Морская жизнь
Эта статья содержит слишком много изображений для ее общего объема . ( декабрь 2023 г. ) |
| Часть серии обзоров по |
| Морская жизнь |
|---|
 |
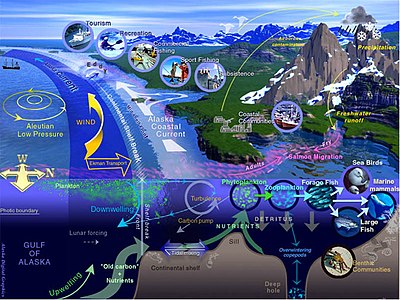

Морская жизнь , морская жизнь или жизнь океана — это растения , животные и другие организмы , которые живут в соленой воде морей океанов или в или солоноватой воде прибрежных эстуариев . На фундаментальном уровне морская жизнь влияет на природу планеты. Морские организмы, в основном микроорганизмы , производят кислород и связывают углерод . Морская жизнь частично формирует и защищает береговую линию, а некоторые морские организмы даже помогают создавать новые суши (например, рифы ) коралловые .
Большинство форм жизни первоначально развилось в морской среде обитания . По объему океаны обеспечивают около 90% жизненного пространства на планете. [ 2 ] Первые позвоночные появились в виде рыб . [ 3 ] которые живут исключительно в воде. Некоторые из них превратились в амфибий , которые проводят часть своей жизни в воде, а часть — на суше. Одна группа амфибий превратилась в рептилий и млекопитающих, а несколько подгрупп каждой из них вернулись в океан в виде морских змей , морских черепах , тюленей , ламантинов и китов . Растительные формы, такие как ламинария и другие водоросли, растут в воде и являются основой некоторых подводных экосистем. Планктон океана образует общую основу пищевой цепи , особенно фитопланктон , который является ключевыми первичными продуцентами .
Морские беспозвоночные демонстрируют широкий спектр модификаций, позволяющих выжить в водах с низким содержанием кислорода, включая дыхательные трубки, такие как сифоны моллюсков . У рыб есть жабры вместо легких , хотя у некоторых видов рыб, например двоякодышащих , есть и то, и другое. Морским млекопитающим (например, дельфинам, китам, выдрам и тюленям) необходимо периодически всплывать на поверхность, чтобы подышать воздухом.
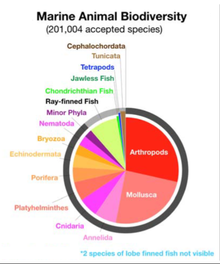
По состоянию на 2023 год [update]было зарегистрировано более 242 000 морских видов , и, возможно, два миллиона морских видов еще не зарегистрированы. В среднем описывается 2332 новых вида в год. [ 4 ] [ 5 ]
Размеры морских видов варьируются от микроскопических, таких как фитопланктон , размер которых может достигать 0,02 микрометра , до огромных китообразных, таких как синий кит – самое большое известное животное, достигающее 33 м (108 футов) в длину. [ 6 ] [ 7 ] Морские микроорганизмы, включая протистов , бактерии и связанные с ними вирусы , по разным оценкам составляют около 70% [ 8 ] или около 90% [ 9 ] [ 1 ] от общей морской биомассы . Морская жизнь изучается с научной точки зрения как в морской биологии , так и в биологической океанографии . Термин «морской» происходит от латинского mare , что означает «море» или «океан».
Вода
[ редактировать ]
Без воды нет жизни. [ 10 ] Его называют универсальным растворителем из-за его способности растворять многие вещества. [ 11 ] [ 12 ] и как растворитель жизни . [ 13 ] Вода — единственное общее вещество, которое существует в твердом , жидком и газообразном состоянии в условиях, нормальных для жизни на Земле. [ 14 ] Лауреат Нобелевской премии Альберт Сент - Дьёрдьи называл воду «матерью и матрицей» : матерью и лоном жизни. [ 15 ]

Обилие поверхностных вод на Земле является уникальной особенностью Солнечной системы . Земли Гидросфера состоит в основном из океанов, но технически включает в себя все водные поверхности мира, включая внутренние моря, озера, реки и подземные воды на глубину до 2000 метров (6600 футов). Самое глубокое подводное место — это Бездна Челленджера в Марианской впадине в Тихом океане , глубина которой составляет 10 900 метров (6,8 миль). [ примечание 1 ] [ 16 ]
Условно планета разделена на пять отдельных океанов, однако все эти океаны соединяются в единый мировой океан . [ 17 ] Масса этого мирового океана составляет 1,35 × 10 18 метрические тонны или около 1/4400 общей массы Земли. Мировой океан занимает площадь 3,618 × 10. 8 км 2 со средней глубиной 3682 м , что дает расчетный объем 1,332 × 10 9 км 3 . [ 18 ] Если бы вся поверхность земной коры находилась на одной высоте с гладкой сферой, глубина образовавшегося мирового океана составила бы около 2,7 километров (1,7 мили). [ 19 ] [ 20 ]
Около 97,5% воды на Земле солёная ; оставшиеся 2,5% — пресная вода . Большая часть пресной воды – около 69% – присутствует в виде льда в ледяных шапках и ледниках . [ 21 ] Средняя соленость океанов Земли составляет около 35 граммов (1,2 унции) соли на килограмм морской воды (3,5% соли). [ 22 ] Большая часть соли в океане образуется в результате выветривания и эрозии горных пород на суше. [ 23 ] Некоторые соли выделяются в результате вулканической деятельности или добываются из холодных магматических пород . [ 24 ]
Океаны также являются резервуаром растворенных атмосферных газов, которые необходимы для выживания многих водных форм жизни. [ 25 ] Морская вода оказывает важное влияние на мировой климат, а океаны служат большим резервуаром тепла . [ 26 ] Сдвиги в распределении температуры океана могут вызвать значительные изменения погоды, такие как Эль-Ниньо и Южное колебание . [ 27 ]

Всего океан занимает 71 процент мировой поверхности. [ 2 ] в среднем около 3,7 км (2,3 мили) в глубину. [ 28 ] По объему океан обеспечивает около 90 процентов жизненного пространства на планете. [ 2 ] Писатель-фантаст Артур Кларк отметил, что было бы более уместно называть планету Земля планетой Океан. [ 29 ] [ 30 ]
Однако вода встречается и в других частях Солнечной системы. Европа , одна из лун, вращающихся вокруг Юпитера , немного меньше земной Луны . Существует большая вероятность того, что под его ледяной поверхностью существует большой соленый океан. [ 31 ] Подсчитано, что внешняя кора твердого льда имеет толщину около 10–30 км (6–19 миль), а глубина жидкого океана под ней составляет около 100 км (60 миль). [ 32 ] Это сделало бы океан Европы в два раза больше земного океана. Высказывались предположения, что океан Европы может поддерживать жизнь . [ 33 ] [ 34 ] и может быть способен поддерживать многоклеточные микроорганизмы , если гидротермальные источники . на дне океана активны [ 35 ] Энцелад , небольшой ледяной спутник Сатурна, также имеет подземный океан , который активно выбрасывает теплую воду с поверхности Луны. [ 36 ]
Эволюция
[ редактировать ]−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||||||||||||||||||||
Историческое развитие
[ редактировать ]Земле . около 4,54 миллиарда лет [ 37 ] [ 38 ] [ 39 ] Самые ранние неоспоримые доказательства существования жизни на Земле датируются по меньшей мере 3,5 миллиарда лет назад. [ 40 ] [ 41 ] во время Эоархейской эры после того, как геологическая кора начала затвердевать после более раннего расплавленного Гадейского эона. микробного мата Окаменелости возрастом 3,48 миллиарда лет были обнаружены в песчанике в Западной Австралии . [ 42 ] [ 43 ] Другим ранним физическим свидетельством биогенного вещества является графит возрастом 3,7 миллиарда лет, в метаосадочных породах обнаруженных в Западной Гренландии. [ 44 ] а также «остатки биотической жизни », обнаруженные в скалах возрастом 4,1 миллиарда лет в Западной Австралии. [ 45 ] [ 46 ] По мнению одного из исследователей, «Если бы жизнь возникла на Земле сравнительно быстро… то она могла бы быть обычным явлением во Вселенной ». [ 45 ]
Все организмы на Земле произошли от общего предка или генофонда предков . [ 47 ] [ 48 ] Считается, что высокоэнергетическая химия создала самовоспроизводящуюся молекулу около 4 миллиардов лет назад, а полмиллиарда лет спустя существовал последний общий предок всей жизни . [ 49 ] В настоящее время научный консенсус заключается в том, что сложная биохимия, из которой состоит жизнь, возникла в результате более простых химических реакций. [ 50 ] Начало жизни могло включать в себя самовоспроизводящиеся молекулы, такие как РНК. [ 51 ] и сборка простых клеток. [ 52 ] В 2016 году ученые сообщили о наборе из 355 генов от последнего универсального общего предка (LUCA) всей жизни , включая микроорганизмы, живущие на Земле . [ 53 ]
Современные виды представляют собой этап процесса эволюции, а их разнообразие является продуктом длинной серии событий видообразования и вымирания. [ 54 ] Общее происхождение организмов было впервые выведено из четырех простых фактов об организмах: во-первых, они имеют географическое распространение, которое нельзя объяснить местной адаптацией. Во-вторых, разнообразие жизни – это не набор уникальных организмов, а организмы, имеющие морфологическое сходство . В-третьих, рудиментарные черты без четкой цели напоминают функциональные наследственные черты и, наконец, на основе этих сходств организмы можно классифицировать в иерархию вложенных групп, подобную генеалогическому древу. [ 55 ] Однако современные исследования показали, что из-за горизонтального переноса генов это «дерево жизни» может быть более сложным, чем простое ветвящееся дерево, поскольку некоторые гены распространились независимо между отдаленно родственными видами. [ 56 ] [ 57 ]
Предыдущие виды также оставили записи своей эволюционной истории. Окаменелости, наряду со сравнительной анатомией современных организмов, составляют морфологическую или анатомическую летопись. [ 58 ] Сравнивая анатомию современных и вымерших видов, палеонтологи могут сделать вывод о происхождении этих видов. Однако этот подход наиболее успешен для организмов, у которых были твердые части тела, такие как раковины, кости или зубы. Кроме того, поскольку прокариоты, такие как бактерии и археи, имеют ограниченный набор общих морфологий, их окаменелости не предоставляют информации об их происхождении.

Совсем недавно доказательства общего происхождения были получены в результате изучения биохимического сходства между организмами. Например, все живые клетки используют один и тот же базовый набор нуклеотидов и аминокислот . [ 60 ] Развитие молекулярной генетики выявило следы эволюции, оставшиеся в геномах организмов: датирование момента, когда виды разошлись по молекулярным часам , вызванным мутациями. [ 61 ] Например, сравнение последовательностей ДНК показало, что люди и шимпанзе имеют общие геномы на 98%, а анализ тех немногих областей, где они различаются, помогает пролить свет на то, когда существовал общий предок этих видов. [ 62 ]
Прокариоты населяли Землю примерно 3–4 миллиарда лет назад. [ 63 ] [ 64 ] Никаких очевидных изменений в морфологии или клеточной организации у этих организмов в течение следующих нескольких миллиардов лет не произошло. [ 65 ] Эукариотические клетки возникли между 1,6 и 2,7 миллиарда лет назад. Следующее серьезное изменение в клеточной структуре произошло, когда бактерии были поглощены эукариотическими клетками, образовав кооперативную ассоциацию, называемую эндосимбиозом . [ 66 ] [ 67 ] Поглощенные бактерии и клетка-хозяин затем подверглись совместной эволюции, при этом бактерии эволюционировали либо в митохондрии, либо в гидрогеносомы . [ 68 ] Очередное поглощение цианобактериоподобных организмов привело к образованию хлоропластов у водорослей и растений. [ 69 ]

История жизни была историей одноклеточных период в океанах начали появляться многоклеточные организмы эукариот, прокариотов и архей примерно до 610 миллионов лет назад, когда в эдиакарский . [ 63 ] [ 70 ] Эволюция многоклеточности происходила в ходе множества независимых событий в таких разнообразных организмах, как губки , бурые водоросли , цианобактерии , слизевики и миксобактерии . [ 71 ] В 2016 году ученые сообщили, что около 800 миллионов лет назад незначительное генетическое изменение в одной молекуле под названием GK-PID, возможно, позволило организмам перейти от одноклеточного организма к одной из многих клеток. [ 72 ]
Вскоре после появления этих первых многоклеточных организмов в течение примерно 10 миллионов лет появилось значительное биологическое разнообразие в результате события, названного Кембрийским взрывом . Здесь в палеонтологической летописи появилось большинство типов современных животных, а также уникальные линии, впоследствии вымершие. [ 73 ] Были предложены различные причины кембрийского взрыва, в том числе накопление кислорода в атмосфере в результате фотосинтеза. [ 74 ]
Около 500 миллионов лет назад растения и грибы начали колонизировать землю. Свидетельства появления первых наземных растений встречаются в ордовике , около 450 миллионов лет назад , в виде ископаемых спор. [ 75 ] Наземные растения начали диверсифицироваться в позднем силуре , примерно 430 миллионов лет назад . [ 76 ] За колонизацией суши растениями вскоре последовали членистоногие и другие животные. [ 77 ] Насекомые добились особенного успеха и даже сегодня составляют большинство видов животных. [ 78 ] Земноводные впервые появились около 364 миллионов лет назад, за ними последовали ранние амниоты и птицы около 155 миллионов лет назад (оба из « рептилийных » линий), млекопитающие около 129 миллионов лет назад, гоминины около 10 миллионов лет назад и современные люди около 250 000 лет назад. назад. [ 79 ] [ 80 ] [ 81 ] Однако, несмотря на эволюцию этих крупных животных, более мелкие организмы, подобные тем типам, которые развились на ранних стадиях этого процесса, продолжают быть весьма успешными и доминировать на Земле, причем большая часть как биомассы, так и видов является прокариотами. [ 82 ]
По оценкам, численность современных видов на Земле колеблется от 10 до 14 миллионов. [ 83 ] из них около 1,2 миллиона задокументированы, а более 86 процентов еще не описаны. [ 84 ]
Микроорганизмы
[ редактировать ]

Микроорганизмы составляют около 70% морской биомассы . [ 8 ] Микроорганизм слишком маленький , или микроб — это микроскопический организм, чтобы его можно было распознать невооруженным глазом. Это может быть одноклеточный [ 85 ] или многоклеточные . Микроорганизмы разнообразны и включают все бактерии и археи , большинство простейших, таких как водоросли , грибы и некоторых микроскопических животных, таких как коловратки .
Многие макроскопические животные и растения имеют микроскопические ювенильные стадии . Некоторые микробиологи также относят вирусы (и вироиды ) к микроорганизмам, но другие считают их неживыми. [ 86 ] [ 87 ]
Микроорганизмы играют решающую роль в переработке питательных веществ в экосистемах, поскольку они действуют как разлагатели . Некоторые микроорганизмы являются патогенными , вызывая заболевания и даже смерть растений и животных. [ 88 ] Будучи обитателями крупнейшей окружающей среды на Земле, морские микробные системы вызывают изменения во всех глобальных системах. Микробы ответственны практически за весь фотосинтез , происходящий в океане, а также за круговорот углерода , азота , фосфора , других питательных веществ и микроэлементов. [ 89 ]



Подводная микроскопическая жизнь разнообразна и до сих пор плохо изучена, например, роль вирусов в морских экосистемах. [ 90 ] Большинство морских вирусов представляют собой бактериофаги , которые безвредны для растений и животных, но необходимы для регулирования морских и пресноводных экосистем. [ 91 ] : 5 Они заражают и уничтожают бактерии в водных микробных сообществах и являются важнейшим механизмом переработки углерода в морской среде. Органические молекулы, высвобождаемые из мертвых бактериальных клеток, стимулируют рост новых бактерий и водорослей. [ 91 ] : 593 Вирусная активность также может способствовать биологическому насосу — процессу, посредством которого в глубинах углерод улавливается океана. [ 92 ]


Поток переносимых по воздуху микроорганизмов кружит над планетой над погодными системами, но под коммерческими воздушными путями. [ 93 ] Некоторые странствующие микроорганизмы переносятся земными пыльными бурями, но большинство происходит из морских микроорганизмов, содержащихся в морских брызгах . В 2018 году ученые сообщили, что сотни миллионов вирусов и десятки миллионов бактерий ежедневно оседают на каждом квадратном метре планеты. [ 94 ] [ 95 ]
Микроскопические организмы обитают по всей биосфере . Масса прокариотных микроорганизмов, в которую входят бактерии и археи, но не ядросодержащие эукариотные микроорганизмы , может достигать 0,8 триллиона тонн углерода (от общей массы биосферы , оцениваемой от 1 до 4 триллионов тонн). [ 96 ] Одноклеточные барофильные морские микробы были обнаружены на глубине 10 900 м (35 800 футов) в Марианской впадине , самом глубоком месте мирового океана. [ 97 ] [ 98 ] Микроорганизмы живут внутри скал на глубине 580 м (1900 футов) ниже морского дна, на глубине 2590 м (8500 футов) океана у побережья северо-запада США. [ 97 ] [ 99 ] а также на глубине 2400 м (7900 футов; 1,5 мили) под морским дном у берегов Японии. [ 100 ] Самая высокая известная температура, при которой может существовать микробная жизнь, составляет 122 ° C (252 ° F) ( Methanopyrus kandleri ). [ 101 ] В 2014 году учёные подтвердили существование микроорганизмов, живущих на глубине 800 м (2600 футов) подо льдом Антарктиды . [ 102 ] [ 103 ] По словам одного исследователя: «Микробы можно найти повсюду — они чрезвычайно адаптируются к условиям и выживают, где бы они ни находились». [ 97 ]
Морские вирусы
[ редактировать ]Вирусы — это небольшие инфекционные агенты , не имеющие собственного метаболизма и способные размножаться только внутри живых клеток других организмов . [ 104 ] Вирусы могут инфицировать все виды форм жизни , от животных и растений до микроорганизмов , включая бактерии и археи . [ 105 ] Линейный размер среднего вируса составляет примерно одну сотую размера средней бактерии . Большинство вирусов невозможно увидеть в оптический микроскоп , поэтому электронные микроскопы . вместо него используются [ 106 ]
Вирусы встречаются везде, где есть жизнь, и, вероятно, существовали с момента появления живых клеток. [ 107 ] Происхождение вирусов неясно, поскольку они не образуют окаменелостей, поэтому молекулярные методы использовались для сравнения ДНК или РНК вирусов и являются полезным средством изучения того, как они возникают. [ 108 ]
Сегодня вирусы признаны древними и имеют происхождение, предшествовавшее расколу жизни на три домена . [ 109 ] Но происхождение вирусов в эволюционной истории жизни неясно: некоторые, возможно, произошли от плазмид — фрагментов ДНК, которые могут перемещаться между клетками, — тогда как другие, возможно, произошли от бактерий. В эволюции вирусы являются важным средством горизонтального переноса генов , увеличивающего генетическое разнообразие . [ 110 ]



Мнения относительно того, являются ли вирусы формой жизни или органическими структурами, взаимодействующими с живыми организмами, расходятся. [ 111 ] Некоторые считают их формой жизни, поскольку они несут генетический материал, размножаются, создавая множество своих копий посредством самосборки, и развиваются посредством естественного отбора . Однако им не хватает ключевых характеристик, таких как клеточная структура, которая обычно считается необходимой для того, чтобы считаться жизнью. Поскольку вирусы обладают некоторыми, но не всеми такими качествами, их называют репликаторами. [ 111 ] и как «организмы на грани жизни». [ 112 ]

Бактериофаги , часто называемые просто фагами , представляют собой вирусы, паразитирующие на бактериях и археях. Морские фаги паразитируют на морских бактериях и археях, таких как цианобактерии . [ 113 ] Они представляют собой распространенную и разнообразную группу вирусов и являются наиболее распространенным биологическим объектом в морской среде, поскольку их хозяева, бактерии, обычно представляют собой численно доминирующую клеточную жизнь в море. Обычно в каждом мл морской воды содержится от 1 до 10 миллионов вирусов, или примерно в десять раз больше вирусов с двухцепочечной ДНК, чем клеточных организмов. [ 114 ] [ 115 ] хотя оценки численности вирусов в морской воде могут варьироваться в широком диапазоне. [ 116 ] [ 117 ] Хвостатые бактериофаги, по-видимому, доминируют в морских экосистемах по количеству и разнообразию организмов. [ 113 ] Бактериофаги, принадлежащие к семействам Corticoviridae , [ 118 ] Иновириды [ 119 ] и микровирусы [ 120 ] Также известно, что они заражают разнообразные морские бактерии.
Микроорганизмы составляют около 70% морской биомассы. [ 8 ] По оценкам, вирусы убивают 20% этой биомассы каждый день, а в океанах вирусов в 15 раз больше, чем бактерий и архей. Вирусы являются основными агентами, ответственными за быстрое уничтожение вредоносного цветения водорослей . [ 115 ] которые часто убивают других морских обитателей. [ 121 ] Число вирусов в океанах уменьшается по мере удаления от берега и глубже в воду, где меньше организмов-хозяев. [ 92 ]
Существуют также архейные вирусы , которые реплицируются внутри архей : это вирусы с двухцепочечной ДНК необычной, а иногда и уникальной формы. [ 122 ] [ 123 ] Наиболее подробно эти вирусы изучены на термофильных археях, в частности на порядках Sulfolobales и Thermoproteales . [ 124 ]
Вирусы являются важным естественным средством передачи генов между различными видами, что увеличивает генетическое разнообразие и стимулирует эволюцию. [ 110 ] Считается, что вирусы играли центральную роль в ранней эволюции, до диверсификации бактерий, архей и эукариот, во времена последнего универсального общего предка жизни на Земле. [ 125 ] Вирусы по-прежнему являются одним из крупнейших резервуаров неизведанного генетического разнообразия на Земле. [ 92 ]
Морские бактерии
[ редактировать ]

Бактерии обширную область прокариотических микроорганизмов составляют . Обычно бактерии имеют длину несколько микрометров и имеют разную форму: от сфер до палочек и спиралей. Бактерии были одними из первых форм жизни, появившихся на Земле , и присутствуют в большинстве ее сред обитания . Бактерии обитают в почве, воде, кислых горячих источниках , радиоактивных отходах . [ 126 ] и глубокие части земной коры . Бактерии также живут в симбиотических и паразитических отношениях с растениями и животными.
Когда-то бактерии считались растениями класса шизомицетов , а теперь их относят к прокариотам . В отличие от клеток животных и других эукариот , бактериальные клетки не содержат ядра и редко содержат мембраносвязанные органеллы . Хотя термин «бактерии» традиционно включал в себя всех прокариотов, научная классификация изменилась после открытия в 1990-х годах того, что прокариоты состоят из двух очень разных групп организмов, которые произошли от одного древнего общего предка. Эти эволюционные домены называются Бактерии и Археи . [ 127 ]
Предками современных бактерий были одноклеточные микроорганизмы, которые были первыми формами жизни , появившимися на Земле около 4 миллиардов лет назад. На протяжении примерно 3 миллиардов лет большинство организмов были микроскопическими, а доминирующими формами жизни были бактерии и археи. [ 65 ] [ 128 ] Хотя бактериальные окаменелости существуют, такие как строматолиты , отсутствие у них отличительной морфологии не позволяет использовать их для изучения истории эволюции бактерий или для определения времени происхождения определенного вида бактерий. Однако последовательности генов могут быть использованы для реконструкции бактериальной филогении , и эти исследования показывают, что бактерии сначала отделились от архейной/эукариотической линии. [ 129 ] Бактерии также участвовали во втором великом эволюционном расхождении – археях и эукариотах. Здесь эукариоты возникли в результате вступления древних бактерий в эндосимбиотические ассоциации с предками эукариотических клеток, которые сами, возможно, были родственны археям . [ 67 ] [ 66 ] Это включало поглощение протоэукариотическими клетками альфапротеобактериальных симбионтов с образованием либо митохондрий , либо гидрогеносом , которые до сих пор встречаются у всех известных эукариев. Позже некоторые эукариоты, уже содержащие митохондрии, также поглотили цианобактериоподобные организмы. Это привело к образованию хлоропластов у водорослей и растений. Есть также некоторые водоросли, возникшие в результате еще более поздних эндосимбиотических событий. Здесь эукариоты поглотили эукариотические водоросли, которые развились в пластиду «второго поколения». [ 130 ] [ 131 ] Это известно как вторичный эндосимбиоз .
-
 Морская Thiomargarita namibiensis , крупнейшая известная бактерия.
Морская Thiomargarita namibiensis , крупнейшая известная бактерия. -

-
 Хлоропласты эндосимбиотическом глаукофитов происхождении имеют слой пептидогликана что свидетельствует об их цианобактерий от , . [ 132 ]
Хлоропласты эндосимбиотическом глаукофитов происхождении имеют слой пептидогликана что свидетельствует об их цианобактерий от , . [ 132 ] -
 Бактерии могут быть полезными. Этот червь Помпеи , экстремофил, встречающийся только в гидротермальных источниках , имеет защитную оболочку из бактерий.
Бактерии могут быть полезными. Этот червь Помпеи , экстремофил, встречающийся только в гидротермальных источниках , имеет защитную оболочку из бактерий.




Самая крупная известная бактерия, морская Thiomargarita namibiensis , видна невооруженным глазом и иногда достигает 0,75 мм (750 мкм). [ 133 ] [ 134 ]
Морские археи
[ редактировать ]
Архея ( по-гречески древний [ 136 ] составляют домен и царство одноклеточных микроорганизмов ) . Эти микробы являются прокариотами , то есть в их клетках нет клеточного ядра или каких-либо других мембраносвязанных органелл .
Первоначально архей относили к бактериям , но эта классификация устарела. [ 137 ] Клетки архей обладают уникальными свойствами, отличающими их от двух других областей жизни: бактерий и эукариот . Археи далее делятся на несколько признанных типов . Классификация сложна, поскольку большинство из них не были изолированы в лаборатории и были обнаружены только путем анализа их нуклеиновых кислот в образцах из окружающей среды.
Археи и бактерии в целом схожи по размеру и форме, хотя некоторые археи имеют очень странную форму, например, плоские и квадратные клетки Haloquadratum walsbyi . [ 138 ] Несмотря на это морфологическое сходство с бактериями, археи обладают генами и несколькими метаболическими путями , которые более тесно связаны с таковыми у эукариот, в частности, ферментами, участвующими в транскрипции и трансляции . Другие аспекты биохимии архей уникальны, например, их зависимость от эфирных липидов в клеточных мембранах , таких как археолы . Археи используют больше источников энергии, чем эукариоты: они варьируются от органических соединений , таких как сахара, до аммиака , ионов металлов или даже газообразного водорода . Солеустойчивые археи ( Haloarchaea ) используют солнечный свет в качестве источника энергии, а другие виды архей фиксируют углерод ; однако, в отличие от растений и цианобактерий , ни один известный вид архей не обладает обоими свойствами. Археи размножаются бесполым путем бинарным делением , фрагментацией или почкованием ; в отличие от бактерий и эукариотов, ни один известный вид не образует спор .
Археи особенно многочисленны в океанах, а археи в планктоне могут быть одной из самых многочисленных групп организмов на планете. Археи являются основной частью жизни на Земле и могут играть роль как в углеродном, так и в азотном цикле .
-
 Галобактерии , обнаруженные в воде, близкой к насыщенной солью, теперь признаны архей.
Галобактерии , обнаруженные в воде, близкой к насыщенной солью, теперь признаны архей. -
 Плоские квадратные клетки архей Haloquadratum walsbyi.
Плоские квадратные клетки архей Haloquadratum walsbyi. -
 Methanosarcina barkeri — морская архея, производящая метан.
Methanosarcina barkeri — морская архея, производящая метан. -
 Термофилы , такие как Pyrolobus fumarii , выживают при температуре выше 100 °C.
Термофилы , такие как Pyrolobus fumarii , выживают при температуре выше 100 °C. -
 Рисунок другого морского термофила, Pyrococcus Furiosus.
Рисунок другого морского термофила, Pyrococcus Furiosus.





Морские протисты
[ редактировать ]Протисты — это эукариоты, которых нельзя отнести к растениям, грибам или животным. Обычно они одноклеточные и микроскопические. Жизнь зародилась как одноклеточные прокариоты ( бактерии и археи ), а позже развилась в более сложные эукариоты . Эукариоты — это более развитые формы жизни, известные как растения, животные, грибы и протисты. Термин «протист» исторически вошёл в употребление как удобный термин для обозначения эукариот, которых нельзя строго отнести к растениям, животным или грибам. Они не являются частью современной кладистики, поскольку являются парафилетическими (не имеют общего предка). Простейших можно разделить на четыре группы в зависимости от того, какое у них питание: растительное, животное, грибное или [ 139 ] или их смесь. [ 140 ]
Протисты по способу добывания пищи
| |||||||
|---|---|---|---|---|---|---|---|
| Тип простейшего | Описание | Пример | Другие примеры | ||||
| Растительный | Автотрофные протисты, которые производят себе пищу без необходимости потреблять другие организмы, обычно с помощью фотосинтеза. | 
|
Красные водоросли Cyanidium sp. | Зеленые водоросли , бурые водоросли , диатомовые водоросли и некоторые динофлагелляты . Растительноподобные протисты являются важными компонентами фитопланктона, обсуждаемыми ниже . | |||
| Животный | Гетеротрофные протисты, добывающие пищу, потребляя другие организмы. | 
|
Радиолярий- протист, рисунок Геккеля | Фораминиферы , а также некоторые морские амебы , инфузории и жгутиконосцы . | |||
| Грибоподобный | Сапротрофные протисты, получающие пищу из останков разложившихся и разложившихся организмов. | 
|
Сети морской слизи образуют лабиринтные сети трубок, по которым могут перемещаться амебы без ложноножек. | Морские лишайники | |||
| Миксотропы | Различный
|
Миксотрофные и осмотрофные протисты, которые получают пищу за счет комбинации вышеперечисленных веществ. | 
|
Эвглена мутабельная — фотосинтезирующий жгутиконосец. | Многие морские миксотропы встречаются среди простейших, в том числе среди инфузорий, ризарий и динофлагеллят. [ 141 ] | ||
Протисты — это очень разнообразные организмы, которые в настоящее время разделены на 18 типов, но их нелегко классифицировать. [ 143 ] [ 144 ] Исследования показали, что большое разнообразие протистов существует в океанах, глубоких морских жерлах и речных отложениях, что позволяет предположить, что большое количество эукариотических микробных сообществ еще не обнаружено. [ 145 ] [ 146 ] протистов мало исследовали Миксотрофных , но недавние исследования в морской среде показали, что миксотрофные протесты составляют значительную часть биомассы протистов . [ 141 ]
- Одноклеточные и микроскопические протисты
-
![Диатомовые водоросли являются основной группой водорослей, производящей около 20% мирового производства кислорода.[147]](//upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg) Диатомовые водоросли — основная группа водорослей, производящая около 20% мирового производства кислорода. [ 147 ]
Диатомовые водоросли — основная группа водорослей, производящая около 20% мирового производства кислорода. [ 147 ] -
![У диатомовых водорослей есть стеклянные клеточные стенки, состоящие из кремнезема и называемые панцирями.[148]](//upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg) У диатомовых водорослей стеклянные клеточные стенки состоят из кремнезема и называются панцирями . [ 148 ]
У диатомовых водорослей стеклянные клеточные стенки состоят из кремнезема и называются панцирями . [ 148 ] -
 Ископаемые панцири диатомей от 32 до 40 млн лет назад
Ископаемые панцири диатомей от 32 до 40 млн лет назад -

-
 Одноклеточная водоросль Gephyrocapsa Oceanica.
Одноклеточная водоросль Gephyrocapsa Oceanica. -
 Две динофлагелляты
Две динофлагелляты -
 Зооксантеллы — это фотосинтезирующие водоросли, живущие внутри хозяев, таких как кораллы .
Зооксантеллы — это фотосинтезирующие водоросли, живущие внутри хозяев, таких как кораллы . -

-

-
 Эта инфузория переваривает цианобактерии . Цитостом . или рот находится внизу справа
Эта инфузория переваривает цианобактерии . Цитостом . или рот находится внизу справа
![Диатомовые водоросли являются основной группой водорослей, производящей около 20% мирового производства кислорода.[147]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg)
![У диатомовых водорослей есть стеклянные клеточные стенки, состоящие из кремнезема и называемые панцирями.[148]](http://upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg)








В отличие от клеток прокариот клетки эукариот высокоорганизованы. Растения, животные и грибы обычно многоклеточные и обычно макроскопические . Большинство протистов одноклеточные и микроскопические. Но есть исключения. Некоторые одноклеточные морские простейшие макроскопичны. Некоторые морские слизевики имеют уникальный жизненный цикл, включающий переключение между одноклеточными, колониальными и многоклеточными формами. [ 149 ] Другие морские протисты не являются ни одноклеточными, ни микроскопическими, например морские водоросли .
- Макроскопические протисты (см. также одноклеточные макроводоросли → )
-
 Одноклеточная гигантская амеба имеет до 1000 ядер и достигает длины 5 мм (0,20 дюйма).
Одноклеточная гигантская амеба имеет до 1000 ядер и достигает длины 5 мм (0,20 дюйма). -
![Gromia sphaerica — крупная сферическая раковинная амеба, оставляющая грязевые следы. Его диаметр составляет до 3,8 см (1,5 дюйма).[150]](//upload.wikimedia.org/wikipedia/commons/thumb/9/99/Gromia_in_situ_closeup.png/266px-Gromia_in_situ_closeup.png) Gromia sphaerica — крупная сферическая раковинная амеба , оставляющая грязевые следы. Его диаметр составляет до 3,8 см (1,5 дюйма). [ 150 ]
Gromia sphaerica — крупная сферическая раковинная амеба , оставляющая грязевые следы. Его диаметр составляет до 3,8 см (1,5 дюйма). [ 150 ] -
 Spiculosiphon Oceana — одноклеточный фораминифер , внешним видом и образом жизни имитирующий губку , вырастает до 5 см в длину.
Spiculosiphon Oceana — одноклеточный фораминифер , внешним видом и образом жизни имитирующий губку , вырастает до 5 см в длину. -
 Ксенофиофор абиссальных , еще один одноклеточный фораминифер, обитает в зонах . У него гигантская раковина диаметром до 20 см (7,9 дюйма). [ 151 ]
Ксенофиофор абиссальных , еще один одноклеточный фораминифер, обитает в зонах . У него гигантская раковина диаметром до 20 см (7,9 дюйма). [ 151 ] -
 Гигантская ламинария , бурая водоросль , не является настоящим растением, однако она многоклеточная и может достигать 50 м в высоту.
Гигантская ламинария , бурая водоросль , не является настоящим растением, однако она многоклеточная и может достигать 50 м в высоту.

![Gromia sphaerica — крупная сферическая раковинная амеба, оставляющая грязевые следы. Его диаметр составляет до 3,8 см (1,5 дюйма).[150]](http://upload.wikimedia.org/wikipedia/commons/thumb/9/99/Gromia_in_situ_closeup.png/266px-Gromia_in_situ_closeup.png)



Протистов описывают как таксономическую сумку, куда все, что не вписывается ни в одно из основных биологических царств . можно поместить [ 152 ] Некоторые современные авторы предпочитают исключать многоклеточные организмы из традиционного определения простейшего, ограничивая простейших одноклеточными организмами. [ 153 ] [ 154 ] Это более ограниченное определение исключает морские водоросли и слизевики . [ 155 ]
Морские микроживотные
[ редактировать ]| Внешние видео | |
|---|---|
В молодом возрасте животные развиваются на микроскопических стадиях, которые могут включать споры , яйца и личинки . По крайней мере, одна микроскопическая группа животных, паразитические книдарии Myxozoa , во взрослой форме являются одноклеточными и включают морские виды. Другие взрослые морские микроживотные многоклеточные. Микроскопические взрослые членистоногие чаще встречаются внутри страны, в пресной воде, но есть и морские виды. К микроскопическим взрослым морским ракообразным относятся некоторые копеподы , ветвистоусые и тихоходки (водяные медведи). Некоторые морские нематоды и коловратки слишком малы, чтобы их можно было распознать невооруженным глазом, как и многие лорициферы , включая недавно обнаруженные анаэробные виды, которые проводят свою жизнь в бескислородной среде. [ 156 ] [ 157 ] Веслоногие раконогие вносят больший вклад во вторичную продуктивность и поглощение углерода Мирового океана, чем любая другая группа организмов. [ 158 ] [ 159 ] Хотя клещей обычно не считают морскими организмами, большинство видов семейства Halacaridae обитают в море. [ 160 ]
- Морские микроживотные
-
 Более 10 000 морских видов относятся к веслоногим ракообразным , маленьким, часто микроскопическим ракообразным.
Более 10 000 морских видов относятся к веслоногим ракообразным , маленьким, часто микроскопическим ракообразным. -
 Фотография в темном поле гастротриха . , червеобразного животного, живущего между частицами отложений
Фотография в темном поле гастротриха . , червеобразного животного, живущего между частицами отложений -
 Панцирные Pliciloricus enigmaticus длиной около 0,2 мм обитают в пространствах между морским гравием.
Панцирные Pliciloricus enigmaticus длиной около 0,2 мм обитают в пространствах между морским гравием. -
 Рисунок тихоходки (водяного медведя) на песчинке
Рисунок тихоходки (водяного медведя) на песчинке -
 Коловушки , обычно длиной 0,1–0,5 мм, могут выглядеть как протисты, но имеют много клеток и принадлежат к отряду Animalia.
Коловушки , обычно длиной 0,1–0,5 мм, могут выглядеть как протисты, но имеют много клеток и принадлежат к отряду Animalia.





Грибы
[ редактировать ]

более 1500 видов грибов . Из морской среды известно [ 161 ] Они паразитируют на морских водорослях или животных или представляют собой сапробы, питающиеся мертвыми органическими веществами водорослей, кораллов, цист простейших, морских трав, древесины и других субстратов. [ 162 ] Споры многих видов имеют специальные придатки, облегчающие прикрепление к субстрату. [ 163 ] Морские грибы также можно найти в морской пене и вокруг гидротермальных зон океана. [ 164 ] разнообразный спектр необычных вторичных метаболитов . Морские грибы производят [ 165 ]
Микопланктон — сапротропный член планктонных сообществ морских и пресноводных экосистем . [ 166 ] [ 167 ] Они состоят из нитчатых свободноживущих грибов и дрожжей, связанных с планктонными частицами или фитопланктоном . [ 168 ] Подобно бактериопланктону , эти водные грибы играют значительную роль в гетеротрофной минерализации и круговороте питательных веществ . [ 169 ] Микопланктон может достигать 20 мм в диаметре и более 50 мм в длину. [ 170 ]
Типичный миллилитр морской воды содержит около 10 3 до 10 4 грибковые клетки. [ 171 ] Это число больше в прибрежных экосистемах и эстуариях из-за стока питательных веществ от наземных сообществ. Более высокое разнообразие микопланктона встречается вокруг побережий и в поверхностных водах на глубине до 1000 метров, вертикальный профиль которого зависит от обилия фитопланктона . [ 172 ] [ 173 ] Этот профиль меняется в зависимости от сезона из-за изменений в доступности питательных веществ. [ 174 ] Морские грибы выживают в среде с постоянным дефицитом кислорода и, следовательно, зависят от диффузии кислорода за счет турбулентности и кислорода, вырабатываемого фотосинтезирующими организмами . [ 175 ]
Морские грибы можно разделить на: [ 175 ]
- Низшие грибы – приспособленные к морским местообитаниям ( зооспоровые грибы, в том числе мастигомицеты: оомицеты и хитридиомицеты )
- Высшие грибы — нитчатые, модифицированные к планктонному образу жизни ( гифомицеты , аскомицеты , базидиомицеты ). Большинство видов микопланктона относятся к высшим грибам. [ 172 ]
Лишайники — это мутуалистические ассоциации между грибом, обычно аскомицетом , и водорослью или цианобактерией . Некоторые лишайники встречаются в морской среде. [ 176 ] Еще больше их встречается в зоне всплеска , где они занимают разные вертикальные зоны в зависимости от того, насколько они устойчивы к погружению. [ 177 ] Некоторые лишайники живут долго; возраст одного вида составляет 8600 лет. [ 178 ] Однако продолжительность их жизни трудно измерить, поскольку определение одного и того же лишайника не является точным. [ 179 ] Лишайники растут путем вегетативного отрыва части, которая может быть определена, а может и не быть определена как один и тот же лишайник, и два лишайника разного возраста могут сливаться, что поднимает вопрос, является ли это одним и тем же лишайником. [ 179 ] Морская улитка Littoraria irrorata повреждает растения Спартины в морских болотах, где она обитает, что позволяет спорам литоральных аскомицетовых грибов колонизировать растение. Затем улитка поедает грибковые наросты, а не саму траву. [ 180 ]
Согласно летописям окаменелостей, грибы относятся к позднему протерозою 900-570 миллионов лет назад. Ископаемые морские лишайники возрастом 600 миллионов лет были обнаружены в Китае. [ 181 ] Была выдвинута гипотеза, что микопланктон произошел от наземных грибов, вероятно, в палеозойскую эру (390 миллионов лет назад). [ 182 ]
Происхождение животных
[ редактировать ]
Самыми ранними животными были морские беспозвоночные , то есть позвоночные появились позже. Животные – многоклеточные эукариоты , [ примечание 2 ] и отличаются от растений, водорослей и грибов отсутствием клеточных стенок . [ 183 ] Морские беспозвоночные — это животные, населяющие морскую среду, помимо позвоночных представителей типа хордовых ; у беспозвоночных отсутствует позвоночный столб . Некоторые из них развили панцирь или твердый экзоскелет .
Самые ранние окаменелости животных могут принадлежать к роду Dickinsonia . [ 184 ] 571–541 миллион лет назад. [ 185 ] Отдельные дикинсонии обычно напоминают двусторонне-симметричный ребристый овал. Они продолжали расти, пока не покрылись осадком или не погибли иным образом. [ 186 ] и провели большую часть своей жизни, твердо привязав свои тела к осадку. [ 187 ] Их таксономическое сходство в настоящее время неизвестно, но способ их роста соответствует билатеральному сходству. [ 188 ]
Помимо дикинсонии , самыми ранними широко признанными окаменелостями животных являются довольно современные на вид книдарии (группа, в которую входят кораллы , медузы , морские анемоны и гидры ), возможно, датируемые примерно 580 млн лет назад. [ 189 ] , Биота Эдиакары процветавшая последние 40 миллионов лет до начала кембрия , [ 190 ] были первыми животными длиной более нескольких сантиметров. Подобно Дикинсонии , многие из них были плоскими, со «стеганным» внешним видом, и казались настолько странными, что появилось предложение отнести их к отдельному царству , Вендозоа . [ 191 ] Другие, однако, были интерпретированы как ранние моллюски ( Kimberella [ 192 ] [ 193 ] ), иглокожие ( Arkarua [ 194 ] ), и членистоногих ( Сприггина , [ 195 ] Парванкорин [ 196 ] ). До сих пор ведутся споры о классификации этих образцов, главным образом потому, что диагностические особенности, которые позволяют систематикам классифицировать более поздние организмы, такие как сходство с живыми организмами, обычно отсутствуют у эдиакарцев. Однако мало кто сомневается в том, что кимберелла была, по крайней мере, трехблобластным двусторонним животным, другими словами, животным, значительно более сложным, чем книдарии. [ 197 ]
Небольшая ракушечная фауна представляет собой очень смешанную коллекцию окаменелостей, обнаруженных между позднем эдиакарским и средним кембрийским периодами. Самый ранний, Cloudina , демонстрирует признаки успешной защиты от хищников и может указывать на начало эволюционной гонки вооружений . Некоторые крошечные раннекембрийские раковины почти наверняка принадлежали моллюскам, а владельцы некоторых «броневых пластин» — Halkieria и Microdictyon были найдены более полные экземпляры — в конечном итоге были идентифицированы, когда в кембрийских лагерштеттенах , сохранившие мягкотелых животных. [ 198 ]
Планы тела и типы
[ редактировать ]
Беспозвоночные сгруппированы в разные типы . Неформально типы можно рассматривать как способ группировки организмов в соответствии со строением их тела . [ 199 ] [ 200 ] : 33 План тела относится к проекту, который описывает форму или морфологию организма, такую как его симметрия , сегментация и расположение его придатков . Идея строения тела возникла у позвоночных , которые были сгруппированы в один тип. Но строение тела позвоночных — лишь одно из многих, а беспозвоночные состоят из многих типов или строений тела. Историю открытия строения тела можно рассматривать как движение от мировоззрения, сосредоточенного на позвоночных, к рассмотрению позвоночных как одного из многих строений тела. первопроходцев Среди зоологов- Линней выделил два строения тела за пределами позвоночных; Кювье назвал троих; у Геккеля было четыре, а у Протиста еще восемь, всего двенадцать. Для сравнения, число типов, признанных современными зоологами, возросло до 35 . [ 200 ]

Исторически считалось, что структуры тела быстро изменились во время кембрийского взрыва . [ 204 ] но более детальное понимание эволюции животных предполагает постепенное развитие строения тела на протяжении раннего палеозоя и далее. [ 205 ] В более общем смысле тип можно определить двумя способами: как описано выше, как группу организмов с определенной степенью морфологического сходства или сходства в развитии ( фенетическое определение) или как группу организмов с определенной степенью эволюционного родства ( филогенетическое определение). определение). [ 205 ]
В 1970-х годах уже велись споры о том, было ли появление современных типов «взрывным» или постепенным, но скрытым нехваткой окаменелостей докембрийских животных. [ 198 ] Повторный анализ окаменелостей из лагерштетте в сланцах Берджесс повысил интерес к этому вопросу, когда выявил животных, таких как Opabinia , которые не вписывались ни в один известный тип . В то время это было интерпретировано как свидетельство того, что современные типы очень быстро развились в результате кембрийского взрыва и что «странные чудеса» сланцев Бёрджесс показали, что ранний кембрий был уникальным экспериментальным периодом эволюции животных. [ 206 ] Более поздние открытия подобных животных и развитие новых теоретических подходов привели к выводу, что многие из «странных чудес» были эволюционными «тетками» или «кузенами» современных групп. [ 207 ] - например, что Опабиния была членом лобоподов , группы, в которую входят предки членистоногих, и что она, возможно, была тесно связана с современными тихоходками . [ 208 ] Тем не менее, до сих пор ведется много споров о том, действительно ли кембрийский взрыв был взрывным, и если да, то как и почему он произошел и почему он кажется уникальным в истории животных. [ 209 ]
Самые ранние животные
[ редактировать ]Животные с самым глубоким ветвлением — самые ранние животные, появившиеся в ходе эволюции, — это морские беспозвоночные организмы. Самыми ранними типами животных являются Porifera , Ctenophora , Placozoa и Cnidaria . Ни один из представителей этих клад не имеет строения тела с двусторонней симметрией .
| ||||||||||||||||||||||||||||||||||


Морские губки
[ редактировать ]
Губки — животные типа Porifera ( от современного латинского языка — несущие поры) . [ 213 ] ). Это многоклеточные организмы, тела которых полны пор и каналов, позволяющих воде циркулировать через них, и состоят из желеобразного мезоила, зажатого между двумя тонкими слоями клеток . У них есть неспециализированные клетки, которые могут трансформироваться в другие типы и которые при этом часто мигрируют между основными клеточными слоями и мезохилом. У губок нет нервной , пищеварительной и кровеносной систем . Вместо этого большинство полагаются на поддержание постоянного потока воды через свое тело для получения пищи и кислорода, а также для удаления отходов.
Губки похожи на других животных тем, что они многоклеточные , гетеротрофные , лишены клеточных стенок и производят сперматозоиды . В отличие от других животных, у них отсутствуют настоящие ткани и органы , а также отсутствует симметрия тела . Формы их тел приспособлены для максимальной эффективности потока воды через центральную полость, где она откладывает питательные вещества, и выходит через отверстие, называемое оскулумом . Многие губки имеют внутренний скелет из спонгина и/или спикулы из карбоната кальция или диоксида кремния . Все губки — сидячие водные животные. Хотя существуют пресноводные виды, подавляющее большинство из них являются морскими (солеными) видами, варьирующимися от приливных зон до глубин, превышающих 8800 м (5,5 миль). Некоторые губки живут очень долго; Есть свидетельства того, что глубоководная стеклянная губка Monorhaphis chuni жила около 11 000 лет. [ 214 ] [ 215 ]
В то время как большинство из примерно 5 000–10 000 известных видов питаются бактериями и другими пищевыми частицами в воде, некоторые из них являются хозяевами фотосинтезирующих микроорганизмов в качестве эндосимбионтов , и эти союзы часто производят больше пищи и кислорода, чем потребляют. Некоторые виды губок, обитающие в условиях скудной пищи, превратились в хищников , питающихся в основном мелкими ракообразными . [ 216 ]
-
 Биоразнообразие губок. На этой фотографии четыре вида губок.
Биоразнообразие губок. На этой фотографии четыре вида губок. -

-
 Цветочная корзина Венеры на глубине 2572 метра
Цветочная корзина Венеры на глубине 2572 метра -

-
 Долгоживущий Monorhaphis chuni.
Долгоживущий Monorhaphis chuni.





Линней ошибочно отнес губки к растениям отряда Водоросли . [ 217 ] Долгое время после этого губки были отнесены к отдельному подцарству Паразоа (то есть рядом с животными ). [ 218 ] Сейчас их классифицируют как парафилетический тип , из которого произошли высшие животные. [ 219 ]
Гребневики
[ редактировать ]Гребенчатые (от греческого « носитель гребня »), широко известные как гребневики, представляют собой тип, обитающий по всему миру в морских водах. Это самые крупные неколониальные животные, плавающие с помощью ресничек (волосков или гребней). [ 220 ] Прибрежные виды должны быть достаточно прочными, чтобы противостоять волнам и бурлящим осадкам, но некоторые океанические виды настолько хрупкие и прозрачные, что их очень трудно поймать в целости и сохранности для изучения. [ 221 ] Раньше считалось, что гребневики присутствуют в океане лишь в незначительной степени, но теперь известно, что они часто составляют значительную и даже доминирующую часть планктонной биомассы. [ 222 ] : 269
Тип насчитывает около 150 известных видов с широким диапазоном форм тела. Размеры варьируются от нескольких миллиметров до 1,5 м (4 фута 11 дюймов). Цидиппиды имеют яйцевидную форму, их реснички расположены в восемь радиальных гребенчатых рядов, а для захвата добычи они используют втягивающиеся щупальца. Бентические платиктениды обычно безгребешковые и плоские. У прибрежных бероидов зияющая пасть и отсутствие щупалец. Большинство взрослых гребневиков охотятся на микроскопических личинок, коловраток и мелких ракообразных, но бероиды охотятся на других гребневиков.
-
 Свет преломляется вдоль рядов гребней цидиппида, левое щупальце развернуто, правое втянуто.
Свет преломляется вдоль рядов гребней цидиппида, левое щупальце развернуто, правое втянуто. -
 Висячие щупальца глубоководных гребневиков усеяны тентиллами ( субщупальцами).
Висячие щупальца глубоководных гребневиков усеяны тентиллами ( субщупальцами). -
 яйцевидной формы Цидиппидный гребневик
яйцевидной формы Цидиппидный гребневик -
 Группа маленьких донных ползающих гребневиков, пускающих щупальца и симбиотически живущих на морской звезде.
Группа маленьких донных ползающих гребневиков, пускающих щупальца и симбиотически живущих на морской звезде. -
 Лобата сп. с парными толстыми долями
Лобата сп. с парными толстыми долями -
![У морского ореха есть временный анус, который образуется только тогда, когда ему необходимо испражняться.[223]](//upload.wikimedia.org/wikipedia/commons/thumb/f/f8/Mnemiopsis_leidyi_2.jpg/255px-Mnemiopsis_leidyi_2.jpg)





![У морского ореха есть временный анус, который образуется только тогда, когда ему необходимо испражняться.[223]](http://upload.wikimedia.org/wikipedia/commons/thumb/f/f8/Mnemiopsis_leidyi_2.jpg/255px-Mnemiopsis_leidyi_2.jpg)
Ранние писатели объединяли гребневиков с книдариями . Гребневики напоминают книдарий тем, что полагаются на поток воды через полость тела как для пищеварения, так и для дыхания, а также имеют децентрализованную нервную сеть, а не мозг. Также, как и книдарии, тела гребневиков состоят из массы желе, с одним слоем клеток снаружи и другим, выстилающим внутреннюю полость. Однако у гребневиков эти слои имеют глубину в две клетки, а у книдарий - только в одну клетку. В то время как книдарии обладают радиальной симметрией , гребневики имеют два анальных канала, которые обладают бирадиальной симметрией (полуоборотная вращательная симметрия). [ 224 ] Положение гребневиков в эволюционном генеалогическом древе животных уже давно обсуждается, и в настоящее время большинство взглядов, основанных на молекулярной филогенетике , заключается в том, что книдарии и билатерии более тесно связаны друг с другом, чем любой из них с гребневиками. [ 222 ] : 222
| Внешние видео | |
|---|---|
Плакозоа
[ редактировать ]Плакозоа (от греческого « плоские животные ») имеют самое простое строение среди всех животных. Они представляют собой базальную форму свободноживущего (непаразитического) многоклеточного организма. [ 225 ] которые еще не имеют общего названия. [ 226 ] Они обитают в морской среде и образуют тип, содержащий пока только три описанных вида, из которых первый, классический Trichoplax adhaerens , был открыт в 1883 году. [ 227 ] С 2017 года были открыты еще два вида. [ 228 ] [ 229 ] а генетические методы показывают, что этот тип включает еще от 100 до 200 неописанных видов . [ 230 ]

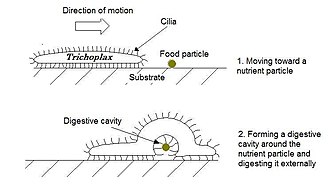
Трихоплакс — небольшое уплощенное животное диаметром около одного мм и обычно толщиной около 25 мкм. Подобно амебам, на которые они внешне похожи, они постоянно меняют свою внешнюю форму. Кроме того, иногда образуются сферические фазы, которые могут облегчить движение. У трихоплакса отсутствуют ткани и органы. Явной симметрии тела нет, поэтому невозможно отличить переднюю часть от задней или левую от правой. Он состоит из нескольких тысяч клеток шести типов в трех отдельных слоях. [ 231 ] Наружный слой простых эпителиальных клеток несет реснички , которые животное использует, чтобы ползать по морскому дну. [ 232 ] Трихоплаксы питаются, заглатывая и поглощая частицы пищи – преимущественно микробы и органический детрит – своей нижней стороной.
Морские книдарии
[ редактировать ]
Книдарии (от греческого крапива ) отличаются наличием стрекательных клеток — специализированных клеток, которые они используют в основном для захвата добычи. Книдарии включают кораллы , морские анемоны , медузы и гидрозои . Они образуют тип, содержащий более 10 000 [ 233 ] виды животных , обитающие исключительно в водной (преимущественно морской) среде. Их тела состоят из мезоглеи , неживого желеобразного вещества, зажатого между двумя слоями эпителия , толщиной в основном в одну клетку . У них есть две основные формы тела: плавающие медузы и сидячие полипы , оба из которых радиально симметричны, а рты окружены щупальцами, несущими книдоциты. Обе формы имеют одно отверстие и полость тела, которые используются для пищеварения и дыхания .
Ископаемые книдарии были обнаружены в горных породах, образовавшихся около 580 миллионов лет назад . Окаменелости книдарий, не образующих минерализованных структур, редки. В настоящее время ученые считают, что книдарии, гребневики и билатерии более тесно связаны с известковыми губками, чем с другими губками , и что антозойи являются эволюционными «тетями» или «сестрами» других книдарий и наиболее тесно связаны с билатериями.
Книдарии — простейшие животные, у которых клетки организованы в ткани. [ 234 ] Звездочка актинии используется в качестве модельного организма в исследованиях. [ 235 ] За ним легко ухаживать в лаборатории, и был разработан протокол , позволяющий ежедневно получать большое количество эмбрионов. [ 236 ] Существует поразительная степень сходства в сохранении и сложности последовательности генов между морскими анемонами и позвоночными. [ 236 ] В частности, у анемона присутствуют гены, участвующие в формировании головы у позвоночных. [ 237 ] [ 238 ]
-
 Морские анемоны распространены в приливных бассейнах .
Морские анемоны распространены в приливных бассейнах . -
 Их щупальца жалят и парализуют мелкую рыбу.
Их щупальца жалят и парализуют мелкую рыбу. -

-
 Если остров опускается под воду, рост кораллов может поспевать за поднимающейся водой и образовать атолл .
Если остров опускается под воду, рост кораллов может поспевать за поднимающейся водой и образовать атолл . -
![Мантия медузы-красного бумажного фонаря сминается и расширяется, как бумажный фонарь.[239]](//upload.wikimedia.org/wikipedia/commons/thumb/e/e5/Red-paper-lantern-jellyfish-Karen-Osborn-Smithsonian-Institution.png/321px-Red-paper-lantern-jellyfish-Karen-Osborn-Smithsonian-Institution.png) Мантия медузы красного бумажного фонаря сминается и расширяется, как бумажный фонарь. [ 239 ]
Мантия медузы красного бумажного фонаря сминается и расширяется, как бумажный фонарь. [ 239 ]




![Мантия медузы-красного бумажного фонаря сминается и расширяется, как бумажный фонарь.[239]](http://upload.wikimedia.org/wikipedia/commons/thumb/e/e5/Red-paper-lantern-jellyfish-Karen-Osborn-Smithsonian-Institution.png/321px-Red-paper-lantern-jellyfish-Karen-Osborn-Smithsonian-Institution.png)
-

-
 Marrusортоканна — еще один колониальный сифонофор, собранный из двух типов зооидов .
Marrusортоканна — еще один колониальный сифонофор, собранный из двух типов зооидов . -
![Porpita porpita состоит из колонии гидроидов[240].](//upload.wikimedia.org/wikipedia/commons/thumb/2/23/Porpita_porpita.jpg/252px-Porpita_porpita.jpg)
-

-
![Turritopsis dohrnii достигает биологического бессмертия, перенося свои клетки обратно в детство.[242][243]](//upload.wikimedia.org/wikipedia/commons/b/b0/Turritopsis_dohrnii_%28cropped%29.jpg) Turritopsis dohrnii достигает биологического бессмертия , перенося свои клетки обратно в детство. [ 242 ] [ 243 ]
Turritopsis dohrnii достигает биологического бессмертия , перенося свои клетки обратно в детство. [ 242 ] [ 243 ] -
![Морская оса — самая смертоносная медуза в мире.[244]](//upload.wikimedia.org/wikipedia/commons/thumb/7/75/Chironex_fleckeri_%28sea_wasp%29.jpg/255px-Chironex_fleckeri_%28sea_wasp%29.jpg)


![Porpita porpita состоит из колонии гидроидов[240].](http://upload.wikimedia.org/wikipedia/commons/thumb/2/23/Porpita_porpita.jpg/252px-Porpita_porpita.jpg)

![Turritopsis dohrnii достигает биологического бессмертия, перенося свои клетки обратно в детство.[242][243]](http://upload.wikimedia.org/wikipedia/commons/b/b0/Turritopsis_dohrnii_%28cropped%29.jpg)
![Морская оса — самая смертоносная медуза в мире.[244]](http://upload.wikimedia.org/wikipedia/commons/thumb/7/75/Chironex_fleckeri_%28sea_wasp%29.jpg/255px-Chironex_fleckeri_%28sea_wasp%29.jpg)
Двусторонние беспозвоночные животные
[ редактировать ]
Некоторые из самых ранних билатерий были червеобразными, а первоначальные билатерии, возможно, были донными червями с одним отверстием в теле. [ 245 ] Двустороннее тело можно представить как цилиндр с кишкой, проходящей между двумя отверстиями: ртом и анусом. Вокруг кишки имеется внутренняя полость тела — целом или псевдоцелом. [ а ] Животные с таким двусторонне-симметричным строением тела имеют головной (передний) конец и хвостовой (задний) конец, а также спину (дорсальный) и брюшной (вентральный); следовательно, у них также есть левая и правая стороны. [ 246 ] [ 247 ]
Наличие передней части означает, что эта часть тела подвергается воздействию раздражителей, таких как еда, что способствует цефализации , развитию головы с органами чувств и рта. [ 248 ] Тело вытягивается назад от головы, и у многих билатерий есть комбинация круговых мышц , сжимающих тело, делая его длиннее, и противоположный набор продольных мышц, укорачивающих тело; [ 247 ] они позволяют мягкотелым животным с гидростатическим скелетом двигаться за счет перистальтики . [ 249 ] У них также есть кишечник, который проходит через цилиндрическое тело от рта до ануса. Многие билатерийные типы имеют первичные личинки , которые плавают с ресничками и имеют апикальный орган, содержащий сенсорные клетки. Однако из каждой из этих характеристик есть исключения; например, взрослые иглокожие радиально симметричны (в отличие от их личинок), а некоторые паразитические черви имеют чрезвычайно упрощенное строение тела. [ 246 ] [ 247 ]

| ← билатерии |
| |||||||||||||||||||||
Протостомы
[ редактировать ]Перворотые (от греческого « первый рот ) — супертип животных » . Это сестринская клада вторичноротых ( от греческого « второй рот »), вместе с которой она образует кладу Nephrozoa . Протостомы отличаются от вторичноротых по способу развития зародышей . У протостомов первым развивающимся отверстием становится рот , а у вторичноротых — анальное отверстие. [ 251 ] [ 252 ]
| ← Перворотые |
| ||||||||||||||||||
| (сохранившийся) |


Морские черви
[ редактировать ]
Черви (древнеанглийское слово « змеи ») образуют ряд типов. Различные группы морских червей связаны лишь отдаленно, поэтому они встречаются в нескольких разных типах, таких как Annelida (сегментированные черви), Chaetognatha (черви-стрелы), Phoronida (подковообразные черви) и Hemichordata . Все черви, кроме Hemichordata, являются протостомами. Полухордовые - это вторичноротые и обсуждаются в отдельном разделе ниже.
Типичное строение тела червя включает длинные цилиндрические трубчатые тела и отсутствие конечностей . Морские черви различаются по размеру от микроскопических до более 1 метра (3,3 фута) в длину для некоторых морских многощетинковых червей ( щетинковых червей ). [ 253 ] и до 58 метров (190 футов) для морского немертинского червя ( шнурочного червя ). [ 254 ] Некоторые морские черви занимают небольшое количество паразитических ниш, живя внутри тел других животных, в то время как другие живут более свободно в морской среде или роются под землей. Многие из этих червей имеют специальные щупальца, используемые для обмена кислорода и углекислого газа, а также могут использоваться для размножения. Некоторые морские черви являются трубчатыми , например, гигантский трубчатый червь , который живет в водах вблизи подводных вулканов и может выдерживать температуру до 90 градусов по Цельсию . Платихельминты (плоские черви) образуют еще один тип червей, включающий класс паразитических ленточных червей. Морской ленточный червь Polygonoporus giganticus , обитающий в кишечнике кашалотов , может вырасти до более чем 30 м (100 футов). [ 255 ] [ 256 ]
Нематоды (круглые черви) представляют собой еще один тип червей с трубчатой пищеварительной системой и отверстиями на обоих концах. [ 257 ] [ 258 ] Описано более 25 000 видов нематод. [ 259 ] [ 260 ] из них более половины являются паразитическими. По оценкам, еще миллион остается неописанным. [ 261 ] Они повсеместно распространены в морской, пресноводной и наземной среде, где часто превосходят численностью других животных как по численности, так и по численности видов. Земли Они встречаются во всех частях литосферы , от вершин гор до дна океанических впадин . [ 262 ] По подсчетам они составляют 90% всех животных на дне океана . [ 263 ] Их численное преобладание, часто превышающее миллион особей на квадратный метр и составляющее около 80% всех отдельных животных на Земле, разнообразие жизненных циклов и присутствие на различных трофических уровнях указывают на важную роль во многих экосистемах. [ 264 ]
-
 Гигантские трубчатые черви собираются вокруг гидротермальных источников .
Гигантские трубчатые черви собираются вокруг гидротермальных источников . -
 Нематоды — это вездесущие псевдоцеломаты , которые могут паразитировать на морских растениях и животных.
Нематоды — это вездесущие псевдоцеломаты , которые могут паразитировать на морских растениях и животных. -
 Мотыль обычно обитает на дне мелких морских вод.
Мотыль обычно обитает на дне мелких морских вод.



Морские моллюски
[ редактировать ]


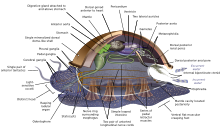
Моллюски (лат. « мягкие ») образуют тип , насчитывающий около 85 000 современных признанных видов . [ 267 ] Это самый крупный морской тип по количеству видов, содержащий около 23% всех названных морских организмов . [ 268 ] Моллюски имеют более разнообразные формы, чем другие типы беспозвоночных. Они весьма разнообразны не только по размеру и анатомическому строению, но и по поведению и среде обитания.
Тип моллюсков делится на 9—10 таксономических классов . К этим классам относятся брюхоногие моллюски , двустворчатые моллюски и головоногие моллюски , а также другие менее известные, но самобытные классы. Брюхоногих моллюсков с защитным панцирем называют улитками , а брюхоногих моллюсков без защитного панциря — слизняками . Брюхоногие моллюски на сегодняшний день являются самыми многочисленными моллюсками с точки зрения видов. [ 269 ] К двустворчатым моллюскам относятся моллюски , устрицы , моллюски , мидии , морские гребешки и многие другие семейства . Насчитывается около 8000 видов морских двустворчатых моллюсков (включая солоноватоводные и устьевые виды). глубоководный моллюск Сообщается, что куахог прожил 507 лет. [ 270 ] что делает его самой продолжительной зарегистрированной жизнью среди всех животных, за исключением колониальных животных или почти колониальных животных, таких как губки . [ 214 ]
- Брюхоногие моллюски и двустворчатые моллюски
-
 Морские брюхоногие моллюски — это морские улитки или морские слизни . Этот голожаберный моллюск — морской слизень.
Морские брюхоногие моллюски — это морские улитки или морские слизни . Этот голожаберный моллюск — морской слизень. -
 Морская улитка Syrinx aruanus имеет раковину длиной до 91 см, самую крупную из ныне живущих брюхоногих моллюсков.
Морская улитка Syrinx aruanus имеет раковину длиной до 91 см, самую крупную из ныне живущих брюхоногих моллюсков. -
 У моллюсков обычно есть глаза. Окаймляющий край мантии гребешка , двустворчатого моллюска , может быть свыше 100 простых глаз .
У моллюсков обычно есть глаза. Окаймляющий край мантии гребешка , двустворчатого моллюска , может быть свыше 100 простых глаз . -
 Обыкновенная мидия , еще один двустворчатый моллюск
Обыкновенная мидия , еще один двустворчатый моллюск




К головоногим относятся осьминоги , кальмары и каракатицы . Выявлено около 800 ныне живущих видов морских головоногих моллюсков. [ 271 ] около 11 000 вымерших таксонов . и описано [ 272 ] Они встречаются во всех океанах, но полностью пресноводных головоногих не существует. [ 273 ]
- Головоногие моллюски
-
![Наутилус — это живое ископаемое, мало изменившееся с тех пор, как он появился 500 миллионов лет назад как одно из первых головоногих моллюсков.[274][275][276]](//upload.wikimedia.org/wikipedia/commons/thumb/d/d5/Nautilus_Palau.JPG/300px-Nautilus_Palau.JPG) Наутилус мало — это живое ископаемое, изменившееся с тех пор, как он появился 500 миллионов лет назад как одно из первых головоногих моллюсков . [ 274 ] [ 275 ] [ 276 ]
Наутилус мало — это живое ископаемое, изменившееся с тех пор, как он появился 500 миллионов лет назад как одно из первых головоногих моллюсков . [ 274 ] [ 275 ] [ 276 ] -
 Реконструкция аммонита , очень успешного раннего головоногого моллюска, появившегося 400 млн лет назад .
Реконструкция аммонита , очень успешного раннего головоногого моллюска, появившегося 400 млн лет назад . -
 Головоногие моллюски, как и эта каракатица , используют свою мантийную полость для реактивного движения .
Головоногие моллюски, как и эта каракатица , используют свою мантийную полость для реактивного движения . -
![Колоссальный кальмар, самый крупный из всех беспозвоночных[277]](//upload.wikimedia.org/wikipedia/commons/thumb/3/37/Calmarcolossal.jpg/157px-Calmarcolossal.jpg) Колоссальный кальмар , самый крупный из всех беспозвоночных [ 277 ]
Колоссальный кальмар , самый крупный из всех беспозвоночных [ 277 ]
![Наутилус — это живое ископаемое, мало изменившееся с тех пор, как он появился 500 миллионов лет назад как одно из первых головоногих моллюсков.[274][275][276]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/d5/Nautilus_Palau.JPG/300px-Nautilus_Palau.JPG)


![Колоссальный кальмар, самый крупный из всех беспозвоночных[277]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/37/Calmarcolossal.jpg/157px-Calmarcolossal.jpg)
Моллюски имеют настолько разнообразные формы, что многие учебники основывают свои описания анатомии моллюсков на обобщенных или гипотетических предках моллюсков . Этот генерализованный моллюск несегментирован и двусторонне-симметричен , нижняя сторона состоит из одной мускулистой ножки . Помимо этого, у него есть еще три ключевые особенности. Во-первых, у него есть мускулистый покров, называемый мантией, покрывающий его внутренности и содержащий значительную полость, используемую для дыхания и выделения . Раковина , выделяемая мантией, покрывает верхнюю поверхность. Во-вторых (кроме двустворчатых моллюсков) у них есть грубый язык, называемый радулой, используемый для питания. В-третьих, у него есть нервная система , включая сложную пищеварительную систему, использующую микроскопические волоски с мускульной силой, называемые ресничками, для выделения слизи . Генерализованный моллюск имеет две парные нервные цепочки (у двустворчатых моллюсков — три). Мозг у видов , у которых он есть, окружает пищевод . У большинства моллюсков есть глаза , и у всех есть датчики, реагирующие на химические вещества, вибрацию и прикосновение. [ 278 ] [ 279 ]
Существуют убедительные доказательства появления морских брюхоногих, головоногих и двустворчатых моллюсков в кембрийский период от 538,8 до 485,4 миллионов лет назад .
Морские членистоногие
[ редактировать ]

Членистоногие (по-гречески « членистые лапы ») имеют экзоскелет (внешний скелет ), сегментированное тело и суставные придатки (парные придатки). Они образуют тип , который включает насекомых , паукообразных , многоножек и ракообразных . Членистоногие характеризуются членистыми конечностями и кутикулой из хитина , часто минерализованного карбонатом кальция . членистоногих Строение тела состоит из сегментов , каждый из которых имеет пару придатков . Жесткая кутикула тормозит рост, поэтому членистоногие периодически заменяют ее путем линьки . Их универсальность позволила им стать самыми богатыми видами среди всех экологических гильдий в большинстве сред.
Эволюционное происхождение членистоногих восходит к кембрийскому периоду и обычно считается монофилетическим . Однако базальные связи членистоногих с вымершими типами, такими как лобоподии . в последнее время обсуждаются [ 282 ] [ 283 ]
| панартроподы |
| ||||||||||||


- Окаменелости членистоногих и живые ископаемые
-
![Ископаемый трилобит. Трилобиты впервые появились около 521 млн лет назад. Они имели большой успех и были обнаружены повсюду в океане в течение 270 млн лет назад.[285]](//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Cheirurus_ingricus.png/301px-Cheirurus_ingricus.png) Ископаемый трилобит . Трилобиты впервые появились около 521 млн лет назад . Они имели большой успех и были обнаружены повсюду в океане на протяжении 270 млн лет назад. [ 285 ]
Ископаемый трилобит . Трилобиты впервые появились около 521 млн лет назад . Они имели большой успех и были обнаружены повсюду в океане на протяжении 270 млн лет назад. [ 285 ] -
 Anomalocaris (« аномальные креветки») был одним из первых высших хищников и впервые появился около 515 млн лет назад.
Anomalocaris (« аномальные креветки») был одним из первых высших хищников и впервые появился около 515 млн лет назад. -
 Самое крупное из известных членистоногих, морской скорпион Jaekelopterus rhenaniae , было обнаружено в слоях эстуариев примерно 390 млн лет назад. Его длина составляла до 2,5 м (8,2 фута). [ 286 ] [ 287 ]
Самое крупное из известных членистоногих, морской скорпион Jaekelopterus rhenaniae , было обнаружено в слоях эстуариев примерно 390 млн лет назад. Его длина составляла до 2,5 м (8,2 фута). [ 286 ] [ 287 ] -
![Xiphosurans, группа, включающая современных мечехвостов, появилась около 480 млн лет назад.[288]](//upload.wikimedia.org/wikipedia/commons/thumb/d/d4/Limulus_%28cropped%29.jpg/257px-Limulus_%28cropped%29.jpg) Xiphosurans , группа, включающая современных мечехвостов, появилась около 480 млн лет назад. [ 288 ]
Xiphosurans , группа, включающая современных мечехвостов, появилась около 480 млн лет назад. [ 288 ]
![Ископаемый трилобит. Трилобиты впервые появились около 521 млн лет назад. Они имели большой успех и были обнаружены повсюду в океане в течение 270 млн лет назад.[285]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Cheirurus_ingricus.png/301px-Cheirurus_ingricus.png)


![Xiphosurans, группа, включающая современных мечехвостов, появилась около 480 млн лет назад.[288]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/d4/Limulus_%28cropped%29.jpg/257px-Limulus_%28cropped%29.jpg)
Размеры современных морских членистоногих варьируются от микроскопического ракообразного Stygotantulus до японского краба-паука . Первичная внутренняя полость членистоногих представляет собой гемоцель , в котором расположены их внутренние органы и через который циркулирует их гемолимфа — аналог крови ; у них открытая система кровообращения . Как и их внешний вид, внутренние органы членистоногих обычно состоят из повторяющихся сегментов. Их нервная система имеет «лестничную форму»: парные вентральные нервные шнуры проходят через все сегменты и образуют парные ганглии в каждом сегменте . Их головы образуются в результате слияния различного числа сегментов, а мозг формируется в результате слияния ганглиев этих сегментов и окружает пищевод . Дыхательная подтипа и выделительная системы членистоногих различаются в зависимости как от окружающей среды, так и от , к которому они принадлежат.
- Современные ракообразные
-
 Многие ракообразные очень маленькие, как этот крошечный амфипод океана , и составляют значительную часть зоопланктона .
Многие ракообразные очень маленькие, как этот крошечный амфипод океана , и составляют значительную часть зоопланктона . -
![У японского краба-паука самый длинный размах ног среди членистоногих: длина от клешни до клешни достигает 5,5 метров (18 футов).[289]](//upload.wikimedia.org/wikipedia/commons/thumb/a/a7/Macrocheira_kaempferi.jpg/314px-Macrocheira_kaempferi.jpg) У японского краба-паука самый длинный размах ног среди членистоногих: длина от клешни до клешни достигает 5,5 метров (18 футов). [ 289 ]
У японского краба-паука самый длинный размах ног среди членистоногих: длина от клешни до клешни достигает 5,5 метров (18 футов). [ 289 ] -
![Тасманский гигантский краб долгожитель и медленно растет, что делает его уязвимым для чрезмерного вылова рыбы.[290]](//upload.wikimedia.org/wikipedia/commons/thumb/2/2c/J_J_Wild_Pseudocarcinus_cropped.jpg/330px-J_J_Wild_Pseudocarcinus_cropped.jpg) Тасманский гигантский краб долгоживущий и медленнорастущий, что делает его уязвимым для чрезмерного вылова рыбы. [ 290 ]
Тасманский гигантский краб долгоживущий и медленнорастущий, что делает его уязвимым для чрезмерного вылова рыбы. [ 290 ] -
![У креветок-богомолов самые развитые глаза в животном мире,[291] они разбивают добычу, размахивая своими булавообразными хищными когтями.[292]](//upload.wikimedia.org/wikipedia/commons/thumb/0/03/Odontodactylus_scyllarus_2.png/298px-Odontodactylus_scyllarus_2.png) У креветок-богомолов самые развитые глаза в животном мире. [ 291 ] и разбивают добычу, размахивая своими булавообразными хищными когтями. [ 292 ]
У креветок-богомолов самые развитые глаза в животном мире. [ 291 ] и разбивают добычу, размахивая своими булавообразными хищными когтями. [ 292 ]

![У японского краба-паука самый длинный размах ног среди членистоногих: длина от клешни до клешни достигает 5,5 метров (18 футов).[289]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/a7/Macrocheira_kaempferi.jpg/314px-Macrocheira_kaempferi.jpg)
![Тасманский гигантский краб долгожитель и медленно растет, что делает его уязвимым для чрезмерного вылова рыбы.[290]](http://upload.wikimedia.org/wikipedia/commons/thumb/2/2c/J_J_Wild_Pseudocarcinus_cropped.jpg/330px-J_J_Wild_Pseudocarcinus_cropped.jpg)
![У креветок-богомолов самые развитые глаза в животном мире,[291] они разбивают добычу, размахивая своими булавообразными хищными когтями.[292]](http://upload.wikimedia.org/wikipedia/commons/thumb/0/03/Odontodactylus_scyllarus_2.png/298px-Odontodactylus_scyllarus_2.png)
Зрение членистоногих зависит от различных комбинаций сложных глаз с пигментными ямками и глазков : у большинства видов глазки могут определять только направление, откуда исходит свет, а сложные глаза являются основным источником информации. Членистоногие также обладают широким спектром химических и механических сенсоров, в основном основанных на модификациях множества щетинок (щетинок), которые выступают через их кутикулу. Методы размножения членистоногих разнообразны: наземные виды используют ту или иную форму внутреннего оплодотворения, тогда как морские виды откладывают яйца, используя либо внутреннее, либо внешнее оплодотворение . Детеныши членистоногих варьируются от миниатюрных взрослых особей до личинок, у которых нет членистых конечностей, и в конечном итоге претерпевают полную метаморфозу , образуя взрослую форму.
вторичноротые
[ редактировать ]У вторичноротых первым отверстием, развивающимся у растущего зародыша, становится анус , а у протостомов — рот. образуют надтип животных Вторыеротые и являются сестринской кладой протостомов . [ 251 ] [ 252 ] Когда-то считалось, что самые ранние известные вторичноротые — это окаменелости Saccorhytus, датированные примерно 540 миллионами лет назад. [ 293 ] Однако другое исследование показало, что Saccorhytus, скорее всего, является экдизозоем . [ 294 ]
| ← вторичноротые | |
| (сохранившийся) |

Иглокожие
[ редактировать ]
Иглокожие (по-гречески « шипистая кожа ») — это тип, в состав которого входят только морские беспозвоночные. Тип насчитывает около 7000 живых видов . [ 295 ] что делает его второй по величине группой вторичноротых после хордовых .
Взрослые иглокожие узнаваемы по радиальной симметрии (обычно пятиточечной) и включают морские звезды , морские ежи , песчаные доллары и морские огурцы , а также морские лилии . [ 296 ] Иглокожие встречаются на любой глубине океана, от приливной зоны до абиссальной зоны . Они уникальны среди животных, поскольку имеют двустороннюю симметрию на личиночной стадии и пятикратную симметрию ( пентамеризм , особый тип радиальной симметрии) во взрослом возрасте. [ 297 ]
Иглокожие важны как с биологической, так и с геологической точки зрения. есть несколько других группировок, которые были бы так многочисленны С биологической точки зрения, в биотической пустыне глубокого моря , а также в более мелких океанах . Большинство иглокожих способны к регенерации тканей, органов, конечностей, а также размножаются бесполым путем ; в некоторых случаях они могут подвергнуться полной регенерации из одной конечности. С геологической точки зрения ценность иглокожих заключается в их окостеневших скелетах , которые вносят основной вклад во многие известняковые образования и могут дать ценную информацию о геологической среде. Они были наиболее часто используемыми видами в регенеративных исследованиях в 19 и 20 веках.
-
 Иглокожие буквально означает «колючая кожа», как показывает этот арбузный морской еж .
Иглокожие буквально означает «колючая кожа», как показывает этот арбузный морской еж . -
![Охряная морская звезда была первым краеугольным хищником, которого удалось изучить. Они ограничивают численность мидий, которые могут уничтожить приливные сообщества.[298]](//upload.wikimedia.org/wikipedia/commons/thumb/8/89/Ochre_sea_star.jpg/360px-Ochre_sea_star.jpg) Охряная морская звезда была первым краеугольным хищником, которого удалось изучить. Они ограничивают численность мидий , которые могут уничтожить приливные сообщества. [ 298 ]
Охряная морская звезда была первым краеугольным хищником, которого удалось изучить. Они ограничивают численность мидий , которые могут уничтожить приливные сообщества. [ 298 ] -
 Красочные морские лилии на мелководье
Красочные морские лилии на мелководье

![Охряная морская звезда была первым краеугольным хищником, которого удалось изучить. Они ограничивают численность мидий, которые могут уничтожить приливные сообщества.[298]](http://upload.wikimedia.org/wikipedia/commons/thumb/8/89/Ochre_sea_star.jpg/360px-Ochre_sea_star.jpg)

-
 Морские огурцы фильтруют питание планктоном и взвешенными веществами.
Морские огурцы фильтруют питание планктоном и взвешенными веществами. -
 Морская свинья , глубоководный морской огурец, — единственное иглокожее животное, которое передвигается на ногах.
Морская свинья , глубоководный морской огурец, — единственное иглокожее животное, которое передвигается на ногах. -
 Бентопелагический и биолюминесцентный морской огурец , плавающий на глубине 3200 метров.
Бентопелагический и биолюминесцентный морской огурец , плавающий на глубине 3200 метров.



Некоторые учёные полагают, что радиация иглокожих была ответственна за Мезозойскую морскую революцию . Помимо трудно классифицируемого аркаруа ( докембрийского животного с иглокожей пятичленной радиальной симметрией), первые окончательные представители этого типа появились примерно в начале кембрия .
полухордовые
[ редактировать ]
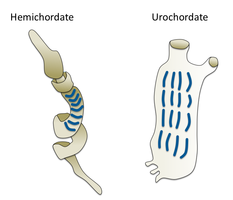
Полухордовые образуют сестринский тип иглокожих . Это одиночные червеобразные организмы, которые люди редко видят из-за своего образа жизни. Они включают две основные группы: желудевые черви и Pterobranchia . Птеробранхии образуют класс, включающий около 30 видов мелких червеобразных животных, обитающих в секретируемых трубках на дне океана. Желудевые черви образуют класс, насчитывающий около 111 видов, которые обычно обитают в U-образных норах на морском дне, от береговой линии до глубины 3000 метров. Черви лежат там, высунув хобот из одного отверстия в норе, питаясь отложениями или суспензией. Предполагается, что предки желудевых червей раньше жили в трубках, как и их родственники, Pterobranchia, но со временем начали вести более безопасный и защищенный образ жизни в норах из отложений. [ 302 ] Некоторые из этих червей могут вырасти очень длинными; один конкретный вид может достигать длины 2,5 метра (8 футов 2 дюйма), хотя большинство желудевых червей намного меньше.
Желудевые черви более узкоспециализированы и развиты, чем другие червеобразные организмы. У них есть система кровообращения с сердцем, которое также функционирует как почка. Желудевые черви имеют жаберные структуры, которые они используют для дыхания, похожие на жабры рыб. Поэтому иногда говорят, что желудевые черви являются связующим звеном между классическими беспозвоночными и позвоночными . Желудевые черви по мере роста размеров постоянно образуют новые жаберные щели, а у некоторых пожилых особей их бывает более сотни с каждой стороны. Каждая щель состоит из жаберной камеры, открывающейся в глотку через U-образную щель. Реснички проталкивают воду через щели, поддерживая постоянный поток, как у рыб. [ 303 ] У некоторых желудевых червей также есть постанальный хвост, который может быть гомологичен постанальному хвосту позвоночных.
Трехчастное строение тела желудевого червя больше не присутствует у позвоночных, за исключением анатомии лобной нервной трубки, которая позже развилась в мозг, разделенный на три части. Это означает, что часть исходной анатомии ранних предков хордовых все еще присутствует у позвоночных, даже если она не всегда видна. Одна из теорий состоит в том, что трехчастное тело произошло от раннего общего предка вторичноротых и, возможно, даже от общего двустороннего предка как вторичноротых, так и протостомов. Исследования показали, что экспрессия генов у эмбриона имеет три одинаковых сигнальных центра, которые формируют мозг всех позвоночных, но вместо того, чтобы принимать участие в формировании их нервной системы, [ 304 ] они контролируют развитие различных областей тела. [ 305 ]
Морские хордовые
[ редактировать ]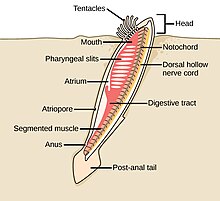
Тип хордовых включает три подтипа, один из которых — позвоночные (см. Ниже). Два других подтипа — морские беспозвоночные: оболочники ( сальпы и асцидии ) и головохордовые (например, ланцетники ). Беспозвоночные хордовые — близкие родственники позвоночных. В частности, обсуждался вопрос о том, насколько тесно некоторые вымершие морские виды, такие как Pikaiidae , Palaeospondylus , Zhongxiniscus и Vetulicolia , могли быть связаны предками с позвоночными.
- Беспозвоночные хордовые — близкие родственники позвоночных.
-
![Ланцетник, небольшое полупрозрачное рыбоподобное головохордовое животное, является одним из ближайших ныне живущих беспозвоночных родственников позвоночных.[307][308]](//upload.wikimedia.org/wikipedia/commons/thumb/5/54/Amphioxus.png/332px-Amphioxus.png) Ланцетник , является одним из ближайших , небольшое полупрозрачное рыбоподобное головохордовое животное ныне живущих беспозвоночных родственников позвоночных. [ 307 ] [ 308 ]
Ланцетник , является одним из ближайших , небольшое полупрозрачное рыбоподобное головохордовое животное ныне живущих беспозвоночных родственников позвоночных. [ 307 ] [ 308 ] -
![Оболочники, такие как эти флуоресцентные асцидии, могут дать ключ к разгадке происхождения позвоночных, а, следовательно, и человека.[309]](//upload.wikimedia.org/wikipedia/commons/thumb/8/8c/Ascidian_%28Rhopalaea_Crassa%29_%284_cm%29.png/280px-Ascidian_%28Rhopalaea_Crassa%29_%284_cm%29.png) Оболочники, как и эти флюоресцентные асцидии , могут дать ключ к разгадке происхождения позвоночных, а, следовательно, и человека. [ 309 ]
Оболочники, как и эти флюоресцентные асцидии , могут дать ключ к разгадке происхождения позвоночных, а, следовательно, и человека. [ 309 ] -
 Пиросомы представляют собой свободно плавающие биолюминесцентные оболочки, состоящие из сотен особей.
Пиросомы представляют собой свободно плавающие биолюминесцентные оболочки, состоящие из сотен особей. -
 Сальповая цепь
Сальповая цепь
![Ланцетник, небольшое полупрозрачное рыбоподобное головохордовое животное, является одним из ближайших ныне живущих беспозвоночных родственников позвоночных.[307][308]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/54/Amphioxus.png/332px-Amphioxus.png)
![Оболочники, такие как эти флуоресцентные асцидии, могут дать ключ к разгадке происхождения позвоночных, а, следовательно, и человека.[309]](http://upload.wikimedia.org/wikipedia/commons/thumb/8/8c/Ascidian_%28Rhopalaea_Crassa%29_%284_cm%29.png/280px-Ascidian_%28Rhopalaea_Crassa%29_%284_cm%29.png)


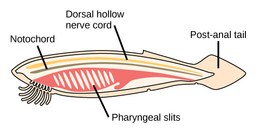
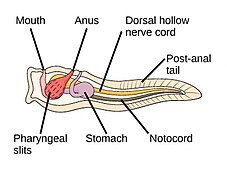
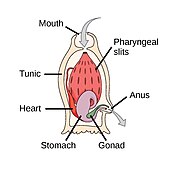
Позвоночные животные
[ редактировать ]
Позвоночные (лат. позвоночника ) — подтип хордовых суставы . Это хордовые животные, у которых есть позвоночник (позвоночник). Позвоночник обеспечивает центральную опорную структуру для внутреннего скелета , которая придает форму, поддержку и защиту телу и может служить средством крепления плавников или конечностей к телу. Позвоночник также служит для размещения и защиты спинного мозга , находящегося внутри позвоночного столба.
Морских позвоночных можно разделить на морских рыб и морских четвероногих .
Морская рыба
[ редактировать ]Рыбы обычно дышат, извлекая кислород из воды через жабры , а их кожа защищена чешуей и слизистой . Они используют плавники , чтобы двигаться и стабилизировать себя в воде, и обычно имеют двухкамерное сердце и глаза, хорошо приспособленные к видению под водой, а также другие сенсорные системы . По состоянию на 2017 год описано более 33 000 видов рыб. [ 310 ] из них около 20 000 – морские рыбы. [ 311 ]
| ← позвоночные животные | |
| (сохранившийся) |
Бесчелюстная рыба
[ редактировать ]
Ранние рыбы не имели челюстей . Большинство из них вымерло, когда их вытеснили челюстные рыбы (внизу), но выжили две группы: миксины и миноги . Миксы образуют класс, насчитывающий около 20 видов угревидных морских рыб , производящих слизь . Это единственные известные живые животные, у которых есть череп , но нет позвоночника . Миноги образуют надкласс, включающий 38 известных современных видов бесчелюстных рыб . [ 312 ] Для взрослой миноги характерен зубчатый воронкообразный присасывающийся рот. Хотя они хорошо известны тем, что вгрызаются в плоть других рыб, чтобы высосать из них кровь , [ 313 ] только 18 видов миног являются фактически паразитическими. [ 314 ] Вместе миксины и миноги являются сестринской группой позвоночных. Живые миксины остаются похожими на миксин, живших примерно 300 миллионов лет назад. [ 315 ] Миноги — очень древняя линия позвоночных, хотя их точное родство с миксинами и челюстными позвоночными до сих пор остается предметом споров. [ 316 ] Молекулярный анализ, проведенный с 1992 года, показал, что миксины наиболее тесно связаны с миногами. [ 317 ] то же самое относится и к позвоночным в монофилетическом смысле. Другие считают их сестринской группой позвоночных животных общего таксона craniata. [ 318 ]
Монстр Талли — вымерший род мягкотелых двулатерий, обитавший в тропических эстуариях около 300 миллионов лет назад. С 2016 года ведутся споры о том, было ли это животное позвоночным или беспозвоночным. [ 319 ] [ 320 ] В 2020 году исследователи нашли «убедительные доказательства» того, что монстр Талли был позвоночным и бесчелюстной рыбой в линии миноги . [ 321 ] [ 322 ] в то время как в 2023 году другие исследователи обнаружили, что 3D-сканирование окаменелостей не подтверждает эти выводы. [ 323 ]
-

-
 Миноги часто являются паразитами и имеют зубчатый воронкообразный присасывающийся рот.
Миноги часто являются паразитами и имеют зубчатый воронкообразный присасывающийся рот. -
 Вымершие птераспидоморфы , предки челюстных позвоночных.
Вымершие птераспидоморфы , предки челюстных позвоночных.



Pteraspidomorphi — вымерший класс ранних бесчелюстных рыб, предков челюстных позвоночных. Те немногие характеристики, которые они разделяют с последними, теперь считаются примитивными для всех позвоночных .
Примерно в начале девона начали появляться рыбы с глубокой перестройкой черепа позвоночных, в результате которой появилась челюсть . [ 324 ] Все челюсти позвоночных, включая человеческую, произошли от этих ранних рыбьих челюстей. Появление челюстей ранних позвоночных было описано как «возможно, самый глубокий и радикальный эволюционный шаг в истории позвоночных». [ 325 ] [ 326 ] Челюсти позволяют захватывать, удерживать и пережевывать добычу. Рыбам без челюстей было труднее выжить, чем рыбам с челюстями, и большинство бесчелюстных рыб вымерло в триасовый период.
Хрящевая рыба
[ редактировать ]Челюстные рыбы делятся на две основные группы: рыбы с костным внутренним скелетом и рыбы с хрящевым внутренним скелетом . У хрящевых рыб, таких как акулы и скаты , челюсти и скелет состоят из хряща, а не из костей . Мегалодон — вымерший вид акул, живший примерно от 28 до 1,5 млн лет назад. Возможно, она очень напоминала коренастую версию большой белой акулы , но была намного крупнее: предполагаемая длина достигала 20,3 метра (67 футов). [ 327 ] Встречается во всех океанах [ 328 ] это был один из самых крупных и могущественных хищников в истории позвоночных. [ 327 ] и, вероятно, оказали глубокое влияние на морскую жизнь. [ 329 ] Гренландская акула имеет самую длинную известную продолжительность жизни среди всех позвоночных - около 400 лет. [ 330 ] Некоторые акулы, такие как большая белая, частично теплокровны и рожают живорождений. Скат манта , крупнейший скат в мире, стал объектом рыболовства и теперь уязвим . [ 331 ]
- Хрящевые рыбы
-
 Хрящевые рыбы, возможно, произошли от колючих акул .
Хрящевые рыбы, возможно, произошли от колючих акул . -

-
 Манта , самый крупный скат
Манта , самый крупный скат -
![Рыба-пила, скаты с длинными рострумами, напоминающими пилу. Все виды сейчас находятся под угрозой исчезновения.[332]](//upload.wikimedia.org/wikipedia/commons/thumb/4/48/Pristis_clavata_2.jpg/407px-Pristis_clavata_2.jpg) Рыба-пила , скаты с длинными рострумами, напоминающими пилу. Все виды сейчас находятся под угрозой исчезновения . [ 332 ]
Рыба-пила , скаты с длинными рострумами, напоминающими пилу. Все виды сейчас находятся под угрозой исчезновения . [ 332 ]



![Рыба-пила, скаты с длинными рострумами, напоминающими пилу. Все виды сейчас находятся под угрозой исчезновения.[332]](http://upload.wikimedia.org/wikipedia/commons/thumb/4/48/Pristis_clavata_2.jpg/407px-Pristis_clavata_2.jpg)
-
 Вымерший мегалодон напоминал гигантскую большую белую акулу .
Вымерший мегалодон напоминал гигантскую большую белую акулу . -
 живет Гренландская акула дольше, чем любое другое позвоночное животное.
живет Гренландская акула дольше, чем любое другое позвоночное животное. -




Костистая рыба
[ редактировать ]


У костистых рыб челюсти и скелет состоят из костей, а не хрящей . У костных рыб также есть твердые костные пластины, называемые жаберными крышками , которые помогают им дышать и защищают жабры, а также у них часто есть плавательный пузырь , который они используют для лучшего контроля своей плавучести. Костных рыб можно разделить на рыб с лопастными плавниками и с лучевыми плавниками . Примерные даты на филогенетическом дереве взяты из Near et al., 2012. [ 333 ] и Чжу и др., 2009. [ 334 ]
| ← костистая рыба |
| ||||||||||||||||||
| (сохранившийся) |
Лопастные плавники имеют форму мясистых лопастей, поддерживаемых костными стеблями, отходящими от тела. [ 335 ] Гуйю онейрос , самая ранняя из известных костистых рыб, жила в позднем силуре 419 миллионов лет назад. Он сочетает в себе как лучепёрые, так и лопастноперые черты, хотя анализ совокупности признаков приближает его к лопастеперым рыбам. [ 334 ] Лопастные плавники превратились в ноги первых четвероногих наземных позвоночных, поэтому ранним предком человека были рыбы с лопастными плавниками. Помимо целакантов и двоякодышащих рыб, сейчас вымерли лопастноперые рыбы.
Остальные костистые рыбы имеют лучевые плавники. Они состоят из переплетений кожи, поддерживаемых костными или роговыми шипами (лучами), которые можно поднимать, чтобы контролировать жесткость плавников.
- Основной отличительной особенностью хондростеев ( осетр , веслонос , бичир и камыш ) — хрящевой характер их скелетов. Предками хондростеев считаются костистые рыбы, однако в более позднем эволюционном развитии признак окостеневшего скелета был утерян, что привело к облегчению тела. [ 336 ]
- Неоптериги (от греческого « новые плавники ») появились где-то в поздней перми, до появления динозавров. Это была очень успешная группа рыб, потому что они могли двигаться быстрее, чем их предки. В ходе эволюции их чешуя и скелет начали светлеть, а челюсти стали более мощными и эффективными. [ 337 ]
костистые кости
[ редактировать ]
Около 96% всех современных видов рыб являются костистыми. [ 338 ] из них около 14 000 являются морскими видами. [ 339 ] Костистых рыб можно отличить от других костистых рыб по гомоцеркальному хвосту — хвосту, верхняя половина которого отражает нижнюю половину. [ 340 ] Еще одно отличие заключается в костях челюстей: у костистых рыб есть изменения в мускулатуре челюстей, которые позволяют им выдвигать челюсти . Это позволяет им хватать добычу и втягивать ее в рот . [ 340 ] В целом костистые кости, как правило, более быстрые и гибкие, чем более базальнокостные рыбы. Их скелетная структура эволюционировала в сторону большей легкости. Хотя костные кости хорошо кальцинированы , они состоят из каркаса из распорок, а не из плотных губчатых костей голостовых рыб . [ 341 ]
Костистые кости встречаются почти во всех морских средах обитания . [ 342 ] Они имеют огромное разнообразие и варьируются по размеру от взрослых бычков длиной 8 мм. [ 343 ] океанской солнечной рыбы весом более 2000 кг. [ 344 ] Следующие изображения демонстрируют разнообразие форм и окраски современных морских костистых рыб...








Почти половина всех современных видов позвоночных — костистые. [ 345 ]
Морские четвероногие
[ редактировать ]
Четвероногий конечности (по-гречески четыре ноги ) — позвоночное животное, имеющее ( ступни). Четвероногие произошли от древних лопастных рыб около 400 миллионов лет назад, в девонский период , когда их самые ранние предки вышли из моря и приспособились к жизни на суше. [ 346 ] Это изменение от плана тела для дыхания и навигации в воде с нейтральной гравитацией к плану тела с механизмами, позволяющими животному вдыхать воздух без обезвоживания и передвигаться по суше, является одним из самых глубоких известных эволюционных изменений. [ 347 ] [ 348 ] Четвероногих можно разделить на четыре класса: амфибии , рептилии , птицы и млекопитающие .
| ← четвероногие |
| ||||||||||||||||||
Морские четвероногие — четвероногие, которые снова вернулись с суши обратно в море. Первые возвращения в океан могли произойти еще в каменноугольном периоде. [ 349 ] тогда как другие возвращения произошли совсем недавно, в кайнозое , например, у китообразных, ластоногих , [ 350 ] и несколько современных амфибий . [ 351 ] Земноводные (от греческого «оба вида жизни ») проводят часть своей жизни в воде, а часть — на суше. Для размножения им в основном требуется пресная вода. Некоторые обитают в солоноватой воде, но настоящих морских амфибий не существует. [ 352 ] Однако поступали сообщения о вторжении амфибий в морские воды, например, о вторжении в Черное море естественного гибрида Pelophylax esculentus, о котором сообщалось в 2010 году. [ 353 ]
Рептилии
[ редактировать ]Рептилии (позднелатинское слово «ползание» или «ползание ») не имеют водной личиночной стадии и в этом отношении отличаются от амфибий. Большинство рептилий яйцекладущие, хотя некоторые виды чешуйчатых живородящие , как и некоторые вымершие водные клады. [ 354 ] — плод развивается внутри матери, находясь в плаценте, а не в яичной скорлупе . Яйца рептилий, как и амниоты , окружены оболочками для защиты и транспортировки, которые адаптируют их к размножению на суше. Многие из живородящих видов выкармливают своих зародышей через различные формы плаценты, аналогичные плацентам млекопитающих , а некоторые обеспечивают первоначальный уход за вылупившимися птенцами.
Некоторые рептилии более тесно связаны с птицами, чем другие рептилии, и многие ученые предпочитают считать Reptilia монофилетической группой, включающей птиц. [ 355 ] [ 356 ] [ 357 ] [ 358 ] К современным нептичьим рептилиям, населяющим или часто посещающим море, относятся морские черепахи , морские змеи , черепахи , морская игуана и морской крокодил . В настоящее время из примерно 12 000 существующих рептилий только около 100 относятся к морским рептилиям. видов и подвидов [ 359 ]
За исключением некоторых морских змей, большинство современных морских рептилий являются яйцекладущими , и им необходимо возвращаться на сушу, чтобы отложить яйца. За исключением морских черепах, эти виды обычно проводят большую часть своей жизни на суше или вблизи нее, а не в океане. Морские змеи обычно предпочитают мелководье вблизи суши, вокруг островов, особенно в несколько защищенных водах, а также вблизи эстуариев. [ 360 ] [ 361 ] В отличие от наземных змей, морские змеи развили уплощенный хвост, который помогает им плавать. [ 362 ]
-

-

-

-
 У морских змей приплюснутые хвосты.
У морских змей приплюснутые хвосты. -
 Древний Ichthyosaurus communis независимо развил ласты, похожие на дельфины.
Древний Ichthyosaurus communis независимо развил ласты, похожие на дельфины.





Некоторые вымершие морские рептилии, такие как ихтиозавры , в ходе эволюции стали живородящими и им не требовалось возвращаться на сушу. Ихтиозавры напоминали дельфинов. Впервые они появились около 245 миллионов лет назад и исчезли около 90 миллионов лет назад. Наземный предок ихтиозавра не имел на спине или хвосте никаких особенностей, которые могли бы помочь в эволюционном процессе. Тем не менее, у ихтиозавра развился спинной и хвостовой плавник , что улучшило его способность плавать. [ 363 ] Биолог Стивен Джей Гулд сказал, что ихтиозавр был его любимым примером конвергентной эволюции . [ 364 ] Первые морские рептилии возникли в перми . В течение мезозоя многие группы рептилий приспособились к жизни в морях, в том числе ихтиозавры , плезиозавры , мозазавры , нотозавры , плакодонты , морские черепахи , талаттозавры и талаттозухи . Морские рептилии стали менее многочисленными после массового вымирания в конце мелового периода .
Птицы
[ редактировать ]
Морские птицы приспособлены к жизни в морской среде. Их часто называют морскими птицами . Хотя морские птицы сильно различаются по образу жизни, поведению и физиологии, они часто демонстрируют поразительную конвергентную эволюцию , поскольку одни и те же экологические проблемы и кормовые ниши привели к схожим адаптациям. Примеры включают альбатросов , пингвинов , олушей и гагарок .
В целом морские птицы живут дольше, размножаются позже и рожают меньше детенышей, чем наземные птицы, но они уделяют много времени своему потомству. Большинство видов гнездятся колониями , размер которых может варьироваться от нескольких десятков птиц до миллионов. Многие виды известны своими длительными ежегодными миграциями , в некоторых случаях пересекающими экватор или вокруг Земли. Они питаются как на поверхности океана, так и под ним, и даже питаются друг другом. Морские птицы могут вести пелагический , прибрежный образ жизни или, в некоторых случаях, проводить часть года полностью вдали от моря. Некоторые морские птицы падают с высоты, ныряя в воду, оставляя за собой следы, похожие на пар, похожие на следы истребителей. [ 365 ] Олуши падают в воду со скоростью до 100 километров в час (60 миль в час). Под кожей на лице и груди у них есть воздушные мешки, которые действуют как пузырчатая пленка , смягчая удары водой.
-
 Европейская сельдевая чайка нападает на косяки сельди сверху.
Европейская сельдевая чайка нападает на косяки сельди сверху. -
 Гентуский пингвин плавает под водой
Гентуский пингвин плавает под водой -
 Альбатросы обитают на огромных территориях океана, а некоторые даже облетают земной шар.
Альбатросы обитают на огромных территориях океана, а некоторые даже облетают земной шар. -
 Олуши «пикирующие бомбы» на большой скорости
Олуши «пикирующие бомбы» на большой скорости




Первые морские птицы появились в меловом периоде , а современные семейства морских птиц возникли в палеогене .
Млекопитающие
[ редактировать ]
Млекопитающие (от латинского « грудь ») характеризуются наличием молочных желез , которые у самок производят молоко для вскармливания (выкармливания) детенышей. Здесь обитает около 130 ныне живущих и недавно вымерших видов морских млекопитающих, таких как тюлени , дельфины , киты , ламантины , каланы и белые медведи . [ 366 ] Они не представляют собой отдельный таксон или систематическую группу, а объединены тем, что в качестве источника питания они полагаются на морскую среду. И китообразные, и сирены полностью водны и поэтому являются обязательными обитателями воды. Тюлени и морские львы ведут полуводный образ жизни; они проводят большую часть своего времени в воде, но им необходимо вернуться на сушу для важных действий, таких как спаривание , размножение и линька . Напротив, и выдры, и белый медведь гораздо менее приспособлены к жизни в воде. Их рацион также значительно различается: некоторые могут питаться зоопланктоном ; другие могут есть рыбу, кальмаров, моллюсков и морскую траву; и некоторые могут есть других млекопитающих.
В процессе конвергентной эволюции морские млекопитающие, особенно китообразные, такие как дельфины и киты, изменили строение своего тела , чтобы оно стало параллельным обтекаемому веретенообразному строению тела пелагических рыб . Передние ноги превратились в ласты , задние исчезли, снова появился спинной плавник , а хвост превратился в мощный горизонтальный плавник . Такое строение тела является адаптацией к активному хищнику в условиях высокого сопротивления . Параллельная конвергенция произошла с ныне вымершей морской рептилией ихтиозавром . [ 367 ]
-
![Находящийся под угрозой исчезновения синий кит, самое крупное из ныне живущих животных[368]](//upload.wikimedia.org/wikipedia/commons/thumb/c/cd/Bluewhale2_noaa.jpg/270px-Bluewhale2_noaa.jpg)
-
![У афалины самая высокая энцефализация среди животных после человека[369]](//upload.wikimedia.org/wikipedia/commons/thumb/1/10/Tursiops_truncatus_01.jpg/271px-Tursiops_truncatus_01.jpg)
-

-
 Дюгонь пасется на морской траве
Дюгонь пасется на морской траве -

-

![Находящийся под угрозой исчезновения синий кит, самое крупное из ныне живущих животных[368]](http://upload.wikimedia.org/wikipedia/commons/thumb/c/cd/Bluewhale2_noaa.jpg/270px-Bluewhale2_noaa.jpg)
![У афалины самая высокая энцефализация среди животных после человека[369]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/10/Tursiops_truncatus_01.jpg/271px-Tursiops_truncatus_01.jpg)




Первичные производители
[ редактировать ]
Первичные продуценты — это автотрофные организмы, которые производят себе пищу, а не поедают другие организмы. Это означает, что первичные продуценты становятся отправной точкой в пищевой цепи для гетеротрофных организмов, которые поедают другие организмы. Некоторые морские первичные продуценты представляют собой специализированные бактерии и археи, которые являются хемотрофами , добывающими себе пищу, собираясь вокруг гидротермальных источников и холодных выходов и используя хемосинтез . Однако большая часть первичной морской продукции происходит от организмов, которые используют фотосинтез на углекислом газе, растворенном в воде. Этот процесс использует энергию солнечного света для преобразования воды и углекислого газа. [ 370 ] : 186–187 в сахара, которые можно использовать как источник химической энергии, так и органических молекул, входящих в состав структурных компонентов клеток. [ 370 ] : 1242 Морские первичные производители важны, потому что они лежат в основе почти всей жизни морских животных, генерируя большую часть кислорода и пищи, которые обеспечивают другие организмы химической энергией, необходимой им для существования.
Основными морскими первичными продуцентами являются цианобактерии , водоросли и морские растения. Кислород , выделяемый как побочный продукт фотосинтеза, необходим почти всем живым существам для осуществления клеточного дыхания . Кроме того, первичные производители оказывают влияние на глобальные углеродный и водный циклы. Они стабилизируют прибрежные районы и могут обеспечить среду обитания для морских животных. Термин «деление» традиционно использовался вместо типа «тип» при обсуждении первичных продуцентов, но Международный кодекс номенклатуры водорослей, грибов и растений теперь принимает оба термина как эквиваленты. [ 371 ]
Цианобактерии
[ редактировать ]

Цианобактерии были первыми организмами, развившими способность превращать солнечный свет в химическую энергию. Они образуют тип (подразделение) бактерий, который варьируется от одноклеточных до нитчатых и включает колониальные виды . Они встречаются практически повсюду на земле: во влажной почве, как в пресноводной, так и в морской среде и даже на антарктических скалах. [ 372 ] В частности, некоторые виды встречаются в виде дрейфующих клеток, плавающих в океане, и поэтому были одними из первых представителей фитопланктона .
Первыми первичными продуцентами, использовавшими фотосинтез, были океанические цианобактерии около 2,3 миллиарда лет назад. [ 373 ] [ 374 ] Выделение молекулярного кислорода цианобактериями как побочного продукта фотосинтеза вызвало глобальные изменения в окружающей среде Земли. Поскольку в то время кислород был токсичен для большей части жизни на Земле, это привело к почти исчезновению организмов, непереносимых к кислороду , — резкое изменение , которое изменило направление эволюции основных видов животных и растений. [ 375 ]
Крошечная морская цианобактерия Prochromococcus , открытая в 1986 году, сегодня составляет часть основы пищевой цепи океана и отвечает за большую часть фотосинтеза в открытом океане. [ 376 ] и примерно 20% кислорода в атмосфере Земли. [ 377 ] Возможно, это самый многочисленный род на Земле: один миллилитр поверхностной морской воды может содержать 100 000 и более клеток. [ 378 ]
Первоначально биологи классифицировали цианобактерии как водоросли и называли их «сине-зелеными водорослями». Более поздняя точка зрения состоит в том, что цианобактерии — это бактерии и, следовательно, даже не относятся к тому же царству , что и водоросли. Сегодня большинство авторитетных источников исключают всех прокариотов и, следовательно, цианобактерий из определения водорослей. [ 379 ] [ 380 ]
Водоросли
[ редактировать ]


Водоросли — неофициальный термин для широко распространенной и разнообразной группы фотосинтезирующих протистов , которые не обязательно тесно связаны между собой и, таким образом, являются полифилетическими . Морские водоросли можно разделить на шесть групп:
- зеленые водоросли , неофициальная группа, содержащая около 8000 признанных видов. [ 381 ] Многие виды живут большую часть своей жизни в виде одиночных клеток или являются нитевидными, тогда как другие образуют колонии, состоящие из длинных цепочек клеток, или представляют собой высокодифференцированные макроскопические водоросли.
- красные водоросли , (спорный) тип, содержащий около 7000 признанных видов, [ 382 ] в основном многоклеточные и включают множество известных морских водорослей . [ 382 ] [ 383 ]
- бурые водоросли , класс , содержащий около 2000 признанных видов, [ 384 ] в основном многоклеточные и включают множество морских водорослей, в том числе ламинарию.
- диатомовые водоросли — (спорный) тип, содержащий около 100 000 признанных видов преимущественно одноклеточных водорослей. Диатомовые водоросли производят около 20 процентов кислорода, производимого на планете каждый год. [ 147 ] поглощают более 6,7 миллиардов тонн кремния из вод, в которых они обитают, ежегодно [ 385 ] и составляют почти половину органического материала, обнаруженного в океанах. Раковины ( панцири ) мертвых диатомей могут достигать глубины до полумили на дне океана . [ 386 ]
- динофлагелляты — тип одноклеточных жгутиконосцев, насчитывающий около 2000 морских видов. [ 387 ] Известно, что многие динофлагелляты фотосинтезируют , но большая часть из них на самом деле миксотрофны , сочетая фотосинтез с поеданием добычи ( фаготрофия ). [ 388 ] Некоторые виды являются эндосимбионтами морских животных и играют важную роль в биологии коралловых рифов . Другие предшествуют другим простейшим, а некоторые формы являются паразитическими.
- эвгленофиты — тип одноклеточных жгутиконосцев, состоящий лишь из нескольких морских представителей.
В отличие от высших растений, у водорослей отсутствуют корни, стебли и листья. По размеру их можно разделить на микроводоросли и макроводоросли .
Микроводоросли — это микроскопические виды водорослей, невидимые невооруженным глазом. В основном это одноклеточные виды, существующие по отдельности, цепочками или группами, хотя некоторые из них являются многоклеточными . Микроводоросли являются важными компонентами морских простейших ( обсуждаемых выше ), а также фитопланктона ( обсуждаемых ниже ). Они очень разнообразны . По оценкам, существует 200 000–800 000 видов, из которых описано около 50 000 видов. [ 389 ] В зависимости от вида их размеры колеблются от нескольких микрометров (мкм) до нескольких сотен микрометров. Они специально адаптированы к среде, в которой преобладают силы вязкости.
-
 Chlamydomonas globosa , одноклеточная зеленая водоросль с двумя жгутиками , едва заметными внизу слева.
Chlamydomonas globosa , одноклеточная зеленая водоросль с двумя жгутиками , едва заметными внизу слева. -
![Chlorella vulgaris, обычная зеленая микроводоросль, в эндосимбиозе с инфузорией[390]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b6/%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg/293px-%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg)
-
 Центрическая диатомовая водоросль
Центрическая диатомовая водоросль -
 Динофлагелляты
Динофлагелляты

![Chlorella vulgaris, обычная зеленая микроводоросль, в эндосимбиозе с инфузорией[390]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b6/%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg/293px-%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg)


Макроводоросли — это более крупные, многоклеточные и более заметные виды водорослей, обычно называемые морскими водорослями . Морские водоросли обычно растут на мелководье в прибрежных водах, где они прикреплены к морскому дну с помощью крепления . Морские водоросли, которые плывут по течению, могут быть выброшены на пляжи. Келп — это крупная бурая морская водоросль, образующая обширные подводные леса, покрывающие около 25% береговой линии мира. [ 391 ] Они входят в число самых продуктивных и динамичных экосистем на Земле. [ 392 ] Некоторые водоросли Саргассума являются планктонными (свободно плавающими). Как и микроводоросли, макроводоросли (водоросли) технически являются морскими протистами , поскольку они не являются настоящими растениями.
-

-
 Саргассум – это планктонная бурая водоросль с воздушными пузырями, которые помогают ей плавать.
Саргассум – это планктонная бурая водоросль с воздушными пузырями, которые помогают ей плавать. -
 Рыбы саргассумы замаскированы под дрейфующие водоросли саргассумов .
Рыбы саргассумы замаскированы под дрейфующие водоросли саргассумов .




- Одноклеточные макроводоросли (см. также макроскопические протисты ← )
-
![Одноклеточные пузырьковые водоросли обитают в приливных зонах. Его диаметр может достигать 4 см.[393]](//upload.wikimedia.org/wikipedia/commons/thumb/3/3d/Ventricaria_ventricosa.JPG/248px-Ventricaria_ventricosa.JPG) Одноклеточные пузырьковые водоросли обитают в приливных зонах . Он может иметь диаметр 4 см. [ 393 ]
Одноклеточные пузырьковые водоросли обитают в приливных зонах . Он может иметь диаметр 4 см. [ 393 ] -
 Бокалы одноклеточной русалки представляют собой грибовидные водоросли, вырастающие до 10 см в высоту.
Бокалы одноклеточной русалки представляют собой грибовидные водоросли, вырастающие до 10 см в высоту. -
![Водоросли-киллеры представляют собой одноклеточные организмы, но похожи на папоротники и имеют стебли длиной до 80 см.[394]](//upload.wikimedia.org/wikipedia/commons/thumb/e/e7/CaulerpaTaxifolia.jpg/316px-CaulerpaTaxifolia.jpg) Водоросли-убийцы представляют собой одноклеточные организмы, но внешне напоминают папоротники и имеют стебли длиной до 80 см. [ 394 ]
Водоросли-убийцы представляют собой одноклеточные организмы, но внешне напоминают папоротники и имеют стебли длиной до 80 см. [ 394 ]
![Одноклеточные пузырьковые водоросли обитают в приливных зонах. Его диаметр может достигать 4 см.[393]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/3d/Ventricaria_ventricosa.JPG/248px-Ventricaria_ventricosa.JPG)

![Водоросли-киллеры представляют собой одноклеточные организмы, но похожи на папоротники и имеют стебли длиной до 80 см.[394]](http://upload.wikimedia.org/wikipedia/commons/thumb/e/e7/CaulerpaTaxifolia.jpg/316px-CaulerpaTaxifolia.jpg)
Одноклеточные организмы обычно микроскопические, длиной менее одной десятой миллиметра. Есть исключения. Бокал Русалки , род субтропических зеленых водорослей , является одноклеточным, но удивительно большим и сложным по форме с одним большим ядром, что делает его модельным организмом для изучения клеточной биологии . [ 395 ] Другая одноклеточная водоросль, Caulerpa Taxifolia , имеет вид сосудистого растения с «листьями», аккуратно расположенными на стеблях, как у папоротника. Селективное разведение в аквариумах с целью получения более выносливых штаммов привело к случайному попаданию в Средиземное море, где они превратились в инвазивный вид, известный в просторечии как водоросли-убийцы . [ 396 ]
Происхождение растений
[ редактировать ]
Еще в силурийском периоде часть фитопланктона развилась в красные , бурые и зеленые водоросли . Затем эти водоросли вторглись на сушу и начали развиваться в наземные растения, которые мы знаем сегодня. Позже, в меловом периоде , некоторые из этих наземных растений вернулись в море в виде морских растений, таких как мангровые заросли и морские травы . [ 397 ]
Морские растения можно найти в приливных зонах и на мелководье, например морские травы , такие как угри и черепаховая трава , талассия . Эти растения приспособились к высокой солености океанской среды. Растительная жизнь также может процветать в солоноватой воде эстуариев , где могут расти мангровые заросли , кордграсс или трава пляжная .
-

-

-
![Морские драконы, замаскированные под плавающие водоросли, обитают в зарослях водорослей и на лугах, заросших водорослями.[398]](//upload.wikimedia.org/wikipedia/commons/thumb/2/2a/Leafy_Sea_Dragon_SA.jpg/120px-Leafy_Sea_Dragon_SA.jpg) Морские драконы , замаскированные под плавающие водоросли, живут в лесах из водорослей и на лугах, заросших водорослями. [ 398 ]
Морские драконы , замаскированные под плавающие водоросли, живут в лесах из водорослей и на лугах, заросших водорослями. [ 398 ]


![Морские драконы, замаскированные под плавающие водоросли, обитают в зарослях водорослей и на лугах, заросших водорослями.[398]](http://upload.wikimedia.org/wikipedia/commons/thumb/2/2a/Leafy_Sea_Dragon_SA.jpg/120px-Leafy_Sea_Dragon_SA.jpg)
Общая мировая площадь мангровых лесов оценивалась в 2010 году в 134 257 квадратных километров (51 837 квадратных миль) (на основе спутниковых данных). [ 399 ] [ 400 ] Общую мировую площадь лугов с водорослями определить труднее, но в 2003 году она по консервативным оценкам составляла 177 000 квадратных километров (68 000 квадратных миль). [ 401 ]
Мангровые заросли и морские травы являются важными местами нагула морской жизни, выступая в качестве укрытия и места кормления для личинок и молодых форм более крупных рыб и беспозвоночных. [ 402 ]
Планктон и трофические взаимодействия
[ редактировать ]
Планктон (от греческого — странники ) — разнообразная группа организмов, обитающих в толще воды крупных водоемов, но не умеющих плавать против течения. В результате они блуждают или дрейфуют по течению. [ 403 ] Планктон определяется своей экологической нишей , а не какой-либо филогенетической или таксономической классификацией. Они являются важнейшим источником пищи для многих морских животных, от кормовой рыбы до китов . Планктон можно разделить на растительный компонент и животный компонент.
Фитопланктон
[ редактировать ]Фитопланктон — это растительные компоненты планктонного сообщества («фито» происходит от греческого слова « растение »). Они автотрофны (самопитаются), то есть производят собственную пищу и им не нужно потреблять другие организмы.
Фитопланктон состоит в основном из микроскопических фотосинтезирующих эукариот , обитающих в верхнем освещенном солнцем слое во всех океанах. Им нужен солнечный свет, чтобы они могли фотосинтезировать. Большая часть фитопланктона представляет собой одноклеточные водоросли, но другая часть фитопланктона — это бактерии, а некоторые — протисты . [ 404 ] Группы фитопланктона включают цианобактерии (вверху) , диатомовые водоросли , различные другие виды водорослей (красные, зеленые, бурые и желто-зеленые), динофлагелляты , эвгленоиды , кокколитофориды , криптомонады , хризофиты , хлорофиты , празинофиты и силикофлагелляты . Они составляют основу первичного производства океана , которое управляет пищевой сетью , и на их долю приходится половина текущего мирового первичного производства, больше, чем наземные леса. [ 405 ]


- Фитопланктон
-
 Фитопланктон является основой пищевой цепи океана.
Фитопланктон является основой пищевой цепи океана. -
 Фитопланктон бывает разных форм и размеров.
Фитопланктон бывает разных форм и размеров. -
 Диатомовые водоросли — один из наиболее распространенных видов фитопланктона.
Диатомовые водоросли — один из наиболее распространенных видов фитопланктона. -
 Колониальный фитопланктон
Колониальный фитопланктон




-
 Цианобактерия Prochromococcus обеспечивает большую часть первичной продукции океана.
Цианобактерия Prochromococcus обеспечивает большую часть первичной продукции океана. -
 Зеленые цианобактерии выброшены на скалу в Калифорнии
Зеленые цианобактерии выброшены на скалу в Калифорнии -
 Гиродиний , одна из немногих голых динофлагеллят, лишенных панциря.
Гиродиний , одна из немногих голых динофлагеллят, лишенных панциря. -
 Зоохлореллы (зеленые), живущие внутри инфузории Stichotricha secunda .
Зоохлореллы (зеленые), живущие внутри инфузории Stichotricha secunda .




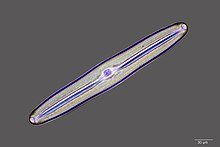
-
 Кокколитофоры названы в честь документального сериала BBC. Голубая планета
Кокколитофоры названы в честь документального сериала BBC. Голубая планета -
 Кокколитофор . Emiliania huxleyi
Кокколитофор . Emiliania huxleyi -
 Цветение водорослей Emiliania huxleyi у южного побережья Англии.
Цветение водорослей Emiliania huxleyi у южного побережья Англии. -
![Guinardia delicatula, диатомовая водоросль, ответственная за цветение водорослей в Северном море и Ла-Манше[407]](//upload.wikimedia.org/wikipedia/commons/thumb/4/43/Fjouenne_sbrmvr012w_20070924163039_small.jpg/260px-Fjouenne_sbrmvr012w_20070924163039_small.jpg) Guinardia delicatula , диатомовая водоросль, ответственная за цветение водорослей в Северном море и Ла-Манше. [ 407 ]
Guinardia delicatula , диатомовая водоросль, ответственная за цветение водорослей в Северном море и Ла-Манше. [ 407 ]



![Guinardia delicatula, диатомовая водоросль, ответственная за цветение водорослей в Северном море и Ла-Манше[407]](http://upload.wikimedia.org/wikipedia/commons/thumb/4/43/Fjouenne_sbrmvr012w_20070924163039_small.jpg/260px-Fjouenne_sbrmvr012w_20070924163039_small.jpg)
Зоопланктон
[ редактировать ]

Зоопланктон — животный компонент планктонного сообщества («зоопарк» происходит от греческого « животное »). Они гетеротрофны (питаются другими), что означает, что они не могут производить собственную пищу и вместо этого должны потреблять в пищу другие растения или животных. В частности, это означает, что они едят фитопланктон.



Зоопланктон обычно крупнее фитопланктона, в основном микроскопический, но некоторые из них можно увидеть невооруженным глазом. Многие простейшие (одноклеточные протисты , питающиеся другой микроскопической жизнью) представляют собой зоопланктон, включая зоофлагелляты , фораминиферы , радиолярии и некоторые динофлагелляты . Другие динофлагелляты миксотрофны и также могут быть отнесены к фитопланктону; Различие между растениями и животными часто нарушается у очень маленьких организмов. Другой зоопланктон включает пелагических книдарий , гребневиков , моллюсков , членистоногих и оболочников , а также планктонных стрелковых червей и щетинковых червей .
Радиолярии — одноклеточные протисты со сложной кремнеземной оболочкой.
Микрозоопланктон: основные пожиратели планктона.
-
 Радиолярии бывают разных форм.
Радиолярии бывают разных форм. -
 Группа планктонных фораминифер.
Группа планктонных фораминифер. -
 Копеподы питаются фитопланктоном. Этот несет яйца.
Копеподы питаются фитопланктоном. Этот несет яйца. -
 Динофлагеллята Protoperidinium выдвигает большую кормовую вуаль для захвата добычи.
Динофлагеллята Protoperidinium выдвигает большую кормовую вуаль для захвата добычи.




Более крупный зоопланктон может охотиться на более мелкий зоопланктон.
Макрозоопланктон
-

-

-

-

-
 Морской амфипод
Морской амфипод -

-
 Пелагический морской огурец
Пелагический морской огурец







| Внешние видео | |
|---|---|
Многие морские животные начинают жизнь как зоопланктон в форме яиц или личинок, прежде чем они превращаются во взрослых особей. Это меропланктоники , то есть планктонные только часть своей жизни.
- Личинки и молодь
-
 Личинка лосося, вылупляющаяся из икры
Личинка лосося, вылупляющаяся из икры -
 морской солнечной рыбы Личинка
морской солнечной рыбы Личинка -
 Молодые планктонные кальмары
Молодые планктонные кальмары -
 Стадия личинки лангуста
Стадия личинки лангуста




Миксотрофный планктон
[ редактировать ]

Динофлагелляты часто миксотрофны или живут в симбиозе с другими организмами.
- Миксопланктон
-
 Тинтинниды инфузории Фавелла
Тинтинниды инфузории Фавелла -
 Эвглена мутабельная — фотосинтезирующий жгутиконосец.
Эвглена мутабельная — фотосинтезирующий жгутиконосец. -
 Noctiluca scintillans , биолюминесцентная динофлагеллята.
Noctiluca scintillans , биолюминесцентная динофлагеллята.



Некоторые динофлагелляты биолюминесцентны . Ночью океанская вода может светиться изнутри и сверкать синим светом из-за этих динофлагеллят. [ 411 ] [ 412 ] Биолюминесцентные динофлагелляты обладают сцинтиллонами , отдельными цитоплазматическими тельцами, содержащими динофлагеллятлюциферазу , основной фермент, участвующий в люминесценции. Люминесценция, иногда называемая фосфоресценцией моря , возникает в виде коротких (0,1 секунды) синих вспышек или искр при стимуляции отдельных сцинтиллонов, обычно в результате механических воздействий, например, от лодки, пловца или прибоя. [ 413 ]
Морская пищевая сеть
[ редактировать ]
По сравнению с наземной средой, морская среда имеет пирамиды биомассы, перевернутые в основании. В частности, биомасса консументов (веслоногих рачков, криля, креветок, кормовых рыб) превышает биомассу первичных продуцентов. Это происходит потому, что основными производителями океана являются крошечные фитопланктоны, которые, как правило, являются r-стратегами , быстро растут и размножаются, поэтому небольшая масса может иметь высокие темпы первичного производства. Напротив, наземные первичные производители, такие как зрелые леса, часто являются K-стратегами , которые растут и размножаются медленно, поэтому для достижения той же скорости первичного производства необходима гораздо большая масса.
Из-за этой инверсии именно зоопланктон составляет большую часть биомассы морских животных . Как первичные потребители , они являются важнейшим связующим звеном между первичными производителями (в основном фитопланктоном) и остальной частью морской пищевой сети ( вторичными потребителями ). [ 414 ]
Если фитопланктон погибает до того, как его съедят, он спускается через эвфотическую зону в составе морского снега и оседает в морские глубины. Таким образом, фитопланктон ежегодно поглощает в океан около 2 миллиардов тонн углекислого газа, в результате чего океан становится поглотителем углекислого газа, в котором содержится около 90% всего поглощенного углерода. [ 415 ]
В 2010 году исследователи обнаружили, что киты переносят питательные вещества из глубин океана обратно на поверхность, используя процесс, который они назвали « китовым насосом» . [ 416 ] Киты кормятся на более глубоких уровнях океана, где водится криль , но регулярно возвращаются на поверхность, чтобы подышать. Там киты испражняются жидкостью, богатой азотом и железом. Вместо того, чтобы тонуть, жидкость остается на поверхности, где ее поглощает фитопланктон . В заливе Мэн китовый насос обеспечивает больше азота, чем реки. [ 417 ]
Другие взаимодействия
[ редактировать ]Биогеохимические циклы
[ редактировать ]
В целом океаны образуют единую морскую систему, где вода – «универсальный растворитель». [ 418 ] – растворяет питательные вещества и вещества, содержащие такие элементы, как кислород, углерод, азот и фосфор. Эти вещества бесконечно цикличны и переработаны, химически соединяются, а затем снова расщепляются, растворяются, а затем осаждаются или испаряются, импортируются и экспортируются обратно на сушу, в атмосферу и на дно океана. , основанные как на биологической активности морских организмов, так и на естественном воздействии солнца, приливов и отливов и движений в земной коре Морские биогеохимические циклы . [ 419 ] [ 420 ]
![Морской углеродный цикл[421]](http://upload.wikimedia.org/wikipedia/commons/thumb/0/03/OceanCarbonCycle.jpg/223px-OceanCarbonCycle.jpg)
-
-
 Морской фосфорный цикл
Морской фосфорный цикл


Отложения и биогенный ил
[ редактировать ]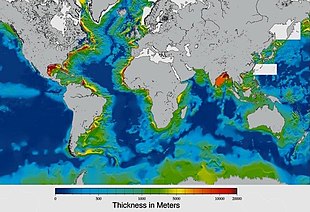
Отложения на дне океана имеют два основных происхождения: терригенное и биогенное. Терригенные отложения составляют около 45% от общего количества морских отложений и возникают в результате эрозии горных пород на суше, переноса реками и земельными стоками, переносимой ветром пыли, вулканов или измельчения ледниками.
Биогенные отложения составляют остальные 55% от общего количества осадков и происходят из скелетных останков морских протистов (одноклеточных планктонных и бентосных организмов). Также могут присутствовать гораздо меньшие количества осажденных минералов и метеорной пыли. В контексте морских отложений «ил» относится не к консистенции осадка, а к его биологическому происхождению. Термин «ил» первоначально использовался Джоном Мюрреем , «отцом современной океанографии», который предложил термин «радиолярийный ил» для обозначения кремнеземных отложений раковин радиолярий, вынесенных на поверхность во время экспедиции «Челленджер» . [ 422 ] Биогенный ил — это пелагический осадок, содержащий не менее 30 процентов скелетных останков морских организмов.
Основные виды биогенных илов
| |||||||
|---|---|---|---|---|---|---|---|
| тип | минерал формы |
протист ответственный |
имя скелет |
описание | |||
| Кремнистый ил | SiO 2 кварц стекло опал chert |
диатомовые водоросли | 
|
кусок | Размеры отдельных диатомей варьируются от 0,002 до 0,2 мм. [ 423 ] | ||
| радиолярии | 
|
скелет | Радиолярии — это простейшие с диаметром обычно от 0,1 до 0,2 мм, которые образуют сложные минеральные скелеты, обычно состоящие из кремнезема. | ||||
| Известковый ил | СаСО 3 кальцит арагонит известняк мел |
фораминиферы | 
|
тест | На сегодняшний день насчитывается около 10 000 видов фораминифер. [ 424 ] обычно размером менее 1 мм. | ||
| кокколитофоры | 
|
кокколит | Кокколитофоры представляют собой сферические клетки диаметром менее 0,1 мм, окруженные известковыми пластинками, называемыми кокколитами. [ 425 ] Кокколиты — важные микрофоссилии . Они являются крупнейшим глобальным источником биогенного карбоната кальция и вносят значительный вклад в глобальный углеродный цикл. [ 426 ] Они являются основной составляющей меловых отложений, таких как белые скалы Дувра . | ||||
-
 Сложный минеральный скелет радиолярии, сделанный из кремнезема.
Сложный минеральный скелет радиолярии, сделанный из кремнезема. -
 Диатомовые водоросли, основные компоненты морского планктона, также имеют кремнеземные скелеты, называемые панцирями .
Диатомовые водоросли, основные компоненты морского планктона, также имеют кремнеземные скелеты, называемые панцирями . -
 Кокколитофоры имеют пластинки или чешуйки, состоящие из карбоната кальция, называемые кокколитами.
Кокколитофоры имеют пластинки или чешуйки, состоящие из карбоната кальция, называемые кокколитами. -
 Кальцифицированная проба планктонного фораминиферана
Кальцифицированная проба планктонного фораминиферана




-
 диатомовой водоросли Микроископаемое , жившее 40 миллионов лет назад.
диатомовой водоросли Микроископаемое , жившее 40 миллионов лет назад. -
 Кизельгур — это мягкая кремнистая состоящая осадочная порода, из микроокаменелостей в форме панцирей ( раковин) одноклеточных диатомей (нажмите, чтобы увеличить).
Кизельгур — это мягкая кремнистая состоящая осадочная порода, из микроокаменелостей в форме панцирей ( раковин) одноклеточных диатомей (нажмите, чтобы увеличить). -
 Иллюстрация Глобигерины ила
Иллюстрация Глобигерины ила -
 Раковины ( раковины ), обычно состоящие из карбоната кальция, из ила фораминифер на глубоком дне океана.
Раковины ( раковины ), обычно состоящие из карбоната кальция, из ила фораминифер на глубоком дне океана.




Земельные взаимодействия
[ редактировать ]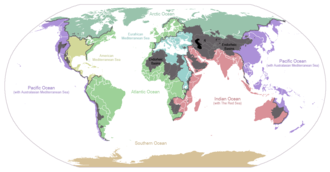
Взаимодействие с сушей влияет на морскую жизнь во многих отношениях. Береговые линии обычно имеют континентальные шельфы , простирающиеся на некоторое расстояние от берега. Они образуют обширные мелководья, освещенные солнцем до морского дна, что обеспечивает фотосинтез и создает среду обитания для лугов с морской травой, коралловых рифов, лесов из водорослей и другой донной жизни . Дальше от берега континентальный шельф спускается в сторону глубокой воды. Ветер , дующий на поверхность океана , или глубокие океанские течения могут привести к тому, что холодные и богатые питательными веществами воды из абиссальных глубин движутся вверх по континентальным склонам . Это может привести к апвеллингам вдоль внешних границ континентальных шельфов, создавая условия для цветения фитопланктона .
Вода, испаряемая солнцем с поверхности океана, может осаждаться на суше и в конечном итоге возвращаться в океан в виде стоков или стоков рек, обогащенная питательными веществами и загрязняющими веществами . Когда реки впадают в устья , пресная вода смешивается с соленой и становится солоноватой . Это создает еще одну среду обитания на мелководье, где мангровые леса и устьевая рыба процветают . В целом жизнь во внутренних озерах может развиваться с большим разнообразием, чем в море, поскольку пресноводные среды обитания сами по себе разнообразны и разделены на части, чего нет в морской среде обитания. Некоторые водные обитатели, такие как лосось и угри , мигрируют между пресноводными и морскими средами обитания. Эти миграции могут привести к обмену патогенами и повлиять на развитие жизни в океане.
Антропогенное воздействие
[ редактировать ]
Деятельность человека влияет на морскую жизнь и морскую среду обитания посредством чрезмерного вылова рыбы , загрязнения , закисления и внедрения инвазивных видов . Это влияет на морские экосистемы и пищевые сети и может привести к пока еще нераспознанным последствиям для биоразнообразия и продолжения морских форм жизни. [ 428 ]
Биоразнообразие и события вымирания
[ редактировать ]

Биоразнообразие является результатом более чем трех миллиардов лет эволюции . Примерно 600 миллионов лет назад вся жизнь состояла из архей , бактерий , простейших и подобных одноклеточных организмов . История биоразнообразия в фанерозое (последние 540 миллионов лет) начинается с быстрого роста во время кембрийского взрыва – периода, в течение которого впервые появились почти все многоклеточных типы организмов . В течение следующих 400 миллионов лет или около того разнообразие беспозвоночных демонстрировало небольшую общую тенденцию, а разнообразие позвоночных демонстрировало общую экспоненциальную тенденцию. [ 430 ]
Однако более 99 процентов всех видов, когда-либо живших на Земле, а их число составляет более пяти миллиардов видов, [ 431 ] считаются вымершими . [ 432 ] [ 433 ] Эти вымирания происходят с неравномерной скоростью. Резкий рост разнообразия был отмечен периодическими массовыми потерями разнообразия, которые классифицируются как события массового вымирания . [ 430 ] События массового вымирания происходят, когда жизнь претерпевает стремительный глобальный упадок. Наибольшее разнообразие и биомасса на Земле приходится на микроорганизмы , которые трудно измерить. Таким образом, зарегистрированные события вымирания основаны на более легко наблюдаемых изменениях разнообразия и численности более крупных многоклеточных организмов , а не на общем разнообразии и изобилии жизни. [ 434 ] Морские окаменелости в основном используются для измерения темпов вымирания из-за их превосходной летописи окаменелостей и стратиграфического диапазона по сравнению с наземными организмами.
Судя по летописи окаменелостей , фоновая скорость вымирания на Земле составляет от двух до пяти таксономических семейств морских животных каждый миллион лет. Великое событие оксигенации было, пожалуй, первым крупным событием вымирания. После кембрийского взрыва пять крупных массовых вымираний значительно превысили фоновые темпы вымирания. [ 435 ] Худшим событием было пермско-триасовое вымирание , произошедшее 251 миллион лет назад. По общему мнению, массовые вымирания «Большой пятерки» фанерозоя (последние 540 миллионов лет) уничтожили более 40% морских родов и, вероятно, более 70% морских видов. [ 436 ] Нынешнее голоценовое вымирание , вызванное деятельностью человека и теперь называемое «шестым вымиранием», может в конечном итоге оказаться еще более разрушительным.
Исследование и изучение морской жизни
[ редактировать ]Исследования и исследования
[ редактировать ]Чтобы проводить исследования и обогащать знания о морской жизни, ученые используют различные методы, чтобы достичь и исследовать глубины океана. Для этой цели используется несколько высокотехнологичных инструментов и транспортных средств. [ 437 ]
- Автономные подводные аппараты (АНПА) — подводные роботы, используемые для исследования океана. АНПА — это независимые роботы, которые могут исследовать мир без экипажа. Их выпускают с корабля и управляют с поверхности. [ 438 ]
- Глубоководные аппараты (DTV) — транспортные средства, буксируемые за исследовательскими судами, предлагающие более простую альтернативу более совершенным подводным аппаратам. Они служат универсальными платформами для использования океанографических инструментов для измерения различных параметров океана, а конкретные модели, такие как DTV BRIDGET, используются для изучения шлейфов гидротермальных жерл, перемещаясь вблизи дна океана. [ 439 ]
- Пилотируемые подводные лодки — пилотируемые подводные аппараты, используемые для исследований, экспериментов и часто используемые армией. [ 437 ]
- Исследовательские суда (НИС) — лодки или корабли, используемые для проведения исследований в течение длительного периода времени. Он способен перевозить разнообразное оборудование для отбора проб и обследования. Исследовательские суда обычно имеют на борту лабораторные помещения, что позволяет исследователям оперативно анализировать материалы, собранные во время круизов.
- Дистанционно управляемые транспортные средства (ROV) – беспилотные транспортные средства. способны достигать большей глубины под водой, чтобы собирать более разнообразную информацию. [ 437 ] [ 440 ]
См. также
[ редактировать ]- Голубая планета - британский документальный телесериал о природе 2001 года - Дэвид Аттенборо
- Голубая планета II - британский документальный телесериал о природе, 2017 г.
- Перепись морской жизни – 10-летняя международная морская биологическая программа
- Колонизация земель
- Экология морских личинок
- Таксономия беспозвоночных - Система классификации животных с упором на беспозвоночных.
Примечания
[ редактировать ]- ^ Это измерение, проведенное судном «Кайко» в марте 1995 года, и оно считается самым точным на сегодняшний день. можно найти в статье Challenger Deep . Более подробную информацию
- ^ Считалось, что миксозоа являются исключением, но теперь считаются сильно модифицированными представителями Cnidaria . Хименес-Гури Э., Филипп Х., Окамура Б., Холланд П.В. (июль 2007 г.). «Будденброкия — книдарийный червь». Наука . 317 (5834): 116–8. Бибкод : 2007Sci...317..116J . дои : 10.1126/science.1142024 . ПМИД 17615357 . S2CID 5170702 .
Ссылки
[ редактировать ]- ^ Jump up to: а б Кавиччиоли Р., Риппл В.Дж., Тиммис К.Н., Азам Ф., Баккен Л.Р., Бэйлис М. и др. (сентябрь 2019 г.). «Предупреждение ученых человечеству: микроорганизмы и изменение климата» . Обзоры природы. Микробиология . 17 (9): 569–586. дои : 10.1038/s41579-019-0222-5 . ПМЦ 7136171 . ПМИД 31213707 .
 Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б с «Национальное управление океанических и атмосферных исследований – Океан» . НОАА . Проверено 20 февраля 2019 г.
- ^ «Маленькая рыбка может быть предком почти всех ныне живущих позвоночных» . Живая наука . 11 июня 2014 г.
- ^ Дрогин Б. (2 августа 2009 г.). «Картирование океана видов» . Лос-Анджелес Таймс . Проверено 18 августа 2009 г.
- ^ Буше, Филипп; Декок, Вим; Лонневиль, Бритт; Ванхоорн, Барт; Вандепитт, Лин (июнь 2023 г.). «Открытие морского биоразнообразия: метрики описаний новых видов» . Границы морской науки . 10 (3389). doi : 10.3389/fmars.2023.929989 — через ResearchGate .
- ^ Пол, GS (2010). «Эволюция динозавров и их мира». Принстонский полевой справочник по динозаврам . Принстон: Издательство Принстонского университета . п. 19 . ISBN 978-0-691-13720-9 .
- ^ Бортолотти, Дэн (2008). в мире Дикий синий: естественная история самого большого животного Нью-Йорк: Книги Томаса Данна. ISBN 978-0-312-38387-9 . OCLC 213451450 .
- ^ Jump up to: а б с Бар-Он Ю.М., Филлипс Р., Майло Р. (июнь 2018 г.). «Распределение биомассы на Земле» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (25): 6506–6511. Бибкод : 2018PNAS..115.6506B . дои : 10.1073/pnas.1711842115 . ПМК 6016768 . ПМИД 29784790 .
- ^ «Перепись морской жизни» . Смитсоновский институт . 30 апреля 2018 года . Проверено 29 октября 2020 г.
- ^ Сяо-Фэн П. (2014). «Глава 5». Вода: молекулярная структура и свойства . Всемирная научная. стр. 390–461. ISBN 9789814440448 .
- ^ Гринвуд Н.Н. , Эрншоу А. (1997). Химия элементов (2-е изд.). Баттерворт-Хайнеманн . п. 620. ИСБН 978-0-08-037941-8 .
- ^ «Вода – универсальный растворитель» . Геологическая служба США . Архивировано из оригинала 9 июля 2017 года . Проверено 27 июня 2017 г.
- ^ Рис Дж.Б. (31 октября 2013 г.). Кэмпбелл Биология (10-е изд.). Пирсон . п. 48. ИСБН 9780321775658 .
- ^ Рис Дж.Б. (31 октября 2013 г.). Кэмпбелл Биология (10-е изд.). Пирсон . п. 44. ИСБН 9780321775658 .
- ^ Коллинз Дж. К. (1991). Матрица жизни: взгляд на природные молекулы с точки зрения воды в окружающей среде . Молекулярные презентации. ISBN 978-0-9629719-0-7 .
- ^ «Дистанционно управляемый автомобиль класса 7000 м KAIKO 7000 » . Японское агентство морских наук и технологий о Земле (JAMSTEC). Архивировано из оригинала 10 апреля 2020 года . Проверено 7 июня 2008 г.
- ^ «Сколько океанов?» . НОАА . 9 апреля 2020 г.
- ^ Шаретт М.А., Смит У.Х. (июнь 2010 г.). «Объем земного океана» . Океанография . 23 (2): 112–14. дои : 10.5670/oceanog.2010.51 . hdl : 1912/3862 .
- ^ Даксбери AC, Дженедез C (7 мая 2021 г.). «Сфера глубин океана – гидрология» . Британская энциклопедия . Проверено 12 апреля 2015 г.
- ^ «Третий камень от Солнца – беспокойная Земля» . Космос НАСА . Проверено 12 апреля 2015 г.
- ^ Перлман Х (17 марта 2014 г.). «Вода мира» . Школа водных наук Геологической службы США . Проверено 12 апреля 2015 г.
- ^ Кенниш К.Дж. (2001). Практический справочник по морским наукам . Серия морских наук (3-е изд.). ЦРК Пресс. п. 35. ISBN 978-0-8493-2391-1 .
- ^ «Почему океан соленый?» .
- ^ Маллен Л. (11 июня 2002 г.). «Соль Ранней Земли» . Журнал НАСА по астробиологии. Архивировано из оригинала 30 июня 2007 года . Проверено 14 марта 2007 г.
- ^ Моррис Р.М. «Океанические процессы» . Журнал НАСА по астробиологии. Архивировано из оригинала 15 апреля 2009 года . Проверено 14 марта 2007 г.
- ^ Скотт М. (24 апреля 2006 г.). «Большое тепловое ведро Земли» . Земная обсерватория НАСА . Проверено 14 марта 2007 г.
- ^ Образец S (21 июня 2005 г.). «Температура поверхности моря» . НАСА. Архивировано из оригинала 6 апреля 2013 года . Проверено 21 апреля 2007 г.
- ^ «Объемы Мирового океана по данным ETOPO1» . НОАА . Архивировано из оригинала 11 марта 2015 года . Проверено 20 февраля 2019 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ «Планета «Земля»: Нам следовало назвать ее «Море» » . Цитата Инвертигатор . 25 января 2017 г.
- ^ «Открытие планеты Океан» . ПНАСА Наука . 14 марта 2002 г. Архивировано из оригинала 8 октября 2022 г. Проверено 19 февраля 2019 г.
- ^ Дайчес П., Браун Д. (12 мая 2015 г.). «Исследования НАСА показывают, что загадочный темный материал Европы может быть морской солью» . НАСА . Проверено 12 мая 2015 г.
- ^ Адаму З. (1 октября 2012 г.). «Вода у поверхности спутника Юпитера лишь временная» . CNN Световые годы . Архивировано из оригинала 5 октября 2012 года . Проверено 24 апреля 2019 г.
- ^ Тритт CS (2002). «Возможность жизни на Европе» . Инженерная школа Милуоки. Архивировано из оригинала 9 июня 2007 года . Проверено 10 августа 2007 г.
- ^ Шульце-Макух Д., Ирвин Л.Н. (2001). «Альтернативные источники энергии могут поддержать жизнь на Европе» (PDF) . Кафедры геологических и биологических наук Техасского университета в Эль-Пасо . Архивировано из оригинала (PDF) 3 июля 2006 года . Проверено 21 декабря 2007 г.
- ^ Фридман Л. (14 декабря 2005 г.). «Проекты: Кампания миссии Европы» . Планетарное общество. Архивировано из оригинала 11 августа 2011 года . Проверено 8 августа 2011 г.
- ^ «Океан внутри Энцелада может быть источником гидротермальной активности» . Пресс-релиз НАСА . 11 марта 2015 г.
- ^ «Возраст Земли» . Геологическая служба США . 9 июля 2007 года . Проверено 31 мая 2015 г.
- ^ Далримпл ГБ (январь 2001 г.). «Возраст Земли в двадцатом веке: проблема (в основном) решена». Специальные публикации . 190 (1). Лондон: Геологическое общество: 205–21. Бибкод : 2001GSLSP.190..205D . дои : 10.1144/ГСЛ.СП.2001.190.01.14 . S2CID 130092094 .
- ^ Манеш Ж., Аллегре С.Дж. , Дюпре Б., Хамелен Б. (май 1980 г.). «Изотопное исследование свинца основных-ультраосновных слоистых комплексов: предположения о возрасте Земли и характеристиках примитивной мантии». Письма о Земле и планетологии . 47 (3): 370–382. Бибкод : 1980E&PSL..47..370M . дои : 10.1016/0012-821X(80)90024-2 . ISSN 0012-821X .
- ^ Шопф Й.В. , Кудрявцев А.Б., Чая А.Д., Трипати А.Б. (5 октября 2007 г.). «Свидетельства архейской жизни: строматолиты и микроокаменелости». Докембрийские исследования . 158 (3–4): 141–155. Бибкод : 2007PreR..158..141S . doi : 10.1016/j.precamres.2007.04.009 . ISSN 0301-9268 .
- ^ Рэйвен П.Х., Джонсон ГБ (2002). Биология (6-е изд.). Бостон: МакГроу-Хилл. п. 68. ИСБН 978-0-07-112261-0 .
- ^ Баумгартнер Р.Дж., Ван Кранендонк М.Дж., Уэйси Д., Фиорентини М.Л., Сондерс М., Карузо С. и др. (2019). «Нанопористый пирит и органическое вещество в строматолитах возрастом 3,5 миллиарда лет свидетельствуют о первозданной жизни» (PDF) . Геология . 47 (11): 1039–1043. Бибкод : 2019Geo....47.1039B . дои : 10.1130/G46365.1 . S2CID 204258554 .
- ^ «Самые ранние признаки жизни: ученые находят остатки микробов в древних камнях» . Физика.орг . 26 сентября 2019 г.
- ^ Отомо И., Какегава Т., Исида А., Нагасе Т., Розинг М.Т. (январь 2014 г.). «Доказательства наличия биогенного графита в метаосадочных породах раннего архейского периода Иисуса». Природа Геонауки . 7 (1): 25–28. Бибкод : 2014NatGe...7... 25O дои : 10.1038/ngeo2025 . ISSN 1752-0894 .
- ^ Jump up to: а б Боренштейн С. (19 октября 2015 г.). «Намеки на жизнь на ранней Земле, которая считалась пустынной» . Ассошиэйтед Пресс . Проверено 9 октября 2018 г.
- ^ Белл Э.А., Бёнке П., Харрисон Т.М., Мао В.Л. (ноябрь 2015 г.). «Потенциально биогенный углерод сохранился в цирконе возрастом 4,1 миллиарда лет» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (47): 14518–21. Бибкод : 2015PNAS..11214518B . дои : 10.1073/pnas.1517557112 . ПМЦ 4664351 . ПМИД 26483481 .
- ^ Пенни Д., Пул А. (декабрь 1999 г.). «Природа последнего всеобщего общего предка». Текущее мнение в области генетики и развития . 9 (6): 672–7. дои : 10.1016/S0959-437X(99)00020-9 . ПМИД 10607605 .
- ^ Теобальд Д.Л. (май 2010 г.). «Формальная проверка теории универсального общего происхождения». Природа . 465 (7295): 219–22. Бибкод : 2010Natur.465..219T . дои : 10.1038/nature09014 . ПМИД 20463738 . S2CID 4422345 .
- ^ Дулитл В.Ф. (февраль 2000 г.). «Искоренение древа жизни» (PDF) . Научный американец . 282 (2): 90–5. Бибкод : 2000SciAm.282b..90D . doi : 10.1038/scientificamerican0200-90 . ПМИД 10710791 . Архивировано из оригинала (PDF) 7 сентября 2006 года . Проверено 5 апреля 2015 г.
- ^ Перето Дж. (март 2005 г.). «Споры о происхождении жизни» (PDF) . Международная микробиология . 8 (1): 23–31. ПМИД 15906258 . Архивировано из оригинала (PDF) 24 августа 2015 года.
- ^ Джойс Г.Ф. (июль 2002 г.). «Древность эволюции на основе РНК». Природа . 418 (6894): 214–21. Бибкод : 2002Natur.418..214J . дои : 10.1038/418214a . ПМИД 12110897 . S2CID 4331004 .
- ^ Треворс Дж.Т., Псеннер Р. (декабрь 2001 г.). «От самосборки жизни до современных бактерий: возможная роль наноклеток» . Обзоры микробиологии FEMS . 25 (5): 573–82. дои : 10.1111/j.1574-6976.2001.tb00592.x . ПМИД 11742692 .
- ^ Уэйд Н. (25 июля 2016 г.). «Знакомьтесь, Лука, предок всего живого» . Нью-Йорк Таймс . Проверено 25 июля 2016 г.
- ^ Баптест Э., Уолш Д.А. (июнь 2005 г.). «Звучит ли «Кольцо жизни» правдиво?». Тенденции в микробиологии . 13 (6): 256–61. дои : 10.1016/j.tim.2005.03.012 . ПМИД 15936656 .
- ^ Дарвин С. (1859 г.). «О происхождении видов» . Лондон: Джон Мюррей.
- ^ Дулитл В.Ф., Баптест Э. (февраль 2007 г.). «Плюрализм моделей и гипотеза Древа жизни» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (7): 2043–9. Бибкод : 2007PNAS..104.2043D . дои : 10.1073/pnas.0610699104 . ЧВК 1892968 . ПМИД 17261804 .
- ^ Кунин В., Голдовский Л., Дарзентас Н., Узунис К.А. (июль 2005 г.). «Сеть жизни: реконструкция микробной филогенетической сети» . Геномные исследования . 15 (7): 954–9. дои : 10.1101/гр.3666505 . ПМЦ 1172039 . ПМИД 15965028 .
- ^ Яблонски Д. (июнь 1999 г.). «Будущее летописи окаменелостей». Наука . 284 (5423): 2114–6. дои : 10.1126/science.284.5423.2114 . ПМИД 10381868 . S2CID 43388925 .
- ^ Сиссарелли Ф.Д., Доркс Т., фон Меринг С., Криви С.Дж., Снел Б., Борк П. (март 2006 г.). «К автоматической реконструкции высокоразрешенного древа жизни». Наука . 311 (5765): 1283–7. Бибкод : 2006Sci...311.1283C . CiteSeerX 10.1.1.381.9514 . дои : 10.1126/science.1123061 . ПМИД 16513982 . S2CID 1615592 .
- ^ Мейсон С.Ф. (6 сентября 1984 г.). «Истоки биомолекулярной рукости». Природа . 311 (5981): 19–23. Бибкод : 1984Natur.311...19M . дои : 10.1038/311019a0 . ПМИД 6472461 . S2CID 103653 .
- ^ Вольф Ю.И., Рогозин И.Б., Гришин Н.В., Кунин Е.В. (сентябрь 2002 г.). «Геномные деревья и древо жизни». Тенденции в генетике . 18 (9): 472–9. дои : 10.1016/S0168-9525(02)02744-0 . ПМИД 12175808 .
- ^ Варки А. , Альтейде Т.К. (декабрь 2005 г.). «Сравнение геномов человека и шимпанзе: поиск иголок в стоге сена» . Геномные исследования . 15 (12): 1746–58. дои : 10.1101/гр.3737405 . ПМИД 16339373 .
- ^ Jump up to: а б Кавалер-Смит Т. (июнь 2006 г.). «Эволюция клеток и история Земли: застой и революция» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 361 (1470): 969–1006. дои : 10.1098/rstb.2006.1842 . ПМЦ 1578732 . ПМИД 16754610 .
- ^ Шопф JW (июнь 2006 г.). «Ископаемые свидетельства архейской жизни» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 361 (1470): 869–85. дои : 10.1098/rstb.2006.1834 . ПМЦ 1578735 . ПМИД 16754604 .
- Альтерманн В., Казмерчак Дж. (ноябрь 2003 г.). «Архейские микроокаменелости: переоценка ранней жизни на Земле» . Исследования в области микробиологии . 154 (9): 611–7. дои : 10.1016/j.resmic.2003.08.006 . ПМИД 14596897 .
- ^ Jump up to: а б Шопф JW (июль 1994 г.). «Разные темпы, разные судьбы: темп и способ эволюции менялись от докембрия к фанерозою» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (15): 6735–42. Бибкод : 1994PNAS...91.6735S . дои : 10.1073/pnas.91.15.6735 . ПМЦ 44277 . ПМИД 8041691 .
- ^ Jump up to: а б Пул А.М., Пенни Д. (январь 2007 г.). «Оценка гипотез происхождения эукариот». Биоэссе . 29 (1): 74–84. doi : 10.1002/bies.20516 . ПМИД 17187354 .
- ^ Jump up to: а б Дьялл С.Д., Браун М.Т., Джонсон П.Дж. (апрель 2004 г.). «Древние вторжения: от эндосимбионтов к органеллам». Наука . 304 (5668): 253–7. Бибкод : 2004Sci...304..253D . дои : 10.1126/science.1094884 . ПМИД 15073369 . S2CID 19424594 .
- ^ Мартин В. (октябрь 2005 г.). «Недостающее звено между гидрогеносомами и митохондриями». Тенденции в микробиологии . 13 (10): 457–9. дои : 10.1016/j.tim.2005.08.005 . ПМИД 16109488 .
- ^ Ланг Б.Ф., Грей М.В., Бургер Дж. (декабрь 1999 г.). «Эволюция митохондриального генома и происхождение эукариот». Ежегодный обзор генетики . 33 : 351–97. дои : 10.1146/annurev.genet.33.1.351 . ПМИД 10690412 .
- Макфадден Дж.И. (декабрь 1999 г.). «Эндосимбиоз и эволюция растительной клетки». Современное мнение в области биологии растений . 2 (6): 513–9. дои : 10.1016/S1369-5266(99)00025-4 . ПМИД 10607659 .
- ^ Делонг Э.Ф. , Пейс Н.Р. (август 2001 г.). «Экологическое разнообразие бактерий и архей». Систематическая биология . 50 (4): 470–8. CiteSeerX 10.1.1.321.8828 . дои : 10.1080/106351501750435040 . ПМИД 12116647 .
- ^ Кайзер Д. (декабрь 2001 г.). «Построение многоклеточного организма». Ежегодный обзор генетики . 35 : 103–23. дои : 10.1146/annurev.genet.35.102401.090145 . ПМИД 11700279 . S2CID 18276422 .
- ^ Циммер С (7 января 2016 г.). «Генетический переворот помог организмам перейти от одной клетки к множеству» . Нью-Йорк Таймс . Проверено 7 января 2016 г.
- ^ Валентайн Дж.В. , Яблонски Д., Эрвин Д.Х. (февраль 1999 г.). «Ископаемые, молекулы и эмбрионы: новые взгляды на кембрийский взрыв». Разработка . 126 (5): 851–9. дои : 10.1242/dev.126.5.851 . ПМИД 9927587 .
- ^ Оно С. (январь 1997 г.). «Причина, а также последствия кембрийского взрыва в эволюции животных». Журнал молекулярной эволюции . 44 (Приложение 1): С23-7. Бибкод : 1997JMolE..44S..23O . дои : 10.1007/PL00000055 . ПМИД 9071008 . S2CID 21879320 .
- Валентин Дж.В., Яблонски Д. (2003). «Морфологическая макроэволюция и развитие: палеонтологическая перспектива» . Международный журнал биологии развития . 47 (7–8): 517–22. ПМИД 14756327 . Проверено 30 декабря 2014 г.
- ^ Веллман Ч., Остерлофф П.Л., Мохиуддин У (сентябрь 2003 г.). «Фрагменты древнейших наземных растений» (PDF) . Природа . 425 (6955): 282–5. Бибкод : 2003Natur.425..282W . дои : 10.1038/nature01884 . ПМИД 13679913 . S2CID 4383813 .
- ^ Бартон Н. (2007). Эволюция . ЦШЛ Пресс. стр. 273–274. ISBN 9780199226320 . Проверено 30 сентября 2012 г.
- ^ Уотерс ER (декабрь 2003 г.). «Молекулярная адаптация и происхождение наземных растений». Молекулярная филогенетика и эволюция . 29 (3): 456–63. дои : 10.1016/j.ympev.2003.07.018 . ПМИД 14615186 .
- ^ Мэйхью П.Дж. (август 2007 г.). «Почему существует так много видов насекомых? Перспективы окаменелостей и филогении». Биологические обзоры Кембриджского философского общества . 82 (3): 425–54. дои : 10.1111/j.1469-185X.2007.00018.x . ПМИД 17624962 . S2CID 9356614 .
- ^ Кэрролл Р.Л. (май 2007 г.). «Палеозойское происхождение саламандр, лягушек и червяг» . Зоологический журнал Линнеевского общества . 150 (Приложение с1): 1–140. дои : 10.1111/j.1096-3642.2007.00246.x . ISSN 1096-3642 .
- ^ Уибл-младший, Ружье Г.В., Новачек М.Дж., Ашер Р.Дж. (июнь 2007 г.). «Меловые эвтерии и лавразийское происхождение плацентарных млекопитающих вблизи границы K/T». Природа . 447 (7147): 1003–6. Бибкод : 2007Natur.447.1003W . дои : 10.1038/nature05854 . ПМИД 17581585 . S2CID 4334424 .
- ^ Витмер Л.М. (июль 2011 г.). «Палеонтология: икона, сбитая с насеста». Природа . 475 (7357): 458–9. дои : 10.1038/475458a . ПМИД 21796198 . S2CID 205066360 .
- ^ Шлосс П.Д., Хандельсман Дж. (декабрь 2004 г.). «Состояние микробной переписи» . Обзоры микробиологии и молекулярной биологии . 68 (4): 686–91. дои : 10.1128/MMBR.68.4.686-691.2004 . ПМК 539005 . ПМИД 15590780 .
- ^ Миллер Г.Т., Спулман С. (январь 2012 г.). «Глава 4.1: Что такое биоразнообразие и почему оно важно?» . Экологическая наука . Cengage Обучение. п. 62. ИСБН 978-1-133-70787-5 .
- ^ Мора С, Титтенсор ДП, Адл С, Симпсон А.Г., Червь Б (август 2011 г.). «Сколько видов существует на Земле и в океане?» . ПЛОС Биология . 9 (8): e1001127. дои : 10.1371/journal.pbio.1001127 . ПМК 3160336 . ПМИД 21886479 .
- ^ Мэдиган М., Мартинко Дж., ред. (2006). Брок Биология микроорганизмов (13-е изд.). Пирсон Образование. п. 1096. ИСБН 978-0-321-73551-5 .
- ^ Рыбицкий Е.П. (1990). «Классификация организмов на пороге жизни, или Проблемы вирусной систематики». Южноафриканский научный журнал . 86 : 182–6. ISSN 0038-2353 .
- ^ Львофф А. (октябрь 1957 г.). «Понятие вируса» . Журнал общей микробиологии . 17 (2): 239–53. дои : 10.1099/00221287-17-2-239 . ПМИД 13481308 .
- ^ «Данные ВОЗ о смертности за 2002 год» . Архивировано из оригинала 19 августа 2006 года . Проверено 20 января 2007 г.
- ^ «Функции микробиома глобального океана являются ключом к пониманию изменений окружающей среды» . www.sciencedaily.com . Университет Джорджии. 10 декабря 2015 года . Проверено 11 декабря 2015 г.
- ^ Саттл, Калифорния (сентябрь 2005 г.). «Вирусы в море». Природа . 437 (7057): 356–61. Бибкод : 2005Natur.437..356S . дои : 10.1038/nature04160 . ПМИД 16163346 . S2CID 4370363 .
- ^ Jump up to: а б Шорс Т (2017). Понимание вирусов (3-е изд.). Издательство Джонс и Бартлетт. ISBN 978-1-284-02592-7 .
- ^ Jump up to: а б с Саттл, Калифорния (октябрь 2007 г.). «Морские вирусы — основные игроки глобальной экосистемы». Обзоры природы. Микробиология . 5 (10): 801–12. дои : 10.1038/nrmicro1750 . ПМИД 17853907 . S2CID 4658457 .
- ^ Моррисон Дж. (11 января 2016 г.). «Живые бактерии движутся в воздушных потоках Земли» . Смитсоновский журнал .
- ^ Роббинс Дж. (13 апреля 2018 г.). «Триллионы и триллионы вирусов падают с неба каждый день» . Нью-Йорк Таймс . Проверено 14 апреля 2018 г.
- ^ Рече И., Д'Орта Г., Младенов Н., Вингет Д.М., Саттл, Калифорния (апрель 2018 г.). «Скорость осаждения вирусов и бактерий над пограничным слоем атмосферы» . Журнал ISME . 12 (4): 1154–1162. дои : 10.1038/s41396-017-0042-4 . ПМК 5864199 . ПМИД 29379178 .
- ^ Персонал (2014). «Биосфера» . Аспенский институт глобальных изменений . Архивировано из оригинала 2 сентября 2010 года . Проверено 10 ноября 2014 г.
- ^ Jump up to: а б с Чой CQ (17 марта 2013 г.). «Микробы процветают в самом глубоком месте на Земле» . ЖиваяНаука . Проверено 17 марта 2013 г.
- ^ Глуд Р.Н., Венцхёфер Ф., Мидделбо М., Огури К., Турневич Р., Кэнфилд Д.Э., Китазато Х (17 марта 2013 г.). «Высокие темпы микробного круговорота углерода в отложениях самой глубокой океанической впадины на Земле». Природа Геонауки . 6 (4): 284–288. Бибкод : 2013NatGe...6..284G . дои : 10.1038/ngeo1773 .
- ^ Оськин Б. (14 марта 2013 г.). «Инопланетяне: жизнь процветает на дне океана» . ЖиваяНаука . Проверено 17 марта 2013 г.
- ^ Морель Р. (15 декабря 2014 г.). «Анализ микробов, обнаруженных с помощью самой глубоководной морской буровой установки» . Новости Би-би-си . Проверено 15 декабря 2014 г.
- ^ Такай К., Накамура К., Токи Т., Цуногай У., Миядзаки М., Миядзаки Дж. и др. (август 2008 г.). «Пролиферация клеток при 122 градусах Цельсия и производство изотопно-тяжелого CH4 гипертермофильным метаногеном при культивировании под высоким давлением» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (31): 10949–54. Бибкод : 2008PNAS..10510949T . дои : 10.1073/pnas.0712334105 . ПМК 2490668 . ПМИД 18664583 .
- ^ Фокс Д. (август 2014 г.). «Озера подо льдом: тайный сад Антарктиды» . Природа . 512 (7514): 244–6. Бибкод : 2014Natur.512..244F . дои : 10.1038/512244a . ПМИД 25143097 .
- ^ Мак Э. (20 августа 2014 г.). «Жизнь подтверждена подо льдом Антарктики; следующий космос?» . Форбс . Проверено 21 августа 2014 г.
- ^ Виммер Э., Мюллер С., Тумпей Т.М., Таубенбергер Дж.К. (декабрь 2009 г.). «Синтетические вирусы: новая возможность понять и предотвратить вирусные заболевания» . Природная биотехнология . 27 (12): 1163–72. дои : 10.1038/nbt.1593 . ПМК 2819212 . ПМИД 20010599 .
- ^ Кунин Е.В., Сенкевич Т.Г., Доля В.В. (сентябрь 2006 г.). «Древний мир вирусов и эволюция клеток» . Биология Директ . 1:29 . дои : 10.1186/1745-6150-1-29 . ПМК 1594570 . ПМИД 16984643 .
- ^ Топли В.В., Уилсон Г.С., Коллиер Л.Х., Балоуз А., Сассман М. (1998). Махи Б.В., Кольер Л. (ред.). Микробиология и микробные инфекции Топли и Уилсона . Том. 1 (9-е изд.). Лондон: Арнольд. стр. 33–37. ISBN 978-0-340-66316-5 .
- ^ Айер Л.М., Баладжи С., Кунин Е.В., Аравинд Л. (апрель 2006 г.). «Эволюционная геномика ядерно-цитоплазматических крупных ДНК-вирусов» . Вирусные исследования . 117 (1): 156–84. doi : 10.1016/j.virusres.2006.01.009 . ПМИД 16494962 .
- ^ Санхуан Р., Небот М.Р., Кирико Н., Мански Л.М., Белшоу Р. (октябрь 2010 г.). «Темпы вирусных мутаций» . Журнал вирусологии . 84 (19): 9733–48. дои : 10.1128/JVI.00694-10 . ПМЦ 2937809 . ПМИД 20660197 .
- ^ Махи У.Дж., Ван Регенмортель М.Х., ред. (2009). Настольная энциклопедия общей вирусологии . Оксфорд: Академическая пресса. п. 28. ISBN 978-0-12-375146-1 .
- ^ Jump up to: а б Канчая С., Фурнус Г., Чибани-Шеннуфи С., Диллманн М.Л., Брюссов Х. (август 2003 г.). «Фаг как агент латерального переноса генов». Современное мнение в микробиологии . 6 (4): 417–24. дои : 10.1016/S1369-5274(03)00086-9 . ПМИД 12941415 .
- ^ Jump up to: а б Кунин Е.В., Старокадомский П. (октябрь 2016 г.). «Живы ли вирусы? Парадигма репликатора проливает решающий свет на старый, но ошибочный вопрос» . Исследования по истории и философии биологических и биомедицинских наук . 59 : 125–34. дои : 10.1016/j.shpsc.2016.02.016 . ПМК 5406846 . ПМИД 26965225 .
- ^ Рыбицкий Е.П. (1990). «Классификация организмов на пороге жизни, или Проблемы вирусной систематики». Южноафриканский научный журнал . 86 : 182–186.
- ^ Jump up to: а б Манн, Нью-Хэмпшир (май 2005 г.). «Третья эра фага» . ПЛОС Биология . 3 (5): е182. дои : 10.1371/journal.pbio.0030182 . ПМК 1110918 . ПМИД 15884981 .
- ^ Уоммак К.Е., Колвелл Р.Р. (март 2000 г.). «Вириопланктон: вирусы в водных экосистемах» . Обзоры микробиологии и молекулярной биологии . 64 (1): 69–114. дои : 10.1128/ММБР.64.1.69-114.2000 . ПМК 98987 . ПМИД 10704475 .
- ^ Jump up to: а б Саттл, Калифорния (сентябрь 2005 г.). «Вирусы в море». Природа . 437 (7057): 356–61. Бибкод : 2005Natur.437..356S . дои : 10.1038/nature04160 . ПМИД 16163346 . S2CID 4370363 .
- ^ Берг О., Борсхайм К.Ю., Братбак Г., Хелдал М. (август 1989 г.). «Высокое количество вирусов обнаружено в водной среде». Природа . 340 (6233): 467–8. Бибкод : 1989Natur.340..467B . дои : 10.1038/340467a0 . ПМИД 2755508 . S2CID 4271861 .
- ^ Вигингтон Ч., Зондереггер Д., Бруссаард К.П., Бьюкен А., Финке Дж.Ф., Фурман Дж.А. и др. (январь 2016 г.). «Повторное исследование взаимосвязи между численностью морских вирусов и микробных клеток» (PDF) . Природная микробиология . 1 (3): 15024. doi : 10.1038/nmicrobiol.2015.24 . ПМИД 27572161 . S2CID 52829633 .
- ^ Крупович М., Бэмфорд Д.Х. (июль 2007 г.). «Предположительные профаги, родственные литическому бесхвостому морскому фагу дцДНК PM2, широко распространены в геномах водных бактерий» . БМК Геномика . 8 : 236. дои : 10.1186/1471-2164-8-236 . ЧВК 1950889 . ПМИД 17634101 .
- ^ Сюэ Х, Сюй Ю, Буше Ю, Полц МФ (январь 2012 г.). «Высокая частота появления нового нитчатого фага VCY φ в окружающей популяции холерного вибриона» . Прикладная и экологическая микробиология . 78 (1): 28–33. Бибкод : 2012ApEnM..78...28X . дои : 10.1128/АЕМ.06297-11 . ПМЦ 3255608 . ПМИД 22020507 .
- ^ Ру С., Крупович М., Пуле А., Деброа Д., Эно Ф. (2012). «Эволюция и разнообразие вирусного семейства Microviridae посредством коллекции из 81 нового полного генома, собранного из считываний вирома» . ПЛОС ОДИН . 7 (7): е40418. Бибкод : 2012PLoSO...740418R . дои : 10.1371/journal.pone.0040418 . ПМЦ 3394797 . ПМИД 22808158 .
- ^ «Вредное цветение водорослей: Красный прилив: Домой | CDC HSB» . www.cdc.gov . Проверено 19 декабря 2014 г.
- ^ Лоуренс С.М., Менон С., Эйлерс Б.Дж., Ботнер Б., Хаят Р., Дуглас Т., Янг М.Дж. (май 2009 г.). «Структурно-функциональные исследования архейных вирусов» . Журнал биологической химии . 284 (19): 12599–603. дои : 10.1074/jbc.R800078200 . ПМК 2675988 . ПМИД 19158076 .
- ^ Прангишвили Д., Фортер П., Гаррет Р.А. (ноябрь 2006 г.). «Вирусы архей: объединяющий взгляд». Обзоры природы. Микробиология . 4 (11): 837–48. дои : 10.1038/nrmicro1527 . ПМИД 17041631 . S2CID 9915859 .
- ^ Прангишвили Д., Гаррет Р.А. (апрель 2004 г.). «Исключительно разнообразные морфотипы и геномы кренархейных гипертермофильных вирусов» (PDF) . Труды Биохимического общества . 32 (Часть 2): 204–8. дои : 10.1042/BST0320204 . ПМИД 15046572 . S2CID 20018642 .
- ^ Фортер П., Филипп Х (июнь 1999 г.). «Последний универсальный общий предок (LUCA), простой или сложный?». Биологический вестник . 196 (3): 373–5, обсуждение 375–7. дои : 10.2307/1542973 . JSTOR 1542973 . ПМИД 11536914 .
- ^ Фредриксон Дж.К., Захара Дж.М., Балквилл Д.Л., Кеннеди Д., Ли С.М., Костандаритес Х.М. и др. (июль 2004 г.). «Геомикробиология вадозных отложений, сильно загрязненных ядерными отходами, на полигоне в Хэнфорде, штат Вашингтон» . Прикладная и экологическая микробиология . 70 (7): 4230–41. Бибкод : 2004ApEnM..70.4230F . дои : 10.1128/АЕМ.70.7.4230-4241.2004 . ПМК 444790 . ПМИД 15240306 .
- ^ Вёзе Ч.Р., Кандлер О., Уилис М.Л. (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–9. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМЦ 54159 . ПМИД 2112744 .
- ^ Делонг Э.Ф., Пейс Н.Р. (август 2001 г.). «Экологическое разнообразие бактерий и архей». Систематическая биология . 50 (4): 470–8. CiteSeerX 10.1.1.321.8828 . дои : 10.1080/106351501750435040 . ПМИД 12116647 .
- ^ Браун-младший, Дулиттл В.Ф. (декабрь 1997 г.). «Археи и переход от прокариот к эукариотам» . Обзоры микробиологии и молекулярной биологии . 61 (4): 456–502. doi : 10.1128/mmbr.61.4.456-502.1997 . ПМК 232621 . ПМИД 9409149 .
- ^ Ланг Б.Ф., Грей М.В., Бургер Дж. (1999). «Эволюция митохондриального генома и происхождение эукариот». Ежегодный обзор генетики . 33 : 351–97. дои : 10.1146/annurev.genet.33.1.351 . ПМИД 10690412 .
- ^ Макфадден Дж.И. (декабрь 1999 г.). «Эндосимбиоз и эволюция растительной клетки». Современное мнение в области биологии растений . 2 (6): 513–9. дои : 10.1016/S1369-5266(99)00025-4 . ПМИД 10607659 .
- ^ Килинг П.Дж. (2004). «Разнообразие и история эволюции пластид и их хозяев». Американский журнал ботаники . 91 (10): 1481–1493. дои : 10.3732/ajb.91.10.1481 . ПМИД 21652304 . S2CID 17522125 .
- ^ «Самая большая бактерия: ученый обнаруживает новую бактериальную форму жизни у африканского побережья» , Институт морской микробиологии Макса Планка , 8 апреля 1999 г., заархивировано из оригинала 20 января 2010 г.
- ^ Список названий прокариот, стоящих в номенклатуре - Род Thiomargarita
- ^ Банг С., Шмитц Р.А. (сентябрь 2015 г.). «Археи, связанные с человеческими поверхностями: нельзя недооценивать» . Обзоры микробиологии FEMS . 39 (5): 631–48. дои : 10.1093/femsre/fuv010 . ПМИД 25907112 .
- ^ «Архея» . Интернет-словарь этимологии . Проверено 17 августа 2016 г.
- ^ Пейс НР (май 2006 г.). «Время перемен» . Природа . 441 (7091): 289. Бибкод : 2006Natur.441..289P . дои : 10.1038/441289a . ПМИД 16710401 . S2CID 4431143 .
- ^ Стоекениус В. (октябрь 1981 г.). «Квадратная бактерия Уолсби: тонкая структура ортогонального прокариота» . Журнал бактериологии . 148 (1): 352–60. дои : 10.1128/JB.148.1.352-360.1981 . ПМК 216199 . ПМИД 7287626 .
- ^ Уиттакер Р.Х., Маргулис Л. (апрель 1978 г.). «Классификация протистов и царства организмов». Биосистемы . 10 (1–2): 3–18. дои : 10.1016/0303-2647(78)90023-0 . ПМИД 418827 .
- ^ Фор Э, Нот Ф, Бенуастон А.С., Лабади К., Биттнер Л., Аята С.Д. (апрель 2019 г.). «Миксотрофные протисты демонстрируют контрастную биогеографию в мировом океане» . Журнал ISME . 13 (4): 1072–1083. дои : 10.1038/s41396-018-0340-5 . ПМК 6461780 . ПМИД 30643201 .
- ^ Jump up to: а б Лелес С.Г., Митра А., Флинн К.Дж., Стокер Д.К., Хансен П.Дж., Калбет А. и др. (август 2017 г.). «Океанические протисты с различными формами приобретенной фототрофии демонстрируют контрастную биогеографию и численность» . Слушания. Биологические науки . 284 (1860): 20170664. doi : 10.1098/rspb.2017.0664 . ПМЦ 5563798 . ПМИД 28768886 .
- ^ Бадд Дж.Э., Дженсен С. (февраль 2017 г.). «Происхождение животных и гипотеза «саванны» ранней двусторонней эволюции» . Биологические обзоры Кембриджского философского общества . 92 (1): 446–473. дои : 10.1111/brv.12239 . ПМИД 26588818 .
- ^ Кавалер-Смит Т. (декабрь 1993 г.). «Царство простейших и его 18 типов» . Микробиологические обзоры . 57 (4): 953–94. дои : 10.1128/MMBR.57.4.953-994.1993 . ПМК 372943 . ПМИД 8302218 .
- ^ Корлисс Дж.О. (1992). «Должен ли быть отдельный кодекс номенклатуры для протистов?». Биосистемы . 28 (1–3): 1–14. дои : 10.1016/0303-2647(92)90003-H . ПМИД 1292654 .
- ^ Слапета Х., Морейра Д., Лопес-Гарсия П. (октябрь 2005 г.). «Степень разнообразия протистов: выводы из молекулярной экологии пресноводных эукариот» . Слушания. Биологические науки . 272 (1576): 2073–81. дои : 10.1098/rspb.2005.3195 . ПМЦ 1559898 . ПМИД 16191619 .
- ^ Морейра Д., Лопес-Гарсия П. (январь 2002 г.). «Молекулярная экология микробных эукариот открывает скрытый мир». Тенденции в микробиологии . 10 (1): 31–8. дои : 10.1016/S0966-842X(01)02257-0 . ПМИД 11755083 .
- ^ Jump up to: а б Алверсон А. (11 июня 2014 г.). «Воздух, которым вы дышите? Это сделала диатомовая водоросль» . Живая наука .
- ^ «Подробнее о диатомовых водорослях» . Музей палеонтологии Калифорнийского университета . Архивировано из оригинала 4 октября 2012 года . Проверено 27 июня 2019 г.
- ^ Девреотес П. (сентябрь 1989 г.). «Dictyostelium discoideum: модельная система межклеточных взаимодействий в развитии». Наука . 245 (4922): 1054–8. Бибкод : 1989Sci...245.1054D . дои : 10.1126/science.2672337 . ПМИД 2672337 .
- ^ Мац М.В., Фрэнк Т.М., Маршалл Н.Дж., Виддер Э.А., Джонсен С. (декабрь 2008 г.). «Гигантский глубоководный протист оставляет следы, похожие на двусторонние». Современная биология . 18 (23). Elsevier Ltd: 1849–54. дои : 10.1016/j.cub.2008.10.028 . ПМИД 19026540 . S2CID 8819675 .
- ^ Гудэй А.Дж., Да Силва А.А., Павловски Дж. (1 декабря 2011 г.). «Ксенофиофоры (Rhizaria, Foraminifera) из каньона Назаре (окраина Португалии, северо-восток Атлантики)». Глубоководные исследования. Часть II: Актуальные исследования в океанографии . Геология, геохимия и биология подводных каньонов к западу от Португалии. 58 (23–24): 2401–2419. Бибкод : 2011DSRII..58.2401G . дои : 10.1016/j.dsr2.2011.04.005 .
- ^ Нил А.С., Рис Дж.Б., Саймон Э.Дж. (2004). Основная биология с физиологией . Пирсон/Бенджамин Каммингс. п. 291. ИСБН 978-0-8053-7503-9 .
- ^ О'Мэлли М.А., Симпсон А.Г., Роджер А.Дж. (2012). «Другие эукариоты в свете эволюционной протистологии». Биология и философия . 28 (2): 299–330. дои : 10.1007/s10539-012-9354-y . S2CID 85406712 .
- ^ Адл С.М., Симпсон А.Г., Фармер М.А., Андерсен Р.А., Андерсон О.Р., Барта Дж.Р. и др. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию протистов» . Журнал эукариотической микробиологии . 52 (5): 399–451. дои : 10.1111/j.1550-7408.2005.00053.x . ПМИД 16248873 . S2CID 8060916 .
- ^ Маргулис Л., Чепмен М.Дж. (19 марта 2009 г.). Королевства и владения: Иллюстрированный путеводитель по типам жизни на Земле . Академическая пресса. ISBN 9780080920146 .
- ^ Фанг Дж (апрель 2010 г.). «Животные процветают без кислорода на морском дне» . Природа . 464 (7290): 825. Бибкод : 2010Natur.464..825F . дои : 10.1038/464825b . ПМИД 20376121 .
- ^ «Глубокий соленый бассейн может быть домом для животных, живущих без кислорода» . Новости науки . 23 сентября 2013 г.
- ^ Йонасдоттир, Сигрун Хульд; Виссер, Андре В.; Ричардсон, Кэтрин; Хит, Майкл Р. (29 сентября 2015 г.). «Сезонный липидный насос веслоногих ракообразных способствует секвестрации углерода в глубинах Северной Атлантики» . Труды Национальной академии наук . 112 (39): 12122–12126. дои : 10.1073/pnas.1512110112 . ISSN 0027-8424 . ПМЦ 4593097 . ПМИД 26338976 .
- ^ Пинти, Жером; Йонасдоттир, Сигрун Х.; Запись, Николас Р.; Виссер, Андре В. (7 марта 2023 г.). «Глобальный вклад сезонно мигрирующих копепод в биологический углеродный насос» . Лимнология и океанография . 68 (5): 1147–1160. Бибкод : 2023LimOc..68.1147P . дои : 10.1002/lno.12335 . ISSN 0024-3590 . S2CID 257422956 .
- ^ Пепато, Альмир Р.; Видигал, Теофания HDA; Климов, Павел Б. (2018). «Молекулярная филогения морских клещей (Acariformes: Halacaridae), древнейшей разновидности современных вторично морских животных» . Молекулярная филогенетика и эволюция . 129 : 182–188. дои : 10.1016/j.ympev.2018.08.012 . ПМИД 30172010 . S2CID 52145427 .
- ^ Хайд К.Д., Джонс Э.Г., Леаньо Э., Пойнтинг С.Б., Пунит А.Д., Вреймоед Л.Л. (1998). «Роль грибов в морских экосистемах». Биоразнообразие и сохранение . 7 (9): 1147–1161. дои : 10.1023/А:1008823515157 . S2CID 22264931 .
- ^ Кирк П.М., Кэннон П.Ф., Минтер Д.В., Сталперс Дж. (2008). Словарь грибов (10-е изд.). КАБИ.
- ^ Хайд К.Д., Гринвуд Р., Джонс Э.Г. (1989). «Прикрепление спор у морских грибов». Ботаника Марина . 32 (3): 205–218. дои : 10.1515/ботм.1989.32.3.205 . S2CID 84879817 .
- ^ Ле Кальвес Т., Бурго Г., Маэ С., Барбье Г., Ванденкокорнхейс П. (октябрь 2009 г.). «Грибное разнообразие в глубоководных гидротермальных экосистемах» . Прикладная и экологическая микробиология . 75 (20): 6415–21. Бибкод : 2009ApEnM..75.6415L . дои : 10.1128/АЕМ.00653-09 . ПМЦ 2765129 . ПМИД 19633124 .
- ^ Сан-Мартин А., Оредхарена С., Галлардо С., Сильва М., Бесерра Дж., Рейносо Р.О., Чами М.К., Вергара К., Ровироса Дж. (2008). «Стероиды морского гриба Geotrichum sp» . Журнал Чилийского химического общества . 53 (1): 1377–1378. дои : 10.4067/S0717-97072008000100011 .
- ^ Джонс Э.Б., Хайд К.Д., Панг К.Л., ред. (2014). Пресноводные грибы: и грибоподобные организмы . Берлин/Бостон: Де Грюйтер. ISBN 9783110333480 .
- ^ Джонс Э.Б., Панг К.Л., ред. (2012). Морские грибы и грибоподобные организмы . Морская и пресноводная ботаника. Берлин, Бостон: Де Грюйтер (опубликовано в августе 2012 г.). дои : 10.1515/9783110264067 . ISBN 978-3-11-026406-7 . Проверено 3 сентября 2015 г.
- ^ Ван X, Сингх П., Гао Z, Чжан X, Джонсон ZI, Ван G (2014). «Распространение и разнообразие планктонных грибов в теплом бассейне западной части Тихого океана» . ПЛОС ОДИН . 9 (7): e101523. Бибкод : 2014PLoSO...9j1523W . дои : 10.1371/journal.pone.0101523.s001 . ПМК 4081592 . ПМИД 24992154 .
- ^ Ван Г, Ван Х, Лю Х, Ли Ц (2012). «Разнообразие и биогеохимическая функция планктонных грибов океана» . В Рагукумаре С. (ред.). Биология морских грибов . Прогресс молекулярной и субклеточной биологии. Том. 53. Берлин, Гейдельберг: Springer-Verlag. стр. 71–88. дои : 10.1007/978-3-642-23342-5 . ISBN 978-3-642-23341-8 . S2CID 39378040 . Проверено 3 сентября 2015 г.
- ^ Дамаре С., Рагукумар С. (июль 2008 г.). «Грибы и макроагрегация в глубоководных отложениях». Микробная экология . 56 (1): 168–77. дои : 10.1007/s00248-007-9334-y . ПМИД 17994287 . S2CID 21288251 .
- ^ Кубанек Дж., Дженсен П.Р., Кейфер П.А., Саллардс М.К., Коллинз Д.О., Феникал В. (июнь 2003 г.). «Устойчивость морских водорослей к микробному воздействию: целенаправленная химическая защита от морских грибов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (12): 6916–21. Бибкод : 2003PNAS..100.6916K . дои : 10.1073/pnas.1131855100 . ПМК 165804 . ПМИД 12756301 .
- ^ Jump up to: а б Гао З., Джонсон З.И., Ван Дж. (январь 2010 г.). «Молекулярная характеристика пространственного разнообразия и новых линий микопланктона в прибрежных водах Гавайских островов» . Журнал ISME . 4 (1): 111–20. дои : 10.1038/ismej.2009.87 . ПМИД 19641535 .
- ^ Панцер К., Йилмаз П., Вайс М., Райх Л., Рихтер М., Визе Дж. и др. (30 июля 2015 г.). «Идентификация биомов водных грибковых сообществ, специфичных для среды обитания, с использованием комплексного почти полноразмерного набора данных 18S рРНК, обогащенного контекстуальными данными» . ПЛОС ОДИН . 10 (7): e0134377. Бибкод : 2015PLoSO..1034377P . дои : 10.1371/journal.pone.0134377 . ПМЦ 4520555 . ПМИД 26226014 .
- ^ Гутьеррес М.Х., Пантоха С., Хинонес Р.А., Гонсалес Р.Р. (2010). «Первая запись нитчатых грибов в прибрежной экосистеме апвеллинга у центрального Чили». Гаяна (на испанском языке). 74 (1): 66–73.
- ^ Jump up to: а б Шридхар КР (2009). «10. Водные грибы – они планктонные?». Планктонная динамика вод Индии . Джайпур, Индия: Публикации Пратикши. стр. 133–148.
- ^ «Виды высших морских грибов» . Университет Миссисипи. Архивировано из оригинала 22 апреля 2013 года . Проверено 5 февраля 2012 г.
- ^ Хоксворт Д.Л. (2000). «Пресноводные и морские лишайниковообразующие грибы» (PDF) . Грибное разнообразие . 5 : 1–7.
- ^ «Лишайники» . Служба национальных парков, Министерство внутренних дел США, Правительство США. 22 мая 2016 года . Проверено 4 апреля 2018 г.
- ^ Jump up to: а б «Сеть земной жизни, рост и развитие лишайников» . Earthlife.net. 14 февраля 2020 г.
- ^ Силлиман Б.Р., Ньюэлл С.Ю. (декабрь 2003 г.). «Грибное хозяйство улитки» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (26): 15643–8. Бибкод : 2003PNAS..10015643S . дои : 10.1073/pnas.2535227100 . ПМК 307621 . ПМИД 14657360 .
- ^ Юань X, Сяо С., Тейлор Т.Н. (май 2005 г.). «Лишайноподобный симбиоз 600 миллионов лет назад». Наука . 308 (5724): 1017–20. Бибкод : 2005Sci...308.1017Y . дои : 10.1126/science.1111347 . ПМИД 15890881 . S2CID 27083645 .
- ^ Джонс Э.Б., Панг К.Л. (31 августа 2012 г.). Морские грибы: и грибоподобные организмы . Вальтер де Грютер. ISBN 9783110264067 .
- ^ Дэвидсон М.В. (26 мая 2005 г.). «Структура животных клеток» . Молекулярные выражения . Таллахасси, Флорида: Университет штата Флорида . Проверено 3 сентября 2008 г.
- ^ Фогель Г. (20 сентября 2018 г.). «Это ископаемое является одним из самых ранних животных в мире, если судить по молекулам жира, сохранившимся в течение полумиллиарда лет» . Наука . АААС . Проверено 21 сентября 2018 г.
- ^ Бобровский И., Хоуп Дж.М., Иванцов А., Неттерсхайм Б.Дж., Холлманн С., Брокс Дж.Дж. (сентябрь 2018 г.). «Древние стероиды делают эдиакарскую ископаемую дикинсонию одним из самых ранних животных» . Наука . 361 (6408): 1246–1249. Бибкод : 2018Sci...361.1246B . дои : 10.1126/science.aat7228 . hdl : 1885/230014 . ПМИД 30237355 .
- ^ Реталлак Г.Дж. (2007). «Рост, распад и уплотнение захоронений дикинсонии, культовой окаменелости Эдиакарского региона» (PDF) . Алчеринга: Австралазийский журнал палеонтологии . 31 (3): 215–240. Бибкод : 2007Алч...31..215Р . дои : 10.1080/03115510701484705 . S2CID 17181699 .
- ^ Сперлинг Э.А., Винтер Дж (2010). «Сродство плакозой к дикинсонии и эволюция способов питания многоклеточных животных позднего протерозоя». Эволюция и развитие . 12 (2): 201–9. дои : 10.1111/j.1525-142X.2010.00404.x . ПМИД 20433459 . S2CID 38559058 .
- ^ Голд Д.А., Раннегар Б., Гелинг Дж.Г., Джейкобс Д.К. (2015). «Реконструкция онтогенеза наследственного состояния подтверждает двустороннюю близость к дикинсонии». Эволюция и развитие . 17 (6): 315–24. дои : 10.1111/ede.12168 . ПМИД 26492825 . S2CID 26099557 .
- ^ Чен Дж.Ю., Оливери П., Гао Ф., Дорнбос С.К., Ли К.В., Боттьер DJ, Дэвидсон Э.Х. (август 2002 г.). «Животный мир докембрия: вероятные формы развития и взрослые книдарии из Юго-Западного Китая» (PDF) . Биология развития . 248 (1): 182–96. дои : 10.1006/dbio.2002.0714 . ПМИД 12142030 . Архивировано из оригинала (PDF) 26 мая 2013 года . Проверено 4 февраля 2015 г.
- ^ Гражданкин Д (июнь 2004 г.). «Схемы распространения в эдиакарской биоте: фации против биогеографии и эволюции». Палеобиология . 30 (2): 203–221. doi : 10.1666/0094-8373(2004)030<0203:PODITE>2.0.CO;2 . ISSN 0094-8373 . S2CID 129376371 .
- ^ Сейлахер А (август 1992 г.). «Вендобионта и Псаммокораллия: утраченные конструкции докембрийской эволюции» . Журнал Геологического общества . 149 (4): 607–613. Бибкод : 1992JGSoc.149..607S . дои : 10.1144/gsjgs.149.4.0607 . ISSN 0016-7649 . S2CID 128681462 . Проверено 4 февраля 2015 г.
- ^ Мартин М.В., Гражданкин Д.В., Боуринг С.А., Эванс Д.А., Федонкин М.А., Киршвинк Дж.Л. (май 2000 г.). «Возраст неопротерозойского тела двулатария и следы окаменелостей, Белое море, Россия: значение для эволюции многоклеточных животных». Наука . 288 (5467): 841–5. Бибкод : 2000Sci...288..841M . дои : 10.1126/science.288.5467.841 . ПМИД 10797002 . S2CID 1019572 .
- ^ Федонкин М.А., Вагонер Б.М. (28 августа 1997 г.). позднего докембрия «Ископаемая кимберелла представляет собой двусторонний организм, похожий на моллюска» . Природа . 388 (6645): 868–871. Бибкод : 1997Natur.388..868F . дои : 10.1038/42242 . ISSN 0028-0836 . S2CID 4395089 .
- ^ Муи Р., Дэвид Б. (декабрь 1998 г.). «Эволюция внутри причудливого типа: гомологии первых иглокожих» . Американский зоолог . 38 (6): 965–974. дои : 10.1093/icb/38.6.965 . ISSN 1540-7063 .
- ^ Макменамин М.А. (сентябрь 2003 г.). Сприггина — трилобитоидное экдизозойное животное . Геонаучные горизонты, Сиэтл, 2003 . Рефераты с программами. Том. 35. Боулдер, Колорадо: Геологическое общество Америки. п. 105. OCLC 249088612 . Архивировано из оригинала 12 апреля 2016 года . Проверено 24 ноября 2007 г. Документ № 40-2, представленный на ежегодном собрании Геологического общества Америки в Сиэтле 2003 г. (2–5 ноября 2003 г.) 2 ноября 2003 г. в Конференц-центре штата Вашингтон .
- ^ Линь Дж.П., Гон III С.М., Гелинг Дж.Г., Бэбкок Л.Е., Чжао Ю.Л., Чжан С.Л. и др. (2006). « Парванкориноподобное членистоногое из кембрия Южного Китая». Историческая биология: Международный журнал палеобиологии . 18 (1): 33–45. дои : 10.1080/08912960500508689 . ISSN 1029-2381 . S2CID 85821717 .
- ^ Баттерфилд, штат Нью-Джерси (декабрь 2006 г.). «Зацепка некоторых «червей» стволовой группы: ископаемые лофотрохозойи в сланцах Берджесс». Биоэссе . 28 (12): 1161–6. doi : 10.1002/bies.20507 . ПМИД 17120226 . S2CID 29130876 .
- ^ Jump up to: а б Бенгтсон С. (ноябрь 2004 г.). «Ранние скелетные окаменелости». Документы Палеонтологического общества . 10 : 67–78. дои : 10.1017/S1089332600002345 .
- ^ Валентин Дж.В. (2004). О происхождении Фила . Чикаго: Издательство Чикагского университета. п. 7. ISBN 978-0-226-84548-7 .
Классификации организмов в иерархических системах использовались в семнадцатом и восемнадцатом веках. Обычно организмы группировались в соответствии с их морфологическим сходством, как это воспринималось теми ранними исследователями, а затем эти группы группировались в соответствии с их сходством и так далее, чтобы сформировать иерархию.
- ^ Jump up to: а б Валентин JW (18 июня 2004 г.). О происхождении Фила . Издательство Чикагского университета. ISBN 9780226845487 .
- ^ «WoRMS — Всемирный реестр морских видов» . www.marinespecies.org .
- ^ Новак Б.Дж., Фрейзер Д., Мэлони Т.Х. (февраль 2020 г.). «Преобразование охраны океана: применение набора инструментов генетического спасения» . Гены . 11 (2): 209. doi : 10.3390/genes11020209 . ПМК 7074136 . ПМИД 32085502 .
- ^ Гулд С.Дж. (1990). Прекрасная жизнь: сланцы Бёрджесс и природа истории . WW Нортон. ISBN 978-0-393-30700-9 .
- ^ Эрвин Д., Валентайн Дж., Яблонски Д. (1997). «Недавние находки окаменелостей и новое понимание развития животных открывают новый взгляд на загадку массового расцвета животных в раннем кембрии» . Американский ученый (март – апрель).
- ^ Jump up to: а б Бадд Дж.Э., Дженсен С. (май 2000 г.). «Критическая переоценка летописи окаменелостей двусторонних типов». Биологические обзоры Кембриджского философского общества . 75 (2): 253–95. дои : 10.1111/j.1469-185X.1999.tb00046.x . ПМИД 10881389 . S2CID 39772232 .
- ^ Гулд С.Дж. (1989). Прекрасная жизнь: сланцы Берджесс и природа истории (Первое изд.). Нью-Йорк. ISBN 978-0-393-02705-1 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Бадд Дж. Э. (февраль 2003 г.). «Кембрийская летопись окаменелостей и происхождение типов» . Интегративная и сравнительная биология . 43 (1): 157–65. дои : 10.1093/icb/43.1.157 . ПМИД 21680420 .
- ^ Бадд Дж. Э. (март 1996 г.). «Морфология Opabinia regalis и реконструкция стеблевой группы членистоногих». Летайя . 29 (1): 1–14. дои : 10.1111/j.1502-3931.1996.tb01831.x . ISSN 0024-1164 .
- ^ Маршалл CR (май 2006 г.). «Объяснение кембрийского «взрыва» животных». Ежегодный обзор наук о Земле и планетах . 34 : 355–384. Бибкод : 2006AREPS..34..355M . doi : 10.1146/annurev.earth.33.031504.103001 . ISSN 1545-4495 . S2CID 85623607 .
- ^ Кинг Н., Рокас А. (октябрь 2017 г.). «Принятие неопределенности в реконструкции ранней эволюции животных» . Современная биология . 27 (19): Р1081–Р1088. дои : 10.1016/j.cub.2017.08.054 . ПМК 5679448 . ПМИД 29017048 .
- ^ Феуда Р., Дорманн М., Петт В., Филипп Х., Рота-Стабелли О., Лартильо Н. и др. (декабрь 2017 г.). «Улучшенное моделирование композиционной неоднородности подтверждает, что губки являются сестрами всех других животных» . Современная биология . 27 (24): 3864–3870.e4. дои : 10.1016/j.cub.2017.11.008 . hdl : 11572/302898 . ПМИД 29199080 .
- ^ Нильсен С (июль 2019 г.). «Ранняя эволюция животных: взгляд морфолога» . Королевское общество открытой науки . 6 (7): 190638. Бибкод : 2019RSOS....690638N . дои : 10.1098/rsos.190638 . ПМК 6689584 . ПМИД 31417759 .
- ^ «Порифера (н.)» . Интернет-словарь этимологии . Проверено 18 августа 2016 г.
- ^ Jump up to: а б Петралия Р.С., Мэттсон, член парламента, Яо П.Дж. (июль 2014 г.). «Старение и долголетие у простейших животных и поиски бессмертия» . Обзоры исследований старения . 16 :66–82. дои : 10.1016/J.arr.2014.05.003 . ПМЦ 4133289 . ПМИД 24910306 .
- ^ Йохум КП, Ван Х, Веннеманн Т.В., Синха Б., Мюллер В.Е. (2012). «Кремневая глубоководная губка Monorhaphis chuni : потенциальный архив палеоклимата древних животных». Химическая геология . 300 : 143–151. Бибкод : 2012ЧГео.300..143J . doi : 10.1016/j.chemgeo.2012.01.009 .
- ^ Васелет Дж., Дюпор Э. (ноябрь 2004 г.). «Захват и переваривание добычи у плотоядной губки Asbestopluma hypogea (Porifera: Demospongiae)». Зооморфология . 123 (4): 179–90. дои : 10.1007/s00435-004-0100-0 . S2CID 24484610 .
- ^ « Губка Линней , 1759 год» . Всемирный регистр морских видов . Проверено 18 июля 2012 г.
- ^ Роуленд С.М., Стивенс Т. (2001). «Археоциаты: история филогенетической интерпретации». Журнал палеонтологии . 75 (6): 1065–1078. doi : 10.1666/0022-3360(2001)075<1065:AAHOPI>2.0.CO;2 . JSTOR 1307076 . S2CID 86211946 .
- ^ Сперлинг Э.А., Пизани Д., Петерсон К.Дж. (1 января 2007 г.). «Пориферановая парафилия и ее значение для палеобиологии докембрия» (PDF) . Геологическое общество, Лондон, специальные публикации . 286 (1): 355–368. Бибкод : 2007GSLSP.286..355S . дои : 10.1144/SP286.25 . S2CID 34175521 . Архивировано из оригинала (PDF) 9 мая 2009 года . Проверено 22 августа 2012 г.
- ^ Руперт Э.Э., Фокс Р.С., Барнс Р.Д. (2004). Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 182–195 . ISBN 978-0-03-025982-1 .
- ^ Миллс CE. «Гребневики – некоторые заметки эксперта» . Проверено 5 февраля 2009 г.
- ^ Jump up to: а б Бруска Р.К., Бруска Г.Дж. (2003). Беспозвоночные (Второе изд.). Синауэр Ассошиэйтс. ISBN 978-0-87893-097-5 .
- ^ Ле Пейдж М (март 2019 г.). «Животное с анусом, который приходит и уходит, может показать, как развивался наш анус» . Новый учёный .
- ^ Мартиндейл MQ, Финнерти-младший, Генри JQ (сентябрь 2002 г.). «Ладиаты и эволюционное происхождение двустороннего строения тела». Молекулярная филогенетика и эволюция . 24 (3): 358–65. дои : 10.1016/s1055-7903(02)00208-7 . ПМИД 12220977 .
- ^ Placozoa в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- ^ Венер Р., Геринг В. (июнь 2007 г.). Зоология (на немецком языке) (24-е изд.). Штутгарт: Тиме. п. 696.
- ^ Шульце Ф.Э. (1883). « Трихоплакс прилипший , нс». Зоологический вестник . Том 6. Амстердам и Йена: Эльзевир. п. 92.
- ^ Эйтель М., Фрэнсис В.Р., Осигус Х.Дж., Кребс С., Варгас С., Блюм Х. и др. (13 октября 2017 г.). «Таксогеномный подход открывает новый род в типе Placozoa» . bioRxiv : 202119. doi : 10.1101/202119 . S2CID 89829846 .
- ^ Осигус Х.Дж., Рольфес С., Херцог Р., Камм К., Шируотер Б. (март 2019 г.). «Polyplacotoma mediterranea — новый разветвленный вид плакозойных» . Современная биология . 29 (5): Р148–Р149. дои : 10.1016/j.cub.2019.01.068 . ПМИД 30836080 .
- ^ «Прилипание трихоплакса» . ВОРМС . 2009 год
- ^ Смит К.Л., Варокко Ф., Киттельманн М., Аззам Р.Н., Купер Б., Уинтерс К.А. и др. (июль 2014 г.). «Новые типы клеток, нейросекреторные клетки и строение тела ранних дивергентных многоклеточных животных Trichoplax adhaerens» . Современная биология . 24 (14): 1565–1572. дои : 10.1016/j.cub.2014.05.046 . ПМЦ 4128346 . ПМИД 24954051 .
- ^ Барнс Р.Д. (1982). Зоология беспозвоночных . Филадельфия: Холт-Сондерс Интернэшнл. стр. 84–85. ISBN 978-0-03-056747-6 .
- ^ Чжан ZQ (2011). «Биоразнообразие животных: введение в классификацию более высокого уровня и таксономическое богатство» (PDF) . Зоотакса . 3148 : 7–12. дои : 10.11646/zootaxa.3148.1.3 .
- ^ « Нематостелла vectensis v1.0» . Геномный портал . Калифорнийский университет . Проверено 19 января 2014 г.
- ^ « Нематостелла » . Nematostella.org. Архивировано из оригинала 8 мая 2006 года . Проверено 18 января 2014 г.
- ^ Jump up to: а б Генихович Г, Технау У (сентябрь 2009 г.). «Старлетка актиния Nematostella vectensis: модельный организм антозоев для исследований в области сравнительной геномики и функциональной эволюционной биологии развития». Протоколы Колд-Спринг-Харбора . 2009 (9): pdb.emo129. дои : 10.1101/pdb.emo129 . ПМИД 20147257 .
- ^ «Откуда берется наша голова? Безмозглая актиния проливает новый свет на эволюционное происхождение головы» . Наука Дейли . 12 февраля 2013 года . Проверено 18 января 2014 г.
- ^ Синигалья К., Бусенгдал Х., Леклер Л., Технау У., Ренцш Ф. (2013). «Ген формирования билатерального паттерна головы шесть3/6 контролирует развитие аборального домена у книдарий» . ПЛОС Биология . 11 (2): e1001488. дои : 10.1371/journal.pbio.1001488 . ПМЦ 3586664 . ПМИД 23483856 .
- ^ «Медуза-красный бумажный фонарь» . Настоящие чудовища . Проверено 25 октября 2015 г.
- ^ «Голубые кнопки во Флориде» . BeachHunter.net .
- ^ Карлескинт Дж., Тернер Р., Смолл Дж. (2012). Введение в морскую биологию (4-е изд.). Cengage Обучение. п. 445. ИСБН 978-1-133-36446-7 .
- ^ Бавестрелло Дж., Соммер С., Сара М. (1992). «Двунаправленное преобразование у Turritopsis nutricula (Hydrozoa)». Морская наука . 56 (2–3): 137–140.
- ^ Пираино С., Боэро Ф., Эшбах Б., Шмид В. (1996). «Обращение жизненного цикла: медузы превращаются в полипы и трансдифференцировка клеток у Turritopsis nutricula (Cnidaria, Hydrozoa)». Биологический бюллетень . 190 (3): 302–312. дои : 10.2307/1543022 . JSTOR 1543022 . ПМИД 29227703 . S2CID 3956265 .
- ^ Феннер П.Дж., Уильямсон Дж.А. (1996). «Смерть во всем мире и серьезные отравления от укусов медуз». Медицинский журнал Австралии . 165 (11–12): 658–61. дои : 10.5694/j.1326-5377.1996.tb138679.x . ПМИД 8985452 . S2CID 45032896 .
- ^ Jump up to: а б с Кэннон Дж.Т., Веллутини Б.С., Смит Дж., Ронквист Ф., Джонделиус Ю., Хейнол А. (февраль 2016 г.). «Xenacoelomorpha — сестринская группа Nephrozoa» . Природа . 530 (7588): 89–93. Бибкод : 2016Natur.530...89C . дои : 10.1038/nature16520 . ПМИД 26842059 . S2CID 205247296 .
- ^ Jump up to: а б Минелли А (2009). Перспективы филогении и эволюции животных . Издательство Оксфордского университета. п. 53. ИСБН 978-0-19-856620-5 .
- ^ Jump up to: а б с Бруска RC (2016). Введение в Bilateria и тип Xenacoelomorpha | Триплобластия и двусторонняя симметрия открывают новые возможности для облучения животных (PDF) . Синауэр Ассошиэйтс. стр. 345–372. ISBN 978-1605353753 .
{{cite book}}:|work=игнорируется ( помогите ) - ^ Финнерти-младший (ноябрь 2005 г.). «Способствовал ли внутренний транспорт, а не направленное передвижение, развитию двусторонней симметрии у животных?» (PDF) . Биоэссе . 27 (11): 1174–80. дои : 10.1002/bies.20299 . ПМИД 16237677 . Архивировано из оригинала (PDF) 10 августа 2014 года . Проверено 27 августа 2019 г.
- ^ Куиллин К.Дж. (май 1998 г.). «Онтогенетическое масштабирование гидростатических скелетов: масштабирование геометрического, статического и динамического стресса дождевого червя lumbricus terrestris» . Журнал экспериментальной биологии . 201 (12): 1871–83. дои : 10.1242/jeb.201.12.1871 . ПМИД 9600869 .
- ^ Спектор Б (26 марта 2020 г.). «Этот первобытный червь может быть предком всех животных]» . Живая наука» .
- ^ Jump up to: а б Уэйд Н. (30 января 2017 г.). «Этот доисторический предок человека был весь рот» . Нью-Йорк Таймс . Проверено 31 января 2017 г.
- ^ Jump up to: а б Хан Дж., Моррис СК, Оу Ц, Шу Д., Хуан Х. (февраль 2017 г.). «Мейофауна вторичноротых из базального кембрия Шэньси (Китай)». Природа . 542 (7640): 228–231. Бибкод : 2017Natur.542..228H . дои : 10.1038/nature21072 . ПМИД 28135722 . S2CID 353780 .
- ^ «Корнуолл – Природа – Червь-суперзвезда» . Би-би-си .
- ^ Марк Карвардин (1995) Книга рекордов Гиннеса животных . Издательство Гиннесс. п. 232.
- ^ «Постоянные паразиты» . Время . 8 апреля 1957 года. Архивировано из оригинала 27 июня 2008 года.
- ^ Харгис В., изд. (1985). Паразитология и патология морских организмов Мирового океана (Технический отчет). Национальное управление океанических и атмосферных исследований .
- ^ «Классификация паразитов животных» . Архивировано из оригинала 14 сентября 2006 года.
- ^ Гарсия Л.С. (октябрь 1999 г.). «Классификация паразитов, переносчиков и подобных организмов человека» . Клинические инфекционные болезни . 29 (4): 734–6. дои : 10.1086/520425 . ПМИД 10589879 .
- ^ Ходда М (2011). «Тип Nematoda Cobb, 1932. В: Чжан, З.-К. Биоразнообразие животных: обзор классификации более высокого уровня и обзор таксономического богатства». Зоотакса . 3148 : 63–95. дои : 10.11646/zootaxa.3148.1.11 .
- ^ Чжан Цзы (2013). «Биоразнообразие животных: обновление классификации и разнообразия в 2013 году. В: Чжан, З.-К. (ред.) Биоразнообразие животных: план классификации более высокого уровня и исследование таксономического богатства (Дополнения, 2013 г.)» . Зоотакса . 3703 (1): 5–11. дои : 10.11646/zootaxa.3703.1.3 . S2CID 85252974 .
- ^ Ламбсхед П.Дж. (1993). «Последние достижения в исследованиях биоразнообразия морского бентоса». Океанис . 19 (6): 5–24.
- ^ Боргони Г., Гарсиа-Мояно А., Литтауэр Д., Берт В., Бестер А., ван Херден Е. и др. (июнь 2011 г.). «Нематода из земных глубоких недр Южной Африки » Природа 474 (7349): 79–82. Бибкод : 2011Природа.474...79Б . дои : 10.1038/nature09974 . hdl : 1854/LU-1269676 . ПМИД 21637257 . S2CID 4399763 .
- ^ Дановаро Р., Гамби С., Делл'Анно А., Коринальдези С., Фраскетти С., Ванреузель А. и др. (январь 2008 г.). «Экспоненциальное снижение функционирования глубоководных экосистем, связанное с утратой донного биоразнообразия» . Современная биология . 18 (1): 1–8. дои : 10.1016/j.cub.2007.11.056 . ПМИД 18164201 . S2CID 15272791 .
- «Исчезновение глубоководных видов может привести к коллапсу океанов, предполагает исследование» . ЭврекАлерт! (Пресс-релиз). 27 декабря 2007 г.
- ^ Платт Х.М. (1994). «предисловие». В Лоренцен С., Лоренцен С.А. (ред.). Филогенетическая систематика свободноживущих нематод . Лондон: Общество Рэев. ISBN 978-0-903874-22-9 .
- ^ Барнс Р.С., Калоу П., Олив П.Дж. (2001). Беспозвоночные, синтез (3-е изд.). Великобритания: Blackwell Science.
- ^ «Тирийский пурпур» . Зеленый Лев . 28 февраля 2014 г. Архивировано из оригинала 28 февраля 2014 г.
- ^ Чепмен А.Д. (2009). Численность живых видов в Австралии и мире (2-е изд.). Канберра: Исследование биологических ресурсов Австралии. ISBN 978-0-642-56860-1 .
- ^ Хэнкок Р. (2008). «Признание исследований моллюсков» . Австралийский музей. Архивировано из оригинала 30 мая 2009 года . Проверено 9 марта 2009 г.
- ^ Подумайте В.Ф., Линдберг Д.Р., ред. (2008). Филогения и эволюция моллюсков . Беркли: Издательство Калифорнийского университета. п. 481. ИСБН 978-0-520-25092-5 .
- ^ Манро Д., Блиер П.У. (октябрь 2012 г.). «Чрезвычайное долголетие Arctica Islandica связано с повышенной устойчивостью к перекисному окислению в митохондриальных мембранах» . Стареющая клетка . 11 (5): 845–55. дои : 10.1111/j.1474-9726.2012.00847.x . ПМИД 22708840 . S2CID 205634828 .
- ^ «Добро пожаловать в CephBase» . CephBase . Архивировано из оригинала 12 января 2016 года . Проверено 29 января 2016 г.
- ^ Уилбур К.М., Кларк, М.Р., Труман, ER, ред. (1985), Моллюска , том. 12. Палеонтология и неонтология головоногих моллюсков, Нью-Йорк: Academic Press, ISBN. 0-12-728702-7
- ^ «Есть ли пресноводные головоногие моллюски?» . Австралийская радиовещательная корпорация . 16 января 2013 г.
- ^ Каллауэй Э (2 июня 2008 г.). «Простодушный Наутилус демонстрирует вспышку памяти» . Новый учёный . Проверено 7 марта 2012 г.
- ^ Филлипс К. (15 июня 2008 г.). «Живые ископаемые воспоминания». Журнал экспериментальной биологии . 211 (12): iii. дои : 10.1242/jeb.020370 . S2CID 84279320 .
- ^ Крук Р., Бэзил Дж (2008). «Двухфазная кривая памяти у наутилуса с камерами Nautilus pompilius L. (Cephalopoda: Nautiloidea)». Журнал экспериментальной биологии . 211 (12): 1992–1998. дои : 10.1242/jeb.018531 . ПМИД 18515730 . S2CID 6305526 .
- ^ Черный R (26 апреля 2008 г.). «Колоссальный кальмар из морозилки» . Новости Би-би-си .
- ^ Jump up to: а б Руперт Р.Э., Фокс Р.С., Барнс Р.Д. (2004). Зоология беспозвоночных (7-е изд.). Cengage Обучение. ISBN 978-81-315-0104-7 .
- ^ Хейворд П.Дж. (1996). Справочник морской фауны Северо-Западной Европы . Издательство Оксфордского университета. ISBN 978-0-19-854055-7 .
- ^ Уилсон Х.М., Андерсон Л.И. (январь 2004 г.). «Морфология и систематика палеозойских многоножек (Diplopoda: Chilognatha: Archipolypoda) из Шотландии» . Журнал палеонтологии . 78 (1): 169–184. doi : 10.1666/0022-3360(2004)078<0169:MATOPM>2.0.CO;2 . S2CID 131201588 .
- ^ Суарес С.Е., Брукфилд М.Э., Катлос Э.Дж., Штёкли Д.Ф. (2017). «U-Pb возраст циркона самого старого зарегистрированного наземного животного, дышащего воздухом» . ПЛОС ОДИН . 12 (6): e0179262. Бибкод : 2017PLoSO..1279262S . дои : 10.1371/journal.pone.0179262 . ПМЦ 5489152 . ПМИД 28658320 .
- ^ Кэмпбелл Л.И., Рота-Стабелли О., Эджкомб Г.Д., Маркиоро Т., Лонгхорн С.Дж., Телфорд М.Дж. и др. (сентябрь 2011 г.). «МикроРНК и филогеномика раскрывают взаимоотношения тихоходок и позволяют предположить, что бархатные черви являются сестринской группой членистоногих» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (38): 15920–4. Бибкод : 2011PNAS..10815920C . дои : 10.1073/pnas.1105499108 . ПМК 3179045 . ПМИД 21896763 .
- ^ Смит Ф.В., Гольдштейн Б. (май 2017 г.). «Сегментация тихоходок и разнообразие сегментных моделей панартропод». Строение и развитие членистоногих . 46 (3): 328–340. дои : 10.1016/j.asd.2016.10.005 . ПМИД 27725256 .
- ^ Бадд Дж.Э. (2001). «Почему членистоногие сегментированы?». Эволюция и развитие . 3 (5): 332–42. дои : 10.1046/j.1525-142X.2001.01041.x . ПМИД 11710765 . S2CID 37935884 .
- ^ «Первая жизнь Дэвида Аттенборо» . Архивировано из оригинала 26 января 2011 года . Проверено 10 марта 2011 г.
- ^ Брэдди С.Дж., Пошманн М., Тетли О.Э. (февраль 2008 г.). «Гигантский коготь обнаружил самое большое членистоногое в истории» . Письма по биологии . 4 (1): 106–9. дои : 10.1098/rsbl.2007.0491 . ПМЦ 2412931 . ПМИД 18029297 .
- ^ Дэниел С. (21 ноября 2007 г.). «Обнаружен гигантский морской скорпион» . Природа . дои : 10.1038/news.2007.272 . Проверено 10 июня 2013 г.
- ^ Бикнелл, Рассел, округ Колумбия; Пейтс, Стивен (2020). «Иллюстрированный атлас ископаемых и современных мечехвостов с акцентом на Xiphosurida» . Границы в науках о Земле . 8 : 98. Бибкод : 2020FrEaS...8...98B . дои : 10.3389/feart.2020.00098 . ISSN 2296-6463 .
- ^ «Уродливый гигантский краб Японии» . Популярная наука . 96 (6): 42. 1920.
- ^ Карри Д.Р., Уорд ТМ (2009). Южно-Австралийский гигантский краб ( Pseudocarcinus gigas ) Промысел (PDF) . Южно-Австралийский институт исследований и разработок . Отчет об оценке рыболовства для PIRSA. Архивировано из оригинала (PDF) 28 марта 2012 года . Проверено 9 декабря 2013 г.
- ^ Килдей П. (28 сентября 2005 г.). «Креветка-богомол может похвастаться самыми развитыми глазами» . Ежедневный калифорнийец . Архивировано из оригинала 29 сентября 2012 года . Проверено 23 сентября 2016 г.
- ^ Патек С.Н., Колдуэлл Р.Л. (2005). «Чрезвычайные силы удара и кавитации биологического молота: ударные силы креветки-богомола-павлина» . Журнал экспериментальной биологии . 208 (19): 3655–3664. дои : 10.1242/jeb.01831 . ПМИД 16169943 . S2CID 312009 .
- ^ Хан, Цзянь; Моррис, Саймон Конвей; Оу, Цян; Шу, Деган; Хуан, Хай (2017). «Мейофауна вторичноротых из базального кембрия Шэньси (Китай)» . Природа . 542 (7640): 228–231. Бибкод : 2017Natur.542..228H . дои : 10.1038/nature21072 . ISSN 1476-4687 . ПМИД 28135722 . S2CID 353780 .
- ^ Лю, Юньхуань; Карлайл, Эмили; Чжан, Хуацяо; Ян, Бен; Штайнер, Майкл; Шао, Тецюань; Дуань, Байчуань; Мароне, Федерика; Сяо, Шухай; Донохью, Филип CJ (17 августа 2022 г.). «Saccorhytus — ранний экдисозой, а не самый ранний вторичноротый» . Природа . 609 (7927): 541–546. Бибкод : 2022Natur.609..541L . дои : 10.1038/s41586-022-05107-z . hdl : 1983/454e7bec-4cd4-4121-933e-abeab69e96c1 . ISSN 1476-4687 . ПМИД 35978194 . S2CID 251646316 .
- ^ «Сеть разнообразия животных - Иглокожие» . Зоологический музей Мичиганского университета . Проверено 26 августа 2012 г.
- ^ «Иглокожие | Определение, характеристики, виды и факты | Британника» . www.britanica.com . 8 июня 2023 г. Проверено 24 июня 2023 г.
- ^ Фокс Р. « Asterias forbesi » . Анатомия беспозвоночных онлайн . Ландерский университет . Проверено 14 июня 2014 г.
- ^ Холсингер, К. (2005). Краеугольные виды. Получено 10 мая 2010 г. с сайта Холсингер К. (11 октября 2005 г.). «Ключевые виды» . Университет Коннектикута . Архивировано из оригинала 30 июня 2010 года . Проверено 12 мая 2010 г.
- ^ Симаков О., Кавасима Т., Марлетаз Ф., Дженкинс Дж., Коянаги Р., Митрос Т. и др. (ноябрь 2015 г.). «Гемихордовые геномы и происхождение вторичноротых» . Природа 527 (7579): 459–65. Бибкод : 2015Nature.527..459S . дои : 10.1038/nature16150 . ПМК 4729200 . ПМИД 26580012 .
- ^ «Как люди получили глотку от этого «уродливого зверя» » . Будущее . 23 ноября 2015 г.
- ^ Jump up to: а б с д Кларк М.А., Чой Дж., Дуглас М. (2018). «Хордовые» . Биология (2-е изд.). ОпенСтакс. ISBN 978-1-947172-95-1 . . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0.
- ^ «Секрет жизни в Оезии: доисторические черви строили трубчатые «дома» на морском дне» . физ.орг .
- ^ Барнс Р.Д. (1982). Зоология беспозвоночных . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 1018–1026. ISBN 978-0-03-056747-6 .
- ^ «Вторичные организаторы раннего мозга и расположение мезодиэнцефальных дофаминергических клеток-предшественников» . Карта жизни . Архивировано из оригинала 10 марта 2014 года . Проверено 10 марта 2014 г.
- ^ Пани А.М., Малларки Э.Э., Аронович Дж., Асимакопулос С., Гроув Э.А., Лоу С.Дж. (март 2012 г.). «Древнее вторичноротое происхождение сигнальных центров мозга позвоночных» . Природа . 483 (7389). НаукаЖизнь: 289–94. Бибкод : 2012Natur.483..289P . дои : 10.1038/nature10838 . ПМЦ 3719855 . ПМИД 22422262 .
- ^ «Хордовые» . ОпенСтакс . 9 мая 2019 года.
- ^ Гевин В. (2005). «Функциональная геномика расширяет биологический сюжет» . ПЛОС Биология . 3 (6): е219. дои : 10.1371/journal.pbio.0030219 . ПМЦ 1149496 . ПМИД 15941356 .
- ^ Тиммер Дж (19 июня 2008 г.). «Геном ланцетника (амфиоксуса) и происхождение позвоночных» . Арс Техника .
- ^ Лемэр П. (2011). «Эволюционный перекресток биологии развития: оболочники». Разработка . 138 (11): 2143–2152. дои : 10.1242/dev.048975 . ПМИД 21558365 . S2CID 40452112 .
- ^ «FishBase: Глобальная информационная система по рыбам» . ФишБаза . Проверено 17 января 2017 г.
- ^ «Сколько рыбы в море? Перепись морской жизни представляет первый отчет» . Наука Дейли . Проверено 17 января 2017 г.
- ^ Docker MF (ноябрь 2006 г.). «Вклад Билла Бимиша в исследования миноги и последние достижения в этой области» . Обзоры гвельфской ихтиологии . 7 .
- ^ Хардисти М.В., Поттер И.С. (1971). Хардисти М.В., Поттер И.С. (ред.). Биология миног (1-е изд.). Академическая пресса. ISBN 978-0-123-24801-5 .
- ^ Гилл Х.С., Рено С.Б., Шапло Ф., Мэйден Р.Л., Поттер И.С. (2003). «Филогения живых паразитических миног (Petromyzontiformes) на основе морфологических данных». Копейя . 2003 (4): 687–703. дои : 10.1643/IA02-085.1 . S2CID 85969032 .
- ^ «Миксини» . Музей палеонтологии Калифорнийского университета. Архивировано из оригинала 15 декабря 2017 года . Проверено 17 января 2017 г.
- ^ Грин С.А., Броннер МЭ (2014). «Минога: модельная система бесчелюстных позвоночных для изучения происхождения нервного гребня и других черт позвоночных» . Дифференциация; Исследования биологического разнообразия . 87 (1–2): 44–51. doi : 10.1016/j.diff.2014.02.001 . ПМЦ 3995830 . ПМИД 24560767 .
- ^ Stock DW, Whitt GS (август 1992 г.). «Свидетельства на основе последовательностей рибосомальной РНК 18S о том, что миноги и миксины образуют естественную группу». Наука . 257 (5071): 787–9. Бибкод : 1992Sci...257..787S . дои : 10.1126/science.1496398 . ПМИД 1496398 .
- ^ Николлс Х. (сентябрь 2009 г.). «Эволюция: Из уст в уста» . Природа . 461 (7261): 164–6. дои : 10.1038/461164a . ПМИД 19741680 .
- ^ Маккой В.Е., Саупе Э.Э., Ламсделл Дж.К., Тархан Л.Г., МакМахон С., Лидгард С. и др. (апрель 2016 г.). «Монстр Талли» — позвоночное животное» . Природа . 532 (7600): 496–9. Бибкод : 2016Natur.532..496M . дои : 10.1038/nature16992 . ПМИД 26982721 . S2CID 205247805 .
- ^ Саллан Л., Джайлз С., Сэнсом Р.С., Кларк Дж.Т., Йохансон З., Сэнсом И.Дж. и др. (20 февраля 2017 г.). «Монстр Талли» не является позвоночным: характеры, конвергенция и тафономия у проблемных животных палеозоя» (PDF) . Палеонтология . 60 (2): 149–157. Бибкод : 2017Palgy..60..149S . дои : 10.1111/пала.12282 .
- ^ Геггель Л. (4 мая 2020 г.). «Древний «монстр Талли» был позвоночным, а не бесхарактерной каплей, утверждает исследование» . Живая наука .
- ^ Маккой В.Е., Виманн Дж., Ламсделл Дж.К., Уэлен К.Д., Лидгард С., Майер П. и др. (сентябрь 2020 г.). «Химические признаки мягких тканей различают позвоночных и беспозвоночных из каменноугольного лагеря Мазон-Крик в штате Иллинойс». Геобиология . 18 (5): 560–565. Бибкод : 2020Gbio...18..560M . дои : 10.1111/gbi.12397 . ПМИД 32347003 . S2CID 216646333 .
- ^ Миками, Томоюки; Икеда, Такафуми; Мурамия, Юсуке; Хирасава, Тацуя; Ивасаки, Ватару (2023). Чернс, Лесли (ред.). «Трехмерная анатомия монстра Талли ставит под сомнение его предполагаемое сходство с позвоночными» . Палеонтология . 66 (2). дои : 10.1111/пала.12646 . ISSN 0031-0239 . S2CID 258198566 .
- ^ Киммел CB, Миллер CT, Кейнс Р.Дж. (2001). «Формирование нервного гребня и эволюция челюсти» . Журнал анатомии . 199 (Части 1–2): 105–20. дои : 10.1017/S0021878201008068 . ПМК 1594948 . ПМИД 11523812 .
- ^ Гай З, Чжу М (2012). «Происхождение челюсти позвоночных: пересечение модели, основанной на биологии развития, и ископаемых свидетельств» . Китайский научный бюллетень . 57 (30): 3819–3828. Бибкод : 2012ЧСБу..57.3819Г . дои : 10.1007/s11434-012-5372-z .
- ^ Мэйси Дж.Г. (2000). Обнаружение ископаемых рыб . Вествью Пресс. стр. 1–223. ISBN 978-0-8133-3807-1 .
- ^ Jump up to: а б Роу С., Хубер Д.Р., Лоури М., МакГенри С., Морено К., Клаузен П. и др. (2008). «Трехмерный компьютерный анализ механики челюстей белой акулы: насколько сильно может укусить большая белая акула?» (PDF) . Журнал зоологии . 276 (4): 336–342. дои : 10.1111/j.1469-7998.2008.00494.x .
- ^ Пимьенто С., Эрет DJ, Макфадден Б.Дж., Хаббелл Дж. (май 2010 г.). Степанова А (ред.). «Древний питомник вымершей гигантской акулы мегалодона из миоцена Панамы» . ПЛОС ОДИН . 5 (5): е10552. Бибкод : 2010PLoSO...510552P . дои : 10.1371/journal.pone.0010552 . ПМК 2866656 . ПМИД 20479893 .
- ^ Ламберт О, Бьянуччи Дж, Пост К, де Мюзон К, Салас-Жисмонди Р, Урбина М, Роймер Дж (июль 2010 г.). «Гигантский укус нового хищного кашалота из миоценовой эпохи Перу». Природа . 466 (7302): 105–8. Бибкод : 2010Natur.466..105L . дои : 10.1038/nature09067 . ПМИД 20596020 . S2CID 4369352 .
- ^ Нильсен Дж., Хедехольм Р.Б., Хайнемайер Дж., Бушнелл П.Г., Кристиансен Дж.С., Олсен Дж. и др. (август 2016 г.). «Радиоуглеродный анализ хрусталика глаза показывает многовековое долголетие гренландской акулы (Somniosus microcephalus)». Наука . 353 (6300): 702–4. Бибкод : 2016Sci...353..702N . doi : 10.1126/science.aaf1703 . hdl : 2022/26597 . ПМИД 27516602 . S2CID 206647043 .
- Энрико де Лазаро (12 августа 2016 г.). «Гренландские акулы — самые долгоживущие позвоночные на Земле, говорят морские биологи» . Новости науки .
- ^ Маршалл А., Беннетт М.Б., Коджа Г., Инохоса-Альварес С., Гальван-Магана Ф., Хардинг М. и др. (2011). " Одеяло Бирострис Красный список исчезающих видов МСОП . 2011 : e.T198921A9108067. doi : 10.2305/IUCN.UK.2011-2.RLTS.T198921A9108067.en .
- ^ Черный R (11 июня 2007 г.). «Защита пилы обретает зубы» . Новости Би-би-си .
- ^ Рядом с TJ, Эйтан Р.И., Дорнбург А., Кун К.Л., Мур Дж.А., Дэвис М.П. и др. (август 2012 г.). «Разрешение филогении лучепёрых рыб и сроки диверсификации» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (34): 13698–703. Бибкод : 2012PNAS..10913698N . дои : 10.1073/pnas.1206625109 . ПМК 3427055 . ПМИД 22869754 .
- ^ Jump up to: а б Чжу М, Чжао В, Цзя Л, Лу Дж, Цяо Т, Цюй Цюй (март 2009 г.). «Самый старый сочлененный остихтиан демонстрирует мозаичные признаки челюстноротых». Природа 458 (7237): 469–74. Бибкод : 2009Nature.458..469Z . дои : 10.1038/nature07855 . ПМИД 19325627 . S2CID 669711 .
- ^ Клак, JA (2002) Набирая силу . Университет Индианы
- ^ «Хондростеи: родственники осетровых» . палеос.com. Архивировано из оригинала 25 декабря 2010 года.
- ^ Лопес-Арбарелло А (2012). «Филогенетические взаимоотношения гинглимодовых рыб (Actinopterygii: Neopterygii)» . ПЛОС ОДИН . 7 (7): e39370. Бибкод : 2012PLoSO...739370L . дои : 10.1371/journal.pone.0039370 . ПМК 3394768 . ПМИД 22808031 .
- ^ Берра ТМ (2008). Распространение пресноводных рыб . Издательство Чикагского университета . п. 55. ИСБН 978-0-226-04443-9 .
- ^ Лакманн А.Р., Эндрюс А.Х., Батлер М.Г., Белак-Лакманн Э.С., Кларк М.Е. (23 мая 2019 г.). «Большеротый буйвол Ictiobus cyprinellus устанавливает рекорд пресноводной костистости, поскольку улучшенный возрастной анализ показывает столетнее долголетие» . Коммуникационная биология . 2 (1): 197. дои : 10.1038/s42003-019-0452-0 . ПМК 6533251 . ПМИД 31149641 .
- ^ Jump up to: а б Бентон М. (2005). «Эволюция рыб после девона» . Палеонтология позвоночных (3-е изд.). Джон Уайли и сыновья . стр. 175–84. ISBN 978-1-4051-4449-0 .
- ^ Боун Кью, Мур Р. (2008). Биология рыб . Гирляндная наука . п. 29. ISBN 978-0-415-37562-7 .
- ^ Дорит Р., Уокер В.Ф., Барнс Р.Д. (1991). Зоология . Издательство Колледжа Сондерса. стр. 67–69 . ISBN 978-0-03-030504-7 .
- ^ «Ученые описывают самую маленькую и легкую рыбу в мире» . Океанографический институт Скриппса . 20 июля 2004 г. Архивировано из оригинала 5 марта 2016 г. Проверено 9 апреля 2016 г.
- ^ Роуч Дж. (13 мая 2003 г.). «Обнаружена самая тяжелая костистая рыба в мире?» . Национальные географические новости . Архивировано из оригинала 17 мая 2003 года . Проверено 9 января 2016 г.
- ^ «Сводная статистика видов, находящихся под угрозой исчезновения во всем мире» (PDF) . Красный список исчезающих видов МСОП . Всемирный союз охраны природы. Осень 2014 г.
Таблица 1: Численность находящихся под угрозой исчезновения видов по основным группам организмов (1996–2014 гг.)
- ^ Наркевич К., Наркевич М. (январь 2015 г.). «Возраст старейших следов четвероногих из Зачелме, Польша». Летайя . 48 (1): 10–12. дои : 10.1111/лет.12083 . ISSN 0024-1164 .
- ^ Лонг Дж.А., Гордон М.С. (сентябрь – октябрь 2004 г.). «Величайший шаг в истории позвоночных: палеобиологический обзор перехода от рыб к четвероногим» (PDF) . Физиологическая и биохимическая зоология . 77 (5): 700–19. дои : 10.1086/425183 . ПМИД 15547790 . S2CID 1260442 .
- ^ Шубин Н. (2008). Ваша внутренняя рыба: путешествие в 3,5-миллиардную историю человеческого тела . Нью-Йорк: Книги Пантеона. ISBN 978-0-375-42447-2 .
- ^ Лаурин М (2010). Как позвоночные покинули воду . Беркли, Калифорния, США: Издательство Калифорнийского университета. ISBN 978-0-520-26647-6 .
- ^ Кановиль А., Лорин М. (2010). «Эволюция микроанатомии плечевой кости и образа жизни амниот и некоторые комментарии к палеобиологическим выводам» . Биологический журнал Линнеевского общества . 100 (2): 384–406. дои : 10.1111/j.1095-8312.2010.01431.x .
- ^ Лорин М. , Кановиль А., Кильяк А. (август 2009 г.). «Использование палеонтологических и молекулярных данных по супердеревьям для сравнительных исследований: на примере микроанатомии бедренной кости лисамфибий» . Журнал анатомии . 215 (2): 110–23. дои : 10.1111/j.1469-7580.2009.01104.x . ПМК 2740958 . ПМИД 19508493 .
- ^ Хопкинс Г.Р., Эд Броди-младший (2015). «Распространение земноводных в солончаковых средах обитания: обзор и эволюционная перспектива». Герпетологические монографии . 29 (1): 1–27. doi : 10.1655/HERPMONOGRAPHHS-D-14-00006 . S2CID 83659304 .
- ^ Натчев Н., Цанков Н., Гем Р. (2011). «Инвазия зеленых лягушек в Черное море: экология среды обитания популяции комплекса Pelophylax esculentus (Anura, Amphibia) в районе лагуны Шабленска Тузла в Болгарии» (PDF) . Герпетологические заметки . 4 : 347–351. Архивировано из оригинала (PDF) 24 сентября 2015 года . Проверено 11 августа 2016 г.
- ^ Сандер П.М. (август 2012 г.). «Палеонтология. Размножение ранних амниот». Наука . 337 (6096): 806–8. Бибкод : 2012Sci...337..806S . дои : 10.1126/science.1224301 . ПМИД 22904001 . S2CID 7041966 .
- ^ Модесто С.П., Андерсон Дж.С. (октябрь 2004 г.). «Филогенетическое определение рептилий» . Систематическая биология . 53 (5): 815–21. дои : 10.1080/10635150490503026 . ПМИД 15545258 .

- ^ Готье Дж.А., Клюге А.Г., Роу Т. (1988). «Ранняя эволюция амниоты». В Бентоне М.Дж. (ред.). Филогения и классификация четвероногих . Том. 1. Оксфорд: Кларендон Пресс. стр. 103–155. ISBN 978-0-19-857705-8 .
- ^ Лаурин М., Рейш Р.Р. (1995). «Переоценка ранней филогении амниот» (PDF) . Зоологический журнал Линнеевского общества . 113 (2): 165–223. дои : 10.1111/j.1096-3642.1995.tb00932.x . Архивировано из оригинала (PDF) 8 июня 2019 года . Проверено 14 августа 2016 г.
- ^ Модесто СП (1999). «Наблюдения за строением раннепермской рептилии Stereosternum tumidum Cope». Палеонтология Африканская . 35 : 7–19.
- ^ Расмуссен А.Р., Мерфи Дж.К., Омпи М., Гиббонс Дж.В., Уец П. (8 ноября 2011 г.). «Морские рептилии» . ПЛОС ОДИН . 6 (11): e27373. Бибкод : 2011PLoSO...627373R . дои : 10.1371/journal.pone.0027373 . ПМК 3210815 . ПМИД 22087300 .
- ^ Стидворти Дж (1974). Змеи мира . Grosset & Dunlap Inc. с. 160. ИСБН 978-0-448-11856-7 .
- ^ «Морские змеи» (PDF) . Продовольственная и сельскохозяйственная организация Объединенных Наций . Проверено 22 августа 2020 г.
- ^ Расмуссен А.Р., Мерфи Дж.К., Омпи М., Гиббонс Дж.В., Уетц П. (2011). «Морские рептилии» . ПЛОС ОДИН . 6 (11): e27373. Бибкод : 2011PLoSO...627373R . дои : 10.1371/journal.pone.0027373 . ПМК 3210815 . ПМИД 22087300 .
- ^ Мартилл Д.М. (1993). «Суповые субстраты: среда для исключительного сохранения ихтиозавров из сланцев Посидония (нижняя юра) Германии». Каупиа – Дармштадтский вклад в естественную историю , 2 : 77–97.
- ^ Гулд С.Дж. (1993). «Изогнутый из формы» . Восемь поросят: размышления о естествознании . Нортон. стр. 179–94. ISBN 978-0-393-31139-6 .
- ^ «Феномен безумного кормления акул сардинами в Африке» . Архивировано из оригинала 2 декабря 2008 года.
- ^ «Список видов и подвидов Комитета по таксономии Общества морских млекопитающих» . Общество морской маммологии. Октябрь 2015. Архивировано из оригинала 6 января 2015 года . Проверено 23 ноября 2015 г.
- ^ Ромер А.С., Парсонс Т.С. (1986). Тело позвоночного . Издательство колледжа Сандерса. п. 96. ИСБН 978-0-03-058446-6 .
- ^ «Синий кит» . Всемирный фонд природы . Проверено 15 августа 2016 г.
- ^ Марино Л. (2004). «Эволюция мозга китообразных: умножение порождает сложность» (PDF) . Международное общество сравнительной психологии (17): 1–16. Архивировано из оригинала (PDF) 16 сентября 2018 года . Проверено 15 августа 2016 г.
- ^ Jump up to: а б Кэмпбелл Н.А., Рис Дж.Б., Урри Л.А., Кейн М.Л., Вассерман С.А., Минорский П.В., Джексон Р.Б. (2008). Биология (8-е изд.). Сан-Франциско: Пирсон – Бенджамин Каммингс. ISBN 978-0-321-54325-7 .
- ^ Барри Ф.Р., Бак В.Р., Демулен В., Гройтер В., Хоксворт Д.Л., Херендин П.С. и др., ред. (2012). Международный кодекс номенклатуры водорослей, грибов и растений (Мельбурнский кодекс), принятый Восемнадцатым Международным ботаническим конгрессом в Мельбурне, Австралия, июль 2011 г. (электронное издание). Международная ассоциация систематики растений . Проверено 14 мая 2017 г.
- ^ Уолш П.Дж., Смит С., Флеминг Л., Соло-Габриэль Х., Гервик У.Х., ред. (2 сентября 2011 г.). «Цианобактерии и цианобактериальные токсины» . Океаны и здоровье человека: морские риски и средства правовой защиты . Академическая пресса. стр. 271–296. ISBN 978-0-08-087782-2 .
- ^ «Повышение уровня кислорода — журнал астробиологии» . Журнал «Астробиология» . 30 июля 2003 г. Проверено 6 апреля 2016 г.
- ^ Фланнери Д.Т., Уолтер Р.М. (2012). «Архейские тафтинговые микробные маты и Великое событие окисления: новый взгляд на древнюю проблему». Австралийский журнал наук о Земле . 59 (1): 1–11. Бибкод : 2012AuJES..59....1F . дои : 10.1080/08120099.2011.607849 . S2CID 53618061 .
- ^ Ротшильд Л. (сентябрь 2003 г.). «Понять эволюционные механизмы и экологические пределы жизни» . НАСА. Архивировано из оригинала 29 марта 2012 года . Проверено 13 июля 2009 г.
- ^ Надис С. (декабрь 2003 г.). «Клетки, управляющие морями» (PDF) . Научный американец . 289 (6): 52–3. Бибкод : 2003SciAm.289f..52N . doi : 10.1038/scientificamerican1203-52 . ПМИД 14631732 . Архивировано из оригинала (PDF) 19 апреля 2014 года . Проверено 2 июня 2019 г.
- ^ «Самый важный микроб, о котором вы никогда не слышали» . npr.org .
- ^ Фломбаум П., Гальегос Дж.Л., Гордилло Р.А., Корнер Дж., Шафран Л.Л., Цзяо Н. и др. (июнь 2013 г.). «Настоящее и будущее глобальное распространение морских цианобактерий Prochromococcus и Synechococcus» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (24): 9824–9. Бибкод : 2013PNAS..110.9824F . дои : 10.1073/pnas.1307701110 . ПМЦ 3683724 . ПМИД 23703908 .
- ^ Нэборс МВ (2004). Введение в ботанику . Сан-Франциско, Калифорния: ISBN Pearson Education, Inc. 978-0-8053-4416-5 .
- ^ Аллаби М., изд. (1992). «Водоросли». Краткий словарь ботаники . Оксфорд: Издательство Оксфордского университета.
- ^ Гири, доктор медицинских наук (октябрь 2012 г.). «Сколько существует видов водорослей?». Журнал психологии . 48 (5): 1057–63. дои : 10.1111/j.1529-8817.2012.01222.x . ПМИД 27011267 . S2CID 30911529 .
- ^ Jump up to: а б Гири, доктор медицинских наук, Гири, генеральный директор (2016). «Альгебаза» . www.algaebase.org . Проверено 20 ноября 2016 г. .
- ^ Томас Д. (2002). Морские водоросли . Серия «Жизнь». Музей естественной истории , Лондон. ISBN 978-0-565-09175-0 .
- ^ Хук С., Манн Д., Янс Х.М., Янс М. (1995). Водоросли: введение в психологию . Издательство Кембриджского университета. п. 166. ИСБН 9780521316873 . OCLC 443576944 .
- ^ Трегер П., Нельсон Д.М., Ван Беннеком А.Дж., Демастер DJ, Лейнаерт А., Кегинер Б. (апрель 1995 г.). «Баланс кремнезема в мировом океане: переоценка». Наука . 268 (5209): 375–9. Бибкод : 1995Sci...268..375T . дои : 10.1126/science.268.5209.375 . ПМИД 17746543 . S2CID 5672525 .
- ^ «Королевский колледж Лондона — озеро Мегачад» . www.kcl.ac.uk. Проверено 5 мая 2018 г.
- ^ Гомес Ф (2012). «Контрольный список и классификация живых динофлагеллят (Dinoflagellata, Alveolata)» . СИКИМАР Океаниды . 27 (1): 65–140. doi : 10.37543/oceanides.v27i1.111 .
- ^ Стокер Д.К. (1999). «Миксотрофия среди динофлагеллят». Журнал эукариотической микробиологии . 46 (4): 397–401. дои : 10.1111/j.1550-7408.1999.tb04619.x . S2CID 83885629 .
- ^ Старкс С. (31 октября 2012 г.). «Место под солнцем – водоросли – это урожай будущего, по мнению исследователей из Geel] Flanders Today» . Архивировано из оригинала 4 марта 2016 года . Проверено 8 декабря 2012 г.
- ^ Дюваль Б., Маргулис Л. (1995). «Микробное сообщество разносторонних колоний Ophrydium: эндосимбионты, резиденты и арендаторы». Симбиоз . 18 : 181–210. ПМИД 11539474 .
- ^ Вернберг Т., Крумхансл К., Филби-Декстер К. и Педерсен М.Ф. (2019) «Состояние и тенденции мировых лесов из водорослей». В: Мировые моря: экологическая оценка , стр. 57–78). Академическая пресса. два : 10.1016/B978-0-12-805052-1.00003-6 .
- ^ Манн К.Х. (декабрь 1973 г.). «Морские водоросли: их продуктивность и стратегия роста: роль крупных морских водорослей в продуктивности прибрежных районов гораздо важнее, чем предполагалось». Наука . 182 (4116): 975–81. Бибкод : 1973Sci...182..975M . дои : 10.1126/science.182.4116.975 . ПМИД 17833778 . S2CID 26764207 .
- ^ Таннелл Дж.В., Чавес Э.А., Уизерс К. (2007). Коралловые рифы южной части Мексиканского залива . Издательство Техасского университета A&M. п. 91. ИСБН 978-1-58544-617-9 .
- ^ « Каулерпа таксифолия (водоросли-убийцы)» . Справочник инвазивных видов . Международный центр сельского хозяйства и биологических наук. 6 ноября 2018 г.
- ^ Мандоли Д.Ф. (июнь 1998 г.). «Разработка плана тела и фазовый переход во время развития вертлужной впадины: как строится сложная архитектура гигантской одноклеточной клетки?». Ежегодный обзор физиологии растений и молекулярной биологии растений . 49 : 173–198. дои : 10.1146/annurev.arplant.49.1.173 . ПМИД 15012232 . S2CID 6241264 .
- ^ Мадл П., Йип М. (2004). «Обзор литературы Caulerpa Taxifolia» . БУФУС-Инфо . 19 (31). Архивировано из оригинала 8 октября 2022 года . Проверено 18 июля 2017 г.
- ^ Орт Р.Дж., Каррутерс Т.Дж., Деннисон В.К., Дуарте К.М., Фуркурин Дж.В., Хек К.Л. и др. (2006). «Глобальный кризис экосистем морских водорослей» . Бионаука . 56 (12): 987–996. doi : 10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2 . hdl : 10261/88476 . S2CID 4936412 .
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2009). « Phycodurus eques » в FishBase . Версия за июль 2009 года.
- ^ Гири С., Очиенг Э., Тисзен Л.Л., Чжу З., Сингх А., Лавленд Т. и др. (2011). «Состояние и распространение мангровых лесов мира по данным спутникового наблюдения Земли». Глобальная экология и биогеография . 20 (1): 154–159. дои : 10.1111/j.1466-8238.2010.00584.x .
- ^ Томас Н., Лукас Р., Бантинг П., Харди А., Розенквист А., Симард М. (2017). «Распространение и движущие силы глобального изменения мангровых лесов, 1996–2010 годы» . ПЛОС ОДИН . 12 (6): e0179302. Бибкод : 2017PLoSO..1279302T . дои : 10.1371/journal.pone.0179302 . ПМЦ 5464653 . ПМИД 28594908 .
- ^ Короткое FT, Фредерик Т. (2003). Мировой атлас морских трав . Беркли, Калифорния: Издательство Калифорнийского университета. п. 24. ISBN 978-0-520-24047-6 . Архивировано из оригинала 10 июля 2019 года . Проверено 10 июля 2019 г.
- ^ Спалдинг М (2010). Мировой атлас мангровых зарослей . Рутледж. дои : 10.4324/9781849776608 . ISBN 978-1-84977-660-8 .
- ^ Лалли С., Парсонс Т. (1993). Биологическая океанография: Введение . Баттерворт-Хайнеманн. ISBN 0-7506-3384-0 .
- ^ Линдси Р., Скотт М., Симмон Р. (2010). «Что такое фитопланктон» . Земная обсерватория НАСА .
- ^ Филд CB, Беренфельд М.Дж., Рандерсон Дж.Т., Фальковски П. (июль 1998 г.). «Первичная продукция биосферы: интеграция наземных и океанических компонентов» . Наука . 281 (5374): 237–40. Бибкод : 1998Sci...281..237F . дои : 10.1126/science.281.5374.237 . ПМИД 9657713 .
- ^ Рост Б, Рибеселл Ю (2004). «Кокколитофоры и биологический насос: реакция на изменения окружающей среды». Кокколитофоры: от молекулярных процессов к глобальному воздействию . Спрингер. стр. 99–125. ISBN 978-3-662-06278-4 .
- ^ Арсеньев Л., Симон Н., Риго-Жалабер Ф., Ле Галль Ф., Шаффрон С., Корр Е., Ком Е., Бигерд Э., Боду АС (2018). «Первые вирусы, заражающие морскую диатомовую водоросль Guinardia delicatula» . Границы микробиологии . 9 : 3235. дои : 10.3389/fmicb.2018.03235 . ПМК 6334475 . ПМИД 30687251 .
- ^ Вареа С., Арагон Х.Л., Баррио Р.А. (октябрь 1999 г.). «Узоры Тьюринга на сфере». Физический обзор E . 60 (4 части Б): 4588–92. Бибкод : 1999PhRvE..60.4588V . дои : 10.1103/PhysRevE.60.4588 . ПМИД 11970318 .
- ^ Харви Э.Н. (1952). Биолюминесценция . Академическая пресса.
- ^ «Предлагаемое объяснение светящихся морей, включая светящиеся в настоящее время калифорнийские моря» . Национальный научный фонд . 18 октября 2011 г.
- ^ Кастро П., Хубер М.Е. (2010). Морская биология (8-е изд.). МакГроу Хилл. стр. 95 . ISBN 978-0071113021 .
- ^ Гастингс Дж.В. (1996). «Химия и цвета биолюминесцентных реакций: обзор». Джин . 173 (1 номер спецификации): 5–11. дои : 10.1016/0378-1119(95)00676-1 . ПМИД 8707056 .
- ^ Хэддок С.Х., Молин М.А., Кейс Дж.Ф. (2009). «Биолюминесценция в море». Ежегодный обзор морской науки . 2 : 443–93. Бибкод : 2010ARMS....2..443H . doi : 10.1146/annurev-marine-120308-081028 . ПМИД 21141672 . S2CID 3872860 .
- ^ «Круговорот углерода и биосеквестрация» . Министерство энергетики США, Управление науки Министерства энергетики. 2008. с. 81.
Отчет семинара DOE/SC-108.
- ^ Кэмпбелл М. (22 июня 2011 г.). «Роль морского планктона в секвестрации углерода» . ЗемляТаймс . Проверено 22 августа 2014 г.
- ^ Роман Дж., Маккарти Джей-Джей (октябрь 2010 г.). «Китовый насос: морские млекопитающие повышают первичную продуктивность прибрежного бассейна» . ПЛОС ОДИН . 5 (10): е13255. Бибкод : 2010PLoSO...513255R . дои : 10.1371/journal.pone.0013255 . ПМЦ 2952594 . ПМИД 20949007 . е13255.
- ^ Браун Дж. Э. (12 октября 2010 г.). «Китовый экскремент улучшает здоровье океана» . Наука Дейли . Проверено 18 августа 2014 г.
- ^ «Вода – универсальный растворитель» . Геологическая служба США . Архивировано из оригинала 9 июля 2017 года . Проверено 27 июня 2017 г.
- ^ Брам-младший, Моррис Джей-Джей, Десима М., Стукель М.Р. (2014). Глава 2: Смертность в океанах: причины и последствия . Материалы симпозиума Eco-DAS IX. Ассоциация наук лимнологии и океанографии. стр. 16–48. ISBN 978-0-9845591-3-8 . .
- ^ Рис Дж. Б. (2013). Кэмпбелл Биология (10-е изд.). Пирсон . ISBN 978-0-321-77565-8 .
- ^ Прентис IC (2001). «Круговорот углерода и углекислый газ в атмосфере» . Изменение климата 2001: научная основа: вклад Рабочей группы I в Третий оценочный доклад Межправительственной группы экспертов по изменению климата / Хоутон, Дж. Т. [ред.] Проверено 31 мая 2012 г. .
- ^ Томсон CW (2014). Путешествие «Челленджера: Атлантика» . Издательство Кембриджского университета. п. 235. ИСБН 978-1-108-07475-9 .
- ^ Хасле Г.Р., Сиверцен Э.Э., Стейдингер, Танген К. (25 января 1996 г.). «Морские диатомеи» . В Томасе ЧР (ред.). Определение морских диатомей и динофлагеллят . Академическая пресса. стр. 5–385. ISBN 978-0-08-053441-1 . Проверено 13 ноября 2013 г.
- ^ Адл С.М., Леандер Б.С., Симпсон А.Г., Арчибальд Дж.М., Андерсон О.Р., Басс Д. и др. (август 2007 г.). «Разнообразие, номенклатура и систематика протистов» . Систематическая биология . 56 (4): 684–9. дои : 10.1080/10635150701494127 . ПМИД 17661235 .
- ^ Мохеймани Н.Р., Уэбб Дж.П., Боровицка М.А. (октябрь 2012 г.). «Биоремедиация и другие потенциальные применения кокколитофоридных водорослей: обзор». Водорослевые исследования . 1 (2): 120–33. дои : 10.1016/j.algal.2012.06.002 .
- ^ Тейлор А.Р., Кракри А., Уиллер Г., Годдард Х., Браунли С. (июнь 2011 г.). «Потенциал-зависимый канал H+, лежащий в основе гомеостаза pH в кальцинирующих кокколитофорах» . ПЛОС Биология . 9 (6): e1001085. дои : 10.1371/journal.pbio.1001085 . ПМК 3119654 . ПМИД 21713028 .
- ^ Халперн Б.С., Фрейзер М., Аффлербах Дж., Лаундс Дж.С., Мишели Ф., О'Хара С. и др. (август 2019 г.). «Недавние темпы изменения воздействия человека на мировой океан» . Научные отчеты . 9 (1): 11609. Бибкод : 2019NatSR...911609H . дои : 10.1038/s41598-019-47201-9 . ПМК 6691109 . ПМИД 31406130 .
- ^ «Воздействие человека на морские экосистемы» . ГЕОМАР Центр океанических исследований имени Гельмгольца . Архивировано из оригинала 31 октября 2020 года . Проверено 22 октября 2019 г.
- ^ Розинг М.Т., Бёрд Д.К., Sleep NH, Бьеррум С.Дж. (апрель 2010 г.). «Никакого климатического парадокса под слабым ранним Солнцем». Природа . 464 (7289): 744–7. Бибкод : 2010Natur.464..744R . дои : 10.1038/nature08955 . ПМИД 20360739 . S2CID 205220182 .
- ^ Jump up to: а б Сахни С., Бентон М.Дж., Ферри, Пенсильвания (август 2010 г.). «Связи между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–7. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ МакКинни М.Л. (1997). «Как редким видам удается избежать вымирания? Палеонтологический взгляд» . Биология редкости . стр. 110–29. дои : 10.1007/978-94-011-5874-9_7 . ISBN 978-94-010-6483-5 .
- ^ Стернс БП, Стернс СК (1999). Смотрю с края вымирания . Нью-Хейвен, Коннектикут: Издательство Йельского университета. п. х. ISBN 978-0-300-08469-6 .
- ^ Новачек М.Ю. (8 ноября 2014 г.). «Блестящее будущее предыстории» . Нью-Йорк Таймс . Нью-Йорк. ISSN 0362-4331 . Проверено 25 декабря 2014 г.
- ^ Ни С. (август 2004 г.). «Вымирание, слизь и низы» . ПЛОС Биология . 2 (8): Е272. дои : 10.1371/journal.pbio.0020272 . ПМК 509315 . ПМИД 15314670 .
- ^ Уорд ПД (октябрь 2006 г.). «Удар из глубины». Научный американец . 295 (4): 64–71. Бибкод : 2006SciAm.295d..64W . doi : 10.1038/scientificamerican1006-64 (неактивен 26 апреля 2024 г.). ПМИД 16989482 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка ) - ^ Морские вымирания: закономерности и процессы - обзор. 2013. Монография CIESM 45 [1]
- ^ Jump up to: а б с «Исследование морской жизни | Перепись морской жизни» . www.coml.org . Проверено 27 декабря 2023 г.
- ^ «Инструменты исследования: АНПА: Управление по исследованию и исследованию океана НОАА» . Oceanexplorer.noaa.gov . Проверено 27 декабря 2023 г.
- ^ «Буксируемые глубоководные аппараты (DTV) | Перепись морской жизни» . www.coml.org . Проверено 27 декабря 2023 г.
- ^ «Революция дронов под водой» . Расширенная навигация . Проверено 27 декабря 2023 г.
Дальнейшее чтение
[ редактировать ]- Халперн Б.С., Уолбридж С., Селкое К.А., Каппель К.В., Микели Ф., Д'Агроса С. и др. (февраль 2008 г.). «Глобальная карта воздействия человека на морские экосистемы». Наука . 319 (5865): 948–52. Бибкод : 2008Sci...319..948H . дои : 10.1126/science.1149345 . ПМИД 18276889 . S2CID 26206024 .
- Палечный М., Хэммилл Э., Карпузи В., Поли Д. (2015). «Тенденция численности наблюдаемых морских птиц в мире, 1950-2010 гг.» . ПЛОС ОДИН . 10 (6): e0129342. Бибкод : 2015PLoSO..1029342P . дои : 10.1371/journal.pone.0129342 . ПМЦ 4461279 . ПМИД 26058068 .
- Руперт Э.Э., Фокс Р.С., Барнс Р.Д. (2004). Зоология беспозвоночных (7-е изд.). Брукс / Коул. ISBN 978-0-03-025982-1 .
- «После 60 миллионов лет экстремального образа жизни морские птицы терпят крушение» . Хранитель . 22 сентября 2015 г.

