ламаркизм

Ламаркизм , также известный как ламаркианское наследование или неоламаркизм , [ 2 ] Это представление о том, что организм может передавать своему потомству физические характеристики, которые родительский организм приобрел в результате использования или неиспользования в течение своей жизни. Его также называют наследованием приобретенных характеристик или, в последнее время, мягким наследованием . Идея названа в честь французского зоолога Жана-Батиста Ламарка (1744–1829), который включил теорию мягкой наследственности классической эпохи в свою теорию эволюции как дополнение к своей концепции ортогенеза , стремления к сложности .
Во вводных учебниках ламаркизм противопоставляется Чарльза Дарвина теории эволюции посредством естественного отбора . Однако книга Дарвина «Происхождение видов» подтвердила идею наследственных последствий использования и неиспользования, как это сделал Ламарк, а его собственная концепция пангенезиса аналогичным образом подразумевала мягкую наследственность. [ 2 ] [ 3 ]
Many researchers from the 1860s onwards attempted to find evidence for Lamarckian inheritance, but these have all been explained away,[4][5] either by other mechanisms such as genetic contamination or as fraud. August Weismann's experiment, considered definitive in its time, is now considered to have failed to disprove Lamarckism, as it did not address use and disuse. Later, Mendelian genetics supplanted the notion of inheritance of acquired traits, eventually leading to the development of the modern synthesis, and the general abandonment of Lamarckism in biology. Despite this, interest in Lamarckism has continued.
Since c. 2000 new experimental results in the fields of epigenetics, genetics, and somatic hypermutation proved the possibility of transgenerational epigenetic inheritance of traits acquired by the previous generation. These proved a limited validity of Lamarckism. The inheritance of the hologenome, consisting of the genomes of all an organism's symbiotic microbes as well as its own genome, is also somewhat Lamarckian in effect, though entirely Darwinian in its mechanisms.
Early history
[edit]Origins
[edit]
The inheritance of acquired characteristics was proposed in ancient times and remained a current idea for many centuries. The historian of science Conway Zirkle wrote in 1935 that:[6]
Lamarck was neither the first nor the most distinguished biologist to believe in the inheritance of acquired characters. He merely endorsed a belief which had been generally accepted for at least 2,200 years before his time and used it to explain how evolution could have taken place. The inheritance of acquired characters had been accepted previously by Hippocrates, Aristotle, Galen, Roger Bacon, Jerome Cardan, Levinus Lemnius, John Ray, Michael Adanson, Jo. Fried. Blumenbach and Erasmus Darwin among others.[6]
Zirkle noted that Hippocrates described pangenesis, the theory that what is inherited derives from the whole body of the parent, whereas Aristotle thought it impossible; but that all the same, Aristotle implicitly agreed to the inheritance of acquired characteristics, giving the example of the inheritance of a scar, or of blindness, though noting that children do not always resemble their parents. Zirkle recorded that Pliny the Elder thought much the same. Zirkle pointed out that stories involving the idea of inheritance of acquired characteristics appear numerous times in ancient mythology and the Bible and persisted through to Rudyard Kipling's Just So Stories.[7] The idea is mentioned in 18th century sources such as Diderot's D'Alembert's Dream.[8] Erasmus Darwin's Zoonomia (c. 1795) suggested that warm-blooded animals develop from "one living filament... with the power of acquiring new parts" in response to stimuli, with each round of "improvements" being inherited by successive generations.[9]
Darwin's pangenesis
[edit]
Charles Darwin's On the Origin of Species proposed natural selection as the main mechanism for development of species, but (like Lamarck) gave credence to the idea of heritable effects of use and disuse as a supplementary mechanism.[10] Darwin subsequently set out his concept of pangenesis in the final chapter of his book The Variation of Animals and Plants Under Domestication (1868), which gave numerous examples to demonstrate what he thought was the inheritance of acquired characteristics. Pangenesis, which he emphasised was a hypothesis, was based on the idea that somatic cells would, in response to environmental stimulation (use and disuse), throw off 'gemmules' or 'pangenes' which travelled around the body, though not necessarily in the bloodstream. These pangenes were microscopic particles that supposedly contained information about the characteristics of their parent cell, and Darwin believed that they eventually accumulated in the germ cells where they could pass on to the next generation the newly acquired characteristics of the parents.[11][12]
Darwin's half-cousin, Francis Galton, carried out experiments on rabbits, with Darwin's cooperation, in which he transfused the blood of one variety of rabbit into another variety in the expectation that its offspring would show some characteristics of the first. They did not, and Galton declared that he had disproved Darwin's hypothesis of pangenesis, but Darwin objected, in a letter to the scientific journal Nature, that he had done nothing of the sort, since he had never mentioned blood in his writings. He pointed out that he regarded pangenesis as occurring in protozoa and plants, which have no blood, as well as in animals.[13]
Lamarck's evolutionary framework
[edit]
Between 1800 and 1830, Lamarck proposed a systematic theoretical framework for understanding evolution. He saw evolution as comprising four laws:[14][15]
- "Life by its own force, tends to increase the volume of all organs which possess the force of life, and the force of life extends the dimensions of those parts up to an extent that those parts bring to themselves;"
- "The production of a new organ in an animal body, results from a new requirement arising. and which continues to make itself felt, and a new movement which that requirement gives birth to, and its upkeep/maintenance;"
- "The development of the organs, and their ability, are constantly a result of the use of those organs."
- "All that has been acquired, traced, or changed, in the physiology of individuals, during their life, is conserved through the genesis, reproduction, and transmitted to new individuals who are related to those who have undergone those changes."
Lamarck's discussion of heredity
[edit]In 1830, in an aside from his evolutionary framework, Lamarck briefly mentioned two traditional ideas in his discussion of heredity, in his day considered to be generally true. The first was the idea of use versus disuse; he theorized that individuals lose characteristics they do not require, or use, and develop characteristics that are useful. The second was to argue that the acquired traits were heritable. He gave as an imagined illustration the idea that when giraffes stretch their necks to reach leaves high in trees, they would strengthen and gradually lengthen their necks. These giraffes would then have offspring with slightly longer necks. In the same way, he argued, a blacksmith, through his work, strengthens the muscles in his arms, and thus his sons would have similar muscular development when they mature. Lamarck stated the following two laws:[1]
- Première Loi: Dans tout animal qui n' a point dépassé le terme de ses développemens, l' emploi plus fréquent et soutenu d' un organe quelconque, fortifie peu à peu cet organe, le développe, l' agrandit, et lui donne une puissance proportionnée à la durée de cet emploi; tandis que le défaut constant d' usage de tel organe, l'affoiblit insensiblement, le détériore, diminue progressivement ses facultés, et finit par le faire disparoître.[1]
- Deuxième Loi: Tout ce que la nature a fait acquérir ou perdre aux individus par l' influence des circonstances où leur race se trouve depuis long-temps exposée, et, par conséquent, par l' influence de l' emploi prédominant de tel organe, ou par celle d' un défaut constant d' usage de telle partie; elle le conserve par la génération aux nouveaux individus qui en proviennent, pourvu que les changemens acquis soient communs aux deux sexes, ou à ceux qui ont produit ces nouveaux individus.[1]
English translation:
- First Law [Use and Disuse]: In every animal which has not passed the limit of its development, a more frequent and continuous use of any organ gradually strengthens, develops and enlarges that organ, and gives it a power proportional to the length of time it has been so used; while the permanent disuse of any organ imperceptibly weakens and deteriorates it, and progressively diminishes its functional capacity, until it finally disappears.
- Second Law [Soft Inheritance]: All the acquisitions or losses wrought by nature on individuals, through the influence of the environment in which their race has long been placed, and hence through the influence of the predominant use or permanent disuse of any organ; all these are preserved by reproduction to the new individuals which arise, provided that the acquired modifications are common to both sexes, or at least to the individuals which produce the young.[16]
In essence, a change in the environment brings about change in "needs" (besoins), resulting in change in behaviour, causing change in organ usage and development, bringing change in form over time—and thus the gradual transmutation of the species. As the evolutionary biologists and historians of science Conway Zirkle, Michael Ghiselin, and Stephen Jay Gould have pointed out, these ideas were not original to Lamarck.[6][2][17]
Weismann's experiment
[edit]
August Weismann's germ plasm theory held that germline cells in the gonads contain information that passes from one generation to the next, unaffected by experience, and independent of the somatic (body) cells. This implied what came to be known as the Weismann barrier, as it would make Lamarckian inheritance from changes to the body difficult or impossible.[18]
Weismann conducted the experiment of removing the tails of 68 white mice, and those of their offspring over five generations, and reporting that no mice were born in consequence without a tail or even with a shorter tail. In 1889, he stated that "901 young were produced by five generations of artificially mutilated parents, and yet there was not a single example of a rudimentary tail or of any other abnormality in this organ."[19] The experiment, and the theory behind it, were thought at the time to be a refutation of Lamarckism.[18]
The experiment's effectiveness in refuting Lamarck's hypothesis is doubtful, as it did not address the use and disuse of characteristics in response to the environment. The biologist Peter Gauthier noted in 1990 that:[20]
Can Weismann's experiment be considered a case of disuse? Lamarck proposed that when an organ was not used, it slowly, and very gradually atrophied. In time, over the course of many generations, it would gradually disappear as it was inherited in its modified form in each successive generation. Cutting the tails off mice does not seem to meet the qualifications of disuse, but rather falls in a category of accidental misuse... Lamarck's hypothesis has never been proven experimentally and there is no known mechanism to support the idea that somatic change, however acquired, can in some way induce a change in the germplasm. On the other hand it is difficult to disprove Lamarck's idea experimentally, and it seems that Weismann's experiment fails to provide the evidence to deny the Lamarckian hypothesis, since it lacks a key factor, namely the willful exertion of the animal in overcoming environmental obstacles.[20]
Ghiselin also considered the Weismann tail-chopping experiment to have no bearing on the Lamarckian hypothesis, writing in 1994 that:[2]
The acquired characteristics that figured in Lamarck's thinking were changes that resulted from an individual's own drives and actions, not from the actions of external agents. Lamarck was not concerned with wounds, injuries or mutilations, and nothing that Lamarck had set forth was tested or "disproven" by the Weismann tail-chopping experiment.[2]
The historian of science Rasmus Winther stated that Weismann had nuanced views about the role of the environment on the germ plasm. Indeed, like Darwin, he consistently insisted that a variable environment was necessary to cause variation in the hereditary material.[21]
Textbook Lamarckism
[edit]
The identification of Lamarckism with the inheritance of acquired characteristics is regarded by evolutionary biologists including Ghiselin as a falsified artifact of the subsequent history of evolutionary thought, repeated in textbooks without analysis, and wrongly contrasted with a falsified picture of Darwin's thinking. Ghiselin notes that "Darwin accepted the inheritance of acquired characteristics, just as Lamarck did, and Darwin even thought that there was some experimental evidence to support it."[2] Gould wrote that in the late 19th century, evolutionists "re-read Lamarck, cast aside the guts of it ... and elevated one aspect of the mechanics—inheritance of acquired characters—to a central focus it never had for Lamarck himself."[22] He argued that "the restriction of 'Lamarckism' to this relatively small and non-distinctive corner of Lamarck's thought must be labelled as more than a misnomer, and truly a discredit to the memory of a man and his much more comprehensive system."[3][23]
Neo-Lamarckism
[edit]Context
[edit]
The period of the history of evolutionary thought between Darwin's death in the 1880s, and the foundation of population genetics in the 1920s and the beginnings of the modern evolutionary synthesis in the 1930s, is called the eclipse of Darwinism by some historians of science. During that time many scientists and philosophers accepted the reality of evolution but doubted whether natural selection was the main evolutionary mechanism.[24]
Among the most popular alternatives were theories involving the inheritance of characteristics acquired during an organism's lifetime. Scientists who felt that such Lamarckian mechanisms were the key to evolution were called neo-Lamarckians. They included the British botanist George Henslow (1835–1925), who studied the effects of environmental stress on the growth of plants, in the belief that such environmentally-induced variation might explain much of plant evolution, and the American entomologist Alpheus Spring Packard Jr., who studied blind animals living in caves and wrote a book in 1901 about Lamarck and his work.[25][26] Also included were paleontologists like Edward Drinker Cope and Alpheus Hyatt, who observed that the fossil record showed orderly, almost linear, patterns of development that they felt were better explained by Lamarckian mechanisms than by natural selection. Some people, including Cope and the Darwin critic Samuel Butler, felt that inheritance of acquired characteristics would let organisms shape their own evolution, since organisms that acquired new habits would change the use patterns of their organs, which would kick-start Lamarckian evolution. They considered this philosophically superior to Darwin's mechanism of random variation acted on by selective pressures. Lamarckism also appealed to those, like the philosopher Herbert Spencer and the German anatomist Ernst Haeckel, who saw evolution as an inherently progressive process.[25] The German zoologist Theodor Eimer combined Larmarckism with ideas about orthogenesis, the idea that evolution is directed towards a goal.[27]
With the development of the modern synthesis of the theory of evolution, and a lack of evidence for a mechanism for acquiring and passing on new characteristics, or even their heritability, Lamarckism largely fell from favour. Unlike neo-Darwinism, neo-Lamarckism is a loose grouping of largely heterodox theories and mechanisms that emerged after Lamarck's time, rather than a coherent body of theoretical work.[28]
19th century
[edit]
Neo-Lamarckian versions of evolution were widespread in the late 19th century. The idea that living things could to some degree choose the characteristics that would be inherited allowed them to be in charge of their own destiny as opposed to the Darwinian view, which placed them at the mercy of the environment. Such ideas were more popular than natural selection in the late 19th century as it made it possible for biological evolution to fit into a framework of a divine or naturally willed plan, thus the neo-Lamarckian view of evolution was often advocated by proponents of orthogenesis.[29] According to the historian of science Peter J. Bowler, writing in 2003:
One of the most emotionally compelling arguments used by the neo-Lamarckians of the late nineteenth century was the claim that Darwinism was a mechanistic theory which reduced living things to puppets driven by heredity. The selection theory made life into a game of Russian roulette, where life or death was predetermined by the genes one inherited. The individual could do nothing to mitigate bad heredity. Lamarckism, in contrast, allowed the individual to choose a new habit when faced with an environmental challenge and shape the whole future course of evolution.[30]
Scientists from the 1860s onwards conducted numerous experiments that purported to show Lamarckian inheritance. Some examples are described in the table.
| Scientist | Date | Experiment | Claimed result | Rebuttal |
|---|---|---|---|---|
| Charles-Édouard Brown-Séquard | 1869 to 1891 | Cut sciatic nerve and dorsal spinal cord of guinea pigs, causing abnormal nervous condition resembling epilepsy | Epileptic offspring | Not Lamarckism, as no use and disuse in response to environment; results could not be replicated; cause possibly a transmitted disease.[31][32][33][34][35][36] |
| Gaston Bonnier | 1884, 1886 | Transplant plants at different altitudes in Alps, Pyrenees | Acquired adaptations | Not controlled from weeds; likely cause genetic contamination[37] |
| Joseph Thomas Cunningham | 1891, 1893, 1895 | Shine light on underside of flatfish | Inherited production of pigment | Disputed cause[38][39][40][41][42][43] |
| Max Standfuss | 1892 to 1917 | Raise butterflies at low temperature | Variations in offspring even without low temperature | Richard Goldschmidt agreed; Ernst Mayr "difficult to interpret".[44][45][46][47] |
Early 20th century
[edit]
A century after Lamarck, scientists and philosophers continued to seek mechanisms and evidence for the inheritance of acquired characteristics. Experiments were sometimes reported as successful, but from the beginning these were either criticised on scientific grounds or shown to be fakes.[48][49][50][4][5] For instance, in 1906, the philosopher Eugenio Rignano argued for a version that he called "centro-epigenesis",[51][52][53][54][55][56] but it was rejected by most scientists.[57] Some of the experimental approaches are described in the table.
| Scientist | Date | Experiment | Claimed result | Rebuttal |
|---|---|---|---|---|
| William Lawrence Tower | 1907 to 1910 | Colorado potato beetles in extreme humidity, temperature | Heritable changes in size, colour | Criticised by William Bateson; Tower claimed all results lost in fire; William E. Castle visited laboratory, found fire suspicious, doubted claim that steam leak had killed all beetles, concluded faked data.[58][59][60][49][50] |
| Gustav Tornier | 1907 to 1918 | Goldfish, embryos of frogs, newts | Abnormalities inherited | Disputed; possibly an osmotic effect[61][62][63][64] |
| Charles Rupert Stockard | 1910 | Repeated alcohol intoxication of pregnant guinea pigs | Inherited malformations | Raymond Pearl unable to reproduce findings in chickens; Darwinian explanation[65][48] |
| Francis Bertody Sumner | 1921 | Reared mice at different temperatures, humidities | Inherited longer bodies, tails, hind feet | Inconsistent results[66][67] |
| Michael F. Guyer, Elizabeth A. Smith | 1918 to 1924 | Injected fowl serum antibodies for rabbit lens-protein into pregnant rabbits | Eye defects inherited for 8 generations | Disputed, results not replicated[68][69] |
| Paul Kammerer | 1920s | Midwife toad | Black foot-pads inherited | Fraud, ink injected; or, results misinterpreted; case celebrated by Arthur Koestler arguing that opposition was political[4][70] |
| William McDougall | 1920s | Rats solving mazes | Offspring learnt mazes quicker (20 vs 165 trials) | Poor experimental controls[71][72][73][74][75][76][5] |
| John William Heslop-Harrison | 1920s | Peppered moths exposed to soot | Inherited mutations caused by soot | Failure to replicate results; implausible mutation rate[77][78] |
| Ivan Pavlov | 1926 | Conditioned reflex in mice to food and bell | Offspring easier to condition | Pavlov retracted claim; results not replicable[79][80] |
| Coleman Griffith, John Detlefson | 1920 to 1925 | Reared rats on rotating table for 3 months | Inherited balance disorder | Results not replicable; likely cause ear infection[81][82][83][84][85][86] |
| Victor Jollos | 1930s | Heat treatment in Drosophila melanogaster | Directed mutagenesis, a form of orthogenesis | Results not replicable[87][88] |
Late 20th century
[edit]The British anthropologist Frederic Wood Jones and the South African paleontologist Robert Broom supported a neo-Lamarckian view of human evolution. The German anthropologist Hermann Klaatsch relied on a neo-Lamarckian model of evolution to try and explain the origin of bipedalism. Neo-Lamarckism remained influential in biology until the 1940s when the role of natural selection was reasserted in evolution as part of the modern evolutionary synthesis.[89] Herbert Graham Cannon, a British zoologist, defended Lamarckism in his 1959 book Lamarck and Modern Genetics.[90] In the 1960s, "biochemical Lamarckism" was advocated by the embryologist Paul Wintrebert.[91]
Neo-Lamarckism was dominant in French biology for more than a century. French scientists who supported neo-Lamarckism included Edmond Perrier (1844–1921), Alfred Giard (1846–1908), Gaston Bonnier (1853–1922) and Pierre-Paul Grassé (1895–1985). They followed two traditions, one mechanistic, one vitalistic after Henri Bergson's philosophy of evolution.[92]
In 1987, Ryuichi Matsuda coined the term "pan-environmentalism" for his evolutionary theory which he saw as a fusion of Darwinism with neo-Lamarckism. He held that heterochrony is a main mechanism for evolutionary change and that novelty in evolution can be generated by genetic assimilation.[93][94] His views were criticized by Arthur M. Shapiro for providing no solid evidence for his theory. Shapiro noted that "Matsuda himself accepts too much at face value and is prone to wish-fulfilling interpretation."[94]
Ideological neo-Lamarckism
[edit]
A form of Lamarckism was revived in the Soviet Union of the 1930s when Trofim Lysenko promoted the ideologically driven research programme, Lysenkoism; this suited the ideological opposition of Joseph Stalin to genetics. Lysenkoism influenced Soviet agricultural policy which in turn was later blamed for the numerous massive crop failures experienced within Soviet states.[95]
Critique
[edit]George Gaylord Simpson in his book Tempo and Mode in Evolution (1944) claimed that experiments in heredity have failed to corroborate any Lamarckian process.[96] Simpson noted that neo-Lamarckism "stresses a factor that Lamarck rejected: inheritance of direct effects of the environment" and neo-Lamarckism is closer to Darwin's pangenesis than Lamarck's views.[97] Simpson wrote, "the inheritance of acquired characters, failed to meet the tests of observation and has been almost universally discarded by biologists."[98]
Zirkle pointed out that Lamarck did not originate the hypothesis that acquired characteristics could be inherited, so it is incorrect to refer to it as Lamarckism:
Что на самом деле сделал Ламарк, так это принял гипотезу о том, что приобретенные признаки передаются по наследству, идею, которой придерживались почти повсеместно на протяжении более двух тысяч лет и которую его современники принимали как нечто само собой разумеющееся, и предположил, что результаты такого наследования были накапливается от поколения к поколению, создавая, таким образом, со временем новые виды. Его индивидуальный вклад в биологическую теорию заключался в применении к проблеме происхождения видов взгляда, согласно которому приобретенные признаки передаются по наследству, и в показе того, что эволюцию можно логически вывести из принятых биологических гипотез. Он, несомненно, был бы очень удивлен, узнав, что веру в наследование приобретенных признаков теперь называют «ламаркистской», хотя он почти наверняка почувствовал бы себя польщенным, если бы сама эволюция была обозначена так. [ 7 ]
Питер Медавар писал о ламаркизме: «Очень немногие профессиональные биологи верят, что что-то подобное происходит или может произойти, но это представление сохраняется по множеству ненаучных причин». Медавар заявил, что не существует известного механизма, с помощью которого адаптация, приобретенная в течение жизни человека, может быть отпечатана в геноме, и наследование по Ламарку недействительно, если оно не исключает возможность естественного отбора, но это не было продемонстрировано ни в одном эксперименте. [ 99 ]
Мартин Гарднер написал в своей книге «Причуды и заблуждения во имя науки» (1957):
Для проверки ламаркианства было проведено множество экспериментов. Все, что было проверено, оказалось отрицательным. С другой стороны, десятки тысяч экспериментов, о которых сообщалось в журналах и которые тщательно проверялись и перепроверялись генетиками всего мира, доказали правильность теории генных мутаций вне всякого разумного сомнения... Несмотря на быстро растущие доказательства Что касается естественного отбора, то Ламарк никогда не переставал иметь преданных последователей... Действительно, в мысли о том, что каждое небольшое усилие, прилагаемое животным, каким-то образом передается его потомству, действительно есть сильная эмоциональная привлекательность. [ 100 ]
По мнению Эрнста Майра, любая теория Ламарка, касающаяся наследования приобретенных признаков, была опровергнута, поскольку « ДНК не участвует непосредственно в формировании фенотипа, а фенотип, в свою очередь, не контролирует состав ДНК». [ 101 ] Питер Дж. Боулер писал, что, хотя многие ранние ученые серьезно относились к ламаркизму, в начале двадцатого века он был дискредитирован генетикой. [ 102 ]
Механизмы, напоминающие ламаркизм.
[ редактировать ]Исследования в области эпигенетики , генетики и соматических гипермутаций. [ 103 ] [ 104 ] подчеркнули возможное наследование черт, приобретенных предыдущим поколением. [ 105 ] [ 106 ] [ 107 ] [ 108 ] [ 109 ] Однако характеристика этих результатов как ламаркизма оспаривается. [ 110 ] [ 111 ] [ 112 ] [ 113 ]
Трансгенерационное эпигенетическое наследование
[ редактировать ]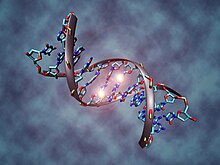
Эпигенетическое наследование , по мнению ученых, в том числе Евы Яблоньки и Мэрион Дж. Лэмб, является ламаркистским. [ 114 ] Эпигенетика основана на наследственных элементах, отличных от генов, которые передаются в половые клетки. К ним относятся паттерны метилирования в ДНК и хроматина метки на белках- гистонах , которые участвуют в регуляции генов . Эти метки реагируют на стимулы окружающей среды, по-разному влияют на экспрессию генов и являются адаптивными, причем фенотипические эффекты сохраняются в течение нескольких поколений. Этот механизм может также обеспечить наследование поведенческих черт, например, у кур. [ 115 ] [ 116 ] [ 117 ] крысы [ 118 ] [ 119 ] и в человеческих популяциях, переживших голодание, метилирование ДНК приводит к изменению функции генов как у голодающего населения, так и у их потомков. [ 120 ] Метилирование аналогичным образом опосредует эпигенетическую наследственность у таких растений, как рис. [ 121 ] [ 122 ] Небольшие молекулы РНК также могут опосредовать наследственную устойчивость к инфекции. [ 123 ] [ 124 ] [ 125 ] Гендель и Рамагопалан отметили, что «эпигенетика допускает мирное сосуществование дарвиновской и ламаркистской эволюции». [ 126 ]
Джозеф Спрингер и Деннис Холли в 2013 году прокомментировали это: [ 127 ]
Ламарк и его идеи были высмеяны и дискредитированы. По странной иронии судьбы Ламарк может смеяться последним. Эпигенетика, новая область генетики, показала, что Ламарк, возможно, был, по крайней мере частично, прав с самого начала. Кажется, что обратимые и наследственные изменения могут происходить без изменения последовательности ДНК (генотипа) и что такие изменения могут быть вызваны спонтанно или в ответ на факторы окружающей среды — «приобретенные черты» Ламарка. Определение того, какие наблюдаемые фенотипы наследуются генетически, а какие вызваны окружающей средой, остается важной и постоянной частью изучения генетики, биологии развития и медицины. [ 127 ]
Прокариотическая могут быть система CRISPR и взаимодействующая с Piwi РНК классифицированы как ламаркианские в рамках дарвиновской теории. [ 128 ] [ 129 ] Однако значение эпигенетики в эволюции неясно. Критики, такие как биолог-эволюционист Джерри Койн, отмечают, что эпигенетическая наследственность длится всего несколько поколений, поэтому она не является стабильной основой для эволюционных изменений. [ 130 ] [ 131 ] [ 132 ] [ 133 ]
Биолог-эволюционист Т. Райан Грегори утверждает, что эпигенетическое наследование не следует считать ламаркианским. По словам Грегори, Ламарк не утверждал, что окружающая среда напрямую влияет на живые существа. Вместо этого Ламарк «утверждал, что окружающая среда создала потребности, на которые организмы реагировали, используя одни функции больше, а другие меньше, что это приводило к тому, что эти функции усиливались или ослаблялись, и что это различие затем унаследовалось потомством». Грегори заявил, что ламаркистская эволюция в эпигенетике больше похожа на точку зрения Дарвина, чем на точку зрения Ламарка. [ 110 ]
В 2007 году Дэвид Хейг написал, что исследования эпигенетических процессов действительно допускают наличие ламаркистского элемента в эволюции, но эти процессы не бросают вызов основным принципам современного эволюционного синтеза, как утверждают современные ламаркисты. Хейг выступал за главенство ДНК и эволюцию эпигенетических переключателей в результате естественного отбора. [ 134 ] Хейг писал, что существует «внутреннее влечение» к эволюции Ламарка со стороны общественности и некоторых ученых, поскольку она наделяет мир смыслом, в котором организмы могут формировать свою собственную эволюционную судьбу. [ 135 ]
Томас Диккенс и Кази Рахман (2012) утверждали, что эпигенетические механизмы, такие как метилирование ДНК и модификация гистонов, генетически наследуются под контролем естественного отбора и не бросают вызов современному синтезу. Они оспаривают утверждения Яблонки и Лэмба об эпигенетических процессах Ламарка. [ 136 ]
В 2015 году Хуршид Икбал и его коллеги обнаружили, что, хотя «эндокринные разрушители оказывают прямое эпигенетическое воздействие на подвергшиеся воздействию зародышевые клетки плода, они корректируются путем перепрограммирования событий в следующем поколении». [ 138 ] Также в 2015 году Адам Вайс утверждал, что возвращение Ламарка в контексте эпигенетики вводит в заблуждение, комментируя: «Мы должны помнить [Ламарка] за то благо, которое он внес в науку, а не за вещи, которые лишь внешне напоминают его теорию. CRISPR и другие явления, подобные ламаркианскому, лишь скрывают простой и элегантный способ, которым на самом деле работает эволюция». [ 139 ]
Соматическая гипермутация и обратная транскрипция в зародышевую линию
[ редактировать ]В 1970-х годах австралийский иммунолог Эдвард Дж. Стил разработал неоламаркистскую теорию соматических гипермутаций в иммунной системе и связал ее с обратной транскрипцией РНК, полученной из клеток организма, в ДНК зародышевых клеток. Этот процесс обратной транскрипции предположительно позволял характеристикам или телесным изменениям, приобретенным в течение жизни, записываться обратно в ДНК и передаваться последующим поколениям. [ 140 ] [ 141 ]
Этот механизм должен был объяснить, почему гомологичные последовательности ДНК из областей гена VDJ родительских мышей обнаруживаются в их зародышевых клетках и, по-видимому, сохраняются в потомстве в течение нескольких поколений. Механизм включал соматическую селекцию и клональную амплификацию вновь приобретенных антител, последовательностей генов генерируемых посредством соматической гипермутации в В-клетках . Продукты информационной РНК этих соматически новых генов захватывались ретровирусами, эндогенными по отношению к В-клеткам, а затем транспортировались через кровоток, где они могли преодолевать барьер Вейсмана или сомато-зародышевый барьер и осуществлять обратную транскрипцию вновь приобретенных генов в зародышевые клетки. линия, наподобие пангенов Дарвина. [ 104 ] [ 103 ] [ 142 ]

Историк биологии Питер Дж. Боулер отметил в 1989 году, что другие ученые не смогли воспроизвести его результаты, и описал научный консенсус того времени: [ 137 ]
Не существует обратной связи между белками и ДНК и, следовательно, нет пути, по которому характеристики, приобретенные в организме, могут передаваться через гены. Работа Теда Стила (1979) вызвала шквал интереса к возможности того, что, в конце концов, могут существовать способы, которыми может иметь место этот обратный поток информации. ... [Его] механизм на самом деле не нарушал принципы молекулярной биологии, но большинство биологов с подозрением отнеслись к утверждениям Стила, и попытки воспроизвести его результаты не увенчались успехом. [ 137 ]
Боулер прокомментировал, что «работа [Стила] в то время подверглась резкой критике со стороны биологов, которые усомнились в результатах его экспериментов и отвергли его гипотетический механизм как неправдоподобный». [ 137 ]
Гологеномная теория эволюции
[ редактировать ]Хологеномная теория эволюции , хотя и дарвинистская, имеет ламаркистские аспекты. Отдельное животное или растение живет в симбиозе со многими микроорганизмами , и вместе они имеют «хологеном», состоящий из всех их геномов. Гологеном может изменяться, как и любой другой геном, в результате мутаций , половой рекомбинации и перестройки хромосом , но, кроме того, он может изменяться, когда популяции микроорганизмов увеличиваются или уменьшаются (напоминая использование и неупотребление Ламарка), а также когда в нем появляются новые виды микроорганизмов (напоминающие ламаркианскую теорию). наследование приобретенных признаков). Эти изменения затем передаются потомству. [ 144 ] Механизм в целом не вызывает сомнений, и естественный отбор иногда действительно происходит на уровне всей системы (хологенома), но неясно, всегда ли это так. [ 143 ]

Эффект Болдуина
[ редактировать ]Эффект Болдуина, названный в честь психолога Джеймса Марка Болдуина Джорджем Гейлордом Симпсоном в 1953 году, предполагает, что способность изучать новое поведение может улучшить репродуктивный успех животного и, следовательно, ход естественного отбора в его генетическом составе. Симпсон заявил, что этот механизм «не противоречит современному синтезу» эволюционной теории. [ 145 ] хотя он сомневался, что это происходило очень часто или что можно было доказать, что это происходит. Он отметил, что эффект Болдуина обеспечил примирение неодарвинистского и неоламаркистского подходов, то, что современный синтез , казалось, сделал ненужным. В частности, этот эффект позволяет животным адаптироваться к новому стрессу в окружающей среде посредством изменения поведения, за которым следуют генетические изменения. Это чем-то напоминает ламаркизм, но без требования к животным наследовать характеристики, приобретенные их родителями. [ 146 ] Эффект Болдуина широко принят дарвинистами. [ 147 ]
В социокультурной эволюции
[ редактировать ]В области культурной эволюции ламаркизм применялся как механизм теории двойного наследования . [ 148 ] Гулд рассматривал культуру как ламарковский процесс, посредством которого старшие поколения передают адаптивную информацию потомству посредством концепции обучения . В истории технологий компоненты ламаркизма использовались, чтобы связать культурное развитие с эволюцией человека, рассматривая технологию как продолжение человеческой анатомии. [ 149 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Ламарк 1830 , с. 235
- ^ Jump up to: а б с д и ж г Гизелин, Майкл Т. (1994). «Воображаемый Ламарк: взгляд на фальшивую «историю» в учебниках» . Письмо из учебника (сентябрь – октябрь 1994 г.). Архивировано из оригинала 12 октября 2000 года . Проверено 17 февраля 2006 г.
- ^ Jump up to: а б Гулд 2002 , стр. 177–178.
- ^ Jump up to: а б с Боулер 2003 , стр. 245–246.
- ^ Jump up to: а б с Медавар 1985 , с. 168
- ^ Jump up to: а б с Зиркль, Конвей (1935). «Наследование приобретенных признаков и предварительная гипотеза пангенезиса». Американский натуралист . 69 (724): 417–445. дои : 10.1086/280617 . S2CID 84729069 .
- ^ Jump up to: а б Зиркль, Конвей (январь 1946 г.). «Ранняя история идеи наследования приобретенных признаков и пангенезиса». Труды Американского философского общества . 35 (2): 91–151. дои : 10.2307/1005592 . JSTOR 1005592 .
- ^ Дидро, Дени; Баллестеро, Мануэль (1992). Мечта Даламбера; и Дополнение к «Путешествию на Бугенвиль» (Первое изд.). Мадрид: Дебаты. ISBN 84-7444-583-3 . OCLC 433436276 .
- ^ Дарвин 1794–1796 , Том I, раздел XXXIX
- ^ Десмонд и Мур 1991 , с. 617: «Но Дарвину не хотелось отказываться от идеи, что хорошо используемый и укрепленный орган может передаваться по наследству».
- ^ Дарвин, Чарльз (27 апреля 1871 г.). «Пангенезис» . Природа . 3 (78): 502–503. Бибкод : 1871Natur...3..502D . дои : 10.1038/003502a0 .
- ^ Холтерхофф, Кейт (2014). «История и восприятие гипотезы пангенезиса Чарльза Дарвина». Журнал истории биологии . 47 (4): 661–695. дои : 10.1007/s10739-014-9377-0 . ПМИД 24570302 . S2CID 207150548 .
- ^ Лю, Юншэн (2008). «Новый взгляд на пангенезис Дарвина». Биологические обзоры . 83 (2): 141–149. дои : 10.1111/j.1469-185x.2008.00036.x . ПМИД 18429766 . S2CID 39953275 .
- ^ Ларсон, Эдвард Дж. (2004). Растущее чувство прогресса . Современная библиотека. стр. 38–41.
{{cite book}}:|work=игнорируется ( помогите ) - ^ Гулд, Стивен (2001). Лежащие камни Марракеша: предпоследние размышления в естественной истории . Винтаж. стр. 119–121. ISBN 978-0-09-928583-0 .
- ^ Ламарк 1914 , с. 113
- ^ Гулд 2002 , стр. 170–191.
- ^ Jump up to: а б Романес, Джордж Джон (1893). Исследование вейсманизма . Открытый суд. ОЛ 23380098М .
- ^ Вейсманн 1889 , «Предполагаемая передача увечий» (1888), стр. 432
- ^ Jump up to: а б Готье, Питер (март – май 1990 г.). «Является ли эксперимент Вейсмана опровержением гипотезы Ламарка?». БИОС . 61 (1/2): 6–8. JSTOR 4608123 .
- ^ Винтер, Расмус (2001). «Август Вейсман о вариациях зародышевой плазмы». Журнал истории биологии . 34 (3): 517–555. дои : 10.1023/А:1012950826540 . ПМИД 11859887 . S2CID 23808208 .
- ^ Гулд 1980 , с. 66
- ^ Гулд, Стивен Джей (4 октября 1979 г.). «Другой взгляд на Ламарка» . Новый учёный . Том. 84, нет. 1175. стр. 38–40 . Проверено 9 ноября 2015 г. [ постоянная мертвая ссылка ]
- ^ Квамен 2006 , с. 216
- ^ Jump up to: а б Боулер 2003 , стр. 236–244.
- ^ Квамен 2006 , стр. 218, 220.
- ^ Квамен 2006 , с. 221
- ^ Боулер, Питер Дж. (1989) [1983]. Эволюция: история идеи (пересмотренная ред.). Издательство Калифорнийского университета. стр. 257, 264, 279–280 . ISBN 978-0520063860 .
- ^ Боулер 1992
- ^ Боулер 2003 , с. 367
- ^ Мамфорд 1921 , с. 209
- ^ Мейсон 1956 , с. 343
- ^ Буркхардт 1995 , с. 166
- ^ Райтьер 2012 , с. 299
- ^ Линвилл и Келли 1906 , с. 108
- ^ Аминофф 2011 , с. 192
- ^ Колер 2002 , с. 167
- ^ Каннингем, Джозеф Томас (1891). «Опыт отсутствия цвета на нижних сторонах камбал». Зоологический обзор . 14 : 27–32.
- ^ Каннингем, Джозеф Томас (май 1893 г.). «Исследования по окраске шкурок плоских рыб» . Журнал Морской биологической ассоциации Соединенного Королевства . 3 (1): 111–118. Бибкод : 1893JMBUK...3..111C . дои : 10.1017/S0025315400049596 . S2CID 84934811 .
- ^ Каннингем, Джозеф Томас (май 1895 г.). «Дополнительные данные о влиянии света на образование пигментов на нижних сторонах плоских рыб» (PDF) . Журнал Морской биологической ассоциации Соединенного Королевства . 4 (1): 53–59. Бибкод : 1895JMBUK...4...53C . дои : 10.1017/S0025315400050761 . S2CID 86159587 .
- ^ Мур, Элдон (15 сентября 1928 г.). «Новый взгляд на менделизм» . Зритель (Рецензия на книгу). Том. 141, нет. 5229. с. 337 . Проверено 24 октября 2015 г. Обзор современной биологии (1928) Дж. Т. Каннингема.
- ^ Кок и Форсдайк 2008 , стр. 132–133.
- ^ Морган 1903 , стр. view=1up, seq=277 257–259
- ^ Гольдшмидт 1940 , стр. 266–267.
- ^ Буркхардт 1998 , «Ламаркизм в Великобритании и Соединенных Штатах», с. 348
- ^ Forel 1934 , p. 36
- ^ Паккард, А.С. (10 июля 1896 г.). «Справочник по крупным бабочкам Палеарктики для исследователей и коллекционеров. Второе полностью переработанное издание, дополненное исследованиями по теории происхождения и т. д. » Наука (Рецензия на книгу). 4 (80): 52-54. дои : 10.1126/science.4.80.52-c . Обзор « Справочника по крупным палеарктическим бабочкам для исследователей и коллекционеров » (1896 г.) Максимилиана Рудольфа Стендфуса .
- ^ Jump up to: а б Делаж и Голдсмит 1912 , с. 210
- ^ Jump up to: а б Колер 2002 , стр. 202–204.
- ^ Jump up to: а б Митман 1992 , с. 219
- ^ Риньяно 1906 г.
- ^ Риньяно и Харви 1911 г.
- ^ Иствуд, М. Лайтфут (октябрь 1912 г.). «Рецензируемая работа: Эухенио Риньяно о наследовании приобретенных персонажей Ч. Харви». Международный журнал этики . 23 (1): 117–118. дои : 10.1086/206715 . JSTOR 2377122 .
- ^ Ньюман 1921 , с. 335
- ^ Риньяно 1926 г.
- ^ Кармайкл, Леонард (23 декабря 1926 г.). «Рецензируемая работа: Биологическая память Эухенио Риньяно, Э.В. Макбрайда». Журнал философии . 23 (26): 718–720. дои : 10.2307/2014451 . JSTOR 2014451 .
- ^ «(1) При наследовании приобретенных признаков (2) Биологические аспекты человеческих проблем». Природа (Рецензия на книгу). 89 (2232): 576–578. 8 августа 1912 г. Бибкод : 1912Natur..89..576. . дои : 10.1038/089576a0 . S2CID 3984855 .
- ^ Бейтсон, Уильям (3 июля 1919 г.). «Свидетельство доктора Каммерера о наследовании приобретенных характеров» . Природа (Письмо в редакцию). 103 (2592): 344–345. Бибкод : 1919Natur.103..344B . дои : 10.1038/103344b0 . S2CID 4146761 .
- ^ Бейтсон 1913 , стр. 219–227.
- ^ Вайнштейн 1998 , «Заметка о работе WL Tower в Лепинотарсе », стр. 352–353.
- ^ Макбрайд, Эрнест (январь 1924 г.). «Работа Торнье как возможное объяснение причин мутаций» . Евгеника Обзор . 15 (4): 545–555. ПМЦ 2942563 . ПМИД 21259774 .
- ^ Каннингем 1928 , стр. 84–97.
- ^ Сладден, Дороти Э. (май 1930 г.). «Экспериментальное искажение развития головастиков амфибий» . Труды Королевского общества Б. 106 (744): 318–325. дои : 10.1098/rspb.1930.0031 .
- ^ Сладден, Дороти Э. (ноябрь 1932 г.). «Экспериментальное искажение развития головастиков земноводных. Часть II» . Труды Королевского общества Б. 112 (774): 1–12. Бибкод : 1932РСПСБ.112....1С . дои : 10.1098/rspb.1932.0072 .
- ^ Блумберг 2010 , стр. 69–70.
- ^ Янг 1922 , с. 249
- ↑ Чайлд 1945 , стр. 146–173.
- ^ Гайер, Майкл Ф .; Смит, Э.А. (март 1920 г.). «Передача дефектов глаз, вызванных у кроликов с помощью сыворотки птицы, сенсибилизированной линзами» . ПНАС . 6 (3): 134–136. Бибкод : 1920PNAS....6..134G . дои : 10.1073/pnas.6.3.134 . ПМЦ 1084447 . ПМИД 16576477 .
- ^ Медавар 1985 , с. 169
- ^ Мур 2002 , с. 330
- ^ Макдугалл, Уильям (апрель 1938 г.). «Четвертый отчет об эксперименте Ламарка». Общий раздел. Британский журнал психологии . 28 (4): 365–395. дои : 10.1111/j.2044-8295.1938.tb00882.x .
- ^ Пантин, Карл Ф.А. (ноябрь 1957 г.). «Оскар Вернер Тигс. 1897–1956». Биографические мемуары членов Королевского общества . 3 : 247–255. дои : 10.1098/rsbm.1957.0017 . S2CID 84312439 .
- ^ Агар, Уилфред Э .; Драммонд, Фрэнк Х.; Тигс, Оскар В. (июль 1935 г.). «Первый отчет об испытаниях ламарковского эксперимента Макдугалла по дрессировке крыс» . Журнал экспериментальной биологии . 12 (3): 191–211. дои : 10.1242/jeb.12.3.191 .
- ^ Агар, Уилфред Э.; Драммонд, Фрэнк Х.; Тигс, Оскар В. (октябрь 1942 г.). «Второй отчет об испытании ламарковского эксперимента Макдугалла по дрессировке крыс» . Журнал экспериментальной биологии . 19 (2): 158–167. дои : 10.1242/jeb.19.2.158 .
- ^ Агар, Уилфред Э.; Драммонд, Фрэнк Х.; Тигс, Оскар В. (июнь 1948 г.). «Третий отчет об испытании ламарковского эксперимента Макдугалла по дрессировке крыс» . Журнал экспериментальной биологии . 25 (2): 103–122. дои : 10.1242/jeb.25.2.103 . Проверено 28 октября 2015 г.
- ^ Агар, Уилфред Э.; Драммонд, Фрэнк Х.; Тигс, Оскар В.; Гансон, Мэри М. (сентябрь 1954 г.). «Четвертый (окончательный) отчет об испытании ламарковского эксперимента Макдугалла по дрессировке крыс» . Журнал экспериментальной биологии . 31 (3): 308–321. дои : 10.1242/jeb.31.3.307 .
- ^ Хаген 2002 , с. 144: «В 1920-е годы энтомолог Дж. У. Хеслоп-Харрисон опубликовал экспериментальные данные, подтверждающие его утверждение о том, что химические вещества в саже вызывают широко распространенные мутации от светлокрылой формы к темнокрылой форме. Поскольку эти мутации предположительно передавались последующим поколениям, Харрисон утверждал, что он задокументировал случай наследования приобретенных признаков. Другие биологи не смогли повторить результаты Харрисона, а Р. А. Фишер отметил, что гипотеза Харрисона требует гораздо более высокой частоты мутаций, чем сообщалось ранее».
- ^ Мур и Декер 2008 , с. 203
- ^ Макдугалл 1934 , с. 180
- ^ Макдауэлл, Э. Карлтон; Викари, Эмилия М. (май 1921 г.). «Алкоголизм и поведение белых крыс. I. Влияние бабушек и дедушек-алкоголиков на поведение в лабиринте» . Журнал экспериментальной зоологии . 33 (1): 208–291. Бибкод : 1921JEZ....33..208M . дои : 10.1002/jez.1400330107 .
- ^ Гриффит, Коулман Р. (ноябрь – декабрь 1920 г.). «Влияние на белую крысу постоянного вращения тела». Американский натуралист . 54 (635): 524–534. дои : 10.1086/279783 . JSTOR 2456346 . S2CID 84453628 .
- ^ Гриффит, Коулман Р. (15 декабря 1922 г.). «Передаются ли по наследству постоянные нарушения равновесия?» . Наука . 56 (1459): 676–678. Бибкод : 1922Sci....56..676G . дои : 10.1126/science.56.1459.676 . ПМИД 17778266 .
- ^ Детлефсен, Джон А. (1923). «Наследуются ли последствия длительного вращения у крыс?». Труды Американского философского общества . 62 (5): 292–300. JSTOR 984462 .
- ^ Детлефсен, Джон А. (апрель 1925 г.). «Наследование приобретенных персонажей». Физиологические обзоры . 5 (2): 224–278. дои : 10.1152/physrev.1925.5.2.244 .
- ^ Доркус, Рой М. (июнь 1933 г.). «Влияние прерывистого вращения на ориентацию и привыкание к нистагму у крыс, а также некоторые наблюдения о влиянии пренатального вращения на постнатальное развитие». Журнал сравнительной психологии . 15 (3): 469–475. дои : 10.1037/h0074715 .
- ^ Мемориальный институт Отона С.А. Спрага, 1940 г. , с. 162
- ^ Йоллос, Виктор [на польском языке] (сентябрь 1934 г.). «Наследственные изменения, вызванные термообработкой у Drosophila melanogaster». Генетика . 16 (5–6): 476–494. дои : 10.1007/BF01984742 . S2CID 34126149 .
- ^ Харвуд 1993 , стр. 121–131.
- ^ Вуд 2013
- ^ Пушка 1975
- ^ Бозигер 1974 , с. 29
- ^ Луазон, Лоран (ноябрь 2011 г.). «Французские корни французских неоламаркизмов, 1879–1985». Журнал истории биологии . 44 (4): 713–744. дои : 10.1007/s10739-010-9240-x . ПМИД 20665089 . S2CID 3398698 .
- ^ Пирсон, Рой Дуглас (март 1988 г.). «Отзывы». Acta Biotheoretica (Рецензия на книгу). 37 (1): 31–36. дои : 10.1007/BF00050806 . Рецензии на книги « Эволюция животных в изменяющейся среде: с особым акцентом на аномальную метаморфозу» (1987) Рюичи Мацуда и «Эволюция индивидуальности» (1987) Лео В. Басса .
- ^ Jump up to: а б Шапиро, Артур М. (1988). «Рецензия на книгу: Эволюция животных в изменяющейся окружающей среде с особым упором на аномальный метаморфоз » (PDF) . Журнал Общества лепидоптерологов (Рецензия на книгу). 42 (2): 146–147 . Проверено 11 декабря 2015 г.
- ^ Бэрд, Скерри и Макинтайр 2006 , с. 166
- ^ Симпсон 1944 , с. 75
- ^ Симпсон 1964 , стр. 14–60.
- ^ Симпсон 1965 , с. 451
- ^ Медавар 1985 , стр. 166–169
- ^ Гарднер 1957 , стр. 142–143.
- ^ Майр 1997 , с. 222: «...признание того, что ДНК не участвует непосредственно в формировании фенотипа и что фенотип, в свою очередь, не контролирует состав ДНК, представляет собой окончательное обесценивание всех теорий, связанных с наследованием приобретенных признаков. Это окончательное опровержение теории эволюционной причинности Ламарка проясняет ситуацию».
- ^ Боулер 2013 , с. 21
- ^ Jump up to: а б Стил, Э.Дж. (2016). «Соматическая гипермутация при иммунитете и раке: критический анализ сигнатур мутаций, смещенных в цепи и контекста кодона». Восстановление ДНК . 45 (2016): 1–2 4. doi : 10.1016/j.dnarep.2016.07.001 . ПМИД 27449479 .
- ^ Jump up to: а б Стил, Э.Дж. (1981). Соматический отбор и адаптивная эволюция: о наследовании приобретенных признаков (2-е изд.). Издательство Чикагского университета.
- ^ Рот, Таня Л.; Любин, Фара Д.; Фанк, Адам Дж.; и др. (май 2009 г.). «Продолжительное эпигенетическое влияние невзгод раннего возраста на ген BDNF » . Биологическая психиатрия . 65 (9): 760–769. doi : 10.1016/j.biopsych.2008.11.028 . ПМК 3056389 . ПМИД 19150054 .
- ^ Арай, Джунко А.; Шаоминь Ли; Хартли, Дин М.; и др. (4 февраля 2009 г.). «Трансгенерационное спасение генетического дефекта в долговременной потенциации и формировании памяти путем ювенильного обогащения» . Журнал неврологии . 29 (5): 1496–1502. doi : 10.1523/JNEUROSCI.5057-08.2009 . ПМК 3408235 . ПМИД 19193896 .
- ^ Хакетт, Джейми А.; Сенгупта, Рупша; Зылич, Ян Дж.; и др. (25 января 2013 г.). «Динамика деметилирования зародышевой ДНК и стирание отпечатка посредством 5-гидроксиметилцитозина» . Наука . 339 (6118): 448–452. Бибкод : 2013Sci...339..448H . дои : 10.1126/science.1229277 . ПМЦ 3847602 . ПМИД 23223451 .
- ^ Бондурянский, Рассел (июнь 2012 г.). «Опять переосмысление наследственности». Тенденции в экологии и эволюции . 27 (6): 330–336. дои : 10.1016/j.tree.2012.02.003 . ПМИД 22445060 .
- ^ Скиннер, Майкл К. (май 2015 г.). «Эпигенетика окружающей среды и единая теория молекулярных аспектов эволюции: неоламаркистская концепция, способствующая неодарвинистской эволюции» . Геномная биология и эволюция . 7 (5): 1296–1302. дои : 10.1093/gbe/evv073 . ПМК 4453068 . ПМИД 25917417 .
- ^ Jump up to: а б Грегори, Т. Райан (8 марта 2009 г.). «Этого не говорил Ламарк, это говорил Дарвин» . Геномикрон (Блог). Архивировано из оригинала 9 февраля 2015 года . Проверено 4 ноября 2015 г.
- ^ Уилкинс 2009 , стр. 295–315.
- ^ Буркхардт, Ричард В. младший (август 2013 г.). «Ламарк, эволюция и наследование приобретенных признаков» . Генетика . 194 (4): 793–805. дои : 10.1534/genetics.113.151852 . ПМЦ 3730912 . ПМИД 23908372 .
- ^ Пенни, Дэвид (июнь 2015 г.). «Эпигенетика, Дарвин и Ламарк» . Геномная биология и эволюция . 7 (6): 1758–1760. дои : 10.1093/gbe/evv107 . ПМК 4494054 . ПМИД 26026157 .
- ^ Яблонка и Лэмб 1995
- ^ Мур 2015
- ^ Ричардс, Эрик Дж. (май 2006 г.). «Наследственная эпигенетическая изменчивость — новый взгляд на мягкое наследование». Обзоры природы Генетика . 7 (5): 395–401. дои : 10.1038/nrg1834 . ПМИД 16534512 . S2CID 21961242 .
- ^ Нэтт, Дэниел; Линдквист, Никлас; Страннехайм, Хенрик; и др. (28 июля 2009 г.). Пиццари, Том (ред.). «Наследование приобретенных адаптаций поведения и экспрессия генов мозга у кур» . ПЛОС ОДИН . 4 (7): е6405. Бибкод : 2009PLoSO...4.6405N . дои : 10.1371/journal.pone.0006405 . ПМЦ 2713434 . ПМИД 19636381 .
- ^ Шеу-Фанг Нг; Лин, Руби Сай; Лэйбатт, Д. Росс; и др. (21 октября 2010 г.). «Хроническая диета с высоким содержанием жиров у отцов программирует дисфункцию β-клеток у потомства самок крыс». Природа . 467 (7318): 963–966. Бибкод : 2010Natur.467..963N . дои : 10.1038/nature09491 . ПМИД 20962845 . S2CID 4308799 .
- ^ Гибсон, Андреа (16 июня 2013 г.). «Мыши-самцы, страдающие ожирением, дают потомство с более высоким уровнем жира в организме» (пресс-релиз). Университет Огайо . Архивировано из оригинала 12 июня 2018 г. Проверено 2 ноября 2015 г.
- ^ Люми, Ламберт Х.; Штейн, Арье Д.; Равелли, Анита CJ (июль 1995 г.). «Время пренатального голодания у женщин и масса тела при рождении их первого и второго ребенка: исследование когорты детей, рожденных в Голландии» . Европейский журнал акушерства, гинекологии и репродуктивной биологии . 61 (1): 23–30. дои : 10.1016/0028-2243(95)02149-М . ПМИД 8549843 . ИНИСТ 3596539 .
- ^ Акимото, Кейко; Катаками, Хацуэ; Хён-Юнг Ким; и др. (август 2007 г.). «Эпигенетическая наследственность у растений риса» . Анналы ботаники . 100 (2): 205–217. дои : 10.1093/aob/mcm110 . ПМЦ 2735323 . ПМИД 17576658 .
- ^ Сано, Хироши (апрель 2010 г.). «Наследование приобретенных признаков у растений: восстановление Ламарка» . Сигнализация и поведение растений . 5 (4): 346–348. дои : 10.4161/psb.5.4.10803 . ПМЦ 2958583 . ПМИД 20118668 .
- ^ Певица Эмили (4 февраля 2009 г.). «Возвращение ламарковской эволюции?» . MIT Technology Review (новости биомедицины). Архивировано из оригинала 27 января 2016 года . Проверено 3 ноября 2015 г.
- ^ Рехави, Одед; Миневич, Григорий; Хоберт, Оливер (9 декабря 2011 г.). «Трансгенерационное наследование приобретенного противовирусного ответа на основе малой РНК у C. Elegans » . Клетка . 147 (6): 1248–1256. дои : 10.1016/j.cell.2011.10.042 . ПМК 3250924 . ПМИД 22119442 .
- ^ Речави, О.; Хури-Зеэви, Л.; Анава, С.; Гох, WS; Керк, Ю.Ю.; Хэннон, Дж.Дж.; Хоберт, О. (17 июля 2014 г.). «Индуцированное голоданием трансгенерационное наследование малых РНК у C. elegans » . Клетка . 158 (2): 277–287. дои : 10.1016/j.cell.2014.06.020 . ПМЦ 4377509 . ПМИД 25018105 .
- ^ Гендель, Адам Э.; Рамагопалан, Шрирам В. (13 мая 2010 г.). «Имеет ли ламаркианская эволюция отношение к медицине?» . BMC Медицинская генетика . 11:73 . дои : 10.1186/1471-2350-11-73 . ПМК 2876149 . ПМИД 20465829 .
- ^ Jump up to: а б Спрингер и Холли, 2013 , с. 94
- ^ Кунин Евгений Владимирович ; Вольф Юрий Иванович (11 ноября 2009 г.). «Является ли эволюция дарвиновской или/и ламаркистской?» . Биология Директ . 4:42 . дои : 10.1186/1745-6150-4-42 . ПМЦ 2781790 . ПМИД 19906303 .
- ^ Кунин Евгений Владимирович (февраль 2019 г.). «CRISPR: новый принцип геномной инженерии, связанный с концептуальными сдвигами в эволюционной биологии» . Биология и философия . 34 (9): 9. дои : 10.1007/s10539-018-9658-7 . ПМК 6404382 . ПМИД 30930513 .
- ^ Койн, Джерри (24 октября 2010 г.). «Эпигенетика: свет и путь?» . Почему эволюция верна (блог) . Проверено 4 ноября 2015 г.
- ^ Койн, Джерри (23 сентября 2013 г.). «Эпигенетическая атака на Guardian» . Почему эволюция верна (блог) . Проверено 4 ноября 2015 г.
- ^ Гонсалес-Ресио, О.; Торо, Массачусетс; Бах, А. (2015). «Прошлое, настоящее и будущее эпигенетики в применении к животноводству» . Границы генетики . 6 : 305. дои : 10.3389/fgene.2015.00305 . ПМЦ 4585102 . ПМИД 26442117 .
- ^ Варона, Луи; Мунилла, Себастьян; Муресан, Елена Флавия; Гонсалес-Родригес, Альдемар; Морено, Чарльз; Алтарь, Джон (2015). «Байесовская модель для анализа трансгенерационных эпигенетических вариаций» . G3: Гены, геномы, генетика . 5 (4): 477–485. дои : 10.1534/g3.115.016725 . ПМЦ 4390564 . ПМИД 25617408 .
- ^ Хейг, Дэвид (июнь 2007 г.). «Правила Вейсмана! Хорошо? Эпигенетика и искушение Ламарка». Биология и философия . 22 (3): 415–428. дои : 10.1007/s10539-006-9033-y . S2CID 16322990 .
Современные неодарвинисты не отрицают, что эпигенетические механизмы играют важную роль в процессе развития, и не отрицают, что эти механизмы обеспечивают разнообразные адаптивные реакции на окружающую среду. Рекуррентные, предсказуемые изменения эпигенетического состояния обеспечивают полезный набор переключателей, которые позволяют генетически идентичным клеткам приобретать дифференцированные функции и позволяют факультативно реагировать генотипа на изменения окружающей среды (при условии, что «подобные» изменения неоднократно происходили в прошлом). Однако большинство неодарвинистов утверждают, что способность адаптивно переключать эпигенетическое состояние является свойством последовательности ДНК (в том смысле, что альтернативные последовательности будут демонстрировать различное поведение при переключении) и что любое увеличение адаптивности в системе происходит за счет процесс естественного отбора.
- ^ Хейг, Дэвид (ноябрь 2011 г.). «Восхождение Ламарка!» . Философия и теория в биологии (Книга-эссе). 3 (е204). дои : 10.3998/ptb.6959004.0003.004 . «Обзор трансформаций ламаркизма: от тонких жидкостей к молекулярной биологии , под редакцией Снайта Б. Гиссиса и Евы Яблонки, MIT Press, 2011»
- ^ Дикинс, Томас Э.; Рахман, Кази (7 августа 2012 г.). «Расширенный эволюционный синтез и роль мягкого наследования в эволюции» . Труды Королевского общества Б. 279 (1740): 2913–2921. дои : 10.1098/rspb.2012.0273 . ПМЦ 3385474 . ПМИД 22593110 .
- ^ Jump up to: а б с д Боулер, Питер Дж. (1989) [1983]. Эволюция: история идеи (пересмотренная ред.). Издательство Калифорнийского университета. стр. 179, 341 . ISBN 978-0520063860 .
- ^ Уайтлоу, Эмма (27 марта 2015 г.). «Спор о ламарковском эпигенетическом наследовании у млекопитающих» . Геномная биология . 16 (60): 60. дои : 10.1186/s13059-015-0626-0 . ПМЦ 4375926 . ПМИД 25853737 .
- ^ Вайс, Адам (октябрь 2015 г.). «Ламарковские иллюзии» . Тенденции в экологии и эволюции . 30 (10): 566–568. дои : 10.1016/j.tree.2015.08.003 . ПМИД 26411613 .
- ^ Стил, Э.Дж. (2016). «Соматическая гипермутация при иммунитете и раке: критический анализ сигнатур мутаций, смещенных в цепи и контекста кодонов». Восстановление ДНК . 45 : 1–24. дои : 10.1016/j.dnarep.2016.07.001 . ПМИД 27449479 .
- ^ Стил, Э.Дж.; Поллард, JW (1987). «Гипотеза: соматическая гипермутация путем преобразования генов посредством подверженной ошибкам информационной петли ДНК-РНК-ДНК». Молекулярная иммунология . 24 (6): 667–673. дои : 10.1016/j.dnarep.2016.07.001 . ПМИД 2443841 .
- ^ Стил, Линдли и Бланден, 1998 г.
- ^ Jump up to: а б Моран, Нэнси А .; Слоан, Дэниел Б. (04 декабря 2015 г.). «Концепция гологенома: полезная или пустая?» . ПЛОС Биология . 13 (12): e1002311. дои : 10.1371/journal.pbio.1002311 . ПМК 4670207 . ПМИД 26636661 .
- ^ Розенберг, Юджин ; Шэрон, Гилл; Зильбер-Розенберг, Илана (декабрь 2009 г.). «Гологеномная теория эволюции содержит ламарковские аспекты в рамках дарвинизма». Экологическая микробиология . 11 (12): 2959–2962. Бибкод : 2009EnvMi..11.2959R . дои : 10.1111/j.1462-2920.2009.01995.x . ПМИД 19573132 .
- ^ Депью, Дэвид Дж. (2003), «Болельщики Болдуина, скептики Болдуина» в: Вебер, Брюс Х.; Депью, Дэвид Дж. (2003). Эволюция и обучение: новый взгляд на эффект Болдуина . МТИ Пресс. стр. 3–31. ISBN 978-0-262-23229-6 .
- ^ Симпсон, Джордж Гейлорд (1953). «Эффект Болдуина». Эволюция . 7 (2): 110–117. дои : 10.2307/2405746 . JSTOR 2405746 .
- ^ Деннетт, Дэниел (2003), «Эффект Болдуина, кран, а не Skyhook» в: Вебер, Брюс Х.; Депью, Дэвид Дж. (2003). Эволюция и обучение: новый взгляд на эффект Болдуина . МТИ Пресс. стр. 69–106. ISBN 978-0-262-23229-6 .
- ^ Кронфельднер, Мария (13 декабря 2005 г.). «Является ли культурная эволюция ламаркистской?». Биология и философия . 22 (4): 493–512. дои : 10.1007/s10539-006-9037-7 . S2CID 85411375 .
- ^ Каллен 2000 , стр. 31–60
Библиография
[ редактировать ]- Аминофф, Майкл Дж. (2011). Браун-Секар: невероятный гений, изменивший медицину . Издательство Оксфордского университета . ISBN 978-0-19-974263-9 . LCCN 2010013439 . ОСЛК 680002156 .
- Бэрд, Дэвис; Шерри, Эрик ; Макинтайр, Ли , ред. (2006). Философия химии: синтез новой дисциплины . Бостонские исследования в области философии науки. Том. 242. Спрингер . ISBN 978-1-4020-3256-1 . LCCN 2006295950 . OCLC 209927684 .
- Бейтсон, Уильям (1913). Проблемы генетики . Издательство Йельского университета . LCCN 13021769 . OCLC 809326988 . Проблемы генетики (1913 г.) в Интернет-архиве.
- Блумберг, Марк С. (2010). Причуды природы: и что они говорят нам об эволюции и развитии (изд. В мягкой обложке). Издательство Оксфордского университета. ISBN 978-0-19-921306-1 . LCCN 2010481198 . OCLC 352916350 .
- Бозигер, Эрнест (1974). «Эволюционные теории после Ламарка и Дарвина». В Аяле, Франсиско Хосе ; Добжанский, Феодосий (ред.). Исследования по философии биологии: редукция и связанные с ней проблемы . Издательство Калифорнийского университета . ISBN 978-0-520-02649-0 . LCCN 73090656 . OCLC 1265669 .
- Боулер, Питер Дж. (1992) [Оригинальное издание в твердом переплете, опубликованное в 1983 году]. Затмение дарвинизма: антидарвиновские теории эволюции за десятилетия около 1900 года (изд. Джона Хопкинса в мягкой обложке). Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-4391-4 . LCCN 82021170 . OCLC 611262030 .
- Боулер, Питер Дж. (2003). Эволюция: история идеи (3-е изд.). Издательство Калифорнийского университета. ISBN 978-0-520-23693-6 . LCCN 2002007569 . OCLC 49824702 .
- Боулер, Питер Дж. (2013). Дарвин удален: представляя мир без Дарвина . Издательство Чикагского университета . ISBN 978-0-226-06867-1 . LCCN 2012033769 . OCLC 808010092 .
- Буркхардт, Ричард В. младший (1995) [первоначально опубликовано в 1977 году]. Дух системы: Ламарк и эволюционная биология: теперь с «Ламарком в 1995 году» (изд. First Harvard University Press в мягкой обложке). Издательство Гарвардского университета . ISBN 978-0-674-83318-0 . LCCN 95010861 . OCLC 32396741 .
- Кэннон, Герберт Грэм (1975) [Первоначально опубликовано в 1959 году: Манчестер, Англия; Издательство Манчестерского университета ]. Ламарк и современная генетика (переиздание). Гринвуд Пресс . ISBN 978-0-8371-8173-8 . LCCN 75010211 . OCLC 1418716 .
- Чайлд, Чарльз Мэннинг (1945). Биографические мемуары Фрэнсиса Бертоди Самнера, 1874–1945 (PDF) . Национальная академия наук, Вашингтон, округ Колумбия. Биографические мемуары. Том. 25. Национальная академия наук . LCCN 52004656 . ОСЛК 11852074 . Проверено 27 октября 2015 г.
- Кок, Алан Г.; Форсдайк, Дональд Р. (2008). Цените свои исключения: наука и жизнь Уильяма Бейтсона . Спрингер. ISBN 978-0-387-75687-5 . LCCN 2008931291 . OCLC 344846261 .
- Каллен, Бен Сэндфорд (2000). Стил, Джеймс; Каллен, Ричард; Чиппиндейл, Кристофер (ред.). Заразительные идеи: об эволюции, культуре, археологии и теории культурного вируса . Книги Оксбоу . ISBN 978-1-84217-014-4 . OCLC 47122736 .
- Каннингем, Джозеф Томас (1928). Современная биология: обзор основных явлений животной жизни в связи с современными концепциями и теориями . Кеган Пол, Trench, Trübner & Co., Ltd. LCCN 29000027 . OCLC 920415 .
- Дарвин, Эразм (1794–1796). Зоономия; или Законы органической жизни . Джозеф Джонсон. LCCN 34036671 . OCLC 670735211 .
- Делаж, Ив ; Голдсмит, Мари [на французском языке] (1912). Теории эволюции . Перевод Андре Тридона. Б. В. Хюбш . LCCN 12031796 . OCLC 522024 . Теории эволюции (1912 г.) в Интернет-архиве.
- Десмонд, Адриан ; Мур, Джеймс Р. (1991). Дарвин . Майкл Джозеф ; Викинг Пингвин . ISBN 978-0-7181-3430-3 . LCCN 92196964 . ОСЛК 26502431 .
- Форель, Огюст (1934) [1922]. Сексуальный вопрос: научное, психологическое, гигиеническое и социологическое исследование . Английская адаптация второго немецкого издания, исправленная и дополненная К. Ф. Маршаллом (пересмотренная редакция). Книжная компания врачей и хирургов. LCCN 22016399 . OCLC 29326677 . Сексуальный вопрос: научное, психологическое, гигиеническое и социологическое исследование (1922 г.) в Интернет-архиве . Проверено 26 октября 2015 г.
- Гарднер, Мартин (1957) [1952]. Причуды и заблуждения во имя науки . Дуврские публикации . LCCN 57003844 . ОСЛК 233892 .
- Гольдшмидт, Ричард (1940). Материальная основа эволюции . Лекции памяти г-жи Хепса Эли Силлиман. Издательство Йельского университета; Издательство Оксфордского университета. LCCN 40012233 . OCLC 595767401 .
- Гулд, Стивен Джей (1980). Большой палец панды: больше размышлений о естествознании (1-е изд.). WW Нортон и компания . ISBN 978-0-393-01380-1 . LCCN 80015952 . OCLC 6331415 .
- Гулд, Стивен Джей (2002). Структура эволюционной теории . Belknap Press издательства Гарвардского университета . ISBN 978-0-674-00613-3 . LCCN 2001043556 . OCLC 47869352 .
- Хаген, Джоэл Б. (2002). «Эксперименты по пересказу: исследования промышленного меланизма у перцовой моли, проведенные HBD Kettlewell». В Гилтроу, Джанет (ред.). Академическое чтение: чтение и письмо по различным дисциплинам (2-е изд.). Бродвью Пресс . ISBN 978-1-55111-393-7 . LCCN 2002514564 . OCLC 46626903 .
- Харвуд, Джонатан (1993). Стили научной мысли: Немецкое генетическое сообщество, 1900–1933 гг . Наука и ее концептуальные основы. Издательство Чикагского университета. ISBN 978-0-226-31882-0 . LCCN 92015321 . OCLC 25746714 .
- Яблонка, Ева ; Лэмб, Мэрион Дж. (1995). Эпигенетическая наследственность и эволюция: ламаркианское измерение . Издательство Оксфордского университета. ISBN 978-0-19-854062-5 . LCCN 94032108 . OCLC 30974876 .
- Колер, Роберт Э. (2002). Пейзажи и лабораторные ландшафты: исследование границы лаборатории и поля в биологии . Издательство Чикагского университета. ISBN 978-0-226-45010-0 . LCCN 2002023331 . OCLC 690162738 .
- Ламарк, Жан-Батист (1830) [Первоначально опубликовано в 1809 году; Париж: Dentu и L'Auteur]. Зоологическая философия (на французском языке) (Новое изд.). Спраут Байьер. LCCN 11003671 . Зоологическая философия (1830 г.) в Интернет-архиве
- —— (1914). Зоологическая философия; Экспозиция по естественной истории животных . Перевод с введением Хью Эллиота. Macmillan and Co., Ltd. LCCN a15000196 . OCLC 1489850 . Зоологическая философия (1914) в Интернет-архиве.
- Линвилл, Генри Р.; Келли, Генри А. (1906). Учебник по общей зоологии . Джинн и компания . LCCN 06023318 . ОСЛК 1041858 . Учебник по общей зоологии (1906 г.) в Интернет-архиве.
- Мейсон, Стивен Финни (1956). Основные течения научной мысли: история наук . Жизнь научной библиотеки. Том. 32 (Переиздание). Абеляр-Шуман. OCLC 732176237 .
- Майр, Эрнст (1997) [первоначально опубликовано в 1976 году]. Эволюция и разнообразие жизни: избранные эссе (изд. First Harvard University Press в мягкой обложке). Belknap Press издательства Гарвардского университета. ISBN 978-0-674-27105-0 . LCCN 75042131 . OCLC 247687824 .
- Майр, Эрнст; Провин, Уильям Б. , ред. (1998). Эволюционный синтез: перспективы унификации биологии . Новое предисловие Эрнста Майра. Издательство Гарвардского университета. ISBN 978-0-674-27226-2 . LCCN 98157613 . OCLC 503188713 .
- Макдугалл, Уильям (1934). Современный материализм и эмерджентная эволюция . Метуэн.
- Медавар, Питер (1985) [первоначально опубликовано в 1983 году]. От Аристотеля до зоопарков: Философский словарь биологии . Оксфордские книги в мягкой обложке (переиздание). Издательство Гарвардского университета. ISBN 978-0-19-283043-2 . LCCN 84016529 . ОСЛК 11030267 .
- Митман, Грегг (1992). Состояние природы: экология, сообщество и американская социальная мысль, 1900–1950 гг . Наука и ее концептуальные основы. Издательство Чикагского университета. ISBN 978-0-226-53236-3 . LCCN 91045638 . ОСЛК 25130594 .
- Мур, Дэвид С. (2015). Развивающийся геном: введение в поведенческую эпигенетику . Издательство Оксфордского университета. ISBN 978-0-19-992234-5 . LCCN 2014049505 . OCLC 894139943 .
- Мур, Рэнди; Декер, Марк Д. (2008). Больше, чем Дарвин: Энциклопедия людей и мест спора между эволюцией и креационизмом . Гринвуд Пресс. ISBN 978-0-313-34155-7 . LCCN 2007044406 . OCLC 177023758 .
- Мур, Джеймс Р., изд. (2002) [Первоначально опубликовано в 1989 году]. История, человечество и эволюция: очерки Джона К. Грина . Издательство Кембриджского университета . ISBN 978-0-521-52478-0 . LCCN 89032583 . OCLC 49784849 .
- Морган, Томас Хант (1903). Эволюция и адаптация . Компания Макмиллан; Macmillan and Co., Ltd. LCCN 03027216 . OCLC 758217701 . Эволюция и адаптация (1903 г.) в Интернет-архиве.
- Мамфорд, Фредерик Блэкмар (1921) [первоначально опубликовано в 1917 году]. Разведение животных . Серия сельских учебников. Компания Макмиллан . LCCN 17007834 . OCLC 5429719 . Разведение животных (1921 г.) в Интернет-архиве.
- Ньюман, Горацио Хакетт (1921). Чтения по эволюции, генетике и евгенике . Издательство Чикагского университета. ЛЦН 21017204 . OCLC 606993 . Чтения по эволюции, генетике и евгенике (1921 г.) в Интернет-архиве.
- Мемориальный институт Отона С.А. Спрага (1940). Исследования Мемориального института Отона С.А. Спрага: Сборник репринтов . Том. 25. Мемориальный институт Отона С.А. Спрага. OCLC 605547177 .
- Кваммен, Дэвид (2006). Неохотный мистер Дарвин: личный портрет Чарльза Дарвина и создание его теории эволюции . Великие открытия (1-е изд.). Атлас Книги/Нортон. ISBN 978-0-393-05981-6 . LCCN 2006009864 . ОСЛК 65400177 .
- Рэтьер, Мартин Н. (2012). Соучастие друзей: как Джордж Элиот, Г. Х. Льюис и Джон Хьюлингс-Джексон закодировали секрет Герберта Спенсера . Издательство Бакнеллского университета . ISBN 978-1-61148-418-2 . LCCN 2012030762 . OCLC 806981125 .
- Риньяно, Эудженио (1906). О передаче приобретенных признаков: гипотеза центроэпигенеза (на французском языке). Феликс Алькан. OCLC 5967582 .
- ——; Харви, Бэзил CH (1911). Эудженио Риньяно о наследовании приобретенных признаков: гипотеза наследственности, развития и ассимиляции . Авторизованный английский перевод Бэзила Ч. Харви. Издательская компания «Открытый суд» . LCCN 11026509 . ОСЛК 1311084 . Эухенио Риньяно о наследовании приобретенных персонажей (1911) в Интернет-архиве
- Риньяно, Эудженио (1926). Биологическая память . Международная библиотека психологии, философии и научного метода. Перевод с предисловием Эрнеста Макбрайда . Кеган Пол, Trench, Trübner & Co.; Харкорт, Брейс и компания . LCCN 26009586 . OCLC 811731 .
- Симпсон, Джордж Гейлорд (1944). «Темп и режим в эволюции» . Труды Нью-Йоркской академии наук . Колумбийская биологическая серия. 15 . Издательство Колумбийского университета : 45–60. дои : 10.1111/j.2164-0947.1945.tb00215.x . LCCN а45000404 . OCLC 993515 . ПМИД 21012247 .
- Симпсон, Джордж Гейлорд (1964). Этот взгляд на жизнь: мир эволюциониста (1-е изд.). Харкорт, Брейс и мир . LCCN 64014636 . ОСЛК 230986 .
- Симпсон, Джордж Гейлорд (1965). Жизнь: Введение в биологию (2-е изд.). Харкорт, Брейс и мир. LCCN 65014384 . ОСЛК 165951 .
- Спрингер, Джозеф Т.; Холли, Деннис (2013). Введение в зоологию (1-е изд.). Джонс и Бартлетт Обучение . ISBN 978-1-4496-4891-6 . LCCN 2011022399 . OCLC 646112356 .
- Стил, Эдвард Дж .; Линдли, Робин А.; Бланден, Роберт В. (1998). Подпись Ламарка: Как ретрогены меняют дарвиновскую парадигму естественного отбора . Книги Спирали; Границы науки. Книги Персея . ISBN 978-0-7382-0014-9 . LCCN 98087900 . OCLC 40449772 .
- Вейсман, август (1889 г.). Поултон, Эдвард Б .; Шёнланд, Зельмар ; Шипли, Артур Э. (ред.). Очерки наследственности и родственных биологических проблем . Кларендон Пресс . LCCN 77010494 . OCLC 488543825 . Проверено 30 октября 2015 г.
- Уилкинс, Джон С. (2009) [Первоначально опубликовано в 2001 году в журнале Laurent, John; Найтингейл, Джон (редакторы), Дарвинизм и эволюционная экономика , глава 8, стр. 160–183; Челтнем, Великобритания: Эдвард Элгар]. «Появление ламаркизма в эволюции культуры». В Ходжсоне, Джеффри М. (ред.). Дарвинизм и экономика . Международная библиотека критических статей по экономике. Том. 233. Эдвард Элгар. ISBN 978-1-84844-072-2 . LCCN 2008939772 . OCLC 271774708 .
- Вуд, Бернард, изд. (2013). Энциклопедия эволюции человека Уайли-Блэквелла (первое издание в мягкой обложке). Уайли-Блэквелл . ISBN 978-1-118-65099-8 . LCCN 2013012756 . OCLC 841039552 .
- Янг, Роберт Томпсон (1922). Биология в Америке . РГ Барсук. LCCN 22019903 . ОСЛК 370597 . Биология в Америке (1922) в Интернет-архиве
Дальнейшее чтение
[ редактировать ]- Бартелеми-Мадол, Мадлен (1982). Ламарк, мифический предшественник: исследование отношений между наукой и идеологией . Английский перевод М. Х. Шэнка. МТИ Пресс . ISBN 978-0-262-02179-1 . LCCN 82010061 . ОСЛК 8533097 . Перевод Ламарка, или Миф о предшественнике (1979)
- Боулер, Питер Дж. (1989). Менделевская революция: появление наследственных концепций в современной науке и обществе . Издательство Университета Джонса Хопкинса . ISBN 978-0-8018-3888-0 . LCCN 89030914 . OCLC 19322402 .
- Беркман, Оливер (19 марта 2010 г.). «Почему все, что вам говорили об эволюции, неверно» . Хранитель .
- Резерфорд, Адам (19 марта 2010 г.). «Помимо заголовка «Дарвин был не прав»» . Хранитель .
- Кук, Джордж М. (декабрь 1999 г.). «Неоламаркский экспериментализм в Америке: истоки и последствия». Ежеквартальный обзор биологии . 74 (4): 417–437. дои : 10.1086/394112 . JSTOR 2664721 . ПМИД 10672643 . S2CID 12954177 .
- Десмонд, Адриан (1989). Политика эволюции: морфология, медицина и реформа в радикальном Лондоне . Наука и ее концептуальные основы. Издательство Чикагского университета. ISBN 978-0-226-14346-0 . LCCN 89005137 . OCLC 709606191 .
- Фехт, Сара (19 октября 2011 г.). «Впервые показано, что долголетие наследуется не по ДНК-механизму» . Научный американец . Проверено 5 ноября 2015 г.
- Гиссис, Снайт Б.; Яблонка, Ева, ред. (2011). Трансформации ламаркизма: от тонких жидкостей к молекулярной биологии . Венская серия по теоретической биологии. Иллюстрации Анны Зелиговской. МТИ Пресс. ISBN 978-0-262-01514-1 . LCCN 2010031344 . OCLC 662152397 .
- Ханивилл, Росс (2008). Эволюция Ламарка: два столетия гения и ревности . Пирс 9 . ISBN 978-1-921208-60-7 . LCCN 2011431766 . OCLC 746154950 .
- Яблонка, Ева ; Лэмб, Мэрион Дж. (2008). «Эпигеном в эволюции: за пределами современного синтеза» (PDF) . Информационный бюллетень ВОГиС . 12 (1/2): 242–254.
- Медавар, Питер (1990). Пайк, Дэвид (ред.). Угроза и слава: размышления о науке и ученых . Предисловие Льюиса Томаса (1-е изд. в США). ХарперКоллинз . ISBN 978-0-06-039112-6 . LCCN 89046107 . OCLC 21977349 . Содержит лекции BBC Reith « Будущее человека ».
- Молино, Жан (2000). «К эволюционной теории музыки и языка» . В Валлине, Нильс Л. [на шведском языке] ; Меркер, Бьорн ; Браун, Стивен (ред.). Истоки музыки . МТИ Пресс. стр. 165–176. ISBN 978-0-262-23206-7 . LCCN 98054088 . OCLC 44963330 . «Состоит из докладов, представленных на семинаре по происхождению музыки, состоявшемся во Фьезоле, Италия, в мае 1997 года, первом из серии под названием «Флорентийские семинары по биомузыкологии».
- Пэн, Уэйн (27 декабря 2011 г.). «Ламаркианская вирусная защита у червей» . Природная генетика . 44:15 . дои : 10.1038/англ.1062 . S2CID 32406225 .
- Пенниси, Элизабет (6 сентября 2013 г.). «Эволюционная ересь? Эпигенетика лежит в основе наследственных признаков растений». Наука . 341 (6150): 1055. doi : 10.1126/science.341.6150.1055 . ПМИД 24009370 .
- Перселл, Стюарт (1999). Неоламаркизм и споры об эволюции во Франции, 1870-1920 гг . Исследования французской цивилизации. Том. 14. Эдвин Меллен Пресс . ISBN 978-0-7734-8275-3 . LCCN 98048633 . OCLC 40193707 .
- Секи, Ёсиюки (апрель 2013 г.). Гроссманн, Роберто Дж.; Ивакири, Ясуко; Таддеи, Тамар Х. (ред.). «Трансгенерационные эффекты, опосредованные сывороткой, на сперму: доказательства ламарковской наследственности?» . Гепатология . 57 (4): 1663–1665. дои : 10.1002/hep.26240 . ПМИД 23568276 . S2CID 5288601 .
- Уоддингтон, Конрад Х. (1961). «Эволюционная система человека». В Бэнтоне, Майкл (ред.). Дарвинизм и изучение общества: столетний симпозиум; Чикаго, Иллинойс . Тавистокские публикации; Четырехугольные книги . LCCN 61007932 . ОСЛК 1003950 . «Очерки... на основе статей, прочитанных на конференции, состоявшейся в Эдинбургском университете... 1959 г.».
- Уорд, Лестер Франк (1891). Неодарвинизм и неоламаркизм . Пресса Gedney & Roberts. LCCN 07037459 . OCLC 4115244 . «Ежегодное обращение президента Вашингтонского биологического общества. Прозвучало 24 января 1891 года. (Из трудов, т. VI.)» Неодарвинизм и неоламаркизм (1891 г.) в Интернет-архиве .
- Уайтлоу, Эмма (февраль 2006 г.). «Эпигенетика: Грехи отцов и их отцов» . Европейский журнал генетики человека . 14 (2): 131–132. дои : 10.1038/sj.ejhg.5201567 . ПМИД 16421606 . S2CID 36863159 .
- Юншэн Лю (сентябрь 2007 г.). «Как отец, так и сын. Свежий обзор наследования приобретенных признаков» . Отчеты ЭМБО . 8 (9): 798–803. дои : 10.1038/sj.embor.7401060 . ЧВК 1973965 . ПМИД 17767188 .