Катенин Бета-1
| Ctnnb1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | Ctnnb1 , Ctnnb, Mrd19, Armadillo, Catenin Beta 1, Evr7, NEDSDV | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Омим : 116806 ; MGI : 88276 ; Гомологен : 1434 ; GeneCards : Ctnnb1 ; OMA : Ctnnb1 - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Катенин бета-1 , также известный как β-катенин ( бета -катенин), является белком , который у людей кодируется CTNNB1 геном .
β-катенин является двойным функциональным белком , участвующим в регуляции и координации клеточной адгезии и транскрипции генов . У людей белок CTNNB1 кодируется CTNNB1 геном . [ 5 ] [ 6 ] У Drosophila гомологичный белок называется Armadillo . β-катенин является субъединицей комплекса белка кадгерина и действует как внутриклеточный сигнальный преобразователь в сигнальном пути Wnt . [ 7 ] [ 8 ] [ 9 ] Он является членом семейства белков катенина и гомологичен γ-катенину , также известному как плакоглобин . β-катенин широко экспрессируется во многих тканях. В сердечной мышцах β-катенин локализуется для прилипания соединений в интеркалированных структурах диска, которые имеют решающее значение для электрической и механической связи между соседними кардиомиоцитами .
Мутации и сверхэкспрессия β-катенина связаны со многими раковыми заболеваниями, включая гепатоцеллюлярную карциному , колоректальную карциному , рак легкого , злокачественные опухоли молочной железы , яичников и рак эндометрия . [ 10 ] Изменения в уровнях локализации и экспрессии β-катенина были связаны с различными формами сердечных заболеваний , включая дилатационную кардиомиопатию . β-катенин регулируется и разрушается комплексом разрушения бета-катенина , и, в частности, белком аденоматозного полипоза Coli (APC), кодируемого APC геном , подавляющей опухоль . Следовательно, генетическая мутация гена APC также тесно связана с раком, и, в частности, колоректальный рак в результате семейного аденоматозного полипоза (FAP).
Открытие
[ редактировать ]β-катенин был первоначально обнаружен в начале 1990-х годов как компонент комплекса адгезии клеток млекопитающих : белок, ответственный за цитоплазматическое закрепление кадгеринов . [ 11 ] Но очень скоро было понято, что белок Drosophila Armadillo , участвуя в опосредовании морфогенных эффектов безвеса/Wnt , является гомологичен β-катенину млекопитающего не только по структуре, но и по функции. [ 12 ] Таким образом, β-катенин стал одним из первых примеров лунного освещения : белка, выполняющего более чем одну радикально отличающуюся клеточную функцию.
Структура
[ редактировать ]Белковая структура
[ редактировать ]Ядро β-катенина состоит из нескольких очень характерных повторов , каждый из которых длится около 40 аминокислот. Называемые повторяющимися Armadillo , все эти элементы объединяются в один жесткий белковый домен с удлиненной формой - называемый домен Armadillo (ARM). Средний повтор -армадилло состоит из трех альфа -спиралей . Первый повтор β-катенина (около N-конца) немного отличается от других-поскольку он имеет удлиненную спираль с изломом, образованной слиянием спиралей 1 и 2. [ 13 ] Из -за сложной формы отдельных повторений весь домен ARM не является прямым стержнем: он обладает небольшой кривизны, так что образуется внешняя (выпуклая) и внутренняя (вогнутая) поверхность. Эта внутренняя поверхность служит лиганд-связывающим сайтом для различных партнеров по взаимодействию доменов ARM.

Сегменты N-концевой и далеко C-терминал для домена ARM не используют какую-либо структуру в решении сами по себе. Тем не менее, эти внутренние неупорядоченные области играют решающую роль в функции β-катенина. N-концевая неупорядоченная область содержит консервативный короткий линейный мотив, ответственный за связывание TRCP1 (также известного как β-TRCP) убиквитин-лигаза E3 -но только тогда, когда она фосфорилирована . Таким образом, деградация β-катенина опосредуется этим N-концевым сегментом. C-концевая область, с другой стороны, является сильным трансактиватором при наборе на ДНК . Этот сегмент не является полностью неупорядоченным: часть С-терминального расширения образует стабильную спираль , которая упакована с доменом ARM, но также может задействовать отдельных партнеров по связыванию. [ 14 ] Этот небольшой структурный элемент (Helixc) ограничивает С-концевой конец домена ARM, защищая его гидрофобные остатки. Helixc не требуется для β-катенина функционировать в клеточной адгезии. С другой стороны, это требуется для передачи сигналов Wnt: возможно, для набора различных коактиваторов, таких как 14-3-3Zeta. [ 15 ] Тем не менее, его точные партнеры среди общих комплексов транскрипции по-прежнему не полностью поняты, и они, вероятно, связаны с тканеспецифическими игроками. [ 16 ] Примечательно, что С-концевой сегмент β-катенина может имитировать эффекты всего пути Wnt, если искусственно сливается с ДНК-связывающим доменом фактора транскрипции LEF1 . [ 17 ]
Плакоглобин (также называемый γ-катенин) имеет поразительно сходную архитектуру с архитектурой β-катенина. Не только их домены ARM напоминают друг друга как в способности к связыванию архитектуры, так и в лиганде, но и N-концевой β-TRCP-связывающий мотив также сохраняется в плакоглобине, подразумевая общее происхождение и общую регуляцию с β-катенином. [ 18 ] Тем не менее, плакоглобин является очень слабым трансактиватором, когда он связан с ДНК-вероятно, это вызвано дивергенцией их С-концевых последовательностей (плакоглобин, по-видимому, отсутствует в трансактиваторных мотивах и, таким образом, ингибирует гены-мишени пути Wnt вместо их активации). [ 19 ]
Партнеры, связывающие с доменом Armadillo
[ редактировать ]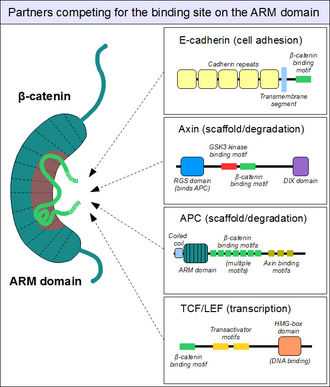
Как указано выше, домен ARM β-катенина действует как платформа, с которой могут связываться конкретные линейные мотивы . Расположенные в структурно разнообразных партнерах, мотивы связывания β-катенина, как правило , расстройства самостоятельно и обычно принимают жесткую структуру при взаимодействии с доменом ARM-как видно на коротких линейных мотивах . Однако взаимодействующие мотивы β-катенина также имеют ряд особых характеристик. Во -первых, они могут достигать или даже превзойти длину 30 аминокислот в длину, и контактировать с доменом руки на чрезмерно большой площади поверхности. Другой необычной особенностью этих мотивов является их часто высокая степень фосфорилирования . Такие события фосфорилирования Ser / Thr значительно усиливают связывание многих β-катенинов, связанных с доменом ARM. [ 20 ]
Структура β-катенина в комплексе с доменом связывания катенина у партнера транскрипционной трансактивации TCF обеспечила начальную структурную дорожную карту того, сколько связывающих партнеров β-катенина может образовывать взаимодействия. [ 21 ] Эта структура продемонстрировала, как в противном случае неупорядоченный N-конец TCF адаптировал то, что казалось жесткой конформацией, причем мотива связывания охватывает многие повторения бета-катенина. Относительно сильное заряженное взаимодействие «горячие точки» были определены (прогнозируемые и позже проверенные, чтобы быть консервативным для взаимодействия β-катенина/E-кадгерина), а также гидрофобные области, считающиеся важными в общем моде связывания и как потенциальные терапевтические малые Молекула ингибитор нацелен на определенные формы рака. Кроме того, после исследований продемонстрировали еще одну особенную характеристику, пластичность в связывании N-конце TCF с бета-катенином. [ 22 ] [ 23 ]
Точно так же мы находим знакомый e-кадгерин , чей цитоплазматический хвост контактирует с доменом ARM таким же каноническим способом. [ 24 ] Аксин белка каркаса (два тесно связанных паралога, аксин 1 и аксин 2 ) содержит сходную мотив взаимодействия на его длинном, неупорядоченном среднем сегменте. [ 25 ] Хотя одна молекула аксина содержит только один мотив рекрутирования β-катенина, его партнер- белок аденоматозного полипоза Coli (APC) содержит 11 таких мотивов в тандемном расположении на каждом протомере, таким образом, способным взаимодействовать с несколькими молекулами β-катенина одновременно. [ 26 ] Поскольку поверхность домена ARM обычно может вместить только один пептидный мотив в любое время, все эти белки конкурируют за один и тот же клеточный пул молекул β-катенина. Этот конкурс является ключом к пониманию того, как работает сигнальный путь Wnt .
Однако этот «основной» сайт связывания на домене ARM β-катенин ни в коем случае не является единственным. Первые спирали домена ARM образуют дополнительный, специальный белок-белковой взаимодействие: это может приспособить линейный мотив, образующий спираль, обнаруженный в коактиваторе BCL9 (или тесно связанном BCL9L )-важный белок, участвующий в передаче сигналов Wnt. [ 27 ] Хотя точные детали гораздо менее ясны, кажется, что тот же сайт используется альфа-катенином, когда β-катенин локализуется в соединениях адгезивы. [ 28 ] Поскольку этот карман отличается от «основного» сайта связывания домена ARM, между альфа-катенином и E-кадгерином нет конкуренции между TCF1 и BCL9, соответственно. [ 29 ] С другой стороны, BCL9 и BCL9L должны конкурировать с α-катенином, чтобы получить доступ к молекулам β-катенина. [ 30 ]
Функция
[ редактировать ]Регуляция деградации посредством фосфорилирования
[ редактировать ]Клеточный уровень β-катенина в основном контролируется его убиквитинированием и протеосомной деградацией . E3 -убиквитин лигаза TRCP1 (также известный как β-TRCP) может распознавать β-катенин в качестве субстрата через короткий линейный мотив на неупорядоченном N-конце. Однако этот мотив (ASP-Ser-Gly-Ile-Ser) β-катенина должен фосфорилироваться на двух серинах , чтобы быть способным связывать β-TRCP. Фосфорилирование мотива выполняется с помощью альфа и бета -гликогена -синтазы 3 (GSK3α и GSK3β). GSK3 являются конститутивно активными ферментами, вовлеченными в несколько важных регуляторных процессов. Однако есть одно требование: субстраты GSK3 должны быть предварительно фосфорилированными четырьмя аминокислотами вниз по течению (C-концевому) фактического целевого сайта. Таким образом, это также требует «заполнительной киназы» для своей деятельности. В случае β-катенина наиболее важной прайминговой киназой является казеинкиназа I (CKI). После того, как насыщенный серино-тиониновый субстрат был «заполнен», GSK3 может «пройти» через него от С-концевого до N-концевого направления, фосфорилируя каждый 4-й серин Треониновые остатки подряд. Этот процесс приведет к двойному фосфорилированию вышеупомянутого мотива распознавания β-TRCP.
Комплекс разрушения бета-катенина
[ редактировать ]Чтобы GSK3 была высокоэффективной киназой на субстрате, предварительное фосфорилирование недостаточно. Существует еще одно требование: аналогично митогеном , активируемым протеинкиназам (MAPK), субстратам необходимо ассоциироваться с этим ферментом с помощью высокоаффинных стыковочных мотивов . β-катенин не содержит таких мотивов, но специальный белок делает: Axin . Более того, его мотив стыковки GSK3 непосредственно примыкает к мотиву связывания β-катенина. [ 25 ] Таким образом, Axin действует как истинный белок каркаса , приводя фермент (GSK3) вместе с его субстратом (β-катенин) в близкую физическую близость.

Но даже Аксин не действует в одиночку. Через свой N-концевой регулятор домена передачи сигналов G-белка (RGS), он рекрутирует белок аденоматозного полипоза Coli (APC). APC похож на огромную «рождественскую елку»: с множеством мотивов связывания β-катенина (одна только молекула APC имеет 11 таких мотивов [ 26 ] ), он может собирать как можно больше молекул β-катенина. [ 31 ] APC может взаимодействовать с множественными молекулами аксина в то же время, когда он имеет три мотива SAMP (Ser-Ala-Met-Pro), чтобы связывать домены RGS, обнаруженные в Axin . Кроме того, Axin также имеет потенциал для олигомеризации через свой C-концевой домен DIX. Результатом является огромная мультимерная белковая сборка, посвященная фосфорилированию β-катенина. Этот комплекс обычно называется комплексом разрушения бета-катенина , хотя он отличается от протеосомного механизма, фактически ответственного за деградацию β-катенина. [ 32 ] Это отмечает только молекулы β-катенина для последующего разрушения.
Передача сигналов Wnt и регулирование разрушения
[ редактировать ]В покоящихся клетках молекулы аксин олигомеризуются друг с другом через свои C-концевые домены DIX, которые имеют два связывающих интерфейса. Таким образом, они могут строить линейные олигомеры или даже полимеры внутри цитоплазмы клеток. Домены DIX уникальны: единственные другие белки, которые, как известно, имеют домен DIX, растрепаны и DIXDC1 . (Одиночный DSH белок Drosophila соответствует трем паралогическим генам, DVL1 , DVL2 и DVL3 у млекопитающих .) DSH ассоциируется с цитоплазматическими областями Frizzled рецепторов с его PDZ и DEP -доменами . Когда молекула Wnt связывается с Frizzled , она вызывает плохо известный каскад событий, что приводит к воздействию домена Disheveled Dix и создания идеального сайта связывания для Axin . Затем аксин титруют от своих олигомерных сборков-комплекса разрушения β-катенина-от DSH . [ 33 ] После того, как он связан с рецепторным комплексом, аксин будет становляться некомпетентным для связывания β-катенина и активности GSK3. цитоплазматические сегменты белков LRP5 и LRP6 , ассоциированных с Важно отметить, что Frizzled Это был настоящий субстрат GSK3. Эти ложно -целевые сайты значительно ингибируют активность GSK3 конкурентоспособной. [ 34 ] Этот способ, связанный с рецептором, отменит опосредование фосфорилирования β-катенина. Поскольку β-катенин больше не обозначен для разрушения, но продолжает производиться, его концентрация будет увеличиваться. Как только уровни β-катенина повышаются достаточно высоко, чтобы насытить все сайты связывания в цитоплазме, он также будет транслироваться в ядро. После участия факторов транскрипции LEF1 , TCF1 , TCF2 или TCF3 , β-катенин заставляет их отключить своих предыдущих партнеров: белки Groucho. В отличие от Groucho , рекрутирующие транскрипционные репрессоры (EG гистон-лизин метилтрансферазы ), β-катенин будет связывать активаторы транскрипции , включив гены-мишени.
Роль в клеточной адгезии
[ редактировать ]
Клеточно -клеточные комплексы адгезионных комплексов необходимы для образования сложных тканей животных. β-катенин является частью белкового комплекса , который образует прилипшие соединения . [ 35 ] Эти клеточные комплексы адгезионной адгезии необходимы для создания и поддержания слоев эпителиальных клеток и барьеров. В качестве компонента комплекса β-катенин может регулировать рост клеток и адгезию между клетками. Он также может быть ответственен за передачу сигнала ингибирования контакта, который заставляет клетки прекращать деление после завершения эпителиального листа. [ 36 ] Комплекс E-Cadherin-β-катенин-α-катенин слабо связан с актиновыми филаментами . Прилейные соединения требуют значительной динамики белка для связи с актиновым цитоскелетом, [ 35 ] тем самым обеспечивает механотрансдукцию . [ 37 ] [ 38 ]
Важным компонентом соединений прилипания являются белки кадгерина. Кадгерины образуют клеточные соединительные структуры, известные как соединения адгезив, а также десмосомы . Кадгерины способны на гомофильные взаимодействия через свои внеклеточные кадгеринские повторные домены в зависимости от Ca2+; Это может удерживать смежные эпителиальные клетки вместе. В то время как в соединении прилипания кадгерины рекрутируют молекулы β-катенина в их внутриклеточные области [ нужно разъяснения ] Анкет β-катенин, в свою очередь, ассоциируется с другим высоко динамическим белком, α-катенином , который напрямую связывается с актиновыми филаментами. [ 39 ] Это возможно, потому что α-катенин и кадгерины связываются в различных сайтах с β-катенином. [ 40 ] Таким образом, комплекс β-катенина-α-катенина может физически образовывать мост между кадгеринами и актиновым цитоскелетом . [ 41 ] Организация комплекса кадгерина -катенина дополнительно регулируется посредством фосфорилирования и эндоцитоза его компонентов. [ Цитация необходима ]
Роли в разработке
[ редактировать ]β-катенин играет центральную роль в направлении нескольких процессов развития, поскольку он может напрямую связывать факторы транскрипции и регулироваться диффузируемым внеклеточным веществом: Wnt. Он действует на ранние эмбрионы, чтобы вызвать целые области тела, а также отдельные клетки на более поздних стадиях развития. Он также регулирует физиологические процессы регенерации.
Раннее эмбриональное паттерн
[ редактировать ]Передача сигналов Wnt и β-катенин-зависимая экспрессия генов играет критическую роль во время образования различных областей тела в раннем эмбрионе. Экспериментально модифицированные эмбрионы, которые не экспрессируют этот белок, не будут развивать мезодерму и инициировать гаструляцию . [ 42 ] Спецификация эндомезодермы ранних эмбрионов также включает в себя активацию β-катенина-зависимой транскрипиональной активности первыми морфогенетическими движениями эмбриогенеза, хотя и процессов механотрансдукции. Эта особенность, которую разделяют позвоночные и членистоногие Bilateria, и Cnidaria, как было предложено, было эволюционно унаследовано от его возможного участия в спецификации эндомезодермы первого метазоа. [ 43 ] [ 44 ] [ 45 ]
Во время стадий бластулы и гаструлы, пути Wnt , а также BMP и FGF будут вызывать образование оси передне-перськой ) [ 46 ]
В ооцитах Xenopus β-катенин первоначально одинаково локализуется во всех областях яйца, но он нацелен на убиквитинирование и деградацию с помощью комплекса разрушения β-катенина. Оплодотворение яйца вызывает вращение наружных слоев коры, движущиеся кластеры белков вьющихся и DSH ближе к экваториальной области. β-катенин будет обогащен локально под влиянием сигнального пути Wnt в клетках, которые наследуют эту часть цитоплазмы. В конечном итоге он будет транслоцироваться в ядро для связывания TCF3 , чтобы активировать несколько генов, которые вызывают характеристики дорсальных клеток. [ 47 ] Эта сигнализация приводит к области клеток, известной как серой полумесяц, которая является классическим организатором эмбрионального развития. Если эта область хирургически удалена из эмбриона, гаструляция вообще не происходит. β-катенин также играет решающую роль в индукции губы Blastopore , которая, в свою очередь, инициирует гаструляцию. [ 48 ] Ингибирование трансляции GSK-3 путем инъекции антисмысловой мРНК может привести к образованию второй бластопоры и лишней оси тела. Аналогичный эффект может быть результатом сверхэкспрессии β-катенина. [ 49 ]
Асимметричное деление клеток
[ редактировать ]β-катенин также участвует в регуляции клеточных существ через асимметричное деление клеток в модельном организме C. elegans . Подобно ооцитам Xenopus , это по сути является результатом неравного распределения DSH , Frizzled , Axin и APC в цитоплазме материнской клетки. [ 50 ]
Обновление стволовых клеток
[ редактировать ]Одним из наиболее важных результатов передачи сигналов Wnt и повышенного уровня β-катенина в определенных типах клеток является поддержание плюрипотентности . [ 46 ] Скорость стволовых клеток в толстой кишке, например, обеспечивается таким накоплением β-катенина, который может стимулировать путем Wnt. [ 51 ] Высокочастотные перистальтические механические штаммы толстой кишки также участвуют в β-катенино-зависимом поддержании гомеостатических уровней стволовых клеток толстой кишки посредством процессов механотрансдукции. Эта особенность патологически усиливается в направлении онкогенной гиперпролиферации в здоровых клетках, сжатых путем давления, полученного генетически измененным гиперпролиферативным опухолевым клетками. [ 52 ]
У других типов клеток и стадий развития β-катенин может способствовать дифференцировке , особенно в отношении мезодермальных линий клеток.
Эпителиально-мезенхимальный переход
[ редактировать ]β-катенин также действует как морфоген на более поздних стадиях эмбрионального развития. Вместе с TGF-β важной ролью β-катенина является индукция морфогенного изменения в эпителиальных клетках. Это побуждает их отказаться от их жесткой адгезии и предполагать более подвижный и слабо связанный мезенхимальный фенотип. Во время этого процесса эпителиальные клетки теряют экспрессию белков, таких как E-Cadherin , Zonula Occludens 1 (ZO1) и цитокератин . В то же время они включают экспрессию виментина , альфа-актина гладких мышц (ACTA2) и фибробласто-специфического белка 1 (FSP1). Они также производят компоненты внеклеточного матрикса, такие как коллаген I типа и фибронектин . Аберрантная активация пути Wnt была вовлечена в патологические процессы, такие как фиброз и рак. [ 53 ] При развитии сердечных мышц β-катенин выполняет двухфазную роль. Первоначально активация Wnt/β-катенина необходима для совершения мезенхимальных клеток в линию сердца; Однако на более поздних стадиях развития требуется подавление β-катенина. [ 54 ] [ 55 ] [ 42 ]
Участие в физиологии сердца
[ редактировать ]В сердечной мышцах β-катенин образует комплекс с N-кадгерином в прилипших соединениях в интеркалированных структурах диска, которые отвечают за электрическое и механическое соединение смежных сердечных клеток. Исследования по модели желудочков кардиомиоцитов взрослых крыс показали, что появление и распределение β-катенина пространственно-временно регулируется во время редактификации этих клеток в культуре. В частности, β-катенин является частью отдельного комплекса с N-кадгерином и альфа-катенином , который в изобилии в соединениях приклеивает на ранних стадиях после выделения кардиомиоцитов для реформации контактов клеток-клеток. [ 56 ] Было показано, что β-катенин образует комплекс с эмерин в кардиомиоцитах в соединениях адгезив в интеркалированных дисках; и это взаимодействие зависит от наличия 3-бета GSK сайтов фосфорилирования на β-катенине. Выбил эмерин значительно изменил локализацию β-катенина и общую интеркалированную архитектуру диска, которая напоминала расширенную фенотип кардиомиопатии . [ 57 ]
На животных моделях сердечного заболевания были представлены функции β-катенина. В модели гвинеи стеноза аорты и гипертрофии левого желудочка , как было показано, β-катенин изменяет субклеточную локализацию от интеркалированных дисков в цитозоль , несмотря на отсутствие изменений в общей численности клеток β-катенина. Винкулин показал аналогичный профиль изменений. N-кадгерин не показал изменений, и не было компенсаторной активации плакоглобина на интеркалированных дисках в отсутствие β-катенина. [ 58 ] В модели хомяка кардиомиопатии и сердечной недостаточности клеточные адгезии были нерегулярными и дезорганизованными, а уровни экспрессии приклеенного соединения/интеркалированного диска и ядерных пулов β-катенина были снижены. [ 59 ] Эти данные свидетельствуют о том, что потеря β-катенина может играть роль в больных интеркалированных дисках, которые были связаны с гипертрофией мышц сердца и сердечной недостаточностью. В крысиной модели миокарда инфаркта аденовирусного перенос гена не фосфорилатируемого , конститутивно-активного β-катенина уменьшал размер ИМ, активировал клеточный цикл и снижал количество апоптоза в кардиомиоцитах и сердечных миофибробластах . Это открытие было координировано с повышенной экспрессией протеинов проживания, выживших и BCL-2 и фактора роста эндотелиального эндотелиального эндотелиального фактора , одновременно способствуя дифференцировке сердечных фибробластов в миофибробласты. Эти данные свидетельствуют о том, что β-катенин может способствовать процессу регенерации и заживления после инфаркта миокарда. [ 60 ] В модели самопроизвольно -гипертонической крысы сердечной недостаточности исследователи обнаружили шаттлинг β-катенина из интеркалированного диска/ сарколеммы в ядро , о чем свидетельствует снижение экспрессии β-катенина во фракции мембранного белка и увеличением фракции ядер. Кроме того, они обнаружили ослабление в ассоциации между гликогеновой синтазойкиназой-3β и β-катенином, что может указывать на измененную стабильность белка. В целом, результаты показывают, что усиленная ядерная локализация β-катенина может быть важной для прогрессирования гипертрофии сердца . [ 61 ]
Что касается механистической роли β-катенина в гипертрофии сердца, исследования трансгенных мышей показали несколько противоречивых результатов относительно того, является ли повышенная регуляция β-катенина полезной или вредной. [ 62 ] [ 63 ] [ 64 ] Недавнее исследование с использованием условной нокаутирующей мыши, у которой либо отсутствовали β-катенин, либо экспрессировали неразлагаемую форму β-катенина в кардиомиоцитах, согласовало потенциальную причину этих расхождений. По-видимому, существует строгий контроль над субклеточной локализацией β-катенина в сердечной мышце. левого желудочка У мышей, у которых отсутствовал β-катенин, не было явного фенотипа в миокарде ; Тем не менее, мыши, несущие стабилизированную форму β-катенина, развивали дилатационную кардиомиопатию , что позволяет предположить, что временная регуляция β-катенина с помощью механизмов деградации белка является критической для нормального функционирования β-катенина в клетках сердца. [ 65 ] В мышиной модели, несущей нокаут десмосомного белка, плакоглобина, участвующего в аритмогенной кардиомиопатии правого желудочка , стабилизация β-катенина также была увеличена, предположительно, чтобы компенсировать потерю своего гомолога плакоглобина. Эти изменения были координированы с активацией Akt и ингибированием гликогенсинтазы киназы 3β , что еще раз предполагает, что аномальная стабилизация β-катенина может быть вовлечена в развитие кардиомиопатии. [ 66 ] В дальнейших исследованиях с использованием двойного нокаута плакоглобина и β-катенина показали, что у двойного нокаута развилась кардиомиопатия, фиброз и аритмии, что приводит к внезапной сердечной смерти . Интеркалированная архитектура диска была серьезно нарушена, и коннексине 43 в разрывы были заметно уменьшены. Измерения электрокардиограммы захватили спонтанные аритмии летальных желудочков у двойных трансгенных животных, что позволяет предположить, что два катенина-β-катенин и плакоглобин-являются критическими и необходимы для механоэлектрической связи в кардиомиоцитах. [ 67 ]
Клиническое значение
[ редактировать ]Роль в депрессии
[ редактировать ]Может ли данные мозга данного человека эффективно справляться со стрессом, и, следовательно, их восприимчивость к депрессии зависит от β-катенина в мозге каждого человека, согласно исследованию, проведенному в Медицинской школе ИКАНА на горе Синай и опубликовано 12 ноября, 2014, в журнале Nature . [ 68 ] Более высокая передача сигналов β-катенина повышает гибкость поведения, тогда как дефектная передача сигналов β-катенина приводит к депрессии и снижению лечения стресса. [ 68 ]
Роль в сердечных заболеваниях
[ редактировать ]Измененные профили экспрессии в β-катенине были связаны с дилатационной кардиомиопатией у людей. Повышенная регуляция экспрессии β-катенина обычно наблюдалась у пациентов с дилатационной кардиомиопатией. [ 69 ] В конкретном исследовании пациенты с дилатано-кардиомиопатией на конечной стадии показали почти удвоенные рецептора эстрогена (ER-альфа) мРНК-мРНК и уровни белка , а также взаимодействие ER-альфа/бета-катенин, присутствующее на интеркалированных дисках контроля, не добываемого человека. Сердцы были потеряны, предполагая, что потеря этого взаимодействия на интеркалированном диске может сыграть роль в прогрессировании сердечной недостаточности. [ 70 ] Вместе с белками Bcl9 и Pygo β -катенин координирует различные аспекты развития слышно, а мутации в Bcl9 или Pygo в модельных организмах, таких как мышь и рыбок данио - вызывают фенотипы, которые очень похожи на человеческие врожденные сердечные расстройства . [ 71 ]
Участие в раке
[ редактировать ]
β-катенин является протоонкогеном . Мутации этого гена обычно встречаются при различных раковых заболеваниях: при первичной гепатоцеллюлярной карциноме , колоректальном раке , раке яичника , раке молочной железы , раке легких и глиобластоме . Было подсчитано, что приблизительно 10% всех образцов ткани, секвенированных от всех раковых заболеваний, демонстрируют мутации в гене CTNNB1. [ 72 ] Большинство из этих мутаций кластер в крошечной области N-концевого сегмента β-катенина: мотив связывания β-TRCP. Мутации потери функции этого мотива по существу делают невозможным убиквитинилирование и деградацию β-катенина. Это приведет к транспортировке β-катенина в ядро без какого-либо внешнего стимула и непрерывно управлять транскрипцией его генов-мишеней. Повышенные уровни ядерного β-катенина также были отмечены при базально-клеточной карциноме (BCC), [ 73 ] Голова и шея плоскоклеточная карцинома (HNSCC), рак предстательной железы (CAP), [ 74 ] Пиломатриксома (PTR) [ 75 ] и медуллобластома (MDB) [ 76 ] Эти наблюдения могут или не могут влиять на мутацию в гене β-катенина: другие компоненты пути Wnt также могут быть неисправными.



Подобные мутации также часто встречаются в мотивах рекрутинга β-катенина APC . Наследственные мутации APC потери функции вызывают состояние, известное как семейный аденоматозный полипоз . Пострадавшие люди развивают сотни полипов в своей большой кишке. Большинство из этих полипов являются доброкачественными по своей природе, но они могут превратиться в смертельный рак с течением времени. Соматические мутации APC при колоректальном раке также не редкость. [ 80 ] β-катенин и APC являются одними из ключевых генов (вместе с другими, такими как K-Ras и Smad4 ), участвующие в развитии колоректального рака. Потенциал β-катенина для изменения ранее эпителиального фенотипа пораженных клеток в инвазивный, мезенхима, подобный типу, вносит значительный вклад в образование метастазирования.
Как терапевтическая цель
[ редактировать ]Из-за участия в развитии рака ингибирование β-катенина продолжает получать значительное внимание. Но нацеливание на сайт связывания на его домене Armadillo не является самой простой задачей из -за его обширной и относительно плоской поверхности. Однако для эффективного ингибирования достаточное связывание с более мелкими «горячими точками» этой поверхности. Таким образом, «сшитый» спиральный пептид, полученный из природного мотива связывания β-катенина, обнаруженного в LEF1, был достаточным для полного ингибирования β-катенина, зависящей от транскрипции. Недавно было также разработано несколько небольших соединений для нацеливания на ту же самую положительно заряженную область домена ARM (CGP049090, PKF118-310, PKF115-584 и ZTM000990). Кроме того, на уровне β-катенина также могут влиять нацеливание вверх по течению компонентов пути Wnt, а также комплекс разрушения β-катенина. [ 81 ] Дополнительный N-концевой карман связывания также важен для активации гена-мишени Wnt (необходимого для рекрутирования BCL9). этот сайт домена ARM может быть фармакологически нацелен на карносовую кислоту . Например, [ 82 ] Этот «вспомогательный» сайт является еще одной привлекательной целью для разработки лекарств. [ 83 ] Несмотря на интенсивные доклинические исследования, в качестве терапевтических агентов пока нет ингибиторов β-катенина. Однако его функция может быть дополнительно изучена с помощью нокдауна siRNA на основе независимой валидации. [ 84 ] Другим терапевтическим подходом к снижению накопления ядер β-катенина является ингибирование галектина-3. [ 85 ] Ингибитор галектина-3 GR-MD-02 в настоящее время проходит клинические испытания в сочетании с одобренной FDA дозой ипилимумаба у пациентов, имеющих продвинутую меланому. [ 86 ] Белки BCL9 и BCL9L были предложены в качестве терапевтических мишеней для колоректального рака, которые представляют гиперактивированную передачу сигналов Wnt, поскольку их делеция не нарушает нормальный гомеостаз, но сильно влияет на поведение метастазов . [ 87 ]
Роль в синдроме алкоголя плода
[ редактировать ]Дестабилизация β-катенина с помощью этанола является одним из двух известных путей, посредством которых воздействие алкоголя индуцирует синдром алкоголя плода (другой-индуцированный этанолом дефицит фолата). Этанол приводит к дестабилизации β-катенина через G-белок-зависимый путь, в котором активировал фосфолипазу Cβ-гидролизу, фосфатидилинозитол- (4,5) -бисфосфат в диацилглицерище и иннозитол- (1,4,5) -трисфосфат. Растворимый иннозитол- (1,4,5) -трисфосфатный триггер выпускает из эндоплазматического ретикулума. Это внезапное увеличение цитоплазматического кальция активирует Ca2+/кальмодулин-зависимую протеинкиназу (CAMKII). Активированный CAMKII дестабилизирует β-катенин через плохо охарактеризованный механизм, но который, вероятно, включает фосфорилирование β-катенина CAMKII. Тем самым подавляется транскрипционная программа β-катенина (которая необходима для нормального развития клеток нервного гребня), что приводит к апоптозу клеток клеток нейронного гребня (гибель клеток). [ 88 ]
Взаимодействия
[ редактировать ]Было показано, что β-катенин взаимодействует :
- APC , [ 89 ] [ 90 ] [ 91 ] [ 92 ] [ 93 ] [ 94 ] [ 95 ] [ 96 ]
- Axin1 , [ 97 ] [ 98 ]
- Андрогенный рецептор , [ 99 ] [ 100 ] [ 101 ] [ 102 ] [ 103 ] [ 104 ]
- Cby1 , [ 105 ]
- CDH1 , [ 24 ] [ 90 ] [ 106 ] [ 107 ] [ 108 ] [ 109 ] [ 110 ] [ 111 ] [ 112 ] [ 113 ] [ 114 ] [ 115 ] [ 116 ] [ 117 ] [ 118 ] [ 119 ] [ 120 ] [ 121 ] [ 122 ] [ 123 ] [ 124 ] [ 125 ] [ 126 ]
- CDH2 , [ 56 ] [ 127 ] [ 128 ]
- CDH3 , [ 125 ] [ 129 ]
- Cdk5r1 , [ 130 ]
- Чук , [ 131 ]
- Ctnnd1 , [ 90 ] [ 111 ]
- Ctnna1 , [ 107 ] [ 116 ] [ 132 ] [ 133 ] [ 134 ]
- EGFR , [ 111 ] [ 120 ] [ 135 ]
- Эмери [ 136 ] [ 137 ]
- ESR1 [ 70 ]
- Fl2 , [ 138 ]
- GSK3B , [ 92 ] [ 139 ]
- HER2/или , [ 112 ] [ 135 ] [ 140 ]
- Hhnf4a , [ 103 ]
- Ikk2 , [ 131 ]
- LEF1 [ 141 ] [ 142 ] [ 143 ] [ 144 ] в том трансгенный числе [ 145 ]
- Magi1 , [ 121 ]
- Muc1 , [ 113 ] [ 146 ] [ 147 ] [ 148 ] [ 149 ] [ 150 ] [ 151 ]
- NR5A1 , [ 152 ] [ 153 ]
- PCAF , [ 154 ]
- PHF17 , [ 155 ]
- Плакоглобин , [ 90 ] [ 111 ]
- Ptpn14 , [ 156 ]
- Ptprf , [ 112 ] [ 157 ]
- Ptprk (ptpkappa), [ 158 ]
- Ptprt (ptprho), [ 159 ]
- PTPRU (PCP-2), [ 160 ] [ 161 ] [ 162 ]
- Psen1 , [ 163 ] [ 164 ] [ 165 ]
- PTK7 [ 166 ]
- Ruvb-like 1 , [ 167 ]
- Smad7 , [ 141 ]
- SMATRCA4 [ 168 ]
- SLC9A3R1 , [ 115 ]
- Usp9x , [ 169 ] и
- Ve-Cadherin . [ 170 ] [ 171 ]
- XIRP1 [ 172 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в GRCH38: Ensembl Release 89: ENSG00000168036 - Ensembl , май 2017 г.
- ^ Jump up to: а беременный в GRCM38: Ensembl Release 89: Ensmusg00000006932 - Ensembl , май 2017 г.
- ^ «Человеческая PubMed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «Мышь Pubmed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ Kraus C, Liehr T, Hülsken J, Behrens J, Birchmeier W, Grzeschik KH, Ballhausen WG (сентябрь 1994). «Локализация гена бета-катенина человека (CTNNB1) до 3P21: область, вовлеченная в развитие опухоли». Геномика . 23 (1): 272–274. doi : 10.1006/geno.1994.1493 . PMID 7829088 .
- ^ Макдональд Б.Т., Тамай К, Хе Х (июль 2009 г.). «Сигнализация Wnt/Beta-Catenin: компоненты, механизмы и заболевания» . Ячейка развития . 17 (1): 9–26. doi : 10.1016/j.devcel.2009.06.016 . PMC 2861485 . PMID 19619488 .
- ^ Peifer M, Rauskolb C, Williams M, Riggleman B, Wieschaus E (апрель 1991 г.). «Ген сегмента полярности Armadillo взаимодействует с бесценным сигнальным путем в формировании эмбриональных и взрослых рисунков». Разработка . 111 (4): 1029–1043. doi : 10.1242/dev.111.4.1029 . PMID 1879348 .
- ^ NORODERMEER J, Klingensmith J, Perrimon N, Nusse R (январь 1994 г.). «Разновидные и Armadillo действуют в бесценном сигнальном пути у дрозофилы». Природа . 367 (6458): 80–83. Bibcode : 1994natur.367 ... 80n . doi : 10.1038/367080a0 . PMID 7906389 . S2CID 4275610 .
- ^ Peifer M, Berg S, Reynolds AB (март 1994 г.). «Повторяющийся аминокислотный мотив, разделяемый белками с разнообразными клеточными ролями». Клетка . 76 (5): 789–791. doi : 10.1016/0092-8674 (94) 90353-0 . PMID 7907279 . S2CID 26528190 .
- ^ Морин П.Дж. (декабрь 1999). «Передача сигналов и рак бета-катенина». Биологии . 21 (12): 1021–1030. doi : 10.1002/(SICI) 1521-1878 (199912) 22: 1 <1021 :: AID-bies6> 3.0.co; 2-p . PMID 10580987 . S2CID 86240312 .
- ^ McCrea PD, Turck CW, Gumbiner B (ноябрь 1991). «Гомолог белка Armadillo у дрозофилы (плакоглобин), связанный с E-кадгерином». Наука . 254 (5036): 1359–1361. Bibcode : 1991sci ... 254.1359M . doi : 10.1126/science.1962194 . PMID 1962194 .
- ^ Кемлер Р. (сентябрь 1993). «От кадгеринов до катенинов: взаимодействие цитоплазматического белка и регуляция клеточной адгезии». Тенденции в генетике . 9 (9): 317–321. doi : 10.1016/0168-9525 (93) 90250-л . PMID 8236461 .
- ^ Готтарди CJ, Peifer M (март 2008 г.). «Терминальные области бета-катенина появляются» . Структура 16 (3): 336–338. doi : 10.1016/j.str.2008.02.005 . PMC 2329800 . PMID 18334207 .
- ^ Xing Y, Takemaru K, Liu J, Berndt JD, Zheng JJ, Moon Rt, Xu W (март 2008 г.). «Кристаллическая структура полного бета-катенина» . Структура 16 (3): 478–487. doi : 10.1016/j.str.2007.12.021 . PMC 4267759 . PMID 18334222 .
- ^ Fang D, Hawke D, Zheng Y, Xia Y, Meisenhelder J, Nika H, et al. (Апрель 2007 г.). «Фосфорилирование бета-катенина AKT способствует транскрипционной активности бета-катенина» . Журнал биологической химии . 282 (15): 11221–11229. doi : 10.1074/jbc.m611871200 . PMC 1850976 . PMID 17287208 .
- ^ Söderholm S, Cantù C (май 2021). «Транскрипция, зависящая от Wnt/β-катенина: тканеспецифический бизнес» . Провода Механизмы болезней . 13 (3): E1511. doi : 10.1002/wsbm.1511 . PMC 9285942 . PMID 33085215 .
- ^ Vleminckx K, Kemler R, Hecht A (март 1999 г.). «С-концевой трансактивационный домен бета-катенина необходим и достаточен для передачи сигналов комплексом LEF-1/бета-катенина в Xenopus laevis» . Механизмы развития . 81 (1–2): 65–74. doi : 10.1016/s0925-4773 (98) 00225-1 . PMID 10330485 . S2CID 15086656 .
- ^ Садот Э., Симча I, Иваи К, Цехановер А., Гейгер Б., Бен-Зеев А (апрель 2000 г.). «Дифференциальное взаимодействие плакоглобина и β-катенина с системой убиквитин-протеасом». Онкоген . 19 (16): 1992–2001. doi : 10.1038/sj.onc.1203519 . PMID 10803460 . S2CID 2872966 .
- ^ Aktary Z, Pasdar M (2012). «Плакоглобин: роль в онкогенезе и метастазировании» . Международный журнал клеточной биологии . 2012 : 189521. DOI : 10.1155/2012/189521 . PMC 3312339 . PMID 22481945 .
- ^ Сюй В., Кимельман Д. (октябрь 2007 г.). «Механистическое понимание структурных исследований β-катенина и его партнеров по связыванию». Журнал сотовой науки . 120 (Pt 19): 3337–3344. doi : 10.1242/jcs.013771 . PMID 17881495 . S2CID 25294495 .
- ^ Грэм Т.А., Уивер С., Мао Ф., Кимельман Д., Сюй В. (декабрь 2000 г.). «Кристаллическая структура комплекса бета-катенина/TCF» . Клетка . 103 (6): 885–896. doi : 10.1016/s0092-8674 (00) 00192-6 . PMID 11136974 . S2CID 16865193 .
- ^ Грэм Т.А., Ферки Д.М., Мао Ф., Кимельман Д., Сюй В. (декабрь 2001 г.). «TCF4 может специально распознавать бета-катенин с использованием альтернативных конформаций». Природа структурная биология . 8 (12): 1048–1052. doi : 10.1038/nsb718 . PMID 11713475 . S2CID 33878077 .
- ^ Poy F, Lepourcelet M, Shivdasani RA, Eck MJ (декабрь 2001 г.). «Структура человеческого комплекса TCF4-бета-катенина». Природа структурная биология . 8 (12): 1053–1057. doi : 10.1038/nsb720 . PMID 11713476 . S2CID 24798619 .
- ^ Jump up to: а беременный Хубер Ах, Вейс Ви (май 2001 г.). «Структура бета-катенина/E-кадгеринового комплекса и молекулярная основа различного распознавания лигандов бета-катенином» . Клетка . 105 (3): 391–402. doi : 10.1016/s0092-8674 (01) 00330-0 . PMID 11348595 . S2CID 364223 .
- ^ Jump up to: а беременный Xing Y, Clements WK, Kimelman D, Xu W (ноябрь 2003 г.). «Кристаллическая структура комплекса бета-катенина/аксин предполагает механизм комплекса разрушения бета-катенина» . Гены и развитие . 17 (22): 2753–2764. doi : 10.1101/gad.1142603 . PMC 280624 . PMID 14600025 .
- ^ Jump up to: а беременный Minde DP, Anvarian Z, Rüdiger SG, Maurice MM (август 2011 г.). «Расширение расстройства: как миссенс -мутации в белке -супрессоре опухоли приводят к раку?» Анкет Молекулярный рак . 10 (1): 101. doi : 10.1186/1476-4598-10-101 . PMC 3170638 . PMID 21859464 .
- ^ Kramps T, Peter O, Brunner E, Nellen D, Frosch B, Chatterjee S, et al. (Апрель 2002 г.). «Сигнализация Wnt/без крыла требует, чтобы BCL9/опосредованная безглетками рекрутировал пигопуса в комплекс ядерного бета-катенина-TCF» . Клетка . 109 (1): 47–60. doi : 10.1016/s0092-8674 (02) 00679-7 . PMID 11955446 . S2CID 16720801 .
- ^ Pokutta S, Weis WI (март 2000 г.). «Структура димеризации и бета-катенин-связывающей области альфа-катенина» . Молекулярная клетка . 5 (3): 533–543. doi : 10.1016/s1097-2765 (00) 80447-5 . PMID 10882138 .
- ^ Sampietro J, Dahlberg CL, Cho US, Hinds TR, Kimelman D, Xu W (октябрь 2006 г.). «Кристаллическая структура комплекса бета-катенина/BCL9/TCF4» . Молекулярная клетка . 24 (2): 293–300. doi : 10.1016/j.molcel.2006.09.001 . PMID 17052462 .
- ^ Brembeck FH, Schwarz-Romond T, Bakkers J, Wilhelm S, Hammerschmidt M, Birchmeier W (сентябрь 2004 г.). «Основная роль BCL9-2 в переключении между клеями и транскрипционными функциями бета-катенина» . Гены и развитие . 18 (18): 2225–2230. doi : 10.1101/gad.317604 . PMC 517514 . PMID 15371335 .
- ^ Лю Дж., Син Ю, Хиндс Т.Р., Чжэн Дж., Сюй В. (июнь 2006 г.). «Третий 20 аминокислотный повтор является самым плотным сайтом связывания APC для бета-катенина». Журнал молекулярной биологии . 360 (1): 133–144. doi : 10.1016/j.jmb.2006.04.064 . PMID 16753179 .
- ^ Kimelman D, Xu W (декабрь 2006 г.). «Комплекс разрушения бета-катенина: понимание и вопросы с структурной точки зрения». Онкоген . 25 (57): 7482–7491. doi : 10.1038/sj.onc.1210055 . PMID 17143292 . S2CID 20529520 .
- ^ Фидлер М., Мендоза-Топаз С., Резерфорд Т.Дж., Миесчзанек Дж., Биенц М. (февраль 2011 г.). «Разновидные взаимодействия взаимодействуют с границей полимеризации домена DIX, чтобы вмешиваться в его функцию в пониженном β-катенине» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (5): 1937–1942. Bibcode : 2011pnas..108.1937f . doi : 10.1073/pnas.1017063108 . PMC 3033301 . PMID 21245303 .
- ^ Metcalfe C, Bienz M (ноябрь 2011 г.). «Ингибирование GSK3 с помощью Wnt Signalling-две контрастные модели» . Журнал сотовой науки . 124 (Pt 21): 3537–3544. doi : 10.1242/jcs.091991 . PMID 22083140 .
- ^ Jump up to: а беременный Brembeck FH, Rosário M, Birchmeier W (февраль 2006 г.). «Балансировать клеточную адгезию и передачу сигналов Wnt, ключевая роль бета-катенина». Текущее мнение в области генетики и развития . 16 (1): 51–59. doi : 10.1016/j.gde.2005.12.007 . PMID 16377174 .
- ^ «Ген Entrez: катенин (ассоциированный кадгерином белок)» .
- ^ Буш М., Алханшали Б.М., Цянь С., Стэнли К.Б., Хеллер В.Т., Мацуи Т. и др. (Октябрь 2019). «Ансамбль гибких конформаций лежит в основе механотрансдукции в комплексе адгезии катенина кадгерина» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (43): 21545–21555. BIBCODE : 2019PNAS..11621545B . doi : 10.1073/pnas.1911489116 . PMC 6815173 . PMID 31591245 .
- ^ Фернандес-Санчес мне, et. (Июль 2018). in vivo"Полем элиф 7 doi : 10.7554/elife . PMC 6053302 . PMID 30024850 .
- ^ Farago B, Nicholl Id, Wang S, Cheng X, Callaway DJ, Bu Z (март 2021 г.). «Активированное наноразмерное движение актин-связывающее домен в комплексе кадгерина Catenin, выявленное с помощью спектроскопии нейтронного спинового эха» . Труды Национальной академии наук Соединенных Штатов Америки . 118 (13): E2025012118. Bibcode : 2021pnas..11825012F . doi : 10.1073/pnas.2025012118 . PMC 8020631 . PMID 33753508 .
- ^ Нельсон WJ (апрель 2008 г.). «Регуляция клеточной адгезии комплексом кадгерина-катенина» . Биохимическое общество транзакций . 36 (Pt 2): 149–155. doi : 10.1042/bst0360149 . PMC 3368607 . PMID 18363555 .
- ^ Bienz M (январь 2005 г.). «Бета-катенин: поворот между клеточной адгезией и передачей сигналов Wnt» . Текущая биология . 15 (2): R64 - R67. Bibcode : 2005cbio ... 15..r64b . doi : 10.1016/j.cub.2004.12.058 . PMID 15668160 . S2CID 12352182 .
- ^ Jump up to: а беременный Haegel H, Larue L, Ohsugi M, Fedorov L, Herrenknecht K, Kemler R (ноябрь 1995 г.). «Отсутствие бета-катенина влияет на развитие мыши при гаструляции». Разработка . 121 (11): 3529–3537. doi : 10.1242/dev.121.11.3529 . PMID 8582267 .
- ^ Фард E (2003). «Механическая индукция поворота в drosophila foregud/stomodeal primordium» . Текущая биология . 13 (16): 1365–1377. Bibcode : 2003cbio ... 13.1365f . doi : 10.1016/s0960-9822 (03) 00576-1 . PMID 1293230 .
- ^ Brunet T, Bouclet A, Ahmadi P, Mitrossilis D, Driquez B, Brunet AC, et al. (2013). «Эволюционное сохранение ранней спецификации мезодермы с помощью механотрансдукции в Bilateria» . Природная связь . 4 : 2821. Bibcode : 2013natco ... 4.2821b . doi : 10.1038/ncomms3821 . PMC 3868206 . PMID 24281726 .
- ^ Nguyen NM, Merle T, Broders-Bondon F, Brunet AC, Battistella A, Land EB, et al. (2022). «Механо-биохимическая морская стимуляция инверсии, гаструляции и спецификации эндомезодермы в многоклеточной эукариоте» . Границы в клеточной биологии и развитии . 10 : 992371. DOI : 10.3389/fcell.2022.992371 . PMC 9754125 . PMID 36531949 .
- ^ Jump up to: а беременный Сокол Си (октябрь 2011 г.). «Поддержание плюрипотентности эмбриональных стволовых клеток с передачей сигналов Wnt» . Разработка . 138 (20): 4341–4350. doi : 10.1242/dev.066209 . PMC 3177306 . PMID 21903672 .
- ^ Schneider S, Steinbeisser H, Warga RM, Hausen P (июль 1996 г.). «Бета-катенин транслокация в ядра разграничивает дорсализирующие центры у лягушек и эмбрионов рыб» . Механизмы развития . 57 (2): 191–198. doi : 10.1016/0925-4773 (96) 00546-1 . PMID 8843396 . S2CID 12694740 .
- ^ Larabell CA, Torres M, Rowning BA, Yost C, Miller JR, Wu M, et al. (Март 1997 г.). «Создание дорсо-вентральной оси у эмбрионов Xenopus представляется ранней асимметрией в β-катенине, которые модулируются сигнальным путем Wnt» . Журнал клеточной биологии . 136 (5): 1123–1136. doi : 10.1083/jcb.136.5.1123 . PMC 2132470 . PMID 9060476 .
- ^ Келли Г.М., Эрезиилмаз Д.Ф., Луна Р.Т. (октябрь 1995). «Индукция вторичной эмбриональной оси у рыбок данио происходит после сверхэкспрессии бета-катенина» . Механизмы развития . 53 (2): 261–273. doi : 10.1016/0925-4773 (95) 00442-4 . PMID 8562427 . S2CID 14885037 .
- ^ Сава Х (2012). «Глава 3: Контроль полярности клеток и асимметричного деления у C. elegans ». В Ян Y (ред.). Планарная полярность клеток во время развития . Современные темы в биологии развития. Тол. 101. С. 55–76. doi : 10.1016/b978-0-12-394592-1.00003-x . ISBN 978-0-12-394592-1 Полем PMID 23140625 .
- ^ Баркер Н., Ван Эс Дж.Х., Куйперс Дж., Куджала П., Ван Ден Борн М., Козинсен М. и др. (Октябрь 2007 г.). «Идентификация стволовых клеток в тонкой кишке и толстой кишке с помощью маркера гена Lgr5». Природа . 449 (7165): 1003–1007. Bibcode : 2007natur.449.1003b . doi : 10.1038/nature06196 . PMID 17934449 . S2CID 4349637 .
- ^ Nguyen Ho-Bouldoires TH, Sollier K, Zamfirov L, Broders-Bondon F, Mitrossilis D, Bermeo S, et al. (Февраль 2022 г.). «Рет-киназа-опосредованная механическая индукция стволовых клеток толстой кишки путем давления роста опухоли стимулирует прогрессирование рака in vivo» . Биология связи . 5 (1): 137. DOI : 10.1038/S42003-022-03079-4 . PMC 8854631 . PMID 35177769 .
- ^ Tian X, Liu Z, Niu B, Zhang J, Tan TK, Lee Sr, et al. (2011). «Комплекс E-Cadherin/β-Catenin и эпителиальный барьер» . Журнал биомедицины и биотехнологии . 2011 : 567305. DOI : 10.1155/2011/567305 . PMC 3191826 . PMID 22007144 .
- ^ Zelarayan L, Gehrke C, Bergmann MW (сентябрь 2007 г.). «Роль бета-катенина в ремоделировании сердца взрослых» . Клеточный цикл . 6 (17): 2120–2126. doi : 10.4161/cc.6.17.4632 . PMID 17786052 .
- ^ Luckert H, Kutsch S, Kanzler B, Tamai Y, Taketo MM, Kemler R (август 2002 г.). «Образование множественных сердец у мышей после удаления бета-катенина в эмбриональной эндодерме» . Ячейка развития . 3 (2): 171–181. doi : 10.1016/s1534-5807 (02) 00206-x . PMID 12194849 .
- ^ Jump up to: а беременный Hertig CM, Butz S, Koch S, Eppenberger-Eberhardt M, Kemler R, Eppenberger HM (январь 1996 г.). «N-кадгерин в кардиомиоцитах взрослых крыс в культуре. II. Пространственно-временное появление белков, участвующих в контакте и коммуникации клеток. Журнал сотовой науки . 109 (1): 11–20. doi : 10.1242/jcs.109.1.11 . PMID 8834786 .
- ^ Уилер М.А., Уорли А., Робертс Р.Г., Элер Е., Эллис Дж. А (март 2010 г.). «Идентификация комплекса эмерин-бета-катенина в сердце, важном для интеркалированной архитектуры диска и локализации бета-катенина» . Клеточные и молекулярные науки о жизни . 67 (5): 781–796. doi : 10.1007/s00018-009-0219-8 . PMC 11115513 . PMID 19997769 . S2CID 27205170 .
- ^ Ван Х, Гердес А.М. (февраль 1999 г.). «Хроническая гипертрофия сердечной перегрузки и неудачи у морских свинок: III. Интеркалированное ремоделирование диска». Журнал молекулярной и клеточной кардиологии . 31 (2): 333–343. doi : 10.1006/jmcc.1998.0886 . PMID 10093046 .
- ^ Йошида М., Окуса Т., Накашима Т., Таканари Х., Яно М., Такемура Г. и др. (Октябрь 2011). «Изменения в соединении адгезии предшествуют ремоделированию разрыва перехода во время развития сердечной недостаточности у кардиомиопатических хомяков» . Сердечно -сосудистые исследования . 92 (1): 95–105. doi : 10.1093/cvr/cvr182 . PMID 21693625 .
- ^ Hahn Jy, Cho HJ, Bae JW, Yuk HS, Kim Ki, Park KW, et al. (Октябрь 2006 г.). «Сверхэкспрессия бета-катенина уменьшает размер инфаркта миокарда за счет дифференциального воздействия на кардиомиоциты и фибробласты сердца» . Журнал биологической химии . 281 (41): 30979–30989. doi : 10.1074/jbc.m603916200 . PMID 16920707 .
- ^ Zheng Q, Chen P, Xu Z, Li F, Yi XP (октябрь 2013). «Экспрессия и перераспределение β-катенина в сердечных миоцитах левого желудочка спонтанно гипертонической крысы». Журнал молекулярной гистологии . 44 (5): 565–573. doi : 10.1007/s10735-013-9507-6 . PMID 23591738 . S2CID 18997718 .
- ^ Бауранд А., Зелараян Л., Бетни Р., Герке С., Дангер С., Ноак С. и др. (Май 2007). «Понижение бета-катенина требуется для адаптивного ремоделирования сердца» . Исследование циркуляции . 100 (9): 1353–1362. doi : 10.1161/01.res.0000266605.63681.5a . PMID 17413044 .
- ^ Chen X, Shevtsov SP, Hsich E, Cui L, Haq S, Aronovitz M, et al. (Июнь 2006 г.). «Сигнальный путь фактора бета-катенина/Т-клеток/энхансерного фактора лимфоцитов необходим для нормальной гипертрофии сердца, вызванной стрессом» . Молекулярная и клеточная биология . 26 (12): 4462–4473. doi : 10.1128/mcb.02157-05 . PMC 1489123 . PMID 16738313 .
- ^ Haq S, Michael A, Andreucci M, Bhattacharya K, Dotto P, Walters B, et al. (Апрель 2003 г.). «Стабилизация бета-катенина с помощью WNT-независимого механизма регулирует рост кардиомиоцитов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (8): 4610–4615. Bibcode : 2003pnas..100.4610H . doi : 10.1073/pnas.0835895100 . PMC 153603 . PMID 12668767 .
- ^ Hirschy A, Croquelois A, Perriard E, Schoenauer R, Agarkova I, Hoerstrup SP, et al. (Сентябрь 2010). «Стабилизированный бета-катенин в постнатальном желудочковом миокарде приводит к расширенной кардиомиопатии и преждевременной смерти» (PDF) . Основные исследования в области кардиологии . 105 (5): 597–608. doi : 10.1007/s00395-010-0101-8 . PMID 20376467 . S2CID 21789076 .
- ^ Li J, Swope D, Raess N, Cheng L, Muller EJ, Radice GL (март 2011 г.). «Сердечная ткань ограниченная делеция плакоглобина приводит к прогрессирующей кардиомиопатии и активации передачи сигналов {бета} -катенина» . Молекулярная и клеточная биология . 31 (6): 1134–1144. doi : 10.1128/mcb.01025-10 . PMC 3067899 . PMID 21245375 .
- ^ Swope D, Cheng L, Gao E, Li J, Radice GL (март 2012 г.). «Потеря кадгерин-связывающих белков β-катенин и плакоглобин в сердце приводит к ремоделированию щелевого соединения и аритмогенезу» . Молекулярная и клеточная биология . 32 (6): 1056–1067. doi : 10.1128/mcb.06188-11 . PMC 3295003 . PMID 22252313 .
- ^ Jump up to: а беременный Dias C, Feng J, Sun H, Shao NY, Mazei-Robison MS, Damez-Erno D, et al. (Декабрь 2014). среднежила «β- катенинтинизированная Природа 516 (7529): 51–5 Bibcode : 2014natur.516 ... 51d Doi : 10.1038/ nature1 PMC 4257892 . PMID 25383518
- ^ Perriard JC, Hirschy A, Ehler E (январь 2003 г.). «Дилатационная кардиомиопатия: болезнь интеркалированного диска?». Тенденции в сердечно -сосудистой медицине . 13 (1): 30–38. doi : 10.1016/S1050-1738 (02) 00209-8 . PMID 12554098 .
- ^ Jump up to: а беременный Элер Е., Элер Е., Хубер О., Мартус П., Дей. (2006). Человек FASEB Journal 20 (7): 926–934. doi : 10.1096/ fj.05-5148com PMID 16675850 . 2246390S2CID
- ^ Cantù C, Felker A, Zimmerli D, Prummel KD, Cabello EM, Chiavacci E, et al. (Ноябрь 2018). «Мутации в генах Bcl9 и Pygo вызывают врожденные дефекты сердца путем тканевого специфического возмущения передачи сигналов Wnt/β-catenin» . Гены и развитие . 32 (21–22): 1443–1458. doi : 10.1101/gad.315531.118 . PMC 6217730 . PMID 30366904 .
- ^ Forbes SA, Bindal N, Bamford S, Cole C, Kok CY, Beare D, et al. (Январь 2011). «Космический: добыча полных геномов рака в каталоге соматических мутаций при раке» . Исследование нуклеиновых кислот . 39 (проблема базы данных): D945 - D950. doi : 10.1093/nar/gkq929 . PMC 3013785 . PMID 20952405 .
- ^ Saldanha G, Ghura V, Potter L, Fletcher A (июль 2004 г.). «Ядерный бета-катенин при карциноме базального клеток коррелирует с повышенной пролиферацией». Британский журнал дерматологии . 151 (1): 157–164. doi : 10.1111/j.1365-2133.2004.06048.x . PMID 15270885 . S2CID 31114274 .
- ^ Kypta RM, Waxman J (август 2012 г.). «Передача сигналов Wnt/β-Catenin при раке предстательной железы». Природные обзоры. Урология . 9 (8): 418–428. doi : 10.1038/nrurol.2012.116 . PMID 22710668 . S2CID 22945223 .
- ^ Hassanein AM, Glanz SM, Kessler HP, Eskin TA, Liu C (ноябрь 2003 г.). «Бета-катенин экспрессируется аберрантно в опухолях, экспрессирующих теневые клетки. Пиломатрома, краниофарингиома и кальцифицирующая одонтогенная киста» . Американский журнал клинической патологии . 120 (5): 732–736. doi : 10.1309/ealeg7ld6w7167px . PMID 14608900 .
- ^ Ellison DW, Onilude OE, Lindsey JC, Lusher ME, Weston CL, Taylor Re, et al. (Ноябрь 2005 г.). «Статус бета-катенина предсказывает благоприятный результат при детской медуллобластоме: Комитет по изучению рака в Соединенном Королевстве». Журнал клинической онкологии . 23 (31): 7951–7957. doi : 10.1200/jco.2005.01.5479 . PMID 16258095 .
- ^ Pooja Navale, MD, Omid Sairari, MD, Joseph F. Tomashefski, Jr., MD, Monika Vyas, MD «Твердое псевдопапиллярное новообразование» . Патология .
{{cite web}}: CS1 Maint: несколько имен: список авторов ( ссылка ) Последнее автор Обновление: 4 марта 2022 г. - ^ Elabeh M, Saha SK, Afrin S, Islam MS, Borahay MA (2021). «Сигнальный путь Wnt/β-катенин в лейомиоме матки: роль в биологии опухоли и возможности нацеливания» . Мол -клеточная биохимия . 476 (9): 3513–3536. doi : 10.1007/s11010-021-04174-6 . PMC 9235413 . PMID 33999334 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Carlson JW, Fletcher CD (2007). «Иммуногистохимия для бета-катенина в дифференциальной диагностике поражений клеток веретена: анализ серии и обзор литературы» . Гистопатология . 51 (4): 509–14. doi : 10.1111/j.1365-2559.2007.02794.x . PMID 17711447 .
- ^ Kobayashi M, Honma T, Matsuda Y, Suzuki Y, Narisawa R, Ajioka Y, Asakura H (май 2000). «Ядерная транслокация бета-катенина при колоректальном раке» . Британский журнал рака . 82 (10): 1689–1693. doi : 10.1054/bjoc.1999.1112 . PMC 2374509 . PMID 10817505 .
- ^ Воронков А., Краусс С. (2013). «Передача сигналов Wnt/Beta-Catenin и ингибиторы малых молекул» . Текущий фармацевтический дизайн . 19 (4): 634–664. doi : 10.2174/1381612811306040634 . PMC 3529405 . PMID 23016862 .
- ^ De La Roche M, Rutherford TJ, Gupta D, Veprintsev DB, Saxty B, Freund SM, Bienz M (февраль 2012 г.). «По сути, лабильная α-спираль, примыкающая к BCL9-связывающему сайту β-катенина, необходим для его ингибирования карнозийной кислотой» . Природная связь . 3 (2): 680. Bibcode : 2012natco ... 3..680d . doi : 10.1038/ncomms1680 . PMC 3293410 . PMID 22353711 .
- ^ Такада К., Чжу Д., Берд Г.Х., Сухдео К., Чжао Дж.Дж., Мани М. и др. (Август 2012 г.). «Целевое нарушение комплекса BCL9/β-катенина ингибирует онкогенную передачу сигналов Wnt» . Научная трансляционная медицина . 4 (148): 148ra117. doi : 10.1126/scitranslmed.3003808 . PMC 3631420 . PMID 22914623 .
- ^ Munkácsy G, Sztupinszki Z, Herman P, Bán B, Monetary Z, Szarvas N, Győrffy B (сентябрь 2016 г.). «Валидация эффективности молчания РНКи с использованием данных генов показывает 18,5% частоту отказов в 429 независимых экспериментах» . Молекулярная терапия. Нуклеиновые кислоты . 5 (9): E366. Doi : 10.1038/mtna.2016.66 . PMC 5056990 . PMID 27673562 .
- ^ Cao Z, Hao Z, Xin M, Yu L, Wang L, Zhang Y, et al. (Декабрь 2018). «Эндогенный и экзогенный галектин-3 способствует адгезии опухолевых клеток с низкой экспрессией MUC1 до HUVEC посредством активации N-кадгерина и CD44» . Лабораторное расследование; Журнал технических методов и патологии . 98 (12): 1642–1656. doi : 10.1038/s41374-018-0119-3 . PMID 30171204 . S2CID 52139917 .
- ^ Номер клинического испытания NCT02117362 для «ингибитора галектина (GR-MD-02) и ипилимумаба у пациентов с метастатической меланомой» на клинических
- ^ Moor AE, Anderle P, Cantù C, Rodriguez P, Wiedemann N, Baruthio F, et al. (Декабрь 2015). «Передача сигналов BCL9/9L-β-катенина связана с плохим исходом при колоректальном раке» . ebiomedicine . 2 (12): 1932–1943. doi : 10.1016/j.ebiom.2015.10.030 . PMC 4703711 . PMID 26844272 .
- ^ Flentke GR, Garic A, Amberger E, Hernandez M, Smith SM (июль 2011 г.). «Кальций-опосредованная репрессия β-катенина и его транскрипционная передача сигналов опосредует гибель клеток нервного гребня в птичьей модели синдрома алкоголя плода» . Врожденные дефекты исследования. Часть A, клиническая и молекулярная тератология . 91 (7): 591–602. doi : 10.1002/bdra.20833 . PMC 4827605 . PMID 21630427 .
- ^ Su Lk, Vogelstein B, Kinzler KW (декабрь 1993). «Ассоциация белка -супрессора APC -опухоли с катенинами». Наука . 262 (5140): 1734–1737. Bibcode : 1993sci ... 262.1734S . doi : 10.1126/science.8259519 . PMID 8259519 .
- ^ Jump up to: а беременный в дюймовый Kucerová D, Sloncová E, Tuhácková от, Vojtechová M, Sovová In (декабрь 2001 г.). «Экспрессия и взаимодействие различных катенинов в клетках колоректальной карциномы». Международный журнал молекулярной медицины . 8 (6): 695–698. Doi : 10.3892/ijmm.8.6.695 . PMID 11712088 .
- ^ Tickenbrock L, Kössmeier K, Rehmann H, Herrmann C, Müller O (март 2003 г.). «Различия между взаимодействием бета-катенина с нефосфорилированным и одному имитированным фосфорилированным 20-аминокислотным остатком повторений белка APC». Журнал молекулярной биологии . 327 (2): 359–367. doi : 10.1016/s0022-2836 (03) 00144-x . PMID 12628243 .
- ^ Jump up to: а беременный Дэвис Г., Цзян В.Г., Мейсон М.Д. (апрель 2001 г.). «Взаимодействие между бета-катенином, GSK3beta и APC после диссоциации клеточной клеток, вызванной мотогеном, и их участие в путях передачи сигнала при раке предстательной железы». Международный журнал онкологии . 18 (4): 843–847. doi : 10.3892/ijo.18.4.843 . PMID 11251183 .
- ^ Ryo A, Nakamura M, Wulf G, Liou YC, Lu KP (сентябрь 2001 г.). «PIN1 регулирует оборот и субклеточную локализацию бета-катенина, ингибируя его взаимодействие с APC». Природная клеточная биология . 3 (9): 793–801. doi : 10.1038/ncb0901-793 . PMID 11533658 . S2CID 664553 .
- ^ Homma Mk, Li D, Krebs EG, Yuasa Y, Homma Y (апрель 2002 г.). «Ассоциация и регуляция активности казеинкиназы 2 с помощью аденоматозного полипоза белка coli» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (9): 5959–5964. Bibcode : 2002pnas ... 99.5959K . doi : 10.1073/pnas.092143199 . PMC 122884 . PMID 11972058 .
- ^ Сато К, Янай Х, Сенда Т., Коху К, Накамура Т., Окумура Н. и др. (Июнь 1997 г.). «DAP-1, новый белок, который взаимодействует с гуанилат-киназоподобными доменами HDLG и PSD-95» . Гены к клеткам . 2 (6): 415–424. doi : 10.1046/j.1365-2443.1997.1310329.x . PMID 9286858 . S2CID 8934092 .
- ^ Eklof Spink K, Fridman SG, Weis WI (ноябрь 2001 г.). «Молекулярные механизмы распознавания бета-катенина с помощью аденоматозного полипоза Coli, выявленные структурой комплекса APC-бета-катенина» . Embo Journal . 20 (22): 6203–6212. doi : 10.1093/emboj/20.22.6203 . PMC 125720 . PMID 11707392 .
- ^ Накамура Т., Хамада Ф., Ишидат Т., Анай К, Кавахара К., Тойошима К., Акияма Т (июнь 1998 г.). «Аксин, ингибитор сигнального пути Wnt, взаимодействует с бета-катенином, GSK-3beta и APC и снижает уровень бета-катенина» . Гены к клеткам . 3 (6): 395–403. doi : 10.1046/j.1365-2443.1998.00198.x . PMID 9734785 . S2CID 10875463 .
- ^ Hocevar BA, Mou F, Rennolds JL, Morris SM, Cooper JA, Howe PH (июнь 2003 г.). «Регуляция сигнального пути Wnt с помощью отключенного 2 (DAB2)» . Embo Journal . 22 (12): 3084–3094. doi : 10.1093/emboj/cdg286 . PMC 162138 . PMID 12805222 .
- ^ Ян Ф., Ли Х, Шарма М., Сасаки Сай, Лонго Д.Л., Лим Б., Солнце З. (март 2002 г.). «Связывание бета-катенина с андрогеновым сигнальным путем» . Журнал биологической химии . 277 (13): 11336–11344. doi : 10.1074/jbc.m1111962200 . PMID 11792709 .
- ^ Masiello D, Chen Sy, Xu Y, Verhoeven MC, Choi E, Hollenberg AN, Balk SP (октябрь 2004 г.). «Рекрутирование бета-катенина рецепторами андрогенов дикого типа или мутанта коррелирует с стимулированным лиганд рост клеток рака предстательной железы» . Молекулярная эндокринология . 18 (10): 2388–2401. doi : 10.1210/me.2003-0436 . PMID 15256534 .
- ^ Song LN, Coghlan M, Gelmann EP (январь 2004 г.). «Антиандрогеновое влияние мифепристона на взаимодействие коактиватора и корепрессора с рецептором андрогена» . Молекулярная эндокринология . 18 (1): 70–85. doi : 10.1210/me.2003-0189 . PMID 14593076 .
- ^ Амир Ал, Баруа М., Макнайт Н.К., Ченг С., Юань Х, Балк С.П. (август 2003 г.). «Прямое бета-катенин-независимое взаимодействие между андрогеновым рецептором и фактором T-клеток 4» . Журнал биологической химии . 278 (33): 30828–30834. doi : 10.1074/jbc.m301208200 . PMID 12799378 .
- ^ Jump up to: а беременный Mulholland DJ, Read JT, Rennie PS, Cox ME, Nelson CC (август 2003 г.). «Функциональная локализация и конкуренция между рецептором андрогена и фактором Т-клеток для ядерного бета-катенина: среднее значение для ингибирования сигнальной оси TCF». Онкоген . 22 (36): 5602–5613. doi : 10.1038/sj.onc.1206802 . PMID 12944908 . S2CID 9301471 .
- ^ Pawlowski JE, Ertel JR, Allen MP, Xu M, Butler C, Wilson EM, Wierman ME (июнь 2002 г.). «Лиговое взаимодействие андрогенового рецептора с бета-катенином: ядерная совместная локализация и модуляция транскрипционной активности в нейрональных клетках» . Журнал биологической химии . 277 (23): 20702–20710. doi : 10.1074/jbc.m200545200 . PMID 11916967 .
- ^ Takemaru K, Yamaguchi S, Lee YS, Zhang Y, Carthew RW, Moon RT (апрель 2003 г.). «Чибби, ядерный бета-катенин, ассоциированный антагонист пути Wnt/без крыла». Природа . 422 (6934): 905–909. Bibcode : 2003natur.422..905t . doi : 10.1038/nature01570 . PMID 12712206 . S2CID 4418716 .
- ^ Дэвис Г., Цзян В.Г., Мейсон М.Д. (апрель 2001 г.). «HGF/SF модифицирует взаимодействие между его рецептором C-MET и комплексом E-кадгерина/катенина в клетках рака простаты». Международный журнал молекулярной медицины . 7 (4): 385–388. doi : 10.3892/ijmm.7.4.385 . PMID 11254878 .
- ^ Jump up to: а беременный Oyama T, Kanai Y, Ochiai A, Akimoto S, Oda T, Yanagihara K, et al. (Декабрь 1994). «Усеченный бета-катенин нарушает взаимодействие между E-кадгерином и альфа-катенином: причина потери межклеточной адгезивности в клеточных линиях рака человека». РАНКА . 54 (23): 6282–6287. PMID 7954478 .
- ^ Хазан Р.Б., Кан Л., Роу С., Борген П.И., Римм Д.Л. (декабрь 1997 г.). «Винкулин связан с комплексом e-кадгериновой адгезии» . Журнал биологической химии . 272 (51): 32448–32453. doi : 10.1074/jbc.272.51.32448 . PMID 9405455 .
- ^ Кинч М.С., Кларк Г.Дж., Дер С.Дж., Берридж К (июль 1995 г.). «Фосфорилирование тирозина регулирует адгезии трансформированного RAS эпителии молочной железы» . Журнал клеточной биологии . 130 (2): 461–471. doi : 10.1083/jcb.130.2.461 . PMC 2199929 . PMID 7542250 .
- ^ Jiang MC, Liao CF, Tai CC (июнь 2002 г.). «CAS/CSE 1 стимулирует E-кадрин-зависимую полярность клеток в эпителиальных клетках толстой кишки HT-29». Биохимическая и биофизическая исследовательская коммуникация . 294 (4): 900–905. doi : 10.1016/s0006-291x (02) 00551-x . PMID 12061792 .
- ^ Jump up to: а беременный в дюймовый Хазан Р.Б., Нортон Л (апрель 1998 г.). «Рецептор эпидермального фактора роста модулирует взаимодействие E-кадгерина с актиновым цитоскелетом» . Журнал биологической химии . 273 (15): 9078–9084. doi : 10.1074/jbc.273.15.9078 . PMID 9535896 .
- ^ Jump up to: а беременный в Бонвини П., В.Г., Розолен А., Нгуен П., Трепель Дж., Гарсия де Херрерос А. и др. (Февраль 2001 г.). «Гелданамицин аннулирует ассоциацию ERBB2 с устойчивым к протеасом бета-катенин в клетках меланомы, увеличивает ассоциацию бета-катенина-E-кадгерина и снижает бета-катенин-чувствительную транскрипцию». РАНКА . 61 (4): 1671–1677. PMID 11245482 .
- ^ Jump up to: а беременный Ли Y, Бхарти А., Чен Д., Гонг Дж., Куфе Д. (декабрь 1998 г.). «Взаимодействие гликогеновой синтазы киназы 3бета с антигеном, ассоциированным с карциномой DF3/MUC1, и бета-катенином» . Молекулярная и клеточная биология . 18 (12): 7216–7224. doi : 10.1128/mcb.18.12.7216 . PMC 109303 . PMID 9819408 .
- ^ Wendeler MW, Praus M, Jung R, Hecking M, Metzig C, Gessner R (апрель 2004 г.). «KSP-Cadherin-это функциональная клеточная молекула адгезии, связанную с Li-Cadherin». Экспериментальные исследования клеток . 294 (2): 345–355. doi : 10.1016/j.yexcr.2003.11.022 . PMID 15023525 .
- ^ Jump up to: а беременный Шибата Т., Чума М., Кокубу А., Сакамото М., Хирохаши С (июль 2003 г.). «EBP50, бета-ассоцированный белок, усиливает передачу сигналов Wnt и чрезмерно экспрессируется при гепатоцеллюлярной карциноме» . Гепатология . 38 (1): 178–186. doi : 10.1053/jhep.2003.50270 . PMID 12830000 . S2CID 10325091 .
- ^ Jump up to: а беременный Павет Дж., Миравет С., Кастаньо Дж. P120 Молекулярный и клеточный биолог 23 (7): 2287–2 doi : 10.1128/ mcb.23.2287-2 PMC 150740 . PMID 1264014 .
- ^ Kang JS, Feinleib JL, Knox S, Ketteringham MA, Krauss RS (апрель 2003 г.). «Промеогенные члены семей Ig и Cadherin связаны с положительной регулированием дифференциации» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (7): 3989–3994. Bibcode : 2003pnas..100.3989K . doi : 10.1073/pnas.0736565100 . PMC 153035 . PMID 12634428 .
- ^ Oneyama C, Nakano H, Sharma SV (март 2002 г.). «UCS15A, новая маленькая молекула, опосредованная доменом SH3-доменом белкового взаимодействия, блокирующее лекарство». Онкоген . 21 (13): 2037–2050. doi : 10.1038/sj.onc.1205271 . PMID 11960376 . S2CID 23869665 .
- ^ Navarro P, Lozano E, Cano A (август 1993 г.). «Экспрессия e- или p-cadherin недостаточно для модификации морфологии и онкогенного поведения клеток карциномы мышиного веретена. Возможное участие плакоглобина». Журнал сотовой науки . 105 (4): 923–934. doi : 10.1242/jcs.105.4.923 . HDL : 10261/78716 . PMID 8227214 .
- ^ Jump up to: а беременный Такахаши К., Сузуки К., Цукатани Ю (июль 1997 г.). «Индукция фосфорилирования тирозина и ассоциации бета-катенина с рецептором EGF при триптическом расщеплении покоящихся клеток при слиянии». Онкоген . 15 (1): 71–78. doi : 10.1038/sj.onc.1201160 . PMID 9233779 . S2CID 10127053 .
- ^ Jump up to: а беременный Dobrosotkaya IY, Джеймс Г.Л. (апрель 2000 г.). «MAGI-1 взаимодействует с бета-катенином и связан с клеточными структурами адгезии». Биохимическая и биофизическая исследовательская коммуникация . 270 (3): 903–909. doi : 10.1006/bbrc.2000.2471 . PMID 10772923 .
- ^ Geng L, Burrow CR, Li HP, Wilson PD (декабрь 2000 г.). «Модификация состава многопротеиновых комплексов полицистина-1 путем фосфорилирования кальция и тирозина». Biochimica et Biophysica Acta (BBA) - молекулярная основа болезни . 1535 (1): 21–35. doi : 10.1016/s0925-4439 (00) 00079-x . PMID 11113628 .
- ^ Shibamoto S, Hayakawa M, Takeuchi K, Hori T, Miyazawa K, Kitamura N, et al. (Март 1995 г.). «Ассоциация P120, субстрат тирозинкиназы, с комплексами E-Cadherin/Catenin» . Журнал клеточной биологии . 128 (5): 949–957. doi : 10.1083/jcb.128.5.949 . PMC 2120395 . PMID 7876318 .
- ^ Рао Р.К., Басурой С., Рао Ву, Карнаки К.Дж., Гупта А (декабрь 2002 г.). «Фосфорилирование тирозина и диссоциация комплексов окклюдина-зо-1 и E-кадгерина-бета-катенина из цитоскелета с помощью окислительного стресса» . Биохимический журнал . 368 (Pt 2): 471–481. doi : 10.1042/bj20011804 . PMC 1222996 . PMID 12169098 .
- ^ Jump up to: а беременный Schmeiser K, Grand RJ (апрель 1999 г.). «Судьба E- и P-кадгерина на ранних стадиях апоптоза» . Гибель клеток и дифференциация . 6 (4): 377–386. doi : 10.1038/sj.cdd.4400504 . PMID 10381631 .
- ^ Пай Р., Данлэп Д., Цин Дж, Мохташеми И., Хотцель К., Французский Д.М. (июль 2008 г.). «Ингибирование фактора роста фибробластов 19 снижает рост опухоли за счет модуляции передачи сигналов бета-катенина» . РАНКА . 68 (13): 5086–5095. doi : 10.1158/0008-5472.can-07-2325 . PMID 18593907 .
- ^ Straub Bk, Boda J, Kuhn C, Schnoelzer M, Korf U, Kempf T, et al. (Декабрь 2003 г.). «Новая клеточная система соединения: кора Adhaerens Mosaic из волоконных клеток линзы» . Журнал сотовой науки . 116 (Pt 24): 4985–4995. doi : 10.1242/jcs.00815 . PMID 14625392 .
- ^ Wahl JK, Kim YJ, Cullen JM, Johnson KR, Wheelock MJ (май 2003). «Комплексы N-кадгерина-катенина образуются перед расщеплением прорегиона и транспортировки в плазматическую мембрану» . Журнал биологической химии . 278 (19): 17269–17276. doi : 10.1074/jbc.m211452200 . PMID 12604612 .
- ^ Klingelhöfer J, Troyanovsky RB, Laur Oy, Troyanovsky S (август 2000 г.). «Амино-концевой домен классических кадгеринов определяет специфичность клеяных взаимодействий». Журнал сотовой науки . 113 (16): 2829–2836. doi : 10.1242/jcs.113.16.2829 . PMID 10910767 .
- ^ Кесавапани С., Лау К.Ф., Маклафлин Д.М., Браунли Дж., Акерли С., Ли П.Н. и др. (Январь 2001). «P35/CDK5 связывается и фосфорилирует бета-катенин и регулирует взаимодействие бета-катенина/пресенилина-1». Европейский журнал нейробиологии . 13 (2): 241–247. doi : 10.1046/j.1460-9568.2001.01376.x (неактивный 2024-07-10). PMID 11168528 .
{{cite journal}}: CS1 Maint: doi неактивен по состоянию на июль 2024 г. ( ссылка ) - ^ Jump up to: а беременный Ламберти С., Лин К.М., Ямамото Ю., Верма У, Верма И.М., Байерс С., Гейнор Р.Б. (ноябрь 2001 г.). «Регуляция функции бета-катенина икапбаб киназы» . Журнал биологической химии . 276 (45): 42276–42286. doi : 10.1074/jbc.m104227200 . PMID 11527961 .
- ^ Roe S, Koslov ER, Rimm DL (июнь 1998 г.). «Мутация в альфа-катенине нарушает адгезию в клетках клона А, не нарушая его активность связывания актина и бета-катенина» . Клеточная адгезия и общение . 5 (4): 283–296. doi : 10.3109/15419069809040298 . PMID 9762469 .
- ^ Aberle H, Butz S, Stappert J, Weissig H, Kemler R, Hoschuetzky H (декабрь 1994 г.). «Сборка комплекса кадгерина-катенина in vitro с рекомбинантными белками». Журнал сотовой науки . 107 (12): 3655–3663. doi : 10.1242/jcs.107.12.3655 . PMID 7706414 .
- ^ Reuver SM, Garner CC (апрель 1998 г.). «E-кадгерин опосредованная клеточная адгезия рекрутирует SAP97 в кортикальный цитоскелет». Журнал сотовой науки . 111 (8): 1071–1080. doi : 10.1242/jcs.111.8.1071 . PMID 9512503 .
- ^ Jump up to: а беременный Schroeder JA, Adriance MC, McConnell EJ, Thompson MC, Pockaj B, Gendler SJ (июнь 2002 г.). «Эрбб-бета-катениновые комплексы связаны с инфильтрирующим протоковым протоковым и мышиным вирусом опухоли молочной железы (MMTV) -WNT-1 и трансгенные карциномы MMTV-C-NEU» . Журнал биологической химии . 277 (25): 22692–22698. doi : 10.1074/jbc.m201975200 . PMID 11950845 .
- ^ Персонажи L, Mr Baelets, Barresi R, Squarezoni S, Sabatels P, Maraldi N и Aldis. (Demember 1997). «Сердечная локализация появляющихся: теперь понимание Emery-Dreifuss Diastrphy» . Молекулярные молекулярные генералы . 6 (13): 2257–2264. doi : 10,1093/hmg/6,13 2257 . PMID 9361031 .
- ^ Markiewicz E, Tilgner K, Barker N, Van De Wetering M, Clevers H, Dorobek M, et al. (Июль 2006 г.). «Внутренний белок ядерной мембраны регулирует активность бета-катенина, ограничивая его накопление в ядре» . Embo Journal . 25 (14): 3275–3285. doi : 10.1038/sj.emboj.7601230 . PMC 1523183 . PMID 16858403 .
- ^ Вей Й., Ренард К.А., Лабалетт С., Ву Й, Леви Л., Нойвейт С. и др. (Февраль 2003 г.). «Идентификация белка LIM FHL2 как коактиватора бета-катенина» . Журнал биологической химии . 278 (7): 5188–5194. doi : 10.1074/jbc.m207216200 . PMID 12466281 .
- ^ Кишида С., Ямамото Х., Хино С., Икеда С., Кишида М., Кикучи А (июнь 1999 г.). «Домены DIX DVL и аксин необходимы для белковых взаимодействий и их способности регулировать стабильность бета-катенина» . Молекулярная и клеточная биология . 19 (6): 4414–4422. doi : 10.1128/mcb.19.6.44414 . PMC 104400 . PMID 10330181 .
- ^ Канай Ю., Очиай А., Шибата Т., Ояма Т., Ушиджима С., Акимото С., Хирохаши С. (март 1995 г.). «Продукт гена C-ERBB-2 напрямую ассоциируется с бета-катенином и плакоглобином». Биохимическая и биофизическая исследовательская коммуникация . 208 (3): 1067–1072. doi : 10.1006/bbrc.1995.1443 . PMID 7702605 .
- ^ Jump up to: а беременный Эдлунд С., Ли Си, Гримсби С., Чжан С., Ассенстрем П., Хернд Ч., Ландстрем М (февраль 2005 г.). «Взаимодействие между SMAD7 и бета-катенином: важность трансформации фактора роста, вызванного бета-индуцированным апоптозом» . Молекулярная и клеточная биология . 25 (4): 1475–1488. doi : 10.1128/mcb.25.4.1475-1488.2005 . PMC 548008 . PMID 15684397 .
- ^ Grueneberg DA, Pablo L, Hu KQ, August P, Weng Z, Papkoff J (июнь 2003 г.). «Функциональный экран в клетках человека идентифицирует UBF2 как фактор транскрипции РНК-полимеразы II, который усиливает путь сигнализации бета-катенина» . Молекулярная и клеточная биология . 23 (11): 3936–3950. doi : 10.1128/mcb.23.11.3936-3950.2003 . PMC 155208 . PMID 12748295 .
- ^ Behrens J, Von Kries JP, Kühl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W (август 1996 г.). «Функциональное взаимодействие бета-катенина с транскрипционным фактором LEF-1». Природа . 382 (6592): 638–642. Bibcode : 1996natur.382..638b . doi : 10.1038/382638A0 . PMID 8757136 . S2CID 4369341 .
- ^ Labbé E, Letamendia A, Attisano L (июль 2000 г.). «Ассоциация SMAD с фактором связывания лимфоидного энхансера 1/T-клеток-специфического фактора опосредует кооперативную передачу сигналов путем трансформирующего фактора роста, бета-бета и Wnt» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (15): 8358–8363. Bibcode : 2000pnas ... 97.8358L . doi : 10.1073/pnas.150152697 . PMC 26952 . PMID 10890911 .
- ^ Barolo S, Posakony JW (май 2002 г.). «Три привычки высокоэффективных сигнальных путей: принципы контроля транскрипции путем передачи сигналов клеток развития» . Гены и развитие . 16 (10). Cold Spring Harbor Laboratory Press & The Genetics Society : 1167–1181. doi : 10.1101/gad.976502 . PMID 12023297 . S2CID 14376483 . п. 1170:
В ... Рыб-рычагах репортерные трансгены, содержащие промотор верхнего сорта, экспрессируются в определенных типах клеток, чувствительных к Wnt (... Dorsky et al. 2002).
- ^ Yamamoto M, Bharti A, Li Y, Kufe D (май 1997). «Взаимодействие антигена и бета-катенина, ассоциированного с карциномой молочной железы DF3/MUC1 в клеточной адгезии» . Журнал биологической химии . 272 (19): 12492–12494. doi : 10.1074/jbc.272.19.12492 . PMID 9139698 .
- ^ Durum SK, Aiello FB (2003). «Интерлейкин-7 индуцирует muc1» . Биология и терапия рака . 2 (2): 194–195. doi : 10.4161/cbt.2.2.351 . PMID 12750562 .
- ^ Schroeder JA, Adriance MC, Thompson MC, Camenisch TD, Gendler SJ (март 2003 г.). «MUC1 изменяет бета-катенин-зависимую образование опухоли и способствует клеточной инвазии». Онкоген . 22 (9): 1324–1332. doi : 10.1038/sj.onc.1206291 . PMID 12618757 . S2CID 25619311 .
- ^ Li Y, Kuwahara H, Ren J, Wen G, Kufe D (март 2001 г.). «Тирозинкиназа C-SRC регулирует передачу сигналов антигена, ассоциированного с карциномой DF3/MUC1, с бета-катеном GSK3 и бета-катеном» . Журнал биологической химии . 276 (9): 6061–6064. doi : 10.1074/jbc.c000754200 . PMID 11152665 .
- ^ Ren J, Li Y, Kufe D (май 2002 г.). «Дельта протеинкиназы С регулирует функцию антигена карциномы DF3/MUC1 в передаче сигналов бета-катенина» . Журнал биологической химии . 277 (20): 17616–17622. doi : 10.1074/jbc.m200436200 . PMID 11877440 .
- ^ Li Y, Ren J, Yu W, Li Q, Kuwahara H, Yin L, et al. (Сентябрь 2001 г.). «Рецептор эпидермального фактора роста регулирует взаимодействие антигена карциномы DF3/MUC1 MUC1 с C-SRC и бета-катенином» . Журнал биологической химии . 276 (38): 35239–35242. doi : 10.1074/jbc.c100359200 . PMID 11483589 .
- ^ Kennell JA, O'Leary EE, Gummow BM, Hammer GD, MacDougald OA (август 2003 г.). «Т-клеточный фактор 4N (TCF-4N), новая изоформа мышиного TCF-4, синергирует с бета-катенином, чтобы коактивировать C/EBPALPHA и стероидогенный фактор 1 транскрипции» . Молекулярная и клеточная биология . 23 (15): 5366–5375. doi : 10.1128/mcb.23.15.5366-5375.2003 . PMC 165725 . PMID 12861022 .
- ^ Mizusaki H, Kawabe K, Mukai T, Ariyoshi E, Kasahara M, Yoshioka H, et al. (Апрель 2003 г.). «Критическая область DAX-1 (чувствительная к дозировке по полу-диспренальной гипоплазии, критическая область гена, гена 1) транскрипция гена 1) регулируется Wnt4 у самок развивающейся гонады» . Молекулярная эндокринология . 17 (4): 507–519. doi : 10.1210/me.2002-0362 . PMID 12554773 .
- ^ GE X, Jin Q, Zhang F, Yan T, Zhai Q (январь 2009 г.). «Ацетилаты PCAF {бета} -catenin и улучшает его стабильность» . Молекулярная биология клетки . 20 (1): 419–427. doi : 10.1091/mbc.e08-08-0792 . PMC 2613091 . PMID 18987336 .
- ^ Беренс Дж (октябрь 2008 г.). «Один удар, два результата для VHL-опосредованного онкогенеза». Природная клеточная биология . 10 (10): 1127–1128. doi : 10.1038/ncb1008-1127 . PMID 18830218 . S2CID 36184371 .
- ^ Wadham C, Gamble JR, Vadas MA, Khew-Goodall Y (июнь 2003 г.). «Белковая тирозинфосфатаза PEZ является основной фосфатазой соединений прилипания и дефосфорилирует бета-катенин» . Молекулярная биология клетки . 14 (6): 2520–2529. doi : 10.1091/mbc.e02-09-0577 . PMC 194899 . PMID 12808048 .
- ^ Aicher B, Lerch MM, Müller T, Schilling J, Ullrich A (август 1997 г.). «Клеточное перераспределение белковых тирозинфосфатаз LAR и птпсигма с помощью индуцибельной протеолитической обработки» . Журнал клеточной биологии . 138 (3): 681–696. doi : 10.1083/jcb.138.3.681 . PMC 2141638 . PMID 9245795 .
- ^ Fuchs M, Müller T, Lerch MM, Ullrich A (июль 1996 г.). «Ассоциация человеческой белко-тирозинфосфатазы каппа с членами семейства Армадильо» . Журнал биологической химии . 271 (28): 16712–16719. doi : 10.1074/jbc.271.28.16712 . PMID 8663237 .
- ^ Besco JA, Hooft van Huijsduijnen R, Frotholm A, Rotter A (октябрь 2006 г.). «Внутриклеточные субстраты обогащенного мозга рецепторный белок тирозинфосфатазы Rho (rptprho/ptprt)». Исследование мозга . 1116 (1): 50–57. doi : 10.1016/j.brainres.2006.07.122 . PMID 16973135 . S2CID 23343123 .
- ^ Ван Б., Кишихара К., Чжан Д., Хара Х, Номото К (февраль 1997 г.). «Молекулярное клонирование и характеристика нового гена тирозинфосфатазы белка рецептора человека, HPTP-J: подавляя экспрессии генов с помощью PMA и кальциевого ионофора в клетках Lyrkat T-лимфомы». Биохимическая и биофизическая исследовательская коммуникация . 231 (1): 77–81. doi : 10.1006/bbrc.1997.6004 . PMID 9070223 .
- ^ Yan HX, He YQ, Dong H, Zhang P, Zeng JZ, Cao HF, et al. (Декабрь 2002 г.). «Физическое и функциональное взаимодействие между рецептор-подобным белковым тирозинфосфатазой PCP-2 и бета-катенином». Биохимия . 41 (52): 15854–15860. doi : 10.1021/bi026095u . PMID 12501215 .
- ^ He Y, Yan H, Dong H, Zhang P, Tang L, Qiu X, et al. (Апрель 2005 г.). «Структурная основа взаимодействия между белковой тирозинфосфатазой PCP-2 и бета-катенином». Science in China Series C: Life Sciences . 48 (2): 163–167. doi : 10.1007/bf02879669 . PMID 15986889 . S2CID 20799629 .
- ^ Tesco G, Kim TW, Diehlmann A, Beyreuther K, Tanzi Re (декабрь 1998 г.). «Отмена взаимодействия презенилина 1/бета-катенина и сохранение гетеродимерного комплекса пресенилина 1 после активации каспазы» . Журнал биологической химии . 273 (51): 33909–33914. doi : 10.1074/jbc.273.51.33909 . PMID 9852041 .
- ^ Кан де, Сориано С., Фрош М.П., Коллинз Т., Нарузе С., Сисодия С.С. и др. (Июнь 1999 г.). «Презенилин 1 облегчает конститутивный оборот бета-катенина: дифференциальная активность мутантов PS1, связанных с болезнью Альцгеймера в пути подписания бета-катенина» . Журнал нейробиологии . 19 (11): 4229–4237. doi : 10.1523/jneurosci.19-11-04229.1999 . PMC 6782616 . PMID 10341227 .
- ^ Мураяма М., Танака С., Палачино Дж., Мураяма О., Хонда Т., Сан Х и др. (Август 1998). «Прямая ассоциация пресенилина-1 с бета-катенином». Письма Febs . 433 (1–2): 73–77. doi : 10.1016/s0014-5793 (98) 00886-2 . PMID 9738936 . S2CID 85416623 .
- ^ Puppo F, Thomé V, Lhoumeau AC, Cibois M, Gangar A, Lembo F, et al. (Январь 2011). «Белковая тирозинкиназа 7 играет консервативную роль в канонической сигнализации Wnt/β-катенина» . Embo сообщает . 12 (1): 43–49. doi : 10.1038/inbom.2010.185 . PMC 3024124 . PMID 21132015 .
- ^ Бауэр А., Хубер О., Кемлер Р. (декабрь 1998 г.). «Pontin52, партнер по взаимодействию бета-катенина, связывается с белком, связывающим TATA,» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (25): 14787–14792. Bibcode : 1998pnas ... 9514787b . doi : 10.1073/pnas.95.25.14787 . PMC 24527 . PMID 9843967 .
- ^ Barker N, Hurlstone A, Musisi H, Miles A, Bienz M, Clevers H (сентябрь 2001 г.). «Фактор ремоделирования хроматина BRG-1 взаимодействует с бета-катенином для продвижения активации гена-мишени» . Embo Journal . 20 (17): 4935–4943. doi : 10.1093/emboj/20.17.4935 . PMC 125268 . PMID 11532957 .
- ^ Тайя С., Ямамото Т., Канай-Азума М., Вуд С.А., Кайбучи К (декабрь 1999 г.). «Деубиквитинирующий фермент FAM взаимодействует и стабилизирует бета-катенин» . Гены к клеткам . 4 (12): 757–767. doi : 10.1046/j.1365-2443.1999.00297.x . PMID 10620020 . S2CID 85747886 .
- ^ Lewalle JM, Bajou K, Desreux J, Mareel M, Dejana E, Noël A, Foidart JM (декабрь 1997 г.). «Изменение межпредвидежных соединений с помощью взаимодействия опухолевых клеток-эндотелиальных клеток in vitro». Экспериментальные исследования клеток . 237 (2): 347–356. doi : 10.1006/excr.1997.3799 . HDL : 2268/61990 . PMID 9434630 .
- ^ Shasby DM, Ries DR, Shasby SS, Winter MC (июнь 2002 г.). «Гистамин стимулирует фосфорилирование белков прилипания соединений и изменяет их связь с виментином». Американский журнал физиологии. Клеточная и молекулярная физиология легких . 282 (6): L1330 - L1338. Citeseerx 10.1.1.1000.5266 . doi : 10.1152/ajplung.00329.2001 . PMID 12003790 .
- ^ Sinn HW, Balsamo J, Lilien J, Lin JJ (сентябрь 2002 г.). «Локализация нового белка XIN в комплекс соединения прилипания в сердечной и скелетной мышце во время развития». Динамика развития . 225 (1): 1–13. doi : 10.1002/dvdy.10131 . PMID 12203715 . S2CID 23393425 .
Дальнейшее чтение
[ редактировать ]- Кикучи А (февраль 2000 г.). «Регуляция передачи сигналов бета-катенина в пути WNT». Биохимическая и биофизическая исследовательская коммуникация . 268 (2): 243–248. doi : 10.1006/bbrc.1999.1860 . PMID 10679188 .
- Уилсон П.Д. (апрель 2001 г.). «Полицистин: новые аспекты структуры, функции и регуляции» . Журнал Американского общества нефрологии . 12 (4): 834–845. doi : 10.1681/asn.v124834 . PMID 11274246 .
- Kalluri R, Neilson EG (декабрь 2003 г.). «Эпителиально-мезенхимальный переход и его последствия для фиброза» . Журнал клинических исследований . 112 (12): 1776–1784. doi : 10.1172/JCI20530 . PMC 297008 . PMID 14679171 .
- De Ferrari GV, Moon RT (декабрь 2006 г.). «UPS и падения передачи сигналов Wnt при распространенных неврологических расстройствах». Онкоген . 25 (57): 7545–7553. doi : 10.1038/sj.onc.1210064 . PMID 17143299 . S2CID 35684619 .
Внешние ссылки
[ редактировать ]- Бета+Катенин в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)
- «Разнообразный набор белков модулирует сигнальный путь канонического Wnt/β-катенина». у Cancer.gov
- «Роль β-катенина в трансдукции сигнала, определение судьбы клеток и трансдифференцирование» при nih.gov
- «Исследователи предлагают первое прямое доказательство того, как артрит разрушает хрящ» на Rochester.edu
- и CTNNB1 CTNNB1 Местоположение генома CTNNB1 страница гена в браузере генома UCSC .
Эта статья включает в себя текст из Национальной медицины Соединенных Штатов , которая находится в общественном достоянии .
PDB Галерея |
|---|















