Митохондрия
| Митохондрия | |
|---|---|
 Две митохондрии из легочной ткани млекопитающих, демонстрирующие свой матрикс и мембраны, как показано с помощью электронной микроскопии. | |
 Схема митохондрий животных | |
| Подробности | |
| Произношение | / ˌ m aɪ t ə ˈ k ɒ n d r i ə n / [1] |
| Часть | Клетка |
| Идентификаторы | |
| латинский | органелла |
| МеШ | D008928 |
| ФМА | 63835 |
| Анатомические термины микроанатомии | |
| Клеточная биология | |
|---|---|
| Диаграмма клеток животных | |
 Компоненты типичной животной клетки:
|
Митохондрия митохондрии ( мн. ) представляет собой органеллу, обнаруженную в клетках большинства эукариот , таких как животные , растения и грибы . Митохондрии имеют двойную мембранную структуру и используют аэробное дыхание для выработки аденозинтрифосфата (АТФ), который используется в клетке в качестве источника химической энергии . [2] Их открыл Альберт фон Кёлликер в 1857 году. [3] в произвольных мышцах насекомых. » , означающий нитевидную гранулу, Термин « митохондрия был придуман Карлом Бендой в 1898 году. Митохондрия в народе получила прозвище «электростанция клетки» — фраза, популяризированная Филипом Зикевицем в Scientific American 1957 года. одноименной статье в [4]
В некоторых клетках некоторых многоклеточных организмов митохондрии отсутствуют (например, зрелые эритроциты млекопитающих ). Известно, что у многоклеточного животного Henneguya salminicola сохранились органеллы, связанные с митохондриями, при полной потере митохондриального генома. [5] [6] [7] Большое количество одноклеточных организмов , таких как микроспоридии , парабазалиды и дипломонады , сократили или трансформировали свои митохондрии в другие структуры, [8] например, гидрогеносомы и митосомы . [9] Оксимонады Monocercomonoides , . Streblomastix и Blattamonas полностью лишились митохондрий [5] [10]
Митохондрии обычно имеют размер от 0,75 до 3 мкм. 2 в поперечном сечении, [11] но значительно различаются по размеру и строению. Если специально не окрашивать , их не видно. Помимо снабжения клеток энергией, митохондрии участвуют в других задачах, таких как передача сигналов , клеточная дифференцировка и гибель клеток , а также поддержание контроля над клеточным циклом и ростом клеток . [12] Митохондриальный биогенез , в свою очередь, во времени координируется с этими клеточными процессами. [13] [14] Митохондрии вовлечены в ряд заболеваний и состояний человека, таких как митохондриальные заболевания , [15] нарушение сердечной деятельности , [16] сердечная недостаточность [17] и аутизм . [18]
Количество митохондрий в клетке может широко варьироваться в зависимости от организма , ткани и типа клеток. Зрелый эритроцит не имеет митохондрий. [19] тогда как клетка печени может иметь более 2000. [20] [21] Митохондрия состоит из отделов, выполняющих специализированные функции. Эти отсеки или области включают внешнюю мембрану, межмембранное пространство , внутреннюю мембрану , кристы и матрикс .
эукариотической клетки Хотя большая часть ДНК содержится в клеточном ядре , митохондрия имеет собственный геном («митогеном»), который существенно похож на бактериальные геномы. [22] Это открытие привело к всеобщему признанию эндосимбиотической гипотезы о том, что свободноживущие прокариотические предки современных митохондрий в далеком прошлом навсегда слились с эукариотическими клетками, эволюционируя таким образом, что современные животные, растения, грибы и другие эукариоты способны дышать , чтобы генерировать клеточная энергия . [23]
Структура
| Клеточная биология | |
|---|---|
| митохондрия | |
 Компоненты типичной митохондрии
3 Ламели
4 Митохондриальная ДНК |
Митохондрии могут иметь разную форму. [24] Митохондрия содержит внешнюю и внутреннюю мембраны, состоящие из бислоев фосфолипидов и белков . [20] Две мембраны имеют разные свойства. Из-за такой двухмембранной организации митохондрия состоит из пяти отдельных частей:
- Наружная митохондриальная мембрана,
- Межмембранное пространство (пространство между наружной и внутренней мембранами),
- Внутренняя мембрана митохондрий,
- Пространство крист (образованное складками внутренней мембраны) и
- Матрица ( пространство внутри внутренней мембраны), представляющая собой жидкость.
Митохондрии имеют складчатость, увеличивающую площадь поверхности, что, в свою очередь, увеличивает выработку АТФ (аденозинтрифосфата).Митохондрии, лишенные внешней мембраны, называются митопластами .
Внешняя мембрана
Внешняя митохондриальная мембрана , окружающая всю органеллу, имеет толщину от 60 до 75 ангстрем (Å). Соотношение белков и фосфолипидов в нем аналогично соотношению клеточной мембраны (около 1:1 по массе). Он содержит большое количество интегральных мембранных белков, называемых поринами . Основным транспортным белком является порообразующий потенциал-зависимый анионный канал (VDAC). VDAC нуклеотидов является основным переносчиком и , ионов и метаболитов между цитозолем межмембранным пространством. [25] [26] Он сформирован в виде бета-цилиндра , охватывающего внешнюю мембрану, аналогично внешней мембране грамотрицательных бактерий . [27] Более крупные белки могут проникать в митохондрии, если сигнальная последовательность на их N-конце связывается с большим многосубъединичным белком, называемым транслоказой, на внешней мембране , который затем активно перемещает их через мембрану. [28] Митохондриальные пробелки импортируются через специализированные транслокационные комплексы.
Наружная мембрана также содержит , в таких разнообразных процессах, как элонгация жирных кислот , окисление адреналина ферменты и деградация триптофана участвующие . Эти ферменты включают моноаминоксидазу , нечувствительную к ротенону НАДН-цитохром-с-редуктазу, кинуренингидроксилазу лигазу и Ко-А- жирных кислот . Разрушение внешней мембраны позволяет белкам из межмембранного пространства проникать в цитозоль, что приводит к гибели клеток. [29] Внешняя митохондриальная мембрана может соединяться с мембраной эндоплазматического ретикулума (ЭР) в структуре, называемой МАМ (митохондриально-ассоциированная ЭР-мембрана). Это важно для передачи сигналов кальция в ЭР-митохондрии и участвует в переносе липидов между ЭР и митохондриями. [30] За пределами внешней мембраны находятся небольшие (диаметром 60 Å) частицы, называемые субъединицами Парсона.
Межмембранное пространство
— Межмембранное пространство митохондрий это пространство между внешней и внутренней мембранами. Его также называют перимитохондриальным пространством. Поскольку внешняя мембрана свободно проницаема для малых молекул, концентрации малых молекул, таких как ионы и сахара, в межмембранном пространстве такие же, как и в цитозоле . [20] Однако крупные белки должны иметь специфическую сигнальную последовательность для транспортировки через внешнюю мембрану, поэтому белковый состав этого пространства отличается от белкового состава цитозоля . Одним из белков , локализующихся таким образом в межмембранном пространстве, является цитохром с . [29]
Внутренняя мембрана
Внутренняя мембрана митохондрий содержит белки с тремя типами функций: [20]
- Те, которые осуществляют цепи переноса электронов . окислительно-восстановительные реакции
- АТФ-синтаза , которая генерирует АТФ в матрице
- Специфические транспортные белки , регулирующие прохождение метаболитов в митохондриальный матрикс и из него.
Он содержит более 151 различных полипептидов и имеет очень высокое соотношение белка к фосфолипидам (более 3:1 по массе, что составляет примерно 1 белок на 15 фосфолипидов). Внутренняя мембрана содержит около 1/5 всего белка митохондрии. [31] Кроме того, внутренняя мембрана богата необычным фосфолипидом кардиолипином . Этот фосфолипид был первоначально обнаружен в коровьем сердце в 1942 году и обычно характерен для митохондриальных и бактериальных плазматических мембран. [32] Кардиолипин содержит четыре жирные кислоты, а не две, и может помочь сделать внутреннюю мембрану непроницаемой. [20] и его нарушение может привести к множеству клинических расстройств, включая неврологические расстройства и рак. [33] В отличие от внешней мембраны внутренняя мембрана не содержит поринов и обладает высокой непроницаемостью для всех молекул. Почти всем ионам и молекулам требуются специальные мембранные транспортеры для входа или выхода из матрикса. Белки переправляются в матрикс через транслоказу комплекса внутренней мембраны (TIM) или через OXA1L . [28] Кроме того, на внутренней мембране существует мембранный потенциал, образующийся под действием ферментов цепи электронов переноса . внутренней мембраны Слияние опосредуется белком внутренней мембраны OPA1 . [34]
Христос

Внутренняя митохондриальная мембрана разделена на многочисленные складки, называемые кристами , которые увеличивают площадь поверхности внутренней митохондриальной мембраны, повышая ее способность вырабатывать АТФ. Для типичных митохондрий печени площадь внутренней мембраны примерно в пять раз больше площади наружной мембраны. Это соотношение является переменным, и митохондрии клеток, которым требуется больше АТФ, например мышечных клеток, содержат еще больше крист. Митохондрии в одной и той же клетке могут иметь существенно разную плотность крист, причем те, которые необходимы для производства большего количества энергии, имеют гораздо большую поверхность крист-мембраны. [35] Эти складки усеяны маленькими круглыми телами, известными как F 1 частицы или оксисомы. [36]
Матрица
Матрица – это пространство, окруженное внутренней мембраной. Он содержит около 2/3 всех белков митохондрии. [20] Матрица важна для производства АТФ с помощью АТФ-синтазы, содержащейся во внутренней мембране. Матрица содержит высококонцентрированную смесь сотен ферментов, особых митохондриальных рибосом , тРНК и нескольких копий генома ДНК митохондриальной . Из ферментов основные функции включают окисление пирувата и жирных кислот , а также цикл лимонной кислоты . [20] Молекулы ДНК упаковываются в нуклеоиды белками, одним из которых является TFAM . [37]
Функция
Наиболее важные роли митохондрий заключаются в производстве энергетической валюты клетки, АТФ (т.е. фосфорилировании АДФ ), посредством дыхания и регуляции клеточного метаболизма . [21] Центральный набор реакций, участвующих в производстве АТФ, известен под общим названием цикл лимонной кислоты или цикл Кребса и окислительное фосфорилирование . Однако помимо производства АТФ у митохондрии есть много других функций.
Преобразование энергии
Доминирующей ролью митохондрий является производство АТФ, о чем свидетельствует большое количество белков во внутренней мембране, выполняющих эту задачу. Это осуществляется путем окисления основных продуктов глюкозы : пирувата и НАДН , которые образуются в цитозоле. [21] Этот тип клеточного дыхания , известный как аэробное дыхание , зависит от присутствия кислорода . Когда кислород ограничен, гликолитические продукты будут метаболизироваться посредством анаэробной ферментации , процесса, который не зависит от митохондрий. [21] Производство АТФ из глюкозы и кислорода при аэробном дыхании примерно в 13 раз выше, чем при ферментации. [38] Митохондрии растений также могут производить ограниченное количество АТФ либо за счет расщепления сахара, образующегося во время фотосинтеза, либо без кислорода, используя альтернативный субстрат — нитрит . [39] АТФ проникает через внутреннюю мембрану с помощью специфического белка и через внешнюю мембрану через порины . [40] После преобразования АТФ в АДФ путем дефосфорилирования , которое высвобождает энергию, АДФ возвращается тем же путем.
Пируват и цикл лимонной кислоты
пирувата, Молекулы образующиеся в результате гликолиза через , активно транспортируются внутреннюю митохондриальную мембрану в матрикс, где они могут либо окисляться , либо соединяться с коферментом А с образованием CO 2 , ацетил-КоА и НАДН . [21] или они могут быть карбоксилированы ( пируваткарбоксилазой ) с образованием оксалоацетата. Эта последняя реакция «восполняет» количество оксалоацетата в цикле лимонной кислоты и, следовательно, является анаплеротической реакцией , увеличивая способность цикла метаболизировать ацетил-КоА, когда энергетические потребности тканей (например, в мышцах ) внезапно возрастают из-за активности. [41]
В цикле лимонной кислоты все промежуточные продукты (например цитрат , , изоцитрат , альфа-кетоглутарат , сукцинат, фумарат , малат и оксалоацетат) регенерируются во время каждого цикла цикла. Таким образом, добавление большего количества любого из этих промежуточных продуктов в митохондрии означает, что дополнительное количество сохраняется в цикле, увеличивая количество всех других промежуточных продуктов по мере превращения одного в другое. Следовательно, добавление любого из них в цикл имеет анаплеротический эффект, а его удаление – катаплеротический эффект. Эти анаплеротические и катаплеротические реакции в течение цикла будут увеличивать или уменьшать количество оксалоацетата, доступного для соединения с ацетил-КоА с образованием лимонной кислоты. Это, в свою очередь, увеличивает или уменьшает скорость производства АТФ митохондриями и, следовательно, доступность АТФ для клетки. [41]
С другой стороны, ацетил-КоА, получаемый в результате окисления пирувата или бета-окисления , жирных кислот является единственным топливом, вступающим в цикл лимонной кислоты. С каждым оборотом цикла на каждую молекулу оксалоацетата, присутствующую в митохондриальном матриксе, расходуется одна молекула ацетил-КоА и никогда не регенерируется. Это окисление ацетатной части ацетил-КоА, в результате которого образуются CO 2 и вода, при этом высвободившаяся энергия улавливается в форме АТФ. [41]
В печени карбоксилирование цитозольного в глюкозу пирувата во внутримитохондриальный оксалоацетат является ранним этапом глюконеогенного пути, который превращает лактат и дезаминированный аланин . [21] [41] под влиянием высокого уровня глюкагона и/или адреналина в крови. [41] Здесь добавление оксалоацетата к митохондриям не имеет общего анаплеротического эффекта, поскольку другой промежуточный продукт цикла лимонной кислоты (малат) немедленно удаляется из митохондрии и превращается в цитозольный оксалоацетат и, в конечном итоге, в глюкозу в процессе, который почти обратный гликолизу . [41]
Ферменты цикла лимонной кислоты расположены в митохондриальном матриксе, за исключением сукцинатдегидрогеназы , которая связана с внутренней мембраной митохондрий в составе Комплекса II. [42] Цикл лимонной кислоты окисляет ацетил-КоА до диоксида углерода и в процессе производит восстановленные кофакторы (три молекулы НАДН и одна молекула ФАДН 2 ), которые являются источником электронов для цепи переноса электронов , а также молекулу ГТФ (который легко преобразуется в АТФ). [21]
O 2 и НАДН: реакции высвобождения энергии
Электроны от НАДН и ФАДН 2 передаются кислороду (О 2 ) и водороду (протонам) в несколько этапов через цепь переноса электронов. Молекулы НАДН и ФАДН 2 производятся внутри матрикса посредством цикла трикарбоновых кислот и в цитоплазме посредством гликолиза . Восстанавливающие эквиваленты из цитоплазмы могут быть импортированы через малат-аспартатную челночную систему белков- антипортеров или поданы в цепь переноса электронов с использованием глицеринфосфатного челнока . [21]
Основные реакции выделения энергии [43] [44] которые делают митохондрии «электростанцией клетки», происходят в белковых комплексах I, III и IV во внутренней мембране митохондрий ( НАДН-дегидрогеназа (убихинон) , цитохром-с-редуктаза и цитохром-с-оксидаза ). В комплексе IV О 2 реагирует с восстановленной формой железа в цитохроме с :


высвободить много свободной энергии [44] [43] из реагентов, не разрывая связей органического топлива. Свободная энергия, затрачиваемая на отрыв электрона от Fe 2+ высвобождается в комплексе III, когда Fe 3+ цитохрома с реагирует с окислением убихинола (QH 2 ):


Образовавшийся убихинон : реагирует (Q) в комплексе I с НАДН


Хотя реакции контролируются цепью переноса электронов, свободные электроны не входят в число реагентов или продуктов в трех показанных реакциях и, следовательно, не влияют на высвобождаемую свободную энергию, которая используется для перекачки протонов (H + ) в межмембранное пространство. Этот процесс эффективен, но небольшой процент электронов может преждевременно восстанавливать кислород, образуя активные формы кислорода, такие как супероксид . [21] Это может вызвать окислительный стресс в митохондриях и способствовать снижению функции митохондрий, связанному со старением. [45]
По мере увеличения концентрации протонов в межмембранном пространстве сильный электрохимический градиент через внутреннюю мембрану устанавливается . Протоны могут возвращаться в матрикс через АТФ-синтазный комплекс, а их потенциальная энергия используется для синтеза АТФ из АДФ и неорганического фосфата (P i ). [21] Этот процесс называется хемиосмосом и впервые был описан Питером Митчеллом . [46] [47] который был удостоен Нобелевской премии по химии 1978 года за свою работу. Позже часть Нобелевской премии по химии 1997 года была присуждена Полу Д. Бойеру и Джону Э. Уокеру за разъяснение рабочего механизма АТФ-синтазы. [48]
Производство тепла
При определенных условиях протоны могут повторно войти в митохондриальный матрикс, не участвуя в синтезе АТФ. Этот процесс известен как утечка протонов или разобщение митохондрий и обусловлен облегченной диффузией протонов в матрикс. градиента протонов В результате этого процесса неиспользованная потенциальная энергия электрохимического выделяется в виде тепла. [21] Этот процесс опосредован протонным каналом, называемым термогенином , или UCP1 . [49] Термогенин в основном содержится в бурой жировой ткани или буром жире и отвечает за несократительный термогенез. Бурая жировая ткань встречается у млекопитающих, ее самый высокий уровень наблюдается в раннем возрасте и у животных, находящихся в спячке. У человека бурая жировая ткань присутствует при рождении и уменьшается с возрастом. [49]
Синтез митохондриальных жирных кислот
Синтез митохондриальных жирных кислот (mtFASII) необходим для клеточного дыхания и митохондриального биогенеза. [50] Также считается, что он играет роль медиатора во внутриклеточной передаче сигналов из-за его влияния на уровни биоактивных липидов, таких как лизофосфолипиды и сфинголипиды . [51]
Октаноил-АСР (С8) считается наиболее важным конечным продуктом mtFASII, который также образует стартовый субстрат биосинтеза липоевой кислоты . [52] Поскольку липоевая кислота является кофактором важных митохондриальных ферментных комплексов, таких как комплекс пируватдегидрогеназы (PDC), комплекс α-кетоглутаратдегидрогеназы (OGDC), комплекс дегидрогеназы α-кетокислот с разветвленной цепью (BCKDC), а также в системе расщепления глицина ( GCS), mtFASII оказывает влияние на энергетический обмен. [53]
Другие продукты mtFASII играют роль в регуляции митохондриальной трансляции, биогенезе кластеров FeS и сборке комплексов окислительного фосфорилирования. [52]
Кроме того, с помощью mtFASII и ацилированного ACP ацетил-КоА регулирует свое потребление в митохондриях. [52]
Поглощение, хранение и высвобождение ионов кальция.

Концентрация свободного кальция в клетке может регулировать ряд реакций и важна для передачи сигнала в клетке. Митохондрии могут временно хранить кальций , что способствует гомеостазу кальция в клетках. [54] [55] Их способность быстро поглощать кальций для последующего высвобождения делает их хорошими «цитозольными буферами» для кальция. [56] [57] [58] Эндоплазматическая сеть (ЭР) является наиболее важным местом хранения кальция. [59] и существует значительное взаимодействие между митохондриями и ЭР в отношении кальция. [60] Кальций поглощается в матрикс митохондриальным унипортером кальция на внутренней митохондриальной мембране . [61] В первую очередь это обусловлено мембранным потенциалом митохондрий . [55] Высвобождение этого кальция обратно во внутреннюю часть клетки может происходить через натрий-кальциевый обменный белок или по путям «индуцированного кальцием высвобождения кальция». [61] Это может инициировать выбросы кальция или волны кальция с большими изменениями мембранного потенциала. Они могут активировать ряд белков системы вторичных мессенджеров , которые могут координировать такие процессы, как высвобождение нейротрансмиттеров в нервных клетках и высвобождение гормонов в эндокринных клетках. [62]
Что 2+ приток к митохондриальному матриксу недавно был признан механизмом регулирования дыхательной биоэнергетики , позволяя электрохимическому потенциалу через мембрану временно «пульсировать» от ΔΨ-доминирования к доминированию pH, способствуя снижению окислительного стресса . [63] В нейронах сопутствующее увеличение цитозольного и митохондриального кальция синхронизирует активность нейронов с митохондриальным энергетическим метаболизмом. Уровни кальция в митохондриальном матриксе могут достигать десятков микромолярных уровней, что необходимо для активации изоцитратдегидрогеназы , одного из ключевых регуляторных ферментов цикла Кребса . [64]
Регуляция клеточной пролиферации
Была исследована связь между клеточной пролиферацией и митохондриями. Опухолевым клеткам требуется достаточное количество АТФ для синтеза биологически активных соединений, таких как липиды , белки и нуклеотиды , для быстрой пролиферации. [65] Большая часть АТФ в опухолевых клетках генерируется посредством пути окислительного фосфорилирования (OxPhos). [66] Взаимодействие с OxPhos вызывает остановку клеточного цикла , что позволяет предположить, что митохондрии играют роль в пролиферации клеток. [66] Производство митохондриального АТФ также жизненно важно для клеток при инфекции. деления и дифференцировки [67] в дополнение к основным функциям в клетке, включая регуляцию объема клетки, концентрации растворенных веществ и клеточной архитектуры. [68] [69] [70] Уровни АТФ различаются на разных стадиях клеточного цикла, что позволяет предположить, что существует взаимосвязь между количеством АТФ и способностью клетки вступать в новый клеточный цикл. [71] Роль АТФ в основных функциях клетки делает клеточный цикл чувствительным к изменениям доступности АТФ, полученного из митохондрий. [71] Вариации уровней АТФ на разных стадиях клеточного цикла подтверждают гипотезу о том, что митохондрии играют важную роль в регуляции клеточного цикла. [71] Хотя конкретные механизмы между митохондриями и регуляцией клеточного цикла не совсем понятны, исследования показали, что контрольные точки низкоэнергетического клеточного цикла контролируют энергетические возможности перед переходом к следующему раунду клеточного деления. [12]
Запрограммированная гибель клеток и врожденный иммунитет
Запрограммированная гибель клеток (ПКС) имеет решающее значение для различных физиологических функций, включая развитие органов и клеточный гомеостаз. Он служит внутренним механизмом предотвращения злокачественной трансформации и играет фундаментальную роль в иммунитете, помогая в противовирусной защите, устранении патогенов, воспалении и рекрутировании иммунных клеток. [72]
Митохондрии давно признаны за их центральную роль во пути апоптоза внутреннем , формы ПКС. [73] В последние десятилетия они также были идентифицированы как сигнальные центры для большей части врожденной иммунной системы . [74] Эндосимбиотическое происхождение митохондрий отличает их от других клеточных компонентов, а воздействие митохондриальных элементов на цитозоль может запускать те же пути, что и маркеры инфекции. Эти пути приводят к апоптозу , аутофагии или индукции провоспалительных генов. [75] [74]
Митохондрии способствуют апоптозу, высвобождая цитохром с , который непосредственно индуцирует образование апоптосом . Кроме того, они являются источником различных молекулярных структур, связанных с повреждениями (DAMP). Эти DAMP часто распознаются теми же рецепторами распознавания образов (PRR), которые реагируют на молекулярные паттерны, связанные с патогенами (PAMP) во время инфекций. [76] Например, митохондриальная мтДНК напоминает бактериальную ДНК из-за отсутствия метилирования CpG и может быть обнаружена с помощью Toll-подобного рецептора 9 и cGAS . [77] Двухцепочечная РНК (дсРНК), образующаяся в результате двунаправленной митохондриальной транскрипции, может активировать пути вирусной чувствительности через RIG-I-подобные рецепторы . [78] Кроме того, N -формилирование митохондриальных белков , аналогичное таковому у бактериальных белков, может распознаваться рецепторами формилпептидов . [79] [80]
В норме эти митохондриальные компоненты изолированы от остальной части клетки, но высвобождаются после проницаемости митохондриальной мембраны во время апоптоза или пассивно после повреждения митохондрий. Однако митохондрии также играют активную роль во врожденном иммунитете, высвобождая мтДНК в ответ на метаболические сигналы. [74] Митохондрии также являются местом локализации иммунных белков и белков, регулирующих апоптоз, таких как BAX , MAVS (расположен на внешней мембране ) и NLRX1 (находится в матриксе ). Эти белки модулируются метаболическим статусом митохондрий и динамикой митохондрий. [74] [81] [82]
Дополнительные функции
Митохондрии играют центральную роль во многих других метаболических задачах, таких как:
- Передача сигналов через митохондриальные активные формы кислорода [83]
- Регуляция мембранного потенциала [21]
- Передача сигналов кальция (включая апоптоз, вызванный кальцием) [84]
- Регуляция клеточного метаболизма [12]
- Определенные гема реакции синтеза [85] (см. также: Порфирин )
- стероидов Синтез [56]
- Гормональная сигнализация [86] – митохондрии чувствительны и реагируют на гормоны, отчасти за счет действия митохондриальных рецепторов эстрогена (mtER). Эти рецепторы были обнаружены в различных тканях и типах клеток, включая мозг. [87] и сердце [88]
- Развитие и функции иммунных клеток [89]
- Нейрональные митохондрии также способствуют контролю качества клеток, сообщая о статусе нейронов микроглии через специализированные соматические соединения. [90]
- Митохондрии развивающихся нейронов способствуют межклеточной передаче сигналов микроглии , связь которой необходима для правильной регуляции развития мозга. [91]
Некоторые митохондриальные функции выполняются только в определенных типах клеток. Например, митохондрии в клетках печени содержат ферменты, которые позволяют им детоксикации аммиака , отхода белкового обмена. Мутация генов, регулирующих любую из этих функций, может привести к митохондриальным заболеваниям .
Митохондриальные белки (белки, транскрибируемые из митохондриальной ДНК) различаются в зависимости от ткани и вида. идентифицировано 615 различных типов белков У человека в митохондриях сердца . [92] тогда как у крыс сообщалось о 940 белках. [93] митохондриальный протеом динамически регулируется. Считается, что [94]
Организация и распространение

Митохондрии (или родственные структуры) встречаются у всех эукариот (кроме оксимонад Monocercomonoides ). [5] Хотя их обычно изображают в виде бобовидных структур, они образуют высокодинамическую сеть в большинстве клеток, где постоянно подвергаются делению и слиянию . Популяция всех митохондрий данной клетки составляет хондриому. [95] Митохондрии различаются по количеству и расположению в зависимости от типа клеток. У одноклеточных организмов часто встречается одна митохондрия, тогда как клетки печени человека содержат около 1000–2000 митохондрий на клетку, что составляет 1/5 объема клетки. [20] Митохондриальное содержимое сходных в остальном клеток может существенно различаться по размеру и мембранному потенциалу. [96] с различиями, возникающими из-за источников, включая неравномерное разделение при делении клеток, что приводит к внешним различиям в уровнях АТФ и последующих клеточных процессах. [97] Митохондрии могут располагаться между миофибриллами мышц спермия или обернуты жгутика вокруг . [20] Часто они образуют сложную трехмерную ветвящуюся сеть внутри клетки с цитоскелетом . Ассоциация с цитоскелетом определяет форму митохондрий, что также может влиять на функцию: [98] Различные структуры митохондриальной сети могут предоставлять популяции различные физические, химические и сигнальные преимущества или недостатки. [99] Митохондрии в клетках всегда распределены вдоль микротрубочек и распределение этих органелл также коррелирует с эндоплазматической сетью . [100] Недавние данные свидетельствуют о том, что виментин , один из компонентов цитоскелета, также имеет решающее значение для связи с цитоскелетом. [101]
Митохондриально-ассоциированная мембрана ЭР (МАМ)
Мембрана ЭР, связанная с митохондриями (МАМ), является еще одним структурным элементом, который все чаще признается за его решающую роль в клеточной физиологии и гомеостазе . Когда-то считавшиеся техническим препятствием в методах фракционирования клеток, предполагаемые примеси везикул ЭР, которые неизменно появлялись в митохондриальной фракции, были повторно идентифицированы как мембранные структуры, полученные из МАМ — интерфейса между митохондриями и ЭР. [102] Физическое соединение между этими двумя органеллами ранее наблюдалось на электронных микрофотографиях, а позже было исследовано с помощью флуоресцентной микроскопии . [102] По оценкам таких исследований, в MAM, который может составлять до 20% внешней мембраны митохондрий, ER и митохондрии разделены всего на 10–25 нм и удерживаются вместе белковыми привязывающими комплексами. [102] [30] [103]
Очищенный МАМ, полученный в результате субклеточного фракционирования, обогащен ферментами, участвующими в обмене фосфолипидов, в дополнение к каналам, связанным с Ca. 2+ сигнализация. [102] [103] Эти намеки на выдающуюся роль MAM в регуляции клеточных липидных запасов и передаче сигнала были подтверждены, что имело значительные последствия для клеточных явлений, связанных с митохондриями, как обсуждается ниже. МАМ не только предоставил понимание механистической основы, лежащей в основе таких физиологических процессов, как внутренний апоптоз и распространение передачи сигналов кальция, но также способствует более точному взгляду на митохондрии. Хотя МАМ часто рассматривается как статичные, изолированные «электростанции», задействованные для клеточного метаболизма посредством древнего эндосимбиотического события, эволюция МАМ подчеркивает степень, в которой митохондрии интегрированы в общую клеточную физиологию, с тесной физической и функциональной связью с эндомембранной системой.
Перенос фосфолипидов
МАМ обогащен ферментами, участвующими в биосинтезе липидов, такими как фосфатидилсеринсинтаза на поверхности ЭР и фосфатидилсериндекарбоксилаза на поверхности митохондрий. [104] [105] Поскольку митохондрии представляют собой динамические органеллы, постоянно подвергающиеся процессам деления и слияния , им требуется постоянное и хорошо регулируемое снабжение фосфолипидами для целостности мембран. [106] [107] Но митохондрии являются не только местом назначения фосфолипидов, синтез которых они завершают; скорее, эта органелла также играет роль в межорганелловом транспорте промежуточных продуктов и продуктов путей биосинтеза фосфолипидов, метаболизме церамидов и холестерина, а также анаболизме гликосфинголипидов. [105] [107]
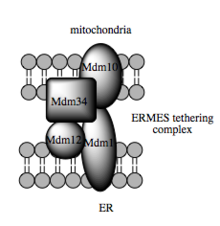
Такая способность транспорта зависит от MAM, который, как было показано, облегчает перенос липидных промежуточных продуктов между органеллами. [104] В отличие от стандартного везикулярного механизма переноса липидов, данные указывают на то, что физическая близость ER и митохондриальных мембран в MAM позволяет осуществлять переворот липидов между противоположными бислоями. [107] Несмотря на этот необычный и, казалось бы, энергетически невыгодный механизм, для такого транспорта не требуется АТФ. [107] Вместо этого у дрожжей было показано, что это зависит от мультибелковой связывающей структуры, называемой структурой встречи ER-митохондрий или ERMES, хотя остается неясным, опосредует ли эта структура непосредственно перенос липидов или она необходима для поддержания мембран в достаточно тесной близости. снизить энергетический барьер для переворота липидов . [107] [108]
МАМ также может быть частью секреторного пути в дополнение к своей роли во внутриклеточном транспорте липидов. В частности, MAM, по-видимому, является промежуточным пунктом назначения между шероховатым ЭР и Гольджи на пути, который приводит к сборке и секреции липопротеинов очень низкой плотности , или ЛПОНП. [105] [109] Таким образом, МАМ служит важнейшим центром обмена веществ и транспорта в липидном обмене.
Передача сигналов кальция
Критическая роль ЭР в передаче сигналов кальция была признана до того, как такая роль митохондрий получила широкое признание, отчасти из-за низкого сродства Ca 2+ каналы, локализованные на внешней митохондриальной мембране, по-видимому, противоречат предполагаемой реакции этой органеллы на изменения внутриклеточного кальция. 2+ поток. [102] [59] Но присутствие МАМ разрешает это очевидное противоречие: тесная физическая связь между двумя органеллами приводит к образованию Ca 2+ микродомены в точках контакта, которые способствуют эффективному Ca 2+ передача из ЭР в митохондрии. [102] Передача происходит в ответ на так называемый «Ca 2+ затяжки», генерируемые спонтанной кластеризацией и активацией IP3R , канонической мембраны ЭР Ca. 2+ канал. [102] [30]
Судьба этих клубков, в частности, остаются ли они ограниченными изолированными местами или интегрированы в Ca. 2+ волны для распространения по клетке — во многом определяется динамикой МАМ. Хотя обратный захват Ca 2+ ЭР (одновременно с его высвобождением) модулирует интенсивность затяжек, таким образом изолируя митохондрии в определенной степени от высокого содержания кальция. 2+ воздействия, MAM часто служит брандмауэром, который по существу буферизует Ca 2+ слойки, действуя как сток, в который могут направляться свободные ионы, высвобождаемые в цитозоль. [102] [110] [111] Это Калифорния 2+ туннелирование происходит через Ca с низким сродством 2+ рецептор VDAC1 , который, как недавно было показано, физически связан с кластерами IP3R на мембране ЭР и обогащен в МАМ. [102] [30] [112] Способность митохондрий служить Ca 2+ сток является результатом электрохимического градиента, возникающего при окислительном фосфорилировании, что делает туннелирование катиона экзергоническим процессом. [112] Нормальный, умеренный приток кальция из цитозоля в митохондриальный матрикс вызывает временную деполяризацию, которая корректируется путем откачки протонов.
Но передача Ca 2+ не является однонаправленным; скорее, это улица с двусторонним движением. [59] Свойства Ca 2+ насос SERCA и канал IP3R, присутствующие на мембране ER, облегчают регуляцию по обратной связи, координируемую функцией MAM. В частности, клиренс Ca 2+ с помощью MAM позволяет формировать пространственно-временной паттерн Ca 2+ сигнализация, потому что Ca 2+ изменяет активность IP3R двухфазным образом. [102] На SERCA также влияет митохондриальная обратная связь: поглощение Ca 2+ МАМ стимулирует выработку АТФ, обеспечивая тем самым энергию, которая позволяет SERCA перезагрузить ЭР Ca. 2+ для продолжения Ca 2+ отток в МАМ. [110] [112] Таким образом, МАМ не является пассивным буфером для Ca. 2+ слойки; скорее, это помогает модулировать дальнейшую Ca 2+ передача сигналов через петли обратной связи, которые влияют на динамику ER.
Регуляция высвобождения Ca в ЭР 2+ на MAM особенно критично, поскольку только определенное окно Ca 2+ поглощение поддерживает митохондрии и, следовательно, клетку в гомеостазе. Достаточное количество внутриорганелл Ca 2+ передача сигналов необходима для стимуляции метаболизма путем активации ферментов дегидрогеназы, имеющих решающее значение для прохождения цикла лимонной кислоты. [113] [114] Однако однажды Ка 2+ передача сигналов в митохондриях достигает определенного порога, она стимулирует внутренний путь апоптоза, частично за счет коллапса потенциала митохондриальной мембраны, необходимого для метаболизма. [102] Исследования, изучающие роль про- и антиапоптотических факторов, подтверждают эту модель; например, было показано, что антиапоптотический фактор Bcl-2 взаимодействует с IP3R, уменьшая Ca 2+ заполнение ЭР, что приводит к уменьшению оттока в МАМ и предотвращению коллапса потенциальных постапоптотических стимулов митохондриальной мембраны. [102] Учитывая необходимость столь тонкой регуляции Ca 2+ передачи сигналов, возможно, неудивительно, что нарушение регуляции митохондриального Ca 2+ был вовлечен в ряд нейродегенеративных заболеваний, в то время как каталог супрессоров опухолей включает несколько препаратов, обогащенных MAM. [112]
Молекулярная основа привязки
Недавние достижения в идентификации связей между митохондриальными и мембранами ER позволяют предположить, что функция каркаса вовлеченных молекулярных элементов является вторичной по отношению к другим, неструктурным функциям. У дрожжей ERMES, мультипротеиновый комплекс взаимодействующих мембранных белков, находящихся в ER и митохондриях, необходим для переноса липидов в MAM и служит примером этого принципа. Например, один из его компонентов также входит в состав белкового комплекса, необходимого для внедрения трансмембранных белков бета-бочонка в липидный бислой. [107] Однако гомолог комплекса ERMES пока не идентифицирован в клетках млекопитающих. Другие белки, вовлеченные в каркас, также имеют функции, независимые от структурной привязки к MAM; например, ER-резидентные и митохондриальные митофузины образуют гетерокомплексы, которые регулируют количество мест контакта между органеллами, хотя митофузины были впервые идентифицированы по их роли в событиях деления и слияния между отдельными митохондриями. [102] Белок 75, связанный с глюкозой (grp75), является еще одним белком с двойной функцией. В дополнение к матриксному пулу grp75, часть служит шапероном, который физически связывает митохондрии и ЭР Ca. 2+ каналы VDAC и IP3R для эффективного Ca 2+ передача в МАМ. [102] [30] Другой потенциальной привязью является Sigma-1R , неопиоидный рецептор, чья стабилизация ER-резидентного IP3R может сохранять связь в MAM во время реакции метаболического стресса. [115] [116]
Перспектива
МАМ является важным узлом передачи сигналов, метаболизма и транспорта в клетке, который обеспечивает интеграцию ЭР и физиологии митохондрий. Связь между этими органеллами является не просто структурной, но и функциональной и имеет решающее значение для общей клеточной физиологии и гомеостаза . Таким образом, MAM предлагает взгляд на митохондрии, который расходится с традиционным представлением об этой органелле как о статической, изолированной единице, предназначенной для метаболических способностей клетки. [117] Вместо этого этот интерфейс митохондрий и ЭР подчеркивает интеграцию митохондрий, продукта эндосимбиотического события, в разнообразные клеточные процессы. Недавно также было показано, что митохондрии и МАМ в нейронах прикреплены к специализированным местам межклеточной коммуникации (так называемым соматическим соединениям). Процессы микроглии контролируют и защищают функции нейронов в этих участках, и предполагается, что MAM-s играют важную роль в этом типе контроля качества клеток. [90]
Происхождение и эволюция
Существуют две гипотезы происхождения митохондрий: эндосимбиотическая и аутогенная . Эндосимбиотическая гипотеза предполагает, что митохондрии изначально были прокариотическими клетками, способными реализовывать окислительные механизмы, которые были невозможны для эукариотических клеток; они стали эндосимбионтами, живущими внутри эукариот. [23] [118] [119] [120] Согласно аутогенной гипотезе, митохондрии возникли в результате отщепления части ДНК от ядра эукариотической клетки в момент ее расхождения с прокариотами; эта часть ДНК была бы окружена мембранами, через которые белки не могли бы пройти. Поскольку митохондрии имеют много общих черт с бактериями , эндосимбиотическая гипотеза является более широко принятой из двух версий. [120] [121]
Митохондрия содержит ДНК , которая организована в виде нескольких копий одной, обычно кольцевой хромосомы . Эта митохондриальная хромосома содержит гены окислительно-восстановительных белков, например белков дыхательной цепи. Гипотеза CoRR предполагает, что такое совместное расположение необходимо для окислительно-восстановительной регуляции. Митохондриальный геном кодирует некоторые РНК рибосом необходимые и 22 тРНК, для трансляции мРНК в белок. Круглая структура встречается и у прокариот. Протомитохондрия , вероятно, была тесно связана с Rickettsia . [122] [123] Однако точное родство предка митохондрий с альфапротеобактериями и образовалась ли митохондрия одновременно с ядром или после ядра, остается спорным. [124] Например, было высказано предположение, что клада бактерий SAR11 имеет относительно недавнего общего предка с митохондриями, [125] в то время как филогеномный анализ показывает, что митохондрии произошли от линии Pseudomonadota или является ее членом , которая тесно связана с альфапротеобактериями . [126] [127] В некоторых статьях митохондрии описываются как сестры альфапротеобактерий, вместе образующие сестринскую группу морских протео1, вместе образующие сестру Magnetococcidae . [128] [129] [130] [131]
Рибосомы, кодируемые митохондриальной ДНК, по размеру и структуре аналогичны рибосомам бактерий. [132] Они очень напоминают бактериальную рибосому 70S , а не рибосомы 80S цитоплазматические , которые кодируются ядерной ДНК.
Эндосимбиотические Линн отношения митохондрий с клетками-хозяевами были популяризированы Маргулис . [133] Эндосимбиотическая гипотеза предполагает, что митохондрии произошли от аэробных бактерий, которые каким-то образом пережили эндоцитоз другой клетки и включились в цитоплазму . Способность этих бактерий осуществлять дыхание в клетках-хозяевах, основанных на гликолизе и ферментации, могла бы обеспечить значительное эволюционное преимущество. Эти симбиотические отношения, вероятно, возникли 1,7–2 миллиарда лет назад. [134] [135]

Некоторые группы одноклеточных эукариот имеют только рудиментарные митохондрии или производные структуры: микроспоридии , метамонады и архамебы . [136] Эти группы выглядят как самые примитивные эукариоты на филогенетических деревьях, построенных с использованием информации рРНК , что когда-то предполагало, что они появились до возникновения митохондрий. Однако сейчас известно, что это результат притяжения длинных ветвей : они представляют собой производные группы и сохраняют гены или органеллы, происходящие из митохондрий (например, митосомы и гидрогеносомы ). [8] Гидрогеносомы, митосомы и родственные органеллы, обнаруженные у некоторых лорицифер (например, Spinoloricus ). [137] [138] и миксозоа (например, Henneguya zschokkei ) вместе классифицируются как MRO, органеллы, связанные с митохондриями. [7] [139]
Monocercomonoides и другие оксимонады, по-видимому, полностью утратили свои митохондрии, и, по крайней мере, некоторые из митохондриальных функций теперь, по-видимому, выполняются цитоплазматическими белками. [5] [140] [10]
Митохондриальная генетика

Митохондрии содержат собственный геном. Митохондриальный геном человека представляет собой кольцевую двухцепочечную молекулу ДНК длиной около 16 тыс. нуклеотидов . [141] Он кодирует 37 генов: 13 для субъединиц дыхательных комплексов I, III, IV и V, 22 для митохондриальной тРНК (для 20 стандартных аминокислот плюс дополнительный ген для лейцина и серина) и 2 для рРНК (12S и 16S рРНК). ). [141] Одна митохондрия может содержать от двух до десяти копий своей ДНК. [142] Одна из двух цепей митохондриальной ДНК (мтДНК) имеет непропорционально более высокое соотношение более тяжелых нуклеотидов аденина и гуанина, и это называется тяжелой цепью (или H-цепью), тогда как другая цепь называется легкой цепью (или L-цепью). . Разница в весе позволяет разделить две нити центрифугированием . мтДНК имеет один длинный некодирующий участок, известный как некодирующая область (NCR), который содержит промотор тяжелой цепи (HSP) и промотор легкой цепи (LSP) для транскрипции РНК, начала репликации H-цепи (OriH). локализованный на L-цепи, трех консервативных последовательностях (CSB 1–3) и последовательности, связанной с терминацией (TAS). Начало репликации L-цепи (OriL) локализовано на H-цепи на 11 000 п.н. ниже OriH, расположенной внутри кластера генов, кодирующих тРНК. [143]
Как и у прокариот, здесь очень высока доля кодирующей ДНК и отсутствуют повторы. Митохондриальные гены транскрибируются в виде мультигенных транскриптов, которые расщепляются и полиаденилируются с образованием зрелых мРНК . Большинство белков, необходимых для функционирования митохондрий, кодируются генами в ядре клетки , и соответствующие белки импортируются в митохондрии. [144] Точное количество генов, кодируемых ядром и митохондриальным геномом, различается у разных видов. Большинство митохондриальных геномов имеют кольцевую форму. [145] Как правило, в митохондриальной ДНК отсутствуют интроны , как и в митохондриальном геноме человека; [144] однако интроны наблюдались в митохондриальной ДНК некоторых эукариот, [146] например, дрожжевой [147] и протисты , [148] в том числе Dictyostelium discoideum . [149] Между областями, кодирующими белок, присутствуют тРНК. Гены митохондриальных тРНК имеют последовательности, отличные от ядерных тРНК, но двойники митохондриальных тРНК были обнаружены в ядерных хромосомах с высоким сходством последовательностей. [150]
У животных митохондриальный геном обычно представляет собой одну кольцевую хромосому длиной примерно 16 т.п.н., содержащую 37 генов. Гены, хотя и высококонсервативны, могут различаться по местоположению. Любопытно, что данная закономерность не обнаружена у человеческой телесной вши ( Pediculus humanus ). Вместо этого этот митохондриальный геном состоит из 18 миникольцевых хромосом, каждая из которых имеет длину 3–4 т.п.н. и содержит от одного до трех генов. [151] Этот образец также встречается у других сосущих вшей , но не у жующих вшей . Было показано, что рекомбинация происходит между минихромосомами.
Генетические исследования человеческой популяции
Практически отсутствие генетической рекомбинации в митохондриальной ДНК делает ее полезным источником информации для изучения популяционной генетики и эволюционной биологии . [152] Поскольку вся митохондриальная ДНК наследуется как одна единица или гаплотип , отношения между митохондриальной ДНК разных людей можно представить в виде генного дерева . Закономерности в этих генных деревьях можно использовать для вывода об эволюционной истории популяций. Классическим примером этого является эволюционная генетика человека , где молекулярные часы можно использовать для определения недавней даты появления митохондриальной Евы . [153] [154] Это часто интерпретируется как решительная поддержка недавней современной человеческой экспансии из Африки . [155] Другой человеческий пример — секвенирование митохондриальной ДНК костей неандертальцев . Относительно большое эволюционное расстояние между последовательностями митохондриальной ДНК неандертальцев и современных людей было интерпретировано как свидетельство отсутствия скрещивания между неандертальцами и современными людьми. [156]
Однако митохондриальная ДНК отражает только историю самок в популяции. Частично это можно преодолеть за счет использования отцовских генетических последовательностей, таких как нерекомбинирующая область Y-хромосомы . [155]
Недавние измерения молекулярных часов митохондриальной ДНК [157] сообщили о значении 1 мутации каждые 7884 года, начиная с самого последнего общего предка человека и обезьяны, что согласуется с оценками частоты мутаций аутосомной ДНК (10 −8 на базу на поколение). [158]
Альтернативный генетический код
| Организм | Кодон | Стандартный | Митохондрии |
|---|---|---|---|
| Млекопитающие | НО, АГГ | Аргинин | Стоп-кодон |
| Беспозвоночные | НО, АГГ | Аргинин | Серин |
| Грибы | ВОЗДУХ | Лейцин | Треонин |
| Все вышеперечисленное | НЕ | изолейцин | Метионин |
| УГА | Стоп-кодон | Триптофан |
Хотя небольшие изменения стандартного генетического кода были предсказаны ранее, [159] ничего не было обнаружено до 1979 года, когда исследователи, изучающие митохондриальные гены человека , установили, что они используют альтернативный код. [160] Тем не менее, митохондрии многих других эукариот, включая большинство растений, используют стандартный код. [161] С тех пор было обнаружено множество небольших вариантов. [161] включая различные альтернативные митохондриальные коды. [162] Кроме того, все кодоны AUA, AUC и AUU являются допустимыми стартовыми кодонами.
Некоторые из этих различий следует рассматривать как псевдоизменения генетического кода, обусловленные явлением редактирования РНК , которое часто встречается в митохондриях. Считалось, что у высших растений CGG кодирует триптофан , а не аргинин ; однако было обнаружено, что кодоном в обработанной РНК является кодон UGG, что соответствует стандартному генетическому коду триптофана. [163] Следует отметить, что митохондриальный генетический код членистоногих претерпел параллельную эволюцию внутри одного типа, при этом некоторые организмы уникальным образом переводят AGG в лизин. [164]
Репликация и наследование
Митохондрии делятся путем деления митохондрий , формы бинарного деления , которое также осуществляется бактериями. [165] хотя этот процесс жестко регулируется эукариотической клеткой-хозяином и включает в себя связь и контакт с несколькими другими органеллами. Регуляция этого деления у эукариот различна. У многих одноклеточных эукариот их рост и деление связаны с клеточным циклом . Например, одна митохондрия может делиться синхронно с ядром. Этот процесс деления и сегрегации необходимо строго контролировать, чтобы каждая дочерняя клетка получила хотя бы одну митохондрия. У других эукариот (например, у млекопитающих) митохондрии могут реплицировать свою ДНК и делиться главным образом в ответ на энергетические потребности клетки, а не в фазе клеточного цикла. Когда энергетические потребности клетки высоки, митохондрии растут и делятся. Когда потребление энергии низкое, митохондрии разрушаются или становятся неактивными. В таких примерах митохондрии, по-видимому, случайным образом распределяются по дочерним клеткам при делении цитоплазмы . Митохондриальная динамика, баланс между Слияние и деление митохондрий является важным фактором патологий, связанных с некоторыми болезненными состояниями. [166]
Гипотеза бинарного деления митохондрий основана на визуализации с помощью флуоресцентной микроскопии и традиционной просвечивающей электронной микроскопии (ПЭМ). Разрешение флуоресцентной микроскопии (≈200 нм) недостаточно для различения структурных деталей, таких как двойная митохондриальная мембрана при делении митохондрий, или даже для различения отдельных митохондрий, когда несколько из них расположены близко друг к другу. Традиционный ТЭМ также имеет некоторые технические ограничения. [ который? ] при проверке деления митохондрий. Криоэлектронная томография недавно была использована для визуализации деления митохондрий в замороженных гидратированных интактных клетках. Выяснилось, что митохондрии делятся путем почкования. [167]
Митохондриальные гены особи наследуются только от матери, за редким исключением. [168] У людей, когда яйцеклетка оплодотворяется спермой, митохондрии и, следовательно, митохондриальная ДНК обычно происходят только из яйцеклетки. Митохондрии сперматозоида попадают в яйцеклетку, но не несут генетическую информацию эмбриону. [169] Вместо этого отцовские митохондрии помечаются убиквитином , чтобы выбрать их для последующего разрушения внутри эмбриона . [170] Яйцеклетка содержит относительно мало митохондрий, но эти митохондрии делятся, заселяя клетки взрослого организма. Этот режим наблюдается у большинства организмов, включая большинство животных. Однако митохондрии у некоторых видов иногда могут наследоваться по отцовской линии. Это норма для некоторых хвойных растений, но не для сосен и тисов . [171] У митилид отцовское наследование происходит только у самцов этого вида. [172] [173] [174] Было высказано предположение, что у людей он встречается на очень низком уровне. [175]
Однородительское наследование приводит к ограничению возможностей генетической рекомбинации между разными линиями митохондрий, хотя одна митохондрия может содержать 2–10 копий своей ДНК. [142] Рекомбинация, которая действительно имеет место, поддерживает генетическую целостность, а не разнообразие. Однако есть исследования, показывающие доказательства рекомбинации в митохондриальной ДНК. Понятно, что ферменты, необходимые для рекомбинации, присутствуют в клетках млекопитающих. [176] Кроме того, данные свидетельствуют о том, что митохондрии животных могут подвергаться рекомбинации. [177] Данные более противоречивы у человека, хотя косвенные доказательства рекомбинации существуют. [178] [179]
Можно ожидать, что сущности, претерпевающие однородительское наследование и практически не имеющие рекомбинации, будут подвержены храповому механизму Мюллера — накоплению вредных мутаций до тех пор, пока функциональность не будет потеряна. Популяции митохондрий животных избегают такого накопления благодаря процессу развития, известному как «узкое место» мтДНК . Узкое место использует стохастические процессы в клетке для увеличения межклеточной изменчивости мутантной нагрузки по мере развития организма: одна яйцеклетка с некоторой долей мутантной мтДНК, таким образом, производит эмбрион, в котором разные клетки имеют разную мутантную нагрузку. Затем отбор на уровне клеток может привести к удалению клеток с большим количеством мутантной мтДНК, что приведет к стабилизации или уменьшению мутантной нагрузки между поколениями. Механизм, лежащий в основе узкого места, обсуждается. [180] [181] [182] недавнее математическое и экспериментальное метаисследование предоставило доказательства сочетания случайного разделения мтДНК при делении клетки и случайного оборота молекул мтДНК внутри клетки. [183]
восстановление ДНК
Митохондрии могут восстанавливать окислительные повреждения ДНК по механизмам, аналогичным тем, которые происходят в ядре клетки . Белки, участвующие в репарации мтДНК , кодируются ядерными генами и транслоцируются в митохондрии. Пути репарации ДНК в митохондриях млекопитающих включают репарацию вырезания оснований , репарацию двухцепочечного разрыва, прямое обращение и репарацию ошибочного спаривания . [184] [185] Альтернативно, повреждение ДНК можно обойти, а не восстановить, путем синтеза транслейкоза.
Из нескольких процессов репарации ДНК в митохондриях наиболее полно изучен путь эксцизионной репарации оснований. [185] Репарация эксцизионного основания осуществляется с помощью последовательности этапов, катализируемых ферментами, которые включают распознавание и вырезание поврежденного основания ДНК, удаление образовавшегося абазического сайта, обработку концов, заполнение пробелов и лигирование. Распространенным повреждением мтДНК, которое восстанавливается путем эксцизионной репарации оснований, является 8-оксогуанин, образующийся в результате окисления гуанина . [186]
Двухцепочечные разрывы могут быть устранены путем гомологичной рекомбинационной репарации как в мтДНК млекопитающих, так и в мтДНК млекопитающих. [187] и посадить мтДНК. [188] Двухцепочечные разрывы мтДНК также можно исправить путем микрогомологического соединения концов . [189] Хотя существуют доказательства процессов репарации прямого обращения и репарации несоответствий в мтДНК, эти процессы недостаточно хорошо изучены. [185]
Отсутствие митохондриальной ДНК
Некоторые организмы вообще потеряли митохондриальную ДНК. В этих случаях гены, кодируемые митохондриальной ДНК, теряются или переносятся в ядро. [141] У криптоспоридий есть митохондрии, в которых отсутствует ДНК, предположительно потому, что все их гены были потеряны или перенесены. [190] У Cryptosporidium митохондрии имеют измененную систему генерации АТФ , которая делает паразита устойчивым ко многим классическим митохондриальным ингибиторам, таким как цианид , азид и атоваквон . [190] Митохондрии, лишенные собственной ДНК, были обнаружены у морских паразитических динофлагеллят из рода Amoebophyra . Этот микроорганизм, A. cerati , имеет функциональные митохондрии, лишенные генома. [191] У родственных видов митохондриальный геном еще имеет три гена, но у A. cerati обнаружен только один митохондриальный ген — ген цитохром с-оксидазы I ( cox1 ), и он мигрировал в геном ядра. [192]
Дисфункция и болезнь
Митохондриальные заболевания
Повреждение и последующая дисфункция митохондрий являются важным фактором ряда заболеваний человека из-за их влияния на клеточный метаболизм. Митохондриальные расстройства часто проявляются как неврологические расстройства, включая аутизм . [18] Они также могут проявляться в виде миопатии , диабета , множественной эндокринопатии и множества других системных нарушений. [193] Заболевания, вызванные мутацией мтДНК, включают синдром Кернса-Сейра , синдром MELAS и наследственную оптическую невропатию Лебера . [194] В подавляющем большинстве случаев эти заболевания передаются женщиной своим детям, поскольку зигота получает митохондрии и, следовательно, мтДНК из яйцеклетки. Считается , что такие заболевания, как синдром Кернса-Сейра, синдром Пирсона и прогрессирующая наружная офтальмоплегия, возникают из-за крупномасштабных перестроек мтДНК, тогда как другие заболевания, такие как синдром MELAS, наследственная нейропатия зрительного нерва Лебера, синдром MERRF и другие, возникают из-за точковых мутаций. в мтДНК. [193]
Также сообщалось, что раковые клетки, толерантные к лекарствам, имеют увеличенное количество и размер митохондрий, что предполагает увеличение митохондриального биогенеза. [195] Исследование, проведенное в журнале Nature Nanotechnology в 2022 году , показало, что раковые клетки могут захватывать митохондрии иммунных клеток с помощью физических туннельных нанотрубок. [196]
При других заболеваниях дефекты ядерных генов приводят к дисфункции митохондриальных белков. Это имеет место при атаксии Фридрейха , наследственной спастической параплегии и болезни Вильсона . [197] Эти заболевания наследуются по принципу доминирования , как и большинство других генетических заболеваний. Различные нарушения могут быть вызваны ядерными мутациями ферментов окислительного фосфорилирования, например, дефицит коэнзима Q10 и синдром Барта . [193] Влияние окружающей среды может взаимодействовать с наследственной предрасположенностью и вызывать митохондриальные заболевания. Например, может существовать связь между воздействием пестицидов и более поздним возникновением болезни Паркинсона . [198] [199] Другие патологии, этиология которых связана с митохондриальной дисфункцией, включают шизофрению , биполярное расстройство , деменцию , болезнь Альцгеймера , [200] [201] Болезнь Паркинсона, эпилепсия , инсульт , сердечно-сосудистые заболевания , миалгический энцефаломиелит/синдром хронической усталости (ME/CFS), пигментный ретинит и сахарный диабет . [202] [203]
Митохондриально-опосредованный окислительный стресс играет роль в кардиомиопатии у диабетиков 2 типа . Увеличение доставки жирных кислот в сердце увеличивает поглощение жирных кислот кардиомиоцитами, что приводит к усилению окисления жирных кислот в этих клетках. Этот процесс увеличивает количество восстанавливающих эквивалентов, доступных для цепи переноса электронов митохондрий, что в конечном итоге увеличивает выработку активных форм кислорода (АФК). АФК увеличивают разобщающие белки (UCP) и усиливают утечку протонов через транслокатор адениновых нуклеотидов (ANT), комбинация которых разъединяет митохондрии. Затем разобщение увеличивает потребление кислорода митохондриями, что усугубляет усиление окисления жирных кислот. Это создает порочный круг разъединения; более того, даже несмотря на то, что потребление кислорода увеличивается, синтез АТФ не увеличивается пропорционально, поскольку митохондрии разобщены. Уменьшение доступности АТФ в конечном итоге приводит к дефициту энергии, проявляющемуся снижением эффективности работы сердца и сократительной дисфункцией. Проблема усугубляется тем, что нарушение высвобождения кальция саркоплазматическим ретикулумом и снижение обратного захвата митохондрий ограничивают пиковые цитозольные уровни важного сигнального иона во время мышечного сокращения. Снижение внутримитохондриальной концентрации кальция увеличивает активацию дегидрогеназы и синтез АТФ. Таким образом, помимо снижения синтеза АТФ из-за окисления жирных кислот, синтез АТФ также нарушается из-за плохой передачи сигналов кальция, что вызывает проблемы с сердцем у диабетиков. [204]
Митохондрии также модулируют такие процессы, как развитие соматических клеток яичек, дифференциация сперматогониальных стволовых клеток, подкисление просвета, выработка тестостерона в семенниках и многое другое. Таким образом, дисфункция митохондрий сперматозоидов может быть причиной бесплодия. [205]
В целях борьбы с митохондриальными заболеваниями митохондриальная заместительная терапия была разработана (МЗТ). В этой форме экстракорпорального оплодотворения используются донорские митохондрии, что позволяет избежать передачи заболеваний, вызванных мутациями митохондриальной ДНК. [206] Однако эта терапия все еще исследуется и может привести к генетической модификации, а также к проблемам с безопасностью. Эти заболевания редки, но могут быть чрезвычайно изнурительными и прогрессирующими заболеваниями, что ставит сложные этические вопросы перед государственной политикой. [207]
Связь со старением
Возможна некоторая утечка электронов, переносимых в дыхательной цепи, с образованием активных форм кислорода . Считалось, что это приводит к значительному окислительному стрессу в митохондриях с высокой скоростью мутаций митохондриальной ДНК. [208] Гипотеза о связи между старением и окислительным стрессом не нова и была предложена в 1956 году. [209] которая позже была уточнена в митохондриальную свободнорадикальную теорию старения . [210] Считалось, что возникает порочный круг, поскольку окислительный стресс приводит к мутациям митохондриальной ДНК, что может привести к ферментативным нарушениям и дальнейшему окислительному стрессу.
В процессе старения в митохондриях может произойти ряд изменений. [211] В тканях пожилых людей наблюдается снижение ферментативной активности белков дыхательной цепи. [212] Однако мутировавшую мтДНК можно обнаружить только примерно в 0,2% очень старых клеток. [213] Было высказано предположение, что большие делеции в митохондриальном геноме приводят к высокому уровню окислительного стресса и гибели нейронов при болезни Паркинсона . [214] Также было показано, что митохондриальная дисфункция возникает при боковом амиотрофическом склерозе . [215] [216]
Поскольку митохондрии играют ключевую роль в функции яичников, обеспечивая АТФ, необходимую для развития от зародышевого пузырька до зрелого ооцита , снижение функции митохондрий может привести к воспалению, что приводит к преждевременной недостаточности яичников и ускоренному старению яичников. Возникающая в результате дисфункция затем отражается в количественных (например, количество копий мтДНК и делеции мтДНК), качественных (например, мутации и разрывы цепей) и окислительных повреждениях (например, дисфункция митохондрий из-за АФК), которые имеют значение не только для старения яичников, но нарушают перекрестные помехи ооцитов и кумулюсов в яичнике, связаны с генетическими нарушениями (такими как Fragile X) и могут мешать отбору эмбрионов. [217]
История
Первые наблюдения внутриклеточных структур, которые, вероятно, представляли собой митохондрии, были опубликованы в 1857 году физиологом Альбертом фон Колликером . [218] [219] Ричард Альтманн в 1890 году определил их как клеточные органеллы и назвал «биобластами». [219] [220] В 1898 году Карл Бенда ввёл термин «митохондрии» от греческого μίτος , mitos , «нить», и χονδρίον , хондрия , «гранулы». [221] [219] [222] В 1900 году Леонор Михаэлис обнаружила, что янус-зеленый можно использовать в качестве прижизненного красителя для митохондрий. [223] В 1904 году Фридрих Мевес впервые зарегистрировал наблюдение митохондрий у растений в клетках белой кувшинки Nymphaea alba . [219] [224] а в 1908 году вместе с Клавдием Рего предположил, что они содержат белки и липиды. Бенджамин Ф. Кингсбери в 1912 году впервые связал их с клеточным дыханием, но почти исключительно на основании морфологических наблюдений. [225] [219] В 1913 году Отто Генрих Варбург связал дыхание с частицами, которые он получил из экстракта печени морской свинки и которые он назвал «грана». [226] Варбург и Генрих Отто Виланд , которые также постулировали аналогичный механизм частиц, разошлись во мнениях относительно химической природы дыхания. Лишь в 1925 году, когда Дэвид Кейлин открыл цитохромы , дыхательная цепь была описана. [219]
В 1939 году эксперименты с использованием измельченных мышечных клеток показали, что клеточное дыхание с использованием одной молекулы кислорода может образовывать четыре молекулы аденозинтрифосфата разработал концепцию фосфатных связей АТФ, являющихся формой энергии в клеточном метаболизме. (АТФ), а в 1941 году Фриц Альберт Липманн . В последующие годы механизм клеточного дыхания получил дальнейшее развитие, хотя его связь с митохондриями не была известна. [219] Введение фракционирования тканей Альбертом Клодом позволило изолировать митохондрии от других клеточных фракций и проводить биохимический анализ только на них. В 1946 году он пришел к выводу, что цитохромоксидаза в митохондриях выделены и другие ферменты, ответственные за дыхательную цепь. Юджин Кеннеди и Альберт Ленинджер в 1948 году обнаружили, что митохондрии являются местом окислительного фосфорилирования у эукариот. Со временем метод фракционирования получил дальнейшее развитие, улучшив качество выделяемых митохондрий, и другие элементы клеточного дыхания . было установлено, что в митохондриях происходят [219]
Первые электронные микрофотографии высокого разрешения появились в 1952 году, заменив окраску Януса Грина как предпочтительный способ визуализации митохондрий. [219] Это привело к более детальному анализу структуры митохондрий, включая подтверждение того, что они окружены мембраной. Исследование также показало наличие второй мембраны внутри митохондрий, которая складывалась в гребни, разделяющие внутреннюю камеру, и что размер и форма митохондрий варьировались от клетки к клетке.
Популярный термин «электростанция клетки» был придуман Филипом Сикевицем в 1957 году. [4] [227]
В 1967 году было обнаружено, что митохондрии содержат рибосомы . [228] В 1968 году были разработаны методы картирования митохондриальных генов, а генетическая и физическая карта митохондриальной ДНК дрожжей была завершена в 1976 году. [219]
См. также
Ссылки
- ^ «митохондрия» . Lexico Британский словарь английского языка . Издательство Оксфордского университета . Архивировано из оригинала 2 января 2020 года.
- ^ Кэмпбелл Н.А., Уильямсон Б., Хейден Р.Дж. (2006). Биология: исследование жизни . Бостон, Массачусетс: Пирсон / Прентис Холл . ISBN 978-0132508827 . Архивировано из оригинала 2 ноября 2014 года . Проверено 6 января 2009 г.
- ^ «Могучие митохондрии и нейродегенеративные заболевания» . Наука в новостях . 1 февраля 2012 года. Архивировано из оригинала 6 апреля 2022 года . Проверено 24 апреля 2022 г.
- ^ Перейти обратно: а б Сикевиц П. (1957). «Электростанция клетки». Научный американец . 197 (1): 131–140. Бибкод : 1957SciAm.197a.131S . doi : 10.1038/scientificamerican0757-131 .
- ^ Перейти обратно: а б с д Карнковска А., Вацек В., Зубакова З., Трейтли С.К., Петржелкова Р., Эме Л. и др. (май 2016 г.). «Эукариот без митохондриальной органеллы» . Современная биология . 26 (10): 1274–1284. Бибкод : 2016CBio...26.1274K . дои : 10.1016/j.cub.2016.03.053 . ПМИД 27185558 .
- ^ Ле Пейдж М. «Животное, которому для выживания не нужен кислород, открыто New Scientist» . Новый учёный . Архивировано из оригинала 26 февраля 2020 года . Проверено 25 февраля 2020 г.
- ^ Перейти обратно: а б Яхаломи Д., Аткинсон С.Д., Нойхоф М., Чанг Э.С., Филипп Х., Картрайт П. и др. (март 2020 г.). «У книдарийного паразита лосося (Myxozoa: Henneguya ) отсутствует митохондриальный геном» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (10): 5358–5363. Бибкод : 2020PNAS..117.5358Y . дои : 10.1073/pnas.1909907117 . ПМК 7071853 . ПМИД 32094163 .
- ^ Перейти обратно: а б Хенце К., Мартин В. (ноябрь 2003 г.). «Эволюционная биология: сущность митохондрий» . Природа . 426 (6963): 127–128. Бибкод : 2003Natur.426..127H . дои : 10.1038/426127a . ПМИД 14614484 . S2CID 862398 .
- ^ Леже М.М., Колиско М., Камикава Р., Лестница К.В., Куме К., Чепичка И. и др. (апрель 2017 г.). «Органеллы, проливающие свет на происхождение гидрогеносом трихомонады и лямблий митосом » . Экология и эволюция природы . 1 (4): 0092. Бибкод : 2017NatEE...1...92L . дои : 10.1038/s41559-017-0092 . ПМК 5411260 . ПМИД 28474007 .
- ^ Перейти обратно: а б Новак Л.В., Трейтли С.С., Пирих Дж., Халакуц П., Пипалия С.В., Вацек В. и др. (декабрь 2023 г.). Датчер С.К. (ред.). «Геномика жгутиконосцев Preaxostyla освещает путь к потере митохондрий» . ПЛОС Генетика . 19 (12): e1011050. дои : 10.1371/journal.pgen.1011050 . ПМЦ 10703272 . ПМИД 38060519 .
- ^ Вимерслаге Л., Ли Д. (март 2016 г.). «Количественная оценка морфологии митохондрий в нейритах дофаминергических нейронов с использованием нескольких параметров» . Журнал методов нейробиологии . 262 : 56–65. doi : 10.1016/j.jneumeth.2016.01.008 . ПМЦ 4775301 . ПМИД 26777473 .
- ^ Перейти обратно: а б с Макбрайд Х.М., Нойшпиль М., Васиак С. (июль 2006 г.). «Митохондрии: больше, чем просто источник энергии» . Современная биология . 16 (14): 551–560 р. Бибкод : 2006CBio...16.R551M . дои : 10.1016/j.cub.2006.06.054 . ПМИД 16860735 . S2CID 16252290 .
- ^ Валеро Т (2014). «Митохондриальный биогенез: фармакологические подходы». Текущий фармацевтический дизайн . 20 (35): 5507–5509. дои : 10.2174/138161282035140911142118 . hdl : 10454/13341 . ПМИД 24606795 .
Таким образом, митохондриальный биогенез определяется как процесс, посредством которого клетки увеличивают свою индивидуальную митохондриальную массу [3]. ... Митохондриальный биогенез происходит путем роста и деления ранее существовавших органелл и временно координируется с событиями клеточного цикла [1].
- ^ Санчис-Гомар Ф, Гарсиа-Хименес ХЛ, Гомес-Кабрера МК, Паллардо ФВ (2014). «Митохондриальный биогенез в здоровье и болезни. Молекулярные и терапевтические подходы». Текущий фармацевтический дизайн . 20 (35): 5619–5633. дои : 10.2174/1381612820666140306095106 . ПМИД 24606801 .
Митохондриальный биогенез (МБ) является важным механизмом, с помощью которого клетки контролируют количество митохондрий.
- ^ Гарднер А., Болес Р.Г. (2005). «Есть ли в будущем «митохондриальная психиатрия»? Обзор». Курс. Психиатрия преп . 1 (3): 255–271. дои : 10.2174/157340005774575064 .
- ^ Леснефски Э.Дж., Могаддас С., Тандлер Б., Кернер Дж., Хоппель К.Л. (июнь 2001 г.). «Митохондриальная дисфункция при сердечных заболеваниях: ишемия-реперфузия, старение и сердечная недостаточность». Журнал молекулярной и клеточной кардиологии . 33 (6): 1065–1089. дои : 10.1006/jmcc.2001.1378 . ПМИД 11444914 .
- ^ Дорн Г.В., Вега Р.Б., Келли Д.П. (октябрь 2015 г.). «Митохондриальный биогенез и динамика в развивающемся и больном сердце» . Гены и развитие . 29 (19): 1981–1991. дои : 10.1101/gad.269894.115 . ПМЦ 4604339 . ПМИД 26443844 .
- ^ Перейти обратно: а б Гриффитс К.К., Леви Р.Дж. (2017). «Свидетельства митохондриальной дисфункции при аутизме: биохимические связи, генетические ассоциации и неэнергетические механизмы» . Окислительная медицина и клеточное долголетие . 2017 : 4314025. doi : 10.1155/2017/4314025 . ПМЦ 5467355 . ПМИД 28630658 .
- ^ Ней П.А. (май 2011 г.). «Нормальное и нарушенное созревание ретикулоцитов» . Современное мнение в гематологии . 18 (3): 152–157. дои : 10.1097/MOH.0b013e328345213e . ПМК 3157046 . ПМИД 21423015 .
- ^ Перейти обратно: а б с д и ж г час я дж Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2005). Молекулярная биология клетки . Garland Publishing Inc. Нью-Йорк: ISBN 978-0815341055 .
- ^ Перейти обратно: а б с д и ж г час я дж к л Voet D, Voet JC, Пратт CW (2006). Основы биохимии (2-е изд.). John Wiley and Sons, Inc., стр. 547, 556 . ISBN 978-0471214953 .
- ^ Андерссон С.Г. , Карлберг О., Канбек Б., Курланд К.Г. (январь 2003 г.). «О происхождении митохондрий: взгляд на геномику» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 358 (1429): 165–77, обсуждение 177–9. дои : 10.1098/rstb.2002.1193 . ПМК 1693097 . ПМИД 12594925 .
- ^ Перейти обратно: а б Габальдон Т. (октябрь 2021 г.). «Происхождение и ранняя эволюция эукариотической клетки». Ежегодный обзор микробиологии . 75 (1): 631–647. doi : 10.1146/annurev-micro-090817-062213 . ПМИД 34343017 . S2CID 236916203 .
- ^ «Митохондрия – гораздо больше, чем преобразователь энергии» . Британское общество клеточной биологии. Архивировано из оригинала 4 апреля 2019 года . Проверено 19 августа 2013 г.
- ^ Блакли-Дайсон Э., Форте М (сентябрь 2001 г.). «Каналы VDAC» . ИУБМБ Жизнь . 52 (3–5): 113–118. дои : 10.1080/15216540152845902 . ПМИД 11798022 . S2CID 38314888 .
- ^ Хугенбум Б.В., Суда К., Энгель А., Фотиадис Д. (июль 2007 г.). «Супрамолекулярные ансамбли потенциалзависимых анионных каналов в нативной мембране». Журнал молекулярной биологии . 370 (2): 246–255. дои : 10.1016/j.jmb.2007.04.073 . ПМИД 17524423 .
- ^ Зет К. (июнь 2010 г.). «Структура и эволюция белков наружной мембраны митохондрий топологии бета-цилиндра» . Biochimica et Biophysica Acta (BBA) – Биоэнергетика . 1797 (6–7): 1292–1299. дои : 10.1016/j.bbabio.2010.04.019 . ПМИД 20450883 .
- ^ Перейти обратно: а б Херрманн Дж. М., Нойперт В. (апрель 2000 г.). «Транспорт белков в митохондрии» (PDF) . Современное мнение в микробиологии . 3 (2): 210–214. дои : 10.1016/S1369-5274(00)00077-1 . ПМИД 10744987 . Архивировано (PDF) из оригинала 16 августа 2022 г. Проверено 10 июля 2022 г.
- ^ Перейти обратно: а б Чипук Дж. Э., Бушье-Хейс Л., Грин Д. Р. (август 2006 г.). «Пермеабилизация внешней мембраны митохондрий во время апоптоза: сценарий невинного свидетеля» . Смерть клеток и дифференцировка . 13 (8): 1396–1402. дои : 10.1038/sj.cdd.4401963 . ПМИД 16710362 .
- ^ Перейти обратно: а б с д и Хаяши Т., Риццуто Р., Хайноцки Г., Су Т.П. (февраль 2009 г.). «МАМ: больше, чем просто домработница» . Тенденции в клеточной биологии . 19 (2): 81–88. дои : 10.1016/j.tcb.2008.12.002 . ПМК 2750097 . ПМИД 19144519 .
- ^ Шенкель Л.К., Бакович М. (январь 2014 г.). «Формирование и регуляция митохондриальных мембран» . Международный журнал клеточной биологии . 2014 : 709828. doi : 10.1155/2014/709828 . ПМЦ 3918842 . ПМИД 24578708 .
- ^ Макмиллин Дж. Б., Доуэн В. (декабрь 2002 г.). «Кардиолипин и апоптоз». Biochimica et Biophysical Acta (BBA) - Молекулярная и клеточная биология липидов . 1585 (2–3): 97–107. дои : 10.1016/S1388-1981(02)00329-3 . ПМИД 12531542 .
- ^ Баутиста Дж.С., Фалабелла М., Фланнери П.Дж., Ханна М.Г., Хилз С.Дж., Поуп С.А. и др. (декабрь 2022 г.). «Достижения в методах анализа кардиолипина и их клиническое применение» . Тенденции в аналитической химии . 157 : 116808. doi : 10.1016/j.trac.2022.116808 . ПМЦ 7614147 . ПМИД 36751553 . S2CID 253211400 .
- ^ Юл Р.Дж., ван дер Блик А.М. (август 2012 г.). «Деление, слияние и стресс митохондрий» . Наука . 337 (6098): 1062–1065. Бибкод : 2012Sci...337.1062Y . дои : 10.1126/science.1219855 . ПМК 4762028 . ПМИД 22936770 .
- ^ Череп С, Посфаи Б, Шварц А.Д., Денес А (2018). «Митохондриальная ультраструктура связана с синаптической деятельностью в местах высвобождения аксонов» . эНейро . 5 (1): ЭНЕВРО.0390–17.2018. дои : 10.1523/ENEURO.0390-17.2018 . ПМЦ 5788698 . ПМИД 29383328 .
- ^ Маннелла, Калифорния (2006). «Структура и динамика крист внутренней мембраны митохондрий». Биохимия и биофизика Acta (BBA) - Исследования молекулярных клеток . 1763 (5–6): 542–548. дои : 10.1016/j.bbamcr.2006.04.006 . ПМИД 16730811 .
- ^ Богенхаген DF (сентябрь 2012 г.). «Структура нуклеоида митохондриальной ДНК». Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1819 (9–10): 914–920. дои : 10.1016/j.bbagrm.2011.11.005 . ПМИД 22142616 .
- ^ Богатый пиар (декабрь 2003 г.). «Молекулярный механизм дыхательной цепи Кейлина». Труды Биохимического общества . 31 (Часть 6): 1095–1105. дои : 10.1042/BST0311095 . ПМИД 14641005 .
- ^ Стойменова М., Игамбердиев А.У., Гупта К.Дж., Хилл Р.Д. (июль 2007 г.). «Анаэробный синтез АТФ, управляемый нитритами, в митохондриях корней ячменя и риса». Планта . 226 (2): 465–474. Бибкод : 2007Завод.226..465С . дои : 10.1007/s00425-007-0496-0 . ПМИД 17333252 . S2CID 8963850 .
- ^ Нойперт В. (1997). «Импорт белка в митохондрии». Ежегодный обзор биохимии . 66 : 863–917. doi : 10.1146/annurev.biochem.66.1.863 . ПМИД 9242927 .
- ^ Перейти обратно: а б с д и ж Страйер Л. (1995). «Цикл лимонной кислоты». В: Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. стр. 509–527, 569–579, 614–616, 638–641, 732–735, 739–748, 770–773. ISBN 0716720094 .
- ^ Кинг А., Селак М.А., Готлиб Э. (август 2006 г.). «Сукцинатдегидрогеназа и фумаратгидратаза: связь митохондриальной дисфункции и рака». Онкоген . 25 (34): 4675–4682. дои : 10.1038/sj.onc.1209594 . ПМИД 16892081 . S2CID 26263513 .
- ^ Перейти обратно: а б Воет Д., Воет Дж.Г. (2004). Биохимия (3-е изд.). Нью-Йорк, штат Нью-Йорк: Уайли. п. 804. ИСБН 978-0-471-19350-0 .
- ^ Перейти обратно: а б Аткинс П., де Паула Дж (2006). «Влияние на биохимию: преобразование энергии в биологических клетках». Физическая химия (8-е изд.). Нью-Йорк: Фриман. стр. 225–229. ISBN 978-0-7167-8759-4 .
- ^ Хуан Х., Мантон К.Г. (май 2004 г.). «Роль окислительного повреждения митохондрий при старении: обзор» (PDF) . Границы бионауки . 9 (1–3): 1100–1117. дои : 10.2741/1298 . ПМИД 14977532 . S2CID 2278219 . Архивировано из оригинала (PDF) 3 марта 2019 г.
- ^ Митчелл П., Мойл Дж. (январь 1967 г.). «Хемиосмотическая гипотеза окислительного фосфорилирования». Природа . 213 (5072): 137–139. Бибкод : 1967Natur.213..137M . дои : 10.1038/213137a0 . ПМИД 4291593 . S2CID 4149605 .
- ^ Митчелл П. (июнь 1967 г.). «Протонный ток в митохондриальных системах». Природа . 214 (5095): 1327–1328. Бибкод : 1967Natur.214.1327M . дои : 10.1038/2141327a0 . ПМИД 6056845 . S2CID 4160146 .
- ^ Нобелевский фонд. «Химия 1997» . Архивировано из оригинала 8 июля 2007 года . Проверено 16 декабря 2007 г.
- ^ Перейти обратно: а б Мозо Дж., Эмре Ю., Буйо Ф., Рикье Д., Крискуоло Ф. (ноябрь 2005 г.). «Терморегуляция: какова роль UCP у млекопитающих и птиц?». Отчеты по биологическим наукам . 25 (3–4): 227–249. дои : 10.1007/s10540-005-2887-4 . ПМИД 16283555 . S2CID 164450 .
- ^ Кастаниотис А.Дж., Аутио К.Дж., Кератар Дж.М., Монтеууис Г., Мякеля А.М., Наир Р.Р. и др. (январь 2017 г.). «Синтез митохондриальных жирных кислот, жирные кислоты и физиология митохондрий». Биохимика и биофизика Acta. Молекулярная и клеточная биология липидов . 1862 (1): 39–48. дои : 10.1016/j.bbalip.2016.08.011 . ПМИД 27553474 .
- ^ Клэй Х.Б., Парл А.К., Митчелл С.Л., Сингх Л., Белл Л.Н., Мердок Д.Г. (март 2016 г.). Петерсон Дж. (ред.). «Изменение пути синтеза митохондриальных жирных кислот (mtFASII) модулирует клеточные метаболические состояния и профили биоактивных липидов, как показано с помощью метаболомного профилирования» . ПЛОС ОДИН . 11 (3): e0151171. Бибкод : 2016PLoSO..1151171C . дои : 10.1371/journal.pone.0151171 . ПМЦ 4786287 . ПМИД 26963735 .
- ^ Перейти обратно: а б с Новински С.М., Ван Вранкен Дж.Г., Дав К.К., Раттер Дж. (октябрь 2018 г.). «Влияние синтеза митохондриальных жирных кислот на митохондриальный биогенез» . Современная биология . 28 (20): Р1212–Р1219. Бибкод : 2018CBio...28R1212N . дои : 10.1016/j.cub.2018.08.022 . ПМК 6258005 . ПМИД 30352195 .
- ^ Вебе З., Берингер С., Алатиби К., Уоткинс Д., Розенблатт Д., Шпикеркоттер У. и др. (ноябрь 2019 г.). «Новая роль митохондриальной синтазы жирных кислот (mtFASII) в регуляции энергетического метаболизма». Биохимика и биофизика Acta. Молекулярная и клеточная биология липидов . 1864 (11): 1629–1643. дои : 10.1016/j.bbalip.2019.07.012 . ПМИД 31376476 . S2CID 199404906 .
- ^ Сантулли Г., Се В., Рейкен С.Р., Маркс А.Р. (сентябрь 2015 г.). «Перегрузка митохондрий кальцием является ключевым фактором, определяющим сердечную недостаточность» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (36): 11389–11394. Бибкод : 2015PNAS..11211389S . дои : 10.1073/pnas.1513047112 . ПМЦ 4568687 . ПМИД 26217001 .
- ^ Перейти обратно: а б Сигел Г.Дж., Агранов Б.В., Фишер С.К., Альберс Р.В., Улер, доктор медицинских наук, ред. (1999). Основная нейрохимия (6-е изд.). Липпинкотт Уильямс и Уилкинс. ISBN 978-0397518203 .
- ^ Перейти обратно: а б Россье МФ (август 2006 г.). «Т-каналы и биосинтез стероидов: в поисках связи с митохондриями». Клеточный кальций . 40 (2): 155–164. дои : 10.1016/j.ceca.2006.04.020 . ПМИД 16759697 .
- ^ Брайтон, Коннектикут, Хант Р.М. (май 1974 г.). «Митохондриальный кальций и его роль в кальцификации. Гистохимическая локализация кальция на электронных микрофотографиях эпифизарной пластинки роста с K-пироантимонатом». Клиническая ортопедия и связанные с ней исследования . 100 (5): 406–416. дои : 10.1097/00003086-197405000-00057 . ПМИД 4134194 .
- ^ Брайтон, Коннектикут, Хант Р.М. (июль 1978 г.). «Роль митохондрий в кальцификации пластинок роста, продемонстрированная на рахитической модели». Журнал костной и суставной хирургии. Американский том . 60 (5): 630–639. дои : 10.2106/00004623-197860050-00007 . ПМИД 681381 .
- ^ Перейти обратно: а б с Сантулли Г., Маркс А.Р. (2015). «Основная роль каналов внутриклеточного высвобождения кальция в мышцах, мозге, обмене веществ и старении». Современная молекулярная фармакология . 8 (2): 206–222. дои : 10.2174/1874467208666150507105105 . ПМИД 25966694 .
- ^ Пиццо П., Поццан Т. (октябрь 2007 г.). «Хореография митохондрий-эндоплазматического ретикулума: структура и динамика передачи сигналов». Тенденции в клеточной биологии . 17 (10): 511–517. дои : 10.1016/j.tcb.2007.07.011 . ПМИД 17851078 .
- ^ Перейти обратно: а б Миллер Р.Дж. (март 1998 г.). «Митохондрии – Кракен просыпается!». Тенденции в нейронауках . 21 (3): 95–97. дои : 10.1016/S0166-2236(97)01206-X . ПМИД 9530913 . S2CID 5193821 .
- ^ Сантулли Г., Пагано Г., Сарду С., Се В., Рейкен С., Д'Асия С.Л. и др. (май 2015 г.). «Канал высвобождения кальция RyR2 регулирует высвобождение инсулина и гомеостаз глюкозы» . Журнал клинических исследований . 125 (5): 1968–1978. дои : 10.1172/JCI79273 . ПМК 4463204 . ПМИД 25844899 .
- ^ Шварцлендер М., Логан Д.С., Джонстон И.Г., Джонс Н.С., Мейер А.Дж., Фрикер М.Д. и др. (март 2012 г.). «Пульсация мембранного потенциала в отдельных митохондриях: вызванный стрессом механизм регулирования дыхательной биоэнергетики у Arabidopsis» . Растительная клетка . 24 (3): 1188–1201. дои : 10.1105/tpc.112.096438 . ПМЦ 3336130 . ПМИД 22395486 .
- ^ Иванников М.В., Маклауд Г.Т. (июнь 2013 г.). «Уровни свободного Ca²⁺ в митохондриях и их влияние на энергетический обмен в окончаниях двигательных нервов дрозофилы» . Биофизический журнал . 104 (11): 2353–2361. Бибкод : 2013BpJ...104.2353I . дои : 10.1016/j.bpj.2013.03.064 . ПМЦ 3672877 . ПМИД 23746507 .
- ^ Вайнберг Ф., Чандель Н.С. (октябрь 2009 г.). «Митохондриальный метаболизм и рак». Анналы Нью-Йоркской академии наук . 1177 (1): 66–73. Бибкод : 2009NYASA1177...66W . дои : 10.1111/j.1749-6632.2009.05039.x . ПМИД 19845608 . S2CID 29827252 .
- ^ Перейти обратно: а б Морено-Санчес Р., Родригес-Энрикес С., Марин-Эрнандес А., Сааведра Э. (март 2007 г.). «Энергетический обмен в опухолевых клетках». ФЭБС Журнал 274 (6): 1393–1418. дои : 10.1111/j.1742-4658.2007.05686.x . ПМИД 17302740 . S2CID 7748115 .
- ^ Мистри Дж.Дж., Марлейн С.Р., Мур Дж.А., Хеллмих С., Войтович Е.Е., Смит Дж.Г. и др. (декабрь 2019 г.). «АФК-опосредованная активация PI3K стимулирует перенос митохондрий из стромальных клеток в гемопоэтические стволовые клетки в ответ на инфекцию» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (49): 24610–24619. Бибкод : 2019PNAS..11624610M . дои : 10.1073/pnas.1913278116 . ПМК 6900710 . ПМИД 31727843 .
- ^ Педерсен П.Л. (декабрь 1994 г.). «АТФ-синтаза. Машина, производящая АТФ». Современная биология . 4 (12): 1138–1141. Бибкод : 1994CBio....4.1138P . дои : 10.1016/S0960-9822(00)00257-8 . ПМИД 7704582 . S2CID 10279742 .
- ^ Паттаппа Дж., Хейвуд Х.К., де Брёйн Дж.Д., Ли Д.А. (октябрь 2011 г.). «Метаболизм мезенхимальных стволовых клеток человека во время пролиферации и дифференцировки». Журнал клеточной физиологии . 226 (10): 2562–2570. дои : 10.1002/jcp.22605 . ПМИД 21792913 . S2CID 22259833 .
- ^ Агарвал Б. (июнь 2011 г.). «Роль анионов в синтезе АТФ и ее молекулярно-механическая интерпретация». Журнал биоэнергетики и биомембран . 43 (3): 299–310. дои : 10.1007/s10863-011-9358-3 . ПМИД 21647635 . S2CID 29715383 .
- ^ Перейти обратно: а б с Sweet S, Сингх Дж. (июль 1999 г.). «Изменения массы митохондрий, мембранного потенциала и содержания клеточного аденозинтрифосфата во время клеточного цикла лейкозных клеток человека (HL-60)». Журнал клеточной физиологии . 180 (1): 91–96. doi : 10.1002/(SICI)1097-4652(199907)180:1<91::AID-JCP10>3.0.CO;2-6 . ПМИД 10362021 . S2CID 25906279 .
- ^ Риера Ромо М (август 2021 г.). «Гибель клеток как часть врожденного иммунитета: причина или следствие?» . Иммунология . 163 (4): 399–415. дои : 10.1111/imm.13325 . ПМЦ 8274179 . ПМИД 33682112 .
- ^ Зеленый ДР (сентябрь 1998 г.). «Пути апоптоза: пути к гибели» . Клетка . 94 (6): 695–698. дои : 10.1016/S0092-8674(00)81728-6 . ПМИД 9753316 . S2CID 16654082 .
- ^ Перейти обратно: а б с д и ж Бахат А., МакВикар Т., Лангер Т. (27 июля 2021 г.). «Метаболизм и врожденный иммунитет встречаются в митохондриях» . Границы клеточной биологии и биологии развития . 9 : 720490. doi : 10.3389/fcell.2021.720490 . ПМЦ 8353256 . ПМИД 34386501 .
- ^ Перейти обратно: а б с д и ж Член парламента Мерфи, О'Нил, Лос-Анджелес (февраль 2024 г.). «Разрыв митохондриального эндосимбиоза как основа воспалительных заболеваний». Природа . 626 (7998): 271–279. Бибкод : 2024Natur.626..271M . дои : 10.1038/s41586-023-06866-z . ПМИД 38326590 .
- ^ Крыско Д.В., Агостинис П., Крыско О., Гарг А.Д., Бахерт С., Ламбрехт Б.Н. и др. (апрель 2011 г.). «Новая роль молекулярных паттернов, связанных с повреждением митохондрий, при воспалении». Тенденции в иммунологии . 32 (4): 157–164. дои : 10.1016/j.it.2011.01.005 . ПМИД 21334975 . ; цитируется в [75]
- ^ Райли Дж.С., Тейт С.В. (апрель 2020 г.). «Митохондриальная ДНК в воспалении и иммунитете» . Отчеты ЭМБО . 21 (4): e49799. дои : 10.15252/эмбр.201949799 . ПМК 7132203 . ПМИД 32202065 . ; цитируется в [75]
- ^ Сет Р.Б., Сунь Л., Эа К.К., Чен З.Дж. (сентябрь 2005 г.). «Идентификация и характеристика MAVS, митохондриального противовирусного сигнального белка, который активирует NF-kappaB и IRF 3». Клетка . 122 (5): 669–682. дои : 10.1016/j.cell.2005.08.012 . ПМИД 16125763 . ; цитируется в [75]
- ^ Дорвард Д.А., Лукас С.Д., Доэрти М.К., Чепмен ГБ, Шоулфилд Э.Дж., Конвей Моррис А. и др. (октябрь 2017 г.). «Новая роль эндогенного митохондриального формилированного пептида, управляемого рецептором формилпептида 1, при остром респираторном дистресс-синдроме» . Торакс . 72 (10): 928–936. doi : 10.1136/thoraxjnl-2017-210030 . ПМЦ 5738532 . ПМИД 28469031 . ; цитируется в [75]
- ^ Кай Н., Гомес-Дюран А., Йонова-Дуинг Е., Кунду К., Берджесс А.И., Голдер З.Дж. и др. (сентябрь 2021 г.). «Варианты митохондриальной ДНК модулируют N-формилметионин, протеостаз и риск заболеваний человека с поздним началом». Природная медицина . 27 (9): 1564–1575. дои : 10.1038/s41591-021-01441-3 . ПМИД 34426706 . ; цитируется в [75]
- ^ Чжан В., Ван Г., Сюй З.Г., Ту Х., Ху Ф., Дай Дж. и др. (июнь 2019 г.). «Лактат является естественным подавителем передачи сигналов RLR путем воздействия на MAVS» . Клетка . 178 (1): 176–189.e15. дои : 10.1016/j.cell.2019.05.003 . ПМК 6625351 . ПМИД 31155231 . ; цитируется в [74]
- ^ Пурсело М., Арну Д. (сентябрь 2014 г.). «Митохондриальная динамика и врожденный противовирусный иммунный ответ». Журнал ФЭБС . 281 (17): 3791–3802. дои : 10.1111/февраль 12940 . ПМИД 25051991 . ; цитируется в [74]
- ^ Ли X, Фанг П., Май Дж., Чой Э.Т., Ван Х., Ян XF (февраль 2013 г.). «Нацеливание на митохондриальные активные формы кислорода в качестве новой терапии воспалительных заболеваний и рака» . Журнал гематологии и онкологии . 6 (19): 19. дои : 10.1186/1756-8722-6-19 . ПМЦ 3599349 . ПМИД 23442817 .
- ^ Хайночки Г., Чордас Г., Дас С., Гарсиа-Перес С., Саотоме М., Синха Рой С. и др. (2006). «Передача сигналов митохондриального кальция и гибель клеток: подходы к оценке роли поглощения митохондриями Ca2+ в апоптозе» . Клеточный кальций . 40 (5–6): 553–560. дои : 10.1016/j.ceca.2006.08.016 . ПМЦ 2692319 . ПМИД 17074387 .
- ^ О-хама Т. (август 1997 г.). «Эволюционное рассмотрение 5-аминолевулинатсинтазы в природе». Происхождение жизни и эволюция биосферы . 27 (4): 405–412. Бибкод : 1997OLEB...27..405O . дои : 10.1023/А:1006583601341 . ПМИД 9249985 . S2CID 13602877 .
- ^ Клинге CM (декабрь 2008 г.). «Эстрогенный контроль функции митохондрий и биогенеза» . Журнал клеточной биохимии . 105 (6): 1342–1351. дои : 10.1002/jcb.21936 . ПМК 2593138 . ПМИД 18846505 .
- ^ Альварес-Дельгадо К., Мендоса-Родригес К.А., Пикасо О., Сербон М. (август 2010 г.). «Различная экспрессия альфа- и бета-митохондриальных рецепторов эстрогена в мозге стареющих крыс: взаимодействие с дыхательным комплексом V». Экспериментальная геронтология . 45 (7–8): 580–585. дои : 10.1016/j.exger.2010.01.015 . ПМИД 20096765 . S2CID 30841790 .
- ^ Павон Н., Мартинес-Абундис Э., Эрнандес Л., Галлардо-Перес Х.К., Альварес-Дельгадо С., Сербон М. и др. (октябрь 2012 г.). «Половые гормоны: влияние на сердечную и митохондриальную активность после ишемии-реперфузии у взрослых крыс. Гендерные различия». Журнал биохимии стероидов и молекулярной биологии . 132 (1–2): 135–146. дои : 10.1016/j.jsbmb.2012.05.003 . ПМИД 22609314 . S2CID 24794040 .
- ^ Бреда К.Н., Даванзо Г.Г., Бассо П.Дж., Сараива Камара Н.О., Мораес-Виейра П.М. (сентябрь 2019 г.). «Митохондрии как центральный узел иммунной системы» . Редокс-биология . 26 : 101255. doi : 10.1016/j.redox.2019.101255 . ПМК 6598836 . ПМИД 31247505 .
- ^ Перейти обратно: а б Череп С., Посфаи Б., Ленарт Н., Фекете Р., Ласло З.И., Леле З. и др. (январь 2020 г.). «Микроглия контролирует и защищает функции нейронов посредством специализированных соматических пуринергических соединений» . Наука . 367 (6477): 528–537. Бибкод : 2020Sci...367..528C . doi : 10.1126/science.aax6752 . ПМИД 31831638 . S2CID 209343260 . Архивировано из оригинала 2 мая 2023 года . Проверено 10 февраля 2022 г.
- ^ Череп С., Шварц А.Д., Посфаи Б., Ласло З.И., Келлермайер А., Корней З. и др. (сентябрь 2022 г.). «Микроглиальный контроль развития нейронов через соматические пуринергические соединения» . Отчеты по ячейкам . 40 (12): 111369. doi : 10.1016/j.celrep.2022.111369 . ПМЦ 9513806 . ПМИД 36130488 . S2CID 252416407 .
- ^ Тейлор С.В., Фэйи Э., Чжан Б., Гленн Г.М., Уорнок Д.Э., Уайли С. и др. (март 2003 г.). «Характеристика митохондриального протеома сердца человека». Природная биотехнология . 21 (3): 281–286. дои : 10.1038/nbt793 . ПМИД 12592411 . S2CID 27329521 .
- ^ Чжан Дж., Ли Х., Мюллер М., Ван Ю., Цзун С., Дэн Н. и др. (апрель 2008 г.). «Систематическая характеристика протеома митохондрий мыши с использованием функционально подтвержденных митохондрий сердца» . Протеомика . 8 (8): 1564–1575. дои : 10.1002/pmic.200700851 . ПМЦ 2799225 . ПМИД 18348319 .
- ^ Чжан Дж., Лием Д.А., Мюллер М., Ван Ю., Цзун С., Дэн Н. и др. (июнь 2008 г.). «Измененная протеомная биология сердечных митохондрий в условиях стресса» . Журнал исследований протеома . 7 (6): 2204–2214. дои : 10.1021/pr070371f . ПМЦ 3805274 . ПМИД 18484766 .
- ^ Логан, округ Колумбия (июнь 2010 г.). «Слияние, деление и расположение митохондрий у растений». Труды Биохимического общества . 38 (3): 789–795. дои : 10.1042/bst0380789 . ПМИД 20491666 .
- ^ дас Невес Р.П., Джонс Н.С., Андреу Л., Гупта Р., Энвер Т., Иборра Ф.Дж. (декабрь 2010 г.). Вайсман Дж.С. (ред.). «Связь изменчивости глобальной скорости транскрипции с изменчивостью митохондрий» . ПЛОС Биология . 8 (12): е1000560. дои : 10.1371/journal.pbio.1000560 . ПМК 3001896 . ПМИД 21179497 .
- ^ Джонстон И.Г., Гаал Б., Невес Р.П., Энвер Т., Иборра Ф.Дж., Джонс Н.С. (2012). Хо Дж. М. (ред.). «Митохондриальная изменчивость как источник внешнего клеточного шума» . PLOS Вычислительная биология . 8 (3): e1002416. arXiv : 1107.4499 . Бибкод : 2012PLSCB...8E2416J . дои : 10.1371/journal.pcbi.1002416 . ПМЦ 3297557 . ПМИД 22412363 .
- ^ Раппапорт Л., Оливьеро П., Сэмюэл Дж.Л. (июль 1998 г.). «Морфология и функция цитоскелета и митохондрий». Молекулярная и клеточная биохимия . 184 (1–2): 101–105. дои : 10.1023/A:1006843113166 . ПМИД 9746315 . S2CID 28165195 .
- ^ Хойцинг Х., Джонстон И.Г., Джонс Н.С. (июнь 2015 г.). «Какова функция митохондриальных сетей? Теоретическая оценка гипотез и предложения для будущих исследований» . Биоэссе . 37 (6): 687–700. doi : 10.1002/bies.201400188 . ПМЦ 4672710 . ПМИД 25847815 .
- ^ Солтыс Б.Дж., Гупта Р.С. (1992). «Взаимоотношения эндоплазматической сети, митохондрий, промежуточных филаментов и микротрубочек - исследование четырехкратного флуоресцентного мечения». Биохимия и клеточная биология = Biochimie et Biologie Cellulaire . 70 (10–11): 1174–1186. дои : 10.1139/o92-163 . ПМИД 1363623 .
- ^ Тан Х.Л., Лунг Х.Л., Ву К.С., Ле АХ, Тан Х.М., Фунг MC (февраль 2008 г.). «Виментин поддерживает морфологию и организацию митохондрий». Биохимический журнал . 410 (1): 141–146. дои : 10.1042/BJ20071072 . ПМИД 17983357 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Риццуто Р., Марчи С., Бонора М., Агиари П., Бонони А., Де Стефани Д. и др. (ноябрь 2009 г.). «Перенос Ca(2+) из ЭР в митохондрии: когда, как и почему» . Biochimica et Biophysica Acta (BBA) – Биоэнергетика . 1787 (11): 1342–1351. дои : 10.1016/j.bbabio.2009.03.015 . ПМК 2730423 . ПМИД 19341702 .
- ^ Перейти обратно: а б де Брито О.М., Скоррано Л. (август 2010 г.). «Интимная связь: пространственная организация взаимоотношений эндоплазматического ретикулума и митохондрий» . Журнал ЭМБО . 29 (16): 2715–2723. дои : 10.1038/emboj.2010.177 . ПМЦ 2924651 . ПМИД 20717141 .
- ^ Перейти обратно: а б Вэнс Дж. Э., Шиао Ю. Дж. (1996). «Внутриклеточный транспорт фосфолипидов: импорт фосфатидилсерина в митохондрии». Противораковые исследования . 16 (3Б): 1333–1339. ПМИД 8694499 .
- ^ Перейти обратно: а б с Лебедзинская М., Шабадкай Г., Джонс А.В., Душинский Дж., Вецковски М.Р. (октябрь 2009 г.). «Взаимодействие между эндоплазматической сетью, митохондриями, плазматической мембраной и другими субклеточными органеллами». Международный журнал биохимии и клеточной биологии . 41 (10): 1805–1816. doi : 10.1016/j.biocel.2009.02.017 . ПМИД 19703651 .
- ^ Твиг Дж., Элорза А., Молина А.Дж., Мохамед Х., Викстром Дж.Д., Уолцер Дж. и др. (январь 2008 г.). «Деление и избирательное слияние управляют сегрегацией и уничтожением митохондрий посредством аутофагии» . Журнал ЭМБО . 27 (2): 433–446. дои : 10.1038/sj.emboj.7601963 . ПМЦ 2234339 . ПМИД 18200046 .
- ^ Перейти обратно: а б с д и ж Осман С., Фёлкер Д.Р., Лангер Т. (январь 2011 г.). «Создание голов или хвостов фосфолипидов в митохондриях» . Журнал клеточной биологии . 192 (1): 7–16. дои : 10.1083/jcb.201006159 . ПМК 3019561 . ПМИД 21220505 .
- ^ Корнманн Б., Карри Э., Коллинз С.Р., Шульдинер М., Нуннари Дж., Вайсман Дж.С. и др. (июль 2009 г.). «Комплекс, связывающий ЭР-митохондрии, выявленный с помощью синтетического биологического анализа» . Наука . 325 (5939): 477–481. Бибкод : 2009Sci...325..477K . дои : 10.1126/science.1175088 . ПМЦ 2933203 . ПМИД 19556461 .
- ^ Русиньол А.Е., Цуй З., Чен М.Х., Вэнс Дж.Е. (ноябрь 1994 г.). «Уникальная мембранная фракция печени крысы, связанная с митохондриями, обладает высокой способностью к синтезу липидов и содержит секреторные белки пре-Гольджи, включая образующиеся липопротеины» . Журнал биологической химии . 269 (44): 27494–27502. дои : 10.1016/S0021-9258(18)47012-3 . ПМИД 7961664 .
- ^ Перейти обратно: а б Копач О, Кругликов И, Пивнева Т, Войтенко Н, Федирко Н (май 2008 г.). «Функциональная связь между рианодиновыми рецепторами, митохондриями и Ca (2+) АТФазами в подчелюстных ацинарных клетках крысы». Клеточный кальций . 43 (5): 469–481. дои : 10.1016/j.ceca.2007.08.001 . ПМИД 17889347 .
- ^ Чордаш Г., Хайночки Г. (апрель 2001 г.). «Сортировка сигналов кальция на стыках эндоплазматической сети и митохондрий». Клеточный кальций . 29 (4): 249–262. дои : 10.1054/ceca.2000.0191 . ПМИД 11243933 .
- ^ Перейти обратно: а б с д Декайпер Ж.П., Монако Г., Бултынк Г., Миссиен Л., Де Смедт Х., Парис Ж.Б. (май 2011 г.). «Связь рецептора IP (3) с митохондриями при апоптозе и аутофагии». Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1813 (5): 1003–1013. дои : 10.1016/j.bbamcr.2010.11.023 . ПМИД 21146562 .
- ^ Диркс Б.П., Флигерт Р., Гузе А.Х. (июнь 2017 г.). «Mag-Fluo4 в Т-клетках: визуализация свободного кальция внутри органелл. 2+ концентрации» . Biochimica et Biophysica Acta. Molecular Cell Research . 1864 (6): 977–986. doi : 10.1016/j.bbamcr.2016.11.026 . PMID 27913206 .
- ^ Хайночки Г., Чордас Г., Йи М. (2011). «Старые игроки в новой роли: митохондриально-ассоциированные мембраны, VDAC и рианодиновые рецепторы как участники распространения сигнала кальция от эндоплазматического ретикулума к митохондриям». Клеточный кальций . 32 (5–6): 363–377. дои : 10.1016/S0143416002001872 . ПМИД 12543096 .
- ^ Marriott KS, Прасад М., Таплиял В., Бозе HS (декабрь 2012 г.). «Рецептор σ-1 на мембране митохондриально-ассоциированного эндоплазматического ретикулума отвечает за регуляцию митохондриального метаболизма» . Журнал фармакологии и экспериментальной терапии . 343 (3): 578–586. дои : 10.1124/jpet.112.198168 . ПМК 3500540 . ПМИД 22923735 .
- ^ Хаяши Т., Су Т.П. (ноябрь 2007 г.). «Шапероны рецептора сигма-1 на границе раздела ЭР-митохондрии регулируют передачу сигналов Ca (2+) и выживаемость клеток» . Клетка . 131 (3): 596–610. дои : 10.1016/j.cell.2007.08.036 . ПМИД 17981125 . S2CID 18885068 .
- ^ Чордас Г., Уивер Д., Хайночки Г. (июль 2018 г.). «Эндоплазматический ретикулум-митохондриальная контактология: структура и сигнальные функции» . Тенденции в клеточной биологии . 28 (7): 523–540. дои : 10.1016/j.tcb.2018.02.009 . ПМК 6005738 . ПМИД 29588129 .
- ^ Маккатчеон Дж.П. (октябрь 2021 г.). «Геномика и клеточная биология внутриклеточных инфекций, полезных для хозяина» . Ежегодный обзор клеточной биологии и биологии развития . 37 (1): 115–142. doi : 10.1146/annurev-cellbio-120219-024122 . ПМИД 34242059 . S2CID 235786110 .
- ^ Каллиер V (8 июня 2022 г.). «Митохондрии и происхождение эукариот» . Знающий журнал . doi : 10.1146/knowable-060822-2 . S2CID 249526889 . Архивировано из оригинала 15 августа 2022 года . Проверено 18 августа 2022 г.
- ^ Перейти обратно: а б Маргулис Л., Саган Д. (1986). Происхождение пола: три миллиарда лет генетической рекомбинации . Нью-Хейвен, Коннектикут: Издательство Йельского университета. стр. 69–71, 87 . ISBN 978-0300033403 .
- ^ Мартин В.Ф., Мюллер М. (2007). Происхождение митохондрий и гидрогеносом . Гейдельберг, Германия: Springer Verlag.
- ^ Емельянов В.В. (апрель 2003 г.). «Митохондриальная связь с происхождением эукариотической клетки» . Европейский журнал биохимии . 270 (8): 1599–1618. дои : 10.1046/j.1432-1033.2003.03499.x . ПМИД 12694174 .
- ^ Мюллер М., Мартин В. (май 1999 г.). «Геном Rickettsia prowazekii и некоторые мысли о происхождении митохондрий и гидрогеносом». Биоэссе . 21 (5): 377–381. doi : 10.1002/(sici)1521-1878(199905)21:5<377::aid-bies4>3.0.co;2-w . ПМИД 10376009 .
- ^ Грей М.В., Бургер Дж., Ланг Б.Ф. (март 1999 г.). «Митохондриальная эволюция» . Наука . 283 (5407): 1476–1481. Бибкод : 1999Sci...283.1476G . дои : 10.1126/science.283.5407.1476 . ПМЦ 3428767 . ПМИД 10066161 .
- ^ Трэш Дж.С., Бойд А., Хаггетт М.Дж., Гроте Дж., Карини П., Йодер Р.Дж. и др. (14 июня 2011 г.). «Филогеномные доказательства общего предка митохондрий и клады SAR11» . Научные отчеты . 1 (1): 13. Бибкод : 2011НатСР...1Е..13Т . дои : 10.1038/srep00013 . ПМК 3216501 . ПМИД 22355532 .
- ^ Мартейн Дж., Воссеберг Дж., Гай Л., Оффре П., Эттема Т.Дж. (май 2018 г.). «Глубокое митохондриальное происхождение за пределами отобранных альфапротеобактерий» . Природа . 557 (7703): 101–105. Бибкод : 2018Natur.557..101M . дои : 10.1038/s41586-018-0059-5 . hdl : 1874/373336 . ПМИД 29695865 . S2CID 13740626 .
- ^ Фань Л., Ву Д., Горемыкин В., Сяо Дж., Сюй Ю., Гарг С. и др. (сентябрь 2020 г.). «Филогенетический анализ с систематическим отбором таксонов показывает, что митохондрии разветвляются внутри Alphaproteobacteria». Экология и эволюция природы . 4 (9). ООО «Спрингер Сайенс энд Бизнес Медиа»: 1213–1219. Бибкод : 2020NatEE...4.1213F . bioRxiv 10.1101/715870 . дои : 10.1038/s41559-020-1239-x . ПМИД 32661403 . S2CID 220507452 .
- ^ Ван С., Ло Х (июнь 2021 г.). «Датирование эволюции альфапротеобактерий по окаменелостям эукариот» . Природные коммуникации . 12 (1): 3324. Бибкод : 2021NatCo..12.3324W . дои : 10.1038/s41467-021-23645-4 . ПМЦ 8175736 . ПМИД 34083540 .
- ^ Эспости, доктор медицинских наук, Гейгер О., Санчес-Флорес А. (16 мая 2022 г.). «О бактериальном происхождении митохондрий: новые идеи с помощью триангулированных подходов». bioRxiv : 2022.05.15.491939. дои : 10.1101/2022.05.15.491939 . S2CID 248836055 .
- ^ Муньос-Гомес С.А., Суско Э., Уильямсон К., Эме Л., Сламовиц С.Х., Морейра Д. и др. (март 2022 г.). «Анализ гетерогенных сайтов и ветвей расширенного набора данных свидетельствует о том, что митохондрии являются сестрами известных альфапротеобактерий» (PDF) . Экология и эволюция природы . 6 (3): 253–262. Бибкод : 2022NatEE...6..253M . дои : 10.1038/s41559-021-01638-2 . ПМИД 35027725 . S2CID 245958471 . Архивировано (PDF) из оригинала 18 ноября 2023 г. Проверено 30 декабря 2023 г.
- ^ Шён М.Е., Мартейн Дж., Воссеберг Дж., Кёстльбахер С., Эттема Т.Дж. (август 2022 г.). «Эволюционное происхождение ассоциации хозяев у Rickettsiales» . Природная микробиология . 7 (8): 1189–1199. дои : 10.1038/s41564-022-01169-x . ПМЦ 9352585 . PMID 35798888 .
- ^ О'Брайен Т.В. (сентябрь 2003 г.). «Свойства митохондриальных рибосом человека». ИУБМБ Жизнь . 55 (9): 505–513. дои : 10.1080/15216540310001626610 . ПМИД 14658756 . S2CID 43415449 .
- ^ Саган Л. (март 1967 г.). «О происхождении митозирующих клеток». Журнал теоретической биологии . 14 (3): 255–274. Бибкод : 1967JThBi..14..225S . дои : 10.1016/0022-5193(67)90079-3 . ПМИД 11541392 .
- ^ Емельянов В.В. (февраль 2001 г.). «Риккетсии, риккетсиоподобные эндосимбионты и происхождение митохондрий». Отчеты по биологическим наукам . 21 (1): 1–17. дои : 10.1023/А:1010409415723 . ПМИД 11508688 . S2CID 10144020 .
- ^ Фэн Д.Ф., Чо Дж., Дулиттл РФ (ноябрь 1997 г.). «Определение времени расхождения с помощью белковых часов: обновление и переоценка» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (24): 13028–13033. Бибкод : 1997PNAS...9413028F . дои : 10.1073/pnas.94.24.13028 . ПМК 24257 . ПМИД 9371794 .
- ^ Кавалер-Смит Т. (1991). «Архамебы: предки эукариотов?». Биосистемы . 25 (1–2): 25–38. Бибкод : 1991BiSys..25...25C . дои : 10.1016/0303-2647(91)90010-I . ПМИД 1854912 .
- ^ Дановаро Р., Делл'Анно А., Пушедду А., Гамби С., Хайнер И., Кристенсен Р.М. (апрель 2010 г.). «Первые многоклеточные животные, живущие в постоянно бескислородных условиях» . БМК Биология . 8:30 . дои : 10.1186/1741-7007-8-30 . ПМЦ 2907586 . ПМИД 20370908 .
- ^ Когхалн А. (7 апреля 2010 г.). «Грязевое существо, живущее без кислорода» . Зоолог. Новый учёный . Архивировано из оригинала 26 февраля 2020 года . Проверено 27 февраля 2020 г.
- ^ Шифлетт А.М., Джонсон П.Дж. (2010). «Органеллы, связанные с митохондриями, у эукариотических простейших» . Ежегодный обзор микробиологии . 64 : 409–429. дои : 10.1146/annurev.micro.62.081307.162826 . ПМК 3208401 . ПМИД 20528687 .
- ^ Карнковска А., Трейтли С.К., Брзонь О., Новак Л., Вацек В., Соукал П. и др. (октябрь 2019 г.). Баттистуцци Ф.У. (ред.). «Геном оксимонад демонстрирует каноническую эукариотическую сложность в отсутствие митохондрии» . Молекулярная биология и эволюция . 36 (10): 2292–2312. дои : 10.1093/molbev/msz147 . ПМК 6759080 . ПМИД 31387118 . Архивировано из оригинала 12 апреля 2024 года . Проверено 14 апреля 2024 г.
- ^ Перейти обратно: а б с Чан, округ Колумбия (июнь 2006 г.). «Митохондрии: динамические органеллы при заболеваниях, старении и развитии» . Клетка . 125 (7): 1241–1252. дои : 10.1016/j.cell.2006.06.010 . ПМИД 16814712 . S2CID 8551160 .
- ^ Перейти обратно: а б Виснер Р.Дж., Рюэгг Дж.К., Морано I (март 1992 г.). «Подсчет целевых молекул с помощью экспоненциальной полимеразной цепной реакции: количество копий митохондриальной ДНК в тканях крысы». Связь с биохимическими и биофизическими исследованиями . 183 (2): 553–559. дои : 10.1016/0006-291X(92)90517-O . ПМИД 1550563 .
- ^ Фалькенберг М. (июль 2018 г.). «Репликация митохондриальной ДНК в клетках млекопитающих: обзор пути» . Очерки по биохимии . 62 (3): 287–296. дои : 10.1042/ebc20170100 . ПМК 6056714 . ПМИД 29880722 .
- ^ Перейти обратно: а б Андерсон С., Банкир А.Т., Баррелл Б.Г., де Брёйн М.Х., Коулсон А.Р., Друэн Дж. и др. (апрель 1981 г.). «Последовательность и организация митохондриального генома человека». Природа . 290 (5806): 457–465. Бибкод : 1981Natur.290..457A . дои : 10.1038/290457a0 . ПМИД 7219534 . S2CID 4355527 .
- ^ Фукухара Х., Сор Ф., Дрисси Р., Диноэль Н., Миякава И., Руссе С. и др. (апрель 1993 г.). «Линейные митохондриальные ДНК дрожжей: частота встречаемости и общие особенности» . Молекулярная и клеточная биология . 13 (4): 2309–2314. дои : 10.1128/mcb.13.4.2309 . ПМЦ 359551 . ПМИД 8455612 .
- ^ Бернарди Дж. (декабрь 1978 г.). «Промежуточные последовательности в митохондриальном геноме». Природа . 276 (5688): 558–559. Бибкод : 1978Natur.276..558B . дои : 10.1038/276558a0 . ПМИД 214710 . S2CID 4314378 .
- ^ Хеббар С.К., Белчер С.М., Перлман П.С. (апрель 1992 г.). «Интрон группы IIA дрожжевых митохондрий, кодирующий матуразу, самосращивается in vitro» . Исследования нуклеиновых кислот . 20 (7): 1747–1754. дои : 10.1093/нар/20.7.1747 . ПМК 312266 . ПМИД 1579468 .
- ^ Грей М.В., Ланг Б.Ф., Седергрен Р., Голдинг Г.Б., Лемье С., Санкофф Д. и др. (февраль 1998 г.). «Структура генома и содержание генов в митохондриальных ДНК простейших» . Исследования нуклеиновых кислот . 26 (4): 865–878. дои : 10.1093/нар/26.4.865 . ПМК 147373 . ПМИД 9461442 .
- ^ Грей М.В., Ланг Б.Ф., Бургер Дж. (2004). «Митохондрии протистов». Ежегодный обзор генетики . 38 : 477–524. дои : 10.1146/annurev.genet.37.110801.142526 . ПМИД 15568984 .
- ^ Телонис А.Г., Лохер П., Кирино Ю., Ригуцос И. (2014). «Ядерные и митохондриальные тРНК-двойники в геноме человека» . Границы генетики . 5 : 344. дои : 10.3389/fgene.2014.00344 . ПМК 4189335 . ПМИД 25339973 .
- ^ Шао Р., Киркнесс Э.Ф., Баркер С.С. (май 2009 г.). «Единственная митохондриальная хромосома, типичная для животных, превратилась в 18 минихромосом у человеческой платяной вши Pediculus humanus» . Геномные исследования . 19 (5): 904–912. дои : 10.1101/гр.083188.108 . ПМЦ 2675979 . ПМИД 19336451 .
- ^ Кастро Х.А., Пикорнелл А., Рамон М. (декабрь 1998 г.). «Митохондриальная ДНК: инструмент исследований популяционной генетики». Международная микробиология . 1 (4): 327–332. ПМИД 10943382 .
- ^ Канн Р.Л., Стоункинг М., Уилсон AC (январь 1987 г.). «Митохондриальная ДНК и эволюция человека». Природа . 325 (6099): 31–36. Бибкод : 1987Natur.325...31C . дои : 10.1038/325031a0 . ПМИД 3025745 . S2CID 4285418 .
- ^ Торрони А., Ахилли А., Маколей В., Ричардс М., Бандельт Х.Дж. (июнь 2006 г.). «Сбор плодов дерева мтДНК человека». Тенденции в генетике . 22 (6): 339–345. дои : 10.1016/j.tig.2006.04.001 . PMID 16678300 .
- ^ Перейти обратно: а б Гарриган Д., Хаммер М.Ф. (сентябрь 2006 г.). «Реконструкция происхождения человека в эпоху генома». Обзоры природы. Генетика . 7 (9): 669–680. дои : 10.1038/nrg1941 . ПМИД 16921345 . S2CID 176541 .
- ^ Крингс М., Стоун А., Шмитц Р.В., Крайницки Х., Стоункинг М., Паабо С. (июль 1997 г.). «Последовательности ДНК неандертальцев и происхождение современного человека». Клетка . 90 (1): 19–30. дои : 10.1016/S0092-8674(00)80310-4 . hdl : 11858/00-001M-0000-0025-0960-8 . ПМИД 9230299 . S2CID 13581775 .
- ^ Соарес П., Эрмини Л., Томсон Н., Мормина М., Рито Т., Рёль А. и др. (июнь 2009 г.). «Поправка на очищающий отбор: улучшенные митохондриальные молекулярные часы человека» . Американский журнал генетики человека . 84 (6): 740–759. дои : 10.1016/j.ajhg.2009.05.001 . ПМЦ 2694979 . ПМИД 19500773 .
- ^ Нахман М.В., Кроуэлл С.Л. (сентябрь 2000 г.). «Оценка частоты мутаций на нуклеотид у человека» . Генетика . 156 (1): 297–304. дои : 10.1093/генетика/156.1.297 . ПМЦ 1461236 . ПМИД 10978293 .
- ^ Крик Ф.Х., Оргель Л.Е. (1973). «Направленная панспермия» (PDF) . Икар . 19 (3): 341–346. Бибкод : 1973Icar...19..341C . дои : 10.1016/0019-1035(73)90110-3 . Архивировано (PDF) из оригинала 29 октября 2011 г. Проверено 21 октября 2014 г.
п. 344: Немного удивительно, что организмы с несколько разными кодами не сосуществуют.
Дальнейшее обсуждение. Архивировано 3 сентября 2011 года в Wayback Machine . - ^ Баррелл Б.Г., Банкир А.Т., Друэн Дж. (ноябрь 1979 г.). «Другой генетический код в митохондриях человека». Природа . 282 (5735): 189–194. Бибкод : 1979Natur.282..189B . дои : 10.1038/282189a0 . ПМИД 226894 . S2CID 4335828 .
- ^ Перейти обратно: а б Эльжановский А., Остелл Дж. (7 января 2019 г.). «Генетические коды» . www.ncbi.nlm.nih.gov . Архивировано из оригинала 13 мая 2011 года . Проверено 10 февраля 2023 г.
- ^ Джукс Т.Х., Осава С. (декабрь 1990 г.). «Генетический код в митохондриях и хлоропластах». Эксперименты . 46 (11–12): 1117–1126. дои : 10.1007/BF01936921 . ПМИД 2253709 . S2CID 19264964 .
- ^ Хизель Р., Виссинджер Б., Шустер В., Бреннике А. (декабрь 1989 г.). «Редактирование РНК в митохондриях растений». Наука . 246 (4937): 1632–1634. Бибкод : 1989Sci...246.1632H . дои : 10.1126/science.2480644 . ПМИД 2480644 .
- ^ Абаскаль Ф., Посада Д., Найт Р.Д., Зардоя Р. (май 2006 г.). «Параллельная эволюция генетического кода в митохондриальных геномах членистоногих» . ПЛОС Биология . 4 (5): е127. doi : 10.1371/journal.pbio.0040127 . ПМК 1440934 . ПМИД 16620150 .
- ^ Пфайффер РФ (2012). Болезнь Паркинсона . ЦРК Пресс. п. 583. ИСБН 978-1439807149 .
- ^ Со А.Ю., Джозеф А.М., Датта Д., Хван Дж.К., Арис Дж.П., Леувенбург К. (август 2010 г.). «Новый взгляд на роль митохондрий в старении: динамика митохондрий и многое другое» . Журнал клеточной науки . 123 (Часть 15): 2533–2542. дои : 10.1242/jcs.070490 . ПМЦ 2912461 . ПМИД 20940129 .
- ^ Ху ГБ (август 2014 г.). «Целоклеточная криоэлектронная томография предполагает деление митохондрий путем почкования». Микроскопия и микроанализ . 20 (4): 1180–1187. Бибкод : 2014MiMic..20.1180H . дои : 10.1017/S1431927614001317 . ПМИД 24870811 . S2CID 7440669 .
- ^ МакВильямс Т.Г., Суомалайнен А. (январь 2019 г.). «Митохондриальная ДНК может быть унаследована от отца, а не только от матери» . Природа . 565 (7739): 296–297. Бибкод : 2019Natur.565..296M . дои : 10.1038/d41586-019-00093-1 . ПМИД 30643304 .
- ^ Кимбалл, JW (2006) «Половое размножение у людей: совокупление и оплодотворение». Архивировано 2 октября 2015 г., в Wayback Machine , страницы биологии Кимбалла (на основе биологии , 6-е изд., 1996 г.).
- ^ Сутовский П., Морено Р.Д., Рамальо-Сантос Дж., Доминко Т., Симерли С., Шаттен Г. (ноябрь 1999 г.). «Убиквитиновая метка для митохондрий сперматозоидов». Природа . 402 (6760): 371–372. Бибкод : 1999Natur.402..371S . дои : 10.1038/46466 . ПМИД 10586873 . S2CID 205054671 . Обсуждается в Science News, архивировано 19 декабря 2007 г. в Wayback Machine .
- ^ Могенсен Х.Л. (1996). «Как и почему цитоплазматическая наследственность у семенных растений». Американский журнал ботаники . 83 (3): 383–404. дои : 10.2307/2446172 . JSTOR 2446172 .
- ^ Зурос Э (декабрь 2000 г.). «Исключительная система митохондриальной ДНК семейства мидий Mytilidae» . Гены и генетические системы . 75 (6): 313–318. дои : 10.1266/ggs.75.313 . ПМИД 11280005 .
- ^ Сазерленд Б., Стюарт Д., Кенчингтон Э.Р., Зурос Э. (январь 1998 г.). «Судьба отцовской митохондриальной ДНК у развивающихся самок мидий Mytilus edulis: значение для механизма двойного однородительского наследования митохондриальной ДНК» . Генетика . 148 (1): 341–347. дои : 10.1093/генетика/148.1.341 . ПМЦ 1459795 . ПМИД 9475744 .
- ↑ голубых мидий (Mytilus edulis) Линии мужской и женской митохондриальной ДНК в группе видов . Архивировано 18 мая 2013 года в Wayback Machine Дональдом Т. Стюартом, Карлосом Сааведрой, Ребеккой Р. Стэнвуд, Эми О. Болл и Элефтериосом Зуросом.
- ^ Джонс Д.Р. (октябрь 2003 г.). «Отцовская передача митохондриальной ДНК (к счастью) редка». Анналы неврологии . 54 (4): 422–424. дои : 10.1002/ана.10771 . ПМИД 14520651 . S2CID 90422 .
- ^ Тьягараджан Б., Падуя Р.А., Кэмпбелл С. (ноябрь 1996 г.). «Митохондрии млекопитающих обладают гомологичной рекомбинационной активностью ДНК» . Журнал биологической химии . 271 (44): 27536–27543. дои : 10.1074/jbc.271.44.27536 . ПМИД 8910339 .
- ^ Лант Д.Х., Хайман, Британская Колумбия (май 1997 г.). «Рекомбинация митохондриальной ДНК животных» . Природа . 387 (6630): 247. Бибкод : 1997Natur.387..247L . дои : 10.1038/387247a0 . ПМИД 9153388 . S2CID 4318161 .
- ^ Эйр-Уокер А., Смит Н.Х., Смит Дж.М. (март 1999 г.). «Насколько клональны митохондрии человека?» . Слушания. Биологические науки . 266 (1418): 477–483. дои : 10.1098/rspb.1999.0662 . ПМЦ 1689787 . ПМИД 10189711 .
- ^ Авадалла П., Эйр-Уокер А., Смит Дж. М. (декабрь 1999 г.). «Неравновесие связей и рекомбинация в митохондриальной ДНК человекообразных». Наука . 286 (5449): 2524–2525. дои : 10.1126/science.286.5449.2524 . ПМИД 10617471 . S2CID 27685285 .
- ^ Кри Л.М., Сэмюэлс Д.С., де Соуза Лопес С.К., Раджасимха Х.К., Воннапинидж П., Манн Дж.Р. и др. (февраль 2008 г.). «Уменьшение молекул митохондриальной ДНК во время эмбриогенеза объясняет быстрое разделение генотипов». Природная генетика . 40 (2): 249–254. дои : 10.1038/ng.2007.63 . ПМИД 18223651 . S2CID 205344980 .
- ^ Цао Л., Шитара Х., Хории Т., Нагао Ю., Имаи Х., Абэ К. и др. (март 2007 г.). «Узкое место в митохондриях возникает без снижения содержания мтДНК в зародышевых клетках самок мышей». Природная генетика . 39 (3): 386–390. дои : 10.1038/ng1970 . ПМИД 17293866 . S2CID 10686347 .
- ^ Вай Т., Теоли Д., Шубридж Э.А. (декабрь 2008 г.). «Генетическое узкое место митохондриальной ДНК возникает в результате репликации субпопуляции геномов». Природная генетика . 40 (12): 1484–1488. дои : 10.1038/ng.258 . ПМИД 19029901 . S2CID 225349 .
- ^ Джонстон И.Г., Бургсталлер Дж.П., Гавличек В., Кольбе Т., Рюлике Т., Брем Г. и др. (июнь 2015 г.). «Стохастическое моделирование, байесовский вывод и новые измерения in vivo проясняют обсуждаемый механизм узкого места мтДНК» . электронная жизнь . 4 : e07464. arXiv : 1512.02988 . дои : 10.7554/eLife.07464 . ПМЦ 4486817 . ПМИД 26035426 .
- ^ Гредилья Р., Гарм С., Стевнснер Т. (2012). «Репарация ядерной и митохондриальной ДНК в некоторых модельных системах старения эукариот» . Окислительная медицина и клеточное долголетие . 2012 : 282438. doi : 10.1155/2012/282438 . ПМЦ 3462412 . ПМИД 23050036 .
- ^ Перейти обратно: а б с Саки М., Пракаш А. (июнь 2017 г.). «Перекрестные помехи, связанные с повреждением ДНК, между ядром и митохондриями» . Свободно-радикальная биология и медицина . 107 : 216–227. doi : 10.1016/j.freeradbiomed.2016.11.050 . ПМЦ 5449269 . ПМИД 27915046 .
- ^ Леон Дж., Сакуми К., Кастильо Э., Шэн З., Ока С., Накабеппу Ю. (февраль 2016 г.). «Накопление 8-оксогуанина в митохондриальной ДНК вызывает митохондриальную дисфункцию и нарушает нейрогенез в культивируемых корковых нейронах взрослых мышей в окислительных условиях» . Научные отчеты . 6 : 22086. Бибкод : 2016NatSR...622086L . дои : 10.1038/srep22086 . ПМЦ 4766534 . ПМИД 26912170 .
- ^ Дахал С., Дубей С., Рагхаван СК (май 2018 г.). «В митохондриях млекопитающих действует репарация двухцепочечных разрывов ДНК, опосредованная гомологичной рекомбинацией» . Клеточные и молекулярные науки о жизни . 75 (9): 1641–1655. дои : 10.1007/s00018-017-2702-y . ПМЦ 11105789 . ПМИД 29116362 . S2CID 4779650 .
- ^ Одахара М., Иноуе Т., Фудзита Т., Хасебе М., Секине Ю. (февраль 2007 г.). «Участие RecA, нацеленного на митохондрии, в восстановлении митохондриальной ДНК мха Physcomitrella patens» . Гены и генетические системы . 82 (1): 43–51. дои : 10.1266/ggs.82.43 . ПМИД 17396019 .
- ^ Тади С.К., Себастьян Р., Дахал С., Бабу Р.К., Чоудхари Б., Рагхаван С.К. (январь 2016 г.). «Соединение концов, опосредованное микрогомологией, является основным медиатором восстановления двухцепочечных разрывов во время повреждений митохондриальной ДНК» . Молекулярная биология клетки . 27 (2): 223–235. doi : 10.1091/mbc.E15-05-0260 . ПМЦ 4713127 . ПМИД 26609070 .
- ^ Перейти обратно: а б Энрикес Ф.Л., Ричардс Т.А., Робертс Ф., МакЛеод Р., Робертс К.В. (февраль 2005 г.). «Необычный митохондриальный отсек Cryptosporidium parvum». Тенденции в паразитологии . 21 (2): 68–74. дои : 10.1016/j.pt.2004.11.010 . ПМИД 15664529 .
- ^ Джон У., Лу Ю., Вольраб С., Грот М., Янушковец Дж., Кохли Г.С. и др. (апрель 2019 г.). «Аэробный эукариотический паразит с функциональными митохондриями, у которого, вероятно, отсутствует митохондриальный геном» . Достижения науки . 5 (4): eaav1110. Бибкод : 2019SciA....5.1110J . дои : 10.1126/sciadv.aav1110 . ПМК 6482013 . ПМИД 31032404 .
- ^ «Настоящая электростанция – даже без ДНК: паразитические водоросли из линии динофлагеллят организовали свой генетический материал беспрецедентным образом» . ScienceDaily . Архивировано из оригинала 24 июня 2019 года . Проверено 8 мая 2019 г.
- ^ Перейти обратно: а б с Зевиани М., Ди Донато С. (октябрь 2004 г.). «Митохондриальные нарушения» . Мозг . 127 (Часть 10): 2153–2172. дои : 10.1093/brain/awh259 . ПМИД 15358637 .
- ^ Тейлор Р.В., Тернбулл DM (май 2005 г.). «Мутации митохондриальной ДНК при заболеваниях человека» . Обзоры природы. Генетика . 6 (5): 389–402. дои : 10.1038/nrg1606 . ПМК 1762815 . ПМИД 15861210 .
- ^ Голдман А., Хисте С., Фрейнкман Е., Дхаван А., Маджумдер Б., Мондал Дж. и др. (август 2019 г.). «Нацеливание на фенотипическую пластичность опухолей и метаболическое ремоделирование при адаптивной перекрестной лекарственной толерантности» . Научная сигнализация . 12 (595). doi : 10.1126/scisignal.aas8779 . ПМЦ 7261372 . ПМИД 31431543 .
- ^ Саха Т., Даш С., Джаябалан Р., Хисте С., Кулкарни А., Курми К. и др. (январь 2022 г.). «Межклеточные нанотрубки опосредуют транспорт митохондрий между раковыми и иммунными клетками» . Природные нанотехнологии . 17 (1): 98–106. Бибкод : 2022НатНа..17...98С . дои : 10.1038/s41565-021-01000-4 . ПМЦ 10071558 . ПМИД 34795441 . S2CID 244349825 .
- ^ Чиннери П.Ф., Шон Е.А. (сентябрь 2003 г.). «Митохондрии» . Журнал неврологии, нейрохирургии и психиатрии . 74 (9): 1188–1199. дои : 10.1136/jnnp.74.9.1188 . ПМЦ 1738655 . ПМИД 12933917 .
- ^ Шерер Т.Б., Бетарбет Р., Гринамир Дж.Т. (июнь 2002 г.). «Окружающая среда, митохондрии и болезнь Паркинсона». Нейробиолог . 8 (3): 192–197. дои : 10.1177/1073858402008003004 . ПМИД 12061498 . S2CID 24318009 .
- ^ Гомес С., Бандез М.Дж., Наварро А. (январь 2007 г.). «Пестициды и нарушение функции митохондрий в связи с паркинсоническим синдромом» . Границы бионауки . 12 : 1079–1093. дои : 10.2741/2128 . ПМИД 17127363 .
- ^ Лим Ю.А., Рейн В., Байсанг Г., Мейер Ф., Поляк А., Рафтери М.Дж. и др. (апрель 2010 г.). «Абета и человеческий амилин имеют общий путь токсичности через митохондриальную дисфункцию». Протеомика . 10 (8): 1621–1633. дои : 10.1002/pmic.200900651 . ПМИД 20186753 . S2CID 33077667 .
- ^ Кинг СП, Лян В.Г., Шерпельц К.П., Шиллинг А.Б., Мередит С.К., Тан В.Дж. (июль 2014 г.). «Молекулярные основы распознавания и деградации субстратов протеазой препоследовательности человека» . Структура . 22 (7): 996–1007. doi : 10.1016/j.str.2014.05.003 . ПМК 4128088 . ПМИД 24931469 .
- ^ Шапира А.Х. (июль 2006 г.). «Митохондриальная болезнь». Ланцет . 368 (9529): 70–82. дои : 10.1016/S0140-6736(06)68970-8 . ПМИД 16815381 . S2CID 27039617 .
- ^ Печеник С.Р., Нойштадт Дж. (август 2007 г.). «Митохондриальная дисфункция и молекулярные пути заболеваний». Экспериментальная и молекулярная патология . 83 (1): 84–92. дои : 10.1016/j.yexmp.2006.09.008 . ПМИД 17239370 .
- ^ Баггер Х., Абель Э.Д. (ноябрь 2010 г.). «Митохондрии в диабетическом сердце» . Сердечно-сосудистые исследования . 88 (2): 229–240. дои : 10.1093/cvr/cvq239 . ПМЦ 2952534 . ПМИД 20639213 .
- ^ Подолак А., Воцлавек-Потоцка И., Лукашук К. (февраль 2022 г.). «Роль митохондрий в фертильности человека и раннем развитии эмбриона: что мы можем узнать для клинического применения оценки и улучшения митохондриальной ДНК?» . Клетки . 11 (5): 797. doi : 10.3390/cells11050797 . ПМЦ 8909547 . ПМИД 35269419 .
- ^ Мэй-Панлуп П., Богене М., Хашем Х.Э., Буэ П.Е., Рейнье П. (январь 2021 г.). «Эмбрион и его митохондрии» . Антиоксиданты . 10 (2): 139. дои : 10.3390/antiox10020139 . ПМЦ 7908991 . ПМИД 33498182 .
- ^ Комитет по рассмотрению этической и социальной политики новых методов профилактики материнской передачи заболеваний митохондриальной ДНК; Совет по политике в области медицинских наук; Медицинский институт; Национальные академии наук, техники и медицины (17 марта 2016 г.), Клэйборн А., Инглиш Р., Кан Дж. (ред.), «Введение» , Методы замены митохондрий: этические, социальные и политические соображения , National Academies Press (США) ) , получено 5 декабря 2023 г.
- ^ Рихтер С., Парк Дж.В., Эймс Б.Н. (сентябрь 1988 г.). «Обычное окислительное повреждение митохондриальной и ядерной ДНК обширно» . Труды Национальной академии наук Соединенных Штатов Америки . 85 (17): 6465–6467. Бибкод : 1988PNAS...85.6465R . дои : 10.1073/pnas.85.17.6465 . ПМК 281993 . ПМИД 3413108 .
- ^ Харман Д. (июль 1956 г.). «Старение: теория, основанная на свободнорадикальной и радиационной химии». Журнал геронтологии . 11 (3): 298–300. CiteSeerX 10.1.1.663.3809 . дои : 10.1093/geronj/11.3.298 . ПМИД 13332224 .
- ^ Харман Д. (апрель 1972 г.). «Биологические часы: митохондрии?». Журнал Американского гериатрического общества . 20 (4): 145–147. дои : 10.1111/j.1532-5415.1972.tb00787.x . ПМИД 5016631 . S2CID 396830 .
- ^ «Митохондрии и старение» . Circuitblue.co. Архивировано из оригинала 29 сентября 2017 года . Проверено 23 октября 2006 г.
- ^ Боффоли Д., Скакко С.К., Вергари Р., Соларино Г., Сантакроче Г., Папа С. (апрель 1994 г.). «Снижение с возрастом активности дыхательной цепи в скелетных мышцах человека». Biochimica et Biophysica Acta (BBA) - Молекулярные основы болезней . 1226 (1): 73–82. дои : 10.1016/0925-4439(94)90061-2 . ПМИД 8155742 .
- ^ де Грей AD (2004). «Митохондриальные мутации при старении млекопитающих: слишком поспешный поворот?». Исследования омоложения . 7 (3): 171–174. дои : 10.1089/rej.2004.7.171 . ПМИД 15588517 .
- ^ Бендер А., Кришнан К.Дж., Моррис К.М., Тейлор Г.А., Рив А.К., Перри Р.Х. и др. (май 2006 г.). «Высокие уровни делеций митохондриальной ДНК в нейронах черной субстанции при старении и болезни Паркинсона». Природная генетика . 38 (5): 515–517. дои : 10.1038/ng1769 . ПМИД 16604074 . S2CID 13956928 .
- ^ Мехта А.Р., Уолтерс Р., Уолдрон Ф.М., Пал С., Сельварадж Б.Т., Маклауд М.Р. и др. (август 2019 г.). «Нацеливание на митохондриальную дисфункцию при боковом амиотрофическом склерозе: систематический обзор и метаанализ» . Мозговые коммуникации . 1 (1): fcz009. doi : 10.1093/braincomms/fcz009 . ПМК 7056361 . ПМИД 32133457 . S2CID 201168692 .
- ^ Мехта А.Р., Грегори Дж.М., Дандо О., Картер Р.Н., Берр К., Нанда Дж. и др. (февраль 2021 г.). «Митохондриальный биоэнергетический дефицит в мотонейронах бокового амиотрофического склероза C9orf72 вызывает дисфункциональный аксональный гомеостаз» . Акта Нейропатологика . 141 (2): 257–279. дои : 10.1007/s00401-020-02252-5 . ПМЦ 7847443 . ПМИД 33398403 .
- ^ Чанг Дж.Л., Шукла П., Пагидас К., Ахмед Н.С., Карри С., Ганн Д.Д. и др. (ноябрь 2020 г.). «Митохондрии в старении яичников и репродуктивном долголетии» . Обзоры исследований старения . 63 : 101168. doi : 10.1016/J.arr.2020.101168 . ПМЦ 9375691 . ПМИД 32896666 . S2CID 221472721 .
- ^ Кёлликер А (1857). «Некоторые замечания об окончаниях кожных нервов и строении мышц» . Журнал научной зоологии (на немецком языке). 8 :311-325. Архивировано из оригинала 22 июня 2023 года . Проверено 22 июня 2023 г. На стр. 316, Кёлликер описал митохондрии, которые он наблюдал в свежих мышцах лягушки: «... очень бледные, округлые гранулы, которые появляются в виде длинных линейных линий [...], как только вы их заметили». (...[это] очень слабые круглые гранулы, которые встраиваются в сократительное вещество [мышцы] длинными линейными рядами. Эти гранулы расположены по всей толщине мышечного волокна, как на поверхности, так и внутри, и [их] так много, что они кажутся немаловажным элементом мышечных волокон, если к ним обратить внимание.) Кёлликер сообщил (стр. 321), что он обнаружил митохондрии в мышцах других животных. На рисунке 3 Таблица XIV. Архивировано 22 июня 2023 года в Wayback Machine . Кёлликер изобразил митохондрии в мышцах лягушки.
- ^ Перейти обратно: а б с д и ж г час я дж Эрнстер Л., Шац Г. (декабрь 1981 г.). «Митохондрии: исторический обзор» . Журнал клеточной биологии . 91 (3, часть 2): 227–255. дои : 10.1083/jcb.91.3.227s . ПМК 2112799 . ПМИД 7033239 .
- ^ Альтманн Р. (1890). ( на немецком языке Элементарные организмы и их связь с клетками ). Лейпциг, Германия: Veit & Co. 125. Архивировано из оригинала 23 июня 2023 года . Проверено 23 июня 2023 г. Из стр. 125: «Поскольку многие другие обстоятельства также указывают на то, что микроорганизмы и гранулы эквивалентны друг другу и представляют собой элементарные организмы, которые встречаются везде, где активируются жизненные силы, мы хотели бы называть их общим названием биобласты». (Поскольку в противном случае некоторые обстоятельства указывают на то, что микроорганизмы и гранулы эквивалентны друг другу и предполагают наличие элементарных организмов, которые можно встретить везде, где высвобождаются живые силы, мы обозначим их собирательным названием «биобласты».)
- ^ «митохондрии» . Интернет-словарь этимологии . Архивировано из оригинала 4 марта 2016 года . Проверено 23 мая 2013 г.
- ^ Бенда С (1898). «О сперматогенезе у позвоночных и высших беспозвоночных. Часть II: Гистиогенез сперматозоидов» [О сперматогенезе у позвоночных и высших беспозвоночных. Часть II: Гистогенез спермы.]. Архив физиологии (на немецком языке). 1898 : 393–398. Архивировано из оригинала 24 февраля 2019 года . Проверено 14 января 2018 г. Из стр. 397: После того, как Бренда заявляет, что «... я уже нашел [их (митохондрии)] во многих типах клеток всех возможных классов животных, ...), он предполагает: «Я хотел бы предварительно предположить, что они резервируют особое положение митохондрий, которое я обосноваю в дальнейшей работе». (Я хотел бы предложить временно зарезервировать для них, как «митохондрий», особый статус, который я обосноваю в дальнейшей работе.)
- Бенда С (1899). «Дальнейшие сообщения о митохондриях » . Архив физиологии: переговоры Берлинского физиологического общества (на немецком языке). 1899 : 376–383. Архивировано из оригинала 23 июня 2023 года . Проверено 23 июня 2023 г.
- ^ Михаэлис Л. (1900). «Жизненное пятно: метод визуализации клеточных гранул». Архив микроскопической анатомии и онтогенеза на ( немецком языке). 55 :558-575. дои : 10.1007/BF02977747 . S2CID 85010030 . Архивировано из оригинала 23 июня 2023 года . Проверено 23 июня 2023 г.
- ^ Цитата Эрнстера Мевес Ф (май 1908 г.). «Хондриосомы как носители наследственной предрасположенности. Цитологические исследования на курином эмбрионе» . Архив микроскопической анатомии . 72 (1): 816–867. дои : 10.1007/BF02982402 . S2CID 85047330 . Архивировано из оригинала 9 января 2020 года . Проверено 24 июня 2019 г. неверно, правильная цитата Мевес Ф (1904). «О возникновении митохондрий или хондромитов в растительных клетках». Бер. Немецкий. Бот . 22 :284-286. дои : 10.1111/j.1438-8677.1904.tb05237.x . S2CID 257822147 . , цитируется в статье Мевеса 1908 года и в Шмидт Э.В. (1913). «Пфланцлихе митохондрии» . Прогресс в ботанике 4 : 164–183 . Проверено 21 сентября 2012 г. , с подтверждением Nymphaea alba
- ^ Кингсбери Б.Ф. (1912). «Цитоплазматическая фиксация» . Анатомическая запись . 6 (2): 39–52. дои : 10.1002/ar.1090060202 . S2CID 221400422 . Архивировано из оригинала 23 июня 2023 года . Проверено 23 июня 2023 г. Из стр. 47: «...митохондрии являются их структурным выражением [т.е. химических восстановительных процессов в цитоплазме]...»
- ^ Варбург О (1913). «О дыхании гранул из клеток печени и о дыхании в беркефельдских фильтратах водных экстрактов печени » . Архив всей физиологии человека и животных Пфлюгера (на немецком языке). 154 : 599–617. Архивировано из оригинала 23 июня 2023 года . Проверено 23 июня 2023 г.
- ^ Милан Л., Триведи М., Сингх А., Талекар М., Амиджи М. (июнь 2015 г.). «Митохондриальная биология, мишени и доставка лекарств». Журнал контролируемого выпуска . 207 : 40–58. дои : 10.1016/j.jconrel.2015.03.036 . ПМИД 25841699 .
- ^ Мартин В.Ф., Гарг С., Зиморски В. (сентябрь 2015 г.). «Эндосимбиотические теории происхождения эукариот» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 370 (1678): 20140330. doi : 10.1098/rstb.2014.0330 . ПМЦ 4571569 . ПМИД 26323761 .
Общий
 В этой статье использованы общедоступные материалы из Научный букварь . НКБИ . Архивировано из оригинала 8 декабря 2009 года.
В этой статье использованы общедоступные материалы из Научный букварь . НКБИ . Архивировано из оригинала 8 декабря 2009 года.
Внешние ссылки
- Лейн Н (2016). Жизненно важный вопрос: энергия, эволюция и происхождение сложной жизни . WW Нортон и компания. ISBN 978-0393352979 .
- Питание клеточных митохондрий. Архивировано 17 августа 2022 года в Wayback Machine - XVIVO Scientific Animation.
- Mitodb.com. Архивировано 3 июля 2013 г. в Wayback Machine — базе данных митохондриальных заболеваний.
- Атлас митохондрий. Архивировано 29 июня 2012 г. в Wayback Machine в Университете Майнца.
- Портал исследований митохондрий на mitochondrial.net
- Митохондрии: архитектура определяет функцию на сайте Cytochemistry.net
- Митохондриальные связи в Университете Алабамы
- MIP. Архивировано 23 мая 2016 г. в Португальском веб-архиве Общества митохондриальной физиологии.
- 3D-структуры белков внутренней митохондриальной мембраны. Архивировано 13 сентября 2011 г. в Wayback Machine . Мичиганского университета
- 3D-структуры белков, связанных с внешней митохондриальной мембраной. Архивировано 13 сентября 2011 г. в Machine Wayback Мичиганского университета.
- Партнерство митохондриального белка в Университете Висконсина
- MitoMiner - база данных митохондриальной протеомики. [ постоянная мертвая ссылка ] в Отделении митохондриальной биологии MRC
- Митохондрии - клеточно-центрированная база данных. Архивировано 7 октября 2011 г. в Wayback Machine.
- Митохондрия, реконструированная с помощью электронной томографии. Архивировано 14 ноября 2012 года в Wayback Machine в Государственном университете Сан-Диего.
- Видеоклип митохондрий печени крысы из криоэлектронной томографии