb.2010.0001 . ПМЦ 2871892 . ПМИД 20439284 .
- ^ Перейти обратно: а б Во, Феликс; Тревик, Стивен А.; Морган-Ричардс, Мэри (2017). «Видообразование в зазеркалье» . Биологический журнал Линнеевского общества . 120 (2): 480–488. дои : 10.1111/bij.12872 .
- ^ Захос 2016 , стр. 77–96.
- ^ Кук, Оратор Ф. (30 марта 1906 г.). «Факторы видообразования» . Наука . 23 (587): 506–507. Бибкод : 1906Sci....23..506C . дои : 10.1126/science.23.587.506 . ПМИД 17789700 .
- ^ Кук, оратор Ф. (ноябрь 1908 г.). «Эволюция без изоляции». Американский натуралист . 42 (503): 727–731. дои : 10.1086/279001 . S2CID 84565616 .
- ^ Виа, Сара (16 июня 2009 г.). «Естественный отбор в действии при видообразовании» . Учеб. Натл. акад. наук. США 106 (Приложение 1): 9939–9946. Бибкод : 2009PNAS..106.9939V . дои : 10.1073/pnas.0901397106 . ПМК 2702801 . ПМИД 19528641 .
- ^ Майр, Эрнст (1982). «Видообразование и макроэволюция» . Эволюция . 36 (6): 1119–1132. дои : 10.1111/j.1558-5646.1982.tb05483.x . ПМИД 28563569 . S2CID 27401899 .
- ^ Пенниси, Элизабет (2004). «Исследователи обмениваются мнениями об обмене генами» (PDF) . Наука . 334–335: 335. Архивировано из оригинала (PDF) 18 февраля 2006 г.
- ^ Жахыбаева Ольга; Питер Гогартен, Дж. (2004). «Кладогенез, слияние и эволюция трех областей жизни» (PDF) . Тенденции в генетике . 20 (4): 182–187. дои : 10.1016/j.tig.2004.02.004 . ПМИД 15041172 . Архивировано (PDF) из оригинала 26 марта 2009 г.
- ^ Вентон, Даниэль (2017). «Основной момент: применение концепции биологических видов ко всей жизни» . Геномная биология и эволюция . 9 (3): 502–503. дои : 10.1093/gbe/evx045 . ПМЦ 5381533 . ПМИД 28391326 .
- ^ Бобей, Луи-Мари; Охман, Ховард (2017). «Биологические виды универсальны во всех сферах жизни» . Геномная биология и эволюция . 9 (3): 491–501. дои : 10.1093/gbe/evx026 . ПМЦ 5381558 . ПМИД 28186559 .
- ^ Кунин, МЫ; Гастон, Кевин, ред. (1996). Биология редкости: причины и последствия редких и общих различий . Спрингер. ISBN 978-0-412-63380-5 . Архивировано из оригинала 5 сентября 2015 года.
- ^ Стернс, Беверли Петерсон; Стернс, Стивен К. (2000). Смотрю с края вымирания . Нью-Хейвен, Лондон: Издательство Йельского университета . п. предисловие х. ISBN 978-0-300-08469-6 .
- ^ Захос 2016 , с. 82.
- ^ Захос, Фрэнк Э. (2015). «Таксономическая инфляция, концепция филогенетических видов и линии на Древе жизни - предостерегающий комментарий о разделении видов» . Журнал зоологической систематики и эволюционных исследований . 53 (2): 180–184. дои : 10.1111/jzs.12088 .
- ^ Агапов, Поль-Майкл; Бининда-Эмондс, Олаф Р.П.; Крэндалл, Кейт А .; Гиттлман, Джон Л.; Мейс, Джорджина М.; Маршалл, Джонатон К.; Первис, Энди (2004). «Влияние концепции видов на исследования биоразнообразия» (PDF) . Ежеквартальный обзор биологии . 79 (2): 161–179. CiteSeerX 10.1.1.535.2974 . дои : 10.1086/383542 . JSTOR 10.1086/383542 . ПМИД 15232950 . S2CID 2698838 . Архивировано (PDF) из оригинала 11 января 2018 г.
- ^ Перейти обратно: а б Привет, Джоди (июль 2001 г.). «Разум видовой проблемы». Тенденции экологии и эволюции . 16 (7): 326–329. дои : 10.1016/S0169-5347(01)02145-0 . ПМИД 11403864 .
- ^ Хейг, Сьюзен М.; Аллендорф, ФРВ (2006). «Гибриды и политика» . В Скотте, Дж. Майкле; Гобл, Д.Д.; Дэвис, Фрэнк В. (ред.). Закон об исчезающих видах, тридцать лет, том 2: Сохранение биоразнообразия в ландшафтах, находящихся под контролем человека . Вашингтон: Island Press. стр. 150–163. Архивировано из оригинала 7 февраля 2018 года.
- ^ Фейт, Дэниел П. (1 января 1992 г.). «Оценка сохранения и филогенетическое разнообразие» . Биологическая консервация . 61 (1): 1–10. Бибкод : 1992BCons..61....1F . дои : 10.1016/0006-3207(92)91201-3 . ISSN 0006-3207 .
- ^ Вейн-Райт, Род-Айленд; Хамфрис, CJ; Уильямс, PH (1991). «Что защищать? – систематика и муки выбора». Биологическая консервация . 55 (3): 235–254. Бибкод : 1991BCons..55..235V . дои : 10.1016/0006-3207(91)90030-D .
- ^ Пеллегрин, Пьер (1986). Классификация животных Аристотеля: биология и концептуальное единство аристотелевского корпуса . Беркли, Калифорния: Univ. Калифорнии Пр. п. xiv + 235. ISBN 0520055020 .
- ^ Лорен, Мишель; Хумар, Марсель (10 января 2022 г.). «Филогенетический сигнал у персонажей «Истории животных» Аристотеля» . Comptes Rendus Palevol (на французском языке) (1). дои : 10.5852/cr-palevol2022v21a1 . S2CID 245863171 .
- ^ Леруа, Арман Мари (2014). Лагуна: как Аристотель изобрел науку . Блумсбери. стр. 88–90. ISBN 978-1-4088-3622-4 .
- ^ Ригато, Эмануэле; Минелли, Алессандро (28 июня 2013 г.). «Великая цепь бытия все еще здесь» . Эволюция: образование и информационно-пропагандистская деятельность . 6 (1): 18. дои : 10.1186/1936-6434-6-18 . ISSN 1936-6434 .
- ^ Рэй, Джон (1686). Всеобщая история растений, Том I, Libr. Я. п. Глава. XX, стр. 40. , цит. Майр, Эрнст (1982). Рост биологической мысли: разнообразие, эволюция и наследственность . Белнап Пресс. п. 256 . ISBN 9780674364455 .
- ^ Дэвис, PH; Хейвуд, В.Х. (1973). Принципы систематики покрытосеменных . Хантингтон, Нью-Йорк: Издательская компания Роберта Э. Кригера. п. 17.
- ^ Маньоль, Питер (1689). Продром общей истории растений, в котором семейства растений расположены таблицами (на латыни). Печ. п. 79.
- ^ Турнефор, Жозеф Питтон де (1656-1708) Автор текста (1694). Элементы ботаники, или Метод познания растений. И. [Текст.] / . Г-н Питтон Турнефор... [Т. I-III] . Париж: L'Imprimerie Royale. п. 562.
{{cite book}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Жюссье, Антуан-Лоран де (1789). Антониус Лаврентий де Жюссье ... Роды растений расположены в соответствии с их естественным порядком: по методу, установленному в саду Парижского региона в 1800 году. (на латыни). У вдовы Эриссант, печатника, через Нову БМ под знаком Золотого Креста. И Теофил Барруа на берег августинцев. п. 498
- ^ Раскрытие, Джеймс Л.; Прингл, Джеймс С. (1993). «7. Таксономическая ботаника и флористика». Флора Северной Америки . Издательство Оксфордского университета. стр. 160–161. ISBN 978-0-19-505713-3 .
- ^ Симпсон, Джордж Гейлорд (1961). Принципы систематики животных . Издательство Колумбийского университета. стр. 56–57.
- ^ Махони, Эдвард П. (1987). «Лавджой и иерархия бытия». Журнал истории идей . 48 (2): 211–230. дои : 10.2307/2709555 . JSTOR 2709555 .
- ^ «Карл Линней (1707–1778)» . UCMP Беркли. Архивировано из оригинала 30 апреля 2011 года . Проверено 24 января 2018 г.
- ^ Гулд, Стивен Джей (2002). Структура эволюционной теории . Гарвард: Белкнап Гарвард. стр. 170–197 . ISBN 978-0-674-00613-3 .
- ^ Боулер, Питер Дж. (2003). Эволюция: история идеи (3-е изд.). Беркли, Калифорния: Издательство Калифорнийского университета . стр. 177–223 и пассим . ISBN 978-0-520-23693-6 .
- ^ Менанд, Луи (2001). Метафизический клуб: история идей в Америке . Фаррар, Штраус и Жиру. стр. 123–124 . ISBN 978-0-374-70638-8 .
Источники
[ редактировать ]- Кларидж, МФ; Дава, штат Ха; Уилсон, MR, ред. (1997). Виды: единицы биоразнообразия . Чепмен и Холл . ISBN 978-0-412-63120-7 .
- Уилер, Квентин ; Мейер, Рудольф, ред. (2000). Концепции видов и филогенетическая теория: дискуссия . Издательство Колумбийского университета . ISBN 978-0-231-10143-1 .
- Захос, Фрэнк Э. (2016). Концепции видов в биологии: историческое развитие, теоретические основы и практическая значимость . Спрингер . ISBN 978-3-319-44964-7 .
Внешние ссылки
[ редактировать ]- Wikispecies — бесплатный каталог видов , который может редактировать каждый. Фонда Викимедиа
- Штрих-кодирование видов
- Каталог жизни
- Европейские названия видов на языке Линнея, чешском, английском, немецком и французском языках.
- Запись «Виды» в Стэнфордской энциклопедии философии.
- Визуальная Такса