Генетика популяции
| Часть серии на |
| Эволюционная биология |
|---|
 |
| Часть серии на |
| Генетическая генеалогия |
|---|
| Концепции |
| Связанные темы |
Генетика популяции является подполе генетики , которая занимается генетическими различиями внутри населения и среди населения , и является частью эволюционной биологии . Исследования в этой отрасли биологии исследуют такие явления, как адаптация , видообразование и структура населения . [ 1 ]
Генетика популяции была жизненно важным ингредиентом в появлении современного эволюционного синтеза . Его основными основателями были Sewall Wright , JBS Haldane и Ronald Fisher , которые также заложили основы для связанной дисциплины количественной генетики . Традиционно высоко математическая дисциплина, современная популяционная генетика охватывает теоретическую, лабораторную и полевую работу. Генетические модели популяции используются как для статистического вывода из данных последовательности ДНК, так и для доказательства/отделения концепции. [ 2 ]
То, что отличает генетику популяции от более новых, более фенотипических подходов к моделированию эволюции, таких как теория эволюционной игры и адаптивная динамика , - это его акцент на таких генетических явлениях, как доминирование , эпистаз , степень, в которой генетическая рекомбинация разбивает неравновесную связь и случайные явления. мутации генетического и дрейфа . Это делает его подходящим для сравнения с данными о геномике популяции .
История
[ редактировать ]Генетика популяции началась как примирение моделей наследования менделев и биостатистики . Естественный отбор достаточно генетических изменений вызовет эволюцию только в том случае, если в популяции . Перед открытием генетики Менделя , одной общей гипотезой смешивалась наследство . Но при смешивании наследования генетическая дисперсия была бы быстро потеряна, что делает эволюцию при естественном или сексуальном отборе неправдоподобным. Принцип Харди -Вайнберга обеспечивает решение для того, как вариация поддерживается в популяции с наследством менделевского. Согласно этому принципу, частоты аллелей (вариации в гене) останутся постоянными в отсутствие отбора, мутации, миграции и генетического дрейфа. [ 3 ]


Следующим ключевым шагом была работа британского биолога и статистика Рональда Фишера . В серии статей, начиная с 1918 года и кульминационной в своей книге 1930 года «Генетическая теория естественного отбора» , Фишер показал, что непрерывные вариации, измеренные биометрическими людьми частоты в популяции, что приводит к эволюции. В серии статей, начиная с 1924 года, другой британский генетитик JBS Haldane разработал математику изменения частоты аллеля в одном локусе гена в широком диапазоне условий. Haldane также применил статистический анализ к реальным примерам естественного отбора, таких как эволюция моли и промышленный меланизм , и показал, что коэффициенты отбора могут быть больше, чем предполагал Фишер, что приводит к более быстрому адаптивной эволюции в качестве стратегии камуфляжа после повышения загрязнения. [ 4 ] [ 5 ]
Американский биолог Сьюалл Райт , который имел опыт работы в экспериментах по разведению животных , сосредоточился на комбинациях взаимодействующих генов и влиянии инбридинга на небольшие, относительно изолированные популяции, которые демонстрировали генетический дрейф. В 1932 году Райт представил концепцию адаптивного ландшафта и утверждал, что генетический дрейф и инбридинги могут отобрать небольшую изолированную субпопуляцию от адаптивного пика, позволяя естественному отбору привести его к различным адаптивным пикам. [ Цитация необходима ]
Работа Фишера, Холдейна и Райта основала дисциплину генетики населения. Этот интегрированный естественный отбор с Mendelian Genetics, который был критическим первым шагом в разработке единой теории того, как работает эволюция. [ 4 ] [ 5 ] Джон Мейнард Смит был учеником Холдейна, в то время как В.Д. Гамильтон находился под влиянием писаний Фишера. Американский Джордж Р. Прайс работал с Гамильтоном и Мейнардом Смитом. Американский Ричард Левонтин и японская мотоо Кимура находились под влиянием Райта и Холдейна. [ Цитация необходима ]
Современный синтез
[ редактировать ]Математика генетики популяции была первоначально развита как начало современного синтеза . Такие авторы, как Битти [ 6 ] утверждают, что генетика популяции определяет ядро современного синтеза. В течение первых нескольких десятилетий 20 -го века большинство полевых натуралистов продолжали верить, что ламаркизм и ортогенез обеспечивали лучшее объяснение сложности, которую они наблюдали в живом мире. [ 7 ] Во время современного синтеза эти идеи были очищены, и были сохранены только эволюционные причины, которые могут быть выражены в математической основе генетики популяции. [ 8 ] Был достигнут консенсус относительно того, какие эволюционные факторы могут влиять на эволюцию, но не на относительную важность различных факторов. [ 8 ]
Теодосий Добханский , постдокторский работник в лаборатории Т. Моргана , оказала влияние работы Сергей российских генетиков, таких как Четвериков . Он помог преодолеть разделение между основаниями микроэволюции, разработанных генетиками популяции, и паттернами макроэволюции, наблюдаемых полевыми биологами, с его генетикой книги 1937 года и происхождением видов . Добжанский исследовал генетическое разнообразие диких популяций и показал, что, в отличие от предположений генетиков популяции, эти популяции имели большое количество генетического разнообразия, с заметными различиями между субпопуляциями. Книга также взяла высоко математическую работу генетиков популяции и поместила ее в более доступную форму. Гораздо больше биологов находились под влиянием генетики популяции через Добжанского, чем смогли прочитать высоко математические произведения в оригинале. [ 9 ]
В Великобритании Эб Форд , пионер экологической генетики , [ 10 ] Продолжение в течение 1930 -х и 1940 -х годов эмпирически продемонстрировать силу отбора из -за экологических факторов, включая способность поддерживать генетическое разнообразие посредством генетических полиморфизмов, таких как типы крови человека . Работа Форда, в сотрудничестве с Фишер, способствовала сдвигу акцента во время современного синтеза к естественному отбору как доминирующей силы. [ 4 ] [ 5 ] [ 11 ] [ 12 ]
Нейтральная теория и динамика фиксирования происхождения
[ редактировать ]Оригинальный, современный синтез -взгляд на генетику популяции предполагает, что мутации обеспечивают достаточно сырья, и фокусируется только на изменении частоты аллелей в популяциях . [ 13 ] Основными процессами, влияющими на частоты аллелей, являются естественный отбор , генетический дрейф , поток генов и рецидивирующая мутация . У Фишера и Райта были некоторые фундаментальные разногласия по поводу относительной роли отбора и дрейфа. [ 14 ] Доступность молекулярных данных по всем генетическим различиям привела к нейтральной теории молекулярной эволюции . С этой точки зрения, многие мутации являются вредными и поэтому никогда не наблюдаются, а большинство остальных являются нейтральными, т.е. не находится под отбором. С судьбой каждой нейтральной мутации, оставленной на случайность (генетический дрейф), направление эволюционных изменений обусловлено, с помощью которых возникают мутации, и поэтому не может быть захвачено моделями изменения частоты (существующих) аллелей. [ 13 ] [ 15 ]
Первоначальный взгляд на генетику популяции обобщает этот подход за пределами строго нейтральных мутаций и видит скорость, с которой конкретное изменение происходит как продукт скорости мутаций и вероятность фиксации . [ 13 ]
Четыре процесса
[ редактировать ]Выбор
[ редактировать ]Естественный отбор , который включает в себя половой отбор , заключается в том, что некоторые черты повышают вероятность организма выживания и воспроизведения . Генетика популяции описывает естественный отбор, определяя пригодность как склонность или вероятность выживания и размножения в конкретной среде. Фитнес обычно определяется символом w = 1- s , где s- коэффициент выбора . Естественный отбор действует на фенотипы , поэтому популяционные генетические модели предполагают относительно простые отношения, чтобы предсказать фенотип и, следовательно, пригодность от аллеля в одном или небольшом количестве локусов. Таким образом, естественный отбор превращает различия в пригодности людей с различными фенотипами в изменения частоты аллелей в популяции в течение последовательных поколений. [ Цитация необходима ]
До появления генетики популяции многие биологи сомневались в том, что небольших различий в физической подготовке были достаточными для того, чтобы иметь большое значение для эволюции. [ 9 ] Генетики популяции решали эту проблему частично, сравнивая отбор с генетическим дрейфом . Отбор может преодолеть генетический дрейф, когда S превышает 1, разделенный на эффективную численность популяции . Когда этот критерий соблюдается, вероятность того, что новый выгодный мутант становится фиксированным , приблизительно равна 2 с . [ 16 ] [ 17 ] Время до фиксации такого аллеля примерно . [ 18 ]

Доминирование
[ редактировать ]Доминирование означает, что фенотипический и/или фитнес -эффект одного аллеля в локусе зависит от того, какой аллель присутствует во второй копии для этого локуса. Рассмотрим три генотипа в одном локусе со следующими значениями пригодности [ 19 ]
| Генотип: | И 1 и 1 | A 1 A 2 | 2 A 2 |
| Относительная фитнеса: | 1 | 1-й | 1-S. |
| Генетика популяции Глоссарий | |
|---|---|
|
S является коэффициентом выбора , а H - коэффициент доминирования. Значение H дает следующую информацию:
| h = 0 | 1 2 , доминирующий рецессивного |
| h = 1 | 2 доминирующее , 1 рецессивное |
| 0 <h <1 | неполное доминирование |
| H <0 | передоминирование |
| H> 1 | Обследование |
Эпистаз
[ редактировать ]
Эпистаз означает, что фенотипический и/или фитнес -эффект аллеля в одном локусе зависит от того, какие аллели присутствуют в других локусах. Отбор действует не ни на одном локусе, а на фенотипе, который возникает посредством развития из полного генотипа. [ 20 ] Тем не менее, многие модели генетики популяции сексуальных видов являются моделями «одиночного локуса», где приспособленность человека рассчитывается как продукт вклада каждого из его локусов, что не предполагает никакого эпистаза.
Фактически, генотип для фитнес -ландшафта более сложный. Генетика популяции должна либо детально моделировать эту сложность, либо захватить ее некоторым более простым средним правилом. Эмпирически полезные мутации, как правило, имеют меньшую пользу для физической подготовки при добавлении к генетическому фону, который уже имеет высокую подготовку: это известно как уменьшающийся эпистаз. [ 21 ] Когда вредные мутации также оказывают меньший эффект подготовки на фоне высокой пригодности, это известно как «синергетический эпистаз». Однако влияние вредных мутаций, как правило, в среднем очень близко к мультипликативному или даже может показать противоположную картину, известную как «антагонистический эпистаз». [ 22 ]
Синергетический эпистаз является центральным в некоторых теориях чистки мутационной нагрузки [ 23 ] и к эволюции сексуального размножения .
Мутация
[ редактировать ]
Генетический процесс мутации происходит внутри человека, что приводит к наследственным изменениям в генетическом материале. Этот процесс часто характеризуется описанием начальных и окончательных состояний или видом изменений, которое произошло на уровне ДНК (например, мутация t-to-c, делеция 1 п.н. Белки (например, нулевая мутация, мутация потери функции) или на более высоком фенотипическом уровне (например, мутация красных глаз). Изменения одноклетотидов часто являются наиболее распространенным типом мутации, но многие другие типы мутаций возможны , и они встречаются с широко различными скоростями, которые могут показывать систематическую асимметрию или смещения ( смещение мутаций ).
Мутации могут включать в себя большие участки ДНК, которые становятся дублированными , обычно посредством генетической рекомбинации . [ 24 ] Это приводит к изменению числа копий в населении. Дубликации являются основным источником сырья для развития новых генов. [ 25 ] Другие типы мутаций иногда создают новые гены из ранее некодирующей ДНК. [ 26 ] [ 27 ]
В распределении фитнес -эффектов (DFE) для новых мутаций выгодно. Мутации с грубыми эффектами обычно вредны. Исследования на мух Drosophila melanogaster показывают, что если мутация изменяет белок, продуцируемый геном, это, вероятно, будет вредным, причем около 70 процентов этих мутаций оказывают вредные эффекты, а остальная часть является либо нейтральной, либо слабо полезной. [ 28 ]
Этот биологический процесс мутации представлен в популяционных моделях одним из двух способов, либо в качестве детерминированного давления рецидивирующей мутации на частотах аллелей, либо источник вариации. В детерминированной теории эволюция начинается с заранее определенного набора аллелей и продолжается путем сдвигов в непрерывных частотах, как если бы популяция бесконечна. Появление мутаций у отдельных лиц представлено «силой» или «давлением» мутации на уровне населения Полем Например, в классической модели баланса мутации и выбора , [ 29 ] Сила давления мутации толкает частоту аллеля вверх, а выбор против его вредных эффектов подталкивает частоту вниз, так что баланс достигается в равновесии, приведенный (в самом простом случае) на f = U/s.
Эта концепция давления мутации в основном полезна для рассмотрения последствий вредной мутации, такой как мутационная нагрузка и ее последствия для эволюции скорости мутации. [ 30 ] Трансформация популяций путем давления мутации маловероятна. Холдейн [ 31 ] утверждал, что это потребует высоких показателей мутаций, не сопровождаемых отбором, и Кимура [ 32 ] еще более пессимистично пришел к выводу, что даже это было маловероятно, так как процесс займет слишком много времени (см. Эволюцию путем давления мутации ).
Тем не менее, эволюция путем давления мутации возможна при некоторых обстоятельствах и давно была предложена в качестве возможной причины потери неиспользованных признаков. [ 33 ] Например, пигменты больше не полезны, когда животные живут в темноте пещер и имеют тенденцию терять. [ 34 ] Экспериментальный пример включает в себя потерю споруляции в экспериментальных популяциях B. subtilis . Споруляция является сложной чертой, кодируемой многими локусами, так что скорость мутации для потери признака была оценена как необычайно высокое значение, . [ 35 ] Потеря споруляции в этом случае может происходить путем рецидивирующей мутации, не требуя отбора для потери способности споруляции. Когда нет отбора для потери функции, скорость, с которой развивается потеря, зависит больше от скорости мутаций, чем от эффективного размера популяции , [ 36 ] Указывая, что это обусловлено большей мутацией, чем генетическим дрейфом.

Роль мутации как источника новизны отличается от этих классических моделей давления мутации. Когда генерические модели населения включают зависимый от скорости процесс мутационного введения или происхождения, т.е. процесс, который вводит новые аллели, включая нейтральные и полезные, то свойства мутации могут оказать более прямое влияние на скорость и направление эволюции. Даже если скорость мутации очень низкая. [ 37 ] [ 38 ] То есть спектр мутации может стать очень важным, в частности, смещения мутаций , предсказуемые различия в скоростях возникновения для различных типов мутаций, поскольку смещение в введении вариации может навязывать смещения на ход эволюции. [ 39 ]
Мутация играет ключевую роль в других классических и недавних теориях, в том числе сфокусирование Мюллера , субфункционализацию , концепцию Эйгена об ошибках катастрофы Линча и гипотезы о опасности .
Генетический дрейф
[ редактировать ]Генетический дрейф - это изменение частот аллелей, вызванные случайной выборкой . [ 40 ] То есть аллели в потомстве являются случайной выборкой среди родителей. [ 41 ] Генетический дрейф может привести к полному исчезновению вариантов генов и тем самым снизить генетическую изменчивость. В отличие от естественного отбора, что делает варианты генов более распространенными или менее распространенными в зависимости от их репродуктивного успеха, [ 42 ] Изменения, вызванные генетическим дрейфом, не обусловлены экологическим или адаптивным давлением и в равной степени могут сделать аллель более распространенным явлением менее распространенным.
Эффект генетического дрейфа больше для аллелей, присутствующих в нескольких копиях, чем когда аллель присутствует во многих копиях. Генетика популяции генетического дрейфа описывается с использованием либо разветвленных процессов , либо уравнения диффузии, описывающего изменения в частоте аллелей. [ 43 ] Райта и Морана Эти подходы обычно применяются к моделям Генетики населения . Предполагая, что генетический дрейф является единственной эволюционной силой, действующей на аллель, после того, как Т -поколения во многих реплицированных популяциях, начиная с частот аллелей P и Q, дисперсия частоты аллелей в этих популяциях является

Рональд Фишер придерживался мнения, что генетический дрейф играет наиболее незначительной роли в эволюции, и это оставалось доминирующим взглядом в течение нескольких десятилетий. Никакая популяционная генетика не давала генетического дрейфа центральной роли само по себе, но некоторые сделали генетический дрейф, важным в сочетании с другой не селективной силой. Теория смещающегося баланса утверждала Сьюолла Райта , что сочетание структуры популяции и генетического дрейфа было важным. Motoo Kimura утверждает Нейтральная теория молекулярной эволюции , что большинство генетических различий внутри и между популяциями вызваны комбинацией нейтральных мутаций и генетического дрейфа. [ 45 ]
Роль генетического дрейфа посредством ошибки отбора проб в эволюции подвергся критике Джоном Хиллеспи. [ 46 ] и придет , [ 47 ] которые утверждают, что выбор на связанных сайтах является более важной стохастической силой, выполняя работу, традиционно приписываемой генетическому дрейфу с помощью ошибки выборки. Математические свойства генетического черновика отличаются от свойств генетического дрейфа. [ 48 ] Направление случайного изменения частоты аллелей автокоррелируется по поколениям. [ 40 ]
Поток генов
[ редактировать ]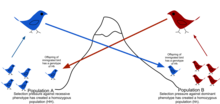
Из -за физических барьеров для миграции, наряду с ограниченной тенденцией для людей перемещать или распространяться ( с варенья ) и тенденцию оставаться или возвращаться в Натал Место ( филопатрия ), естественные популяции редко все промежуточные, как это может предположить в теоретических случайных моделях ( Панмикси ). [ 49 ] Обычно существует географический диапазон, в котором люди более тесно связаны друг с другом, чем те, которые случайно выбираются из населения в целом. Это описывается как степень, в которой популяция генетически структурирована. [ 50 ]

Генетическое структурирование может быть вызвано миграцией из -за исторического изменения климата , расширения диапазона видов или текущей доступности среды обитания . Поток генов затрудняется горными хребтами, океанами и пустынями или даже человеческими структурами, такими как великая стена Китая , которая препятствовала потоку растений. [ 51 ]
Поток генов - это обмен генами между популяциями или видами, разбивая структуру. Примеры потока генов внутри вида включают миграцию, а затем размножение организмов или обмен пыльцой . Перенос генов между видами включает образование гибридных организмов и горизонтальный перенос генов . Генетические модели популяции могут быть использованы для определения того, какие популяции показывают значительную генетическую изоляцию друг от друга, и для восстановления их истории. [ 52 ]
Возмещение населения изоляции приводит к депрессии инбридинга . Миграция в популяцию может ввести новые генетические варианты, [ 53 ] потенциально способствует эволюционному спасению . Если значительная часть людей или гаметов мигрирует, это также может изменить частоты аллелей, например, привести к миграционной нагрузке . [ 54 ]
При наличии потока генов другие барьеры для гибридизации между двумя расходящимися популяциями внешних необходимы видов, чтобы население стало новым видом .
Горизонтальный перенос генов
[ редактировать ]Горизонтальный перенос генов - это перенос генетического материала от одного организма в другой организм, который не является его потомством; Это наиболее распространено среди прокариот . [ 55 ] В медицине это способствует распространению устойчивости к антибиотикам , например, когда одна бактерия приобретает гены устойчивости, она может быстро перенести их на другие виды. [ 56 ] Горизонтальный перенос генов от бактерий к эукариотам, таким как дрожжи Saccharomyces cerevisiae и Adzuki Beang Beetle Callosobruchus chinensis также, также могли произойти. [ 57 ] [ 58 ] Примером более масштабных переводов являются эукариотические бделлоидные вращения , которые, по-видимому, получали ряд генов от бактерий, грибов и растений. [ 59 ] Вирусы также могут нести ДНК между организмами, позволяя переносить генов даже в биологических областях . [ 60 ] Крупномасштабный перенос генов также произошел между предками эукариотических клеток и прокариот во время приобретения хлоропластов и митохондрий . [ 61 ]
Связь
[ редактировать ]Если все гены находятся в равновесии сцепления , эффект аллеля в одном локусе может быть усреднен по всему генному пулу в других локусах. В действительности один аллель часто встречается в неравновесном сцеплении с генами в других локусах, особенно с генами, расположенными поблизости на одной и той же хромосоме. Рекомбинация слишком медленно разбивает эту неравновесную связь, чтобы избежать генетического автостопа , где аллель в одном локусе поднимается до высокой частоты, потому что он связан с аллелем под отбором в соседнем локусе. Связь также замедляет скорость адаптации, даже в сексуальных популяциях. [ 62 ] [ 63 ] [ 64 ] Эффект неравновесного сцепления на замедление скорости адаптивной эволюции возникает из -за сочетания эффекта холма -Робертсона (задержки в объединении полезных мутаций) и фонового отбора (задержки в отделении полезных мутаций от Deleyeterious Hitchhikers ).
Связь является проблемой для популяционных генетических моделей, которые обрабатывают один ген -локус одновременно. Однако он может быть использован как метод обнаружения действия естественного отбора с помощью селективных зачистков .
В крайнем случае небельной популяции , связь полна, и популяционные генетические уравнения могут быть получены и решены с точки зрения движущейся волны частот генотипа вдоль простого ландшафта фитнеса . [ 65 ] Большинство микробов , такие как бактерии , являются бесполые. Генетика популяции их адаптации имеет два контрастных режима. Когда продукт полезного уровня мутаций и численности населения невелик, бесполые популяции следуют «преемственному режиму» динамики фиксации происхождения, при этом частота адаптации сильно зависит от этого продукта. Когда продукт намного больше, бесполые популяции следуют режиму «параллельные мутации» с скоростью адаптации, менее зависящей от продукта, характеризующегося клональными помехами и появлением новой полезной мутации до того, как последняя была фиксирована .
Приложения
[ редактировать ]Объяснение уровней генетической вариации
[ редактировать ]Нейтральная теория предсказывает, что уровень разнообразия нуклеотидов в популяции будет пропорционален продукту размера популяции и уровня нейтральной мутации. Тот факт, что уровни генетического разнообразия варьируются гораздо меньше, чем размеры популяции, известен как «парадокс вариации». [ 66 ] Хотя высокие уровни генетического разнообразия были одним из первоначальных аргументов в пользу нейтральной теории, парадокс вариации был одним из самых сильных аргументов против нейтральной теории.
Понятно, что уровни генетического разнообразия сильно различаются в отношении вида в зависимости от скорости локальной рекомбинации, как из -за генетического автостопа , так и фона . Большинство современных решений парадокса вариации вызывают некоторый уровень отбора на связанных сайтах. [ 67 ] Например, один анализ предполагает, что более крупные популяции имеют более селективные развертки, которые устраняют более нейтральное генетическое разнообразие. [ 68 ] Негативная корреляция между уровнем мутаций и размером популяции также может способствовать. [ 69 ]
История жизни влияет на генетическое разнообразие больше, чем история популяции, например, R-стратеги имеют больше генетического разнообразия. [ 67 ]
Обнаружение выбора
[ редактировать ]Модели генетики популяции используются для вывода, какие гены проходят отбор. Одним из распространенных подходов является поиск областей неравновесного неравновесного сцепления и низкой генетической дисперсии вдоль хромосомы, для обнаружения недавних селективных зачистков .
Вторым распространенным подходом является тест Макдональда -Крейтмана , который сравнивает количество вариаций внутри вида ( полиморфизм ) с дивергенцией между видами (замены) в двух типах сайтов; предполагается, что он нейтрален. Как правило, синонимичные сайты считаются нейтральными. [ 70 ] Гены, перенесшие положительный отбор, имеют избыток дивергентных сайтов по сравнению с полиморфными сайтами. Тест также может быть использован для получения массовой оценки доли замен, которые фиксируются положительным отбором, α. [ 71 ] [ 72 ] Согласно нейтральной теории молекулярной эволюции , это число должно быть почти нулевым. Таким образом, высокие числа были интерпретированы как фальсификация нейтральной теории по всему геному. [ 73 ]
Демографический вывод
[ редактировать ]Самый простой тест на структуру популяции в сексуально воспроизводимых, диплоидных видах-это увидеть, следуют ли частоты генотипа Харди-Вейнберг пропорции как функцию частот аллелей. Например, в простейшем случае одного локуса с двумя аллелями, обозначенными A и A на частотах P и Q , случайное спаривание прогнозирует FREQ ( AA ) = P 2 Для AA гомозигот , freq ( aa ) = q 2 для гомозигот АА и Freq ( AA ) = 2 pq для гетерозигот . В отсутствие структуры населения пропорции Харди-Вайнберга достигаются в пределах 1–2 поколения случайного спаривания. Более того, существует избыток гомозигот, что указывает на структуру популяции. Степень этого избытка может быть определено количественно как коэффициент инбридинга, f .
Люди могут быть сгруппированы в K. субпопуляции [ 74 ] [ 75 ] Степень структуры популяции может быть затем рассчитана с использованием F -ST , которая является мерой доли генетической дисперсии, которая может быть объяснена структурой популяции. Структура генетической популяции может быть связана с географической структурой, и генетическая примеси может быть обнаружена .
Теория коалса связывает генетическое разнообразие в выборке с демографической историей популяции, из которой она была взята. Обычно он предполагает нейтралитет , и поэтому для таких анализов выбираются последовательности из более нейтрально развивающихся частей геномов. Его можно использовать для вывода взаимосвязи между видами ( филогенетикой ), а также структурой популяции, демографической истории (например, узкие места населения , рост популяции ), биологический рассеивание , динамика источника и зрителя [ 76 ] и интрогрессия внутри вида.
Другой подход к демографическому выводу зависит от частотного спектра аллеля . [ 77 ]
Эволюция генетических систем
[ редактировать ]Предполагая, что существуют локусы, которые контролируют саму генетическую систему, создаются генетические модели популяции для описания эволюции доминирования и других форм надежности , эволюции сексуального размножения и скорости рекомбинации, эволюции скорости мутаций , эволюции эволюционных конденсаторов , эволюция дорогостоящих сигнальных признаков , эволюция старения и эволюция сотрудничества . Например, большинство мутаций являются вредными, поэтому оптимальная скорость мутаций для вида может быть компромиссом между повреждением от высокой частоты вредных мутаций и метаболическими затратами на содержание систем для снижения скорости мутаций, таких как ферменты, восстанавливающие ДНК. [ 78 ]
Одним из важных аспектов таких моделей является то, что отбор достаточно силен, только для очистки вредных мутаций и, следовательно, одолеть мутационное смещение в сторону деградации, если коэффициент отбора S больше, чем обратный размер популяции . Это известно как дрейфующий барьер и связан с почти нейтральной теорией молекулярной эволюции . Теория дрейфовых барьеров прогнозирует, что виды с большими эффективными размерами популяции будут иметь высокотехнологичные, эффективные генетические системы, в то время как у людей с небольшими размерами популяции будут раздутые и сложные геномы, содержащие, например, интроны и транспонируемые элементы . [ 79 ] Однако, как это парадоксально, виды с большими размерами популяции могут быть настолько устойчивы к последствиям определенных типов ошибок, что они развивают более высокие частоты ошибок, например, в транскрипции и трансляции , чем небольшие популяции. [ 80 ]
Смотрите также
[ редактировать ]- Аллельный частотный спектр
- Ассортативное спаривание
- Баланс отбора
- Предвзятость в введении вариаций
- Теория коалесценции
- Генетика сохранения
- Конструктивная нейтральная эволюция
- Направленный выбор
- Распределение фитнес -эффектов (DFE)
- Гипотеза Дрифт-Барриер
- Эффективный размер населения
- Ошибка катастрофа
- Эволюционный ландшафт
- Фитнес -ландшафт
- Фиксация
- Эффект основателя
- Поток генов
- Генный пул
- Генетический дрейф
- Генетический автостоп
- Генетическая вариация
- Адорма Хадана
- Харди-Вайнберг Равновесие
- Идеализированное население
- Инбридинга
- Внедовая депрессия
- Модель бесконечных сайтов
- Внутригеномный конфликт
- Кин. Выбор
- Связь неравновесия
- Тест Макдональд -Крейтман
- Мейотический драйв
- Хпилот Мюллера
- Баланс мутации -выбора
- Гипотеза о опасности
- Естественный отбор
- Нейтральная теория молекулярной эволюции
- Панмиксия
- Расширяющая депрессия
- Полиморфизм
- Полигенная адаптация
- Численность населения
- Количественная генетика
- Процесс безудержного (Fisherian Runaway)
- Выборочный разверток
- Эгоистичный генетический элемент
- Сексуальный отбор
- Изменение теории баланса
- Субфункционализация
- Supergene (совместный генный комплекс)
Ссылки
[ редактировать ]- ^ «Генетика населения - последние исследования и новости» . www.nature.com . Получено 2018-01-29 .
- ^ Сандо, Мария Р .; Brandvain, Yaniv; DHOHER, SUMIT; Фицпатрик, Кортни Л.; Голдберг, Эмма Э.; Стерн, Кейтлин А.; Ван Клив, Джереми; Да, Д. Джастин (9 декабря 2014 г.). «Не просто теория - полезность математических моделей в эволюционной биологии» . PLOS Биология . 12 (12): E1002017. doi : 10.1371/journal.pbio.1002017 . PMC 4260780 . PMID 25489940 .
- ^ Ewens, WJ (2004). Математическая популяция генетика (2 -е изд.). Нью -Йорк: Спрингер. ISBN 978-0-387-20191-7 .
- ^ Jump up to: а беременный в Боулер, Питер Дж. (2003). Эволюция: история идеи (3 -е изд.). Беркли: Университет Калифорнийской прессы. С. 325–339 . ISBN 978-0-520-23693-6 .
- ^ Jump up to: а беременный в Ларсон, Эдвард Дж. (2004). Эволюция: замечательная история научной теории (современная библиотека изд.). Нью -Йорк: современная библиотека. С. 221–243 . ISBN 978-0-679-64288-6 .
- ^ Битти, Джон (1986). «Синтез и синтетическая теория». Интеграция научных дисциплин . Наука и философия. Тол. 2. Springer Netherlands. С. 125–135. doi : 10.1007/978-94-010-9435-1_7 . ISBN 9789024733422 .
- ^ Майр, Эрнст ; Provine, William B., eds. (1998). Эволюционный синтез: перспективы объединения биологии ([Новое изд. Эд.). Кембридж, Массачусетс: издательство Гарвардского университета. С. 295–298. ISBN 9780674272262 .
- ^ Jump up to: а беременный Provine, WB (1988). «Прогресс в эволюции и значении в жизни». Эволюционный прогресс . Университет Чикагской Прессы. С. 49–79.
- ^ Jump up to: а беременный Провин, Уильям Б. (1978). «Роль математических генетиков популяции в эволюционном синтезе 1930 -х и 1940 -х годов». Исследования истории биологии . 2 : 167–192. PMID 11610409 .
- ^ Ford, EB (1975) [1964]. Экологическая генетика (4 -е изд.). Лондон: Чепмен и Холл. С. 1ff.
- ^ Mayr, Ernst (1988). На пути к новой философии биологии: наблюдения эволюциониста . Кембридж, штат Массачусетс: издательство Belknap of Harvard University Press . п. 402. ISBN 978-0-674-89665-9 .
- ^ Майр, Эрнст ; Provine, William B., eds. (1998). Эволюционный синтез: перспективы объединения биологии ([Новое изд. Эд.). Кембридж, Массачусетс: издательство Гарвардского университета. С. 338–341. ISBN 9780674272262 .
- ^ Jump up to: а беременный в McCandlish, David M.; Столцфус, Арлин (сентябрь 2014 г.). «Моделирование эволюции с использованием вероятности фиксации: история и последствия». Ежеквартальный обзор биологии . 89 (3): 225–252. doi : 10.1086/677571 . PMID 25195318 . S2CID 19619966 .
- ^ Кроу, Джеймс Ф. (2010). «Райт и Фишер на инбридинге и случайном дрейфе» . Генетика . 184 (3): 609–611. doi : 10.1534/Genetics.109.110023 . ISSN 0016-6731 . PMC 2845331 . PMID 20332416 .
- ^ Коробки, мечта; Барбадилла, Антонио (2017). "Молекулярные генетические народы " Генетика 205 (3): 1003–1035. два 10.1534/genetics.116.196493: 5340319PMC 28270526PMID
- ^ Haldane, JBS (1927). «Математическая теория естественного и искусственного отбора, часть V: отбор и мутация». Математические разбирательства Кембриджского философского общества . 23 (7): 838–844. Bibcode : 1927pcps ... 23..838h . doi : 10.1017/s0305004100015644 . S2CID 86716613 .
- ^ Orr, HA (2010). «Генетика популяции полезных мутаций» . Философские транзакции Королевского общества B: биологические науки . 365 (1544): 1195–1201. doi : 10.1098/rstb.2009.0282 . PMC 2871816 . PMID 20308094 .
- ^ Hermisson, J.; Пеннингс, PS (2005). «Мягкие зачистки: генетика адаптации молекулярной популяции от постоянной генетической вариации» . Генетика . 169 (4): 2335–2352. doi : 10.1534/Genetics.104.036947 . PMC 1449620 . PMID 15716498 .
- ^ Гиллеспи, Джон (2004). Генетика популяции: краткое руководство (2 -е изд.). Johns Hopkins University Press. ISBN 978-0-8018-8008-7 .
- ^ Мико, И. (2008). «Эпистаз: взаимодействие генов и эффекты фенотипа» . Природное образование . 1 (1): 197.
- ^ Бергер, Д.; Postma, E. (13 октября 2014 г.). «Предвзятые оценки снижения возврата эпистаза ? Генетика . 198 (4): 1417–1420. doi : 10.1534/Genetics.114.169870 . PMC 4256761 . PMID 25313131 .
- ^ Kouyos, Roger D.; Сайландер, Олин К.; Бонхоффер, Себастьян (июнь 2007 г.). «Эпистаз между вредными мутациями и эволюцией рекомбинации». Тенденции в экологии и эволюции . 22 (6): 308–315. doi : 10.1016/j.tree.2007.02.014 . PMID 17337087 .
- ^ Кроу, JF (5 августа 1997 г.). «Высокий уровень спонтанной мутации: это риск для здоровья?» Полем Труды Национальной академии наук Соединенных Штатов Америки . 94 (16): 8380–8386. Bibcode : 1997pnas ... 94.8380c . doi : 10.1073/pnas.94.16.8380 . PMC 33757 . PMID 9237985 .
- ^ Гастингс, PJ; Люпский, младший; Розенберг, С.М.; IRA, G. (2009). «Механизмы изменения числа копий генов» . Nature Reviews Genetics . 10 (8): 551–564. doi : 10.1038/nrg2593 . PMC 2864001 . PMID 19597530 .
- ^ М., Лонг; Betrán, E.; Торнтон, К.; Ван, В. (ноябрь 2003 г.). «Происхождение новых генов: проблески от молодых и старых». НАТ Преподобный Генет . 4 (11): 865–75. doi : 10.1038/nrg1204 . PMID 14634634 . S2CID 33999892 .
- ^ Лю, н.; Окамура, К.; Тайлер, DM; Филлипс; Чунг; Лай (2008). «Эволюция и функциональная диверсификация генов микроРНК животных» . Клеточные исследования . 18 (10): 985–996. doi : 10.1038/cr.2008.278 . PMC 2712117 . PMID 18711447 .
- ^ McLysaght, Aoife; Херст, Лоуренс Д. (25 июля 2016 г.). «Открытые вопросы в изучении генов de novo: что, как и почему» . Nature Reviews Genetics . 17 (9): 567–578. doi : 10.1038/nrg.2016.78 . PMID 27452112 . S2CID 6033249 .
- ^ Сойер, SA; Parsch, J.; Zhang, Z.; Hartl, DL (2007). «Распространенность положительного отбора среди почти нейтральных замен аминокислот у дрозофилы» . Труды Национальной академии наук . 104 (16): 6504–6510. Bibcode : 2007pnas..104.6504S . doi : 10.1073/pnas.0701572104 . ISSN 0027-8424 . PMC 1871816 . PMID 17409186 .
- ^ Кроу, Джеймс Ф.; Kimura, Motoo (1970). Введение в теорию генетики популяции ([Перепечатка] изд.). Нью -Джерси: Блэкберн Пресс. ISBN 9781932846126 .
- ^ Линч, Майкл (август 2010 г.). «Эволюция скорости мутации» . Тенденции в генетике . 26 (8): 345–352. doi : 10.1016/j.tig.2010.05.003 . PMC 2910838 . PMID 20594608 .
- ^ JBS Haldane (1932). Причины эволюции . Longmans, Green and Co., Нью -Йорк.
- ^ М. Кимура (1980). «Среднее время до фиксации мутантного аллеля в конечной популяции под постоянным давлением мутации: исследования с помощью аналитических, численных и псевдопользующих методов» . Proc Natl Acad Sci USA . 77 (1): 522–526. Bibcode : 1980pnas ... 77..522K . doi : 10.1073/pnas.77.1.522 . PMC 348304 . PMID 16592764 .
- ^ Haldane, JBS (1933). «Часть, которую играет повторяющаяся мутация в эволюции». Американский натуралист . 67 (708): 5–19. doi : 10.1086/280465 . JSTOR 2457127 . S2CID 84059440 .
- ^ Протас, Мередит; Conrad, M.; Гросс, JB; Табин, C.; Боровский Р. (2007). «Регрессивная эволюция в мексиканской пещере Тетры, Астианакс Мексика» . Текущая биология . 17 (5): 452–454. Bibcode : 2007cbio ... 17..452p . doi : 10.1016/j.cub.2007.01.051 . PMC 2570642 . PMID 17306543 .
- ^ H. Maughan, J. MASEL, CW Birky, Jr. and WL Nicholson (2007). «Роли накопления и отбора мутаций в потере споруляции в экспериментальных популяциях Bacillus subtilis» . Генетика . 177 (2): 937–48. doi : 10.1534/Genetics.107.075663 . PMC 2034656 . PMID 17720926 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ MASEL, J .; Король, OD; Maughan, H. (2007). «Потеря адаптивной пластичности в течение длительных периодов окружающей среды» . Американский натуралист . 169 (1): 38–46. doi : 10.1086/510212 . PMC 1766558 . PMID 17206583 .
- ^ К. Гомес, Дж. Бертрам и Дж. Масел (2020). «Предвзятость мутации может формировать адаптацию в больших бесполых популяциях, испытывающих клональные помехи» . Прокурор R. Soc. Беременный 287 (1937): 20201503. DOI : 10.1098/rspb.2020.1503 . PMC 7661309 . PMID 33081612 .
- ^ AV Cano, H. Rozhonova, A. Stoltzfus, DM McCandlish и JL Payne (2022-02-10). «Мутационный смещение формирует спектр адаптивных замен» . Proc Natl Acad Sci USA . 119 (7). Bibcode : 2022pnas..11919720C . doi : 10.1073/pnas.2119720119 . PMC 8851560 . PMID 35145034 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Stoltzfus, A.; Yampolsky, Ly (2009). «Возможность подъема: мутация как причина нерушины в эволюции» . Журнал наследственности . 100 (5): 637–647. doi : 10.1093/jhered/esp048 . PMID 19625453 .
- ^ Jump up to: а беременный MASEL, J. (2011). «Генетический дрейф» . Текущая биология . 21 (20): R837 - R838. Bibcode : 2011cbio ... 21.r837m . doi : 10.1016/j.cub.2011.08.007 . PMID 22032182 .
- ^ Futuyma, Douglas (1998). Эволюционная биология . Sinauer Associates . п. Глоссарий. ISBN 978-0-87893-189-7 .
- ^ Аверс, Шарлотта (1989). Процесс и шаблон в эволюции . Издательство Оксфордского университета.
- ^ Wahl, LM (2011). «Фиксация, когда N и S различаются: классические подходы дают элегантные новые результаты» . Генетика . 188 (4): 783–785. doi : 10.1534/Genetics.111.131748 . PMC 3176088 . PMID 21828279 .
- ^ Бартон, Николас Х.; Бриггс, Дерек, Эг; Эйзен, Джонатан А.; Гольдштейн, Дэвид Б.; Патель, Нипам Х. (2007). Эволюция Cold Spring Harbor Laboratory Press. п. 417. ISBN 978-0-87969-684-9 .
- ^ Futuyma, Douglas (1998). Эволюционная биология . Sinauer Associates . п. 320. ISBN 978-0-87893-189-7 .
- ^ Гиллеспи, Дж. Х. (2000). «Генетический дрейф в бесконечной популяции: модель псевдохичхийка» . Генетика . 155 (2): 909–919. doi : 10.1093/Genetics/155.2.909 . PMC 1461093 . PMID 10835409 .
- ^ Провина, Уильям Б. ошибка "случайного генетического дрейфа" . CreateSpace.
- ^ Нехер, Ричард А.; Шрайман, Борис И. (август 2011 г.). «Генетический проект и квази-нейтральность в крупных факультативно сексуальных популяциях» . Генетика . 188 (4): 975–996. Arxiv : 1108.1635 . doi : 10.1534/genetics.111.128876 . ISSN 0016-6731 . PMC 3176096 . PMID 21625002 .
- ^ Бустон, премьер -министр; Пилкингтон, JG; и др. (2007). «Являются ли группы клоун -рыб, состоящие из близких родственников? Анализ микросателлитной ДНК -виоации в амфиприонной прокуле ». Молекулярная экология . 12 (3): 733–742. doi : 10.1046/j.1365-294x.2003.01762.x . PMID 12675828 . S2CID 35546810 .
- ^ Repaci, v.; Стоу, AJ; Бриско, Д.А. (2007). «Тонкомасштабная генетическая структура, соучредительное и множественное спаривание в австралийской аллодапиновой пчел ( Ramphocinclus brachyurus )». Журнал зоологии . 270 (4): 687–691. doi : 10.1111/j.1469-7998.2006.00191.x .
- ^ Jump up to: а беременный Su, H.; Qu, L.-J.; Он, К.; Zhang, Z.; Ван, J; Chen, Z.; Гу, Х. (2003). «Великая китайская стена: физический барьер для потока генов?». Наследственность . 90 (3): 212–219. doi : 10.1038/sj.hdy.6800237 . ISSN 0018-067X . PMID 12634804 . S2CID 13367320 .
- ^ Gravel, S. (2012). «Популяционная генетика модели местного происхождения» . Генетика . 1202 (2): 607–619. Arxiv : 1202.4811 . BIBCODE : 2012ARXIV1202.4811G . doi : 10.1534/Genetics.112.139808 . PMC 3374321 . PMID 22491189 .
- ^ Morjan, C.; Ризеберг Л. (2004). «Как виды эволюционируют коллективно: последствия потока генов и отбора для распространения выгодных аллелей» . Молекулярная экология . 13 (6): 1341–56. Bibcode : 2004molec..13.1341M . doi : 10.1111/j.1365-294x.2004.02164.x . PMC 2600545 . PMID 15140081 .
- ^ Болник, Даниэль I.; Носил, Патрик (сентябрь 2007 г.). «Естественный отбор в популяциях, подверженных миграционной нагрузке» . Эволюция 61 (9): 2229–2243. doi : 10.1111/j.1558-5646.2007.00179.x . PMID 17767592 . S2CID 25685919 .
- ^ Баучер, Ян; Дуади, Кристоф Дж.; Папке, Р. Тейн; Уолш, Дэвид А.; Будро, Мэри Эллен Р.; Nesbø, Camilla L.; Дело, Ребекка Дж.; Doolittle, W. Ford (2003). «Боковой перенос генов и происхождение прокариотических групп». Ежегодный обзор генетики . 37 (1): 283–328. doi : 10.1146/annurev.genet.37.050503.084247 . ISSN 0066-4197 . PMID 14616063 .
- ^ Уолш, Т. (2006). «Комбинаторная генетическая эволюция мультизистентности». Текущее мнение о микробиологии . 9 (5): 476–82. doi : 10.1016/j.mib.2006.08.009 . PMID 16942901 .
- ^ Kondo, N.; Nikoh, N.; Ijichi, N.; Shimada, M.; Фукацу Т. (2002). «Фрагмент генома эндосимбионта Wolbachia, перенесенный в x хромосому насекомого хозяина» . Труды Национальной академии наук . 99 (22): 14280–14285. Bibcode : 2002pnas ... 9914280K . doi : 10.1073/pnas.222228199 . ISSN 0027-8424 . PMC 137875 . PMID 12386340 .
- ^ Sprague, G. (1991). «Генетический обмен между королевствами». Текущее мнение в области генетики и развития . 1 (4): 530–533. doi : 10.1016/s0959-437x (05) 80203-5 . PMID 1822285 .
- ^ Гладишев, EA; Meselson, M.; Arkhipova, IR (2008). «Массивный горизонтальный перенос генов у бделлоидных оздоровок» . Наука . 320 (5880): 1210–1213. BIBCODE : 2008SCI ... 320.1210G . doi : 10.1126/science.1156407 . ISSN 0036-8075 . PMID 18511688 . S2CID 11862013 .
- ^ Балдо, А.; McClure, M. (1 сентября 1999 г.). «Эволюция и горизонтальная перенос генов, кодирующих датпазу у вирусов и их хозяев» . Журнал вирусологии . 73 (9): 7710–7721. doi : 10.1128/jvi.73.9.7710-7721.1999 . PMC 104298 . PMID 10438861 .
- ^ Пул, А.; Пенни Д. (2007). «Оценка гипотез для происхождения эукариот». Биологии . 29 (1): 74–84. doi : 10.1002/bies.20516 . PMID 17187354 .
- ^ Вайсман, Д.Б.; Hallatschek, O. (15 января 2014 г.). «Скорость адаптации в больших сексуальных популяциях с линейными хромосомами» . Генетика . 196 (4): 1167–1183. doi : 10.1534/Genetics.113.160705 . PMC 3982688 . PMID 24429280 .
- ^ Вайсман, Даниэль Б.; Бартон, Николас Х.; Маквин, Гил (7 июня 2012 г.). «Ограничения на скорость адаптивной замены в сексуальных популяциях» . PLOS Genetics . 8 (6): E1002740. doi : 10.1371/journal.pgen.1002740 . PMC 3369949 . PMID 22685419 .
- ^ Нехер, Ра; Шрайман, Би; Фишер, DS (30 ноября 2009 г.). «Скорость адаптации в больших сексуальных популяциях» . Генетика . 184 (2): 467–481. Arxiv : 1108.3464 . doi : 10.1534/Genetics.109.109009 . PMC 2828726 . PMID 19948891 .
- ^ Десаи, Майкл М.; Фишер, Даниэль С. (2007). «Полезный баланс выбора мутаций и влияние связи на положительный отбор» . Генетика . 176 (3): 1759–1798. doi : 10.1534/Genetics.106.067678 . PMC 1931526 . PMID 17483432 .
- ^ Lewontin, RC (1973). Генетическая основа эволюционных изменений ([4 -я печать.] Ред.). Нью -Йорк: издательство Колумбийского университета. ISBN 978-0231033923 .
- ^ Jump up to: а беременный Эллегрен, Ганс; Галтье, Николас (6 июня 2016 г.). «Детерминанты генетического разнообразия» . Nature Reviews Genetics . 17 (7): 422–433. doi : 10.1038/nrg.2016.58 . PMID 27265362 . S2CID 23531428 .
- ^ Corbett-Detig, Russell B.; Хартл, Даниэль Л.; Сактон, Тимоти Б.; Бартон, Ник Х. (10 апреля 2015 г.). «Естественный отбор ограничивает нейтральное разнообразие среди широкого спектра видов» . PLOS Биология . 13 (4): E1002112. doi : 10.1371/journal.pbio.1002112 . PMC 4393120 . PMID 25859758 .
- ^ Sung, W.; Акерман, MS; Миллер, SF; Доак, Тг; Линч М. (17 октября 2012 г.). «Гипотеза и эволюция с температурой мутации» (PDF) . Труды Национальной академии наук . 109 (45): 18488–18492. Bibcode : 2012pnas..10918488S . doi : 10.1073/pnas.1216223109 . PMC 3494944 . PMID 23077252 .
- ^ Charlesworth, J. Eyre-Walker (2008). «Тест Макдональда -Крейтмана и слегка вредные мутации» . Молекулярная биология и эволюция . 25 (6): 1007–1015. doi : 10.1093/molbev/msn005 . PMID 18195052 .
- ^ Эйр-Уокер, А. (2006). «Геномная скорость адаптивной эволюции» (PDF) . Тенденции в экологии и эволюции . 21 (10): 569–575. doi : 10.1016/j.tree.2006.06.015 . PMID 16820244 .
- ^ Смит, NGC; Эйр-Уокер, А. (2002). «Адаптивная эволюция белка у дрозофилы». Природа . 415 (6875): 1022–1024. Bibcode : 2002natur.415.1022S . doi : 10.1038/4151022a . PMID 11875568 . S2CID 4426258 .
- ^ Hahn, MW (2008). «На пути к теории отбора молекулярной эволюции» . Эволюция 62 (2): 255–265. doi : 10.1111/j.1558-5646.2007.00308.x . PMID 18302709 . S2CID 5986211 .
- ^ Причард, JK; Стивенс, М.; Доннелли, П. (июнь 2000 г.). «Вывод структуры популяции с использованием данных с мультилокусным генотипом» . Генетика . 155 (2): 945–959. doi : 10.1093/Genetics/155.2.945 . ISSN 0016-6731 . PMC 1461096 . PMID 10835412 .
- ^ Верити, Роберт; Николс, Ричард А. (август 2016 г.). «Оценка количества субпопуляций (k) в структурированных популяциях» . Генетика . 203 (4): 1827–1839. doi : 10.1534/Genetics.115.180992 . ISSN 0016-6731 . PMC 4981280 . PMID 27317680 .
- ^ Манлик, Оливер; Чабан, Дельфина; Даниэль, Клэр; Bejder, Lars; Аллен, Саймон Дж.; Шервин, Уильям Б. (13 ноября 2018 г.). «Демография и генетика предполагают изменение динамики источника дельфина - SINK с последствиями для сохранения». Морские млекопитающие . 35 (3): 732–759. doi : 10.1111/mms.12555 . S2CID 92108810 .
- ^ Гутенкунст, Райан Н.; Эрнандес, Райан Д.; Уильямсон, Скотт Х.; Бустаманте, Карлос Д.; Маквин, Гил (23 октября 2009 г.). «Вывод совместной демографической истории множественных популяций из многомерных данных частоты SNP» . PLOS Genetics . 5 (10): E1000695. Arxiv : 0909.0925 . doi : 10.1371/journal.pgen.1000695 . PMC 2760211 . PMID 19851460 .
- ^ Sniegowski, P. (2000). «Эволюция скорости мутаций: отделение причин от последствий». Биологии . 22 (12). Gerrish P.; Джонсон Т ;.. Бритва А.: 1057–1066. doi : 10.1002/1521-1878 (200012) 22:12 <1057 :: AID-bies3> 3.0.co; 2-W . PMID 11084621 . S2CID 36771934 .
- ^ Линч, Майкл; Конери, Джон С. (2003). «Происхождение сложности генома». Наука . 302 (5649): 1401–1404. Bibcode : 2003sci ... 302.1401L . Citeseerx 10.1.1.135.974 . doi : 10.1126/science.1089370 . PMID 14631042 . S2CID 11246091 .
- ^ Rajon, E.; MASEL, J. (3 января 2011 г.). «Эволюция частоты молекулярных ошибок и последствия для развития» . Труды Национальной академии наук . 108 (3): 1082–1087. Bibcode : 2011pnas..108.1082R . doi : 10.1073/pnas.1012918108 . PMC 3024668 . PMID 21199946 .
Внешние ссылки
[ редактировать ]- Учебники по генетике населения (архивировано 23 января 2015 г.)
- Генетика молекулярной популяции
- База данных частот аллелей в Йельском университете
- Ehstrafd.org - база данных частот аллелей в Земле человека (архивировано 13 июля 2009 г.)
- История генетики популяции
- Как отбор меняется генетический состав популяции , видео лекции Стивена С. Стернса ( Йельский университет )
- National Geographic : Атлас человеческого путешествия ( гаплогруппы ) карты миграции на основе