Сексуальный диморфизм

| Часть серии на |
| Секс |
|---|
 |
| Биологические термины |
| Сексуальное размножение |
| Сексуальность |
| Сексуальная система |
Сексуальный диморфизм - это состояние, при котором полов одного и того же вида демонстрируют разные морфологические характеристики, включая характеристики, не связанные с непосредственно в воспроизведении . [ 1 ] Состояние происходит у большинства диологических видов, которые состоят из большинства животных и некоторых растений. Различия могут включать вторичные половые характеристики , размер, вес, цвет, маркировку или поведенческие или когнитивные признаки. Репродуктивная конкуренция мужского мужского мужчины развила разнообразные сексуально диморфные черты. Агрессивные признаки коммунальных услуг, такие как «боевые» зубы и тупые головы, усиленные в качестве избиения баранов, используются в качестве оружия в агрессивных взаимодействиях между соперниками. Пассивные дисплеи, такие как декоративное перья или вызов песен, также развивались в основном благодаря сексуальному отбору. [ 2 ] Эти различия могут быть тонкими или преувеличенными и могут быть подвергнуты сексуальному отбору и естественному отбору . Противоположностью диморфизма является мономорфизм , когда оба биологических полов фенотипически неотличимы друг от друга. [ 3 ]
Обзор
[ редактировать ]

Украшение и окраска
[ редактировать ]
Общие и легко идентифицированные типы диморфизма состоят из украшения и окраски, хотя и не всегда очевидны. Разница в окраске полов в данном виде называется сексуальным дихроматизмом, который обычно наблюдается у многих видов птиц и рептилий. [ 4 ] Сексуальный отбор приводит к преувеличенным диморфическим признакам, которые используются преимущественно в конкуренции над товарищами. [ 5 ] Повышенная физическая форма, возникающая в результате того, что орнаментовые средства сметают его затраты на производство или поддержание, что свидетельствует о сложных эволюционных последствиях, но затраты и эволюционные последствия варьируются от видов к видам. [ 6 ]
Павфуры представляют собой заметные иллюстрации принципа. Декоративное оперение павлина, используемое на ухационном дисплее, привлекает Peahens . На первый взгляд можно принять павлинов и павхен за совершенно разные виды из -за ярких цветов и огромного размера оперения мужчины; Пиан имеет приглушенную коричневую окраску. [ 7 ] Оперение павлина увеличивает его уязвимость к хищникам, потому что это препятствие в полете, и это делает птицу заметной в целом. [ 7 ] Аналогичные примеры многочисленны, например, в райских птицах [ 8 ] и Аргус Фазанты . [ Цитация необходима ]
Другим примером сексуального дихроматизма является пример синих сисек . Мужчины хроматически более желтые, чем женщины. Считается, что это получается путем приема личинок зеленых чешуекрылых , которые содержат большое количество каротиноидов лютеина и зеаксантина . [ 9 ] Эта диета также влияет на сексуально диморфные цвета в ультрафиолетовом спектре, которым не известно человеку ультрафиолетовый спектр. [ 10 ] [ 11 ] Следовательно, у птиц-мужчин, хотя и появляются желтые для людей, на самом деле имеют фиолетовое оперение, которое видно женщинами. Считается, что это оперение является индикатором мужских способностей родителей. [ 12 ] Возможно, это хороший показатель для женщин, потому что он показывает, что они хорошо получают пищевую запас, из которого получается каротиноид. Существует положительная корреляция между хромами хвоста и грудными перьями и состоянием тела. [ 13 ] Каротиноиды играют важную роль в иммунной функции для многих животных, поэтому каротиноид -зависимые сигналы могут указывать на здоровье. [ 14 ]
Лягушки представляют собой еще одну заметную иллюстрацию принципа. Существует два типа дихроматизма для видов лягушек: онтогенетический и динамичный. Онтогенетические лягушки чаще встречаются и имеют постоянные изменения цвета у мужчин или женщин. Ranoidea Lesueuri является примером динамической лягушки с временными изменениями цвета у мужчин в течение сезона размножения. [ 15 ] Hyperolius ocellatus - это онтогенетическая лягушка с драматическими различиями как в цвете, так и в рисунке между полами. В сексуальной зрелости мужчины демонстрируют ярко -зеленый с белыми дорсолатеральными линиями. [ 16 ] Напротив, женщины ржавые от красного до серебра с небольшими пятнами. Яркая окраска в мужской популяции привлекает женщин и является апосемичным знаком к потенциальным хищникам.
Женщины часто демонстрируют предпочтение преувеличенным вторичным сексуальным характеристикам мужского пола при отборе партнеров. [ 17 ] Гипотеза сексуального сына объясняет, что женщины предпочитают более сложных мужчин и выбирают против мужчин, которые скучно по цвету, независимо от видения вида. [ 18 ]
Подобный половой диморфизм и выбор спаривания также наблюдаются у многих видов рыб. Например, мужские гуппи имеют красочные пятна и украшения, в то время как женщины, как правило, серые. Женские гуппи предпочитают ярко окрашенные мужчины тупым мужчинам. [ 19 ] [ страница необходима ]
В Blennies Redlip только у мужской рыбы развивается орган в анальной области ирогенитальной области, который продуцирует антимикробные вещества. Во время родительской помощи мужчины втирают свои аналогидочные области над внутренними поверхностями своих гнезд, тем самым защищая свои яйца от микробных инфекций, одной из наиболее распространенных причин смертности у молодой рыбы. [ 20 ]
Растения
[ редактировать ]Большинство цветущих растений являются гермафродитными , но приблизительно 6% видов имеют отдельные мужчины и женщины ( Dioecy ). [ 21 ] Сексуальный диморфизм распространен в двудомных растениях [ 22 ] : 403 и диорические виды. [ 23 ] : 71
Мужчины и женщины у насекомых видов, скользящих с насекомыми, обычно похожи друг на друга, потому что растения обеспечивают награды (например, нектар ), которые побуждают опылителей посещать другой подобный цветок , завершая опыление . Орхидеи катазы являются одним интересным исключением из этого правила. Мужские катазоистые орхидеи насильственно прикрепляют опылиния к эвглоссиновых опылителям пчел пчел. Пчелы будут тогда избегать других мужских цветов, но могут посетить самку, которая отличается от мужчин. [ 24 ]
Различные другие исключительные исключения, такие как локсостилис алата , имеют заметно разные полов, с влиянием выявления наиболее эффективного поведения у опылителей, которые затем используют наиболее эффективную стратегию при посещении каждого пола цветка вместо поиска, скажем, для пыльцы в нектаре -Вывая женский цветок. [ Цитация необходима ]
Некоторые растения, такие как некоторые виды гераня, имеют то, что составляет серийный половой диморфизм. Цветы таких видов могут, например, представить свои пыльники на открытии, затем пролить измотанные пыльники через день или два и, возможно, изменить их цвета, а также созревает пеликт ; Специализированные опылители очень склонны сконцентрироваться на точном появлении цветов, которые они служат, что экономит их время и усилия и соответствует интересам растения. Некоторые такие растения идут еще дальше и меняют свой внешний вид после оплодотворения, тем самым препятствуя дальнейшим визитам опылителей. Это выгодно для обеих сторон, потому что это позволяет избежать повреждения развивающихся фруктов и тратить усилия опылителя на неотъемлемые визиты. По сути, стратегия гарантирует, что опылители могут ожидать вознаграждения каждый раз, когда они посещают соответствующую рекламу цветка. [ Цитация необходима ]
У женщин водного растения в Валлиснерии Американа есть плавучие цветы, прикрепленные к длинным цветочным стеблу , которые оплодотворяются, если они связываются с одним из тысяч свободных цветов, выпущенных мужчиной. [ 25 ] [ Лучший источник необходим ] Сексуальный диморфизм чаще всего ассоциируется с переплетом ветра у растений из-за отбора для эффективного рассеивания пыльцы у мужчин против захвата пыльцы у женщин, например, Leucadendron Rubrum . [ 26 ]
Сексуальный диморфизм у растений также может зависеть от репродуктивного развития. Это можно увидеть в каннабисе Sativa , типе конопли, которая имеет более высокие показатели фотосинтеза у мужчин при росте, но более высоких показателях у женщин, как только растения становятся сексуально зрелыми. [ 27 ]
Каждый сексуально воспроизводит существующие виды сосудистого растения имеет чередование поколений; Растения, которые мы видим о нас, обычно, являются диплоидными спорофитами , но их потомство не являются семенами, которые люди обычно признают новым поколением. Семя на самом деле представляет собой потомство гаплоидного генерации микрогаметофитов ( пыльцы ) и мегагаметофитов ( эмбрионные мешки в яйцеклетках ). Каждое пыльцевое зерно соответственно может рассматриваться как мужское растение само по себе; Он производит сперматозоиду и резко отличается от женского растения, мегагаметофита, который производит женскую гамету. [ Цитация необходима ]
Насекомые
[ редактировать ]
Насекомые демонстрируют широкий спектр сексуального диморфизма между таксонами, включая размер, украшение и окраску. [ 28 ] Диморфизм сексуального размера, предназначенный для женщин, наблюдался во многих таксонах, развивался, несмотря на интенсивную конкуренцию мужского мужского мужчины для товарищей. [ 29 ] Например, в Osmia Rufa самка больше/шире, чем мужчины, а мужчины имеют размер 8–10 мм, а женщины - 10–12 мм размер. [ 30 ] В хакерских императорах женщины так же больше, чем мужчины. [ 31 ] Причина сексуального диморфизма связана с массой размера обеспечения, в которой женщины потребляют больше пыльцы, чем мужчины. [ 32 ]
У некоторых видов есть свидетельства мужского диморфизма, но, по -видимому, это для различий ролей. Это видно на видах пчелиновых видов Macrotera Portalis, в котором есть небольшая головная морф, способная к полету и большую головную морф, неспособный к полету, для мужчин. [ 33 ] Anthidium manicatum также демонстрирует сексуальное диморфизм с мужчинами. Выбор для большего размера у мужчин, а не женщин у этого вида, мог быть вызван из -за их агрессивного территориального поведения и последующего успеха в дифференциальном спаривании. [ 34 ] Другим примером является Lasioglossum Hemichalceum , который является видом пота, который показывает радикальные физические диморфизмы между мужским потомством. [ 35 ] Не весь диморфизм должен иметь радикальную разницу между полами. Andrena Agilissima - это горнодобывающая пчела, где женщины имеют только немного большую голову, чем у мужчин. [ 36 ]
Оружие приводит к повышению физической подготовки за счет увеличения успеха в конкуренции мужского пола у многих видов насекомых. [ 37 ] Рога жука в онтофаге -Тельце - это увеличенные рост головы или грудной клетки, экспрессируемых только у мужчин. Коприс Охус также имеет отдельный сексуальный и мужской диморфизм в рогах головы. [ 38 ] Другим жуком с отчетливым связанным с рогом сексуальным диморфизмом является дихотома алломирины, также известная как японская носорога . [ 39 ] Эти структуры впечатляют из -за преувеличенных размеров. [ 40 ] Существует прямая корреляция между длиной мужского рога и размером тела и более высоким доступом к товарищам и пригодности. [ 40 ] У других видов жуков как мужчины, так и женщины могут иметь орнамент, такие как рога. [ 38 ] Как правило, диморфизм сексуального размера насекомых (SSD) внутри видов увеличивается с размером тела. [ 41 ]
Сексуальный диморфизм у насекомых также демонстрируется дихроматизмом. В родах бабочек Bicyclus и Junonia диморфические паттерны крыла развивались из-за выражения по полу, которая опосредует внутрилокусный сексуальный конфликт и приводит к повышению подготовки к мужчинам. [ 42 ] Сексуальная дихроматическая природа Bicyclus Anhynana отражается на женском отборе на основе спин-ультрафиолетовых зрачков для глазок. [ 43 ] Обыкновенный сара также демонстрирует сексуальный дихроматизм; У мужчин желтые и радужные крылья, в то время как женские крылья белые и не являются иридесцентными. [ 44 ] Естественно отобранное отклонение в защитной женской окраске отображается у миметических бабочек. [ 45 ]
Пауки и сексуальные каннибализм
[ редактировать ]

Многие группы арахнидов демонстрируют сексуальный диморфизм, [ 46 ] Но это наиболее широко изучается в пауках. в пауке Zygiella x-notata , у взрослых, имеют больший размер тела Zygiella x-notata. Например, [ 47 ] Диморфизм размера показывает корреляцию с сексуальным каннибализмом , [ 48 ] который является заметным у пауков (он также встречается у насекомых, таких как молятельные мантисы ). В размере диморфического волчьего паука Тигроса Хеллуо , пострадавшие от пищи женщины чаще каннибализируют. [ 49 ] Следовательно, существует высокий риск низкой подготовки к мужчинам из-за доконного каннибализма, который привел к мужскому отбору более крупных женщин по двум причинам: более высокая плодовитость и более низкие показатели каннибализма. [ 49 ] Кроме того, женская плодовитость положительно коррелирует с размером женского тела, и выбирается большой размер женского тела, который наблюдается в семействе Araneidae . Все Argiope виды , включая Argiope Bruennichi , используют этот метод. Некоторые мужчины эволюционировали украшение [ нечеткий ] Включая связывание самки с шелком, имея пропорционально более длинные ноги, модифицируя сеть женщин, спаривание во время кормления женщины, или предоставляя брачный подарок в ответ на сексуальный каннибализм. [ 49 ] Размер мужского тела не находится под отбором из -за каннибализма у всех видов пауков, таких как Нефила Пилипес , но более заметно отбирается для менее диморфных видов пауков, которые часто выбирают для большего размера мужчин. [ 50 ] У вида Maratus Volans мужчины известны своим характерным красочным вентилятором, который привлекает женщин во время спаривания. [ 51 ]
Рыба
[ редактировать ]Рыбные рыбы-древний и разнообразный класс, с самой широкой степенью сексуального диморфизма любого класса животных. Фэрбэрн отмечает, что «женщины, как правило, больше, чем мужчины, но мужчины часто бывают больше по видам с боем мужского пола или мужского по отцовской помощи ... [диапазон размеров] от карликовых мужчин до мужчин, более чем в 12 раз тяжелее женщин». [ 52 ] [ страница необходима ]
Есть случаи, когда мужчины существенно больше, чем женщины. Примером является Lamprologus callipterus , тип цихлидской рыбы. У этой рыбы мужчины характеризуются как в 60 раз больше женщин. Считается, что повышенный размер мужчины является выгодным, потому что мужчины собирают и защищают пустые снаряды улитки, в каждой из которых пород самка. [ 53 ] Мужчины должны быть больше и мощнее, чтобы собрать самые большие раковины. Размер тела женщины должен оставаться маленьким, потому что для того, чтобы она размножалась, она должна отложить яйца в пустые раковины. Если она станет слишком большой, она не будет вписаться в раковины и не сможет размножаться. Маленький размер тела женщины также, вероятно, полезен для ее шансов найти незанятую оболочку. Большие раковины, хотя и предпочтительны у женщин, часто ограничены в доступности. [ 54 ] Следовательно, женщина ограничивается ростом размера оболочки и может фактически изменить скорость ее роста в зависимости от доступности размера оболочки. [ 55 ] Другими словами, способность мужчины собирать большие раковины зависит от его размера. Чем больше мужчина, тем больше раковин, он может собрать. Затем это позволяет женщинам быть больше в его задумчивом гнезде, что делает разницу между размерами полов менее существенными. Соревнования по мужчинам и мужчинам у этих видов рыб также выбирают для большого размера у мужчин. Существует агрессивная конкуренция со стороны мужчин на территории и доступ к более крупным снарядам. Крупные мужчины выигрывают бои и крадут снаряды у конкурентов. Другим примером является Dragonet , в котором мужчины значительно больше, чем женщины, и обладают более длинными плавниками.
Сексуальный диморфизм также встречается у гермафродитной рыбы. Эти виды известны как последовательные гермафродиты . В рыбе репродуктивные истории часто включают изменение пола от женщины к мужчине, где существует тесная связь между ростом, полом человека и системой спаривания, в которой он работает. [ 56 ] В системах протонитных спаривания, где мужчины доминируют в спаривании со многими женщинами, размер играет значительную роль в репродуктивном успехе мужского пола. [ 57 ] Мужчины имеют склонность быть больше, чем у женщин сопоставимого возраста, но неясно, связано ли увеличение размера из -за всплеска роста во время сексуального перехода или из -за истории более быстрого роста у людей, меняющихся. [ 58 ] Крупные мужчины могут задушить рост женщин и контролировать экологические ресурсы. [ Цитация необходима ]
Социальная организация играет большую роль в смене секса со стороны рыбы. Часто видно, что рыба изменит свой пол, когда в социальной иерархии не хватает доминирующего мужчины. Женщины, которые меняют секс, часто являются теми, кто достигает и сохраняет преимущество начального размера в начале жизни. В любом случае, женщины, которые меняют пол для мужчин, больше и часто оказываются хорошим примером диморфизма.
В других случаях с рыбой мужчины будут переживать заметные изменения в размере тела, и женщины пройдут через морфологические изменения, которые можно увидеть только внутри тела. Например, у лосося в социальных сетях у мужчин развивается больший размер тела при зрелости, включая увеличение глубины тела, высоту горба и длину морды. Самки испытывают незначительные изменения в длине морды, но наиболее заметным отличием является огромное увеличение размера гонады , которое составляет около 25% массы тела. [ 59 ]
Сексуальный отбор наблюдался для женского орнамента в Gobiusculus flavescens , известной как гоби из двух сторон. [ 60 ] Традиционные гипотезы предполагают, что соревнования по мужчинам и мужчинам стимулируют отбор. Тем не менее, отбор для украшения в этом виде предполагает, что эффектные женские черты могут быть выбраны с помощью конкуренции за женским и делом для женщин или выбора мужчин. [ 60 ] Поскольку орнамент на основе каротиноидов предполагает качество спаривания, женские два пятна гуппи, которые развивают красочные апельсиновые животы в течение сезона размножения, считаются благоприятными для мужчин. [ 61 ] Мужчины вкладывают значительные средства в потомство во время инкубации, что приводит к сексуальным предпочтениям у разноцветных женщин из -за более высокого качества яиц. [ 61 ]
Амфибии и неавийские рептилии
[ редактировать ]
У амфибий и рептилий степень сексуального диморфизма широко варьируется среди таксономических групп . Сексуальный диморфизм у амфибий и рептилий может быть отражен в любом из следующих: анатомия; относительная длина хвоста; относительный размер головы; общий размер, как у многих видов гадюков и ящериц ; окраска, как у многих амфибий , змей и ящериц, а также в некоторых черепах ; украшение, как во многих новых и ящерицах; Наличие конкретного поведения, связанного с полом, является общим для многих ящериц; и вокальные качества, которые часто наблюдаются у лягушек . [ Цитация необходима ]
Анольские ящерицы показывают заметный диморфизм размера с мужчинами, как правило, значительно больше, чем женщины. Например, средний мужской Anolis Sagrei составлял 53,4 мм против 40 мм у женщин. [ 62 ] Различные размеры голов в анолах были объяснены различиями в пути эстрогена. [ 63 ] Сексуальный диморфизм у ящериц, как правило, объясняется последствиями сексуального отбора, но другие механизмы, включая экологическую дивергенцию и отбор плодовитости, дают альтернативные объяснения. [ 64 ] Развитие цветового диморфизма у ящериц вызвано гормональными изменениями в начале половой зрелости, как видно из Psamodromus algirus , Sceloporus gadoviae и S. uldulates erhthrocheilus . [ 64 ] Половой диморфизм в размерах также наблюдается у видов лягушек, таких как P. bibroni i .
Мужские окрашенные ящерицы драконов, Ctenophorus pictus . ярко заметны в своей размножении, но мужской цвет снижается при старении . Мужская окраска, по-видимому, отражает врожденную антиоксидийную способность, которая защищает от окислительного повреждения ДНК . [ 65 ] Мужская окраска размножения, вероятно, является показателем для женщин базового уровня окислительного повреждения ДНК (значительный компонент старения) у потенциальных товарищей. [ 65 ]
Птицы
[ редактировать ]




Возможные механизмы были предложены для объяснения макроэволюции диморфизма сексуального размера у птиц. К ним относятся половой отбор, отбор для плодовитости у женщин, ниша дивергенция между полами и аллометрия, но их относительная важность до сих пор не до конца понятна. [ 66 ] [ 67 ] Сексуальный диморфизм у птиц может проявляться в различиях в размерах или оперении между полами. Диморфизм сексуального размера варьируется среди таксонов, и мужчины, как правило, больше, хотя это не всегда так, например, птицы добычи , колибри и некоторые виды безлетовых птиц. [ 68 ] [ 69 ] Диморфизм оперения в форме украшения или окраски также варьируется, хотя мужчины, как правило, являются более украшенным или ярко окрашенным полом. [ 70 ] Такие различия были связаны с неравным репродуктивным вкладом полов. [ 71 ] Эта разница дает более сильный выбор женского пола, так как они имеют больший риск в производстве потомства. У некоторых видов вклад мужчины в размножение заканчивается при совокуплении, в то время как у других видов мужчина становится главным (или единственным) попечителем. Полиморфизмы оперения развивались, чтобы отразить эти различия и другие показатели репродуктивной пригодности, такие как состояние тела [ 72 ] или выживание. [ 73 ] Мужской фенотип посылает сигналы женщинам, которые затем выбирают «наиболее подходящий» мужчина.
Сексуальный диморфизм является продуктом как генетики, так и факторов окружающей среды. Пример сексуального полиморфизма, определяемый условиями окружающей среды, существует в сказочных скачках с красными . Красные сказочные мужчины могут быть классифицированы на три категории в течение сезона размножения : черные заводчики, коричневые заводчики и коричневые вспомогательные. [ 72 ] Эти различия возникают в ответ на состояние тела птицы: если они здоровы, они будут производить больше андрогенов, становящихся черными заводчиками, в то время как менее здоровые птицы производят меньше андрогенов и станут коричневыми вспомогательными. [ 72 ] Таким образом, репродуктивный успех мужчины определяется его успехом в течение каждого годового сезона, приводящего к репродуктивному успеху варьироваться с каждым годовым условиями окружающей среды.
Мигрирующие паттерны и поведение также влияют на сексуальные диморфизмы. Этот аспект также возвращается к диморфизму размера у видов. Было показано, что более крупные мужчины лучше справляются с трудностями миграции и, следовательно, более успешны в воспроизведении при достижении места назначения размножения. [ 74 ] При просмотре этого с эволюционной точки зрения, многие теории и объяснения учитываются. Если это результат для каждого сезона миграции и размножения, ожидаемыми результатами должны быть сдвиг в сторону более крупного мужского населения за счет полового отбора. Сексуальный отбор силен, когда фактор отбора окружающей среды также введен. Выбор окружающей среды может поддерживать меньший размер цыплят, если эти цыплят родились в области, которая позволила им расти до большего размера, хотя при нормальных условиях они не смогут достичь этого оптимального размера для миграции. Когда окружающая среда дает преимущества и недостатки такого рода, сила отбора ослаблена, а силы окружающей среды получают больший морфологический вес. Сексуальный диморфизм также может привести к изменению сроков миграции, что приводит к различиям в успехе спаривания в популяции птиц. [ 75 ] Когда диморфизм производит такую большую вариацию между полами и между членами полов, может иметь место множественные эволюционные эффекты. Это время может даже привести к явлению видообразования, если вариация становится сильно радикальной и благоприятной к двум различным результатам. Сексуальный диморфизм поддерживается противодействующим давлением естественного отбора и сексуального отбора. Например, половой диморфизм в окраске увеличивает уязвимость видов птиц к хищничеству европейскими воробьиками в Дании. [ 76 ] Предположительно, повышенный половой диморфизм означает, что мужчины более яркие и заметны, что приводит к увеличению хищничества. [ 76 ] Более того, производство более преувеличенных украшений у мужчин может быть за счет подавленной иммунной функции. [ 72 ] До тех пор, пока репродуктивные выгоды от признака из -за полового отбора больше, чем затраты, налагаемые естественным отбором, тогда признака будет распространяться по всему населению. Репродуктивные выгоды возникают в форме большего числа потомков, в то время как естественный отбор налагает затраты в виде снижения выживаемости. Это означает, что даже если признака заставляет мужчин умирать раньше, черта все еще полезна, пока мужчины с чертой производят больше потомства, чем мужчины, не имеющие этой черты. Этот баланс поддерживает диморфизм в этих видах и гарантирует, что следующее поколение успешных мужчин также будет демонстрировать эти черты, которые привлекательны для женщин.
Такие различия в форме и репродуктивных ролях часто вызывают различия в поведении. Как указывалось ранее, мужчины и женщины часто играют разные роли в размножении. Ухаживание и спаривание мужчин и женщин регулируются в основном гормонами на протяжении всей жизни птицы. [ 77 ] Активационные гормоны встречаются во время полового созревания и взрослой жизни и служат для «активации» определенных поведений, когда это уместно, например, территориальность во время сезона размножения. [ 77 ] Организационные гормоны встречаются только в критический период в начале разработки, либо незадолго до, либо сразу после вылупления у большинства птиц, и определяют закономерности поведения на всю оставшуюся жизнь птицы. [ 77 ] Такие поведенческие различия могут вызвать непропорциональную чувствительность к антропогенному давлению. [ 78 ] Женщины Whinchat в Швейцарии размножаются в сильно управляемых лугах. [ 78 ] Ранее сбор трав во время сезона размножения приводит к большему количеству смертей женщин. [ 78 ] Популяции многих птиц часто встречаются мужчинами, и когда сексуальные различия в поведении увеличивают это соотношение, популяции снижаются более быстрыми темпами. [ 78 ] Кроме того, не все мужские диморфические признаки связаны с гормонами, такими как тестостерон, вместо этого они являются естественной частью развития, например, оперения. [ 79 ] Кроме того, сильное гормональное влияние на фенотипические различия предполагает, что генетический механизм и генетическая основа этих сексуально диморфных признаков могут включать транскрипционные факторы или кофакторы, а не регуляторные последовательности. [ 80 ]
Сексуальный диморфизм также может влиять на различия в инвестициях родителей во время дефицита пищи. Например, в сине-ногах , женские цыпочки растут быстрее, чем мужчины, в результате чего родители, выпускающие меньший пол, мужчины, во время нехватки пищи. Затем это приводит к максимизации репродуктивного успеха в жизни родителей. [ 81 ] В черновостых Limosa Limosa Limosa Limosa женщины также являются более крупным полом, а темпы роста женских цыплят более подвержены ограниченным условиям окружающей среды. [ 82 ]
Сексуальный диморфизм также может появляться только в сезон спаривания; Некоторые виды птиц демонстрируют только диморфные черты в сезонных вариациях. Мужчины этих видов будут раститься в менее яркий или менее преувеличенный цвет в течение неверного сезона. [ 80 ] Это происходит потому, что вид больше сосредоточен на выживании, чем на размножении, что вызывает переход в менее богато украшенное состояние. [ сомнительно - обсудить ]
Следовательно, половой диморфизм имеет важные последствия для сохранения. Тем не менее, половой диморфизм обнаруживается не только у птиц и, таким образом, важен для сохранения многих животных. Такие различия в форме и поведении могут привести к сексуальной сегрегации , определяемой как половые различия в использовании пространства и ресурсов. [ 83 ] Большинство исследований сексуального сегрегации было проведено на кожих, [ 83 ] Но такое исследование распространяется на летучие мыши , [ 84 ] кенгуру , [ 85 ] и птицы. [ 86 ] Половые планы сохранения даже были предложены для видов с выраженной сексуальной сегрегацией. [ 84 ]
Термин сесвиморфизм (латинский цифровой префикс сесвик - означает один с половиной, поэтому на полпути между моно (один) и di - (два)) был предложен для видов птиц, у которых оба пола в основном имеют одинаковое оперение Узор, хотя самка явно различима по причине ее будущего или промытого цвета ». [ 87 ] : 14 Примеры включают Cape Sparrow ( прохожие Меланурус ), [ 87 ] : 67 Rufous Sparrow (подвид P. motinensis motinensis ), [ 87 ] : 80 и Саксл Воробей ( П. Амдендри ). [ 87 ] : 245
Неавийские динозавры
[ редактировать ]Изучение окаменелостей неавийских динозавров в поисках сексуальных диморфных характеристик требует поставки полных и сформулированных скелетных и тканевых остатков. Как наземные организмы, туши динозавров подвержены экологическому и географическому влиянию, которое неизбежно составляет степень сохранения. Доступность хорошо сохранившихся останков не является вероятным результатом в результате разложения и ископаемости . Некоторые палеонтологи искали половой диморфизм среди динозавров, используя статистику и сравнение с экологически или филогенетически связанными современными животными.
Апатозавр и диплодокус
Женский апатозавр и диплодокус имели взаимосвязанные каудальные позвонки , которые позволили им поднять свои хвосты, чтобы помочь при совокупленности. Обнаружение, что это слияние произошло только в 50% скелетов апатозавра и диплодокуса , и 25% скелетов камаразавра показали, что это сексуально диморфная черта.
Теропода
Было выдвинуто предположение, что мужские тероподы обладали выдвижным пенисом, особенностью, похожей на современные крокодильцы . Крокодильские скелеты были исследованы, чтобы определить, существует ли скелетный компонент, который отличается от обоих полов, чтобы помочь дать представление о физических различиях между мужскими и женскими тероподами. Результаты показали, что хвостовые шевроны мужских крокодилов, используемые для закрепления мышц полового члена, были значительно больше, чем у женщин. Были критические замечания в отношении этих выводов, но это остается предметом дебатов среди адвокатов и противников. [ Цитация необходима ]
Ornithopoda
Исследования сексуального диморфизма у адрозавров , как правило, сосредоточены на характерных черепных гребнях , которые, вероятно, обеспечивали функцию в сексуальном дисплее. Биометрическое исследование 36 черепов обнаружило, что половой диморфизм был продемонстрирован в гребне 3 видов хадрозавридов. Гребники могут быть классифицированы как полные (мужские) или узкие (женские) и могли дать некоторое преимущество в интрасексуальном соревнованиях по спариванию.
Сератеры
По словам Скотта Д. Сэмпсона, если бы Ceratopsids проявил половой диморфизм, современные экологические аналоги предполагают, что он будет обнаружен в демонстрационных структурах, таких как рога и изличие. Никаких убедительных доказательств сексуального диморфизма в размере тела или символов спаривания не известно в Ceratopsids, хотя есть доказательства того, что более примитивные цератопсийские протокератопы Эндрюси обладали полами, которые были различимы на основе размера фрила и носового признания. Это согласуется с другими известными тетраподными группами, где животные среднего размера, как правило, демонстрируют заметно более половой диморфизм, чем более крупные. Тем не менее, было предложено, чтобы эти различия могли быть лучше объяснены внутривидовым и онтогенным изменением, а не половым диморфизмом. [ 88 ] Кроме того, многие сексуальные диморфные признаки, которые могли существовать в цератопсах, включают вариации мягких тканей, такие как окраска или росы , которые вряд ли будут сохранены в ископаемом отчете.
Стегозавры
Исследование, проведенное в 2015 году по образцам Hesperosaurus mjosi, обнаружило доказательства сексуального диморфизма в форме дермальных пластин. Были описаны два морфы тарелки: одно было коротким, шириной и овальной формой, другой выше и уже. [ 89 ] [ 90 ]
Млекопитающие
[ редактировать ]У большой части видов млекопитающих мужчины больше, чем женщины. Как гены , так и гормоны влияют на формирование многих животных мозгов до « рождения » (или вылупления ), а также поведения взрослых людей. Гормоны значительно влияют на образование мозга человека, а также на развитие мозга в период полового созревания. Обзор в 2004 году в природе обзор Neuroscience наблюдал, что «поскольку« поскольку легче манипулировать уровнем гормонов, чем экспрессия генов половых хромосом, эффекты гормонов изучались гораздо более широко и гораздо лучше поняты, чем прямые действия в мозге генов сексуальной хромосом ". Он пришел к выводу, что, хотя «дифференцирующие эффекты секреции гонад кажутся доминирующими,« существующий объем исследований »подтверждает идею о том, что половые различия в нейронной экспрессии генов X и Y значительно способствуют половым различиям в функциях мозга и болезнях». [ 91 ]
Выносливые
[ редактировать ]
Морские млекопитающие показывают некоторые из величайших различий в сексуальном размере у млекопитающих из -за сексуального отбора и факторов окружающей среды, таких как местоположение размножения. [ 92 ] Система спаривания выносливов варьируется от многоженства до последовательной моногамии . Выпечны известны ранним дифференциальным ростом и материнскими инвестициями, поскольку единственными питательными веществами для новорожденных щенков является молоко, обеспечиваемое матери. [ 93 ] Например, мужчины значительно больше (примерно на 10% тяжелее и на 2% длиннее), чем женщины при рождении у щенков морских львов. [ 94 ] Схема дифференциальных инвестиций может варьироваться в основном пренатально и пост-натально. [ 95 ] Мираунга Леонина , южный слон Печать , является одним из самых диморфных млекопитающих. [ 96 ]
Приматы
[ редактировать ]Люди
[ редактировать ]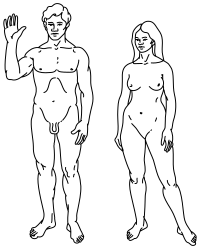 | |
 |
 |
|
Вверху: стилизованная иллюстрация людей на пионерской табличке , показывающая как мужской (слева, и женский (справа).
| |
По словам Кларка Спенсера Ларсена, современные Homo Sapiens демонстрируют ряд сексуального диморфизма, причем средняя масса тела между полами различается примерно на 15%. [ 97 ] Существенное обсуждение в академической литературе учитывает потенциальные эволюционные преимущества, связанные с сексуальной конкуренцией (как внутрисексуальной, так и межсексуальной), а также краткосрочные и долгосрочные сексуальные стратегии. [ 98 ] По словам Дейли и Уилсона, «полы больше различаются у людей, чем у моногамных млекопитающих, но гораздо меньше, чем у чрезвычайно полигамных млекопитающих». [ 99 ]
Средний базальный уровень метаболизма примерно на 6 процентов у подростков выше, чем у женщин, и увеличивается примерно на 10 процентов выше после полового созревания. Женщины, как правило, превращают больше пищи в жир , в то время как мужчины превращаются в мышечные и расходуемые энергетические запасы циркулирующих. По словам Тима Хьюитта, директора по исследованиям в Департаменте спортивной медицины в Медицинском центре Университета штата Огайо Уэкснер , у женщин в среднем 50–60% мужчин и 80-90% силы нижней части тела мужчин, относительно размера тела, но женщины имеют более высокую выносливость , чем мужчины. [ 100 ]
Разница в силе относительно массы тела менее выражена у обученных людей. В олимпийской тяжелой атлетике, мужские записи варьируются от 5,5 × массы тела в категории с самым низким весом до 4,2 × в категории с самым высоким весом, в то время как записи женщин варьируются от 4,4 × до 3,8 ×, разница с поправкой на вес составляет всего 10–20%, и и только 10–20%и Абсолютная разница около 30% (т. Е. 492 кг против 348 кг для неограниченных весовых категорий; см. Олимпийские записи о тяжелой атлетике ). Исследование, проведенное путем анализа ежегодных мировых рейтингов с 1980 по 1996 год, показало, что время бега мужчин было в среднем на 10% быстрее, чем женщины ». [ 101 ]
В раннем подростковом возрасте женщины в среднем выше, чем мужчины (поскольку женщины, как правило, проходят половое созревание раньше ), но мужчины в среднем превосходят их по высоте в более позднем подростковом и взрослой жизни. В Соединенных Штатах взрослые мужчины в среднем на 9% выше [ 102 ] и 16,5% тяжелее [ 103 ] чем взрослые женщины.
Мужчины обычно имеют большие трахей и разветвления бронхов , примерно на 30 процентов больший объем легких на массу тела . В среднем, у мужчин больше сердца , на 10 процентов выше количества эритроцитов , более высокий гемоглобин , следовательно, большую кислородную способность. Они также имеют более высокие коэффициенты свертывания циркуляции ( витамин К , про -тромбин и тромбоциты ). Эти различия приводят к более быстрому заживлению ран и снижению чувствительности к нервной боли после травмы. [ 104 ] У мужчин, вызывающее боль в периферическом нерве происходит через микроглию , в то время как у женщин это происходит через Т-клетки (за исключением беременных женщин, которые следуют мужской схеме). [ 105 ]
У самок обычно больше лейкоцитов (хранящихся и циркулирующих), а также больше гранулоцитов и В и Т -лимфоцитов . Кроме того, они производят больше антител более высокой скоростью, чем у мужчин, поэтому у них развивается меньше инфекционных заболеваний и поддается более коротким периодам. [ 104 ] Этологи утверждают, что женщины, взаимодействующие с другими женщинами и множественные потомства в социальных группах, испытывают такие черты, как селективное преимущество. [ 106 ] [ 107 ] [ 108 ] [ 109 ] [ 110 ] ‹ Шаблон чрезмерной цитаты рассматривается для удаления .› [ чрезмерные цитаты ] Самки обладают более высокой чувствительностью к боли из -за вышеупомянутых различий в нерве, которые увеличивают ощущение, и, таким образом, женщины требуют более высоких уровней обезболивающих после травмы. [ 105 ] Гормональные изменения у женщин влияют на чувствительность к боли, а беременные женщины обладают такой же чувствительностью, что и мужчины. Острая терпимость к боли также более последователен в течение всей жизни у женщин, чем у мужчин, несмотря на эти гормональные изменения. [ 111 ] Несмотря на различия в физическом чувстве, оба пола имеют сходную психологическую терпимость к (или способности справляться и игнорировать) боль. [ 112 ]
В человеческом мозге разница между полами наблюдалась в транскрипции пары генов PCDH11X /Y, уникальной для Homo Sapiens . [ 113 ] Сексуальная дифференциация в человеческом мозге из недифференцированного состояния вызвана тестостероном из яичка плода. Тестостерон превращается в эстроген в мозге посредством действия фермента ароматазы. Тестостерон действует на многих областях мозга, включая SDN-POA , чтобы создать мужской схему мозга. [ 114 ] Мозг беременных женщин, несущих мужские плоды, могут быть защищены от мужского воздействия андрогена за счет действия связующего полового гормона, связывающего глобулин . [ 115 ]
Взаимосвязь между половыми различиями в мозге и человеческим поведением является предметом противоречий в психологии и обществе в целом. [ 116 ] [ 117 ] Многие женщины, как правило, имеют более высокое соотношение серого вещества в левом полушарии мозга по сравнению с мужчинами. [ 118 ] [ 119 ] Мужчины в среднем имеют больший мозг, чем женщины; Однако при корректировке общего объема мозга различия в сером веществе между полами практически отсутствуют. Таким образом, процент серого вещества, по -видимому, больше связано с размером мозга, чем с сексом. [ 120 ] [ 121 ] Различия в физиологии мозга между полами не обязательно связаны с различиями в интеллекте. Haier et al. В исследовании 2004 года «мужчины и женщины, по -видимому, достигают сходных результатов IQ с различными областями мозга, что позволяет предположить, что нет единой, лежащей в основе нейроанатомической структуры в общем интеллекте и что различные типы конструкций мозга могут проявлять эквивалентную интеллектуальную эффективность». [ 122 ] (См. Статью о сексе и интеллекте для получения дополнительной информации по этому вопросу.) Выявленные строгие теоретические графические теоретические анализа связей человеческого мозга [ 123 ] Это в многочисленных теоретических параметрах (например, минимальная ширина двухпоставления, номер края, свойство с графом расширителя , минимальная крышка вершины ), структурный развязки женщин значительно «лучше» подключен, чем соединение мужчин. Это было показано [ 124 ] что теоретические различия графа связаны с полом, а не с различиями в объеме головного мозга путем анализа данных 36 женщин и 36 мужчин, где объем мозга каждого человека в группе был меньше объема мозга каждого женщина в группе.
Сексуальный диморфизм также был описан на уровне генов и показано, что он простирается от половых хромосом. В целом, было обнаружено, что около 6500 генов обладают сексуальной дифференциальной экспрессией по крайней мере в одной ткани. Многие из этих генов напрямую связаны с воспроизводством, а скорее связаны с более общими биологическими особенностями. Кроме того, было показано, что гены с пола-специфической экспрессией подвергаются снижению эффективности отбора, что приводит к более высоким частотам популяции вредных мутаций и способствует распространенности нескольких заболеваний человека. [ 125 ] [ 126 ]
Иммунная функция
[ редактировать ]Сексуальный диморфизм в иммунной функции является общей схемой у позвоночных, а также в ряде беспозвоночных. Чаще всего женщины более «иммунокомпетентны», чем мужчины. Эта черта не является согласованной среди всех животных, но отличается в зависимости от таксономии, причем у насекомых обнаруживается наибольшая иммунная система, представленная на жене. [ 127 ] У млекопитающих это приводит к более частым и тяжелым инфекциям у мужчин и более высоким показателям аутоиммунных расстройств у женщин. Одной из потенциальных причин могут быть различия в экспрессии генов иммунных клеток между полами. [ 128 ] Другое объяснение заключается в том, что эндокринологические различия между полами влияют на иммунную систему - например, тестостерон действует как иммунодепрессивный агент. [ 129 ]
Ячейки
[ редактировать ]Фенотипические различия между полами очевидны даже в культивируемых клетках из тканей. [ 130 ] Например, женские стволовые клетки , полученные из мышц , имеют лучшую эффективность регенерации мышц, чем мужские. [ 131 ] Есть сообщения о нескольких метаболических различиях между мужскими и женскими клетками [ 132 ] И они также реагируют на стресс по -разному. [ 133 ]
Репродуктивно выгодно
[ редактировать ]Теоретически, более крупные самки предпочитают конкуренцию за товарищей, особенно у полигамных видов. Более крупные женщины предлагают преимущество в фертильности, поскольку физиологические требования размножения ограничиваются у женщин. Следовательно, существует теоретическое ожидание того, что женщины, как правило, более крупны по видам, которые являются моногамными. Женщины больше во многих видах насекомых , многих пауков , многих рыб , многих рептилий, сов , хищных птиц и некоторых млекопитающих, таких как пятнистая гиена , и киты, такие как голубые киты . Например, у некоторых видов женщины сидячими, и поэтому мужчины должны искать их. Фриц Воллрат и Джефф Паркер утверждают, что эта разница в поведении приводит к радикально различным давлениям отбора на двух полов, очевидно, предпочитая меньших мужчин. [ 134 ] Случаи, когда мужчина больше, чем самка, также изучались, [ 134 ] и требуют альтернативных объяснений.
Одним из примеров такого типа диморфизма сексуального размера является миотис Bat Myotis Nigricans (Black Myotis Bat), где женщины значительно больше, чем мужчины с точки зрения массы тела, измерения черепа и длины предплечья. [ 135 ] Взаимодействие между полами и энергией, необходимой для производства жизнеспособного потомства, делает его благоприятным для женщин больше в этом виде. Женщины несут энергетическую стоимость производства яиц, что намного больше, чем стоимость изготовления спермы у мужчин. Гипотеза о преимуществах плодовитости гласит, что более крупная женщина может производить больше потомства и давать им более благоприятные условия для обеспечения их выживания; Это верно для большинства эктотерм. Большая женщина может обеспечить родительскую помощь в течение более длительного времени, пока потомство созревает. Периоды беременности и лактации довольно длинные у М. Нигриканов , женщины сосут свое потомство, пока не достигнут почти взрослого размера. [ 136 ] Они не смогут летать и ловить добычу, если в это время не компенсируют дополнительную массу потомства. Меньший размер мужского пола может быть адаптацией для повышения маневренности и ловкости, позволяя мужчинам лучше конкурировать с женщинами за продукты питания и другие ресурсы.

Некоторые виды рыболовов также демонстрируют чрезвычайный половой диморфизм. Женщины более типичны по внешнему виду для других рыб, тогда как мужчины являются крошечными рудиментарными существами с замедленными пищеварительными системами. Мужчина должен найти с ней женщину и слиться с ней: он затем живет паразитически, становясь чуть больше, чем телом, производящего спермы, в том, что составляет эффективно гермафродитный композитный организм. Аналогичная ситуация обнаруживается в Zeus Water Bug PhoreTicoVelia Disparata , где у самой железистой области на спине, которая может служить для кормления мужчины, что цепляется за нее (хотя мужчины могут выжить от женщин, они, как правило ) [ 137 ] Это доведено до логической крайности у ризоцефальских ракообразных, таких как Саккулина , где мужчина вводит себя в тело женщины и становится не чем иным, как клетки, продуцирующие сперму, до такой степени, что супер -заказ принимал за гермафродитическую. [ 138 ]
Некоторые виды растений также демонстрируют диморфизм, в котором самки значительно больше, чем у мужчин, например, в мохе дикранум [ 139 ] и печеночный Sphaerocarpos . [ 140 ] Есть некоторые доказательства того, что в этих родах диморфизм может быть связан с сексуальной хромосомой, [ 140 ] [ 141 ] или химическую передачу сигналов от женщин. [ 142 ]
Еще один сложный пример сексуального диморфизма - в Vespula Squamosa , южной желтой рубеже. У этого вида Wasp работники являются самыми маленькими, мужские работники немного больше, а женские королевы значительно больше, чем ее работники, и мужские коллеги. [ Цитация необходима ]
Эволюция
[ редактировать ]
В 1871 году Чарльз Дарвин выдвинул теорию сексуального отбора , которая связывала половой диморфизм с сексуальным отбором . [ 144 ]
Первым шагом к сексуальному диморфизму является размер дифференциация сперматозоидов и яиц ( анизогамия ). [ 145 ] [ 146 ] [ 147 ] [ 148 ] : 917 Анизогамия и обычно большое количество небольших мужских гамет по сравнению с более крупными женскими гаметами, как правило, заключается в разработке сильной конкуренции спермы , [ 149 ] [ 150 ] Поскольку небольшая сперма позволяет организмам производить большое количество сперматозоидов и создавать мужчин (или мужская функция гермафродитов [ 151 ] ) более избыточно.
Вольвоциновые водоросли были полезны для понимания эволюции сексуального диморфизма [ 152 ] и такие виды, как жук C. maculatus , где женщины больше, чем мужчины, используются для изучения его основных генетических механизмов. [ 153 ]
У многих немоногамных видов польза для репродуктивной пригодности для спаривания мужчины с несколькими женщинами является большим, тогда как польза для репродуктивной подготовки спаривания самки с несколькими мужчинами невелика или не существует. [ 154 ] У этих видов существует давление отбора на любые признаки, позволяющие мужчине иметь больше спасательных явлений. Поэтому у мужчины могут быть разные черты от женщины.

Эти черты могут быть теми, которые позволяют ему бороться с другими мужчинами за контроль над территорией или гаремом , таким как большой размер или оружие; [ 155 ] Или они могут быть чертами, что женщины, по какой -то причине, предпочитают в товарищах. [ 156 ] Соревнования по мужчинам -мужчинам не представляет глубоких теоретических вопросов [ 157 ] Но Mate Choice делает.
Женщины могут выбирать мужчин, которые кажутся сильными и здоровыми, поэтому, вероятно, обладают «хорошими аллелями » и приводят к здоровому потомству. [ 158 ] Однако у некоторых видов женщины, похоже, выбирают мужчин с чертами, которые не улучшают выживаемость потомства, и даже черты, которые уменьшают его (потенциально приводят к таким признакам, как хвост павлина). [ 157 ] Две гипотезы для объяснения этого факта - это гипотеза сексуального сына и принцип гандикапа .
Гипотеза сексуального сына гласит, что женщины могут изначально выбрать черту, потому что она улучшает выживание их молодых, но как только это предпочтение станет широко распространенным, женщины должны продолжать выбирать черту, даже если это становится вредным. У тех, у кого нет сыновей, которые непривлекательны для большинства женщин (поскольку предпочтение широко распространено) и поэтому получают несколько спасений. [ 159 ]
Принцип гандикапа гласит, что мужчина, который выживает, несмотря на то, что обладает каким -то гандикапом, доказывает, что остальные его гены являются «хорошими аллелями». Если мужчины с «плохими аллелями» не могут выжить в гандикапе, женщины могут развиваться, чтобы выбрать мужчин с такого рода гандикапом; Черта действует как трудный сигнал пригодности. [ 160 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Энциклопедия поведения животных . Тол. 2. Академическая пресса. 21 января 2019 г. с. 7. ISBN 978-0-12-813252-4 .
- ^ Ralls K, Mesnick S (2009). «Сексуальный диморфизм» . Академическая пресса . 1 (1): 1005–1011. doi : 10.1016/b978-0-12-373553-9.00233-9 . ISBN 9780123735539 .
- ^ «Словарь эволюции и биологии человека» . Человеческая биология . Архивировано с оригинала 7 ноября 2017 года . Получено 3 ноября 2017 года .
- ^ Armenta JK, Dunn Po, Whittingham LA (август 2008 г.). «Количественная оценка сексуального дихроматизма птиц: сравнение методов» . Журнал экспериментальной биологии . 211 (Pt 15): 2423–30. doi : 10.1242/jeb.013094 . PMID 18626076 .
- ^ Андерссон 1994 , с
- ^ Захави А (сентябрь 1975 г.). «Мат-выбор-выбор для гандикапа» (PDF) . Журнал теоретической биологии . 53 (1): 205–14. Bibcode : 1975jthbi..53..205z . Citeseerx 10.1.1.586.3819 . doi : 10.1016/0022-5193 (75) 90111-3 . PMID 1195756 . Архивировано из оригинала (PDF) 10 августа 2017 года . Получено 14 мая 2011 года .
- ^ Подпрыгнуть до: а беременный Zi J, Yu X, Li Y, Hu X, Xu C, Wang X, et al. (Октябрь 2003 г.). «Стратегии окрашивания в павлине перья» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (22): 12576–8. Bibcode : 2003pnas..10012576Z . doi : 10.1073/pnas.2133313100 . PMC 240659 . PMID 14557541 .
- ^ «Птицы парадиса: короли красоты» . Национальное географическое общество. 19 октября 2023 года . Получено 22 ноября 2023 года .
- ^ Slagsvold T, Lifjeld JT (1985). «Различия в цвете оперения великолепного Parus Major в отношении среды обитания, сезона и еды». Журнал зоологии . 206 (3): 321–328. doi : 10.1111/j.1469-7998.1985.tb05661.x .
- ^ Bowmaker JK, Heath LA, Wilkie SE, Hunt DM (август 1997 г.). «Визуальные пигменты и капли масла из шести классов фоторецептора в сетчатке птиц» . Видение исследования . 37 (16): 2183–94. doi : 10.1098/rspb.1998.0315 . PMC 1688915 . PMID 9578901 .
- ^ Bowmaker JK, Heath LA, Wilkie SE , Hunt DM (август 1997 г.). «Визуальные пигменты и капли масла из шести классов фоторецептора в сетчатке птиц» . Видение исследования . 37 (16): 2183–94. doi : 10.1098/rspb.1998.0316 . JSTOR 50814 . PMC 1688906 . PMID 9578901 .
- ^ Senar JC, Figuerola J, Pascual J (февраль 2002 г.). «Яркие желтые синие сиськи делают лучших родителей» . Разбирательство. Биологические науки . 269 (1488): 257–61. doi : 10.1098/rspb.2001.1882 . PMC 1690890 . PMID 11839194 .
- ^ Johnsen A, Delhey K, Andersson S, Kempenaers B (июнь 2003 г.). «Цвет оперения в синих сиськах: сексуальный дихроматизм, зависимость от состояния и генетические эффекты» . Разбирательство. Биологические науки . 270 (1521): 1263–70. doi : 10.1098/rspb.2003.2375 . JSTOR 3558810 . PMC 1691364 . PMID 12816639 .
- ^ Лозано Г.А. (1994). «Каротиноиды, паразиты и половой отбор» (PDF) . Оикос . 70 (2): 309–311. Bibcode : 1994oikos..70..309L . doi : 10.2307/3545643 . JSTOR 3545643 .
- ^ Donnellan, SC, & Mahony, MJ (2004). Аллезим, хромосомная и морфологическая изменчивость в группе видов LeSueuri (Anura: Hylidae), включая описание нового вида. Австралийский журнал зоологии
- ^ Bell, RC, & Zamudio, KR (2012). Сексуальный дихроматизм у лягушек: естественный отбор, сексуальный отбор и неожиданное разнообразие. Труды Королевского общества B: Биологические науки.
- ^ Райан М.Дж., Рэнд как (апрель 1993 г.). «Распознавание видов и сексуальный отбор как унитарная проблема в общении с животными». Эволюция; Международный журнал органической эволюции . 47 (2): 647–657. doi : 10.2307/2410076 . JSTOR 2410076 . PMID 28568715 .
- ^ Рубелини Д., Спина Ф., Сайно Н. (2004). «Протейндри и сексуальный диморфизм у перелетных птиц Транс-Сахары». Поведенческая экология . 15 (4): 592–601. Citeseerx 10.1.1.498.7541 . doi : 10.1093/beheco/arh048 .
- ^ Короткий Р.В., Балабан Е (4 августа 1994 г.). Различия между полами . Издательство Кембриджского университета. ISBN 9780521448789 Полем Получено 3 ноября 2017 года - через Google Books.
- ^ Джакомелло Е., Маркини Д., Расотто М.Б. (сентябрь 2006 г.). «Мужская сексуальная диморфная черта обеспечивает антимикробные препараты яиц в бренни -рыбе» . Биологические письма . 2 (3): 330–3. doi : 10.1098/rsbl.2006.0492 . PMC 1686180 . PMID 17148395 .
- ^ Renner SS, Ricklefs RE (1995). «Dioecy и ее корреляты в цветущих растениях» . Американский журнал ботаники . 82 (5): 596–606. doi : 10.2307/2445418 . JSTOR 2445418 .
- ^ Behnke H, Lüttge U, Esser K, Kadereit JW, Runge M (6 декабря 2012 г.). Прогресс в ботанике / Прогресс ботаники: структурная ботаника Физиология Генетика Таксономия Геоботаника / Структура Физиология Генетика Систематика Геоботания . Springer Science & Business Media. ISBN 978-3-642-79844-3 .
- ^ Рамават К.Г., Мериллон Дж. М., Шиванна К.Р. (19 апреля 2016 г.). Репродуктивная биология растений . CRC Press. ISBN 978-1-4822-0133-8 .
- ^ Ромеро Г.А., Нельсон К.Е. (июнь 1986 г.). «Сексуальный диморфизм в орхидеях катазетки: насильственное размещение пыльцы и конкуренция мужского цветов». Наука . 232 (4757): 1538–40. Bibcode : 1986sci ... 232.1538r . doi : 10.1126/science.232.4757.1538 . JSTOR 1698050 . PMID 17773505 . S2CID 31296391 .
- ^ «Угорчатая трава (он же дикий сельдерей, ленточная трава)» . Университет Массачусетса . Архивировано из оригинала 12 июля 2011 года.
- ^ Фридман Дж., Барретт С.С. (июнь 2009 г.). «Ветер перемен: новое понимание экологии и эволюции опыления и спаривания у насыщенных ветряными растениями» . Анналы ботаники . 103 (9): 1515–27. doi : 10.1093/aob/mcp035 . PMC 2701749 . PMID 19218583 .
- ^ Гебер М.А. (1999). Пол и половой диморфизм в цветущих растениях . Берлин: Спрингер. ISBN 978-3-540-64597-9 Полем п. 206
- ^ Бондьюрианский Р (январь 2007 г.). «Эволюция зависимого от состояния полового диморфизма». Американский натуралист . 169 (1): 9–19. doi : 10.1086/510214 . PMID 17206580 . S2CID 17439073 .
- ^ Barreto FS, Avise JC (август 2011 г.). «Система генетического спаривания морского паука с диморфизмом сексуального размера, представленного мужчинами: доказательство искажения отцовства, несмотря на случайный успех спаривания» . Поведенческая экология и социобиология . 65 (8): 1595–1604. Bibcode : 2011becos..65.1595b . doi : 10.1007/s00265-011-1170-x . PMC 3134710 . PMID 21874083 .
- ^ Gruber B, Eckel K, Everaars J, Dormannn CF (30 июня 2011 г.). «На управлении пчелой Red Mason (Osmia bicornis) в яблочных садах» (PDF ) апидология 42 (5): 564–5 Doi : 10.1007/s13592-011-0059- z ISSN 0044-8 22935710S2CID
- ^ «Hackberry Emperor - Asterocampa Celtis (Boisduval & Leconte)» . entnemdept.ufl.edu . Получено 15 ноября 2017 года .
- ^ Rust R, Torchio P, Trostle G (1989). «Поздний эмбриогенез и незрелое развитие Osmia Rufa Cornigera (Rossi) (Hymenoptera: Megachilidae)» . Апидология . 20 (4): 359–367. doi : 10.1051/apido: 19890408 .
- ^ Danforth B (1991). «Морфология и поведение диморфных мужчин у Perdita portalis (Hymenoptera: Andrenidae)». Поведенческая экология и социобиология . 29 (4): 235–247. Bibcode : 1991becos..29..235d . doi : 10.1007/bf00163980 . S2CID 37651908 .
- ^ Jaycox Elbert R (1967). «Территориальное поведение среди мужчин Антидия Бамнгенса». Журнал Канзасского энтомологического общества . 40 (4): 565–570.
- ^ Kukuk PF (1 октября 1996 г.). «Мужской диморфизм в ласиоглоссме (чилаликт) полухлице: роль личиночного питания». Журнал Канзасского энтомологического общества . 69 (4): 147–157. JSTOR 25085712 .
- ^ Paxton RJ, Giovanetti F, Scamoni E, Scanni B (1 октября 1999 г.). «Спаривание в коммунальной пчеле, Андена ругает (Hymenoptera andreniidae)» ЭКОЛОГИЯ ИТОЛОГИЯ И ЭВОЛЮЦИЯ . 11 (4): 371–3 Bibcode : 19999tece..11..371p Doi : 10.1080/ 08927014.1999.9522 ISSN 0394-9
- ^ Wang MQ, Yang D (2005). «Сексуальный диморфизм у насекомых». Китайский бюллетень энтомологии . 42 : 721–725.
- ^ Подпрыгнуть до: а беременный Sugiura S, Yamaura Y, Makihara H (ноябрь 2007 г.). «Сексуальный и мужской диморфизм рога у Copris Ochus (Coleoptera: Scarabaeidae)» . Зоологическая наука . 24 (11): 1082–1085. doi : 10.2108/zsj.24.1082 . PMID 18348608 . S2CID 34705415 .
- ^ Хонго, Йошихито (1 декабря 2007 г.). «Эволюция мужской диморфической аллометрии в популяции японской рогатой жука Трипоксил Дихотомис Septentrionalis» . Поведенческая экология и социобиология . 62 (2): 245–253. Bibcode : 2007becos..62..245h . doi : 10.1007/s00265-007-0459-2 . ISSN 1432-0762 .
- ^ Подпрыгнуть до: а беременный Emlen DJ, Marangelo J, Ball B, Cunningham CW (май 2005). «Разнообразие в оружии сексуального отбора: эволюция рога в роде жуков Onthophagus (Coleoptera: Scarabaeidae)». Эволюция; Международный журнал органической эволюции . 59 (5): 1060–1084. Citeseerx 10.1.1.133.7557 . doi : 10.1111/j.0014-3820.2005.tb01044.x . PMID 16136805 . S2CID 221736269 .
- ^ Teder, T. & Tammaru, T. (2005). «Диморфизм сексуального размера внутри видов увеличивается с размером тела у насекомых». Оикос [ ISBN отсутствует ]
- ^ Оливер Дж.С., Монтейро А (июль 2011 г.). «О происхождении сексуального диморфизма у бабочек» . Разбирательство. Биологические науки . 278 (1714): 1981–1988. doi : 10.1098/rspb.2010.2220 . PMC 3107650 . PMID 21123259 .
- ^ Робертсон Ка, Монтейро А (август 2005 г.). «Женский Bicyclus Anynana Butterflies выбирают мужчин на основе своих спинных ультрафиолетовых учеников глаз» . Разбирательство. Биологические науки . 272 (1572): 1541–1546. doi : 10.1098/rspb.2005.3142 . PMC 1559841 . PMID 16048768 .
- ^ Wiklund C, Lindfors V, Forsberg J (1996). «Раннее появление мужчин и репродуктивная фенология взрослого взрослого, заинтересованного в бабочке Godpteryx Rhamni в Швеции». Оикос . 75 (2): 227–240. Bibcode : 1996oikos..75..227w . doi : 10.2307/3546246 . JSTOR 3546246 .
- ^ Kunte K (июль 2008 г.). «Миметические бабочки поддерживают модель сексуального диморфизма Уоллеса» . Разбирательство. Биологические науки . 275 (1643): 1617–1624. doi : 10.1098/rspb.2008.0171 . PMC 2602815 . PMID 18426753 .
- ^ McLean CJ, Garwood RJ, Brassey CA (2018). «Сексуальный диморфизм в арахнидных порядках» . ПЕРЕЙ . 6 : E5751. doi : 10.7717/peerj.5751 . PMC 6225839 . PMID 30416880 .
- ^ Смит Т. Обнаружение повседневной схемы активности Zygiella x-notata и его связь с светом (PDF) (тезис MS).
- ^ Prenter J, Elwood RW, Montgomery WI (декабрь 1999 г.). «Диморфизм сексуального размера и репродуктивные инвестиции женщин -пауков: сравнительный анализ». Эволюция; Международный журнал органической эволюции . 53 (6): 1987–1994. doi : 10.2307/2640458 . JSTOR 2640458 . PMID 28565440 .
- ^ Подпрыгнуть до: а беременный в Wilder SM, Rypstra AL (2008). «Диморфизм сексуального размера опосредует возникновение зависимого от состояния сексуального каннибализма у паука волка». Поведение животных . 76 (2): 447–454. doi : 10.1016/j.anbehav.2007.12.023 . S2CID 54373571 .
- ^ Foellmer MW, Fairbairn DJ (2004). «Мужчины, находящиеся в атаке: сексуальный каннибализм и его последствия для мужской морфологии и поведения в пауке с шаром» . Эволюционные экологические исследования . 6 : 163–181.
- ^ Girard MB, Elias Do, Kasumovic MM (декабрь 2015 г.). «Женское предпочтение мультимодальному ухаживанию: множественные сигналы важны для успеха спаривания мужского пола у пауковых пауков» . Разбирательство. Биологические науки . 282 (1820): 20152222. DOI : 10.1098/rspb.2015.2222 . PMC 4685782 . PMID 26631566 .
- ^ Fairbairn D (28 апреля 2013 г.). Странные пары: необычные различия между полами в животном царстве . Принстон. ISBN 978-0691141961 .
- ^ Ota K, Kohda M, Sato T (июнь 2010 г.). «Необычная аллометрия для диморфизма сексуального размера в цихлиде, где мужчины чрезвычайно больше женщин». Журнал биологических наук . 35 (2): 257–65. doi : 10.1007/s12038-010-0030-6 . PMID 20689182 . S2CID 12396902 .
- ^ Сато Т (1994). «Активное накопление нерестового субстрата: определяет экстремального полигинья в цихлидской рыбе, выращивающей раковину». Поведение животных . 48 (3): 669–678. doi : 10.1006/anbe.1994.1286 . S2CID 53192909 .
- ^ Schütz D, Taborsky M (2005). . ( "Аранаета PDF ) Журнал арахнологии 33 (3): 767–775. doi : 10.1636/s03-56.1 . S2CID S2CID the original (PDF)Марш Получено 14 мая
- ^ McCormick Mi, Ryen CA, Munday PL, Walker SP (май 2010). Бриффа М (ред.). «Различные механизмы лежат в основе сексуального диморфизма по размеру в двух популяциях рыб, изменяющей пол» . Plos один . 5 (5): E10616. Bibcode : 2010ploso ... 510616M . doi : 10.1371/journal.pone.0010616 . PMC 2868897 . PMID 20485547 .
- ^ Warner RR (июнь 1988 г.). «Изменение пола и модель по размеру». Тенденции в экологии и эволюции . 3 (6): 133–6. Bibcode : 1988tecoe ... 3..133W . doi : 10.1016/0169-5347 (88) 90176-0 . PMID 21227182 .
- ^ Адамс С., Уильямс А.Дж. (2001). «Предварительный тест гипотезы переходного роста с использованием протонитной коралловой форели Plectropomus maculatus ». Журнал рыбной биологии . 59 (1): 183–185. Bibcode : 2001jfbio..59..183a . doi : 10.1111/j.1095-8649.2001.tb02350.x .
- ^ Хендри А., Берг ОК (1999). «Вторичные сексуальные персонажи, использование энергии, старение и стоимость воспроизведения в лососе в Sockeye». Канадский журнал зоологии . 77 (11): 1663–1675. doi : 10.1139/cjz-77-11-1663 .
- ^ Подпрыгнуть до: а беременный Amundsen T, Forsgren E (ноябрь 2001 г.). «Выбор мужского пола выбирает женскую окраску в рыбе» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (23): 13155–60. Bibcode : 2001pnas ... 9813155a . doi : 10.1073/pnas.211439298 . PMC 60840 . PMID 11606720 .
- ^ Подпрыгнуть до: а беременный Svensson PA, Pélabon C, Blount JD, Surai PF, Amundsen T (2006). «Отражает ли женская брачная окраска каротиноидов яиц и качества сцепления в Goby с двумя пятнами (Gobiusculus flavescens, Gobiidae)?» Полем Функциональная экология . 20 (4): 689–698. Bibcode : 2006fueco..20..689s . doi : 10.1111/j.1365-2435.2006.01151.x . HDL : 10536/dro/du: 30038904 .
- ^ Батлер М.А., Schoener TW, Losos JB (февраль 2000 г.). «Взаимосвязь между диморфизмом сексуального размера и использованием среды обитания в больших антильских анолис -ящерицах» . Эволюция; Международный журнал органической эволюции . 54 (1): 259–72. doi : 10.1111/j.0014-3820.2000.tb00026.x . PMID 10937202 . S2CID 7887284 .
- ^ Sanger TJ, Seav SM, Tokita M, Langerhans RB, Ross LM, Losos JB, Abzhanov A (июнь 2014 г.). «Путь эстрогена лежит в основе эволюции преувеличенных мужских черепных форм у ящериц анолиса» . Разбирательство. Биологические науки . 281 (1784): 20140329. DOI : 10.1098/rspb.2014.0329 . PMC 4043096 . PMID 24741020 .
- ^ Подпрыгнуть до: а беременный Pinto, A., Wiederhecker, H. и Neck, G. (2005). Сексуальный диморфизм в неотропической ящерице, Tropidurus (Squamata, Tropiduridae). Амфибия-рептилии.
- ^ Подпрыгнуть до: а беременный Олссон М., Тоблер М., Хили М., Перрин С., Уилсон М. (август 2012 г.). «Значительный компонент старения (повреждение ДНК) отражается в исчезающих цветах размножения: экспериментальный тест с использованием врожденных антиоксидантных миметиков у окрашенных ящериц драконов» . Эволюция; Международный журнал органической эволюции . 66 (8): 2475–83. doi : 10.1111/j.1558-5646.2012.01617.x . PMID 22834746 . S2CID 205783815 .
- ^ Caron FS, Pie MR (2024). «Макроэволюция диморфизма сексуального размера у птиц» . Биологический журнал Линневого общества . 141 (5): в печати. doi : 10.1093/biolinnean/blad168 .
- ^ Bravo CR, Bautista-Sopelana LM, Alonso JC (2024). «Пересмотр гипотезы дивергенции ниши у сексуально диморфных птиц: перекрывается ли диета с диморфизмом сексуального размера?». Журнал экологии животных . 93 (4): 460–474. Bibcode : 2024Janec..93..460b . doi : 10.1111/1365-2656.14058 . HDL : 10261/360371 . PMID 38462717 .
- ^ Андерссон 1994 , с
- ^ Berns CM, Adams DC (11 ноября 2012 г.). «Стать другим, но оставаться одинаково: модели сексуального размера и диморфизма формы в счетах колибри». Эволюционная биология . 40 (2): 246–260. doi : 10.1007/s11692-012-9206-3 . ISSN 0071-3260 . S2CID 276492 .
- ^ McGraw KJ, Hill GE, Stradi R, Parker RS (февраль 2002 г.). «Влияние доступа к каротиноидам в рационе на сексуальное дихроматизм и пигментную композицию оперения в американском Goldfinch» (PDF) . Сравнительная биохимия и физиология. Часть B, Биохимия и молекулярная биология . 131 (2): 261–9. doi : 10.1016/s1096-4959 (01) 00500-0 . PMID 11818247 . Архивировано из оригинала (PDF) 28 августа 2005 года.
- ^ Gibbs HL, Weatherhead PJ, Boag PT, White BN, Tabak LM, Hoysak DJ (декабрь 1990 г.). «Реализованный репродуктивный успех полигинозных красных крыльев, выявленных маркерами ДНК» . Наука . 250 (4986): 1394–7. doi : 10.1098/rspb.1998.0308 . JSTOR 50849 . PMC 1688905 . PMID 17754986 .
- ^ Подпрыгнуть до: а беременный в дюймовый Линдси В.Р., Вебстер М.С., Вариан К.В., Швабл Х (2009). «Приобретение и поведение цвета оперения связаны с андрогенами в фенотипически пластической птице». Поведение животных . 77 (6): 1525–1532. doi : 10.1016/j.anbehav.2009.02.027 . S2CID 15799876 .
- ^ Петри М (1994). «Улучшенный рост и выживание потомков павлина с более сложными поездами». Природа . 371 (6498): 598–599. Bibcode : 1994natur.371..598p . doi : 10.1038/371598a0 . S2CID 4316752 .
- ^ Рубелини Д., Спина Ф., Сайно Н. (2004). «Протейндри и сексуальный диморфизм у перелетных птиц Транс-Сахары» . Поведенческая экология . 15 (4): 592–601. doi : 10.1093/beheco/arh048 .
- ^ Kissner KJ, Weatherhead PJ, Francis CM (январь 2003 г.). «Сексуальный размер диморфизм и сроки весенней миграции у птиц». Журнал эволюционной биологии . 16 (1): 154–62. Citeseerx 10.1.1.584.2867 . doi : 10.1046/j.1420-9101.2003.00479.x . PMID 14635890 . S2CID 13830052 .
- ^ Подпрыгнуть до: а беременный Møller AP, Nielsen JT (2006). «Уязвимость добычи в отношении сексуальной окраски добычи». Поведенческая экология и социобиология . 60 (2): 227–233. Bibcode : 2006becos..60..227m . doi : 10.1007/s00265-006-0160-x . S2CID 36836956 .
- ^ Подпрыгнуть до: а беременный в Адкинс-Реган Э. (2007). «Гормоны и развитие половых различий в поведении». Журнал орнитологии . 148 (Дополнение 1): S17 - S26. doi : 10.1007/S10336-007-0188-3 . S2CID 13868097 .
- ^ Подпрыгнуть до: а беременный в дюймовый Martin U, Grüebler HS, Müller M, Spaar R, Horch P, Naef-Daenzer B (2008). «Предвзяточная смертность женщин, вызванная антропогенной потерей гнезда, способствует снижению численности населения и соотношению полов взрослых у птицы луга». Биологическое сохранение . 141 (12): 3040–3049. Bibcode : 2008bcons.141.3040g . doi : 10.1016/j.biocon.2008.09.008 .
- ^ Owens, Ipf, Short, RV,. (1995). Гормональная основа сексуального диморфизма у птиц: последствия для новых теорий сексуального отбора. Тенденции в экологии и эволюции., 10 (Ref), 44.
- ^ Подпрыгнуть до: а беременный Койн Дж.А., Кей Эх, Прутт-Джонс (январь 2008 г.). «Генетическая основа сексуального диморфизма у птиц» . Эволюция; Международный журнал органической эволюции . 62 (1): 214–9. doi : 10.1111/j.1558-5646.2007.00254.x . PMID 18005159 . S2CID 11490688 .
- ^ Веландо А (2002). «Экспериментальная манипуляция с материнскими усилиями дает дифференциальные эффекты у сыновей и дочерей: последствия для адаптивных соотношений полов в сине-ногах» . Поведенческая экология . 13 (4): 443–449. doi : 10.1093/beheco/13.4.443 .
- ^ Loostra AJ, Verhoeven MA, Piersma T (2018). «Специфичный рост у цыплят из сексуально-диморфического чернохвового Годвита» (PDF) . Ибис . 160 (1): 89–100. doi : 10.1111/ibi.12541 . S2CID 90880117 .
- ^ Подпрыгнуть до: а беременный Главный MB (март 2008 г.). «Примирение конкурирующих экологических объяснений сексуальной сегрегации в цветах». Экология . 89 (3): 693–704. Bibcode : 2008ecol ... 89..693m . doi : 10.1890/07-0645.1 . PMID 18459333 .
- ^ Подпрыгнуть до: а беременный Сафи К., Кениг Б., Керт Г. (2007). «Половые различия в генетике популяции, размере домашнего диапазона и использовании среды обитания летучей мыши партий ( Vespertilio Murinus , Linnaeus 1758) в Швейцарии и их последствия для сохранения» (PDF) . Биологическое сохранение . 137 (1): 28–36. Bibcode : 2007bcons.137 ... 28 с . doi : 10.1016/j.biocon.2007.01.011 . Архивировано из оригинала (PDF) 25 сентября 2017 года . Получено 2 февраля 2019 года .
- ^ Coulson G, Macfarlane AM, Parsons SE, Cutter J (2006). «Эволюция сексуальной сегрегации у травоядных млекопитающих: кенгуру как сумчатые модели». Австралийский журнал зоологии . 54 (3): 217–224. doi : 10.1071/Zo05062 .
- ^ González-Solís J, Croxall JP, Wood AG (2000). «Сексуальный диморфизм и сексуальная сегрегация в стратегиях кормления северной гигантской бенров, Macranectes Halli , во время инкубации». Оикос . 90 (2): 390–398. Bibcode : 2000oikos..90..390g . doi : 10.1034/j.1600-0706.2000.900220.x .
- ^ Подпрыгнуть до: а беременный в дюймовый Саммерс-Смит Д.Д. (1988). Воробьи . Калтон, Стаффордшир, Великобритания: T. & Ad Poyser . ISBN 978-0-85661-048-6 .
- ^ Майорино, Леонардо; Фарке, Эндрю А.; Кочакис, Тассос; Пирас, Паоло (7 мая 2015 г.). «Мужчины напоминают женщин: переоценка сексуального диморфизма в протокератопах Эндрюси (Неоцератопсия, Protoceratopsidae)» . Plos один . 10 (5): E0126464. doi : 10.1371/journal.pone.0126464 . ISSN 1932-6203 . PMC 4423778 . PMID 25951329 .
- ^ Сатта, Эван Томас (22 апреля 2015 г.). «Свидетельство о сексуальном диморфизме в высеченном динозавре Стегозавр Мьоси (Ornithischia, Stegosauria) из формирования Morrison (Верхняя юрская величина) западной США» . Plos один . 10 (4): E0123503. BIBCODE : 2015PLOSO..1023503S . doi : 10.1371/journal.pone.0123503 . ISSN 1932-6203 . PMC 4406738 . PMID 25901727 .
- ^ «Стегозавры могли различаться между мужчинами, женщинами» . Scienceday . Получено 12 марта 2024 года .
- ^ Арнольд А.П. (сентябрь 2004 г.). «Секс -хромосомы и пол мозга». Природные обзоры. Нейробиология . 5 (9): 701–8. doi : 10.1038/nrn1494 . PMID 15322528 . S2CID 7419814 .
- ^ Кассини М.Х. (январь 2020 г.). «Смешанная модель эволюции полигинья и диморфизма сексуального размера у млекопитающих». Обзор млекопитающих . 50 (1): 112–120. doi : 10.1111/mam.12171 . ISSN 0305-1838 . S2CID 208557639 .
- ^ Cappozzo HL, Campagna C, Monserrat J (1991). «Сексуальный диморфизм у новорожденных южных морских львов». Морские млекопитающие . 7 (4): 385–394. Bibcode : 1991mms ... 7..385c . doi : 10.1111/j.1748-7692.1991.tb00113.x .
- ^ Салогни Е., Галимберти Ф., Санвито С., Миллер Э.Х. (март 2019 г.). «Мужские и женские щенки очень сексуально диморфного уплотнения северного слона (Mirounga angustirostris) слегка различаются по размеру тела». Канадский журнал зоологии . 97 (3): 241–250. doi : 10.1139/cjz-2018-0220 . ISSN 0008-4301 . S2CID 91796880 .
- ^ Оно Ка, Бонесс DJ (январь 1996 г.). «Сексуальный диморфизм у щенков морских львов: дифференциальные материнские инвестиции или специфические для пола различия в распределении энергии?». Поведенческая экология и социобиология . 38 (1): 31–41. Bibcode : 1996becos..38 ... 31o . doi : 10.1007/s002650050214 . S2CID 25307359 .
- ^ Tarnawski BA, Cassini GH, Flores DA (2014). «Аллометрия черепа и половой диморфизм в онтогенезе южного слона (Мирунга Леонина)». Канадский журнал зоологии . 31 : 19–31. doi : 10.1139/cjz-2013-0106 . HDL : 11336/29893 .
- ^ Ларсен CS (август 2003 г.). «Равенство для полов в эволюции человека? Ранний сексуальный диморфизм гоминидов и последствия для систем спаривания и социального поведения» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (16): 9103–4. Bibcode : 2003pnas..100.9103L . doi : 10.1073/pnas.1633678100 . PMC 170877 . PMID 12886010 .
- ^ Buss DM (2007). «Эволюция спаривания человека» (PDF) . Acta Psychologica Sinica . 39 (3): 502–512. Архивировано из оригинала (PDF) 12 августа 2011 года . Получено 14 мая 2011 года .
- ^ Дейли М., Уилсон М. (1996). «Эволюционная психология и семейный конфликт». В Бусс Д.М. , Маламут Н.М. (ред.). Секс, власть, конфликт: эволюционные и феминистские перспективы . Издательство Оксфордского университета . п. 13 ISBN 978-0-19-510357-1 .
- ^ Реттнер, Рэйчел (3 января 2014 г.). «Почему подтягивания труднее для женщин» . Livescience .
- ^ Sparling PB, O'Donnell EM, Snow TK (декабрь 1998 г.). «Гендерная разница в производительности на расстоянии проходила плато: анализ мировых рейтингов с 1980 по 1996 год» . Медицина и наука в спорте и упражнениях . 30 (12): 1725–9. doi : 10.1097/00005768-199812000-00011 . PMID 9861606 .
- ^ «Национальная статистика здравоохранения» (PDF) . Национальные отчеты о статистике здравоохранения . 10 22 октября 2008 г. Получено 21 апреля 2012 года .
- ^ «Национальное обследование здоровья и питания США, 1999–2002 гг.» (PDF) . Получено 1 мая 2014 года .
- ^ Подпрыгнуть до: а беременный Глюксман А (1981). Сексуальный диморфизм в биологии и патологии человека и млекопитающих . Академическая пресса . С. 66–75. ISBN 978-0-12-286960-0 Полем OCLC 7831448 .
- ^ Подпрыгнуть до: а беременный Танец А (27 марта 2019 г.). «Почему полов не чувствуют боли одинаково» . Природа . 567 (7749): 448–450. Bibcode : 2019natur.567..448d . doi : 10.1038/d41586-019-00895-3 . PMID 30918396 . S2CID 85527866 .
- ^ Durden-Smith J, Desimone D (1983). Секс и мозг . Нью -Йорк: Arbor House . ISBN 978-0-87795-484-2 .
- ^ Герш Э.С. , Герш I (1981). Биология женщин . Тол. 306. Балтимор: Университетский парк Пресс (оригинал из Мичиганского университета). п. 511. Bibcode : 1983natur.306..511. Полем doi : 10.1038/306511b0 . ISBN 978-0-8391-1622-6 Полем S2CID 28060318 .
{{cite book}}:|journal=игнорируется ( помощь ) - ^ Stein JH (1987). Внутренняя медицина (2 -е изд.). Бостон: Маленький, Браун . ISBN 978-0-316-81236-8 .
- ^ McLaughlin M, Shryer T (8 августа 1988 г.). «Мужчины против женщин: новые дебаты о сексуальных различиях». US News & World Report : 50–58.
- ^ McEwen BS (март 1981 г.). «Нейронные гонадные стероидные действия». Наука . 211 (4488): 1303–11. Bibcode : 1981sci ... 211.1303M . doi : 10.1126/science.6259728 . PMID 6259728 .
- ^ «Острая терпимость к боли со временем более последовательно у женщин, чем мужчин, согласно новым исследованиям» . NCCIH . Получено 11 мая 2022 года .
- ^ Возняки К. «Устойчивость к боли и чувствительность у мужчин, женщин, рыжеволоса и других» . Webmd . Получено 11 мая 2022 года .
- ^ Лопес А.М., Росс Н., Закрытие Дж., Дагналл А., Аморим А., Кроу Т.Дж. (апрель 2006 г.). «Статус инактивации PCDH11X: половые диморфизмы в уровнях экспрессии генов в мозге». Человеческая генетика . 119 (3): 267–75. doi : 10.1007/s00439-006-0134-0 . PMID 16425037 . S2CID 19323646 .
- ^ Lombardo MV, Ashwin E, Auyeung B, Chakrabarti B, Taylor K, Hackett G, et al. (Январь 2012 г.). «Тестостерон плода влияет на сексуально диморфное серое вещество в человеческом мозге» . Журнал нейробиологии . 32 (2): 674–80. doi : 10.1523/jneurosci.4389-11.2012 . PMC 3306238 . PMID 22238103 .
- ^ «Разнообразные роли для связывающего полового гормонов глобулина в репродукции» . biolreprod.org . Архивировано с оригинала 23 сентября 2015 года.
- ^ Fine C (август 2010 г.). Благословения пола: как наш разум, общество и неврозексизм создают разницу (1 -е изд.). WW Norton & Company . ISBN 978-0-393-06838-2 .
- ^ Jordan-Young R (сентябрь 2010 г.). Brain Storm: недостатки в науке о сексуальных различиях . Гарвардский университет издательство . ISBN 978-0-674-05730-2 .
- ^ Marner L, Nyengaard Jr, Tang Y, Pakkenberg B (июль 2003 г.). «Отмеченная потеря миелинизированных нервных волокон в человеческом мозге с возрастом». Журнал сравнительной неврологии . 462 (2): 144–52. doi : 10.1002/cne.10714 . PMID 12794739 . S2CID 35293796 .
- ^ Gur RC, Turetsky Bi, Matsui M, Yan M, Bilker W, Hughett P, Gur Re (май 1999). «Половые различия в мозговом сером и белом веществе у здоровых молодых людей: корреляции с когнитивными показателями» . Журнал нейробиологии . 19 (10): 4065–72. doi : 10.1523/jneurosci.19-10-04065.1999 . PMC 6782697 . PMID 10234034 .
- ^ Леонард С.М., Таулер С., Добро пожаловать С., Халдерман Л.К., Отто Р., Экерт М.А., Чиарелло С (декабрь 2008 г.). «Размер вопросов: церебральный объем влияет на половые различия в нейроанатомии» . Кора головного мозга . 18 (12): 2920–31. doi : 10.1093/cercor/bhn052 . PMC 2583156 . PMID 18440950 .
- ^ Людерс Э., Штайнмец Х., Янке Л. (декабрь 2002 г.). «Размер мозга и объем серого вещества в здоровом человеческом мозге». Нейрорепорт . 13 (17): 2371–4. doi : 10.1097/00001756-200212030-00040 . PMID 12488829 .
- ^ Haier RJ, Jung Re, Yeo Ra, Head K, Alkire Mt (март 2005 г.). «Нейроанатомия общего интеллекта: половые вопросы» (PDF) . Нейроамиж . 25 (1): 320–7. doi : 10.1016/j.neuroimage.2004.11.019 . PMID 15734366 . S2CID 4127512 . Архивировано из оригинала (PDF) 24 мая 2010 года.
- ^ Szalkai B, Varga B, Grolmusz V (2015). «Теоретический анализ графика показывает: женские мозги лучше связаны, чем мужчины» . Plos один . 10 (7): E0130045. Arxiv : 1501.00727 . BIBCODE : 2015PLOSO..1030045S . doi : 10.1371/journal.pone.0130045 . PMC 4488527 . PMID 26132764 .
- ^ Szalkai B, Varga B, Grolmusz V (июнь 2018 г.). «У смещения размера мозга компенсированные графические теоретические параметры также лучше в структурных разъемах женщин». Визуализация мозга и поведение . 12 (3): 663–673. doi : 10.1007/s11682-017-9720-0 . PMID 28447246 . S2CID 4028467 .
- ^ Гершони М., Пьетроковский С (февраль 2017 г.). «Ландшафт сексуального транскриптома и его последующий выбор у взрослых» . BMC Biology . 15 (1): 7. doi : 10.1186/s12915-017-0352-z . PMC 5297171 . PMID 28173793 .
- ^ Гершони М., Пьетроковский С (июль 2014 г.). «Снижение отбора и накопления вредных мутаций в генах, экспрессируемых исключительно у мужчин» . Природная связь . 5 : 4438. Bibcode : 2014natco ... 5.4438g . doi : 10.1038/ncomms5438 . PMID 25014762 .
- ^ Celly CD, Stoehr AM, Nunn C, Smyth KN, Prokop ZM (декабрь 2018 г.). «Сексуальный диморфизм в иммунитете между животными: метаанализ» . Экологические письма . 21 (12): 1885–1894. BIBCODE : 2018ECOLL..21.1885K . doi : 10.1111/ele.13164 . PMID 30288910 .
- ^ Gal-OZ ST, Maier B, Yoshida H, Seddu K, Elbaz N, Czysz C, et al. (Сентябрь 2019). «Immgen Report: половой диморфизм в транскриптом иммунной системы» . Природная связь . 10 (1): 4295. Bibcode : 2019natco..10.4295g . doi : 10.1038/s41467-019-12348-6 . PMC 6754408 . PMID 31541153 .
- ^ Гроссман С. (1989). «Возможно, основные механизмы сексуального диморфизма в иммунном ответе, факте и гипотезе». Журнал стероидной биохимии . 34 (1–6): 241–251. doi : 10.1016/0022-4731 (89) 90088-5 . PMID 2696846 .
- ^ Поллилик Е (август 2013 г.). «Биология: клеточный пол имеет значение» . Природа . 500 (7460): 23–4. Bibcode : 2013natur.500 ... 23 с . doi : 10.1038/500023a . PMID 23903733 . S2CID 4318641 .
- ^ Deasy BM, Lu A, Tebbets JC, Feduska JM, Schugar RC, Pollet JB, et al. (Апрель 2007 г.). «Роль для клеточного пола в регенерации стволовых клеток, опосредованной стволовыми клетками: женские клетки имеют более высокую эффективность регенерации мышц» . Журнал клеточной биологии . 177 (1): 73–86. doi : 10.1083/jcb.200612094 . PMC 2064113 . PMID 17420291 .
- ^ Mittelstrass K, Ried JS, Yu Z, Krumsiek J, Gieger C, Prehn C, et al. (Август 2011 г.). Маккарти Ми (ред.). «Открытие сексуальных диморфизмов в метаболических и генетических биомаркерах» . PLOS Genetics . 7 (8): E1002215. doi : 10.1371/journal.pgen.1002215 . PMC 3154959 . PMID 21852955 .
- ^ Penaloza C, Estevez B, Orlanski S, Sikorska M, Walker R, Smith C, et al. (Июнь 2009 г.). «Пол клеток диктует его ответ: дифференциальная экспрессия генов и чувствительность к стрессу, вызывающему гибель клеток в мужских и женских клетках» . FASEB Journal . 23 (6): 1869–79. doi : 10.1096/fj.08-119388 . PMC 2698656 . PMID 19190082 .
- ^ Подпрыгнуть до: а беременный Vollrath F, Parker GA (1992). «Сексуальный диморфизм и искаженные половые соотношения у пауков». Природа . 360 (6400): 156–159. Bibcode : 1992natur.360..156V . doi : 10.1038/360156a0 . S2CID 4320130 .
- ^ Bornholdt R, Oliveira LR, Fabián Me (ноябрь 2008 г.). «Диморфизм сексуального размера у миотис -нигриканов (Schinz, 1821) (Chiroptera: Vespertilionidae) из Южной Бразилии» (PDF) . Бразильский журнал биологии . 68 (4): 897–904. doi : 10.1590/s1519-69842008000400028 . PMID 19197511 .
- ^ Hayssen V, Kunz TH (1996). «Аллометрия массы мусора в летучих мышах: сравнение с размером матери, морфология крыла и филогения» . Журнал млекопитающих . 77 (2): 476–490. doi : 10.2307/1382823 . JSTOR 1382823 .
- ^ Arnqvist G, Jones TM, Elgar Ma (июль 2003 г.). «Поведение насекомых: изменение сексуальных ролей в брачном кормлении» (PDF) . Природа . 424 (6947): 387. Bibcode : 2003natur.424..387a . doi : 10.1038/424387a . PMID 12879056 . S2CID 4382038 . Архивировано из оригинала (PDF) 15 сентября 2004 года.
- ^ Механизм оплодотворения: растения для людей , Под редакцией Брайана Дейла
- ^ Шоу А.Дж. (2000). «Экология населения, генетика населения и микроэволюция». В Шоу А.Дж., Гоффинет Б (ред.). Bryophyte Biology . Кембридж: издательство Кембриджского университета. С. 379–380. ISBN 978-0-521-66097-6 .
- ^ Подпрыгнуть до: а беременный Schuster RM (1984). «Сравнительная анатомия и морфология печени». Новое руководство по браизенту . Тол. 2. Ничинан, Миядзаки, Япония: Ботаническая лаборатория Хаттори. п. 891.
- ^ Crum HA, Anderson Le (1980). Мосс восточной Северной Америки . Тол. 1. Нью -Йорк: издательство Колумбийского университета. п. 196. ISBN 978-0-231-04516-2 .
- ^ Бриггс Д. (1965). «Экспериментальная таксономия некоторых британских видов рода Dicranum » . Новый фитолог . 64 (3): 366–386. doi : 10.1111/j.1469-8137.1965.tb07546.x . JSTOR 2430169 .
- ^ Умирает Alvarez ME, Rushton AW, Gozalo R, Pillola GL, Linan and, Ahlberg P (2010). « Парадоксады Brachyrhachis Linnarsson, 1883 против парадоксидов Mediterraneus pompeckj, 1901: проблематичное определение» . Дар 132 (2): 95–104. Bibcode : 2010gff ... 132 ... 95d . Doi : 10.1080/11035897.2010.481363 . S2CID 129620469 .
- ^ Падиан К, Хорнер -младший (1 ноября 2014 г.). «Сексуальный отбор Дарвина: понимание его идей в контексте» . Comptes rendus palevol . 13 (8): 709–715. Bibcode : 2014crpal..13..709p . doi : 10.1016/j.crpv.2014.09.001 . ISSN 1631-0683 .
- ^ Togashi T, Bartelt JL, Yoshimura J, Tainaka K, Cox PA (август 2012 г.). «Эволюционные траектории объясняют диверсифицированную эволюцию изогамии и анизогамии в морских зеленых водорослях» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (34): 13692–7. BIBCODE : 2012PNAS..10913692T . doi : 10.1073/pnas.1203495109 . PMC 3427103 . PMID 22869736 .
- ^ Ханшен, Эрик Р.; Херрон, Мэтью Д.; Винс, Джон Дж.; Носаки, Хисайоши; Мичод, Ричард Э. (сентябрь 2018 г.). «Многоклеточность стимулирует эволюцию сексуальных признаков» . Американский натуралист . 192 (3): E93 - E105. doi : 10.1086/698301 . ISSN 0003-0147 . PMC 6685534 . PMID 30125231 .
- ^ Сильжестам, Маттиас; Martinossi-Allibert, Ivain (2024). «Анизогамия не всегда способствует эволюции черт конкуренции спаривания у мужчин» . Американский натуралист . 203 (2): 230–253. doi : 10.1086/727968 . PMID 38306281 .
- ^ Реш, Винсент Х .; Cardé, Ring T. (22 июля 2009 г.). Энциклопедия насекомых . Академическая пресса. ISBN 978-0-08-092090-0 .
- ^ Паркер Г.А. (май 1982). «Почему так много крошечных сперматозоидов? Конкуренция спермы и поддержание двух полов». Журнал теоретической биологии . 96 (2): 281–94. Bibcode : 1982jthbi..96..281p . doi : 10.1016/0022-5193 (82) 90225-9 . PMID 7121030 . S2CID 29879237 .
- ^ Ян Дж.Н. (май 2010). «Сотрудничество и эволюция анизогамии». Журнал теоретической биологии . 264 (1): 24–36. Bibcode : 2010jthbi.264 ... 24y . doi : 10.1016/j.jtbi.2010.01.019 . PMID 20097207 .
- ^ Белл Г. (1985). «О функции цветов». Труды Королевского общества B: Биологические науки . 224 (1235): 223–266. Bibcode : 1985rspsb.224..223b . doi : 10.1098/rspb.1985.0031 . JSTOR 36033 . S2CID 84275261 .
- ^ Geng S, De Hoff P, Umen JG (июль 2014 г.). «Эволюция полов из пути спецификации сопряженного типа» . PLOS Биология . 12 (7): E1001904. doi : 10.1371/journal.pbio.1001904 . PMC 4086717 . PMID 25003332 .
- ^ Kaufmann P, Wolak Me, Husby A, Immonen E (октябрь 2021 г.). «Быстрая эволюция диморфизма сексуального размера, облегчаемого Y-связанной генетической дисперсией» . Природа экология и эволюция . 5 (10): 1394–1402. Bibcode : 2021natee ... 5.1394K . doi : 10.1038/s41559-021-01530-z . PMID 34413504 . S2CID 237242736 .
- ^ Futuyma 2005 , p. 330
- ^ Futuyma 2005 , p. 331
- ^ Futuyma 2005 , p. 332
- ^ Подпрыгнуть до: а беременный Ридли 2004 , с. 328
- ^ Futuyma 2005 , p. 335
- ^ Ридли 2004 , с. 330
- ^ Ридли 2004 , с. 332
Источники
[ редактировать ]- Андерссон М.Б. (1994). Сексуальный отбор . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА . ISBN 978-0-691-00057-2 .
- Futuyma D (2005). Эволюция (1 -е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-187-3 .
- Ридли М. (2004). Эволюция (3 -е изд.). Малден, Массачусетс: Blackwell Publishing. ISBN 978-1-4051-0345-9 .
Дальнейшее чтение
[ редактировать ]- Бондьюрианский Р (январь 2007 г.). «Эволюция зависимого от состояния полового диморфизма». Американский натуралист . 169 (1): 9–19. doi : 10.1086/510214 . PMID 17206580 . S2CID 17439073 .
- Figuerola J (1999). «Сравнительное исследование эволюции диморфизма обратного размера в моногамных куликах». Биологический журнал Линневого общества . 67 (1): 1–18. doi : 10.1111/j.1095-8312.1999.tb01926.x . HDL : 10261/44557 . S2CID 85330510 .
- Székely T, Lislevand T, Figuerola J, Fairbairn D, Blanckenhorn W (2007). Пол, размер и гендерные роли: эволюционные исследования диморфизма сексуального размера . С. 16–26.
Внешние ссылки
[ редактировать ]- Пол+диморфизм в Национальной библиотеке медицинской библиотеки Медицинской библиотеки Медицинской библиотеки (сетка)