G-белок-связанный рецептор
| GPCR | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | 7tm_1 | ||
| Pfam | PF00001 | ||
| PFAM клан | CL0192 | ||
| Эко | 5001.1.1 | ||
| InterPro | IPR000276 | ||
| PROSITE | PDOC00210 | ||
| TCDB | 9.A.14 | ||
| OPM Суперсемейство | 6 | ||
| OPM белок | 1GZM | ||
| CDD | CD14964 | ||
| |||
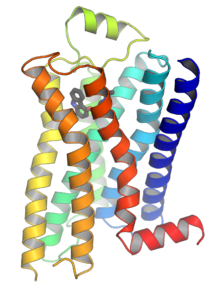
G-белок-связанные рецепторы ( GPCR известные как семи- (проходные) рецепторы доменных доменов , рецепторы 7TM , гептоалелогические рецепторы , серпентинские рецепторы и G - ), также , связанные белок являются рецепторами клеточной поверхности , которые обнаруживают молекулы вне клетки и активируют клеточные ответы. Они в сочетании с G -белками . Они проходят через клеточную мембрану семь раз в форме шести петель [ 2 ] (Три внеклеточных петли, взаимодействующих с молекулами лиганда , три внутриклеточных петли, взаимодействующих с G-белками, N-концевой внеклеточной области и C-концевой внутриклеточной области [ 2 ] ) аминокислотных остатков , поэтому их иногда называют рецепторами семи трансмембран. [ 3 ] Лиганды могут связываться либо с внеклеточным N -конце, так и с петлями (например, глутаматные рецепторы) или с сайтом связывания в трансмембранных спиралях ( Rhodopsin семейство -подобного). Все они активируются агонистами , хотя также наблюдалась спонтанная аутоцивация пустого рецептора. [ 3 ]
G-белковые рецепторы обнаруживаются только у эукариот , включая дрожжи и хонофлагелляты . [ 4 ] Лиганды , которые связывают и активируют эти рецепторы, включают чувствительные к свету соединения, запахи , феромоны , гормоны и нейротрансмиттеры и различаются по размеру от малых молекул до пептидов к большим белкам . G-белковые рецепторы участвуют во многих заболеваниях.
Существуют два основных пути передачи сигнала, включающих рецепторы, связанные с G-белком:
- Сигнальный путь лагеря и
- Путь сигнала фосфатидилинозитола . [ 5 ]
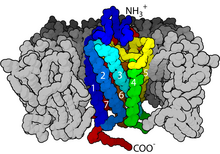
Когда лиганд связывается с GPCR, он вызывает конформационное изменение в GPCR, что позволяет ему действовать как коэффициент обмена нуклеотидов гуанина (GEF). Затем GPCR может активировать связанный G -белок , обменивая ВВП , связанный с GTP, на GTP . Α -субъединица белка, вместе со связанным GTP, может затем диссоциации от субъединиц β и γ, чтобы дополнительно влиять на внутриклеточные сигнальные белки или функциональные белки -мишени непосредственно в зависимости от типа α -субъединицы ( G αS , G α/O , g αq/ 11 , G α12/13 ). [ 6 ] : 1160
GPCRS являются важной целью лекарств и приблизительно 34% [ 7 ] Все утвержденные лекарствами Управления по контролю за продуктами и лекарствами (FDA) нацелены на 108 членов этой семьи. Глобальный объем продаж для этих лекарств оценивается в 180 миллиардов долларов США по состоянию на 2018 год. [update]. [ 7 ] По оценкам, GPCR являются мишенями примерно для 50% лекарств, которые в настоящее время на рынке, в основном из -за их участия в сигнальных путях, связанных со многими заболеваниями, то есть менталиными, метаболическими, включая эндокринологические расстройства, иммунологические, включая вирусные инфекции, сердечно -сосудистые, воспалительные, ощущения, расстройства, расстройства, иммунологические и рак. Давным -давно обнаруженная связь между GPCR и многими эндогенными и экзогенными веществами, приводящими к EG Analgeia, является еще одной динамически развивающейся областью фармацевтических исследований. [ 3 ]
История и значение
[ редактировать ]При определении первой структуры комплекса между G-белком, связанным с рецептором (GPCR) и тримером G-белка (Gαβγ) в 2011 году, была открыта новая глава исследования GPCR для структурных исследований глобальных переключателей с более чем одним белком исследование. Предыдущие прорывы включали определение кристаллической структуры первого GPCR, родопсина, в 2000 году и кристаллической структуры первого GPCR с диффузированным лигандом (β 2 AR) в 2007 году. Путь, в котором семь трансмембранных спиралей GPCR являются Расположенная в пакет был подозревался на основе модели родопсина с низким разрешением лягушки из исследований криогенной электронной микроскопии двумерных кристаллы. Кристаллическая структура родопсина, которая появилась три года спустя, не была неожиданностью, кроме присутствия дополнительной цитоплазматической спирали H8 и точного расположения петли, покрывающего сайт связывания сетчатки. Тем не менее, он обеспечил каркас, который, как было надежда, станет универсальным шаблоном для гомологического моделирования и дизайна лекарств для других GPCR - понятие, которое оказалось слишком оптимистичным.
Семь лет спустя кристаллизация β 2 -адренергического рецептора (β 2 AR) с диффузированным лигандом принесла удивительные результаты, потому что он выявил совершенно другую форму внеклеточной стороны рецептора, чем у родопсина. Эта область важна, потому что она отвечает за связывание лиганда и нацелена на многие лекарства. Более того, сайт связывания лиганда был гораздо более просторным, чем в структуре родопсина, и был открыт для внешнего вида. В других рецепторах кристаллизовались вскоре после этого сторона связывания была еще легче доступна для лиганда. Новые структуры, дополненные биохимическими исследованиями, обнаружили механизмы действия молекулярных переключателей, которые модулируют структуру рецептора, приводящего к состояниям активации для агонистов или к завершению или частичной инактивации для обратных агонистов. [ 3 ]
Нобелевская премия 2012 года по химии была присуждена Брайану Кобилке и Роберту Лефковицу за их работу, которая «имела решающее значение для понимания того, как функционируют рецепторы, связанные с G-белком». [ 8 ] Были по меньшей мере семь других нобелевских призов, присуждаемых за некоторый аспект передачи сигналов, опосредованной G. По состоянию на 2012 год два из десяти мировых бестселлеров ( Advair Diskusus and Abilify ) действуют путем нацеливания на G-белковые рецепторы. [ 9 ]
Классификация
[ редактировать ]
Точный размер сверхсемейства GPCR неизвестен, но, по крайней мере, 831 различные человека гены (или около 4% всего белка-кодирующего генома ) было предсказано, что для них кодирует для них из анализа последовательности генома . [ 10 ] [ 11 ] Хотя были предложены многочисленные схемы классификации, суперсемейство была классически разделена на три основных класса (A, B и C) без обнаруженной гомологии общей последовательности между классами.
Самый большой класс на сегодняшний день - это класс A, который составляет почти 85% генов GPCR. Из GPCR класса A более половины из них предсказывают кодировать обонятельные рецепторы , в то время как оставшиеся рецепторы связаны с известными эндогенными соединениями или классифицируются как сиротские рецепторы . Несмотря на отсутствие гомологии последовательности между классами, все GPCR имеют общую структуру и механизм передачи сигнала . Очень крупная группа родопсина А была дополнительно подразделена на 19 подгрупп ( A1-A19 ). [ 12 ]
Согласно классической системе AF, GPCR могут быть сгруппированы в шесть классов на основе гомологии последовательности и функционального сходства: [ 13 ] [ 14 ] [ 15 ] [ 16 ]
- Класс A (или 1) ( подобный родопсину )
- Класс B (или 2) ( семейство рецепторов секретана )
- Класс C (или 3) ( метаботропный глутамат /феромон)
- Класс D (или 4) ( рецепторы грибкового спаривания феромонов )
- Класс E (или 5) ( циклические рецепторы AMP )
- Класс F (или 6) ( Frizzled / Smoothered )
Совсем недавно альтернативная система классификации под названием GRAFS ( глутамат , родопсин , адгезия , Frizzled / Taste2 , секретарь ) для GPCR. была предложена [ 10 ] Они соответствуют классическим классам C, A, B2, F и B. [ 17 ]
Раннее исследование, основанное на доступной последовательности ДНК, показало, что геном человека кодирует примерно 750 г белковых рецепторов, [ 18 ] Около 350 из которых обнаруживают гормоны, факторы роста и другие эндогенные лиганды. Приблизительно 150 GPCR, обнаруженных в человеческом геноме, имеют неизвестные функции.
Некоторые веб-серверы [ 19 ] и методы прогнозирования биоинформатики [ 20 ] [ 21 ] были использованы для прогнозирования классификации GPCRS в соответствии с их аминокислотной последовательности, посредством подхода псевдокислотного состава .
Физиологические роли
[ редактировать ]GPCR вовлечены в широкий спектр физиологических процессов. Некоторые примеры их физиологических ролей включают:
- Визуальный смысл: опсины используют реакцию фотоизомеризации , чтобы преобразовать электромагнитное излучение в клеточные сигналы. Родопсин , например, использует преобразование 11 -цис -ретиналистов в все транс -ретинальные для этой цели.
- Вкусовой смысл (вкус): GPCR в вкусовых клетках опосредуют высвобождение густдуцина в ответ на вещества с горьким, умми- и сладким вкусом.
- Определение запаха: рецепторы обонятельного эпителий связывают одоранты (обонятельные рецепторы) и феромоны (рецепторы из рвомероназальных)
- Поведенческая регуляция и регуляция настроения: рецепторы в млекопитающих мозге связывают несколько различных нейротрансмиттеров , включая серотонин , дофамин , гистамин , ГАМК и глутамат
- Регуляция активности иммунной системы и воспаления : рецепторы хемокинов связывают лиганды, которые опосредуют межклеточную связь между клетками иммунной системы; Рецепторы, такие как гистаминовые рецепторы, связывают воспалительные медиаторы и привлекают типы клеток -мишеней в воспалительный ответ . GPCR также участвуют в иммунной модуляции, например, регулирование индукции интерлейкина [ 22 ] или подавление TLR -индуцированных иммунных ответов от Т -клеток. [ 23 ]
- Передача вегетативной нервной системы: как симпатические , так и парасимпатические нервные системы регулируются путями GPCR, ответственные за контроль многих автоматических функций организма, таких как артериальное давление, частота сердечных сокращений и пищеварительные процессы
- Ощущение плотности клеток: новая роль GPCR в регуляции чувствительности плотности клеток.
- Модуляция гомеостаза (например, водный баланс). [ 24 ]
- Участвует в росте и метастазировании некоторых типов опухолей . [ 25 ]
- Используется в эндокринной системе для пептидных и аминокислотных производных гормонов, которые связываются с GCPR на клеточной мембране клетки-мишени. Это активирует лагерь, который, в свою очередь, активирует несколько киназ, что позволяет клеточный ответ, такой как транскрипция.
Рецепторная структура
[ редактировать ]GPCR-это интегральные мембранные белки , которые обладают семью мембранными доменами или трансмембранными спиралями . [ 26 ] [ 27 ] Внеклеточные части рецептора могут быть гликозилированы . Эти внеклеточные петли также содержат два высококонсервативных цистеина остатка , которые образуют дисульфидные связи для стабилизации структуры рецептора. Некоторые из семи трансмембранных спиральных белков ( каналродопсин ), которые напоминают GPCR, могут содержать ионные каналы в их белке.
В 2000 году была решена первая кристаллическая структура GPCR млекопитающего, в рамках родопсина бычьего родопсина ( 1F88 ). [ 28 ] В 2007 году была решена первая структура человеческого GPCR [ 29 ] [ 1 ] [ 30 ] Эта структура GPCR β -адренергического рецептора человека оказалась очень похожей на родопсин бычьего родопсина. Структуры активированных или агонистских GPCR также были определены. [ 31 ] [ 32 ] [ 33 ] [ 34 ] Эти структуры указывают на то, как связывание лиганда на внеклеточной стороне рецептора приводит к конформационным изменениям в цитоплазматической стороне рецептора. Самым большим изменением является внешнее движение цитоплазматической части 5 -й и 6 -й трансмембранной спирали (TM5 и TM6). Структура активированного бета-2 адренергического рецептора в комплексе с G S подтвердила, что Gα связывается с полостью, созданным этим движением. [ 35 ]
GPCR демонстрирует структуру, аналогичную некоторым другим белкам с семью трансмембранными доменами , такими как микробные родопсины и рецепторы адипонектина 1 и 2 ( Adipor1 и Adipor2 ). Однако эти рецепторы и каналы и каналы 7TMH (7-трансмембранные спирали) не связываются с G-белками . Кроме того, Adipor1 и Adipor2 ориентированы на противоположности GPCR в мембране (то есть GPCR обычно имеет внеклеточный N-конце , цитоплазматический C-конце , тогда как адипоры перевернуты). [ 36 ]
Структурно -функциональные отношения
[ редактировать ]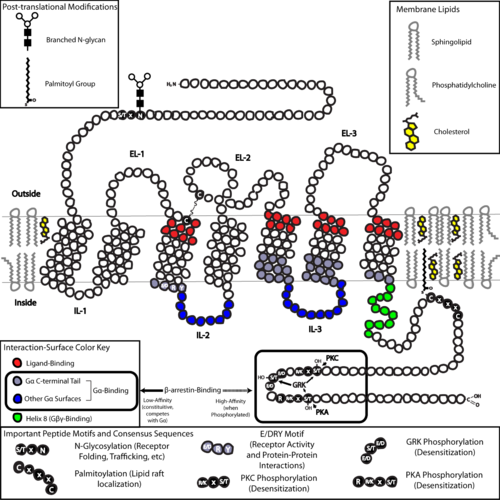
С точки зрения структуры, GPCR характеризуются внеклеточным N-концевым , с последующим семью трансмембранными (7-ТМ) α-спираль (TM-1-TM-7), соединенные тремя внутриклеточными (IL-1-IL-3) и Три внеклеточных петли (EL-1-EL-3) и, наконец, внутриклеточный C-конце . GPCR допускается в третичную структуру , напоминающую ствол, причем семь трансмембранных спиралей образуют полость в плазматической мембране, которая обслуживает лиганд -связывающий домен, который часто покрывается EL -2. Лиганды могут также связываться в другом месте, однако, как и в случае с более массовыми лигандами (например, белками или большими пептидами ), которые вместо этого взаимодействуют с внеклеточными петлями или, как показано метаботропными рецепторами глутамата класса C , N-, N- Терминальный хвост. GPCR класса C отличаются их большим N-концевым хвостом, который также содержит лиганд-связывающий домен. При связывании глутамата с MGLUR N-концевой хвост подвергается конформационному изменению, которое приводит к его взаимодействию с остатками внеклеточных петлей и доменов TM. Возможный эффект всех трех типов Активация, вызванная агонистом , является изменением относительных ориентаций спиралей TM (сравниваемых скручивающим движением), приводящего к более широкой внутриклеточной поверхности и «откровению» остатков внутриклеточных спиралей и доменов TM, имеющих решающее значение для сигнала трансдукции (т.е. -протеиновая связь). Обратные агонисты и антагонисты также могут связываться с рядом различных мест, но возможный эффект должен быть профилактикой этой переориентации спирали TM. [ 3 ]
Структура N- и C-концевых хвостов GPCR также может служить важными функциями за пределами лиганд-связывания. Например, С-концерт мускариновых рецепторов М 3 , и домен с шестью аминокислотами полибазического (KKKRRK) в С-конце необходим для его предотвращения с G Q- белками. [ 37 ] В частности, С-конце часто содержит серина (SER) или треонина остатки (THR), которые при фосфорилировании увеличивают аффинность внутриклеточной поверхности для связывания белков каркаса, называемых β- аррестинами (β-Arr). [ 38 ] После того, как β-арресты стерически предотвращают соединение G-белка и могут рекрутировать другие белки, что приводит к созданию сигнальных комплексов, участвующих в активации пути внеклеточного сигнала киназы ( ERK ) или эндоцитоза рецептора (интернализация). Поскольку фосфорилирование этих остатков Ser и Thr часто происходит в результате активации GPCR, β-ArR-опосредованный G-белок-декорезывание и интернализация GPCR являются важными механизмами десенсибилизации . [ 39 ] Кроме того, интернализованные «мегакомплексы», состоящие из одного GPCR, β-Arr (в конформации хвоста), [ 40 ] [ 41 ] и гетеротримерный G -белок существует и может объяснить передачу сигналов белка из эндосом. [ 42 ] [ 43 ]
Последней общей структурной темой GPCR является пальмитоилирование одного или нескольких участков С-концевого хвоста или внутриклеточных петлей. Палмитоилирование - это ковалентная модификация остатков цистеина (Cys) посредством добавления гидрофобных ацильных групп и оказывает влияние нацеливания рецептора на холестерин и сфинголипид микродомены плазматической мембраны, называемых липидными плотами . Поскольку многие из нижестоящих преобразователей и эффекторных молекул GPCR (включая те, которые участвуют в путях отрицательной обратной связи ), также нацелены на липидные рафты, это имеет эффект облегчения передачи сигналов быстрого рецептора.
GPCR реагируют на внеклеточные сигналы, опосредованные огромным разнообразием агонистов, начиная от белков до биогенных аминов до протонов , но все проводят этот сигнал с помощью механизма связывания G-белка. Это стало возможным благодаря домену гинино -целевого обмена ( GEF ), в основном образованным комбинацией IL-2 и IL-3, а также соседних остатков связанных спиралей TM.
Механизм
[ редактировать ]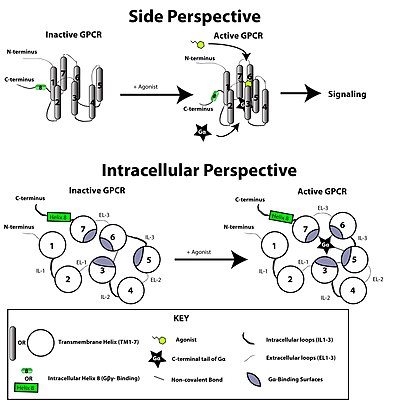
Рецептор, связанный с G-белком, активируется внешним сигналом в форме лиганда или другого сигнального медиатора. Это создает конформационные изменения в рецепторе, вызывая активацию G -белка . Дальнейший эффект зависит от типа G -белка. G -белки впоследствии инактивируются GTPase, активирующими белки, известные как белки RGS .
Связывание лиганда
[ редактировать ]GPCR включают один или несколько рецепторов для следующих лигандов: Сенсорные сигнальные медиаторы (например, световые и обонятельные стимулирующие молекулы); Аденозин , бомбесин , брадикинин , эндотелин , γ-аминобутирическая кислота ( ГАМК ), фактор роста гепатоцитов ( HGF ), меланокортины , нейропептид Y , опиоидные пептиды, опсины , соматостатин , GH , тахикинины , члены вазоактивного интенсивного пептида и вазопопин ; биогенные амины (например, дофамин , адреналин , норэпинефрин , гистамин , серотонин и мелатонин ); глутамат ( метаботропный эффект); глюкагон ; ацетилхолин ( мускариновый эффект); хемокины ; Липидные медиаторы воспаления (например, простагландины , простаноиды , активирующий тромбоциты фактор и лейкотриены ); Пептидные гормоны (например, кальцитонин , анафилатоксин C5A , гормон-стимулирующий фолликул [FSH], гонадотропин-рилизинг-гормон [гнрх], нейрокинин , тиротропин-рилизинг гормон [TRH] и окситоцин ); и эндоканнабиноиды .
GPCR, которые действуют как рецепторы для стимулов, которые еще не были идентифицированы, известны как сиротские рецепторы .
Однако, в отличие от других типов рецепторов, которые были изучены, в которых лиганды связываются внешне с мембраной, лиганды GPCR обычно связываются в трансмембранном домене. Однако активированные протеазой рецепторы активируются путем расщепления части их внеклеточного домена. [ 45 ]
Конформационные изменения
[ редактировать ]
Перевод сигнала через мембрану рецептором не полностью изучена. Известно, что в неактивном состоянии GPCR связан с гетеротримерным комплексом G -белка . Связывание агониста с GPCR приводит к конформационному изменению в рецепторе, которое передается в границе G α -субъединицы гетеротримерного G -белка посредством динамики белкового домена . Активированная субъединица G α обменивается GTP вместо GDP , что, в свою очередь, запускает диссоциацию субъединицы G α из димера G βγ и от рецептора. Дисонцированные субъединицы G α и G βγ взаимодействуют с другими внутриклеточными белками, чтобы продолжить каскад трансдукции сигнала, в то время как Freed GPCR способен переподходить в другой гетеротримерный G -белок с образованием нового комплекса, который готов инициировать другой раунд трансдукции сигнала. [ 46 ]
Считается, что рецепторная молекула существует в конформационном равновесии между активными и неактивными биофизическими состояниями. [ 47 ] Связывание лигандов с рецептором может сдвинуть равновесие в направлении активных рецепторных состояний. Существуют три типа лигандов: агонисты - это лиганды, которые сдвигают равновесие в пользу активных состояний; Обратные агонисты - это лиганды, которые сдвигают равновесие в пользу неактивных состояний; и нейтральные антагонисты - это лиганды, которые не влияют на равновесие. Пока не известно, как именно активные и неактивные состояния отличаются друг от друга.
G-белок активации/цикл дезактивации
[ редактировать ]
Когда рецептор неактивен, домен GEF может быть связан с также неактивной α-субъединицей гетеротримерного G-белка . Эти «G-белок» представляют собой тример субъединиц α, β и γ (известных как Gα, Gβ и Gγ, соответственно), которые становятся неактивными, когда обратимо связаны с гинозинфосфатом (GDP) (или, альтернативно, нет гванового ядлеотеда. ), но активно, когда связывается с гуанозиновым трифосфатом (GTP). После активации рецептора домен GEF, в свою очередь, аллостерически активирует G-белок, облегчая обмен молекулой ВВП для GTP на α-субъединице G-белка. Клетка поддерживает соотношение 10: 1 цитозольного GTP: GDP, поэтому обеспечивается обмен на GTP. В этот момент субъединицы G-белка диссоциируют от рецептора, а также друг друга, чтобы получить мономер Gα-GTP и тесно взаимодействующий димер Gβγ , которые теперь могут свободны модулировать активность других внутриклеточных белков. Степень, в которой они могут диффундировать , однако, ограничена из -за пальмитоилирования Gα и наличия Изопреноидная часть, которая была ковалентно добавлена к С-концам Gγ.
Поскольку gα также обладает медленной способностью GTP → GDP-гидролиза , в конечном итоге регенерируется неактивная форма α-субъединицы (Gα-GDP), что позволяет повторно связываться с димером Gβγ для образования «покоящегося» G-белка, который может снова связываться с GPCR и ожидает активации. Скорость гидролиза GTP часто ускоряется из-за действий другого семейства аллостерических модулирующих белков, называемых регуляторами передачи сигналов G-белка , или RGS-белков, которые являются типом белка, активирующим GTPase , или зазором. Фактически, многие из первичных эффекторных белков (например, аденилатциклазы ), которые активируются/инактивированы при взаимодействии с Gα-GTP, также обладают активностью GAP. Таким образом, даже на этой ранней стадии процесса передача сигналов, инициированная GPCR, обладает способностью к самооценке.
Перекрестные помехи
[ редактировать ]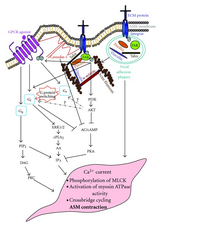
Было показано, что GPCR вниз по течению, возможно, взаимодействуют с интегриновыми сигналами, такими как FAK . [ 48 ] Передача сигналов интегрина будет фосфорилировать FAK, который затем может снизить GPCR G αS активность .
Сигнализация
[ редактировать ]
Если рецептор в активном состоянии встречается с G -белком , он может активировать его. Некоторые данные свидетельствуют о том, что рецепторы и G-белки фактически предварительно связаны. [ 37 ] Например, связывание G -белков с рецепторами влияет на сродство рецептора к лигандам. Активированные G -белки связаны с GTP .
Дальнейшая передача сигнала зависит от типа G -белка. Фермент аденилатциклаза является примером клеточного белка, который может регулироваться белком G, в данном случае G. -белок G Активность аденилатциклазы активируется, когда она связывается с субъединицей активированного G -белка. Активация аденилатциклазы заканчивается, когда G -белок возвращается в состояние GDP .
Аденилатциклазы (из которых 9 мембранных и одну цитозольную форму известны у людей) также могут быть активированы или ингибированы другими способами (например, связывание Ca2+/ кальмодулин ), которые могут модифицировать активность этих ферментов в аддитивной или синергической моде) наряду с G -белками.
Сигнальные пути, активируемые через GPCR, ограничены первичной последовательности и третичной структурой самого GPCR, но в конечном итоге определяются конкретной конформацией, стабилизированной конкретным лигандом , а также наличием молекул преобразователя . В настоящее время считается, что GPCR используют два основных типа преобразователей: G-белковые и β-аресресты . Поскольку β-Arr обладает высоким сродством только к фосфорилированной форме большинства GPCR (см. Выше или ниже), большая часть передачи сигналов в конечном итоге зависит от активации G-белка. Тем не менее, возможность взаимодействия позволяет возникнуть, независимая от G-белка.
G-белок-зависимая передача сигналов
[ редактировать ]Существуют три основных G-белковые сигнальные пути, опосредованные четырьмя подклассами G-белков, отличающимися друг от друга путем гомологии последовательности ( G αS , G αI/O , G αQ/11 и G α12/13 ). Каждый подкласс G-белка состоит из нескольких белков, каждый из которых является продуктом множественных генов или вариаций сплайсинга , которые могут наполнить их различиями, от тонких до отчетливых в отношении сигнальных свойств, но в целом они, по-видимому, разумно сгруппированы в четыре класса. Поскольку сигнальные трансдуцирующие свойства различных возможных комбинаций βγ , по-видимому, не имеют радикально отличаются друг от друга, эти классы определяются в соответствии с изоформой их α-субъединицы. [ 6 ] : 1163
В то время как большинство GPCR способны активировать более одного Gα-Subtype, они также показывают предпочтение одному подтипу над другим. Когда активированный подтип зависит от лиганда, связанного с GPCR, это называется функциональной селективностью (также известной как перенос, направленный агонистами, или специфичный для конформации агонизм). Однако связывание любого отдельного агониста также может инициировать активацию множества различных G-белков, поскольку оно может быть способно стабилизировать более чем одну конформацию GPCR GEF- домена, даже в течение одного взаимодействия. Кроме того, конформация, которая предпочтительно активирует одну изоформу Gα, может активировать другую, если предпочтение будет менее доступна. Кроме того, обратной связи пути может привести к модификациям рецепторов (например, фосфорилирование), которые изменяют предпочтение G-белка. Независимо от этих различных нюансов, предпочтительный партнер GPCR от GPCR обычно определяется в соответствии с G-белком, наиболее явно активируемым эндогенным лигандом в большинстве физиологические или экспериментальные условия.
Передача сигналов Gα
[ редактировать ]- Эффектором как G αS , так и G αi/O является циклическим аденозиновым монофосфатом (ЦАМ-графство), генерирующей ферментной аденилатциклазы или AC. Несмотря на то, что у млекопитающих существует десять различных продуктов гена переменного тока, каждый из которых имеет тонкие различия в тканей распределении или функции , все катализируют преобразование цитозольного аденозин-трифосфата (АТФ) в лагерь, и все они непосредственно стимулированы G-белками класса G αS . Напротив, однако, взаимодействие с субъединицами Gα типа G αI/O ингибирует AC с генерирующим цАМФ. Таким образом, GPCR, связанный с G αS, противодействует действию GPCR, связанного с G αI/O , и наоборот. Уровень цитозольного лагеря может затем определить активность различных ионных каналов , а также членов семейства Ser/Thr-специфической протеинкиназы A (PKA). Таким образом, лагерь считается вторым посланником , а PKA вторичным эффектором .
- Эффектор пути G αQ/11 представляет собой фосфолипазу C-β (PLCβ), который катализирует расщепление мембрановой фосфатидилинозитол 4,5-бисфосфат ) во второй мессенджер . ( PIP2 ) и диацилглицерин (DAG). IP3 действует на рецепторы IP3 , обнаруженные в мембране эндоплазматического ретикулума (ER), чтобы вызвать CA 2+ высвобождается из ER, в то время как DAG диффундирует вдоль плазматической мембраны , где она может активировать любые мембранные локализованные формы второй Ser/Thr -киназы, называемой протеинкиназой C (PKC). Поскольку многие изоформы PKC также активируются увеличением внутриклеточного CA 2+ , оба эти пути также могут сходиться друг к другу, чтобы сигнализировать через один и тот же вторичный эффектор. Повышенный внутриклеточный CA 2+ Также связывается и аллостерически активирует белки, называемые кальмодулинами , которые, в свою очередь, имеют тозолическую малую GTPase , Rho . После того, как он связан с GTP, Rho может продолжать активировать различные белки, ответственные за регуляцию цитоскелета , такие как рокиназа (ROCK). Большинство GPCR, которые от пары до G α12/13 также соединяются с другими подклассами, часто G αQ/11 .
Передача сигналов Gβγ
[ редактировать ]Приведенные выше описания игнорируют эффекты gβγ -опоры, что также может быть важно, в частности в случае активированного G αi/O -священного GPCR. Основными эффекторами Gβγ являются различные ионные каналы, такие как G-белок, регулируемая внутренне выпрямляющей K + каналы (Girks), P / Q -и N-тип , управляемый CA 2+ Каналы , а также некоторые изоформы переменного тока и ПЛК, а также некоторые изоформы фосфоинозитид-3-киназы (PI3K).
G-белок-независимая передача сигналов
[ редактировать ]Хотя они классически рассматриваются только о совместной работе, GPCR могут сигнализировать через G-белок-независимые механизмы, а гетеротримерные G-белок могут играть функциональные роли, независимые от GPCR. GPCR могут сигнализировать независимо через многие белки, уже упомянутые для их роли в G-белонезависимой передаче сигналов, такой как β-ARR , GRK и SRC . Было показано, что такая передача сигналов физиологически актуальна, например, передача сигналов β-арреста , опосредованная рецептором хемокина CXCR3, была необходима для полной эффективности хемотаксис активированных Т-клеток. [ 49 ] Кроме того, дальнейшие белки каркаса, участвующие в субклеточной локализации GPCR (например, белки, содержащие PDZ -домен )), также могут действовать в качестве преобразователей сигнала. Чаще всего эффектор является членом семьи MAPK .
Примеры
[ редактировать ]В конце 1990 -х годов начали накапливаться доказательства, чтобы предположить, что некоторые GPCR могут сигнализировать без G -белков. Митоген -активируемая митогеном протеинкиназа, ключевой медиатор трансдукции сигнала ниже активации рецептора во многих путях, активируется в ответ на ЦВЕТ-опосредованную активацию рецептора в плесени слизки D. Дискостик, несмотря на отсутствие связанного белка G-белка. α- и β-субъединицы. [ 50 ]
Было продемонстрировано , что в клетках млекопитающих был продемонстрирован широко изученный β 2- адренорецептор, активирует путь ERK2 после опосредованного аррестином, опосредованным G-белкопосредованной передачей сигналов. Следовательно, представляется вероятным, что некоторые механизмы, ранее считавшиеся связанными исключительно с рецепторной десенсибилизацией, на самом деле являются примерами рецепторов, переключающих их сигнальный путь, а не просто отключаются.
В клетках почек было показано, что рецептор Bradykinin B2 напрямую взаимодействует с белковой тирозинфосфатазой. Присутствие тирозин-фосфорилированной последовательности ITIM (иммунорецептор-тирозин-ингибирующий мотив) в рецепторе B2 необходим для опосредования этого взаимодействия и впоследствии антипролиферативного эффекта бредикинина. [ 51 ]
GPCR-независимая передача сигналов гетеротримерными G-белками
[ редактировать ]Хотя это относительно незрелая область исследований, представляется, что гетеротримерные G-белок также могут принять участие в передаче сигналов без GPCR. Существуют доказательства ролей в качестве датчиков сигнала практически во всех других типах рецептор-опосредованной передачи сигналов, включая интегрины , рецепторные тирозинкиназы (RTK), рецепторы цитокинов ( JAK/STATS ), а также модуляция различных других «дополнительных» белков, таких как, как GEFS , гуанино-нуклеотидные ингибиторы диссоциаторы (GDI) и белковые фосфатазы . Там может быть даже специфические белки этих классов, первичная функция которых является частью GPCR-независимых путей, называемых активаторами передачи сигналов G-белка (AGS). Как повсеместное распространение этих взаимодействий, так и важность Gα и Gβγ -субъединиц для этих процессов до сих пор неясны.
Детали путей лагеря и PIP2
[ редактировать ]


Существуют два основных пути передачи сигнала, включающих G-белковые рецепторы : путь сигнала CAMP и сигнальный путь фосфатидилинозитола . [ 5 ]
Camp Signal Pathway
[ редактировать ]Трансдукция сигнала CAMP содержит пять основных персонажей: стимулирующий гормональный рецептор (RS) или ингибирующий гормон рецептор (RI); Стимулирующий регулятивный G-белок (GS) или ингибирующий регулятивный G-белок (GI); Аденилилайкклаза ; протеинкиназа А (ПКА); и лагерь фосфодиэстераза .
Стимулирующий рецептор гормонов (RS) является рецептором, который может связываться с молекулами стимулирующих сигналов, в то время как ингибирующий гормон -рецептор (RI) является рецептором, который может связываться с ингибирующими молекулами сигнала.
Стимулирующий регулятивный G-белок представляет собой G-белок, связанный с рецептором стимулирующего гормона (RS), а его α-субъединица при активации может стимулировать активность фермента или другого внутриклеточного метаболизма. Напротив, ингибирующий регулятивный G-белок связан с ингибирующим гормональным рецептором, а его α-субъединица при активации может ингибировать активность фермента или другого внутриклеточного метаболизма.
Аденилилуциклаза представляет собой 12-трансмембранный гликопротеин, который катализирует преобразование АТФ в лагерь с помощью кофактора mg 2+ или mn. 2+ Полем Производившийся лагерь является вторым посланником в клеточном метаболизме и является аллостерическим активатором протеинкиназы A.
Протеинкиназа А является важным ферментом в метаболизме клеток из -за его способности регулировать метаболизм клеток путем фосфорилирования специфических преданных ферментов в метаболическом пути. Он также может регулировать специфическую экспрессию генов, клеточную секрецию и проницаемость мембраны. Белковый фермент содержит две каталитические субъединицы и две регуляторные субъединицы. Когда нет лагеря, комплекс неактивен. Когда лагерь связывается с регуляторными субъединицами, их конформация изменяется, вызывая диссоциацию регуляторных субъединиц, которая активирует протеинкиназу А и обеспечивает дальнейшие биологические эффекты.
Эти сигналы затем могут быть прекращены CAMP-фосфодиэстеразой, которая представляет собой фермент, который разлагает лагерь до 5'-AMP и инактивирует протеинкиназу A.
Фосфатидилинозитол сигнальный путь
[ редактировать ]В пути сигнального пути фосфатидилинозитола внеклеточная сигнальная молекула связывается с рецептором G-белка (G Q ) на клеточной поверхности и активирует фосфолипазу C , которая расположена на плазматической мембране . Липаза ( гидролизует фосфатидилинозитол 4,5-бисфосфат PIP2) в два вторых посланника: Инозитол 1,4,5-трисфосфат (IP3) и диацилглицерин (DAG). IP3 связывается с рецептором IP3 в мембране гладкой эндоплазматической ретикулумы и митохондрий для открытия CA 2+ каналы. DAG помогает активировать протеинкиназу C (PKC), которая фосфорилирует многие другие белки, изменяя их каталитическую активность, что приводит к клеточным реакциям.
Эффект CA 2+ Также замечательны: он сотрудничает с DAG в активации PKC и может активировать путь киназы камеры , в котором кальций-модулированный белком кальмодулин (CAM) связывается CA 2+ претерпевает изменение конформации и активирует камучную киназу II, которая обладает уникальной способностью повысить свою аффинность связывания с CAM путем аутофосфорилирования, делая CAM недоступным для активации других ферментов. Затем киназа фосфорилирует целевые ферменты, регулируя свою деятельность. Два пути сигнала соединены вместе CA 2+ -Кэм, которая также является регуляторной субъединицей аденилатциклазы и фосфодиэстеразы в сигнальном пути ЦАМФ.
Рецепторное регулирование
[ редактировать ]GPCR становятся десенсибилизированными при воздействии их лиганда в течение длительного периода времени. Существуют две признанные формы десенсибилизации: 1) гомологичная десенсибилизация , в которой активированный GPCR подавляется; и 2) гетерологичная десенсибилизация , в которой активированный GPCR вызывает подавление другого GPCR. Ключевой реакцией этого подавления является фосфорилирование внутриклеточного (или цитоплазматического ) домена рецептора протеинкиназами .
Фосфорилирование с помощью цАМФ-зависимых протеинкиназ
[ редактировать ]Циклические ампер-зависимые протеинкиназы ( протеинкиназа А ) активируются сигнальной цепью, поступающей из G-белка (который был активирован рецептором) через аденилатциклазу и циклический AMP (CAMP). В механизме обратной связи эти активированные киназы фосфорилируют рецептор. Чем дольше рецептор остается активным, тем больше киназ активируется и чем больше рецепторов фосфорилируются. В β 2 -адренорецепторах это фосфорилирование приводит к переключению связи с класса G S класса G -белка на G I. класс [ 52 ] ЦАМ-зависимое PKA-опосредованное фосфорилирование может вызвать гетерологичную десенсизирование у рецепторов, отличных от активированных. [ 53 ]
Фосфорилирование GRK
[ редактировать ]GR -рецепторные киназы, связанные с G , представляют собой протеинкиназы, которые фосфорилируют только активные GPCR. [ 54 ] Рецепторные киназы, связанные с G-белком (GRK), являются ключевыми модуляторами передачи сигналов рецептора, связанного с G-белком (GPCR). Они представляют собой семейство семи серин-треониновых протеинкиназ млекопитающих, которые фосфорилируют рецептор, связанный с агонистами. ГРК-опосредованное фосфорилирование рецептора быстро инициирует глубокое нарушение передачи сигналов и десенсибилизации рецептора. Активность ГРК и субклеточного нацеливания жестко регулируется взаимодействием с доменами рецептора, субъединицами G-белка, липидами, закреплением белков и чувствительных к кальцие белкам. [ 55 ]
Фосфорилирование рецептора может иметь два последствия:
- Транслокация : рецептор, наряду с частью мембраны, в которую он встроен, приведен на внутреннюю часть клетки, где он дефосфорилируется в кислой везикулярной среде [ 56 ] а затем вернул. Этот механизм используется для регулирования долгосрочного воздействия, например, на гормон, путем разрешения разрешения следовать десенсисизации. Альтернативно, рецептор может подвергаться лизозомальной деградации или оставаться интернализованным, где, как полагают, он участвует в инициации сигнальных событий, характер которого в зависимости от внутренней локализации субклеточной локализации пузырька. [ 53 ]
- Arrestin Связывание : фосфорилированный рецептор может быть связан с молекулами арестина , которые предотвращают его связывание (и активируют) G -белки, фактически сменяя его на короткий период времени. Этот механизм используется, например, с родопсином в сетчатке для компенсации воздействия яркого света. Во многих случаях связывание Аррестина с рецептором является предпосылкой для транслокации. Например, бета-аррестин, связанный с β 2- адренорецепторами, действует как адаптер для связывания с клатрином и с бета-субъединицей AP2 (молекулы адаптера клатрина); Таким образом, Arrestin здесь действует как каркас, собирающий компоненты, необходимые для клатрин -опосредованного эндоцитоза β 2 -адренорецепторов. [ 57 ] [ 58 ]
Механизмы прекращения сигнала GPCR
[ редактировать ]Как упомянуто выше, G-белки могут прекратить свою собственную активацию из-за их внутренней способности GTP → GDP-гидролиза . Тем не менее, эта реакция продолжается медленной скоростью (≈0,02 раза/сек), и, таким образом, потребуется около 50 секунд для любого отдельного G-белка, чтобы деактивировать, если другие факторы не вступили в игру. Действительно, существует около 30 изоформ белков RGS , которые, когда они связаны с Gα через их домен GAP , ускоряют скорость гидролиза до ≈30 раз/с. Это 1500-кратное увеличение скорости позволяет ячейке реагировать на внешние сигналы с высокой скоростью, а также пространственное разрешение из-за ограниченного количества второго мессенджера , которое можно сгенерировать, и ограниченное расстояние G-белок может диффундировать за 0,03 секунды. По большей части, белки RGS беспорядочны по своей способности деактивировать G-белок, в то время как RGS участвует в данном сигнальном пути, представляется более определяемой ткани и GPCR, чем все остальное. Кроме того, белки RGS имеют дополнительную функцию увеличения скорости обмена GTP-GDP при GPCRS (то есть как своего рода CO-GEF), что еще больше способствует временному разрешению передачи сигналов GPCR.
Кроме того, GPCR может быть десенсибилизирован сам. Это может произойти как:
- Прямой результат занятия лиганда , в котором изменение конформации позволяет рекрутировать GPCR-регулирующие киназы (GRK), которые приводят к фосфорилированию различных остатков серина / треонина IL-3 и C-концевого хвоста. аффиние GPCR к β-аррестину После фосфорилирования GRK повышается (β-аррестин-1/2 в большинстве тканей), после чего β-аррестин может связывать и действовать, чтобы как стерически затруднить соединение G-белка, так и инициировать процесс и инициатор рецепторной интернализации посредством клатрин-опосредованного эндоцитоза . Поскольку только рецептор с лигированным чувствительным чувствительным к этому механизму он называется гомологичной десенсибилизацией
- Аффинность к β-аррестину может быть увеличена в лиганде, и не независимо от ГР, посредством фосфорилирования различных сайтов Ser/Thr (но также IL-3 и C-концевого хвоста) с помощью PKC и PKA. Эти фосфорилирования часто достаточны для самостоятельного ухудшения связей G-белка. [ 59 ]
- Вместо этого PKC/PKA может, фосфорилируйте GRK, что также может привести к фосфорилированию GPCR и связыванию β-аррестина независимым от профессии. Эти последние два механизма позволяют десенсибилизацию одного GPCR из -за деятельности других или гетерологичной десенсибилизации . GRK также могут иметь домены разрыва и, следовательно, также могут способствовать инактивации, а через не киназные также механизмы. Комбинация этих механизмов также может произойти.
Как только β-аррестин связан с GPCR, он претерпевает конформационное изменение, позволяющее ему служить белком для каркаса для адаптерного комплекса, называемого AP-2 , который, в свою очередь, рекрутирует другой белок, называемый клатрин . Если достаточное количество рецепторов в местном районе намекает клатрин, они агрегируют и мембранные пород внутренне в результате взаимодействия между молекулами клатрина, в процессе, называемом опсонизацией . После того, как яма была ущипнута от плазматической мембраны из -за действий двух других белков, называемых амфифизином и динамином , теперь это эндоцитарная пузырька . В этот момент молекулы адаптера и клатрин диссоциировались , и рецептор либо перемещается обратно на плазматическую мембрану, либо нацелен на лизосомы для деградации .
В любой точке этого процесса β-аррестины могут также рекрутировать другие белки, такие как нерецепторная тирозинкиназа (NRTK), C-SRC , которая может активировать ERK1/2 или другую митогенную протеинкиназу (MAPK) Сигнализация, например, фосфорилирование небольшой GTPase , RAS рекрутировать белки каскада ERK или напрямую (IE, RAF-1 , MEK , ERK-1/2), в какой точке сигнализация инициируется из-за их непосредственной близости друг к другу. Другой мишенью C-SRC являются молекулы динамина, участвующие в эндоцитозе. Динамины полимеризуются вокруг шеи входящей пузырьки, а их фосфорилирование с помощью C-SRC обеспечивает энергию, необходимую для конформационного изменения, позволяющего окончательно «зажать» от мембраны.
Клеточная регуляция GPCR
[ редактировать ]Десенсибилизация рецепторов опосредуется посредством комбинированного фосфорилирования, связывания β-Arr и эндоцитоза, как описано выше. Понижающая регуляция возникает, когда эндоцитозированный рецептор встроен в эндосому, которая обезвреживается, чтобы слияние с органеллой, называемой лизосомой. Поскольку лизосомальные мембраны богаты протонными насосами, их интерьеры имеют низкий pH (≈4,8 против цитозоля ph≈7,2), который действует для денатуры GPCR. Кроме того, лизосомы содержат много деградирующих ферментов , включая протеазы, которые могут функционировать только при таком низком pH, и поэтому пептидные связи, соединяющие остатки GPCR вместе. Независимо от того, находится ли дан данным рецептором в лизосому, задерживается в эндосомах или переезжает обратно в плазматическую мембрану, зависит от множества факторов, включая тип рецептора и величину сигнала. Регуляция GPCR дополнительно опосредуется факторами транскрипции генов. Эти факторы могут увеличить или уменьшить транскрипцию генов и, таким образом, увеличивать или уменьшить генерацию новых рецепторов (повышение или понижение), которые перемещаются в клеточную мембрану.
Рецепторная олигомеризация
[ редактировать ]Олигомеризация рецептора, связанная с G-белком, является широко распространенным явлением. Одним из лучших примеров является GABA B. рецептор метаботропный Этот так называемый конститутивный рецептор образуется путем гетеродимеризации субъединиц GABA B R1 и GABA B R2 . Экспрессия GABA B R1 без GABA B R2 в гетерологичных системах приводит к удержанию субъединицы в эндоплазматической ретикулуме . Экспрессия только субъединицы GABA B R2, тем временем, приводит к поверхностной экспрессии субъединицы, хотя без функциональной активности (то есть рецептор не связывает агонист и не может инициировать ответ после воздействия агониста). Экспрессия двух субъединиц вместе приводит к экспрессии плазматической мембраны функционального рецептора. Было показано, что GABA B связывание R2 с GABA B R1 вызывает маскировку сигнала удержания [ 60 ] функциональных рецепторов. [ 61 ]
Происхождение и диверсификация суперсемейства
[ редактировать ]Сигнальная трансдукция, опосредованная суперсемейством GPCR, восходит к происхождению многоклеточности . GPCR, подобные млекопитающим, встречаются в грибах и были классифицированы в соответствии с системой классификации GRAFS на основе отпечатков пальцев GPCR. [ 17 ] Идентификация членов суперсемейства через эукариотическую область и сравнение мотивов, специфичных для семейства, показали, что суперсемейство GPCRS имеет общее происхождение. [ 62 ] Характерные мотивы указывают на то, что три из пяти семейств Grafs, родопсин , адгезия и вьющиеся в Frizzed , развивались из рецепторов Dictyostelium discoideum camp до разделения опишхоконтов . Позже семейство секретанов развивалось из семейства рецепторов GPCR адгезии до разделения нематод . [ 17 ] GPCR насекомых, по -видимому, находятся в их собственной группе, а Taste2 идентифицируется как спуск от родопсина . [ 62 ] Обратите внимание, что разделение секретина / адгезии основано на предполагаемой функции, а не на подписи, поскольку классический класс B (7TM_2, PFAM PF00002 ) используется для идентификации оба в исследованиях.
Смотрите также
[ редактировать ]- G-белковые рецепторы база данных рецепторов
- Список кодов сетки (D12.776)
- Метаботропный рецептор
- Рецептор сироты
- Pepducins , класс кандидатов на наркотики, нацеленные на GPCRS
- Рецептор активирован исключительно синтетическим лигандом , методом контроля передачи сигналов клеток с помощью синтетических GPCRS
- Взял суперсемейство
Ссылки
[ редактировать ]- ^ Подпрыгнуть до: а беременный Cherezov V, Rosenbaum DM, Hanson MA, Rasmussen SG, Thian FS, Kobilka TS, et al. (Ноябрь 2007 г.). «Кристаллическая структура с высоким разрешением инженерного рецептора, связанного с G-белком человека, бета-адренергического, связанного с G» . Наука . 318 (5854): 1258–65. Bibcode : 2007sci ... 318.1258c . doi : 10.1126/science.1150577 . PMC 2583103 . PMID 17962520 .
- ^ Подпрыгнуть до: а беременный Чжан, Цзянь В.; Ли, Лей; Хуан, Цинсенг; Рен, Пей-Ген (1 января 2013 г.). «Глава третья - Рецептор Обестатина в энергетическом гомеостазе и патогенезе ожирения» . В Дао, Я-Синг (ред.). Прогресс в молекулярной биологии и трансляционной науке . G-белковые рецепторы в энергетическом гомеостазе и патогенезе ожирения. Тол. 114. Академическая пресса. С. 89–107. doi : 10.1016/b978-0-12-386933-3.00003-0 . ISBN 9780123869333 Полем PMID 23317783 . Архивировано из оригинала 17 января 2023 года . Получено 24 октября 2023 года .
- ^ Подпрыгнуть до: а беременный в дюймовый и Trzaskowski B, Laatk D, Yuan S, Ghoshdastider U, Debinski A, Filipek S (2012). «Действие молекулярных переключателей в GPCR-теоретические и экспериментальные исследования» . Текущая лекарственная химия . 19 (8): 1090–109. doi : 10.2174/092986712799320556 . PMC 3343417 . PMID 22300046 .
 Текст был скопирован из этого источника, который доступен по атрибуции 2.5 Generic (CC по 2.5) лицензии
Текст был скопирован из этого источника, который доступен по атрибуции 2.5 Generic (CC по 2.5) лицензии
- ^ Кинг Н., Хиттингер К.Т., Кэрролл С.Б. (июль 2003 г.). «Эволюция ключевых клеточных сигналов и семейств адгезии белков предшествует происхождению животных». Наука . 301 (5631): 361–3. Bibcode : 2003sci ... 301..361K . doi : 10.1126/science.1083853 . PMID 12869759 . S2CID 9708224 .
- ^ Подпрыгнуть до: а беременный Гилман А.Г. (1987). «G-белки: преобразователи сигналов, сгенерированных рецептором». Ежегодный обзор биохимии . 56 (1): 615–49. doi : 10.1146/annurev.bi.56.070187.003151 . PMID 3113327 .
- ^ Подпрыгнуть до: а беременный Wettschureck N, Spertmanns S (октябрь 2005 г.). «Белки G млекопитающего G и их специфические функции типа клеток». Физиологические обзоры . 85 (4): 1159–204. doi : 10.1152/physrev.00003.2005 . PMID 16183910 .
- ^ Подпрыгнуть до: а беременный Hauser AS, Chavali S, Crink I, Jahn LJ, Morceanov, Gloriam De, Bubu MM (Janahuary 2018). Полем Смеситель 172 (1–2): 41–54,19. doi : 10.1016/j.cell . PMC 5766829 . PMID 29249361 .
- ^ Королевская шведская академия наук (10 октября 2012 г.). «Нобелевская премия по химии 2012 Роберт Дж. Лефковиц, Брайан К. Кобилка» . Получено 10 октября 2012 года .
- ^ Линдсли CW (июнь 2013 г.). «Лучшие рецептурные препараты 2012 года во всем мире: доминируют биологии, но лекарства CNS с небольшими молекулами держатся на верхних местах» . ACS Химическая нейробиология . 4 (6): 905–7. doi : 10.1021/cn400107y . PMC 3689196 . PMID 24024784 .
- ^ Подпрыгнуть до: а беременный в Bjarnadóttir TK, Gloriam DE, Hellstrand SH, Kristiansson H, Fredriksson R, Schiöth HB (сентябрь 2006 г.). «Комплексный репертуар и филогенетический анализ G-белковых рецепторов у человека и мыши». Геномика . 88 (3): 263–73. doi : 10.1016/j.ygeno.2006.04.001 . PMID 16753280 .
- ^ «Ключевое слово:« G-белок, связанный с рецептором [KW-0297] »и организм:« Homo Sapiens (человек) [9606] »в Uniprotkb» . www.uniprot.org . Архивировано из оригинала 15 сентября 2020 года . Получено 24 июня 2019 года .
- ^ JOOST P, Metner A (октябрь 2002 г.). «Филогенетический анализ 277 рецепторов, связанных с G-белком человека в качестве инструмента для прогнозирования лигандов рецепторов сирот» . Биология генома . 3 (11): Research0063. doi : 10.1186/gb-2002-3-11-research0063 . PMC 133447 . PMID 12429062 .
- ^ Attwood TK, Findlay JB (февраль 1994 г.). «Рецепторы отпечатков отпечатков пальцев G-белок». Протеиновая инженерия . 7 (2): 195–203. doi : 10.1093/белок/7.2.195 . PMID 8170923 .
- ^ Колаковский Л.Ф. (1994). «GCRDB: база данных рецепторов, связанных с G-белком». Рецепторы и каналы . 2 (1): 1–7. PMID 8081729 .
- ^ Foord SM, Bonner Ti, Neubig RR, Rosser EM, PIN JP, Davenport AP, et al. (Июнь 2005 г.). «Международный союз фармакологии. XLVI. G-списка рецепторов, связанных с белком». Фармакологические обзоры . 57 (2): 279–88. doi : 10.1124/pr.57.2.5 . PMID 15914470 . S2CID 34541683 .
- ^ "InterPro" . Архивировано из оригинала 21 февраля 2008 года . Получено 10 декабря 2007 года .
- ^ Подпрыгнуть до: а беременный в Кришнан А., Алмен М.С., Фредрикссон Р., Шиёт Х.Б. (2012). XUE C (ред.). «Происхождение GPCR: идентификация таких млекопитающих, как родопсин, адгезия, глутамат и вьющиеся GPCR в грибах» . Plos один . 7 (1): E29817. BIBCODE : 2012PLOSO ... 729817K . doi : 10.1371/journal.pone.0029817 . PMC 3251606 . PMID 22238661 .
- ^ Vassilatis DK, Hohmann JG, Zeng H, Li F, Ranchalis JE, Mortrud MT, et al. (Апрель 2003 г.). «G-белковые рецепторные репертуары человека и мыши» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (8): 4903–8. Bibcode : 2003pnas..100.4903V . doi : 10.1073/pnas.0230374100 . PMC 153653 . PMID 12679517 .
- ^ Xiao X, Wang P, Chou KC (июль 2009 г.). «GPCR-CA: подход к изображению Automaton Cellular для прогнозирования функциональных классов рецепторов, связанных с G-белком» . Журнал вычислительной химии . 30 (9): 1414–23. doi : 10.1002/jcc.21163 . PMID 19037861 . S2CID 813484 . Архивировано с оригинала 9 апреля 2017 года.
- ^ Qiu JD, Huang JH, Liang RP, Lu XQ (июль 2009 г.). «Прогнозирование классов рецепторов, связанных с G-белком, на основе концепции псевдокислотного состава Чоу: подход от дискретного вейвлет-преобразования». Аналитическая биохимия . 390 (1): 68–73. doi : 10.1016/j.ab.2009.04.009 . PMID 19364489 .
- ^ Gu Q, Ding YS, Zhang TL (май 2010 г.). «Прогнозирование классов рецепторов, связанных с G-белком в низкой гомологии с использованием псевдокислотного состава Чоу с приблизительной энтропией и гидрофобными паттернами». Белковые и пептидные буквы . 17 (5): 559–67. doi : 10.2174/092986610791112693 . PMID 19594431 .
- ^ Сароз Y, Kho DT, Glass M, Graham ES, Grimsey NL (декабрь 2019 г.). «Рецептор каннабиноида 2 (CB 2 ) через G-альфа-S и индуцирует секрецию цитокинов IL-6 и IL-10 в первичных лейкоцитах человека» . ACS Фармакология и трансляционная наука . 2 (6): 414–428. doi : 10.1021/acsptsci.9b00049 . PMC 7088898 . PMID 32259074 .
- ^ Шарма Н., Ахаде А.С., Кадри А (апрель 2013 г.). «Сфингозин-1-фосфат подавляет TLR-индуцированную секрецию CXCL8 из Т-клеток человека». Журнал биологии лейкоцитов . 93 (4): 521–8. doi : 10.1189/jlb.0712328 . PMID 23345392 . S2CID 21897008 .
- ^ Хейзелл Г.Г., Хиндмарч С.К., Папа Г.Р., Ропер Дж. А., Лайтман С.Л., Мерфи Д. и др. (Январь 2012 г.). «G-белковые рецепторы в гипоталамических паравентрикулярных и супраоптических ядрах-сперцентире к нейроэндокринному гомеостазу» . Границы в нейроэндокринологии . 33 (1): 45–66. doi : 10.1016/j.yfrne.2011.07.002 . PMC 3336209 . PMID 21802439 .
- ^ Dorsam RT, Gutkind JS (февраль 2007 г.). «Рецепторы, связанные с G-белком и рак». Природные обзоры. Рак . 7 (2): 79–94. doi : 10.1038/nrc2069 . PMID 17251915 . S2CID 10996598 .
- ^ Venkatakrishnan AJ, Deupi X, Lebon G, Tate CG, Schertler GF, Babu MM (февраль 2013 г.). «Молекулярные подписи рецепторов, связанных с G-белком». Природа . 494 (7436): 185–94. Bibcode : 2013natur.494..185V . doi : 10.1038/nature11896 . PMID 23407534 . S2CID 4423750 .
- ^ Hollenstein K, De Graaf C, Bortolato A, Wang MW, Marshall FH, Stevens RC (январь 2014 г.). «Понимание структуры GPCR класса B» . Тенденции в фармакологических науках . 35 (1): 12–22. doi : 10.1016/j.tips.2013.11.001 . PMC 3931419 . PMID 24359917 .
- ^ Palczewski K, Kumasaka T, Hori T, Behnke CA, Motoshima H, Fox Ba, et al. (Август 2000). «Кристаллическая структура родопсина: рецептор, связанный с белком Ag». Наука . 289 (5480): 739–45. Bibcode : 2000sci ... 289..739p . Citeseerx 10.1.1.1012.2275 . doi : 10.1126/science.289.5480.739 . PMID 10926528 .
- ^ Расмуссен С.Г., Чой Х.Дж., Розенбаум Д.М., Кобилка Т.С., Тиан Ф.С., Эдвардс П.К. и др. (Ноябрь 2007 г.). «Кристаллическая структура человеческого адренергического G-белкового рецептора Beta2 Beta2». Природа . 450 (7168): 383–7. Bibcode : 2007natur.450..383r . doi : 10.1038/nature06325 . PMID 17952055 . S2CID 4407117 .
- ^ Розенбаум Д.М., Черезов В., Хансон М.А., Расмуссен С.Г., Тиан Ф.С., Кобилка Т.С. и др. (Ноябрь 2007 г.). «Инженерная инженерия GPCR дает структурную информацию о функции бета2-адренергического рецептора с высоким разрешением» . Наука . 318 (5854): 1266–73. Bibcode : 2007sci ... 318.1266R . doi : 10.1126/science.1150609 . PMID 17962519 . S2CID 1559802 .
- ^ Rasmussen SG, Choi HJ, Fung JJ, Pardon E, Casarosa P, Chae PS и др. (Январь 2011). «Структура нанободированного активного состояния β (2) адренорецептора» . Природа . 469 (7329): 175–80. Bibcode : 2011natur.469..175r . doi : 10.1038/nature09648 . PMC 3058308 . PMID 21228869 .
- ^ Розенбаум Д.М., Чжан С., Лион Д.А., Холл Р., Арагао Д., Арлоу Д.Х. и др. (Январь 2011). «Структура и функция необратимого агониста-β (2) комплекса адренорецепторов» . Природа . 469 (7329): 236–40. Bibcode : 2011natur.469..236r . doi : 10.1038/nature09665 . PMC 3074335 . PMID 21228876 .
- ^ Warne T, Moukhametzianov R, Baker JG, Nehmé R, Edwards PC, Leslie AG, et al. (Январь 2011). «Структурная основа для агонистского и частичного агонистского действия на β (1)-адренергический рецептор» . Природа . 469 (7329): 241–4. Bibcode : 2011natur.469..241W . doi : 10.1038/nature09746 . PMC 3023143 . PMID 21228877 .
- ^ Xu F, Wu H, Katritch V, Han GW, Jacobson KA, Gao ZG, et al. (Апрель 2011). «Структура агонистского аденозинового рецептора A2A-рецептора A2A» . Наука . 332 (6027): 322–7. Bibcode : 2011sci ... 332..322X . doi : 10.1126/science.1202793 . PMC 3086811 . PMID 21393508 .
- ^ Rasmussen SG, Devree BT, Zou Y, Kruse AC, Chung KY, Kobilka TS и др. (Июль 2011). «Кристаллическая структура белкового комплекса β2-адренергического рецептора-GS» . Природа . 477 (7366): 549–55. Bibcode : 2011natur.477..549r . doi : 10.1038/nature10361 . PMC 3184188 . PMID 21772288 .
- ^ Yauchi T, Kamon J, It Y, Tsuchida A, Yokomizo T, S, S, et al. (Июнь 2003 г.). «Клонирование рецепторов усыновления, что медиабетическая меаболическая эфенса» Природа 423 (6941): 762–9 Bibcode : 2003natur.423..762y Doi : 10.1038/ природа PMID 1280233 52860797S2CID
- ^ Подпрыгнуть до: а беременный Цинь К, Донг С., Ву Г, Ламберт На (август 2011 г.). «Неактивная предварительная символа G (Q)-связанных рецепторов и гетеротримеров G (Q)» . Природная химическая биология . 7 (10): 740–7. doi : 10.1038/nchembio.642 . PMC 3177959 . PMID 21873996 .
- ^ Lohse MJ, Benovic JL, Codina J, Caron MG, Lefkowitz RJ (июнь 1990 г.). «Бета-аррестин: белок, который регулирует функцию бета-адренергического рецептора». Наука . 248 (4962): 1547–50. Bibcode : 1990sci ... 248.1547L . doi : 10.1126/science.2163110 . PMID 2163110 .
- ^ Luttrell LM, Lefkowitz RJ (февраль 2002 г.). «Роль бета-аррестинов в прекращении и трансдукции сигналов рецепторов, связанных с G-белком». Журнал сотовой науки . 115 (Pt 3): 455–65. doi : 10.1242/jcs.115.3.455 . HDL : 10161/7805 . PMID 11861753 .
- ^ Cahill TJ, Thomsen AR, Tarrasch JT, Plouffe B, Nguyen AH, Yang F, et al. (Март 2017). «Отличительные конформации комплексов GPCR-β-аррестина опосредуют десенсибилизацию, передачу сигналов и эндоцитоз» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (10): 2562–2567. Bibcode : 2017pnas..114.2562c . doi : 10.1073/pnas.1701529114 . PMC 5347553 . PMID 28223524 .
- ^ Кумари П., Шривастава А., Банерджи Р., Гош Е., Гупта П., Ранджан Р. и др. (Ноябрь 2016). «Функциональная компетентность частично вовлеченного комплекса GPCR-β-Arrestin» . Природная связь . 7 : 13416. Bibcode : 2016natco ... 713416K . doi : 10.1038/ncomms13416 . PMC 5105198 . PMID 27827372 .
- ^ Томсен А.Р., Плуфф Б., Кэхилл Т.Дж., Шукла А.К., Тарраш Дж. Т., Дозей А.М. и др. (Август 2016 г.). «Super-Complex GPCR-G-белок-β-аррестин опосредует постоянную передачу сигналов G-белка» . Клетка . 166 (4): 907–919. doi : 10.1016/j.cell.2016.07.004 . PMC 5418658 . PMID 27499021 .
- ^ Нгуен А.Х., Кахилл Т.Дж. (Декабрь 2019). GPCR-G . Структура и молекулярный биологический 26 (12): 1123–1 doi : /s4154-030-030-0 10.1038 PMC 7108882 . PMID 31740855 .
- ^ Millar RP, Newton CL (январь 2010 г.). «Год в исследовании рецепторов, связанных с белком,» . Молекулярная эндокринология . 24 (1): 261–74. doi : 10.1210/me.2009-0473 . PMC 5428143 . PMID 20019124 .
- ^ Латунный LF (сентябрь 2003 г.). «Активация тромбина и тромбоцитов». Грудь . 124 (3 Suppl): 18S - 25 с. doi : 10.1378/грудь.124.3_suppl.18s . PMID 12970120 . S2CID 22279536 .
- ^ Digby GJ, Lober RM, Sethi PR, Lambert NA (ноябрь 2006 г.). «Некоторые гетеротримеры G -белка физически диссоциации в живых клетках» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (47): 17789–94. Bibcode : 2006pnas..10317789d . doi : 10.1073/pnas.0607116103 . PMC 1693825 . PMID 17095603 .
- ^ Рубенштейн Л.А., Лансара Р.Г. (1998). «Активация G-белковых рецепторов влечет за собой цистеиновую модуляцию связывания агониста» . Журнал молекулярной структуры: Теохем . 430 : 57–71. doi : 10.1016/s0166-1280 (98) 90217-2 . Архивировано из оригинала 16 мая 2011 года . Получено 14 января 2006 года .
- ^ Teoh CM, Tam JK, Tran T (2012). «Интегран и GPCR перекрестные помехи в регуляции передачи сигналов сокращения ASM при астме» . Журнал аллергии . 2012 : 341282. DOI : 10.1155/2012/341282 . PMC 3465959 . PMID 23056062 .
- ^ Смит Дж.С., Николсон Л.Т., Суванпрадид Дж., Гленн Р.А., Клап Н.М., Алагельс П. и др. (Ноябрь 2018). «Предвзятые агонисты рецептора хемокинов CXCR3 по -разному контролируют хемотаксис и воспаление» . Наука сигнализация . 11 (555): EAAQ1075. doi : 10.1126/scisignal.aaq1075 . PMC 6329291 . PMID 30401786 .
- ^ Ким Дж.Ю., Хаастерт П.В., Девреотс П.Н. (апрель 1996 г.). «Социальные чувства: сигнальные пути, связанные с G-белком рецептора, при диктиостелиевом дикодеуме» . Химия и биология . 3 (4): 239–43. doi : 10.1016/s1074-5521 (96) 90103-9 . PMID 8807851 .
- ^ Duchene J, Schanstra JP, Pecher C, Pizard A, Susini C, Esteve JP, et al. (Октябрь 2002). «Новое взаимодействие белкового белка между G-белковым рецептором и фосфатазой Shp-2 участвует в индуцированном брадикинином ингибированию пролиферации клеток» . Журнал биологической химии . 277 (43): 40375–83. doi : 10.1074/jbc.m202744200 . PMID 12177051 .
- ^ Chen-Izu Y, Xiao RP, Izu LT, Cheng H, Kuschel M, Spurgeon H, Lakatta EG (ноябрь 2000 г.). «G (i) -зависимая локализация передачи сигналов бета (2)-адренергического рецептора на каналы Ca (2+) L-типа» . Биофизический журнал . 79 (5): 2547–56. Bibcode : 2000bpj .... 79.2547c . doi : 10.1016/s0006-3495 (00) 76495-2 . PMC 1301137 . PMID 11053129 .
- ^ Подпрыгнуть до: а беременный Tan CM, Brady AE, Nickols HH, Wang Q, Limbird LE (2004). «Мембранная перенос G-белковых рецепторов». Ежегодный обзор фармакологии и токсикологии . 44 (1): 559–609. doi : 10.1146/annurev.pharmtox.44.101802.121558 . PMID 14744258 .
- ^ Santulli G, Trimarco B, Iaccarino G (март 2013 г.). «Рецепторная киназа, связанная с G-белком и гипертония: молекулярная информация и патофизиологические механизмы». Высокое кровяное давление и сердечно -сосудистая профилактика . 20 (1): 5–12. doi : 10.1007/s40292-013-0001-8 . PMID 23532739 . S2CID 45674941 .
- ^ Пенела П., Рибас С, мэр Ф (ноябрь 2003 г.). Механизмы правил. Клеточная передача сигналов . 15 (11) (11): 973–8 doi : /s 10.1016 PMID 1499340 .
- ^ Krueger KM, Daaka Y, Pitcher JA, Lefkowitz RJ (январь 1997 г.). «Роль секвестрации в рецептор-рецепторе, связанной с G-белком, регуляция дефосфорилирования бета2-адренергического рецептора путем подкисления» . Журнал биологической химии . 272 (1): 5–8. doi : 10.1074/jbc.272.1.5 . PMID 8995214 .
- ^ Laporte SA, Oakley RH, Holt JA, Barak LS, Caron MG (июль 2000 г.). «Взаимодействие бета-аррестана с адаптером AP-2 необходимо для кластеризации бета-2-адренергического рецептора в ямы, покрытые клатрином» . Журнал биологической химии . 275 (30): 23120–6. doi : 10.1074/jbc.m002581200 . PMID 10770944 .
- ^ Laporte SA, Oakley RH, Zhang J, Holt JA, Ferguson SS, Caron MG, Barak LS (март 1999 г.). «Комплекс бета2-адренергического рецептора/бетааррестана рекрутирует адаптер клатрина AP-2 во время эндоцитоза» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (7): 3712–7. Bibcode : 1999pnas ... 96.3712L . doi : 10.1073/pnas.96.7.3712 . PMC 22359 . PMID 10097102 .
- ^ Тобин А.Б. (март 2008 г.). «Фосфорилирование рецептора с G-белком, где, где, когда и кем» . Британский журнал фармакологии . 153 (Suppl 1): S167–76. doi : 10.1038/sj.bjp.0707662 . PMC 2268057 . PMID 18193069 .
- ^ Margeta-Mitrovic M, Jan Yn, Jan Ly (июль 2000 г.). «Контрольная точка от торговли контролирует гетеродимеризация рецептора GABA (B)» . Нейрон . 27 (1): 97–106. doi : 10.1016/s0896-6273 (00) 00012-x . PMID 10939334 . S2CID 15430860 .
- ^ White JH, Wise A, Main MJ, Green A, Fraser NJ, Disney GH, et al. (Декабрь 1998). «Гетеродимеризация требуется для формирования функционального рецептора ГАМК (b)». Природа . 396 (6712): 679–82. Bibcode : 1998natur.396..679W . doi : 10.1038/25354 . PMID 9872316 . S2CID 4406311 .
- ^ Подпрыгнуть до: а беременный Nordström KJ, Sällman Almén M, Edstam MM, Fredriksson R, Schiöth HB (сентябрь 2011 г.). «Независимые HHSearch, Geelleman-анализ на основе Wunsch и мотивы показывают общую иерархию для большинства семейств рецепторов, связанных с G». Молекулярная биология и эволюция . 28 (9): 2471–80. doi : 10.1093/molbev/msr061 . PMID 21402729 .
Дальнейшее чтение
[ редактировать ]- Vassilatis DK, Hohmann JG, Zeng H, Li F, Ranchalis JE, Mortrud MT, et al. (Апрель 2003 г.). «G-белковые рецепторные репертуары человека и мыши» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (8): 4903–8. Bibcode : 2003pnas..100.4903V . doi : 10.1073/pnas.0230374100 . PMC 153653 . PMID 12679517 .
- «Справочная библиотека GPCR» . Получено 11 августа 2008 года .
Ссылка на молекулярные и математические модели для начального ответа рецептора
- «Нобелевская премия по химии 2012» (PDF) . Архивировано (PDF) из оригинала 18 октября 2012 года . Получено 10 октября 2012 года .
Внешние ссылки
[ редактировать ]- G-белок-связанные+рецепторы в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)
- Линия ячейки GPCR Архивирована 3 апреля 2015 года на машине Wayback
- «Руководство IUPHAR/BPS по базе данных фармакологии (GPCR)» . База данных Iuphar . Эдинбургский университет / Международный союз базовой и клинической фармакологии . Получено 6 февраля 2019 года .
- "GPCRDB" .
Данные, диаграммы и веб-инструменты для G-белковых рецепторов (GPCR).
; Munk C. (Июль 2016 г.). GPCRDB: введение » Британский журнал . 173 (14): 2195–2 doi : 10.111/bph.13509 . PMC 4919580 . PMID 27159948 . - «G-белковые рецепторы в сети» . Архивировано из оригинала 23 июля 2011 года . Получено 10 ноября 2010 года .
классификация GPCR
- «Сетевой центр PSI GPCR» . Архивировано из оригинала 25 июля 2013 года . Получено 11 июля 2013 года .
Инициатива по структуре белка: Биологический сетевой центр, направленный на определение трехмерных структур репрезентативных белков семейства GPCR
- GPCR-HGMOD Архивировал 1 февраля 2016 года в The Wayback Machine , базе данных 3D-структурных моделей всех рецепторов, связанных с G-белком человека, построенной GPCR- i-Tasser конвейером Zhang J, Yang J, Jang R, Zhang Y (август 2015 г.). «GPCR-I-Tasser: гибридный подход к моделированию структуры рецептора, связанной с G-белком, и применение к геному человека» . Структура 23 (8): 1538–1549. doi : 10.1016/j.str.2015.06.007 . PMC 4526412 . PMID 26190572 .