Геликаза
Геликазы - это класс ферментов, которые, как считается, жизненно важны для всех организмов . организма Их основная функция - распаковать генетический материал . Геликазы представляют собой моторные белки, которые направляются в направлении вдоль нуклеиновой кислоты основной цепи фосфодиэстера , отделяя две гибридизованные ниши нуклеиновых кислот (отсюда и Helic - + -са ), используя энергию из АТФ гидролиза . Существует много геликаз, представляющих большое разнообразие процессов, в которых необходимо катализировать разделение цепи. Приблизительно 1% эукариотических генов кодируют геликазы. [ 1 ]
Человеческий геном кодирует для 95 не избыточных геликаз: 64 РНК-геликазы и 31 ДНК-геликазы. [ 2 ] Многие клеточные процессы, такие как репликация ДНК , транскрипция , трансляция , рекомбинация , репарация ДНК и биогенез рибосом , включают разделение цепей нуклеиновых кислот, которые требуют использования геликаз. Некоторые специализированные геликазы также участвуют в зондировании вирусных нуклеиновых кислот во время инфекции и выполняют иммунологическую функцию. Геликаза - это фермент, который играет решающую роль в процессах репликации и восстановления ДНК. Его первичная функция-расслабиться на двухцепочечной молекуле ДНК, разбивая водородные связи между комплементарными парами оснований, что позволяет отделить цепи ДНК. Это создает репликацию вилки, которая служит шаблоном для синтеза новых пряди ДНК. Геликаза является важным компонентом клеточных механизмов, который обеспечивает точную репликацию ДНК и поддержание генетической информации. ДНК геликаза катализирует регрессию. Recg и Enzyme Pria работают вместе, чтобы перемотать дуплексную ДНК, создавая холдей. RecG выпускает связанные белки, а PRIA Helicase облегчает перезагрузку ДНК, чтобы возобновить репликацию ДНК. RecG заменяет одноцепочечный связывающий белок (SSB), который регулирует сайты загрузки геликазы-фаркса во время регрессии вилки. Белок SSB взаимодействует с DNA -геликазами PRIA и RecG, чтобы восстановить остановленные вилки репликации ДНК. Эти ферменты должны связываться с SSB-Helicase, чтобы быть загруженным на застопорившие вилки. Тепловой скольжение и дуплексное связывание ДНК, возможно, поддерживаются клиновой доменом ассоциации RecG с линкером SSB. В регрессионной реакции, облегченной Recg и AtphollidayJunctions, создаются для последующей обработки.
Функция
[ редактировать ]Геликазы часто используются для разделения цепей ДНК двойной спирали или самостоятельной молекулы РНК с использованием энергии из гидролиза АТФ , процесса, характеризующегося разрывом водородных связей между отожженными нуклеотидными основаниями . Они также функционируют для удаления белков, связанных с нуклеиновой кислотой, и катализируют гомологичную рекомбинацию ДНК . [ 3 ] Метаболические процессы РНК, такие как трансляция, транскрипция, биогенез рибосом , сплайсинг РНК , транспорт РНК, редактирование РНК и деградацию РНК, облегчены геликазами. [ 3 ] Геликазы постепенно перемещаются вдоль одной нуклеиновой прядью дуплекс с направлением и процессивностью, специфичными для каждого конкретного фермента.
Геликазы принимают разные структуры и состояния олигомеризации . В то время как DNAB -подобные геликазы расстреляют ДНК в форме кольца в качестве гексамеров , было показано, что другие ферменты активны в качестве мономеров или димеров . Исследования показали, что геликазы могут действовать пассивно, ожидая, когда не будет проведено незаметное раскручение, а затем транслокация между перемещенными прядями, [ 4 ] или может играть активную роль в катализировании разделения цепей, используя энергию, генерируемую в гидролизе АТФ. [ 5 ] В последнем случае геликаза действует сравнительно с активным двигателем, раскручиваясь и транслокацию вдоль своего субстрата в качестве прямого результата своей активности АТФазы. [ 6 ] Геликазы могут обрабатывать гораздо быстрее in vivo, чем in vitro из -за наличия вспомогательных белков, которые помогают в дестабилизации развязки вилки. [ 6 ]

Активационный барьер в активности геликазы
[ редактировать ]Ферментативное действие геликазы, такое как разматывание нуклеиновых кислот, достигается за счет снижения барьеры активации ( ) каждого конкретного действия. [ 7 ] [ 5 ] [ 8 ] [ 9 ] Активационный барьер является результатом различных факторов и может быть определен


где
- = количество парных пар размолочных оснований (BPS),
- = свободная энергия формирования пары оснований,
- = снижение свободной энергии из -за геликазы и
- = Снижение свободной энергии из -за поврежденных сил.




Факторы, способствующие высоте барьеры активации, включают в себя: специфическую последовательность нуклеиновой кислоты вовлеченной молекулы, количество участвующих пар оснований, напряжение, присутствующее на вилке репликации и силы дестабилизации. [ 7 ] [ 5 ] [ 8 ] [ 9 ]
Активные и пассивные геликазы
[ редактировать ]Размер активационного барьера для преодоления геликазой способствует его классификации как активной или пассивной геликазы. У пассивных геликаз существует значительный барьер активации (определяется как , где это постоянная Больцманн и температура системы). Из -за этого значительного активационного барьера его раскручивающее прогрессирование в значительной степени влияет последовательность нуклеиновых кислот в молекуле, чтобы расслабиться, и наличие дестабилизационных сил, действующих на вилку репликации. [ 7 ] [ 5 ] [ 8 ] [ 9 ] Некоторые комбинации нуклеиновых кислот будут снижать скорость разматывания (то есть гуанин и цитозин ), в то время как различные дестабилизирующие силы могут увеличить скорость разматывания. [ 5 ] [ 8 ] [ 9 ] В пассивных системах скорость раскручивания ( ) меньше, чем скорость транслокации ( ) (транслокация вдоль одноцепочечной нуклеиновой кислоты, SSNA), из-за ее зависимости от переходного раскрытия пар оснований на вилке репликации, чтобы определить его скорость раскручивания. [ 7 ] [ 5 ] [ 8 ] [ 9 ]





В активных геликазах, , где у системы отсутствует значительный барьер, поскольку геликаза может дестабилизировать нуклеиновые кислоты, раскрывая двойную спираль с постоянной скоростью, независимо от последовательности нуклеиновой кислоты. В активных геликазах, ближе к , из -за активной способности геликазы непосредственно дестабилизировать вилку репликации, чтобы способствовать разматыванию. [ 7 ] [ 5 ] [ 8 ] [ 9 ]



Активные геликазы демонстрируют сходное поведение, когда действуют как на двойные нуклеиновые кислоты, так и на DSNA или SSNA, в отношении скорости раскручивания и скорости транслокации, где в обеих системах и примерно равны.
Эти две категории геликаз также могут быть смоделированы как механизмы. В таких моделях пассивные геликазы концептуализируются в виде бауновых фиксетов, обусловленных тепловыми колебаниями и последующими анизотропными градиентами в решетке ДНК. Активные геликазы, напротив, концептуализируются как ступеньки, также известные как двигатели PowerStroke-используя либо конформационный «червя дюйма», либо механизм «ходьбы» ручной работы для прогресса. [ 10 ] В зависимости от организма, такой прогресс, проводящий спираль, может произойти на скорости вращения в диапазоне 5000 [ 11 ] до 10000 [ 12 ] Rpm
История геликазы ДНК
[ редактировать ]ДНК-геликазы были обнаружены в E.coli в 1976 году. Эта геликаза была описана как «разматывающий фермент ДНК», который «обнаружен денатуру ДНК-дуплексы в АТФ-зависимой реакции, без обнаруженного разложения». [ 13 ] Первая эукариотическая ДНК -геликаза была в 1978 году на растении Лилии. [ 14 ] С тех пор ДНК -геликазы были обнаружены и изолированы у других бактерий, вирусов, дрожжей, мух и более высоких эукариот. [ 15 ] На сегодняшний день, по меньшей мере, 14 различных геликаз были выделены из одноклеточных организмов, 6 геликаз из бактериофагов, 12 из вирусов, 15 из дрожжей, 8 из растений, 11 из тимуса теленка и приблизительно 25 геликаз из клеток человека. [ 16 ] Ниже приведена история обнаружения геликазы:
- 1976 -Открытие и выделение E. coli ДНК -геликазы на основе [ 13 ]
- 1978 - Открытие первых эукариотических ДНК -геликаз, выделенных из растения Лилии [ 14 ]
- 1982 - «Ден гена T4 41» - это первая зарегистрированная бактериофажная ДНК геликаза [ 15 ]
- 1985 - Геликазы первого млекопитающего, выделенные из тимуса теленка [ 17 ]
- 1986 - Большой опухолевой антиген SV40 сообщил о вирусной геликазе (1 -й зарегистрированный вирусный белок, который, как было определено, служит ДНК -геликазой) [ 18 ]
- 1986 - ATPASEIII, дрожжевой белок, определяемый как ДНК -геликаза [ 19 ]
- 1988 - Открытие семи консервативных аминокислотных доменов, определенных как геликазные мотивы
- 1989 - Обозначение суперсемейства ДНК геликазы I и суперсемейства II [ 20 ]
- 1989 - Идентификация семьи Helicase Dead Box [ 21 ]
- 1990 - Выделение человеческой ДНК -геликазы [ 22 ]
- 1992 - Выделение первой зарегистрированной митохондриальной ДНК -геликазы (из бычьего мозга) [ 23 ]
- 1996 - Отчет об обнаружении первой очищенной хлоропластной ДНК -геликазы из гороха [ 24 ]
- 2002 - Выделение и характеристика первого биохимически активного малярийного паразита -геликаза - Plasmodium cynomolgi . [ 25 ]
Структурные особенности
[ редактировать ]Общая функция геликаз учитывает тот факт, что они демонстрируют определенную степень аминокислотной гомологии последовательности ; Все они обладают мотивами последовательностей, расположенными во внутренней части их первичной структуры , участвующей в связывании АТФ , гидролизе АТФ и транслокации вдоль нуклеиновой кислоты субстрата . Переменная часть аминокислотной последовательности связана со специфическими признаками каждой геликазы.
Присутствие этих мотивов геликазы позволяет предположить, что активность геликазы объясняется данным белком, но не обязательно подтверждает его как активную геликазу. Однако консервативные мотивы поддерживают эволюционную гомологию среди ферментов. Основываясь на этих мотивах геликазы, был выделен ряд суперсемейств геликазы.
Суперсемейства
[ редактировать ]Геликазы классифицируются в 6 группах (суперсемей) на основе их общих мотивов последовательности. [ 26 ] Геликазы, не образующие кольцевую структуру, находятся в суперсемействах 1 и 2, а геликазы, образующие кольцо, являются частью суперсемей, 3–6. [ 27 ] Геликазы также классифицируются как α или β в зависимости от того, работают ли они с однократной или двухцепочкой ДНК ; α-геликазы работают с одноцепочечной ДНК , а β-геликазы работают с двойной ДНК . Они также классифицируются по полярности транслокации. Если происходит транслокация 3'-5 ', геликаза-тип A; Если происходит транслокация 5'-3 'Это тип B. [ 26 ]
- Суперсемейство 1 (SF1) : эта суперсемейство может быть дополнительно подразделено на геликазы SF1A и SF1B. [ 26 ] В этой группе геликазы могут иметь либо 3'-5 '(подсемейство SF1A), либо 5'-3' (SF1B-подсемейство) полярность транслокации. [ 26 ] [ 28 ] Наиболее известными геликазами SF1A являются Rep и UVRD в грамотрицательных бактериях и геликазе PCRA из грамположительных бактерий. [ 26 ] Наиболее известными геликазами в группе SF1B являются HELICASES RECD и DDA. [ 26 ] У них есть ядро, похожее на Reca. [ 27 ]
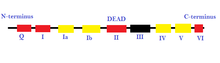
- Суперсемейство 2 (SF2) : Это самая большая группа геликаз, которые участвуют в различных клеточных процессах. [ 26 ] [ 2 ] Они характеризуются наличием девяти консервативных мотивов: Q, I, IA, IB и II через VI. [ 2 ] Эта группа в основном состоит из мертвой РНК-геликаз. [ 27 ] Некоторые другие геликазы, включенные в SF2,-это семейство, подобное Recq, и SNF2-подобные ферменты. [ 26 ] Большинство геликаз SF2 являются типом A за несколькими исключениями, такими как семейство XPD. [ 26 ] У них есть ядро, похожее на Reca. [ 27 ]
- Суперсемейство 3 (SF3) : суперсемейство 3 состоит из AAA+ геликаз, кодируемых в основном небольшими вирусами ДНК и некоторыми крупными нуклеоцитоплазматическими ДНК -вирусами. [ 29 ] [ 30 ] У них есть направление транслокации 3'-5 ', что означает, что все они являются геликазами типа А. [ 26 ] Наиболее известной геликазой SF3 является вирус папилломы E1 Helicase. [ 26 ]
- Суперсемейство 4 (SF4) : все геликазы семейства SF4 имеют полярность типа B (5'-3 '). У них есть складка Reca. [ 26 ] Наиболее изученной SF4 -геликазой является GP4 из бактериофага T7. [ 26 ]
- Суперсемейство 5 (SF5) : белки Rho соответствуют группе SF5. У них есть складка Reca. [ 26 ]
- Суперсемейство 6 (SF6) : они содержат основной AAA+, который не включен в классификацию SF3. [ 26 ] Некоторые белки в группе SF6: поддержание мини -хромосом MCM , RUVB, RUVA и RUVC. [ 26 ]
Все геликазы являются членами Ploop или Walker , содержащей семью.
Геликазы расстройства и болезнь
[ редактировать ]ATRX Helicase Мутации
[ редактировать ]Ген ATRX кодирует АТФ-зависимую геликазу, ATRX (также известный как XH2 и XNP) семейства подгрупп SNF2, который, как считается, отвечает за такие функции, как ремоделирование хроматина, регуляция генов и метилирование ДНК. [ 31 ] [ 32 ] [ 33 ] [ 34 ] Эти функции помогают в профилактике апоптоза, что приводит к регуляции размеров коры, а также к вкладу в выживание гиппокампа и корковых структур, влияющих на память и обучение. [ 31 ] Эта геликаза расположена на X-хромосоме (XQ13.1-Q21.1), в перицентромерном гетерохроматине и связывается с гетерохроматином белка 1 . [ 31 ] [ 33 ] Исследования показали, что ATRX играет роль в метилировании рДНК и имеет важное значение для эмбрионального развития. [ 35 ] Мутации были обнаружены по всему белку ATRX , причем более 90% из них расположены в домене цинковых пальцев и геликазы. [ 36 ] Мутации ATRX могут привести к X-связанной альфа-талассамейской замедлению ( синдром ATR-X ). [ 31 ]
Было обнаружено, что различные типы мутаций, обнаруженных в ATRX, связаны с ATR-X, включая чаще всего одно базовые миссенс-мутации, а также глупость, FrameShift и делеционные мутации. [ 34 ] Характеристики ATR-X включают в себя: микроцефалия, скелетные и лицевые аномалии, умственную отсталость, аномалии половых органов, судороги, ограниченное использование языка и способности, а также альфа-талассемию. [ 31 ] [ 35 ] [ 32 ] Фенотип, наблюдаемый в ATR-X, предполагает, что мутация гена ATRX вызывает подавление экспрессии генов, таких как гены альфа-глобина. [ 32 ] До сих пор неизвестно, что вызывает экспрессию различных характеристик ATR-X у разных пациентов. [ 35 ]
XPD Helicase Point Mutations
[ редактировать ]XPD (ксеродерма пигментный фактор D, также известный как белок ERCC2) представляет собой 5'-3 ', суперсемейство II, АТФ-зависимая геликаза, содержащая железные кластерные домены. [ 26 ] [ 37 ] Было показано, что унаследованные точечные мутации в HPD Helicase связаны с ускоренными нарушениями старения, такими как синдром Кокейна (CS) и трихотиодистрофия (TTD). [ 38 ] Синдром Кокейна и трихотиодистрофия являются нарушениями развития, связанные с чувствительностью к ультрафиолетовому свету и преждевременному старению, а синдром Кокейна демонстрирует тяжелую умственную замедление со времен рождения. [ 38 ] Мутация HPD Helicase также участвовала в ксеродерме Pigmentosum (XP), расстройство, характеризующемся чувствительностью к ультрафиолетовому свету и приводит к нескольким 1000-кратному увеличению развития рака кожи. [ 38 ]
XPD является важным компонентом комплекса TFIIH , фактора транскрипции и восстановления в ячейке. [ 38 ] [ 39 ] [ 40 ] [ 41 ] [ 42 ] В рамках этого комплекса он облегчает восстановление нуклеотидного удаления путем раскручивания ДНК. [ 38 ] TFIIH помогает восстановить поврежденную ДНК, такую как повреждение солнца. [ 38 ] [ 39 ] [ 40 ] [ 41 ] [ 42 ] Мутация в HPD Helicase, которая помогает сформировать этот комплекс и способствует его функции, вызывает чувствительность к солнечному свету, наблюдаемому при всех трех заболеваниях, а также повышенный риск развития рака и преждевременного старения, наблюдаемого при трихотиодистрофии и синдроме Кокайне. [ 38 ]
Мутации HPD Helicase, приводящие к трихотиодистрофии, обнаружены по всему белку в различных местах, участвующих в взаимодействиях белка-белка. [ 38 ] Эта мутация приводит к нестабильному белку из -за его неспособности образовывать стабилизирующие взаимодействия с другими белками в точках мутаций. [ 38 ] Это, в свою очередь, дестабилизирует весь комплекс TFIIH, который приводит к дефектам с механизмами транскрипции и восстановления ячейки. [ 38 ]
Было высказано предположение, что мутации XPD Helicase, приводящие к синдрому Кокейна, могут быть результатом мутаций в пределах XPD, вызывая жесткость белка и последующую неспособность переключаться от функций восстановления на функции транскрипции из -за «блокировки» в режиме восстановления. [ 38 ] Это может привести к тому, что геликаза вырезает сегменты ДНК, предназначенные для транскрипции. [ 38 ] Хотя текущие данные указывают на дефект в HPD Helicase, что приводит к потере гибкости в белке в случаях синдрома Cockayne, до сих пор неясно, как эта структура белка приводит к симптомам, описанным при синдроме Cockayne. [ 38 ]
В Xeroderma pigmentosa мутация HPD Helicase существует в месте связывания АТФ или ДНК. [ 38 ] Это приводит к структурно функциональной геликазе, способной облегчить транскрипцию, однако она ингибирует свою функцию в раскручивании ДНК и репарации ДНК. [ 38 ] Отсутствие способности клетки восстанавливать мутации, такие как мутации, вызванные повреждением солнца, является причиной высокой частоты рака у пациентов с пигментом ксеродермы.
Семейные мутации Recq
[ редактировать ]
Геликазы Recq (3'-5 ') принадлежат к группе геликаз суперсемейства, которые помогают поддерживать стабильность генома и подавлять неподходящую рекомбинацию. [ 43 ] [ 44 ] Недостатки и/или мутации в геликазах семейства CREQ демонстрируют аберрантную генетическую рекомбинацию и/или репликацию ДНК, что приводит к хромосомной нестабильности и общей снижении способности пролиферировать. [ 43 ] Мутации в геликазах семейства Recq BLM, RecQL4 и WRN, которые играют роль в регуляции гомологичной рекомбинации, приводят к синдрому цветущего аутосомно -рецессивных заболеваний (BS), синдрома Ротмунда -Тхосона (RTS) и синдрома Werner (WS - ), соответственно. [ 44 ] [ 45 ]
Синдром Блума характеризуется предрасположенностью к раку с ранним началом со средним возрастом 24 года. [ 44 ] [ 46 ] У пациентов с синдромом Блума наблюдается высокая частота взаимного обмена между родственными хроматидами (SCE) и чрезмерным хромосомным повреждением. [ 47 ] Существуют доказательства того, что BLM играет роль в спасении нарушенной репликации ДНК на репликационных вилках. [ 47 ]
Синдром Вернера - это расстройство преждевременного старения, с симптомами, включая раннее начало атеросклероза и остеопороза и других заболеваний, связанных с возрастом, высокое происхождение саркомы и смерть, часто происходящую от инфаркта миокарда или рака в 4-6 десятилетиях жизни. [ 44 ] [ 48 ] Клетки пациентов с синдромом Вернера демонстрируют снижение репродуктивного срока службы с хромосомными разрывами и транслокациями, а также большие делеции хромосомных компонентов, вызывая нестабильность генома. [ 48 ]
Синдром Ротмунда-Томсона, также известный как врожденная Poikiloderma , характеризуется преждевременным старением, кожей и скелетными аномалиями, сыпи, poikiloderma , ювенильными катарактами и предрасположенностью к раковым заболеваниям, таким как остеосарком. [ 44 ] [ 49 ] Хромосомные перестройки, вызывающие геномную нестабильность, обнаружены в клетках пациентов с синдромом Ротмунда-Томсона. Recq - это семейство ферментов ДНК геликазы, которые встречаются в различных организмах, включая бактерии, археи и эукариоты (как люди). Эти ферменты играют важную роль в метаболизме ДНК во время репликации ДНК, рекомбинации и восстановления. У людей есть пять известных белков Recq Helicase: Recq1, BLM, WRN, Recq4 и Recq5. Мутации в некоторых из этих генов связаны с генетическими нарушениями. Например, мутации в гене BLM вызывают синдром Блума, который характеризуется повышенным риском рака и другими проблемами со здоровьем. [ 50 ] Мутации в гене WRN приводят к синдрому Вернера, состоянию, характеризующемуся преждевременным старением и повышенным риском возрастных заболеваний. Геликазы Recq имеют решающее значение для поддержания геномной стабильности и целостности. Они помогают предотвратить накопление генетических нарушений, которые могут привести к таким заболеваниям, как рак. Целостность генома зависит от семейства DNA -геликазы DECQ, которое включает в себя восстановление ДНК, рекомбинацию, репликацию и процессы транскрипции. Нестабильность генома и раннее старение - это условия, возникающие в результате мутаций в геликазах CREQ человека. [ 51 ] Recq Helicase SGS1 отсутствует в дрожжевых клетках, что делает их полезными моделями для понимания аномалий клеток человека и функции Helicase Recq. [ 52 ] Член семейства Helicase, RecQ1, связан с небольшим количеством необычных генетических расстройств рака у людей. Он участвует в транскрипции, клеточном цикле и репарации ДНК. Согласно недавним исследованиям, миссенс -мутации в гене RecQ1 могут играть роль в развитии семейного рака молочной железы. Геликазы ДНК часто притягиваются к областям повреждения ДНК и необходимы для репликации клеточной ДНК, рекомбинации, восстановления и транскрипции. Химические манипуляции с их молекулярными процессами могут изменить скорость, с которой раковые клетки делятся, а также эффективность транзакций и клеточного гомеостаза. Индуцированная малоколекулярным захватом ДНК-геликаз, типа метаболического белка ДНК, может иметь вредные последствия для быстро пролиферирующих раковых клеток, что может быть эффективным при лечении рака.
Во время мейоза двойные разрывы ДНК и другие повреждения ДНК в хроматиде восстанавливаются гомологичной рекомбинацией с использованием либо сестринской хроматиды , либо гомологичной несосной хроматиды в качестве матрицы. Этот ремонт может привести к кроссоверу (CO) или, чаще, неверновой (NCO) рекомбинантной. В дрожжах Schizosaccharomyces Pombe Fancm -Family DNA Helicase FMI1 направляет образование рекомбинации NCO во время мейоза. [ 53 ] также Helicase rqh1 типа Recq направляет мейотическую рекомбинацию NCO. [ 54 ] Эти геликазы, благодаря своей способности раскручивать промежуточные соединения D-петли , способствуют рекомбинации NCO в процессе отжига, зависящего от синтеза .
На растении Arabidopsis thaliana Fancm Helicase способствует NCO и противодействует образованию рекомбинантов CO. [ 55 ] Другая геликаза, recq4a/b, также независимо снижает Cos. Было высказано предположение, что COS ограничено из -за долгосрочных затрат на рекомбинацию CO, то есть разрыв благоприятной генетической комбинации аллелей, созданных в результате прошлого естественного отбора . [ 55 ]
РНК геликазы
[ редактировать ]
РНК геликазы необходимы для большинства процессов метаболизма РНК, таких как биогенез рибосом , сплайсинг пре-мРНК и инициация трансляции . Они также играют важную роль в восприятии вирусных РНК. [ 56 ] РНК геликазы участвуют в посредничестве противовирусного иммунного ответа, поскольку они могут идентифицировать иностранные РНК у позвоночных. Около 80% всех вирусов представляют собой РНК -вирусы, и они содержат свои собственные РНК -геликазы. [ 57 ] Дефектные РНК-геликазы были связаны с раковыми заболеваниями, инфекционными заболеваниями и нейро-деггенеративными расстройствами. [ 56 ] Некоторые неврологические расстройства, связанные с дефектными РНК-геликазами: амиотрофический боковой склероз , мышечная атрофия позвоночника , спиноцеребеллярная атаксия типа 2 , болезнь Альцгеймера и синдром летальной врожденной контрактуры . [ 57 ]
РНК -геликазы и ДНК -геликазы могут быть найдены вместе во всех суперсемлиях геликазы, за исключением SF6. [ 58 ] [ 59 ] Все эукариотические РНК-геликазы, которые были идентифицированы в настоящее время, представляют собой нетронутые формирование и являются частью SF1 и SF2. С другой стороны, кольцевые РНК-геликазы были обнаружены у бактерий и вирусов. [ 56 ] Однако не все РНК -геликазы проявляют активность геликазы, как определено ферментативной функцией, то есть белки семейства SWI/SNF. Хотя эти белки несут типичные мотивы геликазы, гидролизируйте АТФ в зависимости от нуклеиновой кислоты и построены вокруг ядра геликазы, как правило, не наблюдается разматывающей активности. [ 60 ]
РНК -геликазы, которые проявляют разматывающую активность, характеризовались как минимум двумя разными механизмами: каноническое дуплексное раскрутительное и локальное разделение цепи. Каноническое дуплексное раскручивание - это пошаговое направленное разделение дуплексной цепи, как описано выше, для раскручивания ДНК. Тем не менее, локальное разделение прядей происходит в процессе, в котором фермент геликаза загружается в любом месте вдоль дуплекса. Это обычно помогает одноцепочечная область РНК, а нагрузка фермента сопровождается связыванием АТФ. [ 61 ] Как только геликаза и АТФ связаны, происходит локальное разделение цепи, которое требует связывания АТФ, но не фактического процесса гидролиза АТФ. [ 62 ] Представлено с меньшим количеством пар оснований дуплекс, а затем диссоциирует без дальнейшей помощи фермента. Этот режим разматывания используется Helicases Dead/Deah Box . [ 63 ]
База данных РНК геликазы [ 64 ] В настоящее время доступен онлайн, который содержит полный список РНК -геликаз с такой информацией, как последовательность, структура, биохимические и клеточные функции. [ 56 ]
Диагностические инструменты для измерения геликазы
[ редактировать ]Измерение и мониторинг активности геликазы
[ редактировать ]Различные методы используются для измерения активности геликазы in vitro . Эти методы варьируются от качественных анализов (анализы, которые обычно влекут за собой результаты, которые не включают значения или измерения) до количественных (анализы с численными результатами, которые можно использовать в статистическом и численном анализе). В 1982–1983 годах был разработан первый прямой биохимический анализ для измерения активности геликазы. [ 15 ] [ 65 ] Этот метод был назван «анализом смещения пряди».
- Анализ смещения цепи включает радиоактивную оболочку ДНК -дуплексов. После обработки геликазы одноцепочечная ДНК визуально обнаруживается как отдельная от двойной ДНК с помощью электрофореза на странице без денатурирования . После обнаружения одноцепочечной ДНК количество радиоактивного тега, которая находится на одноцепочечной ДНК, определено количественно, чтобы дать числовое значение для количества двойного разматывания ДНК. Анализ смещения пряди является приемлемым для качественного анализа, его неспособность отобразить результаты в течение более одного времени времени, его потребление времени и его зависимость от радиоактивных соединений для маркировки заслуживает необходимости разработки диагностики, которая может контролировать активность геликазы в режиме реального времени Полем
Позже были разработаны другие методы, которые включали некоторые, если не все следующие: высокопроизводительные механики, использование нерадиоактивной мечения нуклеотидов, более быстрое время реакции/меньшее потребление времени, мониторинг активности геликазы в режиме реального времени (с использованием кинетического измерения вместо этого. конечной точки/анализа единой точки). Эти методологии включают в себя: «метод быстрого потока утоления, анализы на основе флуоресценции, анализы фильтрации, анализ близости сцинтилляции резонанса , анализ передачи энергии резонанса флуоресцентного , анализ, основанный на технологии вспышки, гомогенном анализе погашения флуоресценции, и электрохилуминцент. -Са основаны на геликазе ». [ 16 ] При использовании специализированных математических уравнений можно использовать некоторые из этих анализов для определения того, сколько спаренных нуклеотидов основания геликаза может разбить на гидролиз 1 молекулы АТФ. [ 66 ]
Коммерчески доступные диагностические наборы также доступны. Одним из таких комплектов является диагностический анализ «Trupoint» от Perkinelmer , Inc. Этот анализ представляет собой анализ гашения флуоресценции по времени, который использует технологию Perkinelmer "Signalclimb", которая основана на двух метках, которые связываются в непосредственной близости друг от друга, но на противоположных ДНК пряди. Одна метка - флуоресцентный хелат лантаноида, который служит этикеткой, которая контролируется через достаточный считыватель для считываемости 96/384. Другая этикетка - это молекула органического гасителя. Основой этого анализа является «гашение» или подавление сигнала хелата лантаноида по молекуле органического гасителя, когда они находятся в непосредственной близости - как это было бы, когда ДНК -дуплекс находится в его нативном состоянии. После активности геликазы на дуплексе гасители и лантаноидные этикетки разделяются, когда ДНК раскопана. Эта потеря в близости отрицает способность гашников репрессировать сигнал лантаноида, вызывая обнаруживаемое увеличение флуоресценции, которое является репрезентативным для количества разоблачной ДНК и может использоваться в качестве количественного измерения активности геликазы. Выполнение и использование методов флуоресцентной визуализации с одной молекулой, сосредоточенная на методах, которые включают оптическое захват в сочетании с эпифлуоресцентной визуализацией, а также поверхностной иммобилизации в сочетании с общей визуализацией флуоресценции внутренней отражения. В сочетании с микроканальными проточными клетками и микрофлюидным контролем позволяют отображать и отслеживать отдельные флуоресцентно меченные белки и ДНК-молекулы, обеспечивая измерение раскручивания и транслокации ДНК при разрешении с одной молекулой. [ 67 ]
Определение полярности геликазы
[ редактировать ]Полярность геликазы, которая также считается «направленностью», определяется как направление (характеризуется как 5 '→ 3' или 3 '→ 5') движения геликазы на одной цепи ДНК/РНК, вдоль которого он движется. Это определение полярности жизненно важно у F.Ex. определение того, прикрепляется ли проверенная геликаза к ведущей цепи ДНК или прямой ДНК. Чтобы охарактеризовать эту особенность геликазы, в качестве субстрата используется частично дуплексная ДНК, которая имеет центральную область одноцелевой ДНК с различной длиной дуплексных областей ДНК (одна короткая область, которая проходит 5 '→ 3' и одна более длинная область, которая работает 3 '→ 5') с обеих сторон этого региона. [ 68 ] После того, как геликаза добавлена в эту центральную одноцелевую область, полярность определяется характеристикой на недавно образованной одноцелевой ДНК.
Смотрите также
[ редактировать ]- Хромодомен геликаза ДНК -связывающий белок: CHD1 , CHD1L , CHD2 , CHD3 , CHD4 , CHD5 , CHD6 , CHD7 , CHD8 , CHD9
- Dead Box / Dead / Deah Box Helicase : DDX3X , DDX5 , DDX6 , DDX10 , DDX11 , DDX12 , DDX58 , DHX8 , DHX9 , DHX37 , DHX40 , DHX58
- ASCC3 , BLM , BRIP1 , DNA2 , FBXO18 , FBXO30 , Helb , Hells , Helq , Helz , HFM1 , HLTF , IFIH1 , NAV2 , PIF1 , Recql , RTEL1 , SHPRH , SMARCA4 , Smarcal1 , WRN , WRNIP1
- База данных РНК геликазы
Ссылки
[ редактировать ]- ^ WU Y (2012). "Раскрытие и перемотка: двойные лица геликазы?" Полем Журнал нуклеиновых кислот . 2012 : 140601. DOI : 10.1155/2012/140601 . PMC 3409536 . PMID 22888405 .
- ^ Jump up to: а беременный в Umate P, Tuteja N, Tuteja R (январь 2011 г.). «Обще геном всеобъемлющий анализ геликаз человека» . Коммуникативная и интегративная биология . 4 (1): 118–137. doi : 10.4161/cib.13844 . PMC 3073292 . PMID 21509200 .
- ^ Jump up to: а беременный Patel SS, Donmez I (июль 2006 г.). «Механизмы геликаз» . Журнал биологической химии . 281 (27): 18265–18268. doi : 10.1074/jbc.r60000008200 . PMID 16670085 .
- ^ Lionnet T, Spiering MM, Benkovic SJ, Bensimon D, Croquette V (декабрь 2007 г.). «В реальном времени наблюдение бактериофага T4 GP41 геликаза выявляет разматывающий механизм» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (50): 19790–19795. Bibcode : 2007pnas..10419790L . doi : 10.1073/pnas.0709793104 . PMC 2148377 . PMID 18077411 .
- ^ Jump up to: а беременный в дюймовый и фон глин Johnson DS, Bai L, Smith BY, Patel SS, Wang MD (июнь 2007 г.). «Исследования с одной молекулярной динамикой выявляют динамику разматывания ДНК с помощью кольцевой геликазы T7» . Клетка . 129 (7): 1299–1309. doi : 10.1016/j.cell.2007.04.038 . PMC 2699903 . PMID 17604719 .
- ^ Jump up to: а беременный «Исследователи решают загадку того, как разделяют пряди ДНК» . 2007-07-03 . Получено 2007-07-05 .
- ^ Jump up to: а беременный в дюймовый и Беттертон М.Д., Юлихер Ф. (2005-08-31). «Erratum: открытие двойных цепей нуклеино-кислоты геликазами: активное и пассивное отверстие [Phys. Rev. E 71, 011904 (2005)]» . Физический обзор e . 72 (2): 029906. BIBCODE : 2005FRVE..72B9906B . doi : 10.1103/physreve.72.029906 .
- ^ Jump up to: а беременный в дюймовый и фон Manosas M, Xi XG, Bensimon D, Croquette V (сентябрь 2010). «Активные и пассивные механизмы геликаз» . Исследование нуклеиновых кислот . 38 (16): 5518–5526. doi : 10.1093/nar/gkq273 . PMC 2938219 . PMID 20423906 .
- ^ Jump up to: а беременный в дюймовый и фон Jarillo J, Ibarra B, Cao-García FJ (2021). «Репликация ДНК: in vitro анализ и модели данных манипуляций с одномолекуляцией » . Вычислительный и структурный биотехнологический журнал . 19 : 3765–3778. doi : 10.1016/j.csbj.2021.06.032 . PMC 8267548 . PMID 34285777 .
- ^ Wu, CG и Spies, M.: Обзор: Что такое геликазы? В: Шпионы, М. (ред.): [1] . Springer Science+Business Media, Нью -Йорк, 2013
- ^ «Биохимия Кевина Ахерна (BB 451/551) в Университете штата Орегон» . Oregonstate.edu . Архивировано из оригинала 2021-01-26 . Получено 2024-01-03 .
- ^ 3-D библиотека анимации; Репликация: [2] (продвинутый)
- ^ Jump up to: а беременный Абдель-Монем М., Дюрвальд Х, Хоффманн-Берлинг Х (июнь 1976 г.). «Ферментативное раскручение ДНК. 2. Разделение цепи от АТФ-зависимого ДНК разматывающего фермента» . Европейский журнал биохимии . 65 (2): 441–449. doi : 10.1111/j.1432-1033.1976.tb10359.x . PMID 133023 .
- ^ Jump up to: а беременный Hotta Y, Stern H (май 1978 г.). «ДНК разматывающий белок из мейотических клеток лилия». Биохимия . 17 (10): 1872–1880. doi : 10.1021/bi00603a011 . PMID 207302 .
- ^ Jump up to: а беременный в Venkatesan M, Silver LL, Nossal NG (октябрь 1982). «Белок бактериофага гена 41, необходимый для синтеза РНК -праймеров, также является ДНК -геликазой» . Журнал биологической химии . 257 (20): 12426–12434. doi : 10.1016/s0021-9258 (18) 33731-1 . PMID 6288720 .
- ^ Jump up to: а беременный Tuteja N, Tuteja R (май 2004). «Прокариотические и эукариотические ДНК -геликазы. Основные молекулярные моторные белки для клеточного механизма» . Европейский журнал биохимии . 271 (10): 1835–1848. doi : 10.1111/j.1432-1033.2004.04093.x . PMC 7164108 . PMID 15128294 .
- ^ Hübscher U, Stalder HP (август 1985 г.). «Геликаза млекопитающих» . Исследование нуклеиновых кислот . 13 (15): 5471–5483. doi : 10.1093/nar/13.15.5471 . PMC 321884 . PMID 3162158 .
- ^ Stahl H, Dröge P, Knippers R (август 1986 г.). «ДНК -геликазная активность крупного опухолевого антигена SV40» . Embo Journal . 5 (8): 1939–1944. doi : 10.1002/j.1460-2075.1986.tb04447.x . PMC 1167061 . PMID 3019672 .
- ^ Sugino A, Ryu BH, Sugino T, Naumovski L, Friedberg EC (сентябрь 1986 г.). «Новая ДНК-зависимая АТФаза, которая стимулирует дрожжевую ДНК-полимеразу I и обладает ДНК-несущей активностью» . Журнал биологической химии . 261 (25): 11744–11750. doi : 10.1016/s0021-9258 (18) 67306-5 . PMID 3017945 .
- ^ Gorbalenya AE, Koonin EV, Donchenko AP, Blinov VM (июнь 1989 г.). «Два родственных суперсемит предполагаемых геликаз, участвующих в репликации, рекомбинации, восстановлении и экспрессии геномов ДНК и РНК» . Исследование нуклеиновых кислот . 17 (12): 4713–4730. doi : 10.1093/nar/17.12.4713 . PMC 318027 . PMID 2546125 .
- ^ Linder, P., Lasko, PF, Ashurner, M., Leroy, P., Nielson, PJ, Nishi, K., Schneir, J., Slumsimski, PP (1989) Рождение мертвого бокса. Nature (Лондон) 337, 121-1
- ^ Тутея Н., Тутея Р., Рахман К., Кан Ли, Фаласки А (декабрь 1990 г.). «ДНК -геликаза из клеток человека» . Исследование нуклеиновых кислот . 18 (23): 6785–6792. doi : 10.1093/nar/18.23.6785 . PMC 332732 . PMID 1702201 .
- ^ Hehman GL, Hauswirth WW (сентябрь 1992 г.). «ДНК геликаза из митохондрий млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (18): 8562–8566. Bibcode : 1992pnas ... 89.8562H . doi : 10.1073/pnas.89.18.8562 . PMC 49960 . PMID 1326759 .
- ^ Тутея Н., Фан Т.Н., Тевари К.К. (май 1996). «Очистка и характеристика ДНК-геликазы из хлоропласта гороха, которая транслоцируется в направлении 3'-5 '» . Европейский журнал биохимии . 238 (1): 54–63. doi : 10.1111/j.1432-1033.1996.0054q.x . PMID 8665952 .
- ^ Tuteja R, Malhotra P, Song P, Tuteja N, Chauhan VS (2002). «Выделение и характеристика гомолога EIF-4A из Plasmodium cynomolgi». Молекулярная и биохимическая паразитология . 124 (1–2): 79–83. doi : 10.1016/s0166-6851 (02) 00205-0 . PMID 12387853 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. Singleton MR, Dillingham MS, Wigley DB (2007). «Структура и механизм геликаз и транслоказ нуклеиновой кислоты». Ежегодный обзор биохимии . 76 : 23–50. doi : 10.1146/annurev.biochem.76.052305.115300 . PMID 17506634 .
- ^ Jump up to: а беременный в дюймовый Fairman-Williams Me, Guenther Up, Jankowsky E (июнь 2010 г.). «Геликазы SF1 и SF2: семейные вопросы» . Современное мнение в структурной биологии . 20 (3): 313–324. doi : 10.1016/j.sbi.2010.03.011 . PMC 2916977 . PMID 20456941 .
- ^ Stelter M, Acajjaoui S, McSweeney S, Timmins J (2013). «Структурное и механистическое понимание ДНК раскручиваемой с помощью Deinococcus radiodurans UVRD» . Plos один . 8 (10): E77364. BIBCODE : 2013PLOSO ... 877364S . doi : 10.1371/journal.pone.0077364 . PMC 3797037 . PMID 24143224 .
- ^ Iyer LM, Aravind L, Koonin EV (декабрь 2001 г.). «Общее происхождение четырех разнообразных семейств крупных эукариотических ДНК -вирусов» . Журнал вирусологии . 75 (23): 11720–11734. doi : 10.1128/JVI.75.23.11720-11734.2001 . PMC 114758 . PMID 11689653 .
- ^ Айер Л.М., Лейпе Д.Д., Кунин Э.В., Аравинд Л. (2004). «Эволюционная история и классификация высшего порядка AAA+ ATPases». Журнал структурной биологии . 146 (1–2): 11–31. doi : 10.1016/j.jsb.2003.10.010 . PMID 15037234 .
- ^ Jump up to: а беременный в дюймовый и Ropers HH, Hamel BC (январь 2005 г.). «X-связанная умственная отсталость». Природные обзоры. Генетика . 6 (1): 46–57. doi : 10.1038/nrg1501 . PMID 15630421 . S2CID 427210 .
- ^ Jump up to: а беременный в Gibbons RJ, Picketts DJ, Villard L, Higgs DR (март 1995 г.). «Мутации в предполагаемом глобальном регуляторе транскрипции вызывают X-связанную умственную замедление с альфа-талассемией (синдром ATR-X)» . Клетка . 80 (6): 837–845. doi : 10.1016/0092-8674 (95) 90287-2 . PMID 7697714 .
- ^ Jump up to: а беременный NextProt Online Betloin Database. «Атркс-транскрипционный регулятор ATRX». Получено 12 ноября 2012 года.
- ^ Jump up to: а беременный Пикеттс DJ, Хиггс DR, Bachoo S, Blake DJ, Barell OW, Gibbons RJ (декабрь 1996 г.). «ATRX кодирует нового члена семейства белков SNF2: мутации указывают на общий механизм, лежащий в основе синдрома ATR-X» . Молекулярная генетика человека . 5 (12): 1899–1907. doi : 10.1093/hmg/5.12.1899 . PMID 8968741 .
- ^ Jump up to: а беременный в Gibbons R (май 2006 г.). «Альфа-талассаемия-умственная отставка, x связано» . ОРФАНЕТНЫЙ ЖУРНАЛ РЕДИЧНЫХ БОЛЬШЕ . 1 : 15. DOI : 10.1186/1750-1172-1-15 . PMC 1464382 . PMID 16722615 .
- ^ Стивенсон Р.Е. (1993). Адам MP, Everman DB, Mirzaa GM, Pagon RA, Wallace SE, Bean LJ, Gripp KW, Amemiya A (Eds.). «Альфа-талассемия X-связанный синдром интеллектуальной инвалидности». GenereViews . Сиэтл (Вашингтон): Университет Вашингтона, Сиэтл. PMID 20301622 .
- ^ Рудольф Дж., Руильон С., Шварц-Линек У, Белый М.Ф. (январь 2010 г.). «Геликаза XPD разжигает пузырьковые структуры и не останавливается путем поражения ДНК, удаленных путем восстановления удаления нуклеотида» . Исследование нуклеиновых кислот . 38 (3): 931–941. doi : 10.1093/nar/gkp1058 . PMC 2817471 . PMID 19933257 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а Fan L, Fuss Jo, Cheng QJ, Arvai AS, Hammel M, Roberts VA, et al. (Май 2008 г.). «XPD Helicase Structures and Actood: понимание фенотипов рака и старения от мутаций XPD» . Клетка . 133 (5): 789–800. doi : 10.1016/j.cell.2008.04.030 . PMC 3055247 . PMID 18510924 .
- ^ Jump up to: а беременный Lainé JP, Mocquet V, Egly JM (2006). «TFIIH ферментативная активность в транскрипции и репарации нуклеотидного удаления». Репарация ДНК, часть а . Методы в фермере. Тол. 408. С. 246–263. doi : 10.1016/s0076-6879 (06) 08015-3 . ISBN 9780121828134 Полем PMID 16793373 .
- ^ Jump up to: а беременный TIRODE F, Busso D, Coin F, Egly JM (январь 1999). «Восстановление фактора транскрипции TFIIH: назначение функций для трех ферментативных субъединиц, XPB, XPD и CDK7» . Молекулярная клетка . 3 (1): 87–95. doi : 10.1016/s1097-2765 (00) 80177-x . PMID 10024882 .
- ^ Jump up to: а беременный Sung P, Bailly V, Weber C, Thompson LH, Prakash L, Prakash S (октябрь 1993). «Человеческая ксеродерма пигментная группа D кодирует ДНК -геликазу». Природа . 365 (6449): 852–855. Bibcode : 1993natur.365..852s . doi : 10.1038/365852A0 . PMID 8413672 . S2CID 4334960 .
- ^ Jump up to: а беременный Schefeffer L, Roy R, Hubert S, Moncollin V, Vermeulen W, Hoeijmakers JH, et al. (Апрель 1993 г.). «Ремонтом ДНК геликаза: комбинированный базовый транскрипционный фактор BTF2 (TFIIH)» Наука . 260 (5104): 58–6 Bibcode : 1993sci ... 260 ... 58 с Doi : 10.1126/ science.8465201 8465201PMID
- ^ Jump up to: а беременный Ханада К., Хиксон ID (сентябрь 2007 г.). «Молекулярная генетика расстройств Helicase Recq» . Клеточные и молекулярные науки о жизни . 64 (17): 2306–2322. doi : 10.1007/s00018-007-7121-z . PMC 11136437 . PMID 17571213 . S2CID 29287970 .
- ^ Jump up to: а беременный в дюймовый и Opresko PL, Cheng WH, Bohr VA (апрель 2004 г.). «Биохимия DEC -хеликаза и болезнь человека» . Журнал биологической химии . 279 (18): 18099–18102. doi : 10.1074/jbc.r300034200 . PMID 15023996 .
- ^ Ouyang KJ, Woo Ll, Ellis NA (2008). «Гомологичная рекомбинация и поддержание целостности генома: рак и старение через призму человеческих геликаз». Механизмы старения и развития . 129 (7–8): 425–440. doi : 10.1016/j.mad.2008.03.003 . PMID 18430459 . S2CID 6804631 .
- ^ Ellis NA, Groden J, Ye TZ, Struaghen J, Lennon DJ, Ciocci S, et al. (Ноябрь 1995). «Продукт гена синдрома Bloom гомологичен Recq Helicases» . Клетка . 83 (4): 655–666. doi : 10.1016/0092-8674 (95) 90105-1 . PMID 7585968 .
- ^ Jump up to: а беременный Selak N, Bachrati CZ, Sheveve I, Dietschy T, Van Loon B, Jacob A, et al. (Сентябрь 2008 г.). «Геликаза синдрома Блума (BLM) взаимодействует физически и функционально с P12, самой маленькой субъединицей человеческой ДНК -полимеразной дельты» . Исследование нуклеиновых кислот . 36 (16): 5166–5179. doi : 10.1093/nar/gkn498 . PMC 2532730 . PMID 18682526 .
- ^ Jump up to: а беременный Grey MD, Shen JC, Kamath-Loeb AS, Blank A, Sopher BL, Martin GM, et al. (Сентябрь 1997). «Белок синдрома Вернера является ДНК -геликазой». Природа генетика . 17 (1): 100–103. doi : 10.1038/ng0997-100 . PMID 9288107 . S2CID 20587915 .
- ^ Китао С., Шимамото А., Гото М., Миллер Р.В., Смитсон В.А., Линдор Н.М., Фуруичи Ю (май 1999). «Мутации в RecQL4 вызывают подмножество случаев синдрома Ротмунда-Томсона». Природа генетика . 22 (1): 82–84. doi : 10.1038/8788 . PMID 10319867 . S2CID 195211275 .
- ^ Гупта, Соня Видуши; Шмидт, Кристина Хильдегард (2020-02-18). «Поддержание целостности генома дрожжей с помощью геликазы семейства семейства CREQ» . Гены . 11 (2): 205. doi : 10.3390/genes11020205 . ISSN 2073-4425 . PMC 7074392 . PMID 32085395 .
- ^ Дебнат С., Шарма С. Recq1 Геликаза в геномной стабильности и раке. Гены. 2020: 11. doi: 10.3390/genes11060622], Дебнат С., Шарма С. Recq1 Helicase в геномной стабильности и раке. Гены. 2020: 11. doi: 10.3390/genes11060622].
{{cite journal}}: CITE Journal требует|journal=( помощь ) ; Отсутствует или пусто|title=( Справка ) CS1 Maint: несколько имен: список авторов ( ссылка ) CS1 Maint: числовые имена: список авторов ( ссылка ) - ^ Хендриксон, Вашингтон; Ward, KB (1975-10-27). «Атомные модели для полипептидных костей миогемеритрина и гемритрина» . Биохимическая и биофизическая исследовательская коммуникация . 66 (4): 1349–1356. doi : 10.1016/0006-291x (75) 90508-2 . ISSN 1090-2104 . PMID 5 .
- ^ Лоренц А., Осман Ф., Сан В., Нанди С., Стейнахер Р., Уитби М.К. (июнь 2012 г.). «Ортолог делящихся дрожжей FANCM направляет неверную рекомбинацию во время мейоза» . Наука . 336 (6088): 1585–1588. Bibcode : 2012sci ... 336.1585L . doi : 10.1126/science.1220111 . PMC 3399777 . PMID 22723423 .
- ^ Лоренц А., Мехатс А., Осман Ф., Уитби М.К. (декабрь 2014 г.). «Паралог и медиаторы RAD51/DMC1 выступают против ДНК -геликаз, чтобы ограничить образование гибридной ДНК и способствовать кроссоверам во время мейотической рекомбинации» . Исследование нуклеиновых кислот . 42 (22): 13723–13735. doi : 10.1093/nar/gku1219 . PMC 4267644 . PMID 25414342 .
- ^ Jump up to: а беременный Séguéla-Arnaud M, Crismani W, Larchevêque C, Mazel J, Froger N, Choinard S, et al. (Апрель 2015). «Многочисленные механизмы ограничивают мейотические кроссоверы: TOP3α и два гомологов BLM противодействуют кроссоверам параллельно с FANCM» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (15): 4713–4718. Bibcode : 2015pnas..112.4713S . doi : 10.1073/pnas.1423107112 . PMC 4403193 . PMID 25825745 .
- ^ Jump up to: а беременный в дюймовый Jankowsky A, Guenther Up, Jankowsky E (январь 2011 г.). «База данных РНК геликазы» . Исследование нуклеиновых кислот . 39 (проблема базы данных): D338 - D341. doi : 10.1093/nar/gkq1002 . PMC 3013637 . PMID 21112871 .
- ^ Jankowsky E, Fairman-Williams Me (2010). «Введение в РНК -геликазы: суперсемейства, семьи и основные темы». В Jankowsky E (ред.). РНК геликазы (биомолекулярные науки RSC) . Кембридж, Англия: Королевское химическое общество. п. 5. ISBN 978-1-84755-914-2 .
- ^ Ранджи А., Борис-Лори К. (2010). «РНК -геликазы: новые роли в репликации вируса и врожденного ответа хозяина» . РНК -биология . 7 (6): 775–787. doi : 10.4161/rna.7.6.14249 . PMC 3073335 . PMID 21173576 .
- ^ Jankowsky E (январь 2011 г.). «РНК -геликазы на работе: привязка и перестройка» . Тенденции в биохимических науках . 36 (1): 19–29. doi : 10.1016/j.tibs.2010.07.008 . PMC 3017212 . PMID 20813532 .
- ^ Yang Q, Del Campo M, Lambowitz AM, Jankowsky E (октябрь 2007 г.). «Мертвые белки размалывают дуплексы по локальному разделению цепи» . Молекулярная клетка . 28 (2): 253–263. doi : 10.1016/j.molcel.2007.08.016 . PMID 17964264 .
- ^ Лю Ф., Патнэм А, Янковский Е (декабрь 2008 г.). «Гидролиз АТФ необходим для переработки белка мертвой коробки, но не для дуплексного разматывания» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (51): 20209–20214. Bibcode : 2008pnas..10520209L . doi : 10.1073/pnas.0811115106 . PMC 2629341 . PMID 19088201 .
- ^ Jarmoskaite I, Russell R (2011). «Мертвые белки как РНК геликазы и шапероны» . Wiley междисциплинарные обзоры. РНК . 2 (1): 135–152. doi : 10.1002/wrna.50 . PMC 3032546 . PMID 21297876 .
- ^ "Индекс /" . www.rnahelicase.org . Архивировано из оригинала 2014-12-18 . Получено 2012-12-07 .
- ^ Matson SW, Tabor S, Richardson CC (ноябрь 1983 г.). «Белок гена 4 бактериофага T7. Характеристика активности геликазы» . Журнал биологической химии . 258 (22): 14017–14024. doi : 10.1016/s0021-9258 (17) 44018-x . PMID 6315716 .
- ^ Sarlós K, Gyimesi M, Kovács M (июнь 2012 г.). «Recq Helicase транслоцируется вдоль одноцепочечной ДНК с умеренной процессивностью и плотной механохимической связью» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (25): 9804–9809. Bibcode : 2012pnas..109.9804S . doi : 10.1073/pnas.1114468109 . PMC 3382518 . PMID 22665805 .
- ^ Pavankumar TL, Exell JC, Kowalczykowski SC (1 января 2016 года). «Прямая флуоресцентная визуализация транслокации и раскручивание отдельными геликазами ДНК». Одномолекулярная ферма: флуоресцентные и высокопроизводительные методы . Методы в фермере. Тол. 581. С. 1–32. doi : 10.1016/bs.mie.2016.09.010 . ISBN 9780128092675 Полем PMC 5854184 . PMID 27793277 .
- ^ Borowiec JA (1996). "ДНК геликазы". В Depamphilis ML (ред.). Репликация ДНК в эукариотических клетках . Cold Spring Harbour, NY: Laboratory Press Cold Spring Harbour. С. 545–574. ISBN 978-0-87969-459-3 Полем OCLC 246537432 .
Внешние ссылки
[ редактировать ]- ДНК+геликазы в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)
- РНК+геликазы в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)