Современный синтез (20 век)
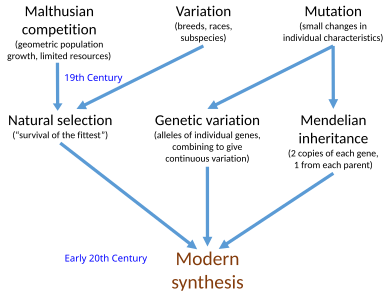
Современный синтез [ а ] представлял собой синтез в начале 20-го века Чарльза Дарвина теории эволюции и идей Грегора Менделя о наследственности в единую математическую структуру. Джулиан Хаксли ввёл этот термин в своей книге 1942 года « Эволюция: современный синтез» . Синтез объединил идеи естественного отбора , менделевской генетики и популяционной генетики . Это также связало широкомасштабную макроэволюцию, наблюдаемую палеонтологами, с мелкомасштабной микроэволюцией местных популяций.
Его основатели определяли синтез по-разному: Эрнст Майр в 1959 году, Г. Ледьярд Стеббинс в 1966 году и Теодосий Добжанский в 1974 году предложили разные основные постулаты, хотя все они включают естественный отбор, работающий над наследственными вариациями, возникающими в результате мутации. Другими важными фигурами в синтезе были Э. Б. Форд , Бернхард Ренш , Иван Шмальхаузен и Джордж Гейлорд Симпсон . Ранним событием в современном синтезе была Р. А. Фишера статья о математической популяционной генетике в 1918 году, хотя Уильям Бейтсон и отдельно Удни Юл уже начали показывать, как менделевская генетика может работать в эволюции, в 1902 году.
Затем последовали различные синтезы, в том числе социальное поведение в Э.О. Уилсона в социобиологии 1975 году, эволюционной биологии развития интеграция эмбриологии с генетикой и эволюцией в Массимо Пильуччи и Гердом Б. Мюллером , начиная с 1977 года, а также предложенный расширенный эволюционный синтез. 2007 г. По мнению биолога-эволюциониста Евгения Кунина , в 2009 г. на смену современному синтезу придет «постмодернистский» синтез, который будет включать революционные изменения в молекулярной биологии , изучении прокариот и возникшего в результате древа жизни , а также геномике . [ 3 ]
События, приведшие к синтезу
[ редактировать ]
Эволюция Дарвина путем естественного отбора, 1859 г.
[ редактировать ]Книга Чарльза Дарвина 1859 года «Происхождение видов » убедила большинство биологов в том, что эволюция имела место, но не в том, что естественный отбор был ее основным механизмом. В XIX и начале XX веков варианты ламаркизма (наследование приобретенных характеристик), ортогенеза (прогрессивная эволюция), сальтационизма (эволюция скачками) и мутационизма (эволюция, вызванная мутациями). в качестве альтернатив обсуждались [ 4 ] Сам Дарвин симпатизировал ламаркизму, но Альфред Рассел Уоллес выступал за естественный отбор и полностью отвергал ламаркизм. [ 5 ] В 1880 году Сэмюэл Батлер назвал точку зрения Уоллеса неодарвинизмом . [ 6 ] [ 7 ]

Затмение дарвинизма, 1880-е годы и далее
[ редактировать ]Начиная с 1880-х годов биологи стали скептически относиться к дарвиновской эволюции. Это затмение дарвинизма (по словам Джулиана Хаксли ) возникло из-за слабостей теории Дарвина относительно его взгляда на наследственность. Дарвин верил в смешанную наследственность , которая подразумевала, что любая новая вариация, даже если она полезна, будет ослабевать на 50% в каждом поколении, как заметил инженер Флиминг Дженкин в 1868 году. [ 8 ] [ 9 ] Это, в свою очередь, означало, что небольшие вариации не сохранялись достаточно долго, чтобы их можно было отобрать. Таким образом, смешение прямо противоречило бы естественному отбору. Кроме того, Дарвин и другие считали вполне возможным ламаркистское наследование приобретенных характеристик, а теория пангенезиса Дарвина 1868 года с вкладом в следующее поколение (геммулы), исходящими из всех частей тела, фактически подразумевала ламаркизм, а также смешение. [ 10 ] [ 11 ] [ 12 ]
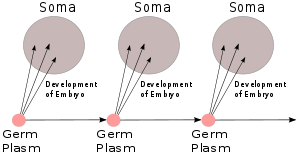
Зародышевая плазма Вейсмана, 1892 г.
[ редактировать ]Идея Августа Вейсмана , изложенная в его книге 1892 года «Зародышевая плазма: теория наследования» , [ 13 ] заключалось в том, что наследственный материал, который он называл зародышевой плазмой , и остальная часть тела ( сома ) имели одностороннюю связь: зародышевая плазма формировала тело, но тело не влияло на зародышевую плазму, за исключением косвенно в его участии в популяции, подверженной естественному отбору. Если это правда, то это сделало пангенезис Дарвина неверным, а наследование по Ламарку — невозможным. Его эксперимент на мышах, в котором он отрезал им хвосты и показал, что их потомство имело нормальные хвосты, продемонстрировал, что наследование является «трудным». [ б ] Он аргументировал сильно и догматично. [ 15 ] за дарвинизм и против ламаркизма, поляризуя мнения среди других ученых. Это усилило антидарвинистские настроения, способствуя его затмению. [ 16 ] [ 17 ]
Спорное начало
[ редактировать ]Генетика, мутационизм и биометрия, 1900–1918 гг.
[ редактировать ]
Проводя селекционные эксперименты с целью выяснения механизма наследования в 1900 году, Хьюго де Врис и Карл Корренс независимо друг от друга заново открыли работу Грегора Менделя . Новость об этом достигла Уильяма Бейтсона в Англии , который сообщил об этой статье во время презентации Королевскому садоводческому обществу в мае 1900 года. [ 18 ] При менделевском наследовании вклад каждого родителя сохраняет свою целостность, а не смешивается с вкладом другого родителя. В случае скрещивания двух настоящих селекционных сортов, таких как горох Менделя круглый и морщинистый, потомство первого поколения все одинаково, в данном случае во всех отношениях. При их скрещивании вновь проявляются первоначальные характеристики (сегрегация): около 3/4 их потомства округлые, 1/4 морщинистые. Существует разрыв между появлением потомства; де Фрис ввел термин «аллель» для обозначения варианта формы наследственной характеристики . [ 19 ] Это усилило основное разделение взглядов, существовавшее уже в 1890-х годах, между градуалистами, последовавшими за Дарвином, и сальтационистами, такими как Бейтсон. [ 20 ]
Этими двумя школами были менделисты, такие как Бейтсон и де Врис, которые выступали за мутационизм, эволюцию, движимую мутациями, основанную на генах, аллели которых разделялись дискретно, как горошины Менделя; [ 21 ] [ 22 ] и биометрическая школа , возглавляемая Карлом Пирсоном и Уолтером Уэлдоном . Биометристы энергично выступали против мутационизма, заявляя, что эмпирические данные указывают на то, что вариации у большинства организмов непрерывны, а не дискретны, как, казалось, предсказывал менделизм; они ошибочно полагали, что менделизм неизбежно подразумевает эволюцию прерывистыми скачками. [ 23 ] [ 24 ]

Традиционная точка зрения состоит в том, что биометристы и менделианцы отвергли естественный отбор и отстаивали свои отдельные теории в течение 20 лет, но споры разрешились только с развитием популяционной генетики. [ 23 ] [ 25 ] Более поздняя точка зрения состоит в том, что Бейтсон, де Врис, Томас Хант Морган и Реджинальд Паннетт к 1918 году сформировали синтез менделизма и мутационизма. Понимание, достигнутое этими генетиками, охватывало действие естественного отбора на аллели (альтернативные формы гена), равновесие Харди-Вайнберга , эволюцию постоянно меняющихся признаков (например, роста) и вероятность того, что новая мутация станет фиксированной. . С этой точки зрения ранние генетики признавали естественный отбор, но отвергали неменделевские идеи Дарвина об изменчивости и наследственности, и синтез начался вскоре после 1900 года. [ 26 ] [ 27 ] Традиционное утверждение, что менделисты отвергли идею непрерывных изменений, неверно; еще в 1902 году Бейтсон и Сондерс писали, что «если бы было хотя бы четыре или пять пар возможных аллеломорфов, различные гомо- и гетерозиготные комбинации могли бы при сериации дать столь близкое приближение к непрерывной кривой». , что о чистоте элементов можно было бы не подозревать». [ 28 ] Также в 1902 году статистик Удни Юл математически показал, что при наличии множества факторов теория Менделя допускает непрерывные изменения. Юл раскритиковал подход Бейтсона как конфронтационный. [ 29 ] но не смог предотвратить ссору между менделистами и биометристами. [ 30 ]
Крысы Касла, 1911 год.
[ редактировать ]Начиная с 1906 года Уильям Касл проводил длительное исследование влияния отбора на окраску шерсти у крыс . Пегий рецессивным или с капюшоном рисунок был по отношению к серому дикому типу. Он скрестил крыс с капюшоном как с дикими, так и с «ирландскими» типами, а затем обратно скрестил потомство с чистыми крысами с капюшоном. Темная полоса на спине была крупнее. Затем он попробовал отобрать разные группы для более крупных или меньших полос в течение 5 поколений и обнаружил, что можно значительно изменить характеристики, выходящие за пределы первоначального диапазона вариаций. Это эффективно опровергло утверждение де Фриза о том, что непрерывные изменения вызваны окружающей средой и не могут передаваться по наследству. К 1911 году Касл заметил, что результаты можно объяснить дарвиновским отбором наследственной вариации достаточного количества менделевских генов. [ 31 ] [ 32 ] [ 33 ]
Плодовые мухи Моргана, 1912 год.
[ редактировать ]Томас Хант Морган начал свою карьеру в области генетики как сальтационист и начал с попыток продемонстрировать, что мутации могут создавать новые виды у плодовых мух. Однако экспериментальная работа в его лаборатории с плодовой мушкой Drosophila melanogaster [ с ] показали, что вместо того, чтобы создавать новые виды за один этап, мутации увеличивают количество генетических вариаций в популяции. [ 34 ] К 1912 году, после многих лет работы над генетикой плодовых мух, Морган показал, что у этих насекомых имеется множество мелких менделевских факторов (обнаруженных как мухи-мутанты), над которыми могла работать дарвиновская эволюция, как если бы вариации были полностью непрерывными. Для генетиков была открыта возможность прийти к выводу, что менделизм поддерживает дарвинизм. [ 35 ]
Препятствие: позитивизм Вуджера, 1929 г.
[ редактировать ]Биолог -теоретик и философ биологии Джозеф Генри Вуджер возглавил внедрение позитивизма в биологию в своей книге «Биологические принципы» 1929 года . Он считал, что зрелая наука характеризуется структурой гипотез , которые можно проверить фактами, установленными экспериментами . Он критиковал традиционный естественноисторический стиль биологии , включая изучение эволюции , как незрелую науку, поскольку она опиралась на повествование . [ 36 ] Вуджер намеревался сыграть роль Роберта Бойля 1661 года химика-скептика , намереваясь превратить предмет биологии в формальную, единую науку и, в конечном итоге, следуя Венскому кружку логических позитивистов, таких как Отто Нейрат и Рудольф Карнап , свести биологию к минимуму. к физике и химии. Его усилия побудили биолога Дж. Б. С. Холдейна настаивать на аксиоматизации биологии и, оказав влияние на таких мыслителей, как Хаксли, помогли осуществить современный синтез. [ 36 ] Позитивистский климат сделал естествознание немодным, а в Америке к концу 1930-х годов научные исследования и преподавание эволюции на университетском уровне практически сошли на нет. Гарвардский физиолог Уильям Джон Крозье говорил своим студентам, что эволюция — это даже не наука: «Нельзя экспериментировать с двумя миллионами лет!» [ 37 ]
Волна мнений изменилась с появлением математического моделирования и контролируемых экспериментов в области популяционной генетики, объединивших генетику, экологию и эволюцию в рамках, приемлемых для позитивизма. [ 38 ]
Элементы синтеза
[ редактировать ]Математическая популяционная генетика Фишера и Холдейна, 1918–1930 гг.
[ редактировать ]В 1918 году Р. А. Фишер написал « Корреляцию между родственниками на основании предположения о менделевском наследовании ». [ 39 ] который показал, как непрерывные вариации могут происходить из ряда дискретных генетических локусов . В этой и других статьях, кульминацией которых стала его книга «Генетическая теория естественного отбора» 1930 года . [ 40 ] Фишер показал, насколько менделевская генетика согласуется с идеей эволюции путем естественного отбора. [ 41 ] [ д ]
В 1920-х годах в серии статей Дж . Б. С. Холдейн проанализировал реальные примеры естественного отбора, такие как эволюция промышленного меланизма у перцовой моли . [ 41 ] и показал, что естественный отбор может работать даже быстрее, чем предполагал Фишер. [ 43 ] И эти ученые, и другие, такие как Добжанский и Райт, хотели поднять биологию до стандартов физических наук, основывая ее на математическом моделировании и эмпирических проверках. Естественный отбор, который когда-то считался непроверяемым, становился предсказуемым, измеримым и проверяемым. [ 44 ]
Эмбриология Де Бира, 1930 г.
[ редактировать ]Традиционная точка зрения состоит в том, что биология развития сыграла небольшую роль в современном синтезе. [ 45 ] но в своей книге 1930 года «Эмбрионы и предки » эволюционный эмбриолог Гэвин де Бир предвосхитил эволюционную биологию развития. [ 46 ] показав, что эволюция может происходить гетерохронно , [ 47 ] например, при сохранении ювенильных черт у взрослых . [ 48 ] Это, как утверждал де Бир, могло вызвать внезапные изменения в летописи окаменелостей , поскольку эмбрионы окаменевают плохо. Поскольку пробелы в летописи окаменелостей использовались в качестве аргумента против постепенной эволюции Дарвина, объяснение де Бира поддержало дарвиновскую позицию. [ 49 ] Однако, несмотря на де Бира, современный синтез в значительной степени игнорировал эмбриональное развитие при объяснении формы организмов, поскольку популяционная генетика оказалась адекватным объяснением того, как развивались такие формы. [ 50 ] [ 51 ] [ и ]
Адаптивный ландшафт Райта, 1932 год.
[ редактировать ]
Популяционный генетик Сьюэлл Райт сосредоточил свое внимание на комбинациях генов, которые взаимодействуют как комплексы, и на влиянии инбридинга на небольшие относительно изолированные популяции, которые могут быть подвержены генетическому дрейфу . В статье 1932 года он представил концепцию адаптивного ландшафта , в которой такие явления, как скрещивание и генетический дрейф в небольших популяциях, могут оттолкнуть их от пиков адаптации, что, в свою очередь, позволит естественному отбору подтолкнуть их к новым пикам адаптации. [ 41 ] [ 53 ] Модель Райта могла бы понравиться полевым натуралистам, таким как Феодосий Добжанский и Эрнст Майр, которые начали осознавать важность географической изоляции в популяциях реального мира. [ 43 ] Работы Фишера, Холдейна и Райта помогли основать дисциплину теоретической популяционной генетики. [ 54 ] [ 55 ] [ 56 ]
Эволюционная генетика Добжанского, 1937 г.
[ редактировать ]
Феодосий Добжанский , иммигрант из Советского Союза в США , работавший в постдокторантуре лаборатории плодовых мух Моргана, был одним из первых, кто применил генетику к естественным популяциям. В основном он работал с Drosophila pseudoobscura . Он многозначительно говорит: «В России разнообразный климат от арктического до субтропического... Исключительно лабораторные работники, которые не обладают и не желают иметь никаких знаний о живых существах в природе, были и находятся в меньшинстве». [ 57 ] Неудивительно, что были и другие российские генетики с подобными идеями, хотя некоторое время их работы были известны лишь немногим на Западе . Его работа 1937 года «Генетика и происхождение видов». [ 58 ] стало ключевым шагом в преодолении разрыва между популяционными генетиками и полевыми натуралистами. В нем были представлены выводы, к которым пришли Фишер, Холдейн и особенно Райт в их высокоматематических статьях, в форме, доступной другим. [ 41 ] [ 43 ] Далее, Добжанский утверждал телесность, а значит, и биологическую реальность механизмов наследственности: эволюция основывалась на материальных генах, расположенных в виде цепочки на физических наследственных структурах — хромосомах — и более или менее прочно связанных друг с другом в зависимости от теории. их фактические физические расстояния на хромосомах. Как и Холдейн и Фишер, «эволюционная генетика» Добжанского [ 59 ] была настоящей наукой, объединяющей теперь клеточную биологию, генетику, а также микро- и макроэволюцию. [ 44 ] В его работе подчеркивалось, что реальные популяции обладают гораздо большей генетической изменчивостью, чем предполагали ранние популяционные генетики в своих моделях, и что важны генетически различные субпопуляции. Добжанский утверждал, что естественный отбор способствует сохранению генетического разнообразия, а также стимулирует изменения. На него повлияло знакомство в 1920-х годах с работами Сергея Четверикова , который изучал роль рецессивных генов в поддержании резервуара генетической изменчивости в популяции, прежде чем его работа была прекращена ростом лысенкоизма в Советском Союзе. Союз . [ 41 ] [ 43 ] К 1937 году Добжанский смог доказать, что мутации были основным источником эволюционных изменений и изменчивости, наряду с перестройками хромосом, влиянием генов на соседей в процессе развития и полиплоидией. Далее, генетический дрейф (он использовал этот термин в 1941 году), отбор, миграция и географическая изоляция могут изменить частоты генов. В-третьих, такие механизмы, как экологическая или сексуальная изоляция и гибридное бесплодие, могут исправить результаты более ранних процессов. [ 60 ]
Экологическая генетика Форда, 1940 г.
[ редактировать ]
Э. Б. Форд был натуралистом-экспериментатором, который хотел проверить естественный отбор в природе, фактически изобретая область экологической генетики . [ 61 ] Его работа по естественному отбору в диких популяциях бабочек и мотыльков впервые показала правильность предсказаний Р. А. Фишера. В 1940 году он первым описал и определил генетический полиморфизм и предсказал, что полиморфизм групп крови человека может поддерживаться в популяции, обеспечивая некоторую защиту от болезней. [ 61 ] [ 62 ] Его книга 1949 года «Менделизм и эволюция». [ 63 ] помог убедить Добжанского сменить акцент в третьем издании его знаменитого учебника «Генетика и происхождение видов» с дрейфа на селекцию. [ 64 ]
Стабилизирующий отбор Шмальгаузена, 1941 г.
[ редактировать ]Иван Шмальгаузен разработал теорию стабилизирующего отбора , идею о том, что отбор может сохранять признак при некоторой ценности, опубликовав в 1941 году статью на русском языке под названием «Стабилизирующий отбор и его место среди факторов эволюции» и монографию « Факторы эволюции: теория Стабилизирующий отбор [ 65 ] в 1945 году. Он развил ее на основе концепции Дж. М. Болдуина 1902 года о том, что изменения, вызванные окружающей средой, в конечном итоге будут заменены наследственными изменениями (включая эффект Болдуина на поведение), что следует из последствий этой теории для их дарвиновского вывода и приводит его в конфликт с лысенкоизмом. Шмальгаузен заметил, что стабилизирующий отбор устранит большинство отклонений от нормы, причем большинство мутаций будут вредными. [ 66 ] [ 67 ] [ 68 ] Добжанский назвал эту работу «важным недостающим звеном в современном взгляде на эволюцию». [ 69 ]
Популяризирующий синтез Хаксли, 1942 г.
[ редактировать ]
В 1942 году Джулиана Хаксли серьезная, но популяризирующая книга [ 70 ] [ 71 ] Эволюция: современный синтез [ 2 ] ввел название для синтеза и намеренно намеревался продвигать «синтетическую точку зрения» на эволюционный процесс. Он представлял себе широкий синтез многих наук: генетики, физиологии развития, экологии, систематики, палеонтологии, цитологии, математического анализа биологии, и предполагал, что эволюция будет протекать неодинаково в разных группах организмов в зависимости от того, как организован их генетический материал и каковы их особенности. стратегии воспроизводства, ведущие к прогрессивным, но различным эволюционным тенденциям. [ 71 ] Его видение было «эволюционным гуманизмом». [ 72 ] с системой этики и значимым местом «Человека» в мире, основанным на единой теории эволюции, которая продемонстрирует прогресс, ведущий к человечеству на его вершине. Естественный отбор, по его мнению, был «фактом природы, который можно проверить путем наблюдения и эксперимента», в то время как «период синтеза» 1920-х и 1930-х годов сформировал «более единую науку». [ 72 ] конкурируя с физикой и способствуя «возрождению дарвинизма». [ 72 ]
Однако книга не была тем исследовательским текстом, которым она казалась. По мнению философа науки Майкла Рьюза и по собственному мнению Хаксли, Хаксли был «универсалом, синтезатором идей, а не специалистом». [ 70 ] Рьюз отмечает, что Хаксли писал так, как если бы он добавлял эмпирические данные к математической схеме, установленной Фишером и популяционными генетиками, но это было не так. Хаксли избегал математики, например, даже не упоминая фундаментальную теорему Фишера о естественном отборе . Вместо этого Хаксли использовал массу примеров, чтобы продемонстрировать, что естественный отбор силен и что он действует на менделевские гены. Книга преуспела в достижении своей цели — убедить читателей в реальности эволюции, эффективно иллюстрируя такие темы, как биогеография островов , видообразование и конкуренция. Хаксли далее показал, что появление долгосрочных ортогенетических тенденций – предсказуемых направлений эволюции – в летописи окаменелостей можно легко объяснить аллометрическим ростом (поскольку части взаимосвязаны). Тем не менее, Хаксли не отвергал ортогенез сразу, а всю свою жизнь сохранял веру в прогресс, рассматривая Homo sapiens в качестве конечной точки, и с 1912 года он находился под влиянием виталист философ- Анри Бергсон , хотя публично он придерживался атеистической позиции в отношении эволюции. [ 70 ] Веру Хаксли в прогресс эволюции и эволюционный гуманизм разделяли в различных формах Добжанский, Майр, Симпсон и Стеббинс, причем все они писали о «будущем человечества». И Хаксли, и Добжанский восхищались священником-палеонтологом Пьером Тейяром де Шарденом , Хаксли написал введение к книге Тейяра об ортогенезе 1955 года « Феномен человека» . Это видение требовало, чтобы эволюция рассматривалась как центральный и руководящий принцип биологии. [ 72 ]
Аллопатрическое видообразование Майра, 1942 г.
[ редактировать ]
Ключевым вкладом Эрнста Майра в синтез стала книга «Систематика и происхождение видов» , опубликованная в 1942 году. [ 73 ] Он утверждал важность и стремился объяснить изменчивость популяций в эволюционных процессах, включая видообразование. В частности, он проанализировал влияние политипических видов, географической изменчивости и изоляции географическими и другими способами. [ 74 ] Майр подчеркнул важность аллопатрического видообразования , при котором географически изолированные субпопуляции расходятся настолько, что возникает репродуктивная изоляция . Он скептически относился к реальности симпатрического видообразования , полагая, что географическая изоляция является предпосылкой для создания внутренних (репродуктивных) изолирующих механизмов. Майр также представил концепцию биологического вида , которая определяла вид как группу скрещивающихся или потенциально скрещивающихся популяций, репродуктивно изолированных от всех других популяций. [ 41 ] [ 43 ] [ 75 ] [ 76 ] Прежде чем уехать из Германии в Соединенные Штаты в 1930 году, Майр находился под влиянием работ немецкого биолога Бернхарда Ренша , который в 1920-х годах анализировал географическое распространение политипических видов, уделяя особое внимание тому, как различия между популяциями коррелируют с такими факторами, как как различия в климате. [ 77 ] [ 78 ] [ 79 ]

Палеонтология Симпсона, 1944 г.
[ редактировать ]Джордж Гейлорд Симпсон продемонстрировал совместимость современного синтеза с палеонтологией в своей книге 1944 года «Темп и модус эволюции» . Работа Симпсона имела решающее значение, поскольку многие палеонтологи не соглашались, в некоторых случаях решительно, с идеей о том, что естественный отбор является основным механизмом эволюции. Оно показало, что тенденции линейного прогресса (например, в эволюции лошади ), которые ранее палеонтологи использовали в качестве подтверждения неоламаркизма и ортогенеза, не выдерживают тщательного изучения. Вместо этого летопись окаменелостей соответствовала нерегулярной, разветвленной и ненаправленной структуре, предсказанной современным синтезом. [ 41 ] [ 43 ]
Общество изучения эволюции, 1946 г.
[ редактировать ]Во время Второй мировой войны Майр редактировал серию бюллетеней Комитета по общим проблемам генетики, палеонтологии и систематики, образованного в 1943 году, в которых сообщалось об обсуждениях «синтетической атаки» на междисциплинарные проблемы эволюции. В 1946 году комитет стал Обществом изучения эволюции, первыми подписавшими его стали Майр, Добжанский и Сьюэлл Райт. Майр стал редактором журнала Evolution . С точки зрения Майра и Добжанского, предполагает историк науки Бетти Смоковитис, дарвинизм возродился, эволюционная биология была узаконена, а генетика и эволюция были синтезированы в новую единую науку. Все укладывалось в новые рамки, за исключением «еретиков», таких как Ричард Гольдшмидт , которые раздражали Майра и Добжанского, настаивая на возможности видообразования путем макромутации , создавая «обнадеживающих монстров». Результатом стали «ожесточенные споры». [ 52 ]

Ботаника Стеббинса, 1950 г.
[ редактировать ]Ботаник Г. Ледьярд Стеббинс расширил синтез, включив в него ботанику . Он описал важные эффекты гибридизации и полиплоидии растений на « видообразование Вариации в своей книге 1950 года и эволюция растений» . Это позволяло эволюции время от времени протекать быстро, причем полиплоидия, в частности, очевидно, могла мгновенно создавать новые виды. [ 41 ] [ 80 ]
Определения учредителей
[ редактировать ]Современный синтез определялся разными его основателями по-разному, с разным количеством основных постулатов, как показано в таблице.
| Компонент | Майр 1959 г. | Стеббинс , 1966 год. | Добжанский , 1974 г. |
|---|---|---|---|
| Мутация | (1) Случайность во всех событиях, приводящих к появлению новых генотипов, например, мутации. [ 81 ] | (1) источник изменчивости, но не направления [ 82 ] | (1) дает генетическое сырье [ 83 ] |
| Рекомбинация | (1) Случайность в рекомбинации, оплодотворении [ 81 ] | (2) источник изменчивости, но не направления [ 82 ] | |
| Хромосомная организация | (3) влияет на генетическую связь, организует вариации в генофонде [ 82 ] | ||
| Естественный отбор | (2) является лишь фактором, задающим направление, [ 81 ] [ 84 ] как видно из адаптации к физической и биотической среде [ 81 ] | (4) направляет изменения в генофонд [ 82 ] | (2) конструирует эволюционные изменения из генетического сырья [ 83 ] |
| Репродуктивная изоляция | (5) ограничивает направление, в котором отбор может направлять популяцию [ 82 ] | (3) делает дивергенцию необратимой у половых организмов [ 83 ] |
После синтеза
[ редактировать ]После синтеза эволюционная биология продолжала развиваться благодаря большому вкладу таких ученых, как У. Д. Гамильтон, [ 85 ] Джордж К. Уильямс, [ 86 ] Э.О. Уилсон, [ 87 ] Эдвард Б. Льюис [ 88 ] и другие.
Инклюзивный фитнес Гамильтона, 1964 год.
[ редактировать ]В 1964 году У. Д. Гамильтон опубликовал две статьи на тему «Генетическая эволюция социального поведения». Они определили инклюзивную приспособленность как количество эквивалентов потомства, которое индивидуум выращивает, спасает или иным образом поддерживает своим поведением. Этому противопоставлялась личная репродуктивная способность, то есть количество потомков, которые человек производит непосредственно. Гамильтон и другие, такие как Джон Мейнард Смит , утверждали, что успех гена состоит в максимизации числа его копий либо путем их рождения, либо косвенно поощряя зачатие от родственных особей, которые разделяют этот ген, — теория родственного отбора . [ 85 ] [ 89 ]
Геноцентрированная эволюция Уильямса, 1966 г.
[ редактировать ]В 1966 году Джордж К. Уильямс опубликовал «Адаптацию и естественный отбор» , изложив геноцентрированный взгляд на эволюцию, следуя концепциям Гамильтона, оспаривая идею эволюционного прогресса и критикуя широко распространенную в то время теорию группового отбора . Уильямс утверждал, что естественный отбор действует путем изменения частоты аллелей и не может работать на уровне групп. [ 90 ] [ 86 ] Эволюция, ориентированная на гены, была популяризирована Ричардом Докинзом в его книге «Эгоистичный ген» 1976 года и развита в его более технических трудах. [ 91 ] [ 92 ]
Социобиология Уилсона, 1975 г.
[ редактировать ]
В 1975 году Э.О. Уилсон опубликовал свою противоречивую работу. [ 93 ] книга «Социобиология: Новый синтез» , подзаголовок намекает на современный синтез [ 87 ] поскольку он пытался привнести изучение животного общества в эволюционный процесс. Это казалось радикально новым, хотя Вильсон следовал Дарвину, Фишеру, Докинзу и другим. [ 87 ] Критики, такие как Герхард Ленски, отмечали, что он следовал подходу Хаксли, Симпсона и Добжанского, который Ленски считал излишне упрощенным с точки зрения человеческого общества. [ 94 ] К 2000 году предложенная дисциплина социобиология превратилась в относительно общепринятую дисциплину эволюционной психологии . [ 87 ]
Гомеозисные гены Льюиса, 1978 г.
[ редактировать ]
В 1977 году технология рекомбинантной ДНК позволила биологам начать изучать генетический контроль развития. Развитие эволюционной биологии развития с 1978 года, когда Эдвард Б. Льюис открыл гомеозисные гены, показало, что многие так называемые гены-инструментарии регулируют развитие, влияя на экспрессию других генов. Также выяснилось, что некоторые регуляторные гены чрезвычайно древние, поэтому такие разные животные, как насекомые и млекопитающие, имеют общие механизмы контроля; например, ген Pax6 участвует в формировании глаз мышей и плодовых мух. Такая глубокая гомология предоставила убедительные доказательства эволюции и указала пути, по которым прошла эволюция. [ 88 ]
Более поздние синтезы
[ редактировать ]В 1982 году вышла историческая заметка к серии книг по эволюционной биологии. [ ж ] мог бы без оговорок заявить, что эволюция является центральным организующим принципом биологии. Смоковитис прокомментировал это так: «То, над чем работали архитекторы синтеза, к 1982 году стало фактом», добавив в сноске, что «центральная роль эволюции, таким образом, стала неявным знанием , частью общепринятой мудрости человечества». профессия». [ 95 ]
Однако к концу 20-го века современный синтез показал свой возраст, и с разных сторон были предложены новые синтезы для исправления его недостатков и заполнения пробелов. Они включали такие разнообразные области, как изучение общества , [ 87 ] биология развития, [ 50 ] эпигенетика, [ 96 ] молекулярная биология , микробиология , геномика , [ 3 ] симбиогенез и горизонтальный перенос генов . [ 97 ] Физиолог Дени Ноубл утверждает, что эти дополнения делают неодарвинизм в смысле современного синтеза начала 20-го века «по крайней мере, неполным как теорию эволюции». [ 97 ] и тот, который был сфальсифицирован более поздними биологическими исследованиями. [ 97 ]
Майкл Роуз и Тодд Окли отмечают, что эволюционная биология, ранее разделенная и « балканизированная », была объединена геномикой. По их мнению, он отверг по крайней мере пять общих предположений современного синтеза, а именно, что геном всегда представляет собой хорошо организованный набор генов; что каждый ген выполняет одну функцию; эти виды биохимически хорошо адаптированы к своим экологическим нишам; что виды являются устойчивыми единицами эволюции, и для них характерны все уровни от организма до органа, клетки и молекулы внутри вида; и что конструкция каждого организма и клетки эффективна. Они утверждают, что «новая биология» объединяет геномику, биоинформатику и эволюционную генетику в универсальный инструментарий для «постмодернистского синтеза». [ 54 ]
Расширенный эволюционный синтез Пильуччи, 2007 г.
[ редактировать ]В 2007 году, более чем через полвека после современного синтеза, Массимо Пильуччи призвал к расширенному эволюционному синтезу , чтобы включить аспекты биологии, которые не были включены или не существовали в середине 20 века. [ 98 ] [ 99 ] Он пересматривает относительную важность различных факторов, бросает вызов предположениям, сделанным в современном синтезе, и добавляет новые факторы. [ 99 ] [ 100 ] такие как многоуровневый отбор , трансгенерационная эпигенетическая наследственность , построение ниш и способность к развитию . [ 101 ] [ 96 ] [ 102 ]
«Постмодернистский» эволюционный синтез Кунина, 2009 г.
[ редактировать ]В 2009 году исполнилось 200 лет со дня рождения Дарвина, 150-летие « Происхождения видов » и 200-летие «раннего эволюционного синтеза» Ламарка. [ 3 ] Philosophie Zoologique , биолог-эволюционист Юджин Кунин заявил, что, хотя «здание Современного Синтеза [начала 20-го века] рухнуло, по-видимому, без возможности восстановления», [ 3 ] можно увидеть новый синтез XXI века. Он утверждал, что в эволюционной биологии произошли три взаимосвязанные революции: молекулярная, микробиологическая и геномная. Молекулярная революция включала в себя нейтральную теорию , согласно которой большинство мутаций нейтральны и что отрицательный отбор происходит чаще, чем положительный , и что вся нынешняя жизнь произошла от одного общего предка . В микробиологии синтез распространился на прокариотов , используя рибосомальную РНК для формирования древа жизни . Наконец, геномика объединила молекулярный и микробиологический синтез — в частности, горизонтальный перенос генов между бактериями показывает, что прокариоты могут свободно обмениваться генами. Многие из этих положений уже были высказаны другими исследователями, такими как Ульрих Кучера и Карл Дж. Никлас . [ 103 ]
На пути к замещающему синтезу
[ редактировать ]
Биологи, наряду с исследователями истории и философии биологии, продолжают обсуждать необходимость и возможную природу замещающего синтеза. Например, в 2017 году Филипп Хьюнеман и Денис М. Уолш заявили в своей книге « Вызов современному синтезу» , что многие теоретики указывали на то, что такие дисциплины, как теория эмбриологического развития, морфология и экология, были опущены. Они отметили, что все подобные аргументы представляют собой продолжающееся желание заменить современный синтез синтезом, который объединит «все биологические области исследований, связанные с эволюцией, адаптацией и разнообразием, в единой теоретической системе». [ 104 ] Далее они отметили, что существует две группы проблем в том, как современный синтез рассматривает наследование. Во-первых, другие способы, такие как эпигенетическое наследование , фенотипическая пластичность , эффект Болдуина и материнский эффект , позволяют возникать и передаваться новым характеристикам, а генам позже догонять новые адаптации. Во-вторых, все подобные механизмы являются частью не системы наследования, а системы развития : фундаментальной единицей является не отдельный эгоистично конкурирующий ген, а сотрудничающая система, которая работает на всех уровнях: от генов и клеток до организмов и культур. направлять эволюцию. [ 105 ] Молекулярный биолог Шон Б. Кэрролл отметил, что, если бы Хаксли имел доступ к эволюционной биологии развития , «эмбриология была бы краеугольным камнем его современного синтеза, и поэтому эво-дево сегодня является ключевым элементом более полного и расширенного эволюционного синтеза. " [ 106 ]
Историография
[ редактировать ]Оглядываясь назад на противоречивые описания современного синтеза, историк Бетти Смоковитис в своей книге 1996 года « Объединяющая биология: эволюционный синтез и эволюционная биология» отмечает , что и историки, и философы биологии пытались понять его научное значение, но нашли его «неправильным». движущаяся цель»; [ 107 ] единственное, в чем они согласились, это то, что это было историческое событие. [ 107 ] По ее словам
«К концу 1980-х годов была признана дурная слава эволюционного синтеза... «синтез» стал настолько печально известным, что немногие серьезные исторически мыслящие аналитики затронули эту тему, не говоря уже о том, чтобы знать, с чего начать разбираться в оставшейся после себя интерпретационной путанице. многочисленными критиками и комментаторами». [ 108 ]
См. также
[ редактировать ]- Теория систем развития
- Геноцентрированный взгляд на эволюцию
- История эволюционной мысли
- Неодарвинизм
- Возражения против эволюции
Примечания
[ редактировать ]- ↑ Также известный как Новый синтез , Современный эволюционный синтез , Эволюционный синтез и неодарвиновский синтез . Эти альтернативные термины неоднозначны, поскольку они, возможно, могут включать более поздние синтезы, поэтому в этой статье используется «современный синтез» Джулиана Хаксли 1942 года. [ 2 ] через.
- ↑ Питер Готье, однако, утверждал, что эксперимент Вейсмана показал только то, что травма не повлияла на зародышевую плазму. Он не проверял эффект ламаркистского использования и неупотребления. [ 14 ]
- ^ Работа Моргана с плодовыми мушками помогла установить связь между менделевской генетикой и хромосомной теорией наследственности, согласно которой наследственный материал воплощался в этих телах внутри клеточного ядра. [ 34 ]
- ↑ Фишер также проанализировал половой отбор в своей книге, но его работа была в значительной степени проигнорирована, а аргументы Дарвина в отношении такого отбора были неправильно поняты, поэтому они не составляли существенной части современного синтеза. [ 42 ]
- ^ Хотя Ч. Х. Уоддингтон в своей статье 1953 года «Эпигенетика и эволюция» призывал эмбриологию . добавить к синтезу [ 52 ]
- ↑ В переиздании книги Добжанского «Генетика и происхождение видов» .
Ссылки
[ редактировать ]- ^ Гулд 2002 , с. 216
- ^ Перейти обратно: а б Хаксли 2010 .
- ^ Перейти обратно: а б с д Кунин, Евгений В. (ноябрь 2009 г.). «Происхождение в 150 лет: виден ли новый эволюционный синтез?» . Тенденции в генетике . 25 (11): 473–475. дои : 10.1016/j.tig.2009.09.007 . ПМК 2784144 . ПМИД 19836100 .
- ^ Боулер 2003 , стр. 236–256.
- ^ Кучера, Ульрих (декабрь 2003 г.). «Сравнительный анализ статей Дарвина-Уоллеса и развитие концепции естественного отбора». Теория в биологических науках . 122 (4): 343–359. дои : 10.1007/s12064-003-0063-6 . S2CID 24297627 .
- ^ Батлер, Сэмюэл (1880). Бессознательная память . Дэвид Бог. п. 280.
Я могу предсказать с некоторой уверенностью, что вскоре мы найдем первоначальный дарвинизм доктора Эразма Дарвина … общепринятым вместо сегодняшнего неодарвинизма, и что вариации, накопление которых приводит к появлению видов, будут признаны обусловленными желаниям и стремлениям живых форм, в которых они появляются, а не приписаны случайности или, другими словами, неизвестным причинам, как в системе г-на Чарльза Дарвина.
- ^ Беккалони, Джордж (2013). «О терминах «дарвинизм» и «неодарвинизм» » . Веб-сайт AR Wallace . Архивировано из оригинала 11 апреля 2019 г. Проверено 14 сентября 2017 г.
- ^ Боулер 2003 , стр. 196–253.
- ^ Ларсон 2004 , стр. 105–129.
- ^ Гайон, Жан (1998). Борьба дарвинизма за выживание: наследственность и гипотеза естественного отбора . Издательство Кембриджского университета. стр. 2–3. ISBN 978-0-521-56250-8 .
- ^ Дарвин, Чарльз (1868). Изменение животных и растений при одомашнивании . Джон Мюррей. ISBN 978-1-4191-8660-8 .
- ^ Холтерхофф, Кейт (2014). «История и восприятие гипотезы пангенезиса Чарльза Дарвина». Журнал истории биологии . 47 (4): 661–695. дои : 10.1007/s10739-014-9377-0 . ПМИД 24570302 . S2CID 207150548 .
- ^ Вейсман, август (1892 г.). теория наследственности Зародышевая плазма: . Йена: Рыбак.
- ^ Готье, Питер (март – май 1990 г.). «Является ли эксперимент Вейсмана опровержением гипотезы Ламарка?». БИОС . 61 (1/2): 6–8. JSTOR 4608123 .
- ^ Боулер 1989 , стр. 248.
- ^ Боулер 2003 , стр. 253–256.
- ^ Боулер 1989 , стр. 247–253, 257.
- ^ Эмброуз, Майк. «Горох Менделя» . Норидж, Великобритания: Отдел ресурсов зародышевой плазмы, Центр Джона Иннеса . Архивировано из оригинала 14 июня 2016 года . Проверено 14 декабря 2017 г.
- ^ «Обзор законов Менделя» . Пирсон . Проверено 14 октября 2017 г.
- ^ Бейтсон 1894 : Мутации (как «спорт») и полиморфизмы были хорошо известны задолго до менделевского восстановления.
- ^ Ларсон 2004 , стр. 157–166.
- ^ Боулер 1989 , стр. 275–276.
- ^ Перейти обратно: а б Графен и Ридли 2006 , с. 69
- ^ Провайн 2001 , стр. 69.
- ^ Олби, Роберт (сентябрь 1989 г.). «Размеры научных противоречий: биометрически-менделевские дебаты». Британский журнал истории науки . 22 (3): 299–320. дои : 10.1017/s0007087400026170 . JSTOR 4026898 . ПМИД 11621982 . S2CID 36719974 .
- ^ Гайон, Дж. (1988). Борьба дарвинизма за выживание: наследственность и гипотеза естественного отбора . Издательство Кембриджского университета.
- ^ Штольцфус, Арлин; Кейбл, Келе (2014). «Менделийский мутационизм: забытый эволюционный синтез» (PDF) . Журнал истории биологии . 47 (4): 501–546. дои : 10.1007/s10739-014-9383-2 . ПМИД 24811736 . S2CID 23263558 . Архивировано (PDF) из оригинала 30 ноября 2014 г.
- ^ Бейтсон, Уильям ; Сондерс, ER (1902). «Экспериментальные исследования по физиологии наследственности» . Королевское общество. Отчеты перед комитетом по эволюции . Архивировано из оригинала 23 апреля 2020 г. Проверено 20 августа 2017 г.
- ^ Юле, Г. Удный (1902). «Законы Менделя и их вероятная связь с межрасовой наследственностью» . Новый фитолог . 1 (10): 226–227. дои : 10.1111/j.1469-8137.1902.tb07336.x .
- ^ Провайн 2001 , стр. 81–82.
- ^ Провайн 2001 , стр. 109–114.
- ^ Замок, МЫ (7 апреля 1905 г.). «Мутационная теория органической эволюции с точки зрения селекции животных» . Наука . 21 (536): 521–525. Бибкод : 1905Sci....21..521C . дои : 10.1126/science.21.536.521 . ISSN 0036-8075 . ПМИД 17770959 .
- ^ Вицедо, Марга (2000). Касл, Уильям Эрнест (1867–1962), биолог . Американская национальная биография онлайн. Издательство Оксфордского университета . дои : 10.1093/anb/9780198606697.article.1302308 .
- ^ Перейти обратно: а б Боулер 2003 , стр. 271–272.
- ^ Провайн 2001 , стр. 120–121.
- ^ Перейти обратно: а б Смоковит 1996 , стр. 100–114
- ^ Смоковит 1996 , стр. 114–119
- ^ Смоковит 1996 , стр. 119–122
- ^ Фишер, Рональд А. (январь 1919 г.). «XV. — Корреляция между родственниками на основании предположения о менделевском наследовании» . Труды Королевского общества Эдинбурга . 52 (2): 399–433. дои : 10.1017/S0080456800012163 . OCLC 4981124 . S2CID 181213898 . «Документ, прочитанный Дж. Артуром Томсоном 8 июля 1918 года Королевскому обществу Эдинбурга».
- ^ Фишер 1999
- ^ Перейти обратно: а б с д и ж г час Ларсон 2004 , стр. 221–243.
- ^ Хоскен, Дэвид Дж.; Хаус, Кларисса М. (25 января 2011 г.). «Половой отбор» . Современная биология . 21 (2): Р62–Р65. дои : 10.1016/j.cub.2010.11.053 . ПМИД 21256434 . S2CID 18470445 .
- ^ Перейти обратно: а б с д и ж Боулер 2003 , стр. 325–339.
- ^ Перейти обратно: а б Смоковит 1996 , стр. 122–132
- ^ Смоковит 1996 , с. 192
- ^ Хелд, Льюис И. (2014). Как Змея потеряла ноги. Любопытные истории с границ Эво-Дево . Издательство Кембриджского университета . п. 67. ИСБН 978-1-107-62139-8 .
- ^ Гулд 1977 , стр. 221–222.
- ^ Холл, БК (2003). «Эво-Дево: эволюционные механизмы развития». Международный журнал биологии развития . 47 (7–8): 491–495. CiteSeerX 10.1.1.113.5158 . ПМИД 14756324 .
- ^ Инго Бригандт (2006). «Гомология и гетерохрония: эволюционный эмбриолог Гэвин Райландс де Бир (1899-1972)» (PDF) . Журнал экспериментальной зоологии . 306Б (4): 317–328. дои : 10.1002/jez.b.21100 . ПМИД 16506229 . Архивировано (PDF) из оригинала 4 ноября 2006 г.
- ^ Перейти обратно: а б Гилберт, Скотт Ф .; Опиц, Дж. М.; Рафф, РА (1996). «Ресинтез эволюционной биологии и биологии развития» . Биология развития . 173 (2): 357–372. дои : 10.1006/dbio.1996.0032 . ПМИД 8605997 .
- ^ Адамс, М. (1991). «Зазеркалье: Эволюция советского дарвинизма». В Уоррене, Л.; Копровски, Х. (ред.). Новые перспективы эволюции . Лисс/Уайли. стр. 37–63 . ISBN 978-0-471-56068-5 .
- ^ Перейти обратно: а б Смоковит 1996 , стр. 153–171
- ^ Райт 1932 , стр. 356–366.
- ^ Перейти обратно: а б Роуз, Майкл Р .; Окли, Тодд Х. (24 ноября 2007 г.). «Новая биология: за пределами современного синтеза» (PDF) . Биология Директ . 2 (30): 30. дои : 10.1186/1745-6150-2-30 . ПМК 2222615 . ПМИД 18036242 . Архивировано (PDF) из оригинала 21 марта 2014 г.
- ^ Хаксли, Джулиан (1942). Эволюция: современный синтез . Аллен и Анвин .
- ^ Ридли, Мэтт (1996). Эволюция (2-е изд.). Блэквелл Наука. ISBN 978-0632042920 .
- ^ Майр и Провин 1998 , стр. 231.
- ^ Добжанский 1937 г.
- ^ Смоковитис 1996 , с. 127
- ^ Элдридж, Найлз (1985). Неоконченный синтез: биологические иерархии и современная эволюционная мысль . Издательство Оксфордского университета . п. 17. ISBN 978-0-19-536513-9 .
- ^ Перейти обратно: а б Форд 1964 года
- ^ Форд 1975 г.
- ^ Форд, Э.Б. (1949). Менделизм и эволюция . Метуэн .
- ^ Добжанский 1951 г.
- ^ Шмальгаузен, Иван Иванович ; Дордик, Исадор (пер.) (1949). Добжанский, Феодосий (ред.). Факторы эволюции. Теория стабилизирующего отбора . Филадельфия и Торонто: Компания Blakiston .
- ^ Левит, Георгий С.; Хоссфельд, Уве; Олссон, Леннарт (2006). «От« современного синтеза »к кибернетике: Иван Иванович Шмальгаузен (1884–1963) и его исследовательская программа по синтезу эволюционной биологии и биологии развития». Журнал экспериментальной зоологии . 306Б (2006): 89–106. дои : 10.1002/jez.b.21087 . ПМИД 16419076 . S2CID 23594114 .
- ^ Адамс, МБ (июнь 1988 г.). «Недостающее звено в эволюционном синтезе. И. И. Шмальгаузен. Факторы эволюции: теория стабилизирующего отбора». Исида . 79 (297): 281–284. дои : 10.1086/354706 . ПМИД 3049441 . S2CID 146660877 .
- ^ Стекло, Бентли (декабрь 1951 г.). «Обзоры и краткие заметки Факторы эволюции. Теория стабилизирующего отбора. И. И. Шмальгаузен, Исадор Дордик, Феодосий Добжанский». Ежеквартальный обзор биологии . 26 (4): 384–385. дои : 10.1086/398434 .
- ^ Майр и Провин 1998 , стр. ix.
- ^ Перейти обратно: а б с Русе 1996 , стр. 328–338.
- ^ Перейти обратно: а б Ламм, Эхуд. «Обзор Джулиана Хаксли, Эволюция: Современный синтез – окончательное издание, с новым предисловием Массимо Пильуччи и Герда Б. Мюллера» (PDF) . МТИ Пресс . Архивировано (PDF) из оригинала 24 декабря 2011 г. Проверено 21 августа 2017 г.
- ^ Перейти обратно: а б с д Смоковит 1996 , стр. 138–153
- ^ Майр 1999 г.
- ^ Привет, Джоди; Фитч, Уолтер М.; Аяла, Франсиско Дж. (2005). «Систематика и происхождение видов: Введение» . ПНАС . 102 (приложение 1): 6515–6519. Бибкод : 2005PNAS..102.6515H . дои : 10.1073/pnas.0501939102 . ПМЦ 1131868 . ПМИД 15851660 .
- ^ Майр и Провайн 1998 , стр. 33–34
- ^ Майр 1982 .
- ^ Ренш 1947 ; Ренш 1959 г.
- ^ Смит, Чарльз Х. «Ренш, Бернхард (Карл Эммануэль) (Германия 1900–1990)» . Некоторые биогеографы, эволюционисты и экологи: хронобиографические очерки . Университет Западного Кентукки . Проверено 14 декабря 2017 г.
- ^ Mayr & Provine 1998 , стр. 298–299, 416.
- ^ Смоковит, В.Б. (2001). «Дж. Ледьярд Стеббинс и эволюционный синтез». Ежегодный обзор генетики . 35 : 803–814. дои : 10.1146/annurev.genet.35.102401.091525 . ПМИД 11700300 .
- ^ Перейти обратно: а б с д Майр Э.: Где мы? Симпозиум по количественной биологии в Колд-Спринг-Харборе 24: 1–14, 1959 г.
- ^ Перейти обратно: а б с д и Стеббинс, Г.Л.: Процессы органической эволюции, с. 12. Прентис Холл, 1966 год.
- ^ Перейти обратно: а б с Добжанский Т.: В: Аяла Ф., Добжанский Т. (ред.) Случай и творчество в эволюции, стр. 307–338. Калифорнийский университет Press , Беркли и Лос-Анджелес (1974)
- ^ Майр, Э.: В: Майр, Э., Провайн, В. (ред.) Некоторые мысли об истории эволюционного синтеза, стр. 1–48. Издательство Гарвардского университета , 1980 г.
- ^ Перейти обратно: а б Гамильтон, WD (1964). «Генетическая эволюция социального поведения I» (PDF) . Журнал теоретической биологии . 7 (1): 1–52. Бибкод : 1964JThBi...7....1H . дои : 10.1016/0022-5193(64)90038-4 . ПМИД 5875341 . Архивировано (PDF) из оригинала 18 декабря 2016 г.
- ^ Перейти обратно: а б Уильямс, Джордж К. (1966). Адаптация и естественный отбор . Издательство Принстонского университета . п. 307. ИСБН 978-0-691-02615-2 .
- ^ Перейти обратно: а б с д и Юделл, Майкл; Десаль, Роб (2000). «Социобиология. Новый синтез. Эдвард О. Уилсон». Журнал истории биологии . 33 (3): 577–584. дои : 10.1023/а:1004845822189 . JSTOR 4331614 . S2CID 88047081 .
- ^ Перейти обратно: а б Гилберт, Скотт Ф. (2000). «Новый эволюционный синтез». Биология развития . Синауэр Ассошиэйтс .
- ^ Мейнард Смит, Джон (1964). «Групповой отбор и родственный отбор». Природа . 201 (4924): 1145–1147. Бибкод : 1964Natur.201.1145S . дои : 10.1038/2011145a0 . S2CID 4177102 .
- ^ Пинкер, Стивен (1994). Языковой инстинкт . Харпер Многолетняя современная классика. п. 294.
- ^ Окаша, Самир (22 сентября 2006 г.). «Популяционная генетика» – через plato.stanford.edu.
- ^ Докинз, Ричард (1982). Расширенный фенотип . Издательство Оксфордского университета . ISBN 978-0-19-288051-2 .
- ^ Фишер, Хелен (16 октября 1994 г.). « Уилсон, — сказали они, — ты весь мокрый!» " . Нью-Йорк Таймс . Проверено 21 июля 2015 г.
- ^ Ленски, Герхард (1976). «Социобиология. Новый синтез. Эдвард О. Уилсон». Социальные силы . 55 (2): 530–531. дои : 10.2307/2576242 . JSTOR 2576242 .
- ^ Смоковит 1996 , стр. 186–187.
- ^ Перейти обратно: а б Пильуччи, Массимо; Финкельман, Леонард (2014). «Дебаты о расширенном (эволюционном) синтезе: где наука встречается с философией» . Бионаука . 64 (6): 511–516. дои : 10.1093/biosci/biu062 .
- ^ Перейти обратно: а б с Благородный, Денис (2011). «Неодарвинизм, современный синтез и эгоистичные гены: полезны ли они в физиологии?» . Журнал физиологии . 589 (5): 1007–1015. дои : 10.1113/jphysicalol.2010.201384 . ПМК 3060581 . ПМИД 21135048 .
- ^ Уэйд, Майкл Дж. (2011). «Неомодернистский синтез: слияние новых данных и объяснительных концепций» . Бионаука . 61 (5): 407–408. дои : 10.1525/bio.2011.61.5.10 .
- ^ Перейти обратно: а б Джон Одлинг-Сми и др . «Расширенный эволюционный синтез: его структура, предположения и предсказания» . Труды Королевского общества B: Биологические науки, август 2015 г.
- ^ Уэйд, Майкл Дж. (2011). «Неомодернистский синтез: слияние новых данных и объяснительных концепций» . Бионаука . 61 (5): 407–408. дои : 10.1525/bio.2011.61.5.10 .
- ^ Данчин, Э.; Шармантье, А.; Шампань, ФА; Месуди, А.; Пухоль, Б.; Бланше, С. (2011). «За пределами ДНК: интеграция инклюзивного наследования в расширенную теорию эволюции». Обзоры природы Генетика . 12 (7): 475–486. дои : 10.1038/nrg3028 . ПМИД 21681209 . S2CID 8837202 .
- ^ Лаубихлер, Манфред Д.; Ренн, Юрген (2015). «Расширенная эволюция: концептуальная основа для интеграции сетей регулирования и создания ниш» . Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 324 (7): 565–577. дои : 10.1002/jez.b.22631 . ПМЦ 4744698 . ПМИД 26097188 .
- ^ Перейти обратно: а б Кучера, Ульрих ; Никлас, Карл Дж. (июнь 2004 г.). «Современная теория биологической эволюции: расширенный синтез». Naturwissenschaften . 91 (6): 255–276. Бибкод : 2004NW.....91..255K . дои : 10.1007/s00114-004-0515-y . ПМИД 15241603 . S2CID 10731711 .
- ^ Хьюнеман, Филипп; Уолш, Денис М. (2017). Вызов современному синтезу: адаптация, развитие и наследование . Издательство Оксфордского университета . стр. 76 (глава 2). ISBN 978-0-19-068145-6 .
- ^ Хьюнеман, Филипп; Уолш, Денис М. (2017). Вызов современному синтезу: адаптация, развитие и наследование . Издательство Оксфордского университета . стр. Введение. ISBN 978-0-19-068145-6 .
- ^ Кэрролл, Шон Б. (2008). «Эво-Дево и расширяющийся эволюционный синтез: генетическая теория морфологической эволюции» . Клетка . 134 (1): 25–36. дои : 10.1016/j.cell.2008.06.030 . ПМИД 18614008 . S2CID 2513041 .
- ^ Перейти обратно: а б Смоковит 1996 , с. 187
- ^ Смоковитис 1996 , с. 43
Источники
[ редактировать ]- Бейтсон, Уильям (1894). Материалы для изучения изменчивости с особым учетом прерывистости происхождения видов . Лондон; Нью-Йорк: Макмиллан . LCCN 07001821 . OCLC 1944285 .
- Боулер, Питер Дж. (2003). Эволюция: История идеи (3-е полностью переработанное и расширенное изд.). Беркли, Калифорния: Издательство Калифорнийского университета . ISBN 978-0-520-23693-6 .
- Боулер, Питер Дж. (1989). Эволюция: история идеи (2-е исправленное изд.). Беркли, Калифорния: Издательство Калифорнийского университета . ISBN 978-0-520-06386-0 .
- Добжанский, Феодосий (1937). Генетика и происхождение видов . Биологическая серия Колумбийского университета . Нью-Йорк: Издательство Колумбийского университета . LCCN 37033383 . OCLC 766405 .
- —— (1951). Генетика и происхождение видов . Биологическая серия Колумбийского университета (3-е исправленное издание). Нью-Йорк: Издательство Колумбийского университета. LCCN 51014816 . OCLC 295774 .
- Фишер, Р.А. (1999) [1930]. Генетическая теория естественного отбора . Отредактировано с предисловием и примечаниями Дж. Х. Беннета (Полное разнообразное издание). Оксфорд, Великобритания: Издательство Оксфордского университета. ISBN 978-0-19-850440-5 .
- Форд, Э.Б. (1964). «Экологическая генетика». Развитие науки . 25 (124). Лондон; Нью-Йорк: Издательство Метуэн ; Джон Уайли и сыновья : 227–35. LCCN 64009038 . OCLC 875507052 . ПМИД 5701915 .
- Гулд, Стивен Джей (1977). Онтогенез и филогения . Belknap Press издательства Гарвардского университета . ISBN 978-0-674-63940-9 .
- Гулд, Стивен Джей (2002). Структура эволюционной теории . Belknap Press издательства Гарвардского университета. ISBN 978-0-674-00613-3 .
- Графен, Алан ; Ридли, Марк , ред. (2006). Ричард Докинз: Как учёный изменил наше мышление . Издательство Оксфордского университета. ISBN 978-0-19-929116-8 .
- Хаксли, Джулиан (2010) [1942]. Эволюция: современный синтез . С новым предисловием Массимо Пильуччи и Герда Б. Мюллера (Окончательное издание). Кембридж, Массачусетс: MIT Press . ISBN 978-0-262-51366-1 .
- Ларсон, Эдвард Дж. (2004). Эволюция: замечательная история научной теории . Современные библиотечные хроники. Том. 17. Нью-Йорк: Современная библиотека . ISBN 978-0-679-64288-6 .
- Майр, Эрнст (1982). Рост биологической мысли: разнообразие, эволюция и наследование . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета. ISBN 978-0-674-36445-5 .
- Майр, Эрнст (1999) [1942]. Систематика и происхождение видов с точки зрения зоолога (1st Harvard University Press, изд.). Издательство Гарвардского университета . ISBN 978-0-674-86250-0 .
- Майр, Эрнст; Провин, Уильям Б. , ред. (1998) [1980]. Эволюционный синтез: перспективы унификации биологии . С новым предисловием Эрнста Майра (1-е изд. в мягкой обложке). Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-27226-2 .
- Провайн, ВБ (2001). Истоки теоретической популяционной генетики с новым послесловием . Издательство Чикагского университета. ISBN 978-0-226-68464-2 .
- Ренш, Бернхард (1947). Новые проблемы теории происхождения. Die трансвидовая эволюция [ Новые проблемы эволюционной теории: трансвидовая эволюция ] (на немецком языке). Штутгарт: Фердинанд Энке Верлаг. ОСЛК 2271422 .
- Русе, Майкл (1996). От Монады человеку: концепция прогресса в эволюционной биологии . Издательство Гарвардского университета. ISBN 978-0-674-03248-4 .
- Смоковит, Василики Бетти (1996). «Объединяющая биология: эволюционный синтез и эволюционная биология». Журнал истории биологии . 25 (1). Принстон, Нью-Джерси: Издательство Принстонского университета: 1–65. дои : 10.1007/bf01947504 . ISBN 978-0-691-03343-3 . ПМИД 11623198 . S2CID 189833728 .
- Райт, Сьюэлл (1932). «Роль мутации, инбридинга, скрещивания и отбора в эволюции». В Джонсе, Дональд Ф. (ред.). Материалы Шестого международного конгресса генетиков . Том. 1. Итака, штат Нью-Йорк: Американское генетическое общество . OCLC 439596433 .
Дальнейшее чтение
[ редактировать ]- Аллен, Гарланд Э. (1978). Томас Хант Морган: Человек и его наука . Принстон, Нью-Джерси: Издательство Принстонского университета . ISBN 978-0-691-08200-4 .
- Докинз, Ричард (1990). Слепой часовщик . Наука пингвинов. Лондон: Книги Пингвинов . ISBN 978-0-14-014481-9 .
- Футуйма, Дуглас Дж. (1998). Эволюционная биология (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates . ISBN 978-0-87893-189-7 .
- Холдейн, JBS (1932). Причины эволюции . Лондон; Нью-Йорк: Longmans, Green & Co. LCCN 32033284 . OCLC 5006266 . «Эта книга основана на серии лекций, прочитанных в январе 1931 года в Прифисгол Симру, Аберистуит, и озаглавлена «Пересмотр дарвинизма».
- Хаксли, Джулиан , изд. (1976) [Первоначально опубликовано в 1940 году; Оксфорд, Великобритания: The Clarendon Press ]. Новая Систематика (Переиздание). Сент-Клер Шорс, Мичиган: Scholarly Press. ISBN 978-0-403-01786-7 .
- Левинсон, Джин (2020). Переосмысление эволюции: революция, которая скрывается на виду . Всемирная научная. ISBN 9781786347268 . Архивировано из оригинала 21 мая 2022 г. Проверено 27 января 2020 г.
- Райт, Сьюэлл (1 марта 1931 г.). «Эволюция менделевских популяций» . Генетика . 16 (2): 97–159. дои : 10.1093/генетика/16.2.97 . ПМК 1201091 . ПМИД 17246615 .
| Базы данных органов управления : Национальные |
|---|