Мутация может привести к множеству различных типов изменений в последовательностях. Мутации в генах не могут иметь никакого эффекта, изменять продукт гена или препятствовать правильному или полному функционированию гена. Мутации также могут возникать в негенных регионах . Исследование генетических вариаций между различными видами дрозофилы , проведенное в 2007 году , показало, что, если мутация изменяет белок, вырабатываемый геном, результат, вероятно, будет вредным: примерно 70% аминокислот полиморфизмов оказывают повреждающее воздействие, а остальные быть либо нейтральным, либо незначительно полезным. [8] Из-за разрушительного воздействия мутаций на гены у организмов есть такие механизмы, как восстановление ДНК , позволяющие предотвратить или исправить мутации, возвращая мутировавшую последовательность обратно в исходное состояние. [5]
Мутации могут включать дублирование больших участков ДНК, обычно посредством генетической рекомбинации . [9] Эти дупликации являются основным источником сырья для развития новых генов: каждые миллион лет в геномах животных дублируются десятки и сотни генов. [10] Большинство генов принадлежат к более крупным семействам генов общего происхождения, что можно обнаружить по гомологии их последовательностей . [11] Новые гены создаются несколькими методами, обычно путем дупликации и мутации предкового гена или путем рекомбинации частей разных генов для образования новых комбинаций с новыми функциями. [12] [13]
Здесь белковые домены действуют как модули, каждый из которых выполняет особую и независимую функцию, которые можно смешивать вместе для получения генов, кодирующих новые белки с новыми свойствами. [14] Например, человеческий глаз использует четыре гена для создания структур, воспринимающих свет: три для колбочек или цветового зрения и один для палочек или ночного зрения; все четыре произошли от одного предкового гена. [15] Еще одно преимущество дублирования гена (или даже всего генома) состоит в том, что это увеличивает инженерную избыточность ; это позволяет одному гену в паре приобрести новую функцию, в то время как другая копия выполняет исходную функцию. [16] [17] Другие типы мутаций иногда создают новые гены из ранее некодирующей ДНК . [18] [19]
Изменения числа хромосом могут включать в себя даже более крупные мутации, когда сегменты ДНК внутри хромосом разрываются, а затем перестраиваются. Например, у Homininae две хромосомы слились, образовав человеческую хромосому 2 ; это слияние не произошло в линии других обезьян , и они сохранили эти отдельные хромосомы. [20] В эволюции наиболее важная роль таких хромосомных перестроек может заключаться в ускорении дивергенции популяции на новые виды за счет снижения вероятности скрещивания популяций и тем самым сохранения генетических различий между этими популяциями. [21]
Последовательности ДНК, которые могут перемещаться по геному, такие как транспозоны , составляют основную часть генетического материала растений и животных и, возможно, сыграли важную роль в эволюции геномов. [22] более миллиона копий последовательности Alu присутствует Например, в геноме человека , и теперь эти последовательности задействованы для выполнения таких функций, как регулирование экспрессии генов . [23] Еще одним эффектом этих мобильных последовательностей ДНК является то, что когда они перемещаются внутри генома, они могут мутировать или удалять существующие гены и тем самым создавать генетическое разнообразие. [6]
Нелетальные мутации накапливаются в генофонде и увеличивают количество генетических вариаций. [24] Обилие некоторых генетических изменений в генофонде может быть уменьшено естественным отбором , в то время как другие «более благоприятные» мутации могут накапливаться и приводить к адаптивным изменениям.
Например, бабочка может произвести потомство с новыми мутациями. Большинство этих мутаций не будут иметь никакого эффекта; но можно изменить цвет одного из потомков бабочки, из-за чего хищникам будет труднее (или легче) его увидеть. Если это изменение цвета выгодно, шансы этой бабочки выжить и произвести собственное потомство немного выше, и со временем количество бабочек с этой мутацией может составить больший процент популяции. [ нужна ссылка ]
Нейтральные мутации определяются как мутации, эффекты которых не влияют на приспособленность человека. Со временем их частота может увеличиваться из-за генетического дрейфа . Считается, что подавляющее большинство мутаций не оказывают существенного влияния на приспособленность организма. [25] [26] Кроме того, механизмы репарации ДНК способны исправить большинство изменений до того, как они станут постоянными мутациями, и у многих организмов есть механизмы, такие как апоптотические пути , для уничтожения соматических клеток, мутировавших в противном случае навсегда . [27]
Полезные мутации могут улучшить репродуктивный успех. [28] [29]
Четыре класса мутаций: (1) спонтанные мутации (молекулярный распад), (2) мутации, вызванные склонным к ошибкам репликационным обходом естественных повреждений ДНК (также называемые склонным к ошибкам транслезным синтезом), (3) ошибки, возникающие во время репарации ДНК, и (4) индуцированные мутации, вызванные мутагены . Ученые также могут намеренно вводить мутантные последовательности посредством манипуляций с ДНК ради научных экспериментов. [ нужна ссылка ]
В одном исследовании 2017 года утверждалось, что 66% мутаций, вызывающих рак, являются случайными, 29% обусловлены окружающей средой (исследуемая популяция охватывала 69 стран) и 5% передаются по наследству. [30]
Люди в среднем передают своим детям 60 новых мутаций, но отцы передают больше мутаций в зависимости от возраста: каждый год у ребенка появляются две новые мутации. [31]
Спонтанные мутации происходят с ненулевой вероятностью даже в здоровой незагрязненной клетке. По оценкам, естественное окислительное повреждение ДНК происходит 10 000 раз на клетку в день у людей и 100 000 раз на клетку в день у крыс . [32] Спонтанные мутации характеризуются специфическими изменениями: [33]
Дезаминирование – гидролиз превращает нормальное основание в атипичное основание, содержащее кетогруппу вместо исходной аминогруппы . Примеры включают C → U и A → HX ( гипоксантин ), которые можно исправить с помощью механизмов репарации ДНК; и 5MeC ( 5-метилцитозин ) → T, который с меньшей вероятностью будет обнаружен как мутация, поскольку тимин является нормальным основанием ДНК.
Неправильное спаривание выскользнувшей цепи - денатурация новой цепи из матрицы во время репликации с последующей ренатурацией в другом месте («проскальзывание»). Это может привести к вставкам или удалениям.
Появляется все больше свидетельств того, что большинство спонтанно возникающих мутаций происходят из-за склонной к ошибкам репликации ( синтеза транслейкоза ) после повреждения ДНК в матричной цепи. У мышей большинство мутаций вызвано синтезом транслейкоза. [36] Аналогично, у дрожжей Kunz et al. [37] обнаружили, что более 60% спонтанных замен и делеций одной пары оснований были вызваны синтезом транслейкоза.
Хотя естественные двухцепочечные разрывы происходят в ДНК сравнительно редко, их восстановление часто приводит к мутациям. Ономологичное соединение концов (NHEJ) является основным путем восстановления двухцепочечных разрывов. NHEJ включает удаление нескольких нуклеотидов , чтобы обеспечить несколько неточное выравнивание двух концов для воссоединения, с последующим добавлением нуклеотидов для заполнения пробелов. Как следствие, NHEJ часто вносит мутации. [38]
Алкилирующие агенты (например, N -этил- N -нитрозомочевина (ENU). Эти агенты могут мутировать как реплицирующуюся, так и нереплицирующуюся ДНК. Напротив, базовый аналог может мутировать ДНК только тогда, когда аналог включен в репликацию ДНК. Каждый Эти классы химических мутагенов оказывают определенные эффекты, которые затем приводят к переходам , трансверсиям или делециям.
Азотистая кислота превращает аминогруппы A и C в диазогруппы , изменяя структуру их водородных связей, что приводит к неправильному спариванию оснований во время репликации.
Если раньше считалось, что мутации происходят случайно или вызываются мутагенами, то молекулярные механизмы мутаций были обнаружены у бактерий и на всем протяжении древа жизни. Как утверждает С. Розенберг: «Эти механизмы демонстрируют картину высокорегулируемого мутагенеза, который временно регулируется реакциями на стресс и активируется, когда клетки/организмы неадаптированы к окружающей среде – при стрессе – потенциально ускоряя адаптацию». [42] Поскольку они представляют собой самоиндуцируемые мутагенные механизмы, которые увеличивают скорость адаптации организмов, их иногда называют механизмами адаптивного мутагенеза и включают SOS-ответ у бактерий. [43] эктопическая внутрихромосомная рекомбинация [44] и другие хромосомные события, такие как дупликации. [42]
Пять типов хромосомных мутаций Типы мелкомасштабных мутаций
Последовательность гена можно изменить разными способами. [45] Генные мутации оказывают различное влияние на здоровье в зависимости от того, где они происходят и изменяют ли они функцию основных белков.Мутации в структуре генов можно разделить на несколько типов. [ нужна ссылка ]
К крупномасштабным мутациям хромосомной структуры относятся:
Амплификации (или дупликации генов ), повторение хромосомного сегмента или наличие дополнительного участка хромосомы, обломок хромосомы может прикрепиться к гомологичной или негомологичной хромосоме, так что некоторые гены присутствуют более чем в двух дозах, что приводит к к множественным копиям всех хромосомных областей, увеличивая дозировку генов, расположенных внутри них.
Полиплоидия , дупликация целых наборов хромосом, потенциально приводящая к отдельной размножающейся популяции и видообразованию .
Делеции крупных хромосомных областей, приводящие к потере генов внутри этих областей.
Мутации, эффект которых заключается в сопоставлении ранее отдельных фрагментов ДНК, потенциально объединяя отдельные гены с образованием функционально различных слитых генов (например, bcr-abl ).
Крупномасштабные изменения в структуре хромосом , называемые хромосомными перестройками , могут привести к снижению приспособленности, а также к видообразованию в изолированных инбредных популяциях. К ним относятся:
Интерстициальные делеции: внутрихромосомная делеция, которая удаляет сегмент ДНК из одной хромосомы, тем самым присоединяя ранее удаленные гены. Например, было обнаружено, что клетки, выделенные из астроцитомы человека , типа опухоли головного мозга, содержат последовательности удаления хромосомной делеции между геном Fused in Glioblastoma (FIG) и рецепторной тирозинкиназой (ROS), продуцирующими слитый белок (FIG- РОС). Аномальный слитый белок Fig-ROS обладает конститутивно активной киназной активностью, которая вызывает онкогенную трансформацию (преобразование нормальных клеток в раковые клетки).
Потеря гетерозиготности : потеря одной аллели в результате делеции или генетической рекомбинации в организме, который ранее имел две разные аллели.
Мелкомасштабные мутации затрагивают ген в одном или нескольких нуклеотидах. (Если затронут только один нуклеотид, их называют точечными мутациями .) К мелкомасштабным мутациям относятся:
Делеции удаляют один или несколько нуклеотидов из ДНК. Как и инсерции, эти мутации могут изменить рамку считывания гена. В общем, они необратимы: хотя теоретически одна и та же последовательность может быть восстановлена путем вставки, мобильные элементы, способные вернуть очень короткую делецию (скажем, 1–2 основания) в любом месте, либо вряд ли существуют, либо существуют. не существует вообще.
Заместительные мутации , часто вызванные химическими веществами или нарушением репликации ДНК, заменяют один нуклеотид на другой. [46] Эти изменения классифицируются как переходы или трансверсии. [47] Наиболее распространенным является переход, при котором пурин заменяется на пурин (A ↔ G) или пиримидин на пиримидин (C ↔ T). Переход может быть вызван азотистой кислотой, неправильным спариванием оснований или мутагенными аналогами оснований, такими как BrdU. Реже встречается трансверсия, при которой пурин заменяется на пиримидин или пиримидин на пурин (C/T ↔ A/G). Примером трансверсии является превращение аденина (А) в цитозин (С). Точечные мутации — это модификации отдельных пар оснований ДНК или других небольших пар оснований внутри гена. Точечная мутация может быть обращена вспять другой точечной мутацией, при которой нуклеотид возвращается в исходное состояние (истинная реверсия), или реверсией второго сайта (дополнительная мутация в другом месте, которая приводит к восстановлению функциональности гена). Как обсуждается ниже , точковые мутации, возникающие в кодирующей белок области гена, можно классифицировать как синонимичные или несинонимичные замены , последние из которых, в свою очередь, можно разделить на миссенс- или нонсенс-мутации. .
Влияние мутации на последовательность белка частично зависит от того, в каком месте генома она происходит, особенно в кодирующей или некодирующей области . Мутации в некодирующих регуляторных последовательностях гена, таких как промоторы, энхансеры и сайленсеры, могут изменять уровни экспрессии гена, но с меньшей вероятностью изменяют последовательность белка. Мутации внутри интронов и в регионах с неизвестной биологической функцией (например, псевдогены , ретротранспозоны ) обычно нейтральны и не влияют на фенотип, хотя мутации интронов могут изменить белковый продукт, если они влияют на сплайсинг мРНК.
Мутации, возникающие в кодирующих областях генома, с большей вероятностью изменяют белковый продукт, и их можно классифицировать по влиянию на аминокислотную последовательность:
Мутация сдвига рамки считывания вызвана вставкой или удалением ряда нуклеотидов, которые не делятся на три в последовательности ДНК. Из-за триплетного характера экспрессии генов кодонами вставка или удаление может нарушить рамку считывания или группировку кодонов, что приведет к совершенно отличной трансляции . от оригинала [49] Чем раньше в последовательности происходит делеция или вставка, тем более изменен вырабатываемый белок. (Например, код CCU GAC UAC CUA кодирует аминокислоты пролин, аспарагиновую кислоту, тирозин и лейцин. Если удалить U в CCU, результирующая последовательность будет CCG ACU ACC UAx, которая вместо этого будет кодировать пролин, треонин, треонин и часть другой аминокислоты или, возможно, стоп-кодон (где x обозначает следующий нуклеотид).) Напротив, любая вставка или делеция, которая делится без остатка на три, называется мутацией в рамке считывания .
Мутация точечной замены приводит к изменению одного нуклеотида и может быть как синонимической, так и несинонимичной.
Синонимичная замена заменяет кодон другим кодоном, кодирующим ту же аминокислоту, так что полученная аминокислотная последовательность не изменяется. Синонимические мутации возникают из-за вырожденной природы генетического кода . Если эта мутация не приводит к каким-либо фенотипическим эффектам, то ее называют молчащей , однако не все синонимичные замены являются молчащими. (Также могут быть молчащие мутации в нуклеотидах за пределами кодирующих областей, таких как интроны, поскольку точная последовательность нуклеотидов не так важна, как в кодирующих областях, но они не считаются синонимичными заменами.)
Несинонимичная замена заменяет кодон другим кодоном, кодирующим другую аминокислоту, в результате чего полученная аминокислотная последовательность модифицируется. Несинонимичные замены можно классифицировать как нонсенс- или миссенс-мутации:
Миссенс -мутация изменяет нуклеотид, вызывая замену другой аминокислоты. Это, в свою очередь, может сделать полученный белок нефункциональным. Такие мутации ответственны за такие заболевания, как буллезный эпидермолиз , серповидноклеточная анемия и SOD1 -опосредованный БАС . [50] С другой стороны, если в кодоне аминокислоты происходит миссенс-мутация, которая приводит к использованию другой, но химически сходной аминокислоты, то иногда в белке происходят незначительные изменения или вообще не происходит никаких изменений. Например, замена ААА на АГА будет кодировать аргинин , химически подобную молекуле предполагаемому лизину . В этом последнем случае мутация будет иметь незначительное влияние на фенотип или вообще не будет иметь его и, следовательно, будет нейтральной .
Нонсенс -мутация — это точечная мутация в последовательности ДНК, которая приводит к появлению преждевременного стоп-кодона или нонсенс-кодона в транскрибируемой мРНК и, возможно, к усеченному и часто нефункциональному белковому продукту. Этот вид мутации связан с различными заболеваниями, такими как врожденная гиперплазия надпочечников . (См. Стоп-кодон .)
Мутация становится следствием функциональной мутации, когда точность функций между мутировавшим белком и его прямым взаимодействием претерпевает изменения. Взаимодействующими веществами могут быть другие белки, молекулы, нуклеиновые кислоты и т. д. Существует множество мутаций, подпадающих под категорию побочных эффектов на функцию, но в зависимости от специфичности изменения могут возникать мутации, перечисленные ниже. [51]
Мутации с потерей функции, также называемые инактивирующими мутациями, приводят к тому, что генный продукт имеет меньшую функцию или вообще не выполняет ее (частично или полностью инактивируется). Когда аллель имеет полную потерю функции ( нулевой аллель ), ее часто называют аморфной или аморфной мутацией в морф Мюллера схеме . Фенотипы, связанные с такими мутациями, чаще всего являются рецессивными . Исключением являются случаи, когда организм гаплоидный или когда уменьшенной дозы нормального генного продукта недостаточно для нормального фенотипа (это называется гаплонедостаточностью ). Заболеванием, вызванным мутацией потери функции, является синдром Гительмана и муковисцидоз. [52]
Мутации с усилением функции, также называемые активирующими мутациями, изменяют продукт гена так, что его эффект становится сильнее (усиленная активация) или даже заменяется другой, аномальной функцией. Когда создается новая аллель, гетерозигота, содержащая вновь созданную аллель, а также исходную, будет экспрессировать новую аллель; генетически это определяет мутации как доминантные фенотипы. Некоторые из морфов Мюллера соответствуют усилению функции, включая гиперморф (повышенная экспрессия генов) и неоморф (новая функция).
Доминантно-негативные мутации (также называемые антиморфными мутациями) имеют измененный генный продукт, который действует антагонистически по отношению к аллелю дикого типа. Эти мутации обычно приводят к изменению молекулярной функции (часто неактивной) и характеризуются доминантным или полудоминантным фенотипом. У людей доминантно-негативные мутации связаны с раком (например, мутации в генах p53 , ATM , CEBPA и PPARgamma ). Синдром Марфана вызван мутациями гена FBN1 , расположенного на хромосоме 15 , который кодирует фибриллин-1, гликопротеиновый компонент внеклеточного матрикса . Синдром Марфана также является примером доминантно-негативной мутации и гаплонедостаточности.
Летальные мутации приводят к быстрой гибели организма, возникая во время развития, и вызывают значительное сокращение продолжительности жизни развитых организмов. Примером заболевания, вызванного доминантной летальной мутацией, является болезнь Хантингтона .
Нулевые мутации, также известные как аморфные мутации, представляют собой форму мутаций с потерей функции, которые полностью запрещают функцию гена. Мутация приводит к полной потере работы на фенотипическом уровне, а также к отсутствию образования генного продукта. Атопическая экзема и синдром дерматита — распространенные заболевания, вызванные нулевой мутацией гена, активирующего филагрин.
Супрессорные мутации — это тип мутации, который приводит к нормальному появлению двойной мутации. При супрессорных мутациях фенотипическая активность другой мутации полностью подавляется, поэтому двойная мутация выглядит нормальной. Существует два типа супрессорных мутаций: внутригенные и экстрагенные супрессорные мутации. Внутригенные мутации возникают в гене, в котором произошла первая мутация, тогда как экстрагенные мутации возникают в гене, который взаимодействует с продуктом первой мутации. Распространенным заболеванием, возникающим в результате этого типа мутации, является болезнь Альцгеймера . [53]
Неоморфные мутации являются частью мутаций приобретения функции и характеризуются контролем синтеза новых белковых продуктов. Вновь синтезированный ген обычно содержит новую экспрессию гена или молекулярную функцию. Результатом неоморфной мутации является то, что ген, в котором возникает мутация, полностью меняет функцию. [54]
Обратная мутация или реверсия — это точечная мутация, которая восстанавливает исходную последовательность и, следовательно, исходный фенотип. [55]
По влиянию на приспособленность (вредные, полезные, нейтральные мутации)
В генетике иногда полезно классифицировать мутации как вредный или полезный (или нейтральный ):
Вредное, или вредны , мутации снижают приспособленность организма. Многие, но не все, мутации незаменимых генов вредны (если мутация не меняет последовательность аминокислот в незаменимом белке, в большинстве случаев она безвредна).
Полезная или выгодная мутация повышает приспособленность организма. Примерами являются мутации, которые приводят к устойчивости бактерий к антибиотикам (которые полезны для бактерий, но обычно не для человека).
Нейтральная мутация не оказывает ни вредного, ни полезного воздействия на организм. Такие мутации происходят с постоянной скоростью, образуя основу молекулярных часов . В нейтральной теории молекулярной эволюции нейтральные мутации обеспечивают генетический дрейф как основу большинства вариаций на молекулярном уровне. У животных и растений большинство мутаций нейтральны, учитывая, что подавляющее большинство их геномов либо некодирующие, либо состоят из повторяющихся последовательностей, не несущих очевидной функции (« мусорная ДНК »). [56]
Крупномасштабные скрининги количественного мутагенеза , в ходе которых проверяются тысячи миллионов мутаций, неизменно обнаруживают, что большая часть мутаций имеет вредные последствия, но всегда также возвращает ряд полезных мутаций. Например, при проверке всех делеций генов в E. coli 80% мутаций были отрицательными, но 20% были положительными, хотя многие из них оказывали очень незначительное влияние на рост (в зависимости от состояния). [57] генов Удаление предполагает удаление целых генов, поэтому точечные мутации почти всегда имеют гораздо меньший эффект. При аналогичном скрининге Streptococcus pneumoniae , но на этот раз со вставками транспозонов , 76% инсерционных мутантов были классифицированы как нейтральные, 16% имели значительно сниженную приспособленность, но 6% имели преимущество. [58]
Эта классификация явно относительна и несколько искусственна: вредная мутация может быстро превратиться в полезную при изменении условий. Кроме того, существует градиент от вредного/полезного к нейтральному, поскольку многие мутации могут иметь небольшие и в основном незначительные последствия, но при определенных условиях станут актуальными. Кроме того, многие признаки определяются сотнями генов (или локусов), так что каждый локус оказывает лишь незначительное влияние. Например, рост человека определяется сотнями генетических вариантов («мутаций»), но каждый из них оказывает очень незначительное влияние на рост. [59] кроме влияния питания . Высота (или размер) сама по себе может быть более или менее полезной, как показывает огромный диапазон размеров в группах животных и растений.
Были предприняты попытки сделать вывод о распределении эффектов приспособленности (DFE) с использованием экспериментов по мутагенезу и теоретических моделей, примененных к данным молекулярных последовательностей. DFE, используемый для определения относительной распространенности различных типов мутаций (т. е. сильно вредных, почти нейтральных или полезных), имеет отношение ко многим эволюционным вопросам, таким как поддержание генетической изменчивости . [60] скорость распада генома , [61] поддержание ауткроссингового полового размножения в отличие от инбридинга [62] и эволюция пола и генетическая рекомбинация . [63] DFE также можно отслеживать, отслеживая неравномерность распределения мутаций с предположительно тяжелыми эффектами по сравнению с распределением мутаций с предположительно легкими или отсутствующими эффектами. [64] Таким образом, DFE играет важную роль в предсказании эволюционной динамики . [65] [66] Для изучения ДФЭ использовались различные подходы, включая теоретические, экспериментальные и аналитические методы.
Эксперимент по мутагенезу. Прямой метод исследования DFE состоит в том, чтобы вызвать мутации, а затем измерить эффекты мутационной приспособленности, что уже было сделано на вирусах, бактериях , дрожжах и дрозофиле . Например, в большинстве исследований DFE вирусов использовался сайт-направленный мутагенез для создания точечных мутаций и измерения относительной приспособленности каждого мутанта. [67] [68] [69] [70] В Escherichia coli одном исследовании использовался мутагенез транспозонов для прямого измерения пригодности случайной вставки производного Tn10 . [71] В дрожжах был разработан комбинированный подход к мутагенезу и глубокому секвенированию для создания высококачественных систематических мутантных библиотек и измерения приспособленности с высокой производительностью. [72] Однако, учитывая, что многие мутации имеют слишком малый эффект, чтобы его можно было обнаружить. [73] и что эксперименты по мутагенезу могут обнаружить только мутации умеренно большого эффекта; ДНК Анализ последовательности может предоставить ценную информацию об этих мутациях.
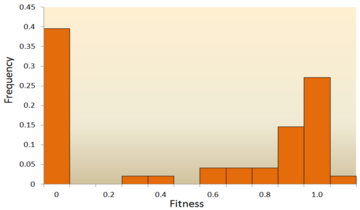
Распределение фитнес-эффектов (DFE) мутаций вируса везикулярного стоматита . В этом эксперименте случайные мутации были введены в вирус путем сайт-направленного мутагенеза, и приспособленность каждого мутанта сравнивалась с предковым типом. Пригодность ноль, меньше единицы, единица и больше единицы соответственно указывает на то, что мутации летальны, вредны, нейтральны и полезны. [67]
На этом рисунке показана упрощенная версия мутаций потери функции, переключения функции, приобретения функции и сохранения функции. Анализ молекулярных последовательностей. Благодаря быстрому развитию технологии секвенирования ДНК доступен огромный объем данных о последовательностях ДНК, и в будущем их будет еще больше. Были разработаны различные методы для вывода DFE на основе данных последовательности ДНК. [74] [75] [76] [77] Изучая различия в последовательностях ДНК внутри и между видами, мы можем сделать вывод о различных характеристиках DFE для нейтральных, вредных и полезных мутаций. [24] Говоря конкретнее, подход анализа последовательности ДНК позволяет нам оценить эффекты мутаций с очень небольшими эффектами, которые трудно обнаружить в экспериментах по мутагенезу.
Одно из первых теоретических исследований распределения эффектов приспособленности было проведено Мотоо Кимурой , влиятельным теоретиком популяционной генетики . Его нейтральная теория молекулярной эволюции предполагает, что большинство новых мутаций будут очень вредными, а небольшая часть будет нейтральной. [25] [78] Более позднее предложение Хироши Акаши предложило бимодальную модель DFE, режимы которой сосредоточены на крайне вредных и нейтральных мутациях. [79] Обе теории сходятся во мнении, что подавляющее большинство новых мутаций нейтральны или вредны, а полезные мутации редки, что подтверждается экспериментальными результатами. Одним из примеров является исследование DFE случайных мутаций вируса везикулярного стоматита . [67] Из всех мутаций 39,6% были летальными, 31,2% — нелетально вредными и 27,1% — нейтральными. Другой пример — эксперимент по высокопроизводительному мутагенезу с дрожжами. [72] В этом эксперименте было показано, что общий DFE является бимодальным, с кластером нейтральных мутаций и широким распространением вредных мутаций.
Хотя относительно небольшое количество мутаций являются полезными, те, которые есть, играют важную роль в эволюционных изменениях. [80] Как и нейтральные мутации, слабо выбранные выгодные мутации могут быть потеряны из-за случайного генетического дрейфа, но сильно выбранные выгодные мутации с большей вероятностью будут исправлены. Знание DFE выгодных мутаций может привести к увеличению способности предсказывать эволюционную динамику. Теоретическая работа по DFE для полезных мутаций была проведена Джоном Х. Гиллеспи. [81] и Х. Аллен Орр . [82] Они предположили, что распределение полезных мутаций должно быть экспоненциальным в широком диапазоне условий, что, в целом, было подтверждено экспериментальными исследованиями, по крайней мере, для тщательно отобранных полезных мутаций. [83] [84] [85]
В целом считается, что большинство мутаций нейтральны или вредны, а полезные мутации встречаются редко; однако доля типов мутаций варьируется между видами. Это указывает на два важных момента: во-первых, доля эффективно нейтральных мутаций, вероятно, будет варьироваться между видами в зависимости от эффективного размера популяции ; во-вторых, средний эффект вредных мутаций резко различается между видами. [24] Кроме того, DFE также различается между кодирующими и некодирующими областями , при этом DFE некодирующей ДНК содержит более слабо выбранные мутации. [24]
Диплоидные организмы (например, люди) содержат две копии каждого гена — отцовскую и материнскую аллели. В зависимости от возникновения мутаций в каждой хромосоме мы можем разделить мутации на три типа. Организм дикого типа или гомозиготный немутантный организм — это организм, в котором ни одна аллель не мутирована.
Гетерозиготная мутация – это мутация только одного аллеля.
Гомозиготная мутация — это идентичная мутация как отцовского, так и материнского аллеля.
Сложные гетерозиготные мутации или генетическое соединение состоит из двух разных мутаций в отцовских и материнских аллелях. [87]
Зародышевая мутация в репродуктивных клетках человека приводит к конституциональной мутации у потомства, то есть мутации, которая присутствует в каждой клетке. Конституционная мутация также может произойти очень скоро после оплодотворения или продолжиться после предыдущей конституциональной мутации у родителя. [88] Мутация зародышевой линии может передаваться через последующие поколения организмов.
Различие между зародышевой линией и соматическими мутациями важно для животных, у которых есть специальная зародышевая линия для производства репродуктивных клеток. Однако это не имеет большого значения для понимания последствий мутаций у растений, у которых отсутствует выделенная зародышевая линия. Различие также размыто у тех животных, которые размножаются бесполым путем с помощью таких механизмов, как почкование , поскольку клетки, дающие начало дочерним организмам, также дают начало зародышевой линии этого организма.
Новая зародышевая мутация, не унаследованная ни от одного из родителей, называется de novo мутацией .
Изменение генетической структуры, не наследуемое от родителя, а также не передающееся потомству, называется соматической мутацией . [86] Соматические мутации не наследуются потомством организма, поскольку не затрагивают зародышевую линию . Однако они передаются всему потомству мутировавшей клетки в одном и том же организме во время митоза. Таким образом, основная часть организма может нести одну и ту же мутацию. Эти типы мутаций обычно вызваны причинами окружающей среды, такими как ультрафиолетовое излучение или любое воздействие определенных вредных химических веществ, и могут вызывать заболевания, включая рак. [89]
На растениях некоторые соматические мутации можно размножать без необходимости получения семян, например, прививкой и стеблевыми черенками. Этот тип мутации привел к появлению новых типов фруктов, таких как «Вкусное» яблоко и «Вашингтонский» пупочный апельсин . [90]
человека и мыши Соматические клетки имеют частоту мутаций более чем в десять раз выше, чем частота мутаций зародышевой линии для обоих видов; мыши имеют более высокий уровень как соматических, так и зародышевых мутаций на деление клеток, чем люди. Неравенство в частоте мутаций между зародышевой линией и соматическими тканями, вероятно, отражает большую важность поддержания генома в зародышевой линии, чем в соме. [91]
Условная мутация — это мутация, которая имеет фенотип дикого типа (или менее тяжелый) при определенных «разрешительных» условиях окружающей среды и мутантный фенотип при определенных «рестриктивных» условиях. Например, чувствительная к температуре мутация может вызвать гибель клеток при высокой температуре (ограничительные условия), но может не иметь вредных последствий при более низкой температуре (разрешительные условия). [92] Эти мутации неавтономны, так как их проявление зависит от наличия определенных условий, в отличие от других мутаций, которые возникают автономно. [93] Разрешающими условиями могут быть температура , [94] определенные химические вещества, [95] свет [95] или мутации в других частях генома . [93] Механизмы in vivo , такие как переключатели транскрипции, могут создавать условные мутации. Например, ассоциация стероидсвязывающего домена может создать переключатель транскрипции, который может изменить экспрессию гена в зависимости от присутствия стероидного лиганда. [96] Условные мутации находят применение в исследованиях, поскольку позволяют контролировать экспрессию генов. Это особенно полезно при изучении заболеваний у взрослых, поскольку позволяет осуществлять экспрессию после определенного периода роста, устраняя тем самым вредный эффект экспрессии генов, наблюдаемый на стадиях развития модельных организмов. [95] Системы ДНК-рекомбиназы, такие как рекомбинация Cre-Lox, используемые в сочетании с промоторами , которые активируются при определенных условиях, могут генерировать условные мутации. Технология двойной рекомбиназы может использоваться для индукции множественных условных мутаций для изучения заболеваний, которые проявляются в результате одновременных мутаций в нескольких генах. [95] Были идентифицированы определенные интеины , которые сращиваются только при определенных допустимых температурах, что приводит к неправильному синтезу белка и, следовательно, к мутациям с потерей функции при других температурах. [97] Условные мутации также могут использоваться в генетических исследованиях, связанных со старением, поскольку экспрессия может меняться через определенный период времени жизни организма. [94]
Чтобы отнести мутацию к категории таковой, «нормальную» последовательность необходимо получить из ДНК «нормального» или «здорового» организма (в отличие от «мутантного» или «больного»), ее следует идентифицировать и сообщил; в идеале он должен быть доступен общественности для прямого сравнения нуклеотидов и согласован научным сообществом или группой экспертов-генетиков и биологов , которые несут ответственность за установление стандарта или так называемого «консенсуса». последовательность. Этот шаг требует огромных научных усилий. Как только консенсусная последовательность станет известна, мутации в геноме можно будет точно определить, описать и классифицировать. Комитет Общества вариаций генома человека (HGVS) разработал стандартную номенклатуру вариантов последовательностей человека. [98] который должен использоваться исследователями и центрами ДНК-диагностики для создания однозначных описаний мутаций. В принципе, эту номенклатуру можно использовать и для описания мутаций в других организмах. Номенклатура указывает тип мутации и изменения оснований или аминокислот.
Нуклеотидная замена (например, 76A>T) – число обозначает положение нуклеотида с 5'-конца; первая буква представляет собой нуклеотид дикого типа, а вторая буква представляет собой нуклеотид, заменивший дикий тип. В данном примере аденин в 76-й позиции заменен тимином.
Если возникает необходимость различать мутации в геномной ДНК , митохондриальной ДНК и РНК , используется простое соглашение. Например, если 100-е основание нуклеотидной последовательности мутировало с G на C, то это будет записано как g.100G>C, если мутация произошла в геномной ДНК, m.100G>C, если мутация произошла в митохондриальной ДНК, или r.100g>c, если мутация произошла в РНК. Обратите внимание, что для мутаций в РНК нуклеотидный код пишется строчными буквами.
Аминокислотная замена (например, D111E). Первая буква — это однобуквенный код аминокислоты дикого типа, число — это положение аминокислоты от N-конца , а вторая буква — это однобуквенный код аминокислоты. аминокислота, присутствующая в мутации. Нонсенс-мутации обозначаются буквой X для второй аминокислоты (например, D111X).
Делеция аминокислоты (например, ΔF508). Греческая буква Δ ( дельта ) обозначает делецию. Буква относится к аминокислоте, присутствующей в диком типе, а число представляет собой положение от N-конца аминокислоты, если бы она присутствовала, как и в диком типе.
Скорость мутаций существенно различается у разных видов, а эволюционные силы, которые обычно определяют мутации, являются предметом продолжающихся исследований.
У людей частота мутаций составляет около 50–90 мутаций de novo на геном на поколение, то есть у каждого человека накапливается около 50–90 новых мутаций, которых не было у его или ее родителей. Это число было установлено путем секвенирования тысяч человеческих троек, то есть двух родителей и как минимум одного ребенка. [99]
Геномы РНК-вирусов основаны на РНК, а не на ДНК. РНК-вирусный геном может быть двухцепочечным (как в ДНК) или одноцепочечным. У некоторых из этих вирусов (например, у одноцепочечного вируса иммунодефицита человека ) репликация происходит быстро, и отсутствуют механизмы проверки генома на точность. Этот подверженный ошибкам процесс часто приводит к мутациям.
Скорость мутаций de novo, будь то зародышевых или соматических, варьируется у разных организмов. [100] Особи одного и того же вида могут даже проявлять различную скорость мутаций. [101] В целом, уровень мутаций de novo низок по сравнению с уровнем наследственных мутаций, что классифицирует их как редкие формы генетических вариаций . [102] Многие наблюдения за частотой мутаций de novo связаны с более высокой частотой мутаций, коррелирующей с возрастом отца. В организмах, размножающихся половым путем, сравнительно более высокая частота делений клеток в зародышевой линии родительской спермы-донора позволяет сделать вывод о том, что скорость мутаций de novo можно отслеживать на общей основе. Частота ошибок в процессе репликации ДНК в ходе гаметогенеза , особенно усиливающаяся при быстром производстве сперматозоидов, может способствовать увеличению возможностей мутаций de novo для репликации, нерегулируемой механизмом репарации ДНК. [103] Это утверждение сочетает в себе наблюдаемые эффекты повышенной вероятности мутаций при быстром сперматогенезе с короткими периодами времени между клеточными делениями, которые ограничивают эффективность механизмов восстановления. [104] Частота мутаций de novo, влияющих на организм в процессе его развития, также может увеличиваться при наличии определенных факторов окружающей среды. Например, определенная интенсивность воздействия радиоактивных элементов может нанести ущерб геному организма, увеличивая скорость мутаций. У людей появление рака кожи в течение жизни вызвано чрезмерным воздействием УФ-излучения , которое вызывает мутации в клеточном и кожном геноме. [105]
Существует широко распространенное предположение, что мутации (полностью) «случайны» в отношении их последствий (с точки зрения вероятности). Было показано, что это неверно, поскольку частота мутаций может варьироваться в зависимости от региона генома, причем такое восстановление ДНК и предвзятость мутаций связаны с различными факторами. Например, Монро и его коллеги продемонстрировали, что в изучаемом растении ( Arabidopsis thaliana ) более важные гены мутируют реже, чем менее важные. Они продемонстрировали, что мутация «неслучайна и приносит пользу растению». [106] [107] Кроме того, было показано, что предыдущие эксперименты, которые обычно использовались для демонстрации случайности мутаций в отношении приспособленности (такие как тест на флуктуации и репликация ), подтверждают только более слабое утверждение о том, что эти мутации являются случайными по отношению к внешним селективным ограничениям, а не приспособленности как фактору приспособленности. весь. [108]
Изменения в ДНК, вызванные мутацией в кодирующей области ДНК, могут вызвать ошибки в последовательности белков, что может привести к частично или полностью нефункциональным белкам. Правильное функционирование каждой клетки зависит от тысяч белков, которые функционируют в нужных местах в нужное время. Когда мутация изменяет белок, который играет решающую роль в организме, это может привести к заболеванию. Одно исследование по сравнению генов разных видов дрозофилы предполагает, что, если мутация действительно изменит белок, эта мутация, скорее всего, будет вредной: примерно 70 процентов полиморфизмов аминокислот оказывают повреждающее воздействие, а остальные являются либо нейтральными, либо нейтральными. слабо выгодно. [8] Некоторые мутации изменяют базовую последовательность ДНК гена, но не меняют белок, вырабатываемый этим геном. Исследования показали, что только 7% точковых мутаций в некодирующей ДНК дрожжей являются вредными, а 12% - в кодирующей ДНК. Остальные мутации либо нейтральны, либо слегка полезны. [109]
Если мутация присутствует в зародышевой клетке , она может дать начало потомству, несущему эту мутацию во всех своих клетках. Это касается наследственных заболеваний. В частности, если в зародышевой клетке есть мутация в гене репарации ДНК, люди, несущие такие мутации зародышевой линии, могут иметь повышенный риск развития рака. Список из 34 таких мутаций зародышевой линии приведен в статье « Нарушение репарации ДНК» . Примером одного из них является альбинизм , мутация, возникающая в гене OCA1 или OCA2 . Люди с этим расстройством более склонны ко многим видам рака и другим расстройствам, а также имеют нарушения зрения.
Повреждение ДНК может вызвать ошибку при репликации ДНК, и эта ошибка репликации может вызвать мутацию гена, которая, в свою очередь, может вызвать генетическое заболевание. Повреждения ДНК восстанавливаются системой репарации ДНК клетки. Каждая клетка имеет ряд путей, с помощью которых ферменты распознают и восстанавливают повреждения ДНК. Поскольку ДНК может быть повреждена разными способами, процесс восстановления ДНК является важным способом защиты организма от болезней. Если повреждение ДНК привело к мутации, мутацию невозможно исправить.
С другой стороны, мутация может возникнуть и в соматической клетке организма. Такие мутации будут присутствовать у всех потомков этой клетки в пределах одного организма. Накопление определенных мутаций на протяжении поколений соматических клеток является одной из причин злокачественной трансформации из нормальной клетки в раковую. [110]
Клетки с гетерозиготными мутациями потери функции (одна хорошая копия гена и одна мутированная копия) могут нормально функционировать с немутированной копией до тех пор, пока хорошая копия не подвергнется спонтанной соматической мутации. Такого рода мутации часто случаются в живых организмах, но их скорость трудно измерить. Измерение этой скорости важно для прогнозирования скорости развития рака у людей. [111]
Точечные мутации могут возникать в результате спонтанных мутаций, возникающих во время репликации ДНК. Скорость мутации может быть увеличена мутагенами. Мутагены могут быть физическими, например, излучением УФ-лучей , рентгеновских лучей или сильной жары, или химическими (молекулы, которые перемещают пары оснований неправильно или нарушают спиральную форму ДНК). Мутагены, связанные с раком, часто изучаются, чтобы узнать о раке и его профилактике.
Хотя мутации, вызывающие изменения в последовательностях белков, могут быть вредными для организма, в некоторых случаях эффект может быть положительным в данной среде. В этом случае мутация может позволить мутантному организму лучше противостоять определенным стрессам окружающей среды, чем организмам дикого типа, или быстрее размножаться. В этих случаях мутация будет иметь тенденцию становиться более распространенной в популяции в результате естественного отбора. Тем не менее, одна и та же мутация может быть полезной в одном состоянии и невыгодной в другом. Примеры включают следующее:
Устойчивость к ВИЧ : специфическая делеция 32 пар оснований в человеческом CCR5 ( CCR5-Δ32 ) придает ВИЧ устойчивость к гомозиготам и задерживает начало СПИДа у гетерозигот. [112] Одним из возможных объяснений этиологии относительно высокой частоты CCR5-Δ32 среди европейского населения является то, что он обеспечивал устойчивость к бубонной чуме середины 14 века в Европе . Люди с этой мутацией имели больше шансов пережить инфекцию; таким образом, его частота среди населения увеличилась. [113] Эта теория могла бы объяснить, почему эта мутация не обнаружена в Южной Африке , которая осталась нетронутой бубонной чумой. Более новая теория предполагает, что избирательное давление на мутацию CCR5 Delta 32 было вызвано оспой, а не бубонной чумой. [114]
Устойчивость к антибиотикам : практически все бактерии развивают устойчивость к антибиотикам при воздействии антибиотиков. Фактически, в бактериальных популяциях уже есть такие мутации, которые отбираются при селекции антибиотиков. [116] Очевидно, что такие мутации полезны только для бактерий, но не для инфицированных.
Стойкость лактазы . Мутация позволила людям экспрессировать фермент лактазу после того, как они естественным образом отлучились от грудного молока, что позволило взрослым переваривать лактозу , что, вероятно, является одной из самых полезных мутаций в недавней эволюции человека . [117]
Привнося новые генетические качества в популяцию организмов, мутации de novo играют решающую роль в объединенных силах эволюционных изменений. Однако вес генетического разнообразия, порожденный мутационными изменениями, часто считается в целом «слабой» эволюционной силой. [101] Хотя случайное возникновение мутаций само по себе обеспечивает основу для генетических вариаций во всей органической жизни, эту силу необходимо принимать во внимание наряду со всеми действующими эволюционными силами. Спонтанные мутации de novo как катастрофические события видообразования зависят от факторов, вызванных естественным отбором , генетическим потоком и генетическим дрейфом . Например, меньшие популяции с тяжелым мутационным воздействием (высокая частота мутаций) склонны к увеличению генетической изменчивости, что приводит к видообразованию в будущих поколениях. Напротив, более крупные популяции, как правило, видят меньшие эффекты от вновь появившихся мутировавших признаков. В этих условиях силы отбора со временем уменьшают частоту мутированных аллелей, которые чаще всего оказываются вредными. [118]
Компенсированные патогенные отклонения относятся к аминокислотным остаткам в последовательности белка, которые являются патогенными у одного вида, но являются остатками дикого типа в функционально эквивалентном белке у другого вида. Хотя аминокислотный остаток является патогенным у первого вида, он не является таковым у второго вида, поскольку его патогенность компенсируется одной или несколькими аминокислотными заменами у второго вида. Компенсаторная мутация может возникнуть в том же белке или в другом белке, с которым он взаимодействует. [119]
Крайне важно понять эффекты компенсаторных мутаций в контексте фиксированных вредных мутаций из-за снижения приспособленности популяции из-за фиксации. [120] Эффективная численность популяции относится к популяции, которая воспроизводится. [121] Увеличение размера этой популяции коррелирует со снижением уровня генетического разнообразия. [121] Положение популяции относительно критического размера популяции важно для определения влияния вредных аллелей на приспособленность. [120] Если популяция меньше критического эффективного размера, приспособленность резко уменьшится, однако, если популяция превышает размер критического эффекта, приспособленность может увеличиться независимо от вредных мутаций, вызванных компенсаторными аллелями. [120]
Поскольку функция молекулы РНК зависит от ее структуры, [122] структура молекул РНК эволюционно консервативна. Следовательно, любая мутация, изменяющая стабильную структуру молекул РНК, должна быть компенсирована другими компенсаторными мутациями. В контексте РНК последовательность РНК можно рассматривать как «генотип», а структуру РНК можно рассматривать как ее «фенотип». Поскольку РНК имеют относительно более простой состав, чем белки, структуру молекул РНК можно предсказать с помощью вычислений с высокой степенью точности. Из-за этого удобства компенсаторные мутации изучались с помощью компьютерного моделирования с использованием алгоритмов сворачивания РНК. [123] [124]
Компенсаторные мутации можно объяснить генетическим феноменом эпистаза, при котором фенотипический эффект одной мутации зависит от мутации (мутаций) в других локусах. Хотя эпистаз изначально был задуман в контексте взаимодействия между различными генами, в последнее время также изучается внутригенный эпистаз. [125] Существование компенсированных патогенных отклонений можно объяснить «признаковым эпистазом», при котором эффекты вредной мутации могут быть компенсированы наличием эпистатической мутации в других локусах. Для данного белка можно рассматривать вредную мутацию (D) и компенсаторную мутацию (C), где C может находиться в том же белке, что и D, или в другом взаимодействующем белке в зависимости от контекста. Эффект приспособленности самого C может быть нейтральным или несколько вредным, так что он все еще может существовать в популяции, а эффект D вреден в той степени, в которой он не может существовать в популяции. Однако когда C и D происходят одновременно, общий эффект приспособленности становится нейтральным или положительным. [119] Таким образом, компенсаторные мутации могут привнести новизну в белки, создавая новые пути эволюции белков: они позволяют людям путешествовать от одного пика приспособленности к другому через долины более низкой приспособленности. [125]
ДеПристо и др. В 2005 г. были выделены две модели, объясняющие динамику компенсаторно-патогенных отклонений (КПД). [126] В первой гипотезе P — патогенная аминокислотная мутация, а C — нейтральная компенсаторная мутация. [126] В этих условиях, если патогенная мутация возникает после компенсаторной мутации, то Р может закрепиться в популяции. [126] Вторая модель CPD утверждает, что P и C являются вредными мутациями, приводящими к долинам приспособленности, когда мутации происходят одновременно. [126] Используя общедоступные данные Ferrer-Costa et al. В 2007 году были получены наборы данных о компенсаторных мутациях и патогенных мутациях человека, которые были охарактеризованы для определения причин CPD. [127] Результаты показывают, что структурные ограничения и расположение в структуре белка определяют, возникнут ли компенсированные мутации. [127]
Лунцер и др. [128] проверили результат замены расходящихся аминокислот между двумя ортологичными белками изопропималатдегидрогеназы (IMDH). Они заменили 168 аминокислот в IMDH Escherichia coli , которые являются остатками дикого типа в IMDH Pseudomonas aeruginosa . Они обнаружили, что более трети этих замен нарушают ферментативную активность IMDH в генетическом фоне Escherichia coli . Это продемонстрировало, что идентичные состояния аминокислот могут приводить к различным фенотипическим состояниям в зависимости от генетического фона. Корриган и др. 2011 год продемонстрировал, как золотистый стафилококк способен нормально расти без присутствия липотейхоевой кислоты из-за компенсаторных мутаций. [129] Результаты полногеномного секвенирования показали, что разрушение циклической ди-АМФ-фосфодиэстеразы (GdpP) в этой бактерии компенсирует исчезновение полимера клеточной стенки, что приводит к нормальному росту клеток. [129]
Исследования показали, что бактерии могут приобретать устойчивость к лекарствам за счет компенсаторных мутаций, которые не препятствуют приспособленности или оказывают незначительное влияние на нее. [130] Предыдущие исследования Gagneux et al. В 2006 году было обнаружено, что выращенные в лаборатории штаммы микобактерий туберкулеза с устойчивостью к рифампицину имеют пониженную пригодность, однако клинические штаммы этих патогенных бактерий, устойчивые к лекарствам, не имеют пониженной приспособленности. [131] Комас и др. В 2012 году было проведено сравнение всего генома клинических штаммов и мутантов, полученных в лаборатории, чтобы определить роль и вклад компенсаторных мутаций в лекарственную устойчивость к рифампицину. [130] Анализ генома показал, что штаммы, устойчивые к рифампицину, имеют мутации в rpoA и rpoC. [130] В аналогичном исследовании изучали бактериальную приспособленность, связанную с компенсаторными мутациями у устойчивой к рифампицину Escherichia coli . [132] Результаты, полученные в этом исследовании, показывают, что устойчивость к лекарствам связана с приспособленностью бактерий, поскольку более высокие затраты на приспособленность связаны с большим количеством ошибок транскрипции. [132]
Гонг и др. [133] собрал полученные данные о генотипе нуклеопротеина гриппа из разных временных шкал и упорядочил их по времени в соответствии со временем происхождения. Затем они выделили 39 аминокислотных замен, произошедших в разные сроки, и заменили их в генетическом фоне, который приблизительно соответствовал генотипу предков. Они обнаружили, что 3 из 39 замен значительно снизили приспособленность предков. Компенсаторные мутации — это новые мутации, которые возникают и оказывают положительное или нейтральное влияние на приспособленность популяции. [134] Предыдущие исследования показали, что популяции могут компенсировать вредные мутации. [119] [134] [135] Берч и Чао проверили геометрическую модель адаптивной эволюции Фишера , проверив, развивается ли бактериофаг φ6 небольшими шагами. [136] Их результаты показали, что приспособленность бактериофага φ6 быстро снижалась и восстанавливалась небольшими шагами. [136] Было показано, что вирусные нуклеопротеины избегают цитотоксических Т-лимфоцитов (CTL) за счет замены аргинина на глицин. [137] Эти замещающие мутации влияют на приспособленность вирусных нуклеопротеинов, однако компенсаторные комутации препятствуют снижению приспособленности и помогают вирусу избежать распознавания со стороны CTL. [137] Мутации могут иметь три разных эффекта; мутации могут иметь пагубные последствия, некоторые из них повышают приспособленность за счет компенсаторных мутаций, и, наконец, мутации могут уравновешивать, приводя к компенсаторным нейтральным мутациям. [138] [132] [131]
В геноме человека частота и характеристики мутаций de novo изучались как важные контекстуальные факторы нашей эволюции. По сравнению с эталонным геномом человека, типичный человеческий геном варьируется примерно от 4,1 до 5,0 миллионов локусов, и большая часть этого генетического разнообразия принадлежит почти 0,5% населения. [139] Типичный геном человека также содержит от 40 000 до 200 000 редких вариантов, наблюдаемых менее чем у 0,5% населения, которые могли возникнуть только в результате хотя бы одной мутации зародышевой линии de novo в истории эволюции человека. [140] Также было исследовано, что мутации de novo играют решающую роль в сохранении генетических заболеваний у людей. Благодаря недавним достижениям в области секвенирования нового поколения (NGS), все типы мутаций de novo в геноме могут быть непосредственно изучены, обнаружение которых обеспечивает глубокое понимание причин как редких, так и распространенных генетических нарушений. В настоящее время наилучшая оценка средней частоты мутаций SNV зародышевой линии человека составляет 1,18 x 10^-8, при этом примерно ~78 новых мутаций на поколение. Возможность проводить полногеномное секвенирование родителей и потомков позволяет сравнивать частоту мутаций между поколениями, сужая возможности происхождения определенных генетических нарушений. [141]
^ Пфоль-Лешкович А., Мандервиль Р.А. (январь 2007 г.). «Охратоксин А: Обзор токсичности и канцерогенности для животных и человека». Молекулярное питание и пищевые исследования . 51 (1): 61–99. дои : 10.1002/mnfr.200600137 . ПМИД 17195275 .
^ Фриз Э. (июнь 1959 г.). «Специфическое мутагенное действие базовых аналогов на фаг Т4». Журнал молекулярной биологии . 1 (2): 87–105. дои : 10.1016/S0022-2836(59)80038-3 .
^ « RB1 Генетика » . Фонд борьбы с раком глаз Дейзи . Оксфорд, Великобритания. Архивировано из оригинала 26 ноября 2011 года . Проверено 9 октября 2015 г.
Arc.Ask3.Ru Номер скриншота №: e7ddf713a89043a549e6316440ab1e32__1720691580 URL1:https://arc.ask3.ru/arc/aa/e7/32/e7ddf713a89043a549e6316440ab1e32.html Заголовок, (Title) документа по адресу, URL1: Mutation - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)