Ампа -приемник

Рецептор α-амино-3-гидрокси-5-метил-4-азоксазолепропионовой кислоты (также известный как AMPA рецептор , AMPAR или Quisqualate Receptor ) является ионотропным трансмембрановым рецептором для глутамата ( игур ). + Ионовый канал, который опосредует быструю синаптическую передачу в центральной нервной системе (ЦНС). Он традиционно классифицируется как рецептор не nmda -типа, а также рецептор каината . Его название происходит от его способности активироваться искусственным аналоговым AMPA . Рецептор был впервые назван « Quisqualate рецептором» Уоткинсом и его коллегами после природного агонистского Quisqualate и был только позже с учетом ярлыка «рецептор Ampa Ampa» после того, как селективный агонист, разработанный Tage Honore и коллегами в Королевской датской школе Даняшской школы в Копенхагене, разработанным Tage Honore и коллегами в Королевской датской школе в Копенхагене, разработанной Tage Honore и коллегами в Королевской датской школе в Копенхагене и колледжах Полем [ 1 ] Кодированный грио2 был -кодированный лиганд -лигандский ядро лиганда (GLUA2 LBD) был первым доменом ионного канала рецептора глутамата, который кристаллизован . [ 2 ]
Структура и функция
[ редактировать ]Субъединица композиция
[ редактировать ]Ampars состоит из четырех типов субъединиц, кодируемых различными генами, обозначенными как Gria1 (также названный Glua1 или Glur1), Gia2 (также названный Glua2 или Glur2), Gia3 (также названный Glua3 или Glur3) и Gria4 (также называемый Glua4 или Glura -D2), который объединяется в образу тетрамеров . [ 3 ] [ 4 ] [ 5 ] Большинство AMPAR являются гетеротетрамерными , состоящие из симметричного «димера димеров» Glua2 и Glua1, Glua3 или Glua4. [ 6 ] [ 7 ] Димеризация начинается в эндоплазматической ретикулуме с взаимодействием N-концевых доменов LIVBP, а затем «застегивает» через лиганд-связывающий домен в пору трансмембранной ионы. [ 7 ]
Конформация белка субъединицы в плазматической мембране в течение некоторого времени вызывала противоречие. В то время как аминокислотная последовательность субъединицы указывала на то, что, по-видимому, существует четыре трансмембранных домена (части белка, которые проходят через плазматическую мембрану), белки, с взаимодействующие субъединицей Термин , казалось, был внутриклеточным. Однако, если каждый из четырех трансмембранных доменов прошел через плазматическую мембрану, то два термина должны были быть на одной и той же стороне мембраны. В конечном итоге было обнаружено, что второй «трансмембранная» домен на самом деле вообще не пересекает мембрану, но переживает себя в мембране и возвращается к внутриклеточной стороне. [ 8 ] Когда четыре субъединицы тетрамера объединяются, этот второй мембранный домен образует ионопроницаемую пор рецептора.
Субъединицы AMPAR различаются наиболее различаются по своей С-концевой последовательности, которая определяет их взаимодействие с белками для каркасов. Все AMPAR содержат PDZ-связывающие домены, но с которыми PDZ-домен они связываются. Например, GLUA1 связывается с SAP97 через домен PDZ Class I SAP97, [ 9 ] в то время как Glua2 связывается с Pick1 [ 10 ] и Grip/ABP . Следует отметить, что AMPAR не могут напрямую связываться с общим синаптическим белком PSD-95 из-за несовместимых доменов PDZ, хотя они взаимодействуют с PSD-95 через Stargazin (прототипный член семейства AMPAR вспомогательных субъединиц). [ 11 ]
Фосфорилирование AMPAR может регулировать локализацию, проводимость и открытую вероятность канала. Glua1 имеет четыре известных сайта фосфорилирования в серине 818 (S818), S831, треонин 840 и S845 (другие субъединицы имеют сходные сайты фосфорилирования, но Glur1 были наиболее тщательно изучены). S818 фосфорилируется протеинкиназой C и необходим для длительного потенцирования (LTP; для роли Glua1 в LTP, см. Ниже). [ 12 ] S831 фосфорилируется CAMKII и PKC во время LTP, что помогает доставить GluA1-содержащий AMPAR в синапс , [ 13 ] и увеличивает их одноканальную проводимость. [ 14 ] Сайт T840 был недавно обнаружен и участвовал в LTD. [ 15 ] Наконец, S845 фосфорилируется PKA, что регулирует его открытую вероятность. [ 16 ]
Ионо -канал функция
[ редактировать ]Каждый AMPA имеет четыре сайта, с которыми может связывать агонист (такой как глутамат), по одному для каждой субъединицы. [ 6 ] Считается, что сайт связывания образуется N-концевым хвостом и внеклеточным петлей между трансмембранными доменами три и четырех. [ 17 ] Когда агонист связывается, эти две петли движутся друг с другом, открывая пор. Канал открывается, когда заняты два сайта, [ 18 ] и увеличивает его ток, поскольку заняты больше связующих сайтов. [ 19 ] После открытия канал может пройти быструю десенсибилизацию, останавливая ток. Считается, что механизм десенсибилизации связан с небольшим изменением угла одной из частей сайта связывания, закрывая пор. [ 20 ] AMPAR открываются и быстро закрываются (1 мс) и, таким образом, ответственны за большую часть быстрой возбуждающей синаптической передачи в центральной нервной системе. [ 18 ] Проницаемость AMPAR для кальция и других катионов , таких как натрий и калий , регулируется субъединицей GLUA2. Если AMPAR не хватает субъединицы GluA2, то он будет проницаемы для натрия, калия и кальция. Присутствие субъединицы Glua2 почти всегда будет делать канал непроницаемым для кальция. Это определяется пост- транскрипционной модификацией- редактирование РНК- сайта редактирования Q - r Glua2 мРНК . Здесь A → I Редактирование изменяет незаряженный аминокислотный глутамин (Q) к положительно заряженному аргинину (R) в ионном канале рецептора. Положительно заряженная аминокислота в критической точке делает ее энергетически неблагоприятной для кальция, чтобы войти в клетку через пор. Почти все субъединицы Glua2 в ЦНС редактируются в форме Glua2 (R). Это означает, что основными ионами, управляемыми AMPAR, являются натрий и калий, что отличает AMPAS от NMDA -рецепторов (другие основные ионотропные глутаматные рецепторы в мозге), которые также позволяют приточить кальция. Однако рецепторы AMPA и NMDA имеют равновесный потенциал около 0 мВ. Профилактика ввода кальция в клетку при активации GluA2-содержащих AMPAR предлагается защищать от экситотоксичности . [ 21 ]
Состав субъединицы AMPAR также важен для того, как модулируется этот рецептор. Если AMPAR не хватает субъединиц GluA2, то он подвержен блокированию в зависимости от напряжения классом молекул, называемых полиаминами . Таким образом, когда нейрон находится в деполяризованном мембранном потенциале, полиамины будут более сильно блокировать канал AMPAR, предотвращая поток ионов калия через пору канала. Таким образом, AMPAR, затрагивающие GluA2, имеют внутреннюю исправляющую кривую I/V , что означает, что они проходят менее внешний ток, чем внутренний ток на эквивалентном расстоянии от потенциала реверса. [ 22 ] Проницаемые ампы кальция обычно встречаются рано во время постнатального развития неокортикальных пирамидальных нейронов , [ 22 ] Некоторые интернейроны, или в дофаминовых нейронах вентральной сегментарной области после воздействия наркотика. [ 23 ]
Наряду с редактированием РНК , альтернативный сплайсинг позволяет ряд функциональных субъединиц рецептора AMPA помимо того, что кодируется в геноме . Другими словами, хотя один ген ( GRIA1 - GRIA4 ) кодируется для каждой субъединицы (GLUA1 -GLUA4), сплайсинг после транскрипции из ДНК позволяет трансмировать взаимозаменяемо, что приводит к нескольким функционально различным субъединицам из каждого гена. [ 24 ]
Последовательность флип/флоп является одним из таких сменных экзонов. 38-аминокислотная последовательность, обнаруженная до (т.е. перед N-конце ) четвертого мембранного домена во всех четырех субъединицах AMPAR, она определяет скорость десенсизации [ 25 ] рецептора, а также скорости, с которой рецептор резативтизируется [ 26 ] и скорость закрытия канала. [ 27 ] Формальная форма присутствует в пренатальных рецепторах AMPA и дает устойчивый ток в ответ на активацию глутамата. [ 28 ]
Синаптическая пластичность
[ редактировать ]Рецепторы AMPA (AMPAR) являются рецепторами глутамата и катионными каналами , которые являются неотъемлемой частью пластичности и синаптической передачи на многих постсинаптических мембранах. Одна из наиболее широко и тщательно исследуемых форм пластичности в нервной системе известна как долгосрочное потенцирование или LTP. Существует два необходимых компонента LTP: пресинаптическое высвобождение глутамата и постсинаптическая деполяризация. Следовательно, LTP может быть индуцирован экспериментально в парной электрофизиологической записи , когда пресинаптическая клетка стимулируется для высвобождения глутамата на постсинаптической клетке, которая деполяризована. Типичный протокол индукции LTP включает стимуляцию «tetanus», которая представляет собой стимуляцию 100 Гц в течение 1 секунды. Когда человек применяет этот протокол к паре клеток, можно увидеть устойчивое увеличение амплитуды возбуждающего постсинаптического потенциала (EPSP) после столбняка. Этот ответ интересен, поскольку считается, что он является физиологическим коррелятом для обучения и памяти в клетке. Фактически, было показано, что после единой парадигмы из парной предотвращения у мышей может быть записана LTP в некоторых гиппокампа Синапсы in vivo . [ 29 ]
Молекулярная основа для LTP была тщательно изучена, и было показано, что AMPAR играют неотъемлемую роль в процессе. И Glur1, и Glur2 играют важную роль в синаптической пластичности. В настоящее время известно, что основной физиологический коррелят для увеличения размера EPSP является постсинаптическая активация AMPAR на мембране, [ 30 ] который достигается через взаимодействие AMPAR со многими клеточными белками.
Самое простое объяснение для LTP заключается в следующем (см. Долгосрочную статью о потенцировании для гораздо более подробной учетной записи). Глутамат связывается с постсинаптическими ампапами и другим рецептором глутамата, рецептором NMDA (NMDAR). Связывание лиганда вызывает открытие AMPAR и NA + течет в постсинаптическую клетку, что приводит к деполяризации. NMDARS, с другой стороны, не открываются непосредственно, потому что их поры закупориваются при покорном мембранном потенциале MG 2+ ионы NMDARS могут открываться только тогда, когда деполяризация от активации AMPAR приводит к отталкиванию MG 2+ Катион в внеклеточное пространство, позволяя пор проходить ток. Однако, в отличие от AMPAR, NMDAR проницаемы для обоих NA + и ca 2+ Полем Ca 2+ Это входит в ячейку запускает активацию AMPAR на мембрану, что приводит к длительному увеличению размера EPSP, лежащего в основе LTP. Вход кальция также фосфорилирует CAMKII , который фосфорилирует AMPAR, увеличивая их одноканальную проводимость.
Ампа -рецептор. Торгоза
[ редактировать ]
Молекулярный и сигнальный ответ на LTP-индуцирующие стимулы
[ редактировать ]Механизм для LTP уже давно стал темой дебатов, но в последнее время механизмы достигли некоторого консенсуса. AMPAR играют ключевую роль в этом процессе, поскольку одним из ключевых показателей индукции LTP является увеличение отношения AMPAR к NMDAR после высокочастотной стимуляции. Идея состоит в том, что AMPARS подвергаются переносу из дендрита в синапс и включены через некоторые серии сигнальных каскадов.
AMPAR первоначально регулируются на уровне транскрипции в их 5 -'промоторных областях. Существуют значительные доказательства, указывающие на контроль транскрипции рецепторов AMPA в долгосрочной памяти с помощью белка связывающего элемента CAMP-ответа ( CREB ) и митогенов, активируемых протеинкиназами (MAPK). [ 31 ] Сообщения переводятся на грубой эндоплазматической ретикулуме (грубой ER) и модифицируются там. Композиции субъединиц определяются во время модификации в грубой ER. [ 10 ] После пост-ER обработки в аппарате Гольджи AMPAR выпускаются в перисинаптическую мембрану как резерв, ожидающий начального процесса LTP.
Первым ключевым этапом в процессе после связывания глутамата с NMDAR является приток кальция через рецепторы NMDA и результирующую активацию CA 2+ /кальмодулин-зависимая протеинкиназа (CAMKII). [ 32 ] Блокирование либо этого притока, либо активации CAMKII предотвращает LTP, показывая, что это необходимые механизмы для LTP. [ 33 ] Кроме того, изобилие CAMKII в синапс вызывает LTP, показывая, что это причинно -следственный и достаточный механизм. [ 34 ]
CAMKII имеет множество режимов активации, чтобы вызвать включение рецепторов AMPA в перисинаптическую мембрану. Фермент CAMKII в конечном итоге отвечает за развитие актинового цитоскелета нейрональных клеток и, в конечном итоге, за развитие дендрита и аксона (синаптическая пластичность). [ 35 ] Первым является прямое фосфорилирование синаптического ассоциированного белка 97 ( SAP97 ). [ 36 ] Во-первых, SAP-97 и миозин-VI, моторный белок, связаны как комплекс с С-конце AMPAR. После фосфорилирования CAMKII комплекс перемещается в перисинаптическую мембрану. [ 37 ] Второй режим активации - через путь MAPK. CAMKII активирует белки Ras, которые продолжают активировать P42/44 MAPK, что приводит к вставке AMPAR непосредственно в перисинаптическую мембрану. [ 38 ]
Торгоза рецептора AMPA в PSD в ответ на LTP
[ редактировать ]После того, как рецепторы AMPA транспортируются в перисинаптическую область через фосфорилирование PKA или SAP97, рецепторы затем переносятся в постсинаптическую плотность (PSD). Тем не менее, этот процесс торговли PSD все еще остается спорным. Одна возможность состоит в том, что во время LTP происходит боковое движение рецепторов AMPA из перисинаптических сайтов непосредственно к PSD. [ 39 ] Другая возможность состоит в том, что экзоцитоз внутриклеточных пузырьков отвечает за транспортировку AMPA в PSD напрямую. [ 40 ] Последние данные свидетельствуют о том, что оба этих процесса происходят после стимула LTP; Однако только боковое движение рецепторов AMPA из перисинаптической области увеличивает количество рецепторов AMPA в PSD. [ 41 ] Точный механизм, ответственный за боковое движение рецепторов AMPA к PSD, еще предстоит обнаружить; Тем не менее, исследования обнаружили несколько важных белков для транспорта рецепторов AMPA. Например, сверхэкспрессия SAP97 приводит к увеличению транспорта рецепторов AMPA до синапсов . [ 42 ] В дополнение к влиянию синаптической локализации, также было обнаружено, что SAP97 влияет на проводимость рецептора AMPA в ответ на глутамат . [ 43 ] Белки миозина являются чувствительными к кальцие моторными белками, которые также были необходимы для транспорта рецепторов AMPA. Нарушение взаимодействия миозина VB с Rab11 и Rab11-FIP2 блокирует рост позвоночника и транспортировку рецепторов AMPA. [ 44 ] Следовательно, возможно, что миозин может привести к боковому движению рецепторов AMPA в перисинаптической области к PSD. Трансмембранные регуляторные белки AMPA -рецептора (TARP) являются семейными белками, которые связываются с рецепторами AMPA и контролируют их торговлю и проводимостью. [ 45 ] CACNG2 (StarGazin) является одним из таких белков и обнаруживается, что он связывает рецепторы AMPA в перисинаптических и постсинаптических областях. [ 46 ] Роль Stargazin в торговле между перизинаптическими и постсинаптическими регионами остается неясной; Тем не менее, Stargazin необходим для иммобилизации рецепторов AMPA в PSD путем взаимодействия с PSD-95. [ 47 ] PSD-95 стабилизирует рецепторы AMPA с синапсом и разрушением взаимодействия Stargazin-PSD-95 подавляемого синаптической передачи. [ 11 ]
Биофизика транспорта рецепторов AMPA
[ редактировать ]Движение рецепторов AMPA на синаптической мембране хорошо аппроксимируется как Брауньян , который, однако, может быть стабилизирован в PSD с помощью сил удержания. Эти силы могут стабилизировать рецепторы трансляции, но позволяют постоянно обменять с перинаптическим доменом. [ 48 ] [ 49 ] Эти силы могут возникнуть в результате местной организации PSD, иногда называйте фазовым разделением .
Конститутивный торговлю и изменениями в композиции субъединиц
[ редактировать ]Рецепторы AMPA постоянно подвергаются обороту (эндоцитозированные, переработанные и повторные вставки) в плазматическую мембрану и из нее . Утилизация эндосом в дендритном отделении позвоночника содержат пулы рецепторов AMPA для такого синаптического повторного введения. [ 50 ] Существуют два разных пути для торговли рецепторами AMPA: регулируемый путь и конститутивный путь. [ 51 ] [ 52 ]
В регулируемом пути рецепторы AMPA-содержащих GluA1 подвергаются перенос в синапс зависимым от активности способом, стимулируемым активацией рецептора NMDA . [ 13 ] В базальных условиях регулируемый путь по существу неактивен, а также временно активируется только при индукции долгосрочного потенцирования . [ 50 ] [ 51 ] Этот путь отвечает за синаптическое укрепление и начальное образование новых воспоминаний. [ 53 ]
В конститутивном пути рецепторы AMPA, переживающие GluA1, обычно гетеромерные рецепторы GluR2-Glur3, замените GluA1-содержащие рецепторы в одном, независимом от активности способа [ 54 ] [ 55 ] Сохранение общего количества рецепторов AMPA в синапсе. [ 50 ] [ 51 ] Этот путь отвечает за поддержание новых воспоминаний, поддержание переходных изменений в результате регулируемого пути. В базальных условиях этот путь обычно активен, поскольку это необходимо также для замены поврежденных рецепторов.
Субъединицы Glua1 и Glua4 состоят из длинного карбокси (C)-хвоста, тогда как субъединицы Glua2 и Glua3 состоят из короткого карбокси-хвоста. Два пути определяются взаимодействиями между C -концами субъединиц рецептора AMPA и синаптическими соединениями и белками. Длинные C-хвост предотвращают вставки GluR1/4 непосредственно в зону постсинаптической плотности (PSDZ) в отсутствие активности, тогда как короткие C-хвост рецепторов GluA2/3 позволяют им вставить непосредственно в PSDZ. [ 39 ] [ 56 ] Термин Clua2 C взаимодействует и связывается с N-этилмалеимидным чувствительным слитым белком , [ 57 ] [ 58 ] [ 59 ] что позволяет быстро вставить GluR2-содержащие рецепторы AMPA в синапсе. [ 60 ] Кроме того, субъединицы GLUR2/3 более стабильно привязаны к синапсу, чем субъединицы GluR1. [ 61 ] [ 62 ] [ 63 ]
Индуцированный ООО эндоцитоз рецепторов AMPA
[ редактировать ]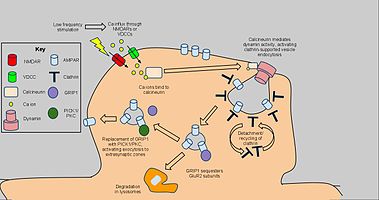
Долгосрочная депрессия вступает в действие механизмов для снижения плотности рецепторов AMPA в выбранных дендритных шипах, зависящих от клатрина и кальциневрина и отличающегося от плоды конститутивного транспорта AMPAR. Начальный сигнал для эндоцитоза AMPAR является NMDAR-зависимым притоком кальция из низкочастотной стимуляции, которая, в свою очередь, активирует белкофосфатазы PP1 и кальциневрин. Тем не менее, эндоцитоз AMPAR также активирован с помощью напряжения, зависимых от кальциевых каналов , агонизма AMPA-рецепторов и введения инсулина , что предполагает общий приток кальция в качестве причины эндоцитоза AMPAR. [ 64 ] Закупорка PP1 не предотвращала эндоцитоз AMPAR, но применение антагониста к кальциневрину привело к значительному ингибированию этого процесса. [ 65 ]
Кальциневрин взаимодействует с эндоцитотическим комплексом в постсинаптической зоне, объясняя его влияние на LTD. [ 66 ] Комплекс, состоящий из ямы, покрытой клатрином, под участком AMPAR-содержащей плазматической мембраны и взаимодействующих белков, является прямым механизмом восстановления AMPAR, в частности, в синапсе, в синапсе. Взаимодействия из кальциневрина активируют активность динамимина GTPase, позволяя клатриновой яме актизировать себя из клеточной мембраны и стать цитоплазматической пузырькой. [ 67 ] Как только клатриновое покрытие отсоединяется, другие белки могут напрямую взаимодействовать с AMPAR, используя PDZ -карбоксильные домены хвоста; Например, белок 1-глутаматного рецептора-взаимодействующего 1 ( GRIP1 ) участвует во внутриклеточной секвестрации AMPAR. [ 68 ] Внутриклеточные ампы впоследствии отсортируются для деградации с помощью лизосомов или утилизации клеточной мембраны. [ 69 ] Для последнего PICK1 и PKC могут вытеснить GRIP1, чтобы вернуть AMPARS на поверхность, обращая вспять эффекты эндоцитоза и LTD. Когда уместно. [ 70 ] Тем не менее, выделенный кальций-зависимый, опосредованный динамин механизм, приведенный выше как ключевой компонент LTD. и как таковые могут иметь приложения для дальнейших поведенческих исследований. [ 71 ]
Роль в припадках
[ редактировать ]Рецепторы AMPA играют ключевую роль в генерации и распространении эпилептических припадков. [ 72 ] Каиновая кислота , судорожный, который широко используется в исследовании эпилепсии, частично вызывает судороги посредством активации рецепторов AMPA [ 73 ]
Молекулярная мишень для терапии эпилепсией
[ редактировать ]Было продемонстрировано, что неконкурентные антагонисты AMPA -рецептора Talampanel и Perampanel обладают активностью при лечении взрослых с частичными припадками, [ 74 ] [ 75 ] Указывая о том, что антагонисты AMPA -рецептора представляют собой потенциальную мишень для лечения эпилепсии. [ 76 ] [ 77 ] Перампанель (торговое название: FYCOMPA) получила одобрение разрешения на маркетинг Европейской комиссией для лечения частичной эпилепсии 27 июля 2012 года. Препарат был утвержден в Соединенных Штатах Управлением по контролю за продуктами и лекарствами (FDA) 22 октября 2012 года. Как и в случае с последними разработанными AED, включая прегабалин , лакозамид и эзогабин , FDA рекомендовал, чтобы Perampanel была классифицирована Управлением по борьбе с наркотиками (DEA) в качестве запланированного препарата. Он был обозначен как контролируемое вещество. 3.
Декановая кислота действует как неконкурентный антагонист AMPA-рецептора в терапевтически значимых концентрациях, в зависимости от напряжения и субъединиц, и этого достаточно для объяснения эффектов его антисиализации. [ 78 ] Это прямое ингибирование возбуждающей нейротрансмиссии декановой кислотой в мозге способствует противосудорожному эффекту с триглицеридом средней цепи кетогенной диеты . [ 78 ] Декановая кислота и антагонист -антагонист AMPA -антагонист, препарат Perampanel, в отдельных участках на рецепторе AMPA, и поэтому возможно, что они оказывают кооперативный эффект на рецепторе AMPA, что позволяет предположить, что Perampanel и кетогенная диета могут быть синергическими. [ 78 ] [ 79 ]
Доклинические исследования показывают, что несколько производных ароматических аминокислот с антиглутаматергическими свойствами, включая антагонизм рецептора AMPA и ингибирование высвобождения глутамата, такого как 3,5-дибром-D-тирозин и 3,5-дибром-L-фенилалнин предполагая использование этих соединений в качестве нового класса противоэпилептических препаратов. [ 80 ] [ 81 ]
Агонисты
[ редактировать ]
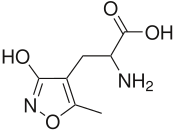
- 5-Fluorowillardiine -синтетическая модификация Willardiine
- AMPA -синтетический агонист, после которого рецептор назван
- Домоевая кислота - встречающийся в природе агонист, который вызывает отравление амнезическими моллюсками
- Глутаминовая кислота (глутамат) - эндогенный агонист
- Иботеновая кислота - природная агонист, обнаруженный в аманите Маскарией
- Quisqualic кислота - природная агонист, обнаруженный у определенных видов
- Willardiine - естественный агонист
Положительные аллостерические модуляторы
[ редактировать ]- Анирацетам
- Циклотиазид
- CX-516
- CX-546
- CX-614
- Навыки (TAK-653)
- CX-717
- Faramptor (CX-691, ORG-24448)
- Hydra-21
- LY-404187
- LY-503430 [ 82 ] [ 83 ]
- Mibampator (LY-451395)
- Org-26576
- Конец -cet
- Бумага
- Pesampator (BIIB-104)
- Пирацетам
- Pramiracetam
- Tulrampator (S-47445, CX-1632)
Антагонисты
[ редактировать ]- Becampanel
- CNQX
- Дасолампанель
- DNQX
- Hamcare (mpqx)
- Кайтоцефалин
- Кинуренновая кислота - эндогенный лиганд
- L-теанин
- NBQX
- 3,5-дибримо-L-фенилаланин, природное галогеничное производное L-фенилаланин [ 84 ]
- Перампанель
- Selurampanel
- Tezampaanel
- Зонампанель
Негативные аллостерические модуляторы
[ редактировать ]
- Барбитураты (например, пентобарбитал , тиопентам натрия )-не селективные
- Этанол -не селективный
- Ингаляционные анестетики (например, циклопропан , энфлуран , галотан , изофлуран , севофлуран )-не селективные
- GYKI-52466
- Импанель
- Перампанель
- Тампалампанель
- PEP1-TGL: Аналог пептида PEP1-TGL GluA1, который ингибирует включение рецептора AMPA в постсинаптическую плотность [ 85 ] [ 86 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Honoré T, Lauridsen J, Krogsgaard-Larsen P (январь 1982 г.). «Связывание [3H] AMPA, структурного аналога глутаминовой кислоты с мембранами мозга крысы» Журнал нейрохимии 38 (1): 173–8 Doi : 10.1111/j.1471-4159.1982.tb10868.x . PMID 6125564 . S2CID 42753770
- ^ Армстронг Н. (2000). «Армстронг и Гуаукс (2000) Механизмы активации и антагонизма рецептора глутамата, чувствительного к ампа,: кристаллические структуры ядра связывающего лиганда GluR2» . Нейрон . 28 (1): 165–181. doi : 10.1016/s0896-6273 (00) 00094-5 . PMID 11086992 . S2CID 3128719 .
- ^ «Рецепторы глутамата: структуры и функции. Университет Бристольского центра синаптической пластичности» . Архивировано из оригинала 15 сентября 2007 года . Получено 2007-09-02 .
- ^ Ши Ш.Х., Хаяси Ю., Петалия Р.С., Заман Ш.Х., Уэдхолд Р.Дж., Свобода К. , Малиноу Р. (июнь 1999 г.). «Быстрая доставка позвоночника и перераспределение рецепторов AMPA после активации синаптического рецептора NMDA». Наука . 284 (5421): 1811–6. Citeseerx 10.1.1.376.3281 . doi : 10.1126/science.284.5421.1811 . PMID 10364548 .
- ^ Песня I, Huganir RL (ноябрь 2002 г.). «Регуляция рецепторов AMPA во время синаптической пластичности». Тенденции в нейронауках . 25 (11): 578–88. doi : 10.1016/s0166-2236 (02) 02270-1 . PMID 12392933 . S2CID 1993509 .
- ^ Jump up to: а беременный Mayer ML (июнь 2005 г.). «Ионные каналы рецептора глутамата» (PDF) . Современное мнение о нейробиологии . 15 (3): 282–8. doi : 10.1016/j.conb.2005.05.004 . PMID 15919192 . S2CID 39812856 .
- ^ Jump up to: а беременный Грегер IH, Ziff EB, Penn AC (август 2007 г.). «Молекулярные детерминанты сборки субъединиц рецептора AMPA». Тенденции в нейронауках . 30 (8): 407–16. doi : 10.1016/j.tins.2007.06.005 . PMID 17629578 . S2CID 7505830 .
- ^ Холлманн М., Марон С., Хейнеманн С (декабрь 1994). «Тегин сайта n-гликозилирования предполагает три трансмембранную топологию домена для глутаматного рецептора GluR1». Нейрон . 13 (6): 1331–43. doi : 10.1016/0896-6273 (94) 90419-7 . PMID 7993626 . S2CID 39682094 .
- ^ Леонард А.С., Даваре М.А., Хорн М.К., Гарнер К.К., Ад Дж.В. (июль 1998 г.). «SAP97 связан с альфа-амино-3-гидрокси-5-метилизоксазол-4-пропионовой кислотой субъединицы GluR1» . Журнал биологической химии . 273 (31): 19518–24. doi : 10.1074/jbc.273.31.19518 . PMID 9677374 .
- ^ Jump up to: а беременный Грегер И.Х., Хатри Л., Зифф Эб (май 2002 г.). «Редактирование РНК в ARG607 контролирует выход рецептора AMPA из эндоплазматической ретикулумы» . Нейрон . 34 (5): 759–72. doi : 10.1016/s0896-6273 (02) 00693-1 . PMID 12062022 . S2CID 15936250 .
- ^ Jump up to: а беременный Bats C, Groc L, Choquet D (март 2007 г.). взаимодействием Между Нейрон 53 (5): 719–34. два 10.1016/j.neuron.2007.01.030: 17329211PMID 16423733S2CID
- ^ Boehm J, Kang MG, Johnson RC, Esteban J, Huganir RL, Malinow R (июль 2006 г.). «Синаптическое включение рецепторов AMPA во время LTP контролируется сайтом фосфорилирования PKC на GLUR1» . Нейрон . 51 (2): 213–25. doi : 10.1016/j.neuron.2006.06.013 . PMID 16846856 . S2CID 16208091 .
- ^ Jump up to: а беременный Хаяси Y, Ши Ш.Х., Эстебан Дж.А., Пикцини А., Понсер Дж.С., Малиноу Р (март 2000 г.). «Вождение рецепторов AMPA в синапсы LTP и CAMKII: потребность в взаимодействии домена GluR1 и PDZ». Наука . 287 (5461): 2262–7. Бибкод : 2000SCI ... 287.2262H . doi : 10.1126/science.287.5461.2262 . PMID 10731148 . S2CID 17001488 .
- ^ Derkach V, Barria A, Soderling TR (март 1999 г.). «CA2+/кальмодулин-киназа II усиливает проводимость канала альфа-амино-3-гидрокси-5-метил-4-азоксазолепропионат глутаматные рецепторы» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (6): 3269–74. doi : 10.1073/pnas.96.6.3269 . PMC 15931 . PMID 10077673 .
- ^ Delgado JY, Coba M, Anderson CN, Thompson KR, Grey EE, Heusner CL, et al. (Ноябрь 2007 г.). «Активация активации рецептора NMDA дефосфорилирует субъединицы 1 рецептора AMPA -рецептора 1 в треонине 840» . Журнал нейробиологии . 27 (48): 13210–21. doi : 10.1523/jneurosci.3056-07.2007 . PMC 2851143 . PMID 18045915 .
- ^ Банке Т.Г., Боуи Д., Ли Х, Аганир Р.Л., Шусбо А., Трейнелис С.Ф. (январь 2000 г.). «Контроль функции рецептора AMPA GLUR1 с помощью CAMP-зависимой протеинкиназы» . Журнал нейробиологии . 20 (1): 89–102. doi : 10.1523/jneurosci.20-01-00089.2000 . PMC 6774102 . PMID 10627585 .
- ^ Армстронг Н., Сан Y, Чен Г.К., Гуаукс Э (октябрь 1998 г.). «Структура глутаматного лиганд-связывающего ядра в комплексе с каинатом». Природа . 395 (6705): 913–7. Bibcode : 1998natur.395..913a . doi : 10.1038/27692 . PMID 9804426 . S2CID 4405926 .
- ^ Jump up to: а беременный Платт С.Р. (март 2007 г.). «Роль глутамата в здоровье и болезнях центральной нервной системы-обзор». Ветеринарный журнал . 173 (2): 278–86. doi : 10.1016/j.tvjl.2005.11.007 . PMID 16376594 .
- ^ Rosenmund C, Stern-Bach Y, Stevens CF (июнь 1998 г.). «Тетрамерная структура канала рецептора глутамата». Наука . 280 (5369): 1596–9. Bibcode : 1998sci ... 280.1596r . doi : 10.1126/science.280.5369.1596 . HDL : 11858/00-001M-0000-0012-FDD8-B . PMID 9616121 .
- ^ Армстронг Н., Джасти Дж., Бейх-Франдсен М., Гуаукс Э (октябрь 2006 г.). «Измерение конформационных изменений, сопровождающих десенсибилизацию в ионотропном рецепторе глутамата» . Клетка . 127 (1): 85–97. doi : 10.1016/j.cell.2006.08.037 . PMID 17018279 . S2CID 16564029 .
- ^ Ким Да, Ким Ш., Чой Х.Б., Мин С., Гваг Б.Дж. (июнь 2001 г.). «Высокая изобилие мРНК GLUR1 и снижение редактирования Q/R мРНК GluR2 в отдельных нейронах NADPH-диафоразы». Молекулярные и клеточные нейронауки . 17 (6): 1025–33. doi : 10.1006/mcne.2001.0988 . PMID 11414791 . S2CID 15351461 .
- ^ Jump up to: а беременный Кумар, Санджай С.; Бакчи, Альберто; Харазия, Виктор; Хугенард, Джон Р. (2002-04-15). «Переключение развития субъединиц рецептора AMPA в неокортикальных пирамидальных нейронах» . Журнал нейробиологии . 22 (8): 3005–3015. doi : 10.1523/jneurosci.22-08-03005.2002 . ISSN 1529-2401 . PMC 6757523 . PMID 11943803 .
- ^ Люшер С., Маленка Р.К. (февраль 2011 г.). «Возникающая наркотики синаптическая пластичность в зависимости: от молекулярных изменений до ремоделирования цепи» . Нейрон . 69 (4): 650–63. doi : 10.1016/j.neuron.2011.01.017 . PMC 4046255 . PMID 21338877 .
- ^ Herbrechter R, Hube N, Buchholz R, Reiner A (июль 2021 г.). «Сплайсинг и редактирование ионотропных глутаматных рецепторов: всесторонний анализ, основанный на данных RNA-seq человека» . Клеточные и молекулярные науки о жизни . 78 (14): 5605–5630. doi : 10.1007/s00018-021-03865-z . PMC 8257547 . PMID 34100982 .
- ^ Mosbacher J, Schoepfer R, Monyer H, Burnashev N, Seeburg PH, Ruppersberg JP (ноябрь 1994 г.). «Молекулярная детерминант для субмиллисекундной десенсибилизации в глутаматных рецепторах». Наука . 266 (5187): 1059–62. Bibcode : 1994sci ... 266.1059m . doi : 10.1126/science.7973663 . PMID 7973663 .
- ^ Sommer B, Keinänen K, Verdoorn TA, Wisden W, Burnashev N, Herb A, et al. (Сентябрь 1990). «Flip and Flop: специфичный функциональный переключатель в глутамате каналах ЦНС». Наука . 249 (4976): 1580–5. Bibcode : 1990sci ... 249.1580s . doi : 10.1126/science.1699275 . PMID 1699275 .
- ^ Pei W, Huang Z, Niu L (февраль 2007 г.). «Glur3 Flip and Flop: различия в кинетике открытия канала». Биохимия . 46 (7): 2027–36. doi : 10.1021/bi062213s . PMID 17256974 .
- ^ Иствуд С.Л., Бернет П.В., Харрисон П.Дж. (февраль 1997 г.). «Изоформы FLIP и флопа-глутамата GluR2-глутамата снижаются в образовании гиппокампа при шизофрении: исследование обратной транскриптазы-полимеразной цепной реакции (RT-PCR)». Исследование мозга. Молекулярное исследование мозга . 44 (1): 92–8. doi : 10.1016/s0169-328x (96) 00195-7 . PMID 9030702 .
- ^ Whitlock JR, Heynen AJ, Shuler MG, Bear MF (август 2006 г.). «Обучение вызывает долгосрочное потенцирование в гиппокампе». Наука . 313 (5790): 1093–7. Bibcode : 2006sci ... 313.1093W . doi : 10.1126/science.1128134 . PMID 16931756 . S2CID 612352 .
- ^ Maren S, Tocco G, Standley S, Baudry M, Thompson RF (октябрь 1993 г.). «Постсинаптические факторы в экспрессии долгосрочного потенцирования (LTP): повышенное связывание рецептора глутамата после индукции LTP in vivo» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (20): 9654–8. Bibcode : 1993pnas ... 90.9654M . doi : 10.1073/pnas.90.20.9654 . PMC 47628 . PMID 8415757 .
- ^ Perkinton MS, Sihra TS, Williams RJ (июль 1999 г.). «CA (2+)-Проницаемые рецепторы AMPA индуцируют фосфорилирование элемента ответа CAMP-элемента, связывающего белок, посредством фосфатидилинозитол 3-киназы, зависимой стимуляции митоген-активированного сигнального каскада протеинкиназы в нейронах» . Журнал нейробиологии . 19 (14): 5861–74. doi : 10.1523/jneurosci.19-14-05861.1999 . PMC 6783096 . PMID 10407026 .
- ^ Fukunaga K, Stoppini L, Miyamoto E, Muller D (апрель 1993 г.). «Долгосрочное потенцирование связано с повышенной активностью Ca2+/кальмодулин-зависимой протеинкиназы II» . Журнал биологической химии . 268 (11): 7863–7. doi : 10.1016/s0021-9258 (18) 53037-4 . PMID 8385124 .
- ^ Лисман Дж., Шульман Х, Клайн Х (март 2002 г.). «Молекулярная основа функции CAMKII в синаптической и поведенческой памяти». Природные обзоры. Нейробиология . 3 (3): 175–90. doi : 10.1038/nrn753 . PMID 11994750 . S2CID 5844720 .
- ^ Маммен Ал, Камеяма К., Рош К.В., Аганир Р.Л. (декабрь 1997 г.). «Фосфорилирование альфа-амино-3-гидрокси-5-метилсоксазол4-пропионовой кислоты субъединицы Glur1 с помощью кальция/кальмодулин-зависимой киназы II» . Журнал биологической химии . 272 (51): 32528–33. doi : 10.1074/jbc.272.51.32528 . PMID 9405465 .
- ^ Ebert DH, Greenberg Me (январь 2013 г.). «Зависимая от активности передача сигналов нейронов и расстройство аутистического спектра» . Природа . 493 (7432): 327–37. Bibcode : 2013natur.493..327e . doi : 10.1038/nature11860 . PMC 3576027 . PMID 23325215 .
- ^ Mauceri D, Cattabeni F, Di Luca M, Gardoni F (май 2004). «Кальций/кальмодулин-зависимая протеинкиназа II фосфорилирование приводит к синапс-ассоциированному белке 97 в шипии» . Журнал биологической химии . 279 (22): 23813–21. doi : 10.1074/jbc.m402796200 . PMID 15044483 .
- ^ Wu H, Nash JE, Zamorano P, Garner CC (август 2002 г.). «Взаимодействие SAP97 с минус-контрольным актиновым моторным миозином VI. Последствия для оборота рецепторов AMPA» . Журнал биологической химии . 277 (34): 30928–34. doi : 10.1074/jbc.m203735200 . PMID 12050163 .
- ^ Чжу Дж.Дж., Цинь Й, Чжао М., Ван Аэлст Л., Малиноу Р. (август 2002 г.). «RAS и RAP -контроль AMPA RECEPTOR RESPICKING во время синаптической пластичности» . Клетка . 110 (4): 443–55. doi : 10.1016/s0092-8674 (02) 00897-8 . PMID 12202034 . S2CID 12858091 .
- ^ Jump up to: а беременный Borgdorff AJ, Choquet D (июнь 2002 г.). «Регуляция боковых движений рецептора AMPA». Природа . 417 (6889): 649–53. Bibcode : 2002natur.417..649b . doi : 10.1038/nature00780 . PMID 12050666 . S2CID 4422115 .
- ^ Park M, Penick EC, Edwards JG, Kauer JA, Ehlers MD (сентябрь 2004 г.). «Утилизация эндосомов снабжает рецепторы AMPA для LTP». Наука . 305 (5692): 1972–5. Bibcode : 2004sci ... 305.1972p . doi : 10.1126/science.1102026 . PMID 15448273 . S2CID 34651431 .
- ^ Макино Х, Малиноу Р (ноябрь 2009 г.). «Включение рецепторов AMPA в синапсы во время LTP: роль бокового движения и экзоцитоза» . Нейрон . 64 (3): 381–90. doi : 10.1016/j.neuron.2009.08.035 . PMC 2999463 . PMID 19914186 .
- ^ Говард М.А., Элиас Г.М., Элиас Л.А., Сват В., Николл Р.А. (февраль 2010 г.). «Роль SAP97 в динамике синаптического глутаматного рецептора» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (8): 3805–10. Bibcode : 2010pnas..107.3805h . doi : 10.1073/pnas.0914422107 . PMC 2840522 . PMID 20133708 .
- ^ Waites CL, Specht CG, Härtel K, Leal-Ortiz S, Genoux D, Li D, et al. (Апрель 2009 г.). «Синаптические изоформы SAP97 регулируют динамику рецептора AMPA и доступ к пресинаптическому глутамату» . Журнал нейробиологии . 29 (14): 4332–45. doi : 10.1523/jneurosci.4431-08.2009 . PMC 3230533 . PMID 19357261 .
- ^ Wang Z, Edwards JG, Riley N, Provance DW, Karcher R, Li XD, et al. (Октябрь 2008 г.). «Myosin VB мобилизует эндосомы переработки и рецепторы AMPA для постсинаптической пластичности» . Клетка . 135 (3): 535–48. doi : 10.1016/j.cell.2008.09.057 . PMC 2585749 . PMID 18984164 .
- ^ Nicoll RA, Tomita S, Bredt DS (март 2006 г.). «Вспомогательные субъединицы помогают рецепторам глутамата AMPA типа». Наука . 311 (5765): 1253–6. Bibcode : 2006sci ... 311.1253n . doi : 10.1126/science.1123339 . PMID 16513974 . S2CID 40782882 .
- ^ Томита С., Чен Л., Кавасаки Ю., Петалия Р.С., Уэдхолд Р.Дж., Николл Р.А., Бредт Д.С. (май 2003 г.). «Функциональные исследования и распределение определяют семейство трансмембранных регуляторных белков рецептора AMPA» . Журнал клеточной биологии . 161 (4): 805–16. doi : 10.1083/jcb.200212116 . PMC 2199354 . PMID 12771129 .
- ^ Чен Л., Четкович Д.М., Петралия Р.С., Суини Н.Т., Кавасаки Ю., Уэдхолд Р.Дж. и др. (2000). «Stargazin регулирует синаптическое нацеливание рецепторов AMPA двумя различными механизмами». Природа . 408 (6815): 936–43. Bibcode : 2000natur.408..936c . doi : 10.1038/35050030 . PMID 11140673 . S2CID 4427689 .
- ^ R, Lounis B, RL, RL, Cognet L, Choquequet D (апрель 2008 г.). Синаптическая передача » . Наука 320 (5873): 201–205. Bibcode : 2008Sci ... 201ch Два : 10,1126/наука . 2715948PMC 18403705PMID
- ^ Hoze N, Nair D, Hosy E, Holcman D (октябрь 2012 г.). «Гетерогенность транспорта AMPA -рецепторов и молекулярных взаимодействий, выявленная в результате анализа суперрезолирования визуализации живых клеток» . Труды Национальной академии наук . 109 (42): 17052–17057. Bibcode : 2012pnas..10917052H . doi : 10.1073/pnas.1204589109 . PMC 3479500 . PMID 23035245 .
- ^ Jump up to: а беременный в Shepherd JD, Huganir RL (2007). «Клеточная биология синаптической пластичности: транспортировка рецепторов AMPA». Ежегодный обзор биологии клеток и развития . 23 : 613–43. doi : 10.1146/annurev.cellbio.23.090506.123516 . PMID 17506699 . S2CID 7048661 .
- ^ Jump up to: а беременный в Малиноу Р., Мейнен З.Ф., Хаяси Ю (июнь 2000 г.). «Механизмы LTP: от молчания до четырехполосного движения». Современное мнение о нейробиологии . 10 (3): 352–7. doi : 10.1016/s0959-4388 (00) 00099-4 . PMID 10851179 . S2CID 511079 .
- ^ Маленка RC (ноябрь 2003 г.). «Синаптическая пластичность и торговля рецепторами AMPA». Анналы нью -йоркской академии наук . 1003 (1): 1–11. Bibcode : 2003nyasa1003 .... 1m . doi : 10.1196/annals.1300.001 . PMID 14684431 . S2CID 22696062 .
- ^ Kessels HW, Malinow R (февраль 2009 г.). «Синаптический рецептор AMPA Пластичность и поведение» . Нейрон . 61 (3): 340–50. doi : 10.1016/j.neuron.2009.01.015 . PMC 3917551 . PMID 19217372 .
- ^ McComack SG, Stornetta RL, Zhu JJ (апрель 2006 г.). «Синаптический обмен рецепторами AMPA поддерживает двунаправленную пластичность» . Нейрон . 50 (1): 75–88. doi : 10.1016/j.neuron.2006.02.027 . PMID 16600857 . S2CID 17478776 .
- ^ Zhu JJ, Esteban JA, Hayashi Y, Malinow R (ноябрь 2000). «Постнатальное синаптическое потенцирование: доставка глюр4-содержащих рецепторов AMPA с помощью спонтанной активности». Nature Neuroscience . 3 (11): 1098–106. doi : 10.1038/80614 . HDL : 10261/47079 . PMID 11036266 . S2CID 16116261 .
- ^ Passafaro M, Piëch V, Sheng M (сентябрь 2001 г.). «Специфичные для субъединицы временные и пространственные паттерны экзоцитоза AMPA-рецептора в нейронах гиппокампа». Nature Neuroscience . 4 (9): 917–26. doi : 10.1038/nn0901-917 . PMID 11528423 . S2CID 32852272 .
- ^ Song I, Kamboj S, Xia J, Dong H, Liao D, Huganir RL (август 1998 г.). «Взаимодействие N-этилмалеимид-чувствительного фактора с рецепторами AMPA» . Нейрон . 21 (2): 393–400. doi : 10.1016/s0896-6273 (00) 80548-6 . PMID 9728920 .
- ^ Остен П., Шривастава С., Инман Г.Дж., Вилим Ф.С., Хатри Л., Ли Л.М. и др. (Июль 1998 г.). «Термин Clur2 C-рецептора AMPA может опосредовать обратимое АТФ-зависимое взаимодействие с NSF и альфа-и бета-SNAP» . Нейрон . 21 (1): 99–110. doi : 10.1016/s0896-6273 (00) 80518-8 . PMID 9697855 . S2CID 18569829 .
- ^ Nishimune A, Isaac JT, Molnar E, Noel J, Nash Sr, Tagaya M, et al. (Июль 1998 г.). «Связывание NSF с GLUR2 регулирует синаптическую передачу». Нейрон . 21 (1): 87–97. doi : 10.1016/s0896-6273 (00) 80517-6 . HDL : 2433/180867 . PMID 9697854 . S2CID 18956893 .
- ^ Beretta F, Sala C, Saglietti L, Hirling H, Sheng M, Passafaro M (апрель 2005 г.). «Взаимодействие NSF важно для прямой вставки GLUR2 в синаптических местах». Молекулярные и клеточные нейронауки . 28 (4): 650–60. doi : 10.1016/j.mcn.2004.11.008 . PMID 15797712 . S2CID 46716417 .
- ^ Cingolani LA, Thalhammer A, Yu LM, Catalano M, Ramos T, Colicos MA, Goda Y (июнь 2008 г.). «Зависимая от активности регуляция состава и численности рецептора синаптического AMPA с помощью бета3 интегринов» . Нейрон . 58 (5): 749–62. doi : 10.1016/j.neuron.2008.04.011 . PMC 2446609 . PMID 18549786 .
- ^ Saglietti L, Dequidt C, Kamieniarz K, Rousset MC, Valnegri P, Thoumine O, et al. (Май 2007). «Внеклеточные взаимодействия между GluR2 и N-кадгерином в регуляции позвоночника» . Нейрон . 54 (3): 461–77. doi : 10.1016/j.neuron.2007.04.012 . PMID 17481398 . S2CID 14600986 .
- ^ Сильверман Дж. Б., Реституито С., Лу В., Ли-Эдвардс Л., Хатри Л., Зифф Э.Б. (август 2007 г.). «Синаптическое закрепление рецепторов AMPA кадгеринами через нервные белковые белки, связанные с нейронным плакофилином AMPA-рецептор-связывающие белковые комплексы» . Журнал нейробиологии . 27 (32): 8505–16. doi : 10.1523/jneurosci.1395-07.2007 . PMC 6672939 . PMID 17687028 .
- ^ Кэрролл Р.К., Битти Е.К., Ся Х, Люшер С., Альтчулер Ю., Николл Р.А. и др. (Ноябрь 1999). «Динамин-зависимый эндоцитоз ионотропных глутаматных рецепторов» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (24): 14112–7. Bibcode : 1999pnas ... 9614112c . doi : 10.1073/pnas.96.24.14112 . PMC 24199 . PMID 10570207 .
- ^ Битти Э.К., Кэрролл Р.К., Ю Х, Моришита В., Ясуда Х, фон Застроу М., Маленка Р.К. (декабрь 2000 г.). «Регуляция эндоцитоза рецептора AMPA путем сигнального механизма, разделяемого с LTD» . Nature Neuroscience . 3 (12): 1291–300. doi : 10.1038/81823 . PMID 11100150 .
- ^ Lai MM, Hong JJ, Ruggiero AM, Burnett PE, Slepnev VI, De Camilli P, Snyder SH (сентябрь 1999 г.). «Комплекс кальциневрина-динамина 1 в качестве датчика кальция для эндоцитоза синаптических пузырьков» . Журнал биологической химии . 274 (37): 25963–6. doi : 10.1074/jbc.274.37.25963 . PMID 10473536 .
- ^ Юнг Н., Хауке В. (сентябрь 2007 г.). «Опосредованный клатрином эндоцитоз в синапсах» . Трафик . 8 (9): 1129–36. doi : 10.1111/j.1600-0854.2007.00595.x . PMID 17547698 .
- ^ Daw Mi, Chittajallu R, Bortolotto Za, Dev KK, Duprat F, Henley JM, et al. (Декабрь 2000 г.). «Белки PDZ, взаимодействующие с C-концевым GLUR2/3, участвуют в PKC-зависимой регуляции рецепторов AMPA в синапсах гиппокампа». Нейрон . 28 (3): 873–86. doi : 10.1016/s0896-6273 (00) 00160-4 . HDL : 2262/89240 . PMID 11163273 . S2CID 13727678 .
- ^ Элерс MD (ноябрь 2000 г.). «Пересмотр или деградация рецепторов AMPA, определяемых в зависимости от активности эндоцитарной сортировки» . Нейрон . 28 (2): 511–25. doi : 10.1016/s0896-6273 (00) 00129-x . PMID 11144360 . S2CID 16333109 .
- ^ Lu W, Ziff EB (август 2005 г.). «PICK1 взаимодействует с ABP/GRIP для регулирования транспорта рецепторов AMPA» . Нейрон . 47 (3): 407–21. doi : 10.1016/j.neuron.2005.07.006 . PMID 16055064 . S2CID 17100359 .
- ^ Ван Ит (март 2008 г.). «Исследование роли эндоцитоза AMPAR и долгосрочной депрессии в поведенческой сенсибилизации: отношение к лечению нарушений мозга, включая наркоманию» . Британский журнал фармакологии . 153 Suppl 1 (S1): S389-95. doi : 10.1038/sj.bjp.0707616 . PMC 2268058 . PMID 18059315 .
- ^ Rogakeski MA (2013). «Рецепторы AMPA как молекулярная мишень при терапии эпилепсией» . Acta Scandinavica. Добавка . 127 (197): 9-18. Doi : 10.1111 / ane.12099 . PMC 4506648 . PMID 23480151 .
- ^ Фрич Б., Рейс Дж., Гасиор М., Камински Р.М., Рогавски Ма (апрель 2014 г.). «Роль рецепторов каинат Gluk1 в судорогах, эпилептических разрядах и эпилептогенеза» . Журнал нейробиологии . 34 (17): 5765–75. doi : 10.1523/jneurosci.5307-13.2014 . PMC 3996208 . PMID 24760837 .
- ^ BiAler M, Johannessen SI, Kupferberg HJ, Levy RH, Perucca E, Tomson T (январь 2007 г.). «Отчет о прогрессе по новым антипилептическим препаратам: краткое изложение восьмой конференции Эйлат (EILAT VIII)». Исследование эпилепсии . 73 (1): 1–52. doi : 10.1016/j.eplepsyres.2006.10.008 . PMID 17158031 . S2CID 45026113 .
- ^ Французский JA, Krauss GL, Biton V, Squillacote D, Yang H, Laurenza A, et al. (Август 2012 г.). «Дополнительная Perampanel для рефрактерных приступов с частичным началом: рандомизированное исследование III фазы 304» . Неврология . 79 (6): 589–96. doi : 10.1212/wnl.0b013e3182635735 . PMC 3413761 . PMID 22843280 .
- ^ Рогавский Ма (март 2011 г.). «Пересмотр рецепторов AMPA в качестве мишени для противоэпилептического лекарственного средства» . Эпилепсия токи . 11 (2): 56–63. doi : 10.5698/1535-7511-11.2.56 . PMC 3117497 . PMID 21686307 .
- ^ Сакай Ф., Игараши Х., Сузуки С., Тазаки Ю (1989). «Церебральный кровоток и гематокрит мозга у пациентов с церебральной ишемией, измеренной с помощью однофотонной эмиссионной компьютерной томографии» . Acta Neurologica Scandinavica. Дополнение . 127 : 9–13. doi : 10.1111/j.1600-0404.1989.tb01805.x . PMID 2631521 . S2CID 30934688 .
- ^ Jump up to: а беременный в Чанг П., Августин К., Боддум К., Уильямс С., Сан М., Тершак Дж. А. и др. (Февраль 2016 г.). «Контроль захвата декановой кислотой посредством прямого ингибирования рецептора AMPA» . Мозг . 139 (Pt 2): 431–43. doi : 10.1093/brain/awv325 . PMC 4805082 . PMID 26608744 .
- ^ Августин, Катрин; Уильямс, Софи; Каннингем, Марк; Девлин, Анита М.; Фридрих, Максимилиан; Jayasekera, Ashan; Хуссейн, Мухаммед А.; Холлиман, Дамиан; Митчелл, Патрик; Дженкинс, Алистер; Чен, Филипп Э.; Уокер, Мэтью С.; Уильямс, Робин С.Б. (2018). «Перампанель и деканоевая кислота демонстрируют синергетическое действие против <scp> ampa </scp> рецепторов и судорог» . Эпилепсия . 59 (11): E172 - E178. doi : 10.1111/epi.14578 . PMID 30324610 .
- ^ Cao W, Shah HP, Glushakov AV, Mecca AP, Shi P, Sumners C, et al. (Декабрь 2009 г.). «Эффективность 3,5-дибром-L-фениланина в моделях инсульта крыс, судорог и дефицита сенсомоторного стробирования » Британский журнал фармакологии 158 (8): 2005–1 Doi : 10.1111/ j.1476-5381.2009.00498.x 2807662PMC 20050189PMID
- ^ Cao W, Glushakov A, Shah HP, Mecca AP, Sumners C, Shi P, et al. (Апрель 2011). «Галогенированная ароматическая аминокислота 3,5-дибром-D:-тирозин оказывает полезное влияние на экспериментальный инсульт и судороги» . Аминокислоты . 40 (4): 1151–8. doi : 10.1007/s00726-010-0739-4 . PMC 8396070 . PMID 20839013 . S2CID 19852158 .
- ^ Murray TK, Whalley K, Robinson CS, Ward MA, Hicks CA, Lodge D, et al. (Август 2003 г.). «LY503430, новый потенциатор рецептора рецептора альфа-амино-3-гидрокси-5-метилизоксазола-4-пропионовой кислоты с функциональным, нейропротекторным и нейротрофическим эффектом на моделях болезни Паркинсона». Журнал фармакологии и экспериментальной терапии . 306 (2): 752–62. doi : 10.1124/jpet.103.049445 . PMID 12730350 . S2CID 86751458 .
- ^ О'Нил М.Дж., Бликман Д., Циммерман Д.М., Нисенбаум Э.С. (июнь 2004 г.). «Потиаторы рецепторов AMPA для лечения расстройств CNS». Современные цели наркотиков. ЦНС и неврологические расстройства . 3 (3): 181–94. doi : 10.2174/1568007043337508 . PMID 15180479 .
- ^ Yarotskyy V, Glushakov AV, Sumners C, Gravenstein N, Dennis DM, Seubert CN, Martynyuk AE (май 2005). «Дифференциальная модуляция глутаматергической передачи 3,5-дибром-L-фенилаланином». Молекулярная фармакология . 67 (5): 1648–54. doi : 10.1124/моль.104.005983 . PMID 15687225 . S2CID 11672391 .
- ^ Ветер, щедрый; Ши, Сонг-Хай; Стивен, Хосе А.; Пиччини, Антонелла; Pancer, Жан-Кристоф; Хорошо, Роберто (24 марта 2000 г.). 287; Наука Наука 287 (5461): 2262–2 Бибкод : 2000SCI . doi : 10.1126/ Science PMID 10731148 .
- ^ Tazerart S, Mitchell DE, Miranda-Rottmann S, Araya R (август 2020 г.). «Правило пластичности, зависящего от шипа, для дендритных шипов» . Природная связь . 11 (1): 4276. Bibcode : 2020natco..11.4276t . doi : 10.1038/s41467-020-17861-7 . PMC 7449969 . PMID 32848151 .