Генетическая история коренных народов Америки
Генетическая история коренных народов Америки разделена на два отдельных периода: первоначальное заселение Америки примерно 20 000–14 000 лет назад (20–14 тысяч лет назад) и контакт с европейцами примерно 500 лет назад. [1] [2] Первый период генетической истории является коренных американцев определяющим фактором для количества генетических линий, мутаций зиготности и основополагающих гаплотипов , присутствующих в сегодняшнем коренном американском населении . [3]
Коренное американское население происходит от древней восточноазиатской линии, которая отделилась от других восточноазиатских народов до последнего ледникового максимума и смешалась с древними северными евразийцами , палеолитическим сибирским населением, имеющим глубокое родство с европейскими охотниками-собирателями (западными евразийцами). [4] [5] [6] [7] Позже они рассеялись по Америке примерно 16 000 лет назад (исключение составляют группы, говорящие на на-дене и эскимосско-алеутах , которые частично произошли от сибирского населения, проникшего в Америку позже). [8] [9]
Анализ генетики коренных народов Америки и Сибири использовался в качестве аргумента в пользу ранней изоляции популяций-основателей Берингии . [10] а в дальнейшем – более быстрая миграция из Сибири через Берингию в Новый Свет . [11] Микросателлитное Южной разнообразие и распространение линии Y, специфичное для Америки, указывают на то, что некоторые коренные популяции Америки были изолированы с момента первоначального заселения региона. [12] На -дене , инуиты и коренные жители Аляски имеют гаплогруппу Q-M242 ; однако они отличаются от других коренных американцев различными мутациями мтДНК и атДНК. [13] [14] [15] Это говорит о том, что народы, впервые поселившиеся на северных окраинах Северной Америки и Гренландии, произошли от более поздних мигрантов, чем те, которые проникли южнее в Америку. [16] [17] Лингвисты и биологи пришли к аналогичному выводу на основе анализа языковых групп коренных американцев и распределения групп крови по системе ABO . [18] [19] [20] [21]
Аутосомная ДНК
[ редактировать ]

Генетическое разнообразие и структура населения на территории Америки также измеряется с использованием аутосомных (атДНК) микросателлитных маркеров генотипированных ; взяты образцы из Северной, Центральной и Южной Америки и проанализированы на основе аналогичных данных, доступных у других коренных народов по всему миру. [23] [24] Коренные популяции Америки демонстрируют меньшее генетическое разнообразие , чем популяции из других континентальных регионов, в то время как их общее самое сильное родство с другими популяциями наблюдается у палеосибирских народов . [24] [5] [6] Наблюдается уменьшение генетического разнообразия по мере географического удаления от Берингова пролива, а также уменьшение генетического сходства с сибирскими популяциями с Аляски (генетическая точка входа). [23] [24] Также наблюдаются свидетельства более высокого уровня разнообразия и меньшего уровня структуры населения в западной части Южной Америки по сравнению с восточной частью Южной Америки. [23] [24] Существует относительное отсутствие дифференциации между мезоамериканским и андским населением, сценарий, который подразумевает, что прибрежные маршруты (в данном случае вдоль побережья Тихого океана) было легче пересекать мигрирующим народам (вносящим больший генетический вклад) по сравнению с внутренними маршрутами. [23]
Общая картина предполагает, что Америка была колонизирована небольшим количеством особей (эффективный размер около 70), которое выросло на многие порядки за 800–1000 лет. [25] [26] Данные также показывают, что между Азией, Арктикой и Гренландией происходил генетический обмен с момента первоначального заселения Америки. [26] [27]
По данным аутосомно-генетического исследования 2012 года, [28] Коренные американцы происходят как минимум от трех основных волн мигрантов из Северной Азии. Большая часть их восходит к одному предковому населению, называемому «первыми американцами». Однако те, кто говорит на языках инуитов из Арктики, унаследовали почти половину своего происхождения от второй волны мигрантов из Восточной Азии, а те, кто говорит на-дене, унаследовали десятую часть своего происхождения от третьей волны мигрантов. За первоначальным заселением Америки последовала быстрая экспансия на юг вдоль западного побережья с небольшим потоком генов позже, особенно в Южной Америке . Единственным исключением из этого правила являются носители языка чибча в Колумбии, чьи предки происходят как из Северной, так и из Южной Америки. [28]
В 2014 году была секвенирована аутосомная ДНК младенца возрастом более 12 500 лет из Монтаны. [29] ДНК была взята из скелета, известного как Анзик-1, найденного в тесной связи с несколькими артефактами Кловиса. Сравнения показали сильное сходство с ДНК из сибирских памятников и практически исключили, что конкретный человек имел какое-либо близкое родство с европейскими источниками (« Солютрейская гипотеза »). ДНК также показала сильное родство со всеми существующими коренными народами Америки, что указывает на то, что все они происходят от древнего населения, жившего в Сибири или рядом с ней. [30]
Лингвистические исследования подкрепили генетические исследования, установив связи между языками, на которых говорят в Сибири, и языками, на которых говорят в Америке. [31]
Два генетических исследования аутосомной ДНК, проведенные в 2015 году, подтвердили сибирское происхождение коренных народов Америки. Однако древний сигнал общего происхождения с австралазийцами (коренными народами Австралии, Меланезии и Андаманских островов ) был обнаружен среди коренных народов региона Амазонки . Этот сигнал также вызывает сомнения, поскольку «популяция Y» в последнее время была связана с глубокой восточноазиатской популяцией, которую можно ассоциировать с человеком Тяньюань и которая является предком современных аборигенов Восточной Азии. Глубокие линии Тяньюань и Восточной Азии образуют сестринскую ветвь андаманского и австралийского населения, причем все они являются ветвями древних восточноевразийцев . Основная миграция из Сибири в Америку произошла 23 000 лет назад. [32] [33] [34] [35] [36] [37] [38]
В исследовании 2018 года были проанализированы древние образцы коренных народов. Генетические данные позволяют предположить, что все коренные американцы в конечном итоге произошли от основной популяции, которая отделилась от аборигенов Восточной Азии и впоследствии смешалась с древними жителями Северной Евразии . Авторы также предоставили доказательства того, что базальные северные и южные ветви коренных американцев, к которым принадлежат все остальные коренные народы, разошлись около 16 000 лет назад. [8] Образец коренных американцев из Айдахо , датируемый 16 000 г. до н.э. , который краниометрически похож на современных коренных американцев, а также на палеосибирцев , оказался генетически «индейцем» и показал высокую близость к определенным восточноазиатским группам, таким как образцы периода Дзёмон из Японии. , подтверждая, что предки коренных американцев отделились от исходной популяции где-то в северо-восточной Сибири. [39]
Исследование, опубликованное в журнале Nature в 2018 году, пришло к выводу, что коренные американцы произошли от единой основной популяции, которая первоначально отделилась от аборигенов Восточной Азии примерно на 36 000 (± 1500) лет назад, при этом поток генов между этой разделенной группой предков коренных американцев и сибиряков сохранялся до тех пор, пока около ~ 25 000 (± 1100) лет назад, до слияния с древними жителями Северной Евразии и последующей изоляции в Америке или Берингии примерно 22 000 лет назад. Коренные народы Северной и Южной Америки отделились друг от друга примерно примерно от 17 500 до 14 600 лет назад. Есть также некоторые свидетельства обратной миграции из Америки в Сибирь примерно через 11 500 лет назад. [8]
В исследовании, опубликованном в журнале Cell в 2019 году, было проанализировано 49 образцов древних коренных американцев со всей Северной и Южной Америки, и был сделан вывод, что все коренные американцы произошли от единого предкового источника, который отделился от древних жителей Восточной Азии и смешался с древним севером. Евразийцы (ANE) и дали начало «предкам коренных американцев», которые позже разделились на различные группы коренных народов. Авторы также отвергли предыдущие утверждения о возможности существования двух отдельных групп населения среди населения Америки. И северные, и южные коренные американцы наиболее близки друг к другу и не имеют признаков примеси с гипотетическими предыдущими популяциями. [40]

Обзорная статья, опубликованная в журнале Nature в 2021 году и обобщающая результаты предыдущих геномных исследований, аналогичным образом пришла к выводу, что все коренные американцы произошли от перемещения людей из Северо-Восточной Азии в Америку. Эти предки американцев, когда-то жившие к югу от континентальных ледниковых щитов, быстро распространились и расширились, разделившись на множество групп, которые позже дали начало основным подгруппам коренного населения Америки. Исследование также отвергло существование, сделанное на основе краниометрических данных, гипотетического отдельного некоренного американского населения (предположительно связанного с коренными австралийцами и папуасами), иногда называемого «палеоамериканцем». [42] [43]
В целом, «предки коренных американцев» произошли от смеси древней восточноазиатской линии, составляющей от 56 до 68% предков, и палеолитического сибирского населения, известного как древние северные евразийцы , составляющего около 32–44% предков. Предки коренных американцев также были тесно связаны с « древними палеосибирцами » и « древними берингийцами ». [44] [40] [6] [5] Коренные американцы (американцы) также описываются как имеющие «преимущественно восточноазиатское происхождение, с меньшим вкладом палеолитического населения Западной Евразии». [45]
ДНК Y-хромосомы
[ редактировать ]
Было постулировано «центральносибирское» происхождение отцовской линии исходных популяций первоначальной миграции в Америку. [46]
Членство в гаплогруппах Q и C3b подразумевает коренное американское происхождение по отцовской линии. [47]
Микросателлитное разнообразие и распространение линии Y , специфичной для Южной Америки, позволяют предположить, что некоторые коренные народы Америки оказались изолированными после первоначальной колонизации своих регионов. [12] На -дене , инуиты и коренные жители Аляски демонстрируют мутации гаплогруппы Q (Y-ДНК) , но отличаются от других коренных американцев различными мутациями мтДНК и аутосомной ДНК (атДНК). [13] [48] [49] Это говорит о том, что самые ранние мигранты в северные районы Северной Америки и Гренландии произошли от более поздних мигрантов. [50] [51]
Гаплогруппа Q
[ редактировать ]
Q-M242 (мутационное имя) является определяющим (SNP) гаплогруппы Q (Y-ДНК) (филогенетическое имя). [53] [54] В Евразии гаплогруппа Q встречается среди древних экземпляров Афонтовой Горы и коренных сибирских популяций , таких как современные чукчи и коряки , а также у некоторых жителей Юго-Восточной Азии, таких как даяки . В частности, большие концентрации мутации Q-M242 наблюдаются в двух группах – кетах (93,8%) и селькупах (66,4%). [55] Кеты считаются единственными оставшимися в живых из древних странников, живших в Сибири. [56] Численность их населения очень мала; насчитывается менее 1500 кетов в России . 2002 [25] Селькупы имеют немного большую численность населения, чем кети, и насчитывают около 4250 особей. [57]
Начиная с палеокоренного периода Америки , произошла миграция в Америку через Берингов пролив ( Берингия ) небольшой популяции, несущей мутацию Q-M242. [14] Член этой исходной популяции претерпел мутацию, которая определяет его потомковую популяцию, известную как мутация Q-M3 (SNP). [58] Эти потомки мигрировали по всей Америке. [53]
Гаплогруппа Q-M3 определяется наличием rs3894 (M3) (SNP). [1] [25] [59] Мутации Q-M3 примерно 15 000 лет, поскольку именно тогда произошла первоначальная миграция палеокоренных американцев в Америку. [60] [61] Q-M3 является преобладающим гаплотипом в Америке, его доля в популяциях Южной Америки составляет 83%. [12] 50% в популяциях на-дене и в североамериканских эскимосско-алеутских популяциях - около 46%. [55] При минимальной обратной миграции Q-M3 в Евразию мутация, вероятно, развилась в восточной Берингии, а точнее на полуострове Сьюард или в западной части Аляски . Берингийский массив начал погружаться под воду, отрезая сухопутные пути. [55] [62] [23]
С момента открытия Q-M3 несколько субкладов было обнаружено популяций, несущих M3. Примером может служить Южная Америка , где в некоторых популяциях наблюдается высокая распространенность (SNP) M19, что определяет субклад Q-M19 . [12] M19 был обнаружен у (59%) мужчин амазонских тикуна и у (10%) мужчин вайуу . [12] Субклад M19, по-видимому, уникален для коренных народов Южной Америки и возник от 5000 до 10 000 лет назад. [12] Это говорит о том, что изоляция населения и, возможно, даже создание племенных групп началась вскоре после миграции в районы Южной Америки. [25] [63] Другие американские субклады включают Q-L54 , Q-Z780 , Q-MEH2 , Q-SA01 и Q-M346 линии . В Канаде были обнаружены еще две линии. Это Q-P89.1 и Q-NWT01 .
Гаплогруппа R1b1a1a2 (M269)
[ редактировать ]R1b1a1a2 (M269) — вторая по распространенности гаплогруппа Y-ДНК, обнаруженная среди коренных американцев, после гаплогруппы Y-ДНК Q. [64]
Линии R1b1a1a2 (M269), обычно встречающиеся у коренных американцев, в большинстве случаев принадлежали к субкладу R1b1a1a2 (M269), наиболее распространенному у западных европейцев, а его самая высокая концентрация обнаруживается среди множества алгонкинских племен на востоке Северной Америки. [65]
Таким образом, по мнению ряда авторов, R1b, скорее всего, был занесен в результате примеси во время после 1492 года . европейского заселения Северной Америки [66] [67] [68]
R1b1a1a2 (M269) встречается преимущественно у североамериканских групп, таких как оджибве (50-79%), семинолы (50%), сиу (50%), чероки (47%), догрибы (40%) и тохоно о'одхам ( Папаго) (38%). Его самая высокая частота наблюдается на северо-востоке Северной Америки, а частота снижается с востока на запад. В индейских племенах юго-запада частота этой гаплогруппы составляет всего 4%. [64] [69]
Гаплогруппа C-P39
[ редактировать ]
Гаплогруппа C-M217 встречается преимущественно у коренных сибиряков, монголов и казахов . Гаплогруппа C-M217 — наиболее распространенная и часто встречающаяся ветвь большой (Y-ДНК) гаплогруппы C-M130 . Потомок гаплогруппы C-M217 C-P39 чаще всего встречается у современных носителей языка на-дене , при этом наибольшая частота встречается среди атабасков - 42%, а с меньшей частотой - у некоторых других групп коренных американцев. [14] Эта отдельная и изолированная ветвь C-P39 включает почти все Y-хромосомы гаплогруппы C-M217, обнаруженные у всех коренных народов Америки. [71]
Некоторые исследователи полагают, что это может указывать на то, что миграция на-дене произошла с Дальнего Востока России после первоначальной колонизации палео-коренных американцев, но до современной экспансии инуитов , инупиатов и юпиков . [14] [13] [72]
Помимо народов на-дене, гаплогруппа C-P39 (C2b1a1a) также встречается среди других коренных американцев, таких как население, говорящее на алгонкинском и сиуанском языках. [73] [74] C-M217 встречается у народа вайу в Колумбии и Венесуэле . [73] [74]
Данные
[ редактировать ]Здесь перечислены известные коренные народы Америки по гаплогруппам ДНК Y-хромосомы человека на основе соответствующих исследований. Выборки берутся у лиц, идентифицированных по этническим и языковым обозначениям в первых двух столбцах, четвертый столбец ( n ) — это размер изучаемой выборки, а в остальных столбцах указан процент конкретной гаплогруппы .
| Группа | Язык | Место | н | С | вопрос | Р1 | Другие | Ссылка |
|---|---|---|---|---|---|---|---|---|
| Алгонкинский [номер 1] | Алгик | Северо-восток Северной Америки | 155 | 7.7 | 33.5 [номер 2] | 38.1 | 20.6 | Больник 2006 г. [75] |
| Апач | На-Дене | ЮЗ США | 96 | 14.6 | 78.1 | 5.2 | 2.1 | Увидимся в 2004 году [14] |
| Атабаскский [номер 3] | На-Дене | Западная часть Северной Америки | 243 | 11.5 | 70.4 | 18.1 | Малхи 2008 г. [64] | |
| Чероки | ирокезский | ЮВ США | 62 | 1.6 | 50.0 [номер 4] | 37.1 | 11.3 | Больник 2006 г. [75] |
| Чероки | ирокезский | Восточная Северная Америка | 30 | 50.0 | 46.7 | 3.3 | Малхи 2008 г. [64] | |
| Шайенн | Алгик | Соединенные Штаты | 44 | 16 | 61 | 16 | 7 | Увидимся в 2004 году [14] |
| Чибчан [номер 5] | Макро-Чибчан | Панама | 26 | 100 | Увидимся в 2004 году [14] | |||
| Чипевян | На-Дене | Канада | 48 | 6 | 31 [номер 6] | 62.5 [номер 7] | Бортолони 2003 г. [12] | |
| Чиппева | Алгик | Восточная Северная Америка | 97 | 4.1 | 15.9 [номер 8] | 50.5 | 29.9 | Больник 2006 г. [75] |
| Догриб | На-Дене | Канада | 15 | 33 | 27 | 40 | Малхи 2008 г. [64] | |
| Догриб | На-Дене | Канада | 37 | 35.1 | 45.9 [номер 9] | 8.1 | 10.8 | Дулик 2012 [76] |
| Макро-J | Бразилия | 51 | 92 [номер 11] | 8 | Бортолони 2003 г. [12] | |||
| Гуарани | Тупиан | Парагвай | 59 | 86 [номер 12] | 9 | 5 | Бортолони 2003 г. [12] | |
| Нет | кечуа | Колумбия | 11 | 78 [номер 13] | 11 | 11 | Бортолони 2003 г. [12] | |
| Инуиты | Эскимосско-алеутский | Североамериканская Арктика | 60 | 80.0 | 11.7 | 8.3 | Увидимся в 2004 году [14] | |
| Нарушенный | Эскимосско-алеутский | Канада | 56 | 1.8 | 55.1 [номер 14] | 33.9 | 8.9 | Дулик 2012 [76] |
| майя | Мезоамерика | 71 | 87.3 | 12.7 | Увидимся в 2004 году [14] | |||
| Смешивание | Миксе-Соке | Мексика | 12 | 100 | Увидимся в 2004 году [14] | |||
| Микстек | Ото-Мангеан | Мексика | 28 | 93 | 7 | Увидимся в 2004 году [14] | ||
| Маскогейский [номер 15] | Маскогейский | ЮВ США | 36 | 2.8 | 75 [номер 16] | 11.1 | 11.1 | Больник 2006 г. [75] |
| Науа | Уто-ацтекский | Мексика | 17 | 94 | 6 | Малхи 2008 г. [64] | ||
| Коренные американцы (США) | Соединенные Штаты | 398 | 9.0 | 58.1 | 22.2 | 10.7 | Хаммер 2005 г. [77] | |
| Навахо | На-Дене | ЮЗ США | 78 | 1.3 | 92.3 | 2.6 | 3.8 | Увидимся в 2004 году [14] |
| Коренные жители Северной Америки | Северная Америка | 530 | 6.0 | 77.2 | 12.5 | 4.3 | Увидимся в 2004 году [14] | |
| попугай | Уто-ацтекский | ЮЗ США | 13 | 61.5 | 38.5 | Малхи 2008 г. [64] | ||
| семинолы | Маскогейский | Восточная Северная Америка | 20 | 45.0 | 50.0 | 5.0 | Малхи 2008 г. [64] | |
| Сиу | Макросиуан | Центральная Северная Америка | 44 | 11 | 25 | 50 | 14 | Увидимся в 2004 году [14] |
| Южная Америка | индеец | Южная Америка | 390 | 92 [номер 17] | 4 | 4 | Бортолони 2003 г. [12] | |
| Рука | На-Дене | Северо-запад Северной Америки | 12 | 42 | 42 | 8 | 8 | Увидимся в 2004 году [14] |
| Тикуна | Тикуна – Юрий | Западный бассейн Амазонки | 33 | 100 [номер 18] | Бортолони 2003 г. [12] | |||
| Тлинкиты | На-Дене | Тихоокеанский северо-запад | 11 | 18 [номер 19] | 82 [номер 20] | Дулик 2012 [76] | ||
| Тупи – Гуарани [номер 21] | Тупиан | Бразилия | 54 | 100 [номер 22] | Бортолони 2003 г. [12] | |||
| Уто-ацтекский [номер 23] | Уто-ацтекский | Мексика , Аризона | 167 | 93.4 | 6.0 | Малхи 2008 г. [64] | ||
| Варао | Варао (изолировать) | Карибский бассейн Южная Америка | 12 | 100 [номер 24] | Бортолони 2003 г. [12] | |||
| Вайуу | Аравакан | Полуостров Гуахира | 19 | 69 [номер 25] | 21 | 10 | Бортолони 2003 г. [12] | |
| Вайуу | Аравакан | Полуостров Гуахира | 25 | 8 | 36 | 44 | 12 | Увидимся в 2004 году [14] |
| Ягуа | Пеба – Ягуань | Перу | 7 | 100 [номер 26] | Бортолони 2003 г. [12] | |||
| Юкпа | карибский | Колумбия | 12 | 100 [номер 27] | Бортолони 2003 г. [12] | |||
| Сапотек | Ото-Мангеан | Мексика | 16 | 75 | 6 | 19 | Увидимся в 2004 году [14] | |
| Зену | вымерший | Колумбия | 30 | 81 [номер 28] | 19 | Бортолони 2003 г. [12] |
Митохондриальная ДНК
[ редактировать ]Давно признано широкое распространение гаплогрупп A, B, C и D мтДНК среди населения Восточной Азии и коренных американцев, а также наличие гаплогруппы X. [78] В целом, наибольшая частота четырех связанных с коренными американцами гаплогрупп встречается в Алтае - Байкальском регионе южной Сибири. [79] Некоторые субклады C и D, более близкие к субкладам коренных американцев, встречаются среди монгольского, амурского, японского, корейского и айнского населения. [78] [80] Исследование ДНК 2023 года показало, что «[в] дополнение к ранее описанным предковым источникам в Сибири, Австрало-Меланезии и Юго-Восточной Азии, ... северный прибрежный Китай также внес свой вклад в генофонд коренных американцев», а также генофонд японцев. люди. [81]

При изучении гаплогруппы митохондриальной ДНК человека результаты показали, что гаплогруппы коренных американцев, включая гаплогруппу X , являются частью единой восточноазиатской популяции. Это также указывает на то, что распределение гаплогрупп мтДНК и уровни расхождения последовательностей среди лингвистически сходных групп были результатом многочисленных предшествующих миграций из популяций Берингова пролива . [82] Вся мтДНК коренных американцев можно отнести к пяти гаплогруппам A , B , C , D и X. : [83] [84] Более конкретно, мтДНК коренных американцев принадлежит к субгаплогруппам A2, B2, C1b, C1c, C1d, D1 и X2a (с минорными группами C4c, D2a и D4h3a). [10] [82] Это предполагает, что 95% мтДНК коренных американцев происходят от минимальной генетической популяции женщин-основателей, включающей субгаплогруппы A2, B2, C1b, C1c, C1d и D1. [83] Остальные 5% состоят из подгаплогрупп X2a, D2a, C4c и D4h3a. [82] [83]
X — одна из пяти гаплогрупп мтДНК, обнаруженных у коренных американцев. Коренные американцы в основном относятся к кладе X2a, которая никогда не встречалась в Старом Свете . [85] По словам Дженнифер Рафф , X2a, вероятно, произошел от той же сибирской популяции, что и остальные четыре материнские линии-основатели. [86]
Генетические последовательности гаплогруппы X разошлись примерно 20 000–30 000 лет назад, образовав две подгруппы: X1 и X2. Субклад X2a X2 встречается только с частотой около 3% от общей численности нынешнего коренного населения Америки. [25] Однако X2a является основным субкладом мтДНК в Северной Америке; у алгонкинских народов он составляет до 25% типов мтДНК. [1] [87] Он также присутствует в меньшем количестве к западу и югу от этой территории — среди сиу (15%), нуу-ча-нулт (11–13%), навахо (7%) и якама (5% ). %). [88] Преобладающая теория появления субгаплогруппы X2a в Северной Америке - это миграция вместе с группами мтДНК A, B, C и D из источника в Горном Алтае в Центральной Азии. [89] [90] [91] [92] Гаплотип X6 присутствовал у тараумара 1,8% (1/53) и уичоли 20% (3/15). [93]
Секвенирование митохондриального генома палеоэскимосских останков (возрастом 3500 лет) отличается от современных коренных американцев и попадает в субгаплогруппу D2a1, группу, наблюдаемую среди сегодняшних жителей Алеутских островов , алеутов и сибирских популяций юпиков . [94] Это говорит о том, что колонизаторы крайнего севера, а впоследствии и Гренландии, произошли от более позднего прибрежного населения. [94] Затем начался генетический обмен на северных крайностях, завезенный народом Туле (протоинуитами) примерно 800–1000 лет назад. [49] [95] Эти последние доколумбовые мигранты представили гаплогруппы A2a и A2b существующему палеоэскимосскому населению Канады и Гренландии, кульминацией которого стали современные инуиты . [49] [95]

Маршрут через Берингию рассматривается как более вероятный, чем солютрейская гипотеза . [96] В реферате в выпуске «Американского журнала физической антропологии» за 2012 год говорится, что «сходство в возрасте и географическом распространении C4c и ранее проанализированной линии X2a подтверждает сценарий двойного происхождения палеокоренных американцев. Принимая во внимание Учитывая, что C4c глубоко укоренен в азиатской части филогении мтДНК и, несомненно, имеет азиатское происхождение, открытие о том, что C4c и X2a характеризуются параллельными генетическими историями, окончательно отвергает спорную гипотезу о пути проникновения атлантических ледников в Северную Америку». [97]
Другое исследование, также сосредоточенное на мтДНК (которая наследуется только по материнской линии), [10] показало, что коренные народы Америки могут проследить свою родословную по материнской линии до нескольких линий-основателей из Восточной Азии, которые могли прийти через Берингов пролив . Согласно этому исследованию, вполне вероятно, что предки коренных американцев какое-то время оставались в районе Берингова пролива , после чего должно было произойти быстрое движение заселения Америки, перенося линии основателей на Юг. Америка .
Согласно исследованию 2016 года, посвященному линиям мтДНК, «небольшая популяция проникла в Америку прибрежным маршрутом около 16 тыс. лет назад, после предыдущей изоляции в восточной Берингии в течение ~ 2,4–9 тысяч лет после отделения от восточно-сибирских популяций. После быстрого перемещения по всей Америке ограниченный поток генов в Южной Америке привел к заметной филогеографической структуре популяций, которая сохранялась во времени. Все древние митохондриальные линии, обнаруженные в этом исследовании, отсутствовали в современных наборах данных, что предполагает высокую скорость вымирания . кроме того, мы применили новый тест множественной логистической регрессии главных компонентов к байесовским серийным слияющим моделям. Анализ подтвердил сценарий, в котором европейская колонизация привела к существенной утрате доколумбовых линий». [98]
Генетическая примесь
[ редактировать ]Древние берингийцы
[ редактировать ]
Недавние археологические находки на Аляске пролили свет на существование ранее неизвестного коренного населения Америки, получившего академическое название « Древние берингийцы ». [99] Хотя среди археологов широко распространено мнение, что первые поселенцы перебрались на Аляску из России по сухопутному мосту через Берингов пролив , вопрос о том, была ли одна группа основателей или несколько волн миграции, является спорным и широко распространенным спором среди ученых в этой области. сегодня. В 2018 году секвенированная ДНК девушки из числа коренных народов, останки которой были найдены на археологическом объекте Апвард-Сан-Ривер на Аляске в 2013 году, оказалась не соответствующей двум признанным ветвям коренных американцев и вместо этого принадлежала раннему населению древних берингийцев. [100] Этот прорыв считается первым прямым геномным доказательством того, что потенциально произошла только одна волна миграции в Америке, причем генетическое ветвление и деление произошли постфактум. По оценкам, волна миграции возникла около 20 000 лет назад. [99] Говорят, что древние берингийцы сегодня являются общей предковой группой среди современного коренного населения Америки, что отличается от результатов, полученных от предыдущих исследований, которые предполагают, что современные популяции являются потомками как северной, так и южной ветвей. [99] Эксперты также смогли использовать более широкие генетические данные, чтобы установить, что раскол между северной и южноамериканской ветвями древних берингийцев на Аляске произошел всего примерно 17 000 и 14 000 лет назад. [28] еще больше бросая вызов концепции множественных миграционных волн, возникающих на самых первых этапах заселения.
Генетические данные о палеокоренных американцах состоят из присутствия явной примеси архаичных линий Сундадонта к отдаленным популяциям тропических лесов Южной Америки, а также в генетике патагонцев-огнеземцев . [101] [102]
Номатто и др. (2009) предполагаемая миграция в Берингию произошла между 40 000 и 30 000 лет назад, при этом миграция до LGM в Америку сопровождалась изоляцией северной популяции после закрытия незамерзающего коридора. [103]
Генетическое исследование коренных народов амазонского региона Бразилии в 2016 году (проведенное Скоглундом и Райхом) показало доказательства примеси отдельной линии неизвестного древнего народа. Эта древняя группа, по-видимому, связана с современными « австралазийскими » народами (т.е. аборигенами Австралии и меланезийцами ). Эта « призрачная популяция » была обнаружена у носителей тупианских языков . Они предварительно назвали эту древнюю группу; «Население Y», от Ypykuéra , «что означает «предок» в языковой семье тупи». [34] Генетическое исследование 2021 года опровергло существование гипотетического австралазийского компонента среди коренных американцев. Сигнал гипотетического австралийского компонента также может быть воспроизведен с использованием мужской выборки из базально-восточноазиатского Тяньюаня и, таким образом, не представляет собой «реальное родство с Австралазией». Авторы объяснили, что предыдущие заявления о возможном австралийском происхождении были основаны на неверно истолкованном генетическом эхо, которое, как выяснилось, представляет собой ранний восточно-евразийский поток генов (представленный старым образцом из Тяньюаня, датируемым 40 000 г. до н. э.) к аборигенам Австралии и папуасам, который был потерян в современные жители Восточной Азии. [42] [43]
Археологические свидетельства присутствия человека в Америке до LGM были впервые представлены в 1970-х годах. [104] [105] в частности, череп « Женщины Лузии », найденный в Бразилии. [106] [107] [108]
Старый мир
[ редактировать ]
Существенное расовое смешение имело место во время и после европейской колонизации Америки . [109] [110]
Южная и Центральная Америка
[ редактировать ]В Латинской Америке, в частности, имело место значительное расовое смешение между коренным американским населением, колониальным населением европейского происхождения и населением стран Африки к югу от Сахары, импортированным в качестве рабов . Примерно с 1700 года возникла латиноамериканская терминология для обозначения различных комбинаций смешанного расового происхождения, возникших в результате этого. [111]
Многие люди, идентифицирующие себя как представители одной расы, демонстрируют генетические свидетельства многорасового происхождения. [ сломанный якорь ] . [112] Европейское завоевание Южной и Центральной Америки, начавшееся в конце 15 века, первоначально осуществлялось мужчинами-солдатами и моряками с Пиренейского полуострова ( Испания и Португалия ). [113] [ ненадежный источник ] Новые солдаты-поселенцы родили детей от американских женщин из числа коренного населения, а затем от африканских рабов . [114] [ ненадежный источник ] этих детей смешанной расы называли Испанские и португальские колонисты Кастас « » . обычно [115]
Северная Америка
[ редактировать ]в Торговля мехом в Северной Америке 16 веке привела к тому, что многие европейцы из Франции , Ирландии и Великобритании женились на женщинах из числа коренных народов Северной Америки. [116] В районах, где эти народы образовали сообщества и развили уникальную синкретическую культуру, их дети стали известны французским колонистам как » или « буа - брюле » « метисы . также называли эти народы « полукровками » или « выходцами из страны » колонисты В некоторых контекстах английские и шотландские . [117]
Коренные американцы в Соединенных Штатах определяются гражданством, культурой и семейными отношениями, а не расой. [118] [119] Никогда не определив индейскую идентичность как расовую, [118] исторически коренные американцы обычно практиковали то, что основное общество определяет как межрасовые браки , что повлияло на расовые представления о количестве крови . [120]
В переписи населения США 2010 года почти 3 миллиона человек указали, что они являются коренными американцами (включая коренных жителей Аляски). [121] Это основано на самоидентификации, поскольку перепись не требует документального подтверждения этого убеждения. Особенно многочисленной была самоидентификация этнического происхождения чероки , [122] явление, получившее название « синдром чероки », когда некоторые американцы полагают, что у них есть «давно потерянный предок чероки», но не могут идентифицировать ни одного чероки или коренных американцев в своем генеалогическом древе или среди своих ныне живущих родственников. [123] [124] Такое культивирование оппортунистической этнической идентичности связано с «престижем», который некоренные жители могут ассоциировать с коренным американским происхождением, никогда не испытывая каких-либо сопутствующих трудностей или угнетения. [125] В частности, на востоке США притворщики распространены. [125] [126]
Некоторые племена приняли требования к количеству крови или Сертификаты степени индийской крови и практикуют исключение членов племени, неспособных доказать, что они являются детьми зарегистрированного члена племени. В этих случаях племя может потребовать проведения теста на отцовство . Для некоторых это стало спорным вопросом в политике резерваций коренных американцев . [127]
Европейские болезни и генетическая модификация
[ редактировать ]Команда под руководством Рипана Малхи, антрополога из Университета Иллинойса в Урбане , провела исследование, в котором они использовали научную технику, известную как секвенирование всего экзома, для тестирования вариантов генов, связанных с иммунитетом, у коренных американцев. [128] Путем анализа древней и современной ДНК коренных народов было обнаружено, что HLA-DQA1 , вариант гена, который кодирует белок, отвечающий за дифференциацию здоровых клеток от вторгающихся вирусов и бактерий, присутствовал почти в 100% древних останков, но только в 36% в современных. Коренные американцы. [128] Эти данные свидетельствуют о том, что эпидемии европейского происхождения, такие как оспа, изменили картину болезней в Америке, в результате чего выжившие после этих вспышек с меньшей вероятностью являются носителями таких вариантов, как HLA-DQA1. Это сделало их менее способными справляться с новыми заболеваниями. По оценкам ученых, изменение в генетическом составе произошло около 175 лет назад, в то время, когда эпидемия оспы распространялась по Америке. [ нужна ссылка ] .
Группы крови
[ редактировать ]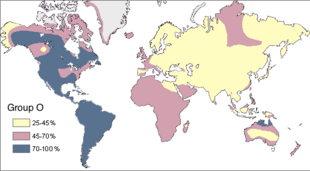
До подтверждения Альфредом Херши и Мартой Чейз в 1952 году ДНК как наследственного материала ученые использовали белки крови для изучения генетических вариаций человека. [129] [130] Считается, что система групп крови АВО была открыта австрийцем Карлом Ландштейнером , который обнаружил три различные группы крови в 1900 году. [131] Группы крови наследуются от обоих родителей. Группа крови АВО контролируется одним геном ( геном АВО ) с тремя аллелями : i , I. А и я Б . [132]
Исследования Людвика и Ханки Хершфельд во время Первой мировой войны показали, что частота групп крови A, B и O сильно различалась от региона к региону. [130] Группа крови «О» (обычно возникающая в результате отсутствия аллелей А и В) очень распространена во всем мире, ее частота составляет 63% во всех человеческих популяциях. [133] Группа «О» является основной группой крови среди коренного населения Америки, особенно среди населения Центральной и Южной Америки, с частотой почти 100%. [133] У коренного населения Северной Америки частота типа «А» колеблется от 16% до 82%. [133] Это еще раз говорит о том, что первоначальные коренные американцы произошли от изолированной популяции с минимальным количеством особей. [134] [135]
Стандартное объяснение такой высокой численности коренных американцев с группой крови O – это генетический дрейф . Поскольку предковая популяция коренных американцев была малочисленной, разнообразие групп крови могло уменьшаться от поколения к поколению из-за эффекта основателя . [136] Другие связанные объяснения включают объяснение «Узкого места», в котором говорится, что среди коренных американцев была высокая частота групп крови A и B, но серьезное сокращение населения в 1500-х и 1600-х годах, вызванное заносом болезней из Европы, привело к огромному числу смертей тех, у кого была кровь. типы A и B. По совпадению, большая часть выживших относилась к типу O. [136]
| ГРУППА ЛЮДЕЙ | О (%) | А (%) | Б (%) | АБ (%) |
|---|---|---|---|---|
| Конфедерация черноногих (коренные жители Северной Америки) | 17 | 82 | 0 | 1 |
| Бороро (Бразилия) | 100 | 0 | 0 | 0 |
| Эскимосы (Аляска) | 38 | 44 | 13 | 5 |
| Инуиты (Восточная Канада и Гренландия) | 54 | 36 | 23 | 8 |
| Гавайцы ( полинезийцы , некоренные американцы) | 37 | 61 | 2 | 1 |
| Коренные жители Северной Америки (в целом коренные народы / первые нации ) | 79 | 16 | 4 | 1 |
| Maya (modern) | 98 | 1 | 1 | 1 |
| Навахо | 73 | 27 | 0 | 0 |
| Перу | 100 | 0 | 0 | 0 |
Ди а Антиген антигенной системы Диего был обнаружен только у коренных народов Америки и Восточной Азии, а также у людей, имеющих некоторое происхождение от этих групп. Частота Ди а Антигена у различных групп коренных народов Америки колеблется почти от 50% до 0%. [138] Различия в частоте встречаемости антигена в популяциях коренных народов Америки коррелируют с основными языковыми семьями, изменяющимися под влиянием условий окружающей среды. [139]
См. также
[ редактировать ]- Введение в генетику
- Археогенетика
- Археология Америки
- Древняя ДНК
- Культура Хлодвига
- Ранние миграции человека
- Генетическая история Африки
- Генетическая история Европы
- Генетическая история Италии
- Генетическая история Северной Африки
- Генетическая история Британских островов
- Генетическая история Пиренейского полуострова
- Генетическая история Ближнего Востока
- Генетика и археогенетика Южной Азии
- Список гаплогрупп исторических людей
- Генетика майя
- Раса и генетика
- Заселение Америки § Оценка геномного возраста
- Список гаплогрупп Y-хромосомы в популяциях мира
Примечания
[ редактировать ]- ^ Алгонкинские этнические группы: оджибве , шайены / арапахо , шауни , микмак , кикапу и мескваки .
- ^ Q-M3=12,9; Q(xM3)=20,6.
- ^ Атабаскские этнические группы: чипевян , тлычу , танана , апачи и навахо .
- ^ Q-M3=32.; Q3(xM3)=17,7.
- ^ Этнические группы чибчан: нгобе и куна . народы
- ^ Q-M3=6; Q(xM3)=25.
- ^ P1 (xQ) 62,5%. Хотя другие исследования идентифицируют это как R(xR2)/R1b ,
эта тема остается спорной (см. Хаммер, Майкл Ф. и др., 2005 г.). - ^ Q-M3=8,2; Q(xQ-M3)=7,2.
- ^ Q-M3=40,5; Q(xM3)=5,4.
- ^ Этнические группы ге: Голотире, Кайганг, Крахо, Мекраноти и Ксикрин.
- ^ Q-M3=90; Q(xM3)=2)
- ^ Q-M3=79; Q(xM3)=7.
- ^ Q-M3=11; Q(xM3)=67.
- ^ Q-M3=10.7; NWT01=44.6.
- ^ Маскогские этнические группы: чикасо , чокто , маскоги и семинолы .
- ^ Q-M3=50,0; Q(xM3)=25,0.
- ^ Q-M3=83; Q(xM3)=9.
- ^ Q-M3=89; Q(xM3)=11.
- ^ С3*=9; C3b=9
- ^ Q-M3=64; Q-MEH2*=9; Q-NWT01=9.
- ^ Тупи-гуарани Бразильские этнические группы: асурини, паракана, каапор и ваямпи .
- ^ Все примеры гаплогруппы Q были Q-M3.
- ^ Уто-ацтекские этнические группы: пима , тохоно о'одхам , тараумара , науа , кора и уичоль .
- ^ Q=M3
- ^ Q-M3=48; Q(xM3)=21.
- ^ Q-M3=86<; Q(xM3)=14.
- ^ Q=M3
- ^ Q-M3=33; Q(xM3)=48.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Венди Тымчук (2008). «Узнайте о гаплогруппе Q Y-ДНК. Учебные пособия по генной базе» . Системы генной базы. Архивировано из оригинала 22 июня 2010 года . Проверено 21 ноября 2009 г.
- ^ Оргел, Лесли Э. (2004). «Пребиотическая химия и происхождение мира РНК». Критические обзоры по биохимии и молекулярной биологии . 39 (2): 99–123. CiteSeerX 10.1.1.537.7679 . дои : 10.1080/10409230490460765 . ПМИД 15217990 . S2CID 4939632 .
- ^ Таллбер, Ким (2014). «Появление, политика и рынок ДНК коренных американцев» . В Кляйнман, Дэниел Ли; Мур, Келли (ред.). Справочник Routledge по науке, технологиям и обществу . Рутледж. п. 23. ISBN 978-1-136-23716-4 . Архивировано из оригинала 30 января 2016 года . Проверено 5 января 2016 г.
- ^ Пост, Козимо; Накацука, Натан; Лазаридис, Иосиф; Скоглунд, Понт; Маллик, Свапан; Ламнидис, Тисей К.; Роланд, Надин; Нэгеле, Кэтрин; Адамски, Николь; Бертолини, Эмили; Брумандхошбахт, Насрин; Купер, Алан; Каллетон, Брендан Дж.; Феррас, Джеймс; Ферри, Мэтью (15 ноября 2018 г.). «Реконструкция глубокой истории населения Центральной и Южной Америки » Ячейка 175 (5):1185–1197.e22. дои : 10.1016/j.cell.2018.10.027 . ISSN 0092-8674 . ПМК 6327247 . ПМИД 30415837 .
- ^ Перейти обратно: а б с Вонг, Эмили Х.М.; Хрунин Андрей; Николс, Лариса; Пушкарев Дмитрий; Хохрин Денис; Вербенко Дмитрий; Евграфов Олег; Ноулз, Джеймс; Новембре, Джон; Лимборская, Светлана; Валуев Антон (27 января 2017 г.). «Реконструкция генетической истории популяций Сибири и Северо-Восточной Европы» . Геномные исследования . 27 (1): 1–14. дои : 10.1101/гр.202945.115 . ISSN 1088-9051 . ПМК 5204334 . ПМИД 27965293 .
- ^ Перейти обратно: а б с Гилл, Хэчан; Ли, Джухён; Чон, Чунгвон (2 апреля 2024 г.). «Реконструкция генетических взаимоотношений между древними и современными популяциями Сибири» . Геномная биология и эволюция . 16 (4): evae063. дои : 10.1093/gbe/evae063 . ISSN 1759-6653 . ПМЦ 10999361 . ПМИД 38526010 .
- ^ Ян, Мелинда А. (6 января 2022 г.). «Генетическая история миграции, диверсификации и примеси в Азии» . Популяционная генетика и геномика человека . 2 (1): 1–32. дои : 10.47248/hpgg2202010001 . ISSN 2770-5005 .
- ^ Перейти обратно: а б с Морено-Майяр, Дж. Виктор; Поттер, Бен А.; Виннер, Лассе; и др. (январь 2018 г.). «Геном Аляски терминального плейстоцена показывает первую популяцию коренных американцев» (PDF) . Природа . 553 (7687): 203–207. Бибкод : 2018Natur.553..203M . дои : 10.1038/nature25173 . ПМИД 29323294 . S2CID 4454580 .
- ^ Арнаис-Виллена, А.; Парга-Лозано, К.; Морено, Э.; Аресес, К.; Рей, Д.; Гомес-Прието, П. (1 апреля 2010 г.). «Происхождение американских индейцев и заселение Америки согласно генам HLA: смешение с народами Азии и Тихого океана» . Современная геномика . 11 (2). Bentham Science Publishers Ltd.: 103–114. дои : 10.2174/138920210790886862 . ISSN 1389-2029 . ПМК 2874220 . ПМИД 20885818 .
- ^ Перейти обратно: а б с Тамм, Эрика; Кивисилд, Томас; Рейдла, Маере; и др. (5 сентября 2007 г.). «Берингийский статус и распространение коренных американцев-основателей» . ПЛОС ОДИН 2 (9):e8 Бибкод : 2007PLoSO...2..829T . дои : 10.1371/journal.pone.0000829 . ПМК 1952074 . ПМИД 17786201 .
- ^ Деренко, Мирослава; Малярчук Борис; Гжибовский, Томаш; и др. (21 декабря 2010 г.). «Происхождение и постледниковое распространение гаплогрупп C и D митохондриальной ДНК в Северной Азии» . ПЛОС ОДИН . 5 (12): e15214. Бибкод : 2010PLoSO...515214D . дои : 10.1371/journal.pone.0015214 . ПМК 3006427 . ПМИД 21203537 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р Бортолини, Мария-Катира; Сальзано, Франсиско М.; Томас, Марк Г.; и др. (сентябрь 2003 г.). «Y-хромосома свидетельствует о различиях в древней демографической истории Америки» . Американский журнал генетики человека . 73 (3): 524–539. дои : 10.1086/377588 . ПМК 1180678 . ПМИД 12900798 .
- ^ Перейти обратно: а б с Рулен, Мерритт (ноябрь 1998 г.). «Происхождение На-Дэне» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (23): 13994–13996. Бибкод : 1998PNAS...9513994R . дои : 10.1073/pnas.95.23.13994 . ПМК 25007 . ПМИД 9811914 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д Зегура, Стивен Л.; Карафет Татьяна М.; Животовский Лев А.; и др. (январь 2004 г.). «SNP высокого разрешения и микросателлитные гаплотипы указывают на единственное недавнее проникновение Y-хромосом коренных американцев в Америку» . Молекулярная биология и эволюция . 21 (1): 164–175. дои : 10.1093/molbev/msh009 . ПМИД 14595095 .
- ^ Сайяр, Джульетта; Форстер, Питер; Линнеруп, Нильс; и др. (2000). «Вариации мтДНК среди гренландских эскимосов. Край берингийской экспансии» . Американский журнал генетики человека . 67 (3): 718–726. дои : 10.1086/303038 . ПМЦ 1287530 . ПМИД 10924403 .
- ^ Шурр, Теодор Г. (21 октября 2004 г.). «Заселение Нового Света: перспективы молекулярной антропологии». Ежегодный обзор антропологии . 33 : 551–583. дои : 10.1146/annurev.anthro.33.070203.143932 .
- ^ Торрони, Антонио; Шурр, Теодор Г.; Ян, Чи-Чуань; и др. (январь 1992 г.). «Анализ митохондриальной ДНК коренных американцев показывает, что популяции америндов и наденов были основаны в результате двух независимых миграций» . Генетика . 30 (1): 153–162. дои : 10.1093/генетика/130.1.153 . ПМК 1204788 . ПМИД 1346260 .
- ^ Уэйд, Николас (12 марта 2014 г.). «В заселении континента наблюдается пауза» . Нью-Йорк Таймс . Архивировано из оригинала 9 апреля 2021 года . Проверено 24 февраля 2017 г.
- ^ Левин, Анатоль В. (1997). «Родные языки Америки» . Введение в языки мира . Оксфордский университет. п. 309. ИСБН 978-0-19-508115-2 . Архивировано из оригинала 10 мая 2021 года . Проверено 7 июня 2020 г.
- ^ Митхун, Марианна (октябрь 1990 г.). «Исследование языков североамериканских индейцев». Ежегодный обзор антропологии . 19 (1): 309–330. дои : 10.1146/annurev.an.19.100190.001521 . JSTOR 2155968 .
- ^ Элис Робертс (2010). Невероятное человеческое путешествие . А&С Черный. стр. 101–03. ISBN 978-1-4088-1091-0 . Архивировано из оригинала 25 января 2021 года . Проверено 5 августа 2019 г.
- ^ Федорова, Сардана А.; Рейдла, Маере; Мецпалу, Эне; Мецпалу, Мейт; Роотси, Сиири; Тамбетс, Кристина; Трофимова, Наталья; Жаданов Сергей Игоревич; Кашани, Бахарак Хушиар; Оливьери, Анна; Воевода Михаил Иванович; Осипова Людмила П.; Платонов Федор А.; Томский Михаил Иванович; Хуснутдинова, Эльза К. (19 июня 2013 г.). «Аутосомные и однородительские портреты коренного населения Саха (Якутии): значение для заселения Северо-Восточной Евразии» . Эволюционная биология BMC . 13 (1): 127. Бибкод : 2013BMCEE..13..127F . дои : 10.1186/1471-2148-13-127 . ISSN 1471-2148 . ПМЦ 3695835 . ПМИД 23782551 .
- ^ Перейти обратно: а б с д и Ван, Сидзя; Льюис, Сесил М. младший; Якобссон, Маттиас; и др. (23 ноября 2007 г.). «Генетическая изменчивость и структура населения коренных американцев» . ПЛОС Генетика . 3 (11): е185. дои : 10.1371/journal.pgen.0030185 . ПМК 2082466 . ПМИД 18039031 .
- ^ Перейти обратно: а б с д Уолш, Брюс; Редд, Алан Дж.; Хаммер, Майкл Ф. (январь 2008 г.). «Совместные вероятности совпадения Y-хромосомных и аутосомных маркеров». Международная судебно-медицинская экспертиза . 174 (2–3): 234–238. doi : 10.1016/j.forsciint.2007.03.014 . ПМИД 17449208 .
- ^ Перейти обратно: а б с д и Уэллс, Спенсер (2002). Путешествие человека: генетическая одиссея . Издательство Принстонского университета. стр. 138–140 . ISBN 978-0-691-11532-0 .
- ^ Перейти обратно: а б Привет, Джоди (2005). «О количестве основателей нового мира: популяционно-генетический портрет населения Америки» . ПЛОС Биология . 3 (6): e193. doi : 10.1371/journal.pbio.0030193 . ПМЦ 1131883 . ПМИД 15898833 .
- ^ Уэйд, Николас (11 февраля 2010 г.). «Геном древнего человека в Гренландии расшифрован» . Нью-Йорк Таймс . Архивировано из оригинала 8 марта 2021 года . Проверено 24 февраля 2017 г.
- ^ Перейти обратно: а б с Райх, Дэвид; Паттерсон, Ник; Кэмпбелл, Десмонд; и др. (16 августа 2012 г.). «Реконструкция истории индейского населения» . Природа . 488 (7411): 370–374. Бибкод : 2012Natur.488..370R . дои : 10.1038/nature11258 . ПМЦ 3615710 . ПМИД 22801491 .
- ^ Расмуссен, Мортен; Анзик, Сара Л.; Уотерс, Майкл Р.; и др. (2014). «Геном человека позднего плейстоцена из захоронения Кловиса в западной Монтане» . Природа . 506 (7487): 225–229. Бибкод : 2014Natur.506..225R . дои : 10.1038/nature13025 . ПМЦ 4878442 . ПМИД 24522598 .
- ^ «Составлен геном древних американцев» . Новости Би-би-си . 14 февраля 2014 года. Архивировано из оригинала 5 мая 2021 года . Проверено 20 июня 2018 г.
- ^ Дедиу, Дэн; Левинсон, Стивен К. (20 сентября 2012 г.). «Абстрактные профили структурной стабильности указывают на универсальные тенденции, семейно-специфические факторы и древние связи между языками» . ПЛОС ОДИН . 7 (9): e45198. Бибкод : 2012PLoSO...745198D . дои : 10.1371/journal.pone.0045198 . ПМЦ 3447929 . ПМИД 23028843 .
- ^ Рагхаван, Маанаса; Штайнрюкен, Матиас; Харрис, Келли; и др. (21 августа 2015 г.). «Геномные данные плейстоцена и недавней истории населения коренных американцев» . Наука . 349 (6250): ааб3884. дои : 10.1126/science.aab3884 . ПМЦ 4733658 . ПМИД 26198033 .
- ^ Скоглунд, Понт; Маллик, Свапан; Бортолини, Мария Катира; Ченнагири, Блэк; Хунемайер, Табита; Петцль-Эрлер, Мария Луиза ; Сальзано, Франсиско Мауро; Паттерсон, Ник; Райх, Дэвид (сентябрь 2015 г.). «Генетические данные двух популяций-основателей Америки» . Природа . 525 (7567): 104–108. Бибкод : 2015Natur.525..104S . дои : 10.1038/nature14895 . ПМЦ 4982469 . ПМИД 26196601 .
- ^ Перейти обратно: а б Скоглунд, Понт; Райх, Дэвид (декабрь 2016 г.). «Геномный взгляд на заселение Америки» . Текущее мнение в области генетики и развития . 41 : 27–35. дои : 10.1016/j.gde.2016.06.016 . ПМК 5161672 . ПМИД 27507099 .
- ^ Валлини и др. 2022 г. (4 июля 2022 г.). «Генетика и материальная культура поддерживают неоднократные экспансии в палеолитическую Евразию из популяционного центра из Африки» . Проверено 16 апреля 2023 г.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Ян, Мелинда А. (6 января 2022 г.). «Генетическая история миграции, диверсификации и примеси в Азии» . Популяционная генетика и геномика человека . 2 (1): 1–32. дои : 10.47248/hpgg2202010001 . ISSN 2770-5005 .
- ^ Нин, Чао; Фернандес, Даниэль; Чангмай, Пия; Флегонтова, Ольга; Юнджю, Эрен; Майер, Роберт; Алтынышик, Н. Эзги; Кассиан Алексей С.; Краузе, Йоханнес (12 октября 2020 г.), Геномное формирование первых американских предков в Восточной и Северо-Восточной Азии , номер doi : 10.1101/2020.10.12.336628 , получено 29 марта 2024 г.
- ^ ФЕРРАС, Тьяго; Суарес Вильягран, Химена; Нэгеле, Катрин; Радзявичюте, Рита; Барбоса Лемес, Ренан; Саласар-Гарсия, Доминго К.; Весоловский, Вероника; Лопес Алвес, Маркони; БАСТОС, Мурило; Рэпп Пи-Дэниел, Энн; ПИНТО ЛИМА, Хелена; Мендес Кардосо, Джессика; ЭСТЕВАМ, Рената; Лирио, Андерсен; Гимарайнш, Геован М. (август 2023 г.). «Генетическая история прибрежных обществ восточной части Южной Америки» . Экология и эволюция природы . 7 (8): 1315–1330. Бибкод : 2023NatEE...7.1315F . дои : 10.1038/s41559-023-02114-9 . ISSN 2397-334X . ПМЦ 10406606 . ПМИД 37524799 .
- ^ Дэвис, Лорен Г.; Мэдсен, Дэвид Б.; Бесерра-Вальдивия, Лорена; и др. (30 августа 2019 г.). «Заселение позднего верхнего палеолита в Куперс-Ферри, Айдахо, США, ~ 16 000 лет назад» . Наука . 365 (6456): 891–897. Бибкод : 2019Sci...365..891D . doi : 10.1126/science.aax9830 . ПМИД 31467216 .
- ^ Перейти обратно: а б Пост, Козимо; Голый, Натан; Лазаридис, Джозеф; и др. (ноябрь 2018 г.). «Реконструкция глубокой истории населения Центральной и Южной Америки» . Клетка . 175 (5): 1185–1197.e22. дои : 10.1016/j.cell.2018.10.027 . ПМК 6327247 . ПМИД 30415837 .
- ^ Майер, Роберт; Флегонтов Павел; Флегонтова, Ольга; Ишилдак, Улаш; Чангмай, Пия; Райх, Дэвид (14 апреля 2023 г.). Нордборг, Магнус; Пшеворски, Молли; Болдинг, Дэвид; Виуф, Карстен (ред.). «О пределах приспособления сложных моделей истории населения к f-статистике» . электронная жизнь . 12 : е85492. doi : 10.7554/eLife.85492 . ISSN 2050-084X . ПМЦ 10310323 . ПМИД 37057893 .
- ^ Перейти обратно: а б Виллерслев, Эске; Мельцер, Дэвид Дж. (17 июня 2021 г.). «Заселение Америки по данным древней геномики». Природа . 594 (7863): 356–364. Бибкод : 2021Natur.594..356W . дои : 10.1038/s41586-021-03499-y . ПМИД 34135521 . S2CID 235460793 .
- ^ Перейти обратно: а б Саркар, Анджали А. (18 июня 2021 г.). «Геномы древнего человека раскрывают заселение Америки» . GEN – Новости генной инженерии и биотехнологии . Проверено 15 сентября 2021 г.
Команда обнаружила, что останки в Пещере Духов принадлежат коренным американцам, отвергнув при этом давнюю теорию о том, что группа под названием палеоамериканцы существовала в Северной Америке до коренных американцев.
- ^ Сапиенс (8 февраля 2022 г.). «Генетическая хроника первых народов Америки» . САПИЕНС . Проверено 3 апреля 2023 г.
ОКОЛО 36 000 лет назад небольшая группа людей, живших в Восточной Азии, начала отделяться от более крупных древних популяций региона. Примерно 25 000 лет назад меньшая группа в самой Восточной Азии разделилась на две части. Один дал начало группе, которую генетики называют древними палеосибирями, оставшимися в Северо-Восточной Азии. Другой стал предком коренных народов Америки.
- ^ Валлини, Леонардо; Зампиери, Карло; Шоаи, Мохамед Джавад; Бортолини, Эухенио; Марчиани, Джулия; Анели, Серена; Пиевани, Тельмо; Бенацци, Стефано; Бараус, Альберто; Меззавилла, Массимо; Петралья, Майкл Д.; Пагани, Лука (25 марта 2024 г.). «Персидское плато служило центром расселения Homo sapiens после основного расселения из Африки» . Природные коммуникации . 15 (1): 1882. Бибкод : 2024NatCo..15.1882V . дои : 10.1038/s41467-024-46161-7 . ISSN 2041-1723 . ПМЦ 10963722 . ПМИД 38528002 .
- ^ Сантос, Фабрисио Р.; Пандия, Арпита; Тайлер-Смит, Крис; и др. (февраль 1999 г.). «Центрально-сибирское происхождение Y-хромосом коренных американцев» . Американский журнал генетики человека . 64 (2): 619–628. дои : 10.1086/302242 . ПМЦ 1377773 . ПМИД 9973301 .
- ^ Белая Верея; Алехандро Хосе. Линии Y-хромосомы человека: популяционно-генетические и судебно-медицинские применения . Университет Сантьяго де Компостела. стр. 135–. GGKEY:JCW0ASCR364. Архивировано из оригинала 30 января 2016 года . Проверено 15 июня 2011 г.
- ^ Зегура, Стивен Л.; Карафет Татьяна М.; Животовский Лев А.; и др. (январь 2004 г.). «SNP высокого разрешения и микросателлитные гаплотипы указывают на единственное недавнее проникновение Y-хромосом коренных американцев в Америку» . Молекулярная биология и эволюция . 21 (1): 164–175. дои : 10.1093/molbev/msh009 . ПМИД 14595095 .
- ^ Перейти обратно: а б с Сайяр, Джульетта; Форстер, Питер; Линнеруп, Нильс; и др. (сентябрь 2000 г.). «Вариации мтДНК среди гренландских эскимосов: край берингийской экспансии» . Американский журнал генетики человека . 67 (3): 718–726. дои : 10.1086/303038 . ПМЦ 1287530 . ПМИД 10924403 .
- ^ Шурр, Теодор Г. (2004). «Население Нового Света - перспективы молекулярной антропологии». Ежегодный обзор антропологии . 33 : 551–583. дои : 10.1146/annurev.anthro.33.070203.143932 . JSTOR 25064865 . S2CID 4647888 .
- ^ Торрони, Антонио; Шурр, Теодор Г.; Ян, Чи-Чуань; и др. (январь 1992 г.). «Анализ митохондриальной ДНК коренных американцев показывает, что популяции америндов и наденов были основаны в результате двух независимых миграций» . Генетика . 130 (1): 153–162. дои : 10.1093/генетика/130.1.153 . ПМК 1204788 . ПМИД 1346260 .
- ^ Балановский Олег; Гурьянов Владимир; Запорожченко Валерий; и др. (февраль 2017 г.). «Филогеография гаплогруппы Q3-L275 Y-хромосомы человека на основе научного и научного сотрудничества» . Эволюционная биология BMC . 17 (S1): 18. Бибкод : 2017BMCEE..17S..18B . дои : 10.1186/s12862-016-0870-2 . ПМЦ 5333174 . ПМИД 28251872 .
- ^ Перейти обратно: а б «Как определяются субклады гаплогруппы Q Y-ДНК» . Системы генной базы. 2009. Архивировано из оригинала 21 января 2010 года . Проверено 22 ноября 2009 г.
- ^ «Дерево гаплогрупп Y-ДНК» . Системы генной базы. 2009. Архивировано из оригинала 25 декабря 2010 года . Проверено 21 ноября 2009 г.
- ^ Перейти обратно: а б с «Частотное распределение гаплогруппы Y-ДНК Q1a3a-M3» . ДжинТри. 2010. Архивировано из оригинала 4 ноября 2009 года . Проверено 30 января 2010 г.
- ^ Флегонтов Павел; Чангмай, Пия; Зидкова Анастасия; Логачева Мария Дмитриевна; Алтынышик, Н. Эзги; Флегонтова, Ольга; Гельфанд Михаил С.; Герасимов Евгений С.; Храмеева Екатерина Евгеньевна; Коновалова Ольга П.; Неретина, Татьяна; Никольский Юрий В.; Старостин, Георгий; Степанова Вита В.; Травинский Игорь Владимирович; Тршиска, Мартин; Тршиска, Питер; Татаринова, Татьяна В. (2016). «Геномное исследование кетов: палео-эскимосской этнической группы со значительным древним североевразийским происхождением» . Научные отчеты . 6 : 20768. arXiv : 1508.03097 . Бибкод : 2016NatSR...620768F . дои : 10.1038/srep20768 . ПМЦ 4750364 . ПМИД 26865217 .
- ^ «Учебный центр :: Учебные пособия по генной базе» . Genebase.com. 22 октября 1964 года. Архивировано из оригинала 17 ноября 2013 года.
- ^ Бонатто, СЛ; Сальзано, FM (4 марта 1997 г.). «Единичная и ранняя миграция населения Америки, подтвержденная данными о последовательностях митохондриальной ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (5): 1866–1871. Бибкод : 1997PNAS...94.1866B . дои : 10.1073/pnas.94.5.1866 . ПМК 20009 . ПМИД 9050871 .
- ^ Смоленяк, Меган; Тернер, Энн (2004). Отследите свои корни с помощью ДНК: использование генетических тестов для изучения вашего генеалогического древа . Родейл. п. 83. ИСБН 978-1-59486-006-5 . Архивировано из оригинала 30 января 2016 года . Проверено 5 января 2016 г.
- ^ Тан, Кер (14 февраля 2008 г.). «Поселенцы Нового Света сделали пит-стоп на 20 000 лет» . Национальные географические новости . Архивировано из оригинала 26 августа 2014 года . Проверено 24 января 2010 г.
- ^ Ловгрен, Стефан (2 февраля 2007 г.). «Первые американцы прибыли недавно и заселили Тихоокеанское побережье, говорится в исследовании ДНК» . Национальные географические новости . Архивировано из оригинала 22 августа 2009 года . Проверено 2 февраля 2010 г.
- ^ «Первые американцы пережили 20 000-летнюю остановку - Дженнифер Виегас, Discovery News» . 10 мая 2017. Архивировано из оригинала 10 октября 2012 года . Проверено 18 ноября 2009 г. страница 2. Архивировано 13 марта 2012 г. в Wayback Machine.
- ^ Гонсалес Бурхард, Эстебан; Боррелл, Луиза Н.; Чоудри, Света; и др. (декабрь 2005 г.). «Латиноамериканское население: уникальная возможность изучения расы, генетики и социальной среды в эпидемиологических исследованиях» . Американский журнал общественного здравоохранения . 95 (12): 2161–2168. дои : 10.2105/AJPH.2005.068668 . ПМЦ 1449501 . ПМИД 16257940 .
- ^ Перейти обратно: а б с д и ж г час я Малхи, Рипан Сингх; Гонсалес-Оливер, Анжелика; Шредер, Кэри Бритт; и др. (декабрь 2008 г.). «Распределение Y-хромосом среди коренных жителей Северной Америки: исследование истории населения Атапаски» . Американский журнал физической антропологии . 137 (4): 412–424. дои : 10.1002/ajpa.20883 . ПМК 2584155 . ПМИД 18618732 .
- ^ Болник, Дебора; Больник, Дэниел; Смит, Дэвид (2006). «Асимметричные мужские и женские генетические истории среди коренных американцев восточной части Северной Америки» . Молекулярная биология и эволюция . 23 (11): 2161–2174. дои : 10.1093/molbev/msl088 . ПМИД 16916941 . «В большинстве случаев существует широко распространенное согласие относительно того, представляет ли конкретная гаплогруппа древнюю индейскую линию или примесь после 1492 года, но статус гаплогруппы R-M173 недавно стал предметом некоторых дискуссий. Некоторые авторы утверждают, что эта гаплогруппа представляет собой основополагающая линия коренных американцев (Лелл и др., 2002; Бортолини и др., 2003), тогда как другие предполагают, что она вместо этого отражает недавнюю европейскую примесь (Тарасона-Сантос и Сантос, 2002; Бош и др., 2003; Зегура и др., 2004). В восточной части Северной Америки характер вариаций гаплотипов внутри этой гаплогруппы подтверждает последнюю гипотезу: гаплотипы R-M173 не группируются по популяциям или культурным территориям, как это делают гаплотипы в других гаплогруппах-основателях, и большинство из них соответствуют или тесно связаны с R- Гаплотипы M173, которые распространены в Европе, но редки в Азии. Эта закономерность противоположна ожидаемой, если гаплотипы R-M173 коренных американцев произошли от азиатских гаплотипов, и предполагает, что недавняя европейская примесь ответственна за присутствие гаплогруппы R-M173 на востоке Севера. Америка."
- ^ Болник, Дебора Энн (2005). Генетическая предыстория восточной части Северной Америки: данные древней и современной ДНК . Калифорнийский университет в Дэвисе. п. 83. «Гаплогруппа R - M173, вероятно, представляет собой недавнюю (после 1492 г.) европейскую примесь, как и P-M45* (Tarazona-Santos and Santos 2002; Bosch et al. 2003; Zegura et al. 2004)»
- ^ Raff 2022 , стр. 59–60: «Гаплогруппы-основатели Y-хромосомы у коренных американцев включают Q-M3 (и его субгаплогруппы, Q-CTS1780) и C3-MPB373 (потенциально C-P39-Z30536). Обнаружены и другие гаплогруппы. [sic] Популяции коренных американцев, такие как R1b, вероятно, возникли в результате постевропейской контактной примеси (44)».
- ^ Малхи и др. 2008 : «Все особи, которые не принадлежали к гаплогруппам Q и C, были исключены из набора данных гаплотипов, поскольку эти гаплотипы, вероятно, являются результатом примеси неместного происхождения (Tarazona-Santos and Santos, 2002; Zegura et al., 2004; Bolnick и др., 2006)...Частота гаплогруппы C наиболее высока на северо-западе Северной Америки, а частота гаплогруппы R, присутствие которой приписывается европейской примеси, достигает максимума на северо-востоке Северной Америки».
- ^ Зегура, С.Л. (31 октября 2003 г.). «SNP высокого разрешения и микросателлитные гаплотипы указывают на единственное недавнее проникновение Y-хромосом коренных американцев в Америку» . Молекулярная биология и эволюция . 21 (1). Издательство Оксфордского университета (OUP): 164–175. дои : 10.1093/molbev/msh009 . ISSN 0737-4038 . ПМИД 14595095 .
- ^ Чжун, Хуа; Ши, Хун; Ци, Сюэ-Бин; и др. (июль 2010 г.). «Глобальное распространение гаплогруппы C Y-хромосомы раскрывает доисторические маршруты миграции африканцев и раннего расселения в Восточной Азии» . Журнал генетики человека . 55 (7): 428–435. дои : 10.1038/jhg.2010.40 . ПМИД 20448651 . S2CID 28609578 .
- ^ Сюэ, Яли; Зержал, Татьяна; Бао, Вэйдун; и др. (1 апреля 2006 г.). «Мужская демография в Восточной Азии: контраст между Севером и Югом во времена роста численности населения» . Генетика . 172 (4): 2431–2439. doi : 10.1534/genetics.105.054270 . ПМЦ 1456369 . ПМИД 16489223 .
- ^ Маринакис, Алики (2008). «Люди На-Дэне» . В Йохансене, Брюс Э.; Прицкер, Барри М. (ред.). Энциклопедия истории американских индейцев . АВС-КЛИО. п. 441. ИСБН 978-1-85109-818-7 . Архивировано из оригинала 30 января 2016 года . Проверено 5 января 2016 г.
- ^ Перейти обратно: а б ISOGG, 2015 «Гаплогруппа C Y-ДНК и ее субклады - 2015». Архивировано 15 августа 2021 г. в Wayback Machine (15 сентября 2015 г.).
- ^ Перейти обратно: а б Зегура, С.Л. (31 октября 2003 г.). «SNP высокого разрешения и микросателлитные гаплотипы указывают на единственное недавнее проникновение Y-хромосом коренных американцев в Америку» . Молекулярная биология и эволюция . 21 (1): 164–175. дои : 10.1093/molbev/msh009 . ПМИД 14595095 .
- ^ Перейти обратно: а б с д Больник, Д.А. (10 августа 2006 г.). «Асимметричные мужские и женские генетические истории среди коренных американцев восточной части Северной Америки». Молекулярная биология и эволюция . 23 (11): 2161–2174. дои : 10.1093/molbev/msl088 . ПМИД 16916941 . S2CID 30220691 .
- ^ Перейти обратно: а б с Дулик, MC; Оуингс, AC; Гайески, Дж.Б.; и др. (14 мая 2012 г.). «Анализ Y-хромосомы выявляет генетическое расхождение и появление новых коренных родословных в популяциях, говорящих на атапасском и эскимосском языках» . Труды Национальной академии наук . 109 (22): 8471–8476. Бибкод : 2012PNAS..109.8471D . дои : 10.1073/pnas.1118760109 . ПМК 3365193 . ПМИД 22586127 .
- ^ Хаммер, Майкл Ф.; Чемберлен, Вероника Ф.; Кирни, Вероника Ф.; и др. (декабрь 2006 г.). «Популяционная структура гаплогрупп SNP Y-хромосомы в Соединенных Штатах и судебно-медицинская экспертиза для создания баз данных STR Y-хромосомы». Международная судебно-медицинская экспертиза . 164 (1): 45–55. doi : 10.1016/j.forsciint.2005.11.013 . ПМИД 16337103 .
- ^ Перейти обратно: а б Шурр, Теодор Г. (май – июнь 2000 г.). «Митохондриальная ДНК и заселение Нового Света». Американский учёный . 88 (3): 246. дои : 10.1511/2000.23.772 .
- ^ Захаров Илья А.; Деренко, Мирослава В.; Малярчук Борис А.; и др. (апрель 2004 г.). «Вариации митохондриальной ДНК аборигенных популяций Алтае-Байкалья: значение для генетической истории Северной Азии и Америки». Анналы Нью-Йоркской академии наук . 1011 (1): 21–35. Бибкод : 2004JAN1011...21Z . дои : 10.1196/анналы.1293.003 . ПМИД 15126280 . S2CID 37139929 .
- ^ Стариковская Елена Борисовна; Сукерник, Рем И.; Дербенева Ольга Александровна; и др. (7 января 2005 г.). «Разнообразие митохондриальной ДНК у коренных народов южной части Сибири и происхождение гаплогрупп коренных американцев» . Анналы генетики человека . 69 (1): 67–89. дои : 10.1046/j.1529-8817.2003.00127.x . ПМЦ 3905771 . ПМИД 15638829 .
- ^ Фелтон, Джеймс (9 мая 2023 г.). «ДНК проливает свет на тайну происхождения коренных американцев» . IFLНаука . Проверено 24 мая 2023 г.
- ^ Перейти обратно: а б с Эшлеман, Джейсон А.; Малхи, Рипан С.; Смит, Дэвид Гленн (14 февраля 2003 г.). «Исследования митохондриальной ДНК коренных американцев - концепции и заблуждения о предыстории населения Америки». Эволюционная антропология . 12 (1): 7–18. дои : 10.1002/evan.10048 . S2CID 17049337 .
- ^ Перейти обратно: а б с Ахилли, Алессандро; Перего, Уго А.; Брави, Клаудио М.; и др. (12 марта 2008 г.). «Филогения четырех панамериканских гаплогрупп мтДНК: значение для исследований эволюции и болезней» . ПЛОС ОДИН . 3 (3): e1764. Бибкод : 2008PLoSO...3.1764A . дои : 10.1371/journal.pone.0001764 . ПМК 2258150 . ПМИД 18335039 .
- ^ Нина Г. Яблонски (2002). Первые американцы: плейстоценовая колонизация Нового Света . Издательство Калифорнийского университета. п. 301. ИСБН 978-0-940228-50-4 . Архивировано из оригинала 30 января 2016 года . Проверено 15 июня 2011 г.
- ^ Дай волю, Роберто; Баттонс, Патриция; Джардина, Бруно (8 марта 2012 г.). Достижения митохондриальной медицины . Springer Science & Business Media. п. 446. ИСБН 978-94-007-2869-1 .
- ^ Рафф, Дженнифер А.; Больник, Дебора А. (октябрь 2015 г.). «Указывает ли митохондриальная гаплогруппа X на древнюю трансатлантическую миграцию в Америку? Критическая переоценка» . ПалеоАмерика . 1 (4): 297–304. дои : 10.1179/2055556315Z.00000000040 . ISSN 2055-5563 . S2CID 85626735 . «Все эти исследования пришли к одному и тому же выводу и предполагают, что гаплогруппа X2a, вероятно, возникла в той же популяции (популяциях), что и другие гаплогруппы-основатели Америки, в силу сопоставимых дат слияния и демографической истории» ... «X2a имеет сопоставимые даты слияния и демографическую историю». не был обнаружен нигде в Евразии, и филогеография не дает нам убедительных оснований полагать, что он, скорее, пришел из Европы, чем из Сибири. Более того, анализ полного генома кенневикского человека, который принадлежит к самой базовой линии X2a, когда-либо идентифицированной. , не дает никаких указаний на недавнее европейское происхождение и перемещает расположение самой глубокой ветви X2a на западное побережье, что соответствует принадлежности X2a к той же наследственной популяции, что и другие митохондриальные гаплогруппы-основатели».
- ^ «Население Америки: генетическое происхождение влияет на здоровье» . Научный американец . 14 августа 2009 года. Архивировано из оригинала 14 сентября 2011 года . Проверено 19 января 2010 г. - через Phys.org.
- ^ Фагундес, Нельсон-младший; Каниц, Рикардо; Эккерт, Роберта; и др. (3 марта 2008 г.). «Геномика митохондриальных популяций подтверждает единое происхождение до Кловиса с прибрежным маршрутом заселения Америки» . Американский журнал генетики человека . 82 (3): 583–592. дои : 10.1016/j.ajhg.2007.11.013 . ПМЦ 2427228 . ПМИД 18313026 .
- ^ Мельцер, Дэвид Дж. (2009). Первые народы в новом мире: колонизация Америки ледникового периода . Издательство Калифорнийского университета. п. 162. ИСБН 978-0-520-94315-5 . Архивировано из оригинала 30 января 2016 года . Проверено 5 января 2016 г.
- ^ «МтДНК-взгляд на заселение мира Homo Sapiens» . Кембриджская ДНК. 2009. Архивировано из оригинала 11 мая 2011 года . Проверено 29 апреля 2010 г.
- ^ Рейдла, Маере; Кивисилд, Тоомас; Мецпалу, Эне; и др. (ноябрь 2003 г.). «Происхождение и распространение гаплогруппы X мтДНК» . Американский журнал генетики человека . 73 (5): 1178–1190. дои : 10.1086/379380 . ПМЦ 1180497 . ПМИД 14574647 .
- ^ «МтДНК-взгляд на заселение мира Homo Sapiens» . Кембриджская служба ДНК. 2007. Архивировано из оригинала 11 мая 2011 года . Проверено 1 июня 2011 г.
- ^ Пеньялоса-Эспиноза, Розенда И.; Сэндс-Аранда, Диего; Бристл-Флауэрс, Ричард М.; и др. (2007). «Характеристика гаплогрупп мтДНК у 14 коренных народов Мексики». человека Биология 79 (3): 313–320. дои : 10.1353/hub.2007.0042 . ПМИД 18078204 . S2CID 35654242 .
- ^ Перейти обратно: а б Феррелл, Роберт Э.; Чакраборти, Ранаджит; Гершовиц, Генри; и др. (июль 1981 г.). «Эскимосы острова Святого Лаврентия: генетическая изменчивость и генетическая дистанция». Американский журнал физической антропологии . 55 (3): 351–358. дои : 10.1002/ajpa.1330550309 . ПМИД 6455922 .
- ^ Перейти обратно: а б Хельгасон, Агнар; Палссон, Гисли; Педерсен, Хеннинг Ленивец; и др. (май 2006 г.). «Вариации мтДНК в популяциях инуитов Гренландии и Канады: история миграции и структура населения». Американский журнал физической антропологии . 130 (1): 123–134. дои : 10.1002/ajpa.20313 . ПМИД 16353217 .
- ^ Рагхаван, Маанаса; Скоглунд, Понт; Граф, Келли Э.; и др. (январь 2014 г.). «Геном Сибири верхнего палеолита свидетельствует о двойном происхождении коренных американцев» . Природа . 505 (7481): 87–91. Бибкод : 2014Natur.505...87R . дои : 10.1038/nature12736 . ПМК 4105016 . ПМИД 24256729 .
- ^ Кашани, Бахарак Хушиар; Перего, Уго А.; Оливьери, Анна; и др. (январь 2012 г.). «Митохондриальная гаплогруппа C4c: редкая линия, попадающая в Америку через незамерзающий коридор?». Американский журнал физической антропологии . 147 (1): 35–39. дои : 10.1002/ajpa.21614 . ПМИД 22024980 .
- ^ Ламас, Бастьен; Ферен-Шмитц, Ларс; Вальверде, Гвидо; и др. (29 апреля 2016 г.). «Древняя митохондриальная ДНК обеспечивает временную шкалу заселения Америки с высоким разрешением» . Достижения науки . 2 (4): e1501385. Бибкод : 2016SciA....2E1385L . дои : 10.1126/sciadv.1501385 . ПМЦ 4820370 . ПМИД 27051878 .
- ^ Перейти обратно: а б с «Прямые генетические данные об основателе населения раскрывают историю первых коренных американцев» . Кембриджский университет . 3 января 2018 г. Архивировано из оригинала 25 сентября 2018 г. Проверено 15 октября 2018 г.
- ^ «Прямые генетические данные об основателе населения раскрывают историю первых коренных американцев» . Архивировано из оригинала 5 ноября 2018 года . Проверено 15 октября 2018 г.
- ^ Гарсиа-Бур, Жауме; Перес-Перес, Александр; Альварес, Сара; и др. (2004). «Ранняя дифференциация населения вымерших аборигенов Огненной Земли-Патагонии: древние последовательности мтДНК и характеристика STR Y-хромосомы». Американский журнал физической антропологии . 123 (4): 361–370. дои : 10.1002/ajpa.10337 . ПМИД 15022364 .
- ^ Невес, Вашингтон; Пауэлл, Дж. Ф.; Озолиньш, Э.Г. (1999). «Внеконтинентальное морфологическое сходство Палли Айке, юг Чили» . Интерсиенсия . 24 (4): 258–263. Архивировано из оригинала 9 июля 2021 года . Проверено 8 июля 2021 г.
- ^ Бонатто, Сандро Л.; Сальзано, Франсиско М. (4 марта 1997 г.). «Единичная и ранняя миграция населения Америки, подтвержденная данными о последовательностях митохондриальной ДНК» . Труды Национальной академии наук . 94 (5): 1866–1871. Бибкод : 1997PNAS...94.1866B . дои : 10.1073/pnas.94.5.1866 . ПМК 20009 . ПМИД 9050871 .
- ^ Синк-Марс, Дж. (1979). «Пещера луфаря I: отложения восточно-берингийской пещеры позднего плейстоцена в Северном Юконе». Канадский журнал археологии (3): 1–32. JSTOR 41102194 .
- ^ Боннихсен, Робсон (1978). «Критические аргументы в пользу плейстоценовых артефактов из бассейна Олд-Кроу, Юкон: предварительное заявление». В Брайане, Алан Лайл (ред.). Ранний человек в Америке с точки зрения Тихоокеанского региона . Международные археологические исследования. стр. 102–118. ISBN 978-0-88864-999-7 .
- ^ Диллехей, Том Д.; Окампо, Карлос (2015). «Новые археологические свидетельства раннего присутствия человека в Монте-Верде, Чили» . ПЛОС ОДИН . 10 (11): e0141923. Бибкод : 2015PLoSO..1041923D . дои : 10.1371/journal.pone.0141923 . ПМЦ 4651426 . ПМИД 26580202 .
- ^ Буржон, Лориан; Берк, Ариана; Хайэм, Томас (6 января 2017 г.). «Самое раннее присутствие человека в Северной Америке, датированное последним ледниковым максимумом: новые радиоуглеродные даты из пещер Блюфиш, Канада» . ПЛОС ОДИН . 12 (1): e0169486. Бибкод : 2017PLoSO..1269486B . дои : 10.1371/journal.pone.0169486 . ПМЦ 5218561 . ПМИД 28060931 .
- ^ Гилберт, М. Томас П.; Дженкинс, Деннис Л.; Гётерстрем, Андерс; и др. (9 мая 2008 г.). «ДНК человеческих копролитов до Кловиса в Орегоне, Северная Америка» . Наука . 320 (5877): 786–789. Бибкод : 2008Sci...320..786G . дои : 10.1126/science.1154116 . ПМИД 18388261 . S2CID 17671309 .
- ^ Ньюман, Ричард (1999). «Смешанные браки». В Кваме Аппиа и Генри Луи Гейтсе-младшем (ред.). Африкана: Энциклопедия африканского и афроамериканского опыта (1-е изд.). Нью-Йорк: Основные книги Civitas. п. 1320 . ISBN 978-0-465-00071-5 .
Смешение рас, термин, обозначающий сексуальные отношения между расами; больше не используется из-за расистского подтекста
- ^ Честин, Джон Чарльз; Вуд, Джеймс А. (2004). Проблемы современной латиноамериканской истории, источники и интерпретации . Сэр Книги. стр. 4–10. ISBN 978-0-8420-5060-9 . Архивировано из оригинала 15 августа 2021 года . Проверено 24 февраля 2010 г.
- ^ Честин (2004:4): «между белой элитой и массой [коренных американцев] и негров к 1700 году существовала тонкая прослойка населения, не подлежащая ни негритянскому рабству, ни опеке [коренных американцев], состоящая из продуктов расовой скрещивание белых, [коренных американцев] и негров и определяемое как метисы, мулаты и замбо (смесь [коренных американцев] и негров) и их многочисленные комбинации».
- ^ Санс, Моника (2000). «Исследования примесей в Латинской Америке: с 20 по 21 век». Биология человека . 72 (1): 155–177. JSTOR 41465813 . ПМИД 10721616 .
- ^ Сладкий, Фрэнк В. (2004). «Афро-европейская генетическая смесь в Соединенных Штатах» . Очерки о цветовой линии и правиле одной капли . Очерки Бакинтайма. Архивировано из оригинала 4 января 2010 года . Проверено 24 февраля 2010 г.
- ^ Сладкий, Фрэнк В. (2005). Юридическая история цветной линии: понятие невидимой черноты . Издательство Бакинтайм. п. 542. ИСБН 978-0-939479-23-8 . Архивировано из оригинала 17 февраля 2010 года . Проверено 24 февраля 2010 г.
- ^ Сунг, Роланд (1999). «Расовые классификации в Латинской Америке» . Латиноамериканские СМИ. Архивировано из оригинала 30 апреля 2010 года . Проверено 25 февраля 2010 г.
- ^ Брео, Гарри Б. (1998). «Телега и тропы Красной реки: Торговля мехом» . Историческое общество Манитобы. Архивировано из оригинала 9 июля 2011 года . Проверено 24 февраля 2010 г.
- ^ «Кто такие метисы?» . Национальный совет метисов . 2001. Архивировано из оригинала 26 февраля 2010 года . Проверено 24 февраля 2010 г.
- ^ Перейти обратно: а б Кимберли ТаллБеар (2003). «ДНК, кровь и расовая принадлежность племени». Обзор Wíčazo Ša . 18 (1). Издательство Университета Миннесоты: 81–107. дои : 10.1353/wic.2003.0008 . JSTOR 140943 . S2CID 201778441 .
- ^ Фурукава, Джулия (22 мая 2023 г.). «Обзор генеалогий и других записей не подтверждает заявления местных лидеров о происхождении абенаков» . Общественное радио Нью-Гэмпшира . Проверено 7 июля 2023 г.
- ^ Адамс, Пол (10 июля 2011 г.). «Количество крови влияет на идентичность коренных американцев» . Новости Би-би-си . Архивировано из оригинала 29 января 2023 года . Проверено 22 октября 2017 г.
- ^ Бюро переписи населения США. «Сайт переписи населения США» . Бюро переписи населения США . Архивировано из оригинала 27 декабря 1996 года . Проверено 22 октября 2017 г.
- ^ «Почему так много людей утверждают, что у них в крови есть чероки? - Нерв» . www.nerve.com . Архивировано из оригинала 22 октября 2017 года . Проверено 22 октября 2017 г.
- ^ Смитерс, Грегори Д. (1 октября 2015 г.). «Почему так много американцев думают, что у них есть кровь чероки?» . Сланец . Архивировано из оригинала 5 октября 2018 года . Проверено 22 октября 2017 г.
- ^ «Синдром чероки — Daily Yonder» . www.dailyyonder.com . 10 февраля 2011 года. Архивировано из оригинала 21 октября 2017 года . Проверено 22 октября 2017 г.
- ^ Перейти обратно: а б Хитт, Джек (21 августа 2005 г.). «Новые индейцы» . Нью-Йорк Таймс . Архивировано из оригинала 21 октября 2017 года . Проверено 22 октября 2017 г. - через www.nytimes.com.
- ^ Ньевес, Эвелин (3 марта 2007 г.). «Вынесение на голосование вопроса: «Кто такой чероки?» " . Нью-Йорк Таймс . Архивировано из оригинала 30 августа 2017 года . Проверено 22 октября 2017 г. - через www.nytimes.com. .
- ^ «На какой процент индейца нужно быть, чтобы быть членом племени или нации? - Сеть средств массовой информации страны Индии» . indiancountrymedianetwork.com . Архивировано из оригинала 21 октября 2017 года . Проверено 22 октября 2017 г. . «Исчезающие индейцы, часть II: расовое лицемерие при принятии решения о зачислении - Сеть средств массовой информации страны Индии» . indiancountrymedianetwork.com . Архивировано из оригинала 22 сентября 2017 года . Проверено 22 октября 2017 г.
- ^ Перейти обратно: а б Прайс, Майкл (15 ноября 2016 г.). «Европейские болезни оставили генетический след у коренных американцев». Наука . дои : 10.1126/science.aal0382 .
- ^ Херши А., Чейз М. (1952). «Независимые функции вирусного белка и нуклеиновой кислоты в росте бактериофага» . J Gen Physiol . 36 (1): 39–56. дои : 10.1085/jgp.36.1.39 . ПМК 2147348 . ПМИД 12981234 .
- ^ Перейти обратно: а б Лилль-Шишкович I (1957). «Развитие исследований плеяд групп крови». Достижения гигиены и экспериментальной медицины (на польском языке). 11 (3): 229–33. OCLC 101713985 . ПМИД 13505351 .
- ^ Ландштейнер, Карл (1900). «Знание об антиферментативном, литическом и агглютинирующем действии сыворотки крови и лимфы». Центральный журнал бактериологии (на немецком языке). 27 :357-362. OCLC 11337636 . НАИД 10008616088 .
- ^ Язер М., Олссон М., Палчич М. (2006). «Фенотип группы крови цис-AB: фундаментальные уроки гликобиологии». Трансфус Мед Рев . 20 (3): 207–17. дои : 10.1016/j.tmrv.2006.03.002 . ПМИД 16787828 .
- ^ Перейти обратно: а б с «Распределение групп крови» . Факультет поведенческих наук Паломарского колледжа. 2010. Архивировано из оригинала 21 февраля 2006 года . Проверено 14 марта 2010 г.
- ^ Эстрада-Мена Б., Эстрада Ф.Дж., Уллоа-Арвизу Р. и др. (май 2010 г.). «Аллели группы крови O у коренных американцев: значение для заселения Америки» . Являюсь. Дж. Физ. Антрополь . 142 (1): 85–94. дои : 10.1002/ajpa.21204 . ПМИД 19862808 .
- ^ Чоун, Брюс; Льюис, Мэрион (июнь 1956 г.). «Гены группы крови индейцев кри и эскимосов района Унгава в Канаде». Американский журнал физической антропологии . 14 (2): 215–224. дои : 10.1002/ajpa.1330140217 . ПМИД 13362488 .
- ^ Перейти обратно: а б Халверсон, Мелисса С.; Больник, Дебора А. (ноябрь 2008 г.). «Древний ДНК-тест на эффект основателя частот групп крови ABO коренных американцев». Американский журнал физической антропологии . 137 (3): 342–347. дои : 10.1002/ajpa.20887 . ПМИД 18618657 .
- ^ «Расовое и этническое распределение групп крови АВО» . Кровавая Книга. 2008. Архивировано из оригинала 4 марта 2010 года . Проверено 31 марта 2010 г.
- ^ Пул Дж (2020). «Система групп крови Диего — обновление» . Иммуногематология . 15 (4): 135–43. doi : 10.21307/иммуногематология-2019-635 . ПМИД 15373634 .
- ^ Бега С., Байи П., Кьярони Ж., Мазьер С. (6 июля 2015 г.). «Возврат к системе групп крови Диего у американских индейцев: доказательства миграции генной культуры» . ПЛОС ОДИН . 10 (7): e0132211. Бибкод : 2015PLoSO..1032211B . дои : 10.1371/journal.pone.0132211 . ПМК 4493026 . ПМИД 26148209 .
Дальнейшее чтение
[ редактировать ]- Питер Н. Джонс (октябрь 2002 г.). мтДНК американских индейцев, генетические данные Y-хромосомы . Институт Исхода. ISBN 978-0-9721349-1-0 .
- Джозеф Фредерик Пауэлл (2005). Первые американцы: раса, эволюция и происхождение коренных американцев . Издательство Кембриджского университета. ISBN 978-0-521-82350-0 . [ постоянная мертвая ссылка ]
- Франсиско М. Сальзано; Мария Катира Бортолини (2002). Эволюция и генетика популяций Латинской Америки . Издательство Кембриджского университета. ISBN 978-0-521-65275-9 . [ постоянная мертвая ссылка ]
- «Население Америки: генетическое происхождение влияет на здоровье» . Американский журнал физической антропологии . Университет Оклахомы. 2009 . Проверено 21 ноября 2009 г.
- Макиннес, Родерик Р. (март 2011 г.). «Обращение президента 2010 г.: Культура: генетики должны изучить немой язык — генетические исследования с участием коренного населения» . Американский журнал генетики человека . 88 (3): 254–261. дои : 10.1016/j.ajhg.2011.02.014 . ПМК 3059421 . ПМИД 21516613 .
- Рафф, Дженнифер (8 февраля 2022 г.). Происхождение: Генетическая история Америки . Издательство Гранд Сентрал. ISBN 978-1-5387-4970-8 .
Генетическая история коренных народов Америки |
|---|


- Коренные народы Америки
- Современная генетическая история человека
- Генетическая генеалогия
- Генетика по этнической принадлежности
- История коренных народов Америки
- История коренных народов Северной Америки
- Коренные темы Карибского бассейна
- Коренные темы Южной Америки
- История Южной Америки
- Индейские темы
- История коренных американцев
- Коренные народы в Соединенных Штатах
- Плейстоцен
- Доколумбовая эпоха
- Коренные народы Канады
- История коренных народов Канады