Катенин бета-1
| CTNNB1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CTNNB1 , CTNNB, MRD19, броненосец, катенин бета 1, EVR7, NEDSDV | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 116806 ; МГИ : 88276 ; Гомологен : 1434 ; Генные карты : CTNNB1 ; OMA : CTNNB1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Катенин бета-1 , также известный как β-катенин ( бета -катенин), представляет собой белок , который у человека кодируется CTNNB1 геном .
β-катенин представляет собой белок с двойной функцией , участвующий в регуляции и координации межклеточной адгезии и транскрипции генов . У человека белок CTNNB1 кодируется CTNNB1 геном . [5] [6] У дрозофилы гомологичный белок называется броненосцем . β-катенин является субъединицей белкового комплекса кадгерина и действует как внутриклеточный преобразователь сигналов в сигнальном пути Wnt . [7] [8] [9] Он является членом семейства белков катенина и гомологичен γ-катенину , также известному как плакоглобин . β-катенин широко экспрессируется во многих тканях. В сердечной мышце β-катенин локализуется в слипчивых соединениях в вставочных структурах диска , которые имеют решающее значение для электрического и механического соединения между соседними кардиомиоцитами .
Мутации и сверхэкспрессия β-катенина связаны со многими видами рака, включая гепатоцеллюлярную карциному , колоректальную карциному , рак легких , злокачественные опухоли молочной железы , яичников и рак эндометрия . [10] Изменения локализации и уровня экспрессии β-катенина связаны с различными формами заболеваний сердца , включая дилатационную кардиомиопатию . β-катенин регулируется и разрушается комплексом разрушения бета-катенина , в частности белком аденоматозного полипоза coli , подавляющим опухоль APC (APC), кодируемым геном . Следовательно, генетическая мутация гена APC также тесно связана с раком, в частности с колоректальным раком, возникающим в результате семейного аденоматозного полипоза (FAP).
Открытие
[ редактировать ]β-катенин был первоначально обнаружен в начале 1990-х годов как компонент адгезионного комплекса клеток млекопитающих : белок, ответственный за цитоплазматическое закрепление кадгеринов . [11] Но очень скоро стало понятно, что белок броненосца дрозофилы , участвующий в обеспечении морфогенных эффектов Wingless/Wnt , гомологичен β-катенину млекопитающих не только по структуре, но и по функциям. [12] Таким образом, β-катенин стал одним из первых примеров подработки : белок, выполняющий более чем одну радикально отличающуюся клеточную функцию.
Структура
[ редактировать ]Структура белка
[ редактировать ]Ядро β-катенина состоит из нескольких очень характерных повторов , каждый длиной примерно 40 аминокислот. Названные повторами броненосца , все эти элементы складываются вместе в один жесткий белковый домен вытянутой формы, называемый доменом броненосца (ARM). Средний повтор броненосца состоит из трех альфа-спиралей . Первый повтор β-катенина (около N-конца) немного отличается от остальных – имеет удлиненную спираль с изломом, образованным слиянием спиралей 1 и 2. [13] Из-за сложной формы отдельных повторов весь ARM-домен не представляет собой прямой стержень: он обладает небольшой кривизной, в результате чего образуется внешняя (выпуклая) и внутренняя (вогнутая) поверхности. Эта внутренняя поверхность служит местом связывания лигандов для различных партнеров по взаимодействию доменов ARM.

Сегменты, N-концевые и дальние C-концевые по отношению к домену ARM, сами по себе не принимают никакой структуры в растворе. Тем не менее, эти внутренне неупорядоченные области играют решающую роль в функции β-катенина. N-концевая неупорядоченная область содержит консервативный короткий линейный мотив, ответственный за связывание TrCP1 (также известной как β-TrCP) убиквитинлигазы E3 – но только тогда, когда она фосфорилирована . Таким образом, деградация β-катенина опосредована этим N-концевым сегментом. С-концевая область, с другой стороны, является сильным трансактиватором при рекрутировании на ДНК . Этот сегмент не полностью неупорядочен: часть C-концевого расширения образует стабильную спираль , которая упаковывается в домен ARM, но может также взаимодействовать с отдельными партнерами по связыванию. [14] Этот небольшой структурный элемент (HelixC) покрывает С-конец домена ARM, защищая его гидрофобные остатки. HelixC не необходим для функционирования β-катенина в межклеточной адгезии. С другой стороны, он необходим для передачи сигналов Wnt: возможно, для рекрутирования различных коактиваторов, таких как 14-3-3zeta. [15] Тем не менее, его точные партнеры среди общих транскрипционных комплексов все еще не полностью изучены, и они, вероятно, связаны с тканеспецифичными игроками. [16] Примечательно, что С-концевой сегмент β-катенина может имитировать эффекты всего пути Wnt, если его искусственно слить с ДНК-связывающим доменом транскрипционного фактора LEF1 . [17]
Плакоглобин (также называемый γ-катенином) имеет поразительно схожую архитектуру с β-катенином. Не только их ARM-домены сходны друг с другом как по архитектуре, так и по способности связывания лигандов, но N-концевой мотив связывания β-TrCP также консервативен в плакоглобине, что подразумевает общее происхождение и общую регуляцию с β-катенином. [18] Однако плакоглобин является очень слабым трансактиватором при связывании с ДНК – это, вероятно, вызвано расхождением их С-концевых последовательностей (по-видимому, плакоглобин лишен мотивов трансактиватора и, таким образом, ингибирует гены-мишени пути Wnt , а не активирует их). [19]
Привязка партнеров к домену броненосца
[ редактировать ]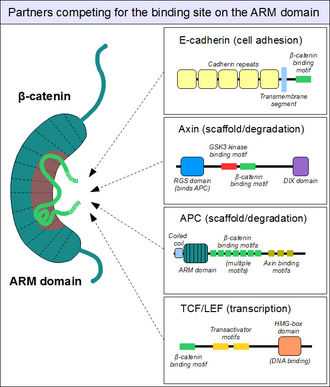
Как показано выше, ARM-домен β-катенина действует как платформа, с которой могут связываться специфические линейные мотивы . Расположенные у структурно различных партнеров, мотивы связывания β-катенина обычно неупорядочены сами по себе и обычно принимают жесткую структуру при взаимодействии с доменом ARM – как это видно для коротких линейных мотивов . Однако мотивы, взаимодействующие с β-катенином, также обладают рядом особенностей. Во-первых, они могут достигать или даже превосходить длину в 30 аминокислот и контактировать с доменом ARM на чрезмерно большой площади поверхности. Другой необычной особенностью этих мотивов является часто высокая степень фосфорилирования . Такие события фосфорилирования Ser / Thr значительно усиливают связывание многих мотивов, связывающих β-катенин, с доменом ARM. [20]
Структура β-катенина в комплексе с катенин-связывающим доменом партнера транскрипционной трансактивации TCF предоставила первоначальную структурную карту того, сколько партнеров по связыванию β-катенина могут образовывать взаимодействия. [21] Эта структура продемонстрировала, как в остальном неупорядоченный N-конец TCF адаптировал то, что казалось жесткой конформацией, с мотивом связывания, охватывающим множество повторов бета-катенина. Были определены «горячие точки» относительно сильного заряженного взаимодействия (предсказано, а затем подтверждено, что они сохраняются для взаимодействия β-катенин/Е-кадгерин), а также гидрофобные области, которые считаются важными для общего способа связывания и как потенциальные терапевтические малые Молекулярный ингибитор нацелен на определенные формы рака. Более того, последующие исследования продемонстрировали еще одну особенность — пластичность связывания N-конца TCF с бета-катенином. [22] [23]
Аналогичным образом мы находим знакомый E-кадгерин , цитоплазматический хвост которого контактирует с доменом ARM таким же каноническим образом. [24] Каркасный белок аксин (два близкородственных паралога, аксин 1 и аксин 2 ) содержит сходный мотив взаимодействия на своем длинном, неупорядоченном среднем сегменте. [25] Хотя одна молекула аксина содержит только один мотив рекрутирования β-катенина, его партнер, белок аденоматозного полипоза coli (APC), содержит 11 таких мотивов в тандемном расположении на протомер, таким образом, он способен взаимодействовать с несколькими молекулами β-катенина одновременно. [26] Поскольку на поверхности домена ARM обычно может разместиться только один пептидный мотив в любой момент времени, все эти белки конкурируют за один и тот же клеточный пул молекул β-катенина. Эта конкуренция является ключом к пониманию того, как работает сигнальный путь Wnt .
Однако этот «основной» сайт связывания β-катенина домена ARM далеко не единственный. Первые спирали домена ARM образуют дополнительный специальный карман межбелкового взаимодействия: он может вмещать образующий спираль линейный мотив, обнаруженный в коактиваторе BCL9 (или близкородственном BCL9L ) – важном белке, участвующем в передаче сигналов Wnt. [27] Хотя точные детали гораздо менее ясны, похоже, что тот же сайт используется альфа-катенином, когда β-катенин локализуется в слипчивых соединениях. [28] Поскольку этот карман отличается от «основного» сайта связывания домена ARM, нет конкуренции между альфа-катенином и E-кадгерином или между TCF1 и BCL9 соответственно. [29] С другой стороны, BCL9 и BCL9L должны конкурировать с α-катенином за доступ к молекулам β-катенина. [30]
Функция
[ редактировать ]Регулирование деградации посредством фосфорилирования
[ редактировать ]Клеточный уровень β-катенина в основном контролируется его убиквитинированием и протеосомной деградацией . Убиквитинлигаза Е3 TrCP1 (также известная как β-TrCP) может распознавать β-катенин в качестве своего субстрата по короткому линейному мотиву на неупорядоченном N-конце. Однако этот мотив (Asp-Ser-Gly-Ile-His-Ser) β-катенина должен быть фосфорилирован по двум серинам , чтобы быть способным связывать β-TrCP. Фосфорилирование мотива осуществляется киназой гликогенсинтазы 3 альфа и бета (GSK3α и GSK3β). GSK3 представляют собой конститутивно активные ферменты, участвующие в нескольких важных регуляторных процессах. Однако есть одно требование: субстраты GSK3 должны быть предварительно фосфорилированы на четыре аминокислоты ниже (С-концево) от фактического сайта-мишени. Таким образом, для его деятельности также требуется «праймирующая киназа». В случае β-катенина наиболее важной праймирующей киназой является казеинкиназа I (CKI). Как только субстрат, богатый серином и треонином, «загрунтован», GSK3 может «пройти» по нему от C-концевого к N-концевому направлению, фосфорилируя каждый четвертый серин или остатки треонина в ряд. Этот процесс также приведет к двойному фосфорилированию вышеупомянутого мотива узнавания β-TrCP.
Комплекс разрушения бета-катенина
[ редактировать ]Чтобы GSK3 была высокоэффективной киназой на субстрате, предварительного фосфорилирования недостаточно. Существует еще одно дополнительное требование: подобно митоген-активируемым протеинкиназам (MAPK), субстраты должны связываться с этим ферментом посредством мотивов стыковки с высоким сродством . β-катенин не содержит таких мотивов, но есть в специальном белке: аксине . Более того, его мотив стыковки GSK3 непосредственно примыкает к мотиву связывания β-катенина. [25] Таким образом, аксин действует как настоящий каркасный белок , приводя фермент (GSK3) вместе с его субстратом (β-катенином) в непосредственную физическую близость.

Но даже аксин не действует в одиночку. Через свой N-концевой регулятор сигнального домена G-белка (RGS) он рекрутирует белок аденоматозного полипоза coli (APC). APC похож на огромную «рождественскую елку»: со множеством мотивов связывания β-катенина (одна только молекула APC обладает 11 такими мотивами). [26] ), он может собрать как можно больше молекул β-катенина. [31] APC может взаимодействовать с несколькими молекулами аксина одновременно, поскольку он имеет три мотива SAMP (Ser-Ala-Met-Pro) для связывания доменов RGS , обнаруженных в аксине . Кроме того, аксин также обладает потенциалом к олигомеризации через свой С-концевой домен DIX. В результате образуется огромная мультимерная белковая сборка, предназначенная для фосфорилирования β-катенина. Этот комплекс обычно называют комплексом разрушения бета-катенина , хотя он отличается от протеосомного механизма, фактически ответственного за деградацию β-катенина. [32] Он лишь маркирует молекулы β-катенина для последующего разрушения.
Передача сигналов Wnt и регуляция разрушения
[ редактировать ]В покоящихся клетках молекулы аксина олигомеризуются друг с другом через свои С-концевые домены DIX, которые имеют два интерфейса связывания. Таким образом, они могут строить линейные олигомеры или даже полимеры внутри цитоплазмы клеток. Домены DIX уникальны: единственные другие белки, которые, как известно, имеют домен DIX, — это Disheveled и DIXDC1 . (Один Dsh белок дрозофилы соответствует трем паралогичным генам: Dvl1 , Dvl2 и Dvl3 у млекопитающих .) Dsh ассоциируется с цитоплазматическими областями рецепторов Frizzled с помощью своих PDZ и DEP доменов . Когда молекула Wnt связывается с Frizzled , она индуцирует малоизвестный каскад событий, которые приводят к обнажению домена DIX растрепанного и созданию идеального сайта связывания для axin . Затем аксин титруется от его олигомерных сборок – комплекса разрушения β-катенина – с помощью Dsh . [33] После связывания с рецепторным комплексом аксин становится некомпетентным для связывания β-катенина и активности GSK3. Важно отметить, что цитоплазматические сегменты ассоциированных с Frizzled белков LRP5 и LRP6 содержат псевдосубстратные последовательности GSK3 (Pro-Pro-Pro-Ser-Pro-x-Ser), соответствующим образом «примированные» (предварительно фосфорилированные) CKI , как если бы это была настоящая подложка GSK3. Эти ложные сайты-мишени значительно конкурентно ингибируют активность GSK3. [34] Таким образом, связанный с рецептором аксин отменит фосфорилирование β-катенина. Поскольку β-катенин больше не предназначен для разрушения, но продолжает вырабатываться, его концентрация будет увеличиваться. Как только уровень β-катенина поднимется достаточно высоко, чтобы насытить все места связывания в цитоплазме, он также переместится в ядро. При задействовании факторов транскрипции LEF1 , TCF1 , TCF2 или TCF3 β-катенин заставляет их отключить своих предыдущих партнеров: белки Groucho. В отличие от Граучо , который рекрутирует репрессоры транскрипции (например, гистон-лизин-метилтрансферазы ), β-катенин связывает активаторы транскрипции , включая гены-мишени.
Роль в межклеточной адгезии
[ редактировать ]
Комплексы межклеточной адгезии необходимы для формирования сложных тканей животных. β-катенин является частью белкового комплекса , образующего адгезионные соединения . [35] Эти комплексы межклеточной адгезии необходимы для создания и поддержания слоев и барьеров эпителиальных клеток . В составе комплекса β-катенин может регулировать рост клеток и адгезию между клетками. Он также может отвечать за передачу сигнала контактного ингибирования, который заставляет клетки прекращать деление после завершения формирования эпителиального листа. [36] Комплекс Е-кадгерин – β-катенин – α-катенин слабо связан с актиновыми нитями . Адгерентные соединения требуют значительной динамики белка для связи с актиновым цитоскелетом. [35] тем самым обеспечивая механотрансдукцию . [37] [38]
Важным компонентом слипчивых соединений являются белки кадгерины. Кадгерины образуют структуры межклеточных соединений, известные как слипчивые соединения, а также десмосомы . Кадгерины способны к гомофильным взаимодействиям через свои внеклеточные домены повторов кадгерина Ca2+-зависимым образом; это может удерживать вместе соседние эпителиальные клетки. Находясь в адгезионном соединении, кадгерины рекрутируют молекулы β-катенина в свои внутриклеточные области. [ нужны разъяснения ] . β-катенин, в свою очередь, связывается с другим высокодинамичным белком , α-катенином , который напрямую связывается с актиновыми нитями. [39] Это возможно, поскольку α-катенин и кадгерины связываются с β-катенином в разных местах. [40] Таким образом, комплекс β-катенин-α-катенин может физически образовывать мост между кадгеринами и актиновым цитоскелетом . [41] Организация кадгерин-катенинового комплекса дополнительно регулируется посредством фосфорилирования и эндоцитоза его компонентов. [ нужна ссылка ]
Роли в разработке
[ редактировать ]β-катенин играет центральную роль в управлении несколькими процессами развития, поскольку он может напрямую связывать факторы транскрипции и регулироваться диффундирующим внеклеточным веществом: Wnt. Он действует на ранние эмбрионы, индуцируя целые области тела, а также отдельные клетки на более поздних стадиях развития. Он также регулирует процессы физиологической регенерации.
Раннее эмбриональное формирование рисунка
[ редактировать ]Передача сигналов Wnt и β-catenin-зависимая экспрессия генов играют решающую роль во время формирования различных областей тела у ранних эмбрионов. Экспериментально модифицированные эмбрионы, которые не экспрессируют этот белок, не смогут развить мезодерму и инициировать гаструляцию . [42] Спецификация эндомезодермы ранних эмбрионов также включает активацию β-catenin-зависимой транскрипционной активности посредством первых морфогенетических движений эмбриогенеза посредством процессов механотрансдукции. Эту особенность разделяют билатерии позвоночных и членистоногих, а также книдарии, и было высказано предположение, что она была эволюционно унаследована от ее возможного участия в энтомезодермальной спецификации первых многоклеточных животных. [43] [44] [45]
На стадиях бластулы и гаструлы Wnt , а также пути BMP и FGF будут индуцировать формирование передне-задней оси, регулировать точное расположение первичной полоски (гаструляция и образование мезодермы), а также процесс нейруляции (развитие центральной нервной системы). ). [46]
В ооцитах Xenopus β-катенин изначально одинаково локализован во всех областях яйцеклетки, но он подвергается убиквитинированию и деградации с помощью комплекса разрушения β-катенина. Оплодотворение яйцеклетки вызывает вращение внешних кортикальных слоев, перемещая кластеры белков Frizzled и Dsh ближе к экваториальной области. β-катенин будет локально обогащаться под влиянием сигнального пути Wnt в клетках, которые наследуют эту часть цитоплазмы. В конечном итоге он переместится в ядро, чтобы связать TCF3 и активировать несколько генов, которые индуцируют характеристики дорсальных клеток. [47] Эта передача сигналов приводит к образованию области клеток, известной как серый полумесяц, которая является классическим организатором эмбрионального развития. Если этот участок хирургически удалить из эмбриона, гаструляция вообще не происходит. β-катенин также играет решающую роль в индукции губы бластопора , которая, в свою очередь, инициирует гаструляцию. [48] Ингибирование трансляции GSK-3 путем инъекции антисмысловой мРНК может привести к образованию второго бластопора и лишней оси тела. Подобный эффект может возникнуть в результате сверхэкспрессии β-катенина. [49]
Асимметричное деление клеток
[ редактировать ]β-катенин также участвует в регуляции судеб клеток посредством асимметричного деления клеток в модельном организме C. elegans . Подобно ооцитам Xenopus , это по существу является результатом неравномерного распределения Dsh , Frizzled , аксина и АПК в цитоплазме материнской клетки. [50]
Обновление стволовых клеток
[ редактировать ]Одним из наиболее важных результатов передачи сигналов Wnt и повышенного уровня β-катенина в определенных типах клеток является поддержание плюрипотентности . [46] Скорость образования стволовых клеток в толстой кишке, например, обеспечивается таким накоплением β-катенина, которое может стимулироваться путем Wnt. [51] Высокочастотные перистальтические механические напряжения толстой кишки также участвуют в β-катенин-зависимом поддержании гомеостатического уровня стволовых клеток толстой кишки посредством процессов механотрансдукции. Эта особенность патологически усиливается в сторону туморогенной гиперпролиферации в здоровых клетках, сжимаемых давлением генетически измененных гиперпролиферативных опухолевых клеток. [52]
В других типах клеток и на стадиях развития β-катенин может способствовать дифференцировке , особенно в сторону мезодермальных клеточных линий.
Эпителиально-мезенхимальный переход
[ редактировать ]β-Катенин также действует как морфоген на более поздних стадиях эмбрионального развития. Вместе с TGF-β важная роль β-катенина заключается в индукции морфогенных изменений в эпителиальных клетках. Это побуждает их отказаться от плотной адгезии и принять более подвижный и слабо ассоциированный мезенхимальный фенотип. Во время этого процесса эпителиальные клетки теряют экспрессию таких белков, как E-кадгерин , Zonula occludens 1 (ZO1) и цитокератин . В то же время они включают экспрессию виментина , альфа-актина гладких мышц (ACTA2) и фибробласт-специфического белка 1 (FSP1). Они также производят компоненты внеклеточного матрикса, такие как коллаген I типа и фибронектин . Аберрантная активация пути Wnt вовлечена в патологические процессы, такие как фиброз и рак. [53] В развитии сердечной мышцы β-катенин выполняет двухфазную роль. Первоначально активация Wnt/β-катенина необходима для коммитирования мезенхимальных клеток в кардиальную линию; однако на более поздних стадиях развития требуется подавление β-катенина. [54] [55] [42]
Участие в физиологии сердца
[ редактировать ]В сердечной мышце β-катенин образует комплекс с N-кадгерином в слипчивых соединениях внутри вставочных структур диска , которые отвечают за электрическое и механическое соединение соседних сердечных клеток. Исследования на модели желудочковых кардиомиоцитов взрослых крыс показали, что появление и распределение β-катенина регулируется пространственно-временно в ходе редифференцировки этих клеток в культуре. В частности, β-катенин является частью отдельного комплекса с N-кадгерином и альфа-катенином , которого много в слипчивых соединениях на ранних стадиях после изоляции кардиомиоцитов для восстановления межклеточных контактов. [56] Показано, что β-катенин образует комплекс с эмерином в кардиомиоцитах в местах слипчивых соединений внутри вставочных дисков; и это взаимодействие зависит от присутствия сайтов 3-бета- фосфорилирования GSK на β-катенине. Нокаут эмерина значительно изменил локализацию β-катенина и общую архитектуру вставочного диска, что напоминало фенотип дилатационной кардиомиопатии . [57]
На животных моделях сердечно-сосудистых заболеваний были раскрыты функции β-катенина. На модели аортального стеноза левого желудочка и гипертрофии на морских свинках было показано, что β-катенин изменяет субклеточную локализацию с вставочных дисков на цитозоль , несмотря на отсутствие изменений в общем клеточном содержании β-катенина. винкулин показал аналогичный профиль изменений. N-кадгерин не выявил изменений, и не наблюдалось компенсаторного повышения уровня плакоглобина в вставочных дисках в отсутствие β-катенина. [58] В модели кардиомиопатии и сердечной недостаточности у хомяков межклеточные спайки были нерегулярными и дезорганизованными, а уровни экспрессии слипчивых соединений/вставочных дисков и ядерных пулов β-катенина были снижены. [59] Эти данные позволяют предположить, что потеря β-катенина может играть роль в заболевании вставочных дисков, которое связано с гипертрофией сердечной мышцы и сердечной недостаточностью. В крысиной модели миокарда инфаркта аденовирусный перенос гена нефосфорилируемого конститутивно активного β-катенина уменьшал размер ИМ, активировал клеточный цикл и уменьшал количество апоптоза в кардиомиоцитах и сердечных миофибробластах . Это открытие было согласовано с усиленной экспрессией белков, способствующих выживанию, сурвивина и Bcl-2 , а также фактора роста эндотелия сосудов, одновременно способствуя дифференцировке сердечных фибробластов в миофибробласты. Эти данные свидетельствуют о том, что β-катенин может способствовать процессу регенерации и заживления после инфаркта миокарда. [60] На модели крыс со спонтанной гипертонической сердечной недостаточностью исследователи обнаружили перемещение β-катенина из вставочного диска/ сарколеммы в ядро , о чем свидетельствует снижение экспрессии β-катенина во фракции мембранных белков и увеличение ядерной фракции. Кроме того, они обнаружили ослабление связи между киназой гликогенсинтазы-3β и β-катенином, что может указывать на изменение стабильности белка. В целом результаты показывают, что усиленная ядерная локализация β-катенина может иметь важное значение для прогрессирования гипертрофии сердца . [61]
Что касается механистической роли β-катенина в сердечной гипертрофии, исследования на трансгенных мышах показали несколько противоречивые результаты относительно того, является ли активация β-катенина полезной или вредной. [62] [63] [64] Недавнее исследование с использованием условно нокаутных мышей, у которых либо вообще отсутствовал β-катенин, либо экспрессировалась неразлагаемая форма β-катенина в кардиомиоцитах, выявило потенциальную причину этих несоответствий. По-видимому, существует строгий контроль над субклеточной локализацией β-катенина в сердечной мышце. левого желудочка Мыши, лишенные β-катенина, не имели явного фенотипа в миокарде ; однако у мышей, несущих стабилизированную форму β-катенина, развилась дилатационная кардиомиопатия , что позволяет предположить, что временная регуляция β-катенина посредством механизмов деградации белка имеет решающее значение для нормального функционирования β-катенина в сердечных клетках. [65] В мышиной модели с нокаутом десмосомального белка плакоглобина, вовлеченного в аритмогенную кардиомиопатию правого желудочка , стабилизация β-катенина также была усилена, предположительно, чтобы компенсировать потерю его гомолога плакоглобина. Эти изменения были согласованы с активацией Akt и ингибированием киназы гликогенсинтазы 3β , что еще раз указывает на то, что аномальная стабилизация β-катенина может быть вовлечена в развитие кардиомиопатии. [66] Дальнейшие исследования с использованием двойного нокаута плакоглобина и β-катенина показали, что двойной нокаут приводит к развитию кардиомиопатии, фиброза и аритмий, что приводит к внезапной сердечной смерти . Архитектура вставочного диска была серьезно нарушена, а коннексина 43 были щелевые соединения заметно уменьшены. Измерения электрокардиограммы выявили спонтанные летальные желудочковые аритмии у двойных трансгенных животных, что позволяет предположить, что два катенина — β-катенин и плакоглобин — имеют решающее значение и незаменимы для механоэлектрического взаимодействия в кардиомиоцитах. [67]
Клиническое значение
[ редактировать ]Роль в депрессии
[ редактировать ]может ли мозг конкретного человека эффективно справляться со стрессом и, следовательно, его восприимчивость к депрессии, зависит от β-катенина в мозгу каждого человека. Согласно исследованию, проведенному в Медицинской школе Икан на горе Синай и опубликованному 12 ноября, 2014, в журнале Nature . [68] Более высокая передача сигналов β-катенина увеличивает поведенческую гибкость, тогда как дефектная передача сигналов β-катенина приводит к депрессии и снижению способности справляться со стрессом. [68]
Роль в сердечных заболеваниях
[ редактировать ]Измененные профили экспрессии β-катенина связаны с дилатационной кардиомиопатией у людей. Повышение экспрессии β-катенина обычно наблюдается у пациентов с дилатационной кардиомиопатией. [69] В конкретном исследовании у пациентов с конечной стадией дилатационной кардиомиопатии наблюдались почти удвоенные рецептора эстрогена альфа (ER-альфа) уровни мРНК и белка , а также взаимодействие ER-альфа/бета-катенин, присутствующее на вставочных дисках контрольного, здорового человека. сердца было потеряно, что позволяет предположить, что потеря этого взаимодействия на вставочном диске может играть роль в прогрессировании сердечной недостаточности. [70] Вместе с белками BCL9 и PYGO β-катенин координирует различные аспекты развития слуха, а мутации в Bcl9 или Pygo в модельных организмах, таких как мыши и рыбки данио, вызывают фенотипы, которые очень похожи на врожденные пороки сердца человека . [71]
Участие в раке
[ редактировать ]
β-катенин является протоонкогеном . Мутации этого гена обычно обнаруживаются при различных видах рака: первичном гепатоцеллюлярном раке , колоректальном раке , раке яичников , раке молочной железы , раке легких и глиобластоме . Было подсчитано, что примерно 10% всех секвенированных образцов тканей всех видов рака обнаруживают мутации в гене CTNNB1. [72] Большинство этих мутаций группируются на крошечной области N-концевого сегмента β-катенина: мотиве связывания β-TrCP. Мутации с потерей функции этого мотива по существу делают невозможным убиквитинилирование и деградацию β-катенина. Это заставит β-катенин перемещаться в ядро без какого-либо внешнего стимула и непрерывно управлять транскрипцией его генов-мишеней. Повышенные уровни ядерного β-катенина также были отмечены при базальноклеточной карциноме (БКК). [73] плоскоклеточный рак головы и шеи (HNSCC), рак предстательной железы (CaP), [74] пиломатриксома (ПТР) [75] и медуллобластома (МДБ) [76] Эти наблюдения могут указывать или не указывать на мутацию в гене β-catenin: другие компоненты пути Wnt также могут быть дефектными.



Подобные мутации также часто наблюдаются в мотивах рекрутирования β-катенина APC . Наследственные мутации APC с потерей функции вызывают состояние, известное как семейный аденоматозный полипоз . появляются сотни полипов У пораженных людей в толстой кишке они могут трансформироваться в смертельный рак . Большинство этих полипов доброкачественны по своей природе, но со временем . Соматические мутации АПК при колоректальном раке также нередки. [80] β-катенин и APC входят в число ключевых генов (наряду с другими, такими как K-Ras и SMAD4 ), участвующих в развитии колоректального рака. Потенциал β-катенина изменять ранее эпителиальный фенотип пораженных клеток на инвазивный, мезенхимоподобный тип в значительной степени способствует образованию метастазов.
В качестве терапевтической цели
[ редактировать ]Из-за его участия в развитии рака ингибирование β-катенина продолжает привлекать значительное внимание. Но нацеливание на сайт связывания на домене броненосца — не самая простая задача из-за его обширной и относительно плоской поверхности. Однако для эффективного ингибирования достаточно связывания с меньшими «горячими точками» этой поверхности. Таким образом, «сшитого» спирального пептида, полученного из природного мотива связывания β-катенина, обнаруженного в LEF1, было достаточно для полного ингибирования β-катенин-зависимой транскрипции. Недавно было также разработано несколько низкомолекулярных соединений, нацеленных на одну и ту же высокоположительно заряженную область домена ARM (CGP049090, PKF118-310, PKF115-584 и ZTM000990). Кроме того, на уровни β-катенина также можно влиять путем воздействия на вышестоящие компоненты пути Wnt, а также на комплекс разрушения β-катенина. [81] Дополнительный N-концевой связывающий карман также важен для активации гена-мишени Wnt (необходимого для рекрутирования BCL9). На этот сайт домена ARM можно фармакологически воздействовать карнозиновой кислотой . , например, [82] Этот «вспомогательный» участок — еще одна привлекательная цель для разработки лекарств. [83] Несмотря на интенсивные доклинические исследования, ингибиторы β-катенина в качестве терапевтических средств пока не доступны. Однако его функция может быть дополнительно изучена путем нокдауна siRNA на основании независимой проверки. [84] Другой терапевтический подход к уменьшению накопления в ядре β-катенина заключается в ингибировании галектина-3. [85] Ингибитор галектина-3 GR-MD-02 в настоящее время проходит клинические испытания в сочетании с одобренной FDA дозой ипилимумаба у пациентов с прогрессирующей меланомой. [86] Белки BCL9 и BCL9L были предложены в качестве терапевтических мишеней для колоректального рака, который обеспечивает гиперактивированную передачу сигналов Wnt, поскольку их делеция не нарушает нормальный гомеостаз, но сильно влияет на поведение метастазов . [87]
Роль в алкогольном синдроме плода
[ редактировать ]Дестабилизация β-катенина этанолом является одним из двух известных путей, посредством которых воздействие алкоголя вызывает алкогольный синдром плода (второй — индуцированный этанолом дефицит фолиевой кислоты). Этанол приводит к дестабилизации β-катенина по G-белково-зависимому пути, при котором активированная фосфолипаза Cβ гидролизует фосфатидилинозитол-(4,5)-бисфосфат до диацилглицерина и инозитол-(1,4,5)-трифосфата. Растворимый инозитол-(1,4,5)-трифосфат вызывает высвобождение кальция из эндоплазматического ретикулума. Это внезапное увеличение цитоплазматического кальция активирует Са2+/кальмодулин-зависимую протеинкиназу (CaMKII). Активированный CaMKII дестабилизирует β-катенин посредством плохо изученного механизма, но, вероятно, включает фосфорилирование β-катенина с помощью CaMKII. Программа транскрипции β-катенина (которая необходима для нормального развития клеток нервного гребня) тем самым подавляется, что приводит к преждевременному апоптозу клеток нервного гребня (гибель клеток). [88]
Взаимодействия
[ редактировать ]Было показано, что β-катенин взаимодействует с:
- БТР , [89] [90] [91] [92] [93] [94] [95] [96]
- АКСИН1 , [97] [98]
- Андрогенный рецептор , [99] [100] [101] [102] [103] [104]
- CBY1 , [105]
- CDH1 , [24] [90] [106] [107] [108] [109] [110] [111] [112] [113] [114] [115] [116] [117] [118] [119] [120] [121] [122] [123] [124] [125] [126]
- CDH2 , [56] [127] [128]
- CDH3 , [125] [129]
- CDK5R1 , [130]
- ЧУК , [131]
- CTNND1 , [90] [111]
- CTNNA1 , [107] [116] [132] [133] [134]
- РЭФР , [111] [120] [135]
- Эмери [136] [137]
- СОЭ1 [70]
- ФХЛ2 , [138]
- ГСК3Б , [92] [139]
- HER2/новый , [112] [135] [140]
- ХНФ4А , [103]
- ИКК2 , [131]
- ЛЕФ1 [141] [142] [143] [144] в том числе трансгенно , [145]
- МАГИ1 , [121]
- МУК1 , [113] [146] [147] [148] [149] [150] [151]
- НР5А1 , [152] [153]
- ПКАФ , [154]
- ПХФ17 , [155]
- Плакоглобин , [90] [111]
- ПТПН14 , [156]
- ПТПРФ , [112] [157]
- ПТПРК (ПТПкаппа), [158]
- ПТПРТ (ПТПрхо), [159]
- ПТПРУ (ПЦП-2), [160] [161] [162]
- ПСЭН1 , [163] [164] [165]
- ПТК7 [166]
- RuvB-подобный 1 , [167]
- СМАД7 , [141]
- SMARCA4 [168]
- SLC9A3R1 , [115]
- USP9X , [169] и
- VE-кадгерин . [170] [171]
- XIRP1 [172]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000168036 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000006932 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Краус С., Лиер Т., Хюльскен Дж., Беренс Дж., Бирчмайер В., Гржещик К.Х., Бальхаузен В.Г. (сентябрь 1994 г.). «Локализация гена бета-катенина человека (CTNNB1) в 3p21: область, участвующая в развитии опухоли». Геномика . 23 (1): 272–274. дои : 10.1006/geno.1994.1493 . ПМИД 7829088 .
- ^ Макдональд Б.Т., Тамай К., Хе Х (июль 2009 г.). «Передача сигналов Wnt/бета-катенина: компоненты, механизмы и заболевания» . Развивающая клетка . 17 (1): 9–26. дои : 10.1016/j.devcel.2009.06.016 . ПМЦ 2861485 . ПМИД 19619488 .
- ^ Пайфер М., Рауколб С., Уильямс М., Ригглман Б., Вишаус Э. (апрель 1991 г.). «Ген полярности сегмента броненосца взаимодействует с бескрылым сигнальным путем как при формировании эмбрионального, так и взрослого паттерна». Разработка . 111 (4): 1029–1043. дои : 10.1242/dev.111.4.1029 . ПМИД 1879348 .
- ^ Ноордермер Дж., Клингенсмит Дж., Перримон Н., Нусс Р. (январь 1994 г.). «Растрепанный и броненосец действует на бескрылом сигнальном пути у дрозофилы». Природа . 367 (6458): 80–83. Бибкод : 1994Natur.367...80N . дои : 10.1038/367080a0 . ПМИД 7906389 . S2CID 4275610 .
- ^ Пайфер М., Берг С., Рейнольдс А.Б. (март 1994 г.). «Повторяющийся аминокислотный мотив, общий для белков с разнообразными клеточными ролями». Клетка . 76 (5): 789–791. дои : 10.1016/0092-8674(94)90353-0 . ПМИД 7907279 . S2CID 26528190 .
- ^ Морин П.Дж. (декабрь 1999 г.). «Передача сигналов бета-катенина и рак». Биоэссе . 21 (12): 1021–1030. doi : 10.1002/(SICI)1521-1878(199912)22:1<1021::AID-BIES6>3.0.CO;2-P . ПМИД 10580987 . S2CID 86240312 .
- ^ МакКри PD, Терк CW, Гамбинер Б (ноябрь 1991 г.). «Гомолог белка броненосца у дрозофилы (плакоглобин), связанный с E-кадгерином». Наука . 254 (5036): 1359–1361. Бибкод : 1991Sci...254.1359M . дои : 10.1126/science.1962194 . ПМИД 1962194 .
- ^ Кемлер Р. (сентябрь 1993 г.). «От кадгеринов к катенинам: взаимодействия цитоплазматических белков и регуляция клеточной адгезии». Тенденции в генетике . 9 (9): 317–321. дои : 10.1016/0168-9525(93)90250-л . ПМИД 8236461 .
- ^ Готтарди CJ, Пайфер М. (март 2008 г.). «Появляются концевые области бета-катенина» . Структура . 16 (3): 336–338. дои : 10.1016/j.str.2008.02.005 . ПМК 2329800 . ПМИД 18334207 .
- ^ Син Ю, Такемару К., Лю Дж., Берндт Дж.Д., Чжэн Дж.Дж., Мун RT, Сюй В. (март 2008 г.). «Кристаллическая структура полноразмерного бета-катенина» . Структура . 16 (3): 478–487. дои : 10.1016/j.str.2007.12.021 . ПМЦ 4267759 . ПМИД 18334222 .
- ^ Фанг Д., Хоук Д., Чжэн Ю., Ся Ю., Мейзенхельдер Дж., Ника Х. и др. (апрель 2007 г.). «Фосфорилирование бета-катенина с помощью АКТ способствует транскрипционной активности бета-катенина» . Журнал биологической химии . 282 (15): 11221–11229. дои : 10.1074/jbc.M611871200 . ПМК 1850976 . ПМИД 17287208 .
- ^ Сёдерхольм С, Канту Ц (май 2021 г.). «Зависимая транскрипция WNT/β-катенин: тканеспецифический процесс» . ПРОВОДА Механизмы заболеваний . 13 (3): e1511. дои : 10.1002/wsbm.1511 . ПМЦ 9285942 . ПМИД 33085215 .
- ^ Влеминкс К., Кемлер Р., Хехт А. (март 1999 г.). «С-концевой домен трансактивации бета-катенина необходим и достаточен для передачи сигналов комплексом LEF-1/бета-катенин у Xenopus laevis» . Механизмы развития . 81 (1–2): 65–74. дои : 10.1016/s0925-4773(98)00225-1 . ПМИД 10330485 . S2CID 15086656 .
- ^ Садот Э., Симха И., Иваи К., Чехановер А., Гейгер Б., Бен-Зеев А. (апрель 2000 г.). «Дифференциальное взаимодействие плакоглобина и β-катенина с системой убиквитин-протеасома». Онкоген . 19 (16): 1992–2001. дои : 10.1038/sj.onc.1203519 . ПМИД 10803460 . S2CID 2872966 .
- ^ Актари З., Пасдар М. (2012). «Плакоглобин: роль в онкогенезе и метастазировании» . Международный журнал клеточной биологии . 2012 : 189521. дои : 10.1155/2012/189521 . ПМЦ 3312339 . ПМИД 22481945 .
- ^ Сюй В., Кимельман Д. (октябрь 2007 г.). «Механистические данные структурных исследований β-катенина и его партнеров по связыванию». Журнал клеточной науки . 120 (Часть 19): 3337–3344. дои : 10.1242/jcs.013771 . ПМИД 17881495 . S2CID 25294495 .
- ^ Грэм Т.А., Уивер С., Мао Ф., Кимельман Д., Сюй В. (декабрь 2000 г.). «Кристаллическая структура комплекса бета-катенин/Tcf» . Клетка . 103 (6): 885–896. дои : 10.1016/S0092-8674(00)00192-6 . ПМИД 11136974 . S2CID 16865193 .
- ^ Грэм Т.А., Ферки Д.М., Мао Ф., Кимельман Д., Сюй В. (декабрь 2001 г.). «Tcf4 может специфически распознавать бета-катенин, используя альтернативные конформации». Структурная биология природы . 8 (12): 1048–1052. дои : 10.1038/nsb718 . ПМИД 11713475 . S2CID 33878077 .
- ^ Пой Ф., Лепурселе М., Шивдасани Р.А., Эк М.Дж. (декабрь 2001 г.). «Структура человеческого комплекса Tcf4-бета-катенин». Структурная биология природы . 8 (12): 1053–1057. дои : 10.1038/nsb720 . ПМИД 11713476 . S2CID 24798619 .
- ^ Перейти обратно: а б Хубер А.Х., Вайс В.И. (май 2001 г.). «Структура комплекса бета-катенин/Е-кадгерин и молекулярные основы распознавания разнообразных лигандов бета-катенином» . Клетка . 105 (3): 391–402. дои : 10.1016/S0092-8674(01)00330-0 . ПМИД 11348595 . S2CID 364223 .
- ^ Перейти обратно: а б Син Ю, Клементс В.К., Кимельман Д., Сюй В. (ноябрь 2003 г.). «Кристаллическая структура комплекса бета-катенин/аксин предполагает механизм разрушения комплекса бета-катенин» . Гены и развитие . 17 (22): 2753–2764. дои : 10.1101/gad.1142603 . ПМК 280624 . ПМИД 14600025 .
- ^ Перейти обратно: а б Минде Д.П., Анвариан З., Рюдигер С.Г., Морис М.М. (август 2011 г.). «Беспорядок: как миссенс-мутации в белке-супрессоре опухолей APC приводят к раку?» . Молекулярный рак . 10 (1): 101. дои : 10.1186/1476-4598-10-101 . ПМК 3170638 . ПМИД 21859464 .
- ^ Крампс Т., Питер О., Бруннер Е., Неллен Д., Фрош Б., Чаттерджи С. и др. (апрель 2002 г.). «Передача сигналов Wnt/wingless требует BCL9/опосредованного рекрутирования пигопуса в ядерный комплекс бета-катенин-TCF» . Клетка . 109 (1): 47–60. дои : 10.1016/S0092-8674(02)00679-7 . ПМИД 11955446 . S2CID 16720801 .
- ^ Покутта С., Вейс В.И. (март 2000 г.). «Структура димеризации и бета-катенин-связывающей области альфа-катенина» . Молекулярная клетка . 5 (3): 533–543. дои : 10.1016/S1097-2765(00)80447-5 . ПМИД 10882138 .
- ^ Сампьетро Дж., Дальберг К.Л., Чо У.С., Хиндс Т.Р., Кимельман Д., Сюй В. (октябрь 2006 г.). «Кристаллическая структура комплекса бета-катенин/BCL9/Tcf4» . Молекулярная клетка . 24 (2): 293–300. doi : 10.1016/j.molcel.2006.09.001 . ПМИД 17052462 .
- ^ Брембек Ф.Х., Шварц-Ромонд Т., Баккерс Дж., Вильгельм С., Хаммершмидт М., Бирчмайер В. (сентябрь 2004 г.). «Основная роль BCL9-2 в переключении между адгезивной и транскрипционной функциями бета-катенина» . Гены и развитие . 18 (18): 2225–2230. дои : 10.1101/gad.317604 . ПМК 517514 . ПМИД 15371335 .
- ^ Лю Дж, Син Ю, Хиндс Т.Р., Чжэн Дж, Сюй В (июнь 2006 г.). «Третий повтор из 20 аминокислот является местом самого тесного связывания APC с бета-катенином». Журнал молекулярной биологии . 360 (1): 133–144. дои : 10.1016/j.jmb.2006.04.064 . ПМИД 16753179 .
- ^ Кимельман Д., Сюй В. (декабрь 2006 г.). «Комплекс разрушения бета-катенина: идеи и вопросы со структурной точки зрения». Онкоген . 25 (57): 7482–7491. дои : 10.1038/sj.onc.1210055 . ПМИД 17143292 . S2CID 20529520 .
- ^ Фидлер М., Мендоса-Топаз С., Резерфорд Т.Дж., Мещанек Дж., Биенц М. (февраль 2011 г.). «Растрепанный взаимодействует с поверхностью полимеризации домена DIX аксина, мешая его функции по подавлению β-катенина» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (5): 1937–1942. Бибкод : 2011PNAS..108.1937F . дои : 10.1073/pnas.1017063108 . ПМЦ 3033301 . ПМИД 21245303 .
- ^ Меткалф К., Биенц М. (ноябрь 2011 г.). «Ингибирование GSK3 передачей сигналов Wnt - две контрастирующие модели» . Журнал клеточной науки . 124 (Часть 21): 3537–3544. дои : 10.1242/jcs.091991 . ПМИД 22083140 .
- ^ Перейти обратно: а б Брембек Ф.Х., Росарио М., Бирчмайер В. (февраль 2006 г.). «Балансирование клеточной адгезии и передачи сигналов Wnt, ключевая роль бета-катенина». Текущее мнение в области генетики и развития . 16 (1): 51–59. дои : 10.1016/j.где.2005.12.007 . ПМИД 16377174 .
- ^ «Ген Энтреза: катенин (кадгерин-ассоциированный белок)» .
- ^ Буш М., Алханшали Б.М., Цянь С., Стэнли С.Б., Хеллер В.Т., Мацуи Т. и др. (октябрь 2019 г.). «Ансамбль гибких конформаций лежит в основе механотрансдукции с помощью адгезионного комплекса кадгерин-катенин» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (43): 21545–21555. Бибкод : 2019PNAS..11621545B . дои : 10.1073/pnas.1911489116 . ПМК 6815173 . ПМИД 31591245 .
- ^ Рёпер Дж.К., Митроссилис Д., Стирнеманн Г., Вахарте Ф., Брито И., Фернандес-Санчес М.Е. и др. (июль 2018 г.). «Основной сайт связывания β-катенина/Е-кадгерина является первичным молекулярным механотрансдуктором дифференцировки in vivo » . электронная жизнь . 7 . дои : 10.7554/eLife.33381 . ПМК 6053302 . ПМИД 30024850 .
- ^ Фараго Б., Николл И.Д., Ван С., Ченг Икс, Каллауэй DJ, Бу Зи (март 2021 г.). «Активированное наномасштабное движение актин-связывающего домена в комплексе катенин-кадгерин, выявленное с помощью нейтронной спин-эхо-спектроскопии» . Труды Национальной академии наук Соединенных Штатов Америки . 118 (13): e2025012118. Бибкод : 2021PNAS..11825012F . дои : 10.1073/pnas.2025012118 . ПМК 8020631 . ПМИД 33753508 .
- ^ Нельсон WJ (апрель 2008 г.). «Регуляция межклеточной адгезии комплексом кадгерин-катенин» . Труды Биохимического общества . 36 (Часть 2): 149–155. дои : 10.1042/BST0360149 . ПМК 3368607 . ПМИД 18363555 .
- ^ Биенц М. (январь 2005 г.). «Бета-катенин: связующее звено между клеточной адгезией и передачей сигналов Wnt» . Современная биология . 15 (2): Р64–Р67. Бибкод : 2005CBio...15..R64B . дои : 10.1016/j.cub.2004.12.058 . ПМИД 15668160 . S2CID 12352182 .
- ^ Перейти обратно: а б Хегель Х., Ларю Л., Осуги М., Федоров Л., Херренкнехт К., Кемлер Р. (ноябрь 1995 г.). «Недостаток бета-катенина влияет на развитие мышей в период гаструляции». Разработка . 121 (11): 3529–3537. дои : 10.1242/dev.121.11.3529 . ПМИД 8582267 .
- ^ Фардж Э (2003). «Механическая индукция скручивания передней кишки / стомодального зачатка дрозофилы» . Современная биология . 13 (16): 1365–1377. Бибкод : 2003CBio...13.1365F . дои : 10.1016/s0960-9822(03)00576-1 . ПМИД 1293230 .
- ^ Брюне Т., Букле А., Ахмади П., Митроссилис Д., Дрикез Б., Брюне А.С. и др. (2013). «Эволюционное сохранение спецификации ранней мезодермы посредством механотрансдукции в Bilateria» . Природные коммуникации . 4 : 2821. Бибкод : 2013NatCo...4.2821B . дои : 10.1038/ncomms3821 . ПМК 3868206 . ПМИД 24281726 .
- ^ Нгуен Н.М., Мерл Т., Бродерс-Бондон Ф., Брюне А.С., Баттистелла А., Лэнд Э.Б. и др. (2022). «Механо-биохимическая морская стимуляция инверсии, гаструляции и спецификации эндомезодермы у многоклеточных эукариот» . Границы клеточной биологии и биологии развития . 10 : 992371. дои : 10.3389/fcell.2022.992371 . ПМЦ 9754125 . ПМИД 36531949 .
- ^ Перейти обратно: а б Сокол С.Ю. (октябрь 2011 г.). «Поддержание плюрипотентности эмбриональных стволовых клеток с помощью передачи сигналов Wnt» . Разработка . 138 (20): 4341–4350. дои : 10.1242/dev.066209 . ПМК 3177306 . ПМИД 21903672 .
- ^ Шнайдер С., Штайнбейсер Х., Варга Р.М., Хаузен П. (июль 1996 г.). «Транслокация бета-катенина в ядра разграничивает дорсализующие центры у эмбрионов лягушки и рыбы» . Механизмы развития . 57 (2): 191–198. дои : 10.1016/0925-4773(96)00546-1 . ПМИД 8843396 . S2CID 12694740 .
- ^ Ларабель К.А., Торрес М., Роунинг Б.А., Йост С., Миллер Дж.Р., Ву М. и др. (март 1997 г.). «Установление дорсо-вентральной оси у эмбрионов Xenopus предвещается ранней асимметрией в β-катенине, которая модулируется сигнальным путем Wnt» . Журнал клеточной биологии . 136 (5): 1123–1136. дои : 10.1083/jcb.136.5.1123 . ПМК 2132470 . ПМИД 9060476 .
- ^ Келли ГМ, Эрезийлмаз ДФ, Мун RT (октябрь 1995 г.). «Индукция вторичной эмбриональной оси у рыбок данио происходит вследствие сверхэкспрессии бета-катенина» . Механизмы развития . 53 (2): 261–273. дои : 10.1016/0925-4773(95)00442-4 . ПМИД 8562427 . S2CID 14885037 .
- ^ Сава Х (2012). «Глава 3: Контроль полярности клеток и асимметричное деление у C. elegans ». Ин Ян Ю (ред.). Плоская полярность клеток во время развития . Текущие темы биологии развития. Том. 101. С. 55–76. дои : 10.1016/B978-0-12-394592-1.00003-X . ISBN 978-0-12-394592-1 . ПМИД 23140625 .
- ^ Баркер Н., ван Эс Дж. Х., Кейперс Дж., Куджала П., ван ден Борн М., Козейнсен М. и др. (октябрь 2007 г.). «Идентификация стволовых клеток в тонкой и толстой кишке по маркерному гену Lgr5». Природа . 449 (7165): 1003–1007. Бибкод : 2007Natur.449.1003B . дои : 10.1038/nature06196 . ПМИД 17934449 . S2CID 4349637 .
- ^ Нгуен Хо-Бульдуар Т.Х., Солье К., Замфиров Л., Бродерс-Бондон Ф., Митроссилис Д., Бермео С. и др. (февраль 2022 г.). «Механическая индукция стволовых клеток толстой кишки, опосредованная Ret-киназой, под действием давления роста опухоли стимулирует прогрессирование рака in vivo» . Коммуникационная биология . 5 (1): 137. дои : 10.1038/s42003-022-03079-4 . ПМЦ 8854631 . ПМИД 35177769 .
- ^ Тянь Х, Лю З, Ню Б, Чжан Дж, Тан ТК, Ли С.Р. и др. (2011). «Комплекс E-кадгерин/β-катенин и эпителиальный барьер» . Журнал биомедицины и биотехнологии . 2011 : 567305. doi : 10.1155/2011/567305 . ПМК 3191826 . ПМИД 22007144 .
- ^ Зеларайян Л., Герке С., Бергманн М.В. (сентябрь 2007 г.). «Роль бета-катенина в ремоделировании сердца у взрослых» . Клеточный цикл . 6 (17): 2120–2126. дои : 10.4161/cc.6.17.4632 . ПМИД 17786052 .
- ^ Ликерт Х., Куч С., Канцлер Б., Тамай Ю., Такето М.М., Кемлер Р. (август 2002 г.). «Формирование множественных сердец у мышей после удаления бета-катенина в эмбриональной энтодерме» . Развивающая клетка . 3 (2): 171–181. дои : 10.1016/s1534-5807(02)00206-x . ПМИД 12194849 .
- ^ Перейти обратно: а б Хертиг С.М., Бутц С., Кох С., Эппенбергер-Эберхардт М., Кемлер Р., Эппенбергер Х.М. (январь 1996 г.). «N-кадгерин в кардиомиоцитах взрослых крыс в культуре. II. Пространственно-временное появление белков, участвующих в межклеточном контакте и общении. Формирование двух различных комплексов N-кадгерин/катенин». Журнал клеточной науки . 109 (1): 11–20. дои : 10.1242/jcs.109.1.11 . ПМИД 8834786 .
- ^ Уилер М.А., Уорли А., Робертс Р.Г., Элер Э., Эллис Дж.А. (март 2010 г.). «Идентификация комплекса эмерин-бета-катенин в сердце, важного для вставочной архитектуры диска и локализации бета-катенина» . Клеточные и молекулярные науки о жизни . 67 (5): 781–796. дои : 10.1007/s00018-009-0219-8 . ПМЦ 11115513 . ПМИД 19997769 . S2CID 27205170 .
- ^ Ван X, Гердес AM (февраль 1999 г.). «Хроническая гипертрофия и недостаточность сердца при перегрузке давлением у морских свинок: III. Вставочное ремоделирование диска». Журнал молекулярной и клеточной кардиологии . 31 (2): 333–343. дои : 10.1006/jmcc.1998.0886 . ПМИД 10093046 .
- ^ Ёсида М., Окуса Т., Накашима Т., Таканари Х., Яно М., Такемура Г. и др. (октябрь 2011 г.). «Изменения в адгезионном соединении предшествуют ремоделированию щелевого соединения во время развития сердечной недостаточности у кардиомиопатических хомяков» . Сердечно-сосудистые исследования . 92 (1): 95–105. дои : 10.1093/cvr/cvr182 . ПМИД 21693625 .
- ^ Хан Дж.И., Чо Х.Дж., Бэ Дж.В., Юк Х.С., Ким К.И., Пак К.В. и др. (октябрь 2006 г.). «Сверхэкспрессия бета-катенина уменьшает размер инфаркта миокарда за счет дифференцированного воздействия на кардиомиоциты и сердечные фибробласты» . Журнал биологической химии . 281 (41): 30979–30989. дои : 10.1074/jbc.M603916200 . ПМИД 16920707 .
- ^ Чжэн Ц, Чен П, Сюй З, Ли Ф, И ХР (октябрь 2013 г.). «Экспрессия и перераспределение β-катенина в кардиальных миоцитах левого желудочка спонтанно гипертонической крысы». Журнал молекулярной гистологии . 44 (5): 565–573. дои : 10.1007/s10735-013-9507-6 . ПМИД 23591738 . S2CID 18997718 .
- ^ Бауранд А., Зеларайан Л., Бетни Р., Герке С., Дангер С., Ноак С. и др. (май 2007 г.). «Понижение уровня бета-катенина необходимо для адаптивного ремоделирования сердца» . Исследование кровообращения . 100 (9): 1353–1362. дои : 10.1161/01.RES.0000266605.63681.5a . ПМИД 17413044 .
- ^ Чен X, Шевцов С.П., Сич Э., Цуй Л., Хак С., Ароновиц М. и др. (июнь 2006 г.). «Сигнальный путь бета-катенин/Т-клеточный фактор/фактор энхансера лимфоцитов необходим для нормальной и вызванной стрессом сердечной гипертрофии» . Молекулярная и клеточная биология . 26 (12): 4462–4473. дои : 10.1128/MCB.02157-05 . ПМЦ 1489123 . ПМИД 16738313 .
- ^ Хак С., Майкл А., Андреуччи М., Бхаттачарья К., Дотто П., Уолтерс Б. и др. (апрель 2003 г.). «Стабилизация бета-катенина Wnt-независимым механизмом регулирует рост кардиомиоцитов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (8): 4610–4615. Бибкод : 2003PNAS..100.4610H . дои : 10.1073/pnas.0835895100 . ПМЦ 153603 . ПМИД 12668767 .
- ^ Хирши А., Крокелуа А., Перриард Е., Шенауэр Р., Агаркова И., Херструп С.П. и др. (сентябрь 2010 г.). «Стабилизированный бета-катенин в постнатальном миокарде желудочков приводит к дилатационной кардиомиопатии и преждевременной смерти» (PDF) . Фундаментальные исследования в кардиологии . 105 (5): 597–608. дои : 10.1007/s00395-010-0101-8 . ПМИД 20376467 . S2CID 21789076 .
- ^ Ли Дж., Своп Д., Раесс Н., Ченг Л., Мюллер Э.Дж., Радиче Г.Л. (март 2011 г.). «Ограниченное сердечной тканью удаление плакоглобина приводит к прогрессирующей кардиомиопатии и активации передачи сигналов бета-катенина» . Молекулярная и клеточная биология . 31 (6): 1134–1144. дои : 10.1128/MCB.01025-10 . ПМК 3067899 . ПМИД 21245375 .
- ^ Своп Д., Ченг Л., Гао Э., Ли Дж., Радиче Г.Л. (март 2012 г.). «Потеря кадгерин-связывающих белков β-катенина и плакоглобина в сердце приводит к ремоделированию щелевых контактов и аритмогенезу» . Молекулярная и клеточная биология . 32 (6): 1056–1067. дои : 10.1128/MCB.06188-11 . ПМК 3295003 . ПМИД 22252313 .
- ^ Перейти обратно: а б Диас С., Фенг Дж., Сунь Х., Шао Нью-Йорк, Мазей-Робисон М.С., Дамес-Верно Д. и др. (декабрь 2014 г.). «β-катенин опосредует устойчивость к стрессу посредством регуляции Dicer1/микроРНК» . Природа . 516 (7529): 51–55. Бибкод : 2014Natur.516...51D . дои : 10.1038/nature13976 . ПМЦ 4257892 . ПМИД 25383518 .
- ^ Перриард Дж.К., Хирши А., Элер Э. (январь 2003 г.). «Дилятационная кардиомиопатия: заболевание вставочного диска?». Тенденции сердечно-сосудистой медицины . 13 (1): 30–38. дои : 10.1016/s1050-1738(02)00209-8 . ПМИД 12554098 .
- ^ Перейти обратно: а б Махмудзаде С., Эдер С., Нордмейер Дж., Элер Э., Хубер О., Мартус П. и др. (май 2006 г.). «Повышение уровня регуляции и перераспределение альфа-рецепторов эстрогена при сердечной недостаточности у человека» . Журнал ФАСЭБ . 20 (7): 926–934. дои : 10.1096/fj.05-5148com . ПМИД 16675850 . S2CID 2246390 .
- ^ Канту С., Фелкер А., Циммерли Д., Пруммель К.Д., Кабельо Э.М., Кьяваччи Е. и др. (ноябрь 2018 г.). «Мутации в генах Bcl9 и Pygo вызывают врожденные пороки сердца из-за тканеспецифического нарушения передачи сигналов Wnt/β-катенина» . Гены и развитие . 32 (21–22): 1443–1458. дои : 10.1101/gad.315531.118 . ПМК 6217730 . ПМИД 30366904 .
- ^ Форбс С.А., Биндал Н., Бэмфорд С., Коул С., Кок С.И., Бир Д. и др. (январь 2011 г.). «COSMIC: исследование полных раковых геномов в Каталоге соматических мутаций рака» . Исследования нуклеиновых кислот . 39 (Проблема с базой данных): D945–D950. дои : 10.1093/nar/gkq929 . ПМК 3013785 . ПМИД 20952405 .
- ^ Салданья Г., Гура В., Поттер Л., Флетчер А. (июль 2004 г.). «Ядерный бета-катенин при базальноклеточной карциноме коррелирует с повышенной пролиферацией». Британский журнал дерматологии . 151 (1): 157–164. дои : 10.1111/j.1365-2133.2004.06048.x . ПМИД 15270885 . S2CID 31114274 .
- ^ Кыпта Р.М., Ваксман Дж. (август 2012 г.). «Передача сигналов Wnt/β-катенина при раке простаты». Обзоры природы. Урология . 9 (8): 418–428. дои : 10.1038/nrurol.2012.116 . ПМИД 22710668 . S2CID 22945223 .
- ^ Хасанейн А.М., Гланц С.М., Кесслер Х.П., Эскин Т.А., Лю С. (ноябрь 2003 г.). «Бета-катенин аберрантно экспрессируется в опухолях, экспрессирующих теневые клетки. Пиломатрикома, краниофарингиома и кальцифицирующая одонтогенная киста» . Американский журнал клинической патологии . 120 (5): 732–736. дои : 10.1309/EALEG7LD6W7167PX . PMID 14608900 .
- ^ Эллисон Д.В., Онилюде О.Е., Линдси Дж.К., Лушер М.Е., Уэстон К.Л., Тейлор Р.Э. и др. (ноябрь 2005 г.). «Статус бета-катенина предсказывает благоприятный исход детской медуллобластомы: Комитет по опухолям головного мозга Детской группы по изучению рака Соединенного Королевства». Журнал клинической онкологии . 23 (31): 7951–7957. дои : 10.1200/JCO.2005.01.5479 . ПМИД 16258095 .
- ^ Пуджа Навале, доктор медицинских наук, Омид Савари, доктор медицинских наук, Джозеф Ф. Томашефски-младший, доктор медицинских наук, Моника Вьяс, доктор медицинских наук «Солидное псевдопапиллярное новообразование» . Очертания патологии .
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) Последнее обновление автора: 4 марта 2022 г. - ^ Эль Сабех М., Саха С.К., Африн С., Ислам М.С., Борахай М.А. (2021). «Сигнальный путь Wnt/β-катенин при лейомиоме матки: роль в биологии опухоли и возможности нацеливания» . Мол Клеточная Биохимия . 476 (9): 3513–3536. дои : 10.1007/s11010-021-04174-6 . ПМЦ 9235413 . ПМИД 33999334 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Карлсон Дж.В., компакт-диск Флетчера (2007). «Иммуногистохимия бета-катенина в дифференциальной диагностике поражений веретенообразных клеток: анализ серии и обзор литературы» . Гистопатология . 51 (4): 509–14. дои : 10.1111/j.1365-2559.2007.02794.x . ПМИД 17711447 .
- ^ Кобаяши М., Хонма Т., Мацуда Ю., Сузуки Ю., Нарисава Р., Аджиока Ю., Асакура Х. (май 2000 г.). «Ядерная транслокация бета-катенина при колоректальном раке» . Британский журнал рака . 82 (10): 1689–1693. дои : 10.1054/bjoc.1999.1112 . ПМК 2374509 . ПМИД 10817505 .
- ^ Воронков А, Краусс С (2013). «Передача сигналов Wnt/бета-катенина и ингибиторы малых молекул» . Текущий фармацевтический дизайн . 19 (4): 634–664. дои : 10.2174/1381612811306040634 . ПМК 3529405 . ПМИД 23016862 .
- ^ де ла Рош М., Резерфорд Т.Дж., Гупта Д., Вепринцев Д.Б., Саксти Б., Фрейнд С.М., Биенц М. (февраль 2012 г.). «По своей природе лабильная α-спираль, примыкающая к сайту связывания BCL9 β-катенина, необходима для его ингибирования карнозиновой кислотой» . Природные коммуникации . 3 (2): 680. Бибкод : 2012NatCo...3..680D . дои : 10.1038/ncomms1680 . ПМК 3293410 . ПМИД 22353711 .
- ^ Такада К., Чжу Д., Берд Г.Х., Сухдео К., Чжао Дж.Дж., Мани М. и др. (август 2012 г.). «Направленное разрушение комплекса BCL9/β-катенин ингибирует онкогенную передачу сигналов Wnt» . Наука трансляционной медицины . 4 (148): 148ра117. doi : 10.1126/scitranslmed.3003808 . ПМЦ 3631420 . ПМИД 22914623 .
- ^ Мункачи Г, Штупински З, Герман П, Бан Б, Пенцвалло З, Сарвас Н, Дьерфи Б (сентябрь 2016 г.). «Подтверждение эффективности подавления РНКи с использованием данных генного массива показывает 18,5% частоты неудач в 429 независимых экспериментах» . Молекулярная терапия. Нуклеиновые кислоты . 5 (9): е366. дои : 10.1038/mtna.2016.66 . ПМК 5056990 . ПМИД 27673562 .
- ^ Цао З, Хао З, Синь М, Ю Л, Ван Л, Чжан Ю и др. (декабрь 2018 г.). «Эндогенный и экзогенный галектин-3 способствует адгезии опухолевых клеток с низкой экспрессией MUC1 к HUVEC посредством активации N-кадгерина и CD44» . Лабораторные исследования; Журнал технических методов и патологии . 98 (12): 1642–1656. дои : 10.1038/s41374-018-0119-3 . ПМИД 30171204 . S2CID 52139917 .
- ^ Номер клинического исследования NCT02117362 «Ингибитор галектина (GR-MD-02) и ипилимумаб у пациентов с метастатической меланомой» на сайте ClinicalTrials.gov.
- ^ Мур А.Е., Андерле П., Канту С., Родригес П., Видеманн Н., Барутио Ф. и др. (декабрь 2015 г.). «Передача сигналов BCL9/9L-β-катенина связана с плохим исходом при колоректальном раке» . Электронная биомедицина . 2 (12): 1932–1943. дои : 10.1016/j.ebiom.2015.10.030 . ПМЦ 4703711 . ПМИД 26844272 .
- ^ Флентке Г.Р., Гарик А., Амбергер Э., Эрнандес М., Смит С.М. (июль 2011 г.). «Опосредованная кальцием репрессия β-катенина и его транскрипционная передача сигналов опосредуют гибель клеток нервного гребня в птичьей модели алкогольного синдрома плода» . Исследование врожденных дефектов. Часть A. Клиническая и молекулярная тератология . 91 (7): 591–602. дои : 10.1002/bdra.20833 . ПМЦ 4827605 . ПМИД 21630427 .
- ^ Су Л.К., Фогельштейн Б., Кинцлер К.В. (декабрь 1993 г.). «Ассоциация белка-супрессора опухоли APC с катенинами». Наука . 262 (5140): 1734–1737. Бибкод : 1993Sci...262.1734S . дои : 10.1126/science.8259519 . ПМИД 8259519 .
- ^ Перейти обратно: а б с д Куцерова Д, Слонцова Е, Тугачкова З, Войтехова М, Совова В (декабрь 2001 г.). «Экспрессия и взаимодействие различных катенинов в клетках колоректальной карциномы». Международный журнал молекулярной медицины . 8 (6): 695–698. дои : 10.3892/ijmm.8.6.695 . ПМИД 11712088 .
- ^ Тикенброк Л., Кессмайер К., Реманн Х., Херрманн С., Мюллер О. (март 2003 г.). «Различия между взаимодействием бета-катенина с нефосфорилированными и одноимитированными фосфорилированными повторами из 20 аминокислотных остатков белка APC». Журнал молекулярной биологии . 327 (2): 359–367. дои : 10.1016/S0022-2836(03)00144-X . ПМИД 12628243 .
- ^ Перейти обратно: а б Дэвис Дж., Цзян В.Г., Мейсон, доктор медицинских наук (апрель 2001 г.). «Взаимодействие между бета-катенином, GSK3beta и APC после мотоген-индуцированной межклеточной диссоциации и их участие в путях передачи сигнала при раке простаты». Международный журнал онкологии . 18 (4): 843–847. дои : 10.3892/ijo.18.4.843 . ПМИД 11251183 .
- ^ Ре А., Накамура М., Вульф Г., Лиу Ю.К., Лу КП (сентябрь 2001 г.). «Pin1 регулирует оборот и субклеточную локализацию бета-катенина, ингибируя его взаимодействие с APC». Природная клеточная биология . 3 (9): 793–801. дои : 10.1038/ncb0901-793 . ПМИД 11533658 . S2CID 664553 .
- ^ Хомма М.К., Ли Д., Кребс Э.Г., Юаса Ю., Хомма Ю. (апрель 2002 г.). «Ассоциация и регуляция активности казеинкиназы 2 белком аденоматозного полипоза coli» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (9): 5959–5964. Бибкод : 2002PNAS...99.5959K . дои : 10.1073/pnas.092143199 . ПМК 122884 . ПМИД 11972058 .
- ^ Сато К., Янаи Х., Сенда Т., Коху К., Накамура Т., Окумура Н. и др. (июнь 1997 г.). «DAP-1, новый белок, который взаимодействует с гуанилаткиназоподобными доменами hDLG и PSD-95» . Гены в клетки . 2 (6): 415–424. дои : 10.1046/j.1365-2443.1997.1310329.x . ПМИД 9286858 . S2CID 8934092 .
- ^ Эклоф Спинк К., Фридман С.Г., Вайс В.И. (ноябрь 2001 г.). «Молекулярные механизмы узнавания бета-катенина аденоматозным полипозом кишечной палочки, выявленные по структуре комплекса АПК-бета-катенин» . Журнал ЭМБО . 20 (22): 6203–6212. дои : 10.1093/emboj/20.22.6203 . ПМК 125720 . ПМИД 11707392 .
- ^ Накамура Т., Хамада Ф., Исидате Т., Анаи К., Кавахара К., Тоёсима К., Акияма Т. (июнь 1998 г.). «Аксин, ингибитор сигнального пути Wnt, взаимодействует с бета-катенином, GSK-3beta и APC и снижает уровень бета-катенина» . Гены в клетки . 3 (6): 395–403. дои : 10.1046/j.1365-2443.1998.00198.x . ПМИД 9734785 . S2CID 10875463 .
- ^ Хочевар Б.А., Моу Ф., Реннольдс Дж.Л., Моррис С.М., Купер Дж.А., Хоу П.Х. (июнь 2003 г.). «Регуляция сигнального пути Wnt с помощью инвалида-2 (Dab2)» . Журнал ЭМБО . 22 (12): 3084–3094. дои : 10.1093/emboj/cdg286 . ПМК 162138 . ПМИД 12805222 .
- ^ Ян Ф., Ли Х, Шарма М., Сасаки С.И., Лонго Д.Л., Лим Б., Сунь З. (март 2002 г.). «Связь бета-катенина с сигнальным путем андрогенов» . Журнал биологической химии . 277 (13): 11336–11344. дои : 10.1074/jbc.M111962200 . ПМИД 11792709 .
- ^ Масиелло Д., Чен С.Ю., Сюй Ю., Верховен М.К., Чой Э., Холленберг А.Н., Балк С.П. (октябрь 2004 г.). «Привлечение бета-катенина диким типом или мутантными андрогенными рецепторами коррелирует со стимулируемым лигандом ростом клеток рака простаты» . Молекулярная эндокринология . 18 (10): 2388–2401. дои : 10.1210/me.2003-0436 . ПМИД 15256534 .
- ^ Сонг Л.Н., Коглан М., Гельманн Э.П. (январь 2004 г.). «Антиандрогенные эффекты мифепристона на коактиваторные и корепрессорные взаимодействия с андрогенными рецепторами» . Молекулярная эндокринология . 18 (1): 70–85. дои : 10.1210/me.2003-0189 . ПМИД 14593076 .
- ^ Амир А.Л., Баруа М., Макнайт Н.К., Ченг С., Юань Х., Балк С.П. (август 2003 г.). «Прямое бета-катенин-независимое взаимодействие между андрогенным рецептором и Т-клеточным фактором 4» . Журнал биологической химии . 278 (33): 30828–30834. дои : 10.1074/jbc.M301208200 . ПМИД 12799378 .
- ^ Перейти обратно: а б Малхолланд DJ, Рид Дж.Т., Ренни П.С., Кокс М.Э., Нельсон CC (август 2003 г.). «Функциональная локализация и конкуренция между андрогенным рецептором и фактором Т-клеток за ядерный бета-катенин: средство ингибирования сигнальной оси Tcf». Онкоген . 22 (36): 5602–5613. дои : 10.1038/sj.onc.1206802 . ПМИД 12944908 . S2CID 9301471 .
- ^ Павловский Дж. Э., Эртель Дж. Р., Аллен М. П., Сюй М., Батлер С., Уилсон Э. М., Вирман М. Е. (июнь 2002 г.). «Взаимодействие лигандированного андрогенного рецептора с бета-катенином: ядерная совместная локализация и модуляция транскрипционной активности в нейрональных клетках» . Журнал биологической химии . 277 (23): 20702–20710. дои : 10.1074/jbc.M200545200 . ПМИД 11916967 .
- ^ Такемару К., Ямагути С., Ли Ю.С., Чжан Ю., Картью Р.В., Мун RT (апрель 2003 г.). «Чибби, ядерный бета-катенин-ассоциированный антагонист пути Wnt/Wingless». Природа . 422 (6934): 905–909. Бибкод : 2003Natur.422..905T . дои : 10.1038/nature01570 . ПМИД 12712206 . S2CID 4418716 .
- ^ Дэвис Дж., Цзян В.Г., Мейсон, доктор медицинских наук (апрель 2001 г.). «HGF/SF изменяет взаимодействие между его рецептором c-Met и комплексом E-кадгерин/катенин в клетках рака простаты». Международный журнал молекулярной медицины . 7 (4): 385–388. дои : 10.3892/ijmm.7.4.385 . ПМИД 11254878 .
- ^ Перейти обратно: а б Ояма Т., Канаи Ю., Отиаи А., Акимото С., Ода Т., Янагихара К. и др. (декабрь 1994 г.). «Укороченный бета-катенин нарушает взаимодействие между E-кадгерином и альфа-катенином: причина потери межклеточной адгезии в линиях раковых клеток человека». Исследования рака . 54 (23): 6282–6287. ПМИД 7954478 .
- ^ Хазан Р.Б., Канг Л., Роу С., Борген П.И., Римм Д.Л. (декабрь 1997 г.). «Винкулин связан с адгезионным комплексом Е-кадгерина» . Журнал биологической химии . 272 (51): 32448–32453. дои : 10.1074/jbc.272.51.32448 . ПМИД 9405455 .
- ^ Кинч М.С., Кларк Г.Дж., Дер СиДжей, Берридж К. (июль 1995 г.). «Фосфорилирование тирозина регулирует спайки рас-трансформированного эпителия молочной железы» . Журнал клеточной биологии . 130 (2): 461–471. дои : 10.1083/jcb.130.2.461 . ПМК 2199929 . ПМИД 7542250 .
- ^ Цзян MC, Ляо CF, Тай CC (июнь 2002 г.). «CAS/CSE 1 стимулирует E-кадрин-зависимую полярность клеток в эпителиальных клетках толстой кишки человека HT-29». Связь с биохимическими и биофизическими исследованиями . 294 (4): 900–905. дои : 10.1016/S0006-291X(02)00551-X . ПМИД 12061792 .
- ^ Перейти обратно: а б с д Хазан Р.Б., Нортон Л. (апрель 1998 г.). «Рецептор эпидермального фактора роста модулирует взаимодействие Е-кадгерина с актиновым цитоскелетом» . Журнал биологической химии . 273 (15): 9078–9084. дои : 10.1074/jbc.273.15.9078 . ПМИД 9535896 .
- ^ Перейти обратно: а б с Бонвини П., Ан В.Г., Розолен А., Нгуен П., Трепель Дж., Гарсия де Эррерос А. и др. (февраль 2001 г.). «Гелданамицин отменяет ассоциацию ErbB2 с резистентным к протеасомам бета-катенином в клетках меланомы, увеличивает ассоциацию бета-катенин-Е-кадгерин и снижает транскрипцию, чувствительную к бета-катенину». Исследования рака . 61 (4): 1671–1677. ПМИД 11245482 .
- ^ Перейти обратно: а б Ли Ю, Бхарти А, Чен Д, Гонг Дж, Куфе Д (декабрь 1998 г.). «Взаимодействие киназы гликогенсинтазы 3бета с антигеном, связанным с карциномой DF3/MUC1, и бета-катенином» . Молекулярная и клеточная биология . 18 (12): 7216–7224. дои : 10.1128/mcb.18.12.7216 . ПМК 109303 . ПМИД 9819408 .
- ^ Венделер М.В., Праус М., Юнг Р., Хекинг М., Мециг С., Гесснер Р. (апрель 2004 г.). «Ksp-кадгерин представляет собой функциональную молекулу межклеточной адгезии, родственную LI-кадгерину». Экспериментальные исследования клеток . 294 (2): 345–355. дои : 10.1016/j.yexcr.2003.11.022 . ПМИД 15023525 .
- ^ Перейти обратно: а б Шибата Т., Чума М., Кокубу А., Сакамото М., Хирохаши С. (июль 2003 г.). «EBP50, белок, ассоциированный с бета-катенином, усиливает передачу сигналов Wnt и сверхэкспрессируется при гепатоцеллюлярной карциноме» . Гепатология . 38 (1): 178–186. дои : 10.1053/jhep.2003.50270 . ПМИД 12830000 . S2CID 10325091 .
- ^ Перейти обратно: а б Пьедра Х., Миравет С., Кастаньо Х., Палмер Х.Г., Хейстеркамп Н., Гарсиа де Эррерос А., Дуньяк М. (апрель 2003 г.). «Связанные с катенином тирозинкиназы Fer и Fyn регулируют фосфорилирование бета-катенина Tyr-142 и взаимодействие бета-катенин-альфа-катенин» . Молекулярная и клеточная биология . 23 (7): 2287–2297. дои : 10.1128/MCB.23.7.2287-2297.2003 . ПМК 150740 . ПМИД 12640114 .
- ^ Канг Дж.С., Фейнлейб Дж.Л., Нокс С., Кеттерингем М.А., Краусс Р.С. (апрель 2003 г.). «Промиогенные члены семейств Ig и кадгеринов взаимодействуют, положительно регулируя дифференцировку» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (7): 3989–3994. Бибкод : 2003PNAS..100.3989K . дои : 10.1073/pnas.0736565100 . ПМК 153035 . ПМИД 12634428 .
- ^ Онеяма С., Накано Х., Шарма С.В. (март 2002 г.). «UCS15A, новая небольшая молекула, препарат, блокирующий белок-белковое взаимодействие, опосредованный доменом SH3». Онкоген . 21 (13): 2037–2050. дои : 10.1038/sj.onc.1205271 . ПМИД 11960376 . S2CID 23869665 .
- ^ Наварро П., Лозано Э., Кано А. (август 1993 г.). «Экспрессии E- или P-кадгерина недостаточно для изменения морфологии и туморогенного поведения клеток веретенообразной карциномы мышей. Возможное участие плакоглобина». Журнал клеточной науки . 105 (4): 923–934. дои : 10.1242/jcs.105.4.923 . hdl : 10261/78716 . ПМИД 8227214 .
- ^ Перейти обратно: а б Такахаши К., Сузуки К., Цукатани Ю. (июль 1997 г.). «Индукция фосфорилирования тирозина и ассоциация бета-катенина с рецептором EGF при триптическом расщеплении покоящихся клеток при слиянии». Онкоген . 15 (1): 71–78. дои : 10.1038/sj.onc.1201160 . ПМИД 9233779 . S2CID 10127053 .
- ^ Перейти обратно: а б Добросоцкая И.Ю., Джеймс Г.Л. (апрель 2000 г.). «MAGI-1 взаимодействует с бета-катенином и связан со структурами межклеточной адгезии». Связь с биохимическими и биофизическими исследованиями . 270 (3): 903–909. дои : 10.1006/bbrc.2000.2471 . ПМИД 10772923 .
- ^ Гэн Л., Берроу Ч.Р., Ли Х.П., Уилсон П.Д. (декабрь 2000 г.). «Модификация состава мультибелковых комплексов полицистина-1 путем фосфорилирования кальция и тирозина». Biochimica et Biophysica Acta (BBA) - Молекулярные основы болезней . 1535 (1): 21–35. дои : 10.1016/S0925-4439(00)00079-X . ПМИД 11113628 .
- ^ Сибамото С., Хаякава М., Такеучи К., Хори Т., Миядзава К., Китамура Н. и др. (март 1995 г.). «Ассоциация p120, субстрата тирозинкиназы, с комплексами E-кадгерин/катенин» . Журнал клеточной биологии . 128 (5): 949–957. дои : 10.1083/jcb.128.5.949 . ПМК 2120395 . ПМИД 7876318 .
- ^ Рао Р.К., Басурой С., Рао В.У., Карнаки К.Дж., Гупта А. (декабрь 2002 г.). «Фосфорилирование тирозина и диссоциация комплексов окклюдин-ZO-1 и E-кадгерин-бета-катенин из цитоскелета под действием окислительного стресса» . Биохимический журнал . 368 (Часть 2): 471–481. дои : 10.1042/BJ20011804 . ПМЦ 1222996 . ПМИД 12169098 .
- ^ Перейти обратно: а б Шмайзер К., Гранд Р.Дж. (апрель 1999 г.). «Судьба Е- и Р-кадгерина на ранних стадиях апоптоза» . Смерть клеток и дифференцировка . 6 (4): 377–386. дои : 10.1038/sj.cdd.4400504 . ПМИД 10381631 .
- ^ Пай Р., Данлэп Д., Цин Дж., Мохташеми И., Хотцель К., французский DM (июль 2008 г.). «Ингибирование фактора роста фибробластов 19 снижает рост опухоли за счет модуляции передачи сигналов бета-катенина» . Исследования рака . 68 (13): 5086–5095. doi : 10.1158/0008-5472.CAN-07-2325 . ПМИД 18593907 .
- ^ Штрауб Б.К., Бода Дж., Кун С., Шнольцер М., Корф У., Кемпф Т. и др. (декабрь 2003 г.). «Новая система межклеточных соединений: мозаика прикреплённой коры из клеток волокон хрусталика» . Журнал клеточной науки . 116 (Часть 24): 4985–4995. дои : 10.1242/jcs.00815 . ПМИД 14625392 .
- ^ Уол Дж.К., Ким Ю.Дж., Каллен Дж.М., Джонсон К.Р., Уилок М.Дж. (май 2003 г.). «Комплексы N-кадгерин-катенин образуются до расщепления прообласти и транспорта к плазматической мембране» . Журнал биологической химии . 278 (19): 17269–17276. дои : 10.1074/jbc.M211452200 . ПМИД 12604612 .
- ^ Клингельхёфер Дж., Трояновский Р.Б., Лаур О.Ю., Трояновский С. (август 2000 г.). «Аминоконцевой домен классических кадгеринов определяет специфичность адгезивных взаимодействий». Журнал клеточной науки . 113 (16): 2829–2836. дои : 10.1242/jcs.113.16.2829 . ПМИД 10910767 .
- ^ Кесавапани С., Лау К.Ф., Маклафлин Д.М., Браунлис Дж., Акерли С., Ли П.Н. и др. (январь 2001 г.). «p35/cdk5 связывает и фосфорилирует бета-катенин и регулирует взаимодействие бета-катенин/пресенилин-1». Европейский журнал неврологии . 13 (2): 241–247. doi : 10.1046/j.1460-9568.2001.01376.x (неактивен 10 июля 2024 г.). ПМИД 11168528 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на июль 2024 г. ( ссылка ) - ^ Перейти обратно: а б Ламберти С., Лин К.М., Ямамото Ю., Верма У., Верма И.М., Байерс С., Гейнор Р.Б. (ноябрь 2001 г.). «Регуляция функции бета-катенина киназами IkappaB» . Журнал биологической химии . 276 (45): 42276–42286. дои : 10.1074/jbc.M104227200 . ПМИД 11527961 .
- ^ Роу С., Козлов Э.Р., Римм Д.Л. (июнь 1998 г.). «Мутация альфа-катенина нарушает адгезию в клетках клона А, не нарушая при этом его активность по связыванию актина и бета-катенина» . Клеточная адгезия и связь . 5 (4): 283–296. дои : 10.3109/15419069809040298 . ПМИД 9762469 .
- ^ Аберле Х., Бутц С., Стаперт Дж., Вайссиг Х., Кемлер Р., Хошуецки Х. (декабрь 1994 г.). «Сборка комплекса кадгерин-катенин in vitro с рекомбинантными белками». Журнал клеточной науки . 107 (12): 3655–3663. дои : 10.1242/jcs.107.12.3655 . ПМИД 7706414 .
- ^ Реувер С.М., Гарнер CC (апрель 1998 г.). «Клеточная адгезия, опосредованная E-кадгерином, привлекает SAP97 в кортикальный цитоскелет». Журнал клеточной науки . 111 (8): 1071–1080. дои : 10.1242/jcs.111.8.1071 . ПМИД 9512503 .
- ^ Перейти обратно: а б Шредер Дж.А., Адрианс MC, МакКоннелл Э.Дж., Томпсон MC, Покадж Б., Гендлер С.Дж. (июнь 2002 г.). «Комплексы ErbB-бета-катенин связаны с инфильтративным вирусом опухоли молочной железы человека и мышиной молочной железы (MMTV)-Wnt-1 и трансгенными карциномами MMTV-c-Neu» . Журнал биологической химии . 277 (25): 22692–22698. дои : 10.1074/jbc.M201975200 . ПМИД 11950845 .
- ^ Картеньи Л., ди Барлетта М.Р., Баррези Р., Скуарзони С., Сабателли П., Маральди Н. и др. (декабрь 1997 г.). «Сердцеспецифичная локализация эмерина: новый взгляд на мышечную дистрофию Эмери-Дрейфуса» . Молекулярная генетика человека . 6 (13): 2257–2264. дои : 10.1093/hmg/6.13.2257 . ПМИД 9361031 .
- ^ Маркевич Э., Тилгнер К., Баркер Н., ван де Ветеринг М., Клеверс Х., Доробек М. и др. (июль 2006 г.). «Белок внутренней ядерной мембраны эмерин регулирует активность бета-катенина, ограничивая его накопление в ядре» . Журнал ЭМБО . 25 (14): 3275–3285. дои : 10.1038/sj.emboj.7601230 . ПМЦ 1523183 . ПМИД 16858403 .
- ^ Вэй Ю., Ренар К.А., Лабалетт С., Ву Ю., Леви Л., Новё С. и др. (февраль 2003 г.). «Идентификация белка LIM FHL2 как коактиватора бета-катенина» . Журнал биологической химии . 278 (7): 5188–5194. дои : 10.1074/jbc.M207216200 . ПМИД 12466281 .
- ^ Кисида С., Ямамото Х., Хино С., Икеда С., Кисида М., Кикучи А. (июнь 1999 г.). «DIX-домены Dvl и аксина необходимы для взаимодействия белков и их способности регулировать стабильность бета-катенина» . Молекулярная и клеточная биология . 19 (6): 4414–4422. дои : 10.1128/mcb.19.6.4414 . ПМК 104400 . ПМИД 10330181 .
- ^ Канаи Ю, Отиаи А, Сибата Т, Ояма Т, Ушиджима С, Акимото С, Хирохаши С (март 1995 г.). «Продукт гена c-erbB-2 напрямую связывается с бета-катенином и плакоглобином». Связь с биохимическими и биофизическими исследованиями . 208 (3): 1067–1072. дои : 10.1006/bbrc.1995.1443 . ПМИД 7702605 .
- ^ Перейти обратно: а б Эдлунд С., Ли С.Ю., Гримсби С., Чжан С., Аспенстрем П., Хелдин Ч., Ландстрем М. (февраль 2005 г.). «Взаимодействие между Smad7 и бета-катенином: важность трансформации апоптоза, индуцированного бета-фактором роста» . Молекулярная и клеточная биология . 25 (4): 1475–1488. дои : 10.1128/MCB.25.4.1475-1488.2005 . ПМК 548008 . ПМИД 15684397 .
- ^ Грюнеберг Д.А., Пабло Л., Ху К.К., Август П., Венг З., Папкофф Дж. (июнь 2003 г.). «Функциональный скрининг в клетках человека идентифицирует UBF2 как фактор транскрипции РНК-полимеразы II, который усиливает сигнальный путь бета-катенина» . Молекулярная и клеточная биология . 23 (11): 3936–3950. дои : 10.1128/MCB.23.11.3936-3950.2003 . ПМК 155208 . ПМИД 12748295 .
- ^ Беренс Дж., фон Крис Дж.П., Кюль М., Брюн Л., Ведлих Д., Гроссшедль Р., Бирхмайер В. (август 1996 г.). «Функциональное взаимодействие бета-катенина с фактором транскрипции LEF-1». Природа . 382 (6592): 638–642. Бибкод : 1996Natur.382..638B . дои : 10.1038/382638a0 . ПМИД 8757136 . S2CID 4369341 .
- ^ Лаббе Э., Летамендия А., Аттизано Л. (июль 2000 г.). «Ассоциация Smads с фактором связывания лимфоидного энхансера 1/специфическим для Т-клеток фактором опосредует совместную передачу сигналов посредством путей трансформирующего фактора роста-бета и wnt» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (15): 8358–8363. Бибкод : 2000PNAS...97.8358L . дои : 10.1073/pnas.150152697 . ПМК 26952 . ПМИД 10890911 .
- ^ Бароло С., Посакони Дж.В. (май 2002 г.). «Три особенности высокоэффективных сигнальных путей: принципы контроля транскрипции посредством передачи сигналов клеток развития» . Гены и развитие . 16 (10). Лабораторное издательство Колд-Спринг-Харбор и Общество генетиков : 1167–1181. дои : 10.1101/gad.976502 . ПМИД 12023297 . S2CID 14376483 . п. 1170:
У... рыбок данио репортерные трансгены, содержащие промотор TOPFLASH, экспрессируются в определенных Wnt-чувствительных типах клеток (...Dorsky et al. 2002).
- ^ Ямамото М., Бхарти А., Ли Й., Куфе Д. (май 1997 г.). «Взаимодействие антигена, ассоциированного с карциномой молочной железы DF3/MUC1, и бета-катенина в клеточной адгезии» . Журнал биологической химии . 272 (19): 12492–12494. дои : 10.1074/jbc.272.19.12492 . ПМИД 9139698 .
- ^ Дурум С.К., Айелло Ф.Б. (2003). «Интерлейкин-7 индуцирует MUC1» . Биология и терапия рака . 2 (2): 194–195. дои : 10.4161/cbt.2.2.351 . ПМИД 12750562 .
- ^ Шредер Дж. А., Адрианс М. К., Томпсон М. К., Камениш Т. Д., Гендлер С. Дж. (март 2003 г.). «MUC1 изменяет бета-катенин-зависимое образование опухолей и способствует клеточной инвазии». Онкоген . 22 (9): 1324–1332. дои : 10.1038/sj.onc.1206291 . ПМИД 12618757 . S2CID 25619311 .
- ^ Ли Ю, Кувахара Х, Рен Дж, Вэнь Г, Куфе Д (март 2001 г.). «Тирозинкиназа c-Src регулирует передачу сигналов человеческого антигена, ассоциированного с карциномой DF3/MUC1, с помощью бета-GSK3 и бета-катенина» . Журнал биологической химии . 276 (9): 6061–6064. дои : 10.1074/jbc.C000754200 . ПМИД 11152665 .
- ^ Рен Дж., Ли Ю., Куфе Д. (май 2002 г.). «Протеинкиназа C дельта регулирует функцию антигена карциномы DF3/MUC1 в передаче сигналов бета-катенина» . Журнал биологической химии . 277 (20): 17616–17622. дои : 10.1074/jbc.M200436200 . ПМИД 11877440 .
- ^ Ли Ю, Рен Дж, Ю В, Ли К, Кувахара Х, Инь Л и др. (сентябрь 2001 г.). «Рецептор эпидермального фактора роста регулирует взаимодействие антигена карциномы человека DF3/MUC1 с c-Src и бета-катенином» . Журнал биологической химии . 276 (38): 35239–35242. дои : 10.1074/jbc.C100359200 . ПМИД 11483589 .
- ^ Кеннелл Дж.А., О'Лири Э.Э., Гаммоу Б.М., Хаммер Г.Д., Макдугалд О.А. (август 2003 г.). «Т-клеточный фактор 4N (TCF-4N), новая изоформа мышиного TCF-4, взаимодействует с бета-катенином, коактивируя факторы транскрипции C/EBPalpha и стероидогенного фактора 1» . Молекулярная и клеточная биология . 23 (15): 5366–5375. дои : 10.1128/MCB.23.15.5366-5375.2003 . ПМЦ 165725 . ПМИД 12861022 .
- ^ Мизусаки Х., Кавабе К., Мукаи Т., Ариеси Э., Касахара М., Ёсиока Х. и др. (апрель 2003 г.). «Транскрипция гена Dax-1 (зависимая от дозы критическая область реверсии пола – врожденной гипоплазии надпочечников на Х-хромосоме, ген 1) регулируется wnt4 в развивающейся гонаде самки» . Молекулярная эндокринология . 17 (4): 507–519. дои : 10.1210/me.2002-0362 . ПМИД 12554773 .
- ^ Гэ Икс, Цзинь Ц, Чжан Ф, Ян Т, Чжай Ц (январь 2009 г.). «PCAF ацетилирует {бета}-катенин и повышает его стабильность» . Молекулярная биология клетки . 20 (1): 419–427. дои : 10.1091/mbc.E08-08-0792 . ПМК 2613091 . ПМИД 18987336 .
- ^ Беренс Дж. (октябрь 2008 г.). «Один удар, два исхода для онкогенеза, опосредованного VHL». Природная клеточная биология . 10 (10): 1127–1128. дои : 10.1038/ncb1008-1127 . ПМИД 18830218 . S2CID 36184371 .
- ^ Уодэм С., Гэмбл-младший, Вадас М.А., Хью-Гудалл Ю. (июнь 2003 г.). «Протеин тирозинфосфатаза Pez является основной фосфатазой слипчивых соединений и дефосфорилирует бета-катенин» . Молекулярная биология клетки . 14 (6): 2520–2529. doi : 10.1091/mbc.E02-09-0577 . ЧВК 194899 . ПМИД 12808048 .
- ^ Айхер Б., Лерх М.М., Мюллер Т., Шиллинг Дж., Ульрих А. (август 1997 г.). «Клеточное перераспределение протеинтирозинфосфатаз LAR и PTPsigma путем индуцируемого протеолитического процессинга» . Журнал клеточной биологии . 138 (3): 681–696. дои : 10.1083/jcb.138.3.681 . ПМК 2141638 . ПМИД 9245795 .
- ^ Фукс М., Мюллер Т., Лерх М.М., Ульрих А. (июль 1996 г.). «Ассоциация человеческой протеин-тирозинфосфатазы каппа с представителями семейства броненосцев» . Журнал биологической химии . 271 (28): 16712–16719. дои : 10.1074/jbc.271.28.16712 . ПМИД 8663237 .
- ^ Беско Дж.А., Хофт ван Хейсдуйнен Р., Фростхольм А., Роттер А. (октябрь 2006 г.). «Внутриклеточные субстраты мозгового рецептора протеина тирозинфосфатазы ро (RPTPrho/PTPRT)». Исследования мозга . 1116 (1): 50–57. дои : 10.1016/j.brainres.2006.07.122 . ПМИД 16973135 . S2CID 23343123 .
- ^ Ван Б., Кишихара К., Чжан Д., Хара Х., Номото К. (февраль 1997 г.). «Молекулярное клонирование и характеристика нового гена человеческого рецепторного белка тирозинфосфатазы, hPTP-J: подавление экспрессии гена с помощью PMA и ионофора кальция в клетках Т-лимфомы Jurkat». Связь с биохимическими и биофизическими исследованиями . 231 (1): 77–81. дои : 10.1006/bbrc.1997.6004 . ПМИД 9070223 .
- ^ Ян Х.Х., Хэ Ю.К., Донг Х., Чжан П., Цзэн Дж.З., Цао Х.Ф. и др. (декабрь 2002 г.). «Физическое и функциональное взаимодействие между рецептороподобным протеином тирозинфосфатазой PCP-2 и бета-катенином». Биохимия . 41 (52): 15854–15860. дои : 10.1021/bi026095u . ПМИД 12501215 .
- ^ Хэ Ю, Ян Х, Донг Х, Чжан П, Тан Л, Цю X и др. (апрель 2005 г.). «Структурные основы взаимодействия протеинтирозинфосфатазы PCP-2 и бета-катенина». Наука в Китае. Серия C: Науки о жизни . 48 (2): 163–167. дои : 10.1007/bf02879669 . ПМИД 15986889 . S2CID 20799629 .
- ^ Теско Г., Ким Т.В., Дильманн А., Бейройтер К., Танци Р.Э. (декабрь 1998 г.). «Отмена взаимодействия пресенилин 1/бета-катенин и сохранение гетеродимерного комплекса пресенилин 1 после активации каспазы» . Журнал биологической химии . 273 (51): 33909–33914. дои : 10.1074/jbc.273.51.33909 . ПМИД 9852041 .
- ^ Канг Д.Э., Сориано С., Фрош М.П., Коллинз Т., Нарусе С., Сисодиа С.С. и др. (июнь 1999 г.). «Пресенилин 1 способствует конститутивному обмену бета-катенина: дифференциальная активность связанных с болезнью Альцгеймера мутантов PS1 в пути передачи сигналов бета-катенина» . Журнал неврологии . 19 (11): 4229–4237. doi : 10.1523/JNEUROSCI.19-11-04229.1999 . ПМК 6782616 . ПМИД 10341227 .
- ^ Мураяма М., Танака С., Паласино Дж., Мураяма О., Хонда Т., Сан Х и др. (август 1998 г.). «Прямая ассоциация пресенилина-1 с бета-катенином». Письма ФЭБС . 433 (1–2): 73–77. дои : 10.1016/S0014-5793(98)00886-2 . ПМИД 9738936 . S2CID 85416623 .
- ^ Пуппо Ф., Томе В., Лумо А.С., Сибуа М., Гангар А., Лембо Ф. и др. (январь 2011 г.). «Протеинтирозинкиназа 7 играет консервативную роль в канонической передаче сигналов Wnt/β-катенин» . Отчеты ЭМБО . 12 (1): 43–49. дои : 10.1038/embor.2010.185 . ПМК 3024124 . ПМИД 21132015 .
- ^ Бауэр А., Хубер О., Кемлер Р. (декабрь 1998 г.). «Pontin52, партнер по взаимодействию бета-катенина, связывается с белком, связывающим ТАТА-бокс» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (25): 14787–14792. Бибкод : 1998PNAS...9514787B . дои : 10.1073/pnas.95.25.14787 . ПМК 24527 . ПМИД 9843967 .
- ^ Баркер Н., Херлстоун А., Мусиси Х., Майлз А., Биенц М., Клеверс Х. (сентябрь 2001 г.). «Фактор ремоделирования хроматина Brg-1 взаимодействует с бета-катенином, способствуя активации целевого гена» . Журнал ЭМБО . 20 (17): 4935–4943. дои : 10.1093/emboj/20.17.4935 . ПМК 125268 . ПМИД 11532957 .
- ^ Тая С., Ямамото Т., Канаи-Адзума М., Вуд С.А., Кайбути К. (декабрь 1999 г.). «Дубиквитинирующий фермент Fam взаимодействует с бета-катенином и стабилизирует его» . Гены в клетки . 4 (12): 757–767. дои : 10.1046/j.1365-2443.1999.00297.x . ПМИД 10620020 . S2CID 85747886 .
- ^ Левалле Дж. М., Бажу К., Десрё Дж., Марил М., Дежана Э., Ноэль А., Фойдар Ж. М. (декабрь 1997 г.). «Изменение межэндотелиальных слипчивых соединений после взаимодействия опухолевых клеток с эндотелиальными клетками in vitro». Экспериментальные исследования клеток . 237 (2): 347–356. дои : 10.1006/excr.1997.3799 . hdl : 2268/61990 . ПМИД 9434630 .
- ^ Шасби Д.М., Райс Д.Р., Шасби СС, Winter MC (июнь 2002 г.). «Гистамин стимулирует фосфорилирование белков адгезионного соединения и изменяет их связь с виментином». Американский журнал физиологии. Клеточная и молекулярная физиология легких . 282 (6): Л1330–Л1338. CiteSeerX 10.1.1.1000.5266 . дои : 10.1152/ajplung.00329.2001 . ПМИД 12003790 .
- ^ Синн Х.В., Бальзамо Дж., Лилиен Дж., Лин Дж.Дж. (сентябрь 2002 г.). «Локализация нового белка Xin в комплексе слипчивых соединений в сердечной и скелетной мышце во время развития». Динамика развития . 225 (1): 1–13. дои : 10.1002/dvdy.10131 . ПМИД 12203715 . S2CID 23393425 .
Дальнейшее чтение
[ редактировать ]- Кикучи А. (февраль 2000 г.). «Регуляция передачи сигналов бета-катенина в пути Wnt». Связь с биохимическими и биофизическими исследованиями . 268 (2): 243–248. дои : 10.1006/bbrc.1999.1860 . ПМИД 10679188 .
- Уилсон П.Д. (апрель 2001 г.). «Полицистин: новые аспекты структуры, функции и регуляции» . Журнал Американского общества нефрологов . 12 (4): 834–845. дои : 10.1681/ASN.V124834 . ПМИД 11274246 .
- Каллури Р., Нилсон Э.Г. (декабрь 2003 г.). «Эпителиально-мезенхимальный переход и его значение для фиброза» . Журнал клинических исследований . 112 (12): 1776–1784. дои : 10.1172/JCI20530 . ПМК 297008 . ПМИД 14679171 .
- Де Феррари Г.В., Moon RT (декабрь 2006 г.). «Взлеты и спады передачи сигналов Wnt при распространенных неврологических расстройствах». Онкоген . 25 (57): 7545–7553. дои : 10.1038/sj.onc.1210064 . ПМИД 17143299 . S2CID 35684619 .
Внешние ссылки
[ редактировать ]- бета + Катенин в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- «Разнообразный набор белков модулирует канонический сигнальный путь Wnt/β-катенин». на сайте рака.gov
- «Роль β-катенина в передаче сигнала, определении судьбы клеток и трансдифференцировке» на сайте nih.gov.
- «Исследователи предлагают первое прямое доказательство того, как артрит разрушает хрящ» на сайте rochester.edu
- человека CTNNB1 Расположение генома CTNNB1 и страница сведений о гене в браузере генома UCSC .
Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .
галерея PDB |
|---|















