Нейромедиатор

Нейромедиатор , — это сигнальная молекула секретируемая нейроном для воздействия на другую клетку через синапс . Клетка, получающая сигнал, или клетка-мишень, может быть другим нейроном, но также может быть железистой или мышечной клеткой . [1]
Нейромедиаторы высвобождаются из синаптических везикул в синаптическую щель , где они могут взаимодействовать с рецепторами нейромедиаторов на клетке-мишени. Эффект нейромедиатора на клетку-мишень определяется рецептором, с которым он связывается. Многие нейромедиаторы синтезируются из простых и многочисленных предшественников, таких как аминокислоты , которые легко доступны и часто требуют небольшого количества стадий биосинтеза для преобразования.
Нейротрансмиттеры необходимы для функционирования сложных нервных систем. Точное количество уникальных нейротрансмиттеров у человека неизвестно, но идентифицировано более 100. [2] Общие нейротрансмиттеры включают глутамат , ГАМК , ацетилхолин , глицин и норадреналин .
Механизм и цикл
[ редактировать ]Синтез
[ редактировать ]Нейротрансмиттеры обычно синтезируются в нейронах и состоят из молекул-предшественников, которые в изобилии обнаруживаются в клетке, или происходят из них. Классы нейромедиаторов включают аминокислоты , моноамины и пептиды . Моноамины синтезируются путем изменения одной аминокислоты. Например, предшественником серотонина является аминокислота триптофан. Пептидные трансмиттеры, или нейропептиды, представляют собой белковые трансмиттеры, которые часто высвобождаются вместе с другими трансмиттерами, оказывая модулирующее действие. [3] Пуриновые нейротрансмиттеры, такие как АТФ, происходят из нуклеиновых кислот. Другие нейротрансмиттеры состоят из продуктов метаболизма, таких как оксид азота и окись углерода . [ нужна ссылка ]
| Примеры | |
|---|---|
| Аминокислота | глицин , глутамат |
| Моноамины | серотонин , адреналин , дофамин |
| Пептиды | вещество Р , опиоиды |
| Пурины | АТФ , ГТП |
| Другой | оксид азота , окись углерода |
Хранилище
[ редактировать ]Нейротрансмиттеры обычно хранятся в синаптических пузырьках , сгруппированных вблизи клеточной мембраны в конце аксона пресинаптического нейрона. Однако некоторые нейротрансмиттеры, такие как метаболические газы окись углерода и оксид азота, синтезируются и высвобождаются сразу после возникновения потенциала действия, даже не откладываясь в везикулах. [4]
Выпускать
[ редактировать ]Как правило, нейромедиатор высвобождается в пресинаптическом терминале в ответ на электрический сигнал, называемый потенциалом действия, в пресинаптическом нейроне. Однако «базовое» высвобождение низкого уровня происходит и без электрической стимуляции. Нейромедиаторы высвобождаются и диффундируют через синаптическую щель , где они связываются со специфическими рецепторами на мембране постсинаптического нейрона. [5]
Рецепторное взаимодействие
[ редактировать ]После попадания в синаптическую щель нейротрансмиттеры диффундируют через синапс, где они могут взаимодействовать с рецепторами клетки-мишени. Эффект нейромедиатора зависит от идентичности рецепторов клетки-мишени, присутствующих в синапсе. В зависимости от рецептора связывание нейротрансмиттеров может вызывать возбуждение , торможение или модуляцию постсинаптического нейрона. Дополнительную информацию см. ниже. [ нужна ссылка ]
Устранение
[ редактировать ]
Чтобы избежать постоянной активации рецепторов постсинаптической клетки или клетки-мишени, нейротрансмиттеры должны быть удалены из синаптической щели. [6] Нейромедиаторы удаляются посредством одного из трех механизмов:
- Диффузия – нейромедиаторы выходят из синаптической щели, где поглощаются глиальными клетками . Эти глиальные клетки, обычно астроциты , поглощают избыток нейромедиаторов.
- Астроциты, тип глиальных клеток головного мозга, активно участвуют в синаптической коммуникации посредством астроцитарной диффузии или глиотрансмиссии . Нейрональная активность вызывает повышение уровня астроцитарного кальция, вызывая высвобождение глиотрансмиттеров, таких как глутамат , АТФ и D-серин. Эти глиотрансмиттеры диффундируют во внеклеточное пространство, взаимодействуя с близлежащими нейронами и влияя на синаптическую передачу. Регулируя уровни внеклеточных нейромедиаторов, астроциты помогают поддерживать правильную синаптическую функцию. Эта двунаправленная связь между астроцитами и нейронами усложняет передачу сигналов в мозгу, что влияет на функции мозга и неврологические расстройства. [7] [8]
- Ферментативная деградация – белки, называемые ферментами, разрушают нейротрансмиттеры.
- Обратный захват – нейромедиаторы реабсорбируются в пресинаптическом нейроне. Транспортеры, или мембранные транспортные белки , перекачивают нейротрансмиттеры из синаптической щели обратно в окончания аксонов (пресинаптический нейрон), где они сохраняются для повторного использования.
Например, ацетилхолин удаляется путем расщепления его ацетильной группы ферментом ацетилхолинэстеразой ; оставшийся холин затем поглощается и перерабатывается пресинаптическим нейроном для синтеза большего количества ацетилхолина . [9] Другие нейротрансмиттеры способны диффундировать из целевых синаптических соединений и выводятся из организма через почки или разрушаются в печени. Каждый нейромедиатор имеет очень специфические пути деградации в регуляторных точках, на которые может воздействовать регуляторная система организма или лекарства. Кокаин блокирует транспортер дофамина, ответственный за обратный захват дофамина. Без транспортера дофамин гораздо медленнее диффундирует из синаптической щели и продолжает активировать дофаминовые рецепторы на клетке-мишени. [10]
Открытие
[ редактировать ]До начала 20-го века ученые предполагали, что большая часть синаптической связи в мозге является электрической. Однако в ходе гистологического исследования Рамона-и-Кахаля был обнаружен разрыв между нейронами размером от 20 до 40 нм, известный сегодня как синаптическая щель . Наличие такого разрыва предполагало связь через химические посланники, пересекающие синаптическую щель, а в 1921 году немецкий фармаколог Отто Леви подтвердил, что нейроны могут общаться, выделяя химические вещества. В ходе серии экспериментов с блуждающими нервами лягушек Леви смог вручную замедлить частоту сердечных сокращений лягушек, контролируя количество физиологического раствора, окружающего блуждающий нерв. По завершении этого эксперимента Лоуи заявил, что симпатическая регуляция сердечной функции может осуществляться посредством изменения концентрации химических веществ. Кроме того, Отто Леви приписывают открытие ацетилхолина (АХ) – первого известного нейромедиатора. [11]
Идентификация
[ редактировать ]Для идентификации нейромедиаторов обычно учитывают следующие критерии:
- Синтез: химическое вещество должно производиться внутри нейрона или присутствовать в нем в виде молекулы-предшественника.
- Высвобождение и реакция. Когда нейрон активируется, химическое вещество должно высвободиться и вызвать реакцию в клетках-мишенях или нейронах.
- Экспериментальный ответ: Применение химического вещества непосредственно к клеткам-мишеням должно вызвать тот же ответ, что и при естественном высвобождении химического вещества из нейронов.
- Механизм удаления: Должен существовать механизм удаления нейромедиатора из места его действия после завершения его сигнальной роли. [12]
Однако, учитывая достижения фармакологии , генетики и химической нейроанатомии , термин «нейромедиатор» можно применять к химическим веществам, которые:
- Переносят сообщения между нейронами посредством воздействия на постсинаптическую мембрану.
- Оказывают незначительное влияние или вообще не влияют на мембранное напряжение, но имеют общую несущую функцию, такую как изменение структуры синапса.
- Общайтесь, отправляя сообщения в обратном направлении, которые влияют на высвобождение или повторный захват передатчиков.
Анатомическую локализацию нейротрансмиттеров обычно определяют с помощью иммуноцитохимических методов, которые определяют расположение либо самих веществ-медиаторов, либо ферментов, участвующих в их синтезе. Иммуноцитохимические методы также показали, что многие медиаторы, особенно нейропептиды , локализованы совместно, то есть нейрон может высвобождать более одного медиатора из своего синаптического окончания . [13] различные методы и эксперименты, такие как окрашивание можно использовать Для идентификации нейротрансмиттеров во всей центральной нервной системе , стимуляция и сбор . [14]
Действия
[ редактировать ]Нейроны общаются друг с другом через синапсы — специализированные точки контакта, через которые нейротрансмиттеры передают сигналы. Когда потенциал действия достигает пресинаптической терминали , потенциал действия может вызвать высвобождение нейротрансмиттеров в синаптическую щель. Эти нейротрансмиттеры затем связываются с рецепторами на постсинаптической мембране, воздействуя на принимающий нейрон либо тормозным , либо возбуждающим образом. Если общие возбуждающие влияния перевешивают тормозящие влияния, принимающий нейрон может генерировать собственный потенциал действия, продолжая передачу информации следующему нейрону в сети. Этот процесс обеспечивает поток информации и формирование сложных нейронных сетей. [15]
Модуляция
[ редактировать ]Нейромедиатор может оказывать возбуждающее, тормозящее или модулирующее действие на клетку-мишень. Эффект определяется рецепторами, с которыми взаимодействует нейромедиатор на постсинаптической мембране. Нейромедиатор влияет на трансмембранный поток ионов, увеличивая (возбуждая) или уменьшая (ингибируя) вероятность того, что клетка, с которой он вступает в контакт, выработает потенциал действия. Синапсы, содержащие рецепторы с возбуждающим эффектом, называются синапсами I типа, а синапсы типа II содержат рецепторы с тормозящим эффектом. [16] Таким образом, несмотря на большое разнообразие синапсов, все они передают сообщения только этих двух типов. Оба типа отличаются внешним видом и преимущественно располагаются на разных участках нейронов, находящихся под его воздействием. [17] Рецепторы с модулирующим эффектом распространяются по всем синаптическим мембранам, а связывание нейротрансмиттеров приводит в движение сигнальные каскады, которые помогают клетке регулировать ее функцию. [18] Связывание нейротрансмиттеров с рецепторами с модулирующим эффектом может иметь множество результатов. Например, это может привести к увеличению или снижению чувствительности к будущим стимулам за счет привлечения большего или меньшего количества рецепторов к синаптической мембране.
Синапсы типа I (возбуждающие) обычно расположены на стержнях или шипах дендритов, тогда как синапсы типа II (тормозящие) обычно расположены на теле клетки. Кроме того, синапсы I типа имеют круглые синаптические везикулы, тогда как везикулы синапсов II типа уплощены. Материал на пресинаптических и постсинаптических мембранах в синапсе I типа более плотный, чем в синапсе II типа, а синаптическая щель I типа шире. Наконец, активная зона синапса типа I больше, чем активная зона синапса типа II.
Разное расположение синапсов типа I и типа II делит нейрон на две зоны: возбуждающее дендритное дерево и тело тормозной клетки. С точки зрения торможения возбуждение поступает по дендритам и распространяется на аксонный бугорок , вызывая потенциал действия . Если сообщение необходимо остановить, лучше всего остановить его, применив торможение к телу клетки, вблизи аксонного холмика, где возникает потенциал действия. Другой способ концептуализировать возбуждающе-тормозное взаимодействие — представить себе, как возбуждение преодолевает торможение. Если тело клетки обычно находится в заторможенном состоянии, единственный способ создать потенциал действия на аксонном холмике — это уменьшить торможение тела клетки. В этой стратегии «открой ворота» возбуждающее сообщение похоже на скаковую лошадь, готовую бежать по дорожке, но сначала необходимо убрать тормозящие стартовые ворота. [19]
Действия нейромедиаторов
[ редактировать ]Как объяснялось выше, единственным прямым действием нейромедиатора является активация рецептора. Следовательно, эффекты нейромедиаторной системы зависят от связей нейронов, использующих медиатор, и химических свойств рецепторов.
- Глутамат используется в подавляющем большинстве быстрых возбуждающих синапсов в головном и спинном мозге. Он также используется в большинстве синапсов, которые являются «модифицируемыми», то есть способными увеличивать или уменьшать силу. Модифицируемые синапсы считаются основными элементами хранения памяти в мозге. Чрезмерное высвобождение глютамата может чрезмерно стимулировать мозг и привести к эксайтотоксичности, вызывающей гибель клеток, что приводит к судорогам или инсультам. [20] Эксайтотоксичность связана с некоторыми хроническими заболеваниями, включая ишемический инсульт , эпилепсию , боковой амиотрофический склероз , болезнь Альцгеймера , болезнь Хантингтона и болезнь Паркинсона . [21]
- ГАМК используется в подавляющем большинстве быстрых тормозных синапсов практически во всех частях мозга. Многие седативные/транквилизирующие препараты усиливают действие ГАМК. [22] Соответственно, глицин является тормозным медиатором в спинном мозге .
- Ацетилхолин был первым нейромедиатором, обнаруженным в периферической и центральной нервной системе. Он активирует скелетные мышцы соматической нервной системы и может как возбуждать, так и тормозить внутренние органы вегетативной системы. [14] Его называют передатчиком в нервно-мышечном соединении, соединяющем двигательные нервы с мышцами. Паралитический кураре, содержащий яд стрел, действует, блокируя передачу в этих синапсах. Ацетилхолин также действует во многих областях мозга, но использует разные типы рецепторов , включая никотиновые и мускариновые рецепторы. [23]
- Дофамин выполняет ряд важных функций в мозге; сюда входит регуляция двигательного поведения, удовольствия, связанные с мотивацией, а также эмоциональное возбуждение. Он играет решающую роль в системе вознаграждения ; Болезнь Паркинсона связана с низким уровнем дофамина, а шизофрения связана с высоким уровнем дофамина. [24]
- Серотонин — моноаминовый нейромедиатор . Большая часть вырабатывается и обнаруживается в кишечнике (приблизительно 90%), а остальная часть — в нейронах центральной нервной системы . Он регулирует аппетит, сон, память и обучение, температуру, настроение, поведение, сокращение мышц, а также функцию сердечно-сосудистой и эндокринной систем . Предполагается, что он играет роль в депрессии, поскольку у некоторых пациентов с депрессией наблюдается более низкая концентрация метаболитов серотонина в спинномозговой жидкости и тканях головного мозга. [25]
- Норадреналин является членом катехоламиновой классификации нейротрансмиттеров. Он синтезируется из аминокислоты тирозина . В периферической нервной системе одна из основных ролей норадреналина заключается в стимуляции высвобождения гормона стресса адреналина (т.е. адреналина ) из надпочечников . [26]
- Адреналин , нейромедиатор и гормон, синтезируется из тирозина . Он высвобождается надпочечниками и играет роль в реакции «бей или беги» . Адреналин оказывает сосудосуживающее действие, что способствует увеличению частоты сердечных сокращений, артериального давления, мобилизации энергии. Вазоконстрикция влияет на метаболизм , способствуя расщеплению глюкозы , попадающей в кровоток. Адреналин также оказывает бронхолитическое действие, то есть расслабляет дыхательные пути. [26]
Типы
[ редактировать ]Существует много разных способов классификации нейротрансмиттеров. Разделения их на аминокислоты , пептиды и моноамины достаточно для некоторых целей классификации. [27]
Основные нейромедиаторы:
- Аминокислоты : глутамат , [28] аспартат , D-серин , гамма-аминомасляная кислота (ГАМК), [номер 1] глицин
- Газотрансмиттеры : оксид азота (NO), оксид углерода (CO), сероводород (H 2 S).
- Моноамины :
- Катехоламины : дофамин (ДА), норадреналин (норадреналин, НЭ), адреналин (адреналин).
- Индоламины : серотонин (5-HT, SER), мелатонин.
- гистамин
- Следовые амины : фенэтиламин , N- метилфенэтиламин , тирамин , 3-йодтиронамин , октопамин , триптамин и др.
- Пептиды : окситоцин , соматостатин , вещество Р , транскрипт, регулируемый кокаином и амфетамином , опиоидные пептиды. [29]
- Пурины : аденозинтрифосфат (АТФ), аденозин.
- Прочие: ацетилхолин (АХ), анандамид и др.
Кроме того, более 100 нейроактивных пептидов , и регулярно открываются новые. обнаружено [30] [31] Многие из них выпускаются совместно с низкомолекулярными передатчиками. Тем не менее, в некоторых случаях пептид является основным передатчиком в синапсе. Бета-эндорфин является относительно известным примером пептидного нейротрансмиттера, поскольку он участвует в весьма специфических взаимодействиях с опиоидными рецепторами в центральной нервной системе .
Одиночные ионы (такие как синаптически высвобождаемый цинк ) некоторые также считают нейротрансмиттерами. [32] а также некоторые газообразные молекулы, такие как оксид азота (NO), окись углерода (CO) и сероводород (H 2 S). [33] Газы производятся в цитоплазме нервов и немедленно диффундируют через клеточную мембрану во внеклеточную жидкость и в близлежащие клетки, стимулируя выработку вторичных мессенджеров. Растворимые газовые нейротрансмиттеры трудно изучать, так как они действуют быстро и мгновенно разрушаются, существуя всего несколько секунд.
Наиболее распространенным передатчиком является глутамат , который возбуждает более 90% синапсов человеческого мозга. [28] Следующей по распространенности является гамма-аминомасляная кислота, или ГАМК, которая оказывает ингибирующее действие более чем на 90% синапсов, не использующих глутамат. Хотя другие трансмиттеры используются в меньшем количестве синапсов, они могут быть очень важны функционально: подавляющее большинство психоактивных препаратов оказывают свое действие, изменяя действия некоторых нейромедиаторных систем, часто действующих через трансмиттеры, отличные от глутамата или ГАМК. Наркотики, вызывающие привыкание, такие как кокаин и амфетамины, оказывают свое воздействие в первую очередь на дофаминовую систему. препараты, вызывающие привыкание, Опиатные оказывают свое действие в первую очередь как функциональные аналоги опиоидных пептидов , которые, в свою очередь, регулируют уровень дофамина.
Список нейротрансмиттеров, пептидов и газообразных сигнальных молекул
[ редактировать ]Нейромедиаторные системы
[ редактировать ]Нейроны, экспрессирующие определенные типы нейротрансмиттеров, иногда образуют отдельные системы, где активация системы влияет на большие объемы мозга, что называется объемной передачей . Основные системы нейромедиаторов включают , среди прочего, систему норадреналина (норадреналина), систему дофамина , систему серотонина и холинергическую систему. Следовые амины оказывают модулирующее действие на нейротрансмиссию в моноаминовых путях (т.е. дофаминовых, норадреналиновых и серотониновых путях) по всему мозгу посредством передачи сигналов через рецепторы, ассоциированные с следовыми аминами 1 . [37] [38] Ниже приводится краткое сравнение этих систем:
| Система | Происхождение пути и прогнозы | Регулируемые когнитивные процессы и поведение |
|---|---|---|
| Норадреналиновая система [39] [40] [41] [42] [43] [44] | Норадренергические пути :
|
|
| Дофаминовая система [41] [42] [43] [45] [46] [47] | Дофаминергические пути :
|
|
| Гистаминовая система [42] [43] [48] | Гистаминергические пути :
|
|
| Серотониновая система [39] [41] [42] [43] [49] [50] [51] | Серотонинергические пути : Каудальные ядра (ХН):
Ростральные ядра (РН):
|
|
| Ацетилхолиновая система [39] [41] [42] [43] [52] | Холинергические пути : Холинергические ядра переднего мозга (FCN):
Стриарные тонически активные холинергические нейроны (TAN) Холинергические ядра ствола мозга (BCN):
|
|
| Адреналиновая система [53] [54] | Адренергические пути :
|
Эффекты лекарств
[ редактировать ]Понимание влияния лекарств на нейротрансмиттеры составляет значительную часть исследовательских инициатив в области нейробиологии . Большинство нейробиологов, занимающихся этой областью исследований, считают, что такие усилия могут еще больше продвинуть наше понимание цепей, ответственных за различные неврологические заболевания и расстройства, а также способов эффективного лечения и, возможно, когда-нибудь предотвращения или лечения таких заболеваний. [55] [ нужна медицинская ссылка ]
Наркотики могут влиять на поведение, изменяя активность нейромедиаторов. Например, лекарства могут снизить скорость синтеза нейротрансмиттеров, воздействуя на синтетический фермент(ы) этого нейромедиатора. Когда синтез нейромедиаторов блокируется, количество нейромедиаторов, доступных для высвобождения, становится существенно меньшим, что приводит к снижению активности нейромедиаторов. Некоторые лекарства блокируют или стимулируют высвобождение определенных нейротрансмиттеров. Альтернативно, лекарства могут предотвратить накопление нейромедиаторов в синаптических везикулах, вызывая утечку мембран синаптических везикул. Препараты, которые препятствуют связыванию нейромедиатора с его рецептором, называются антагонистами рецепторов . Например, препараты, используемые для лечения пациентов с шизофренией, такие как галоперидол, хлорпромазин и клозапин, являются антагонистами рецепторов дофамина в головном мозге. Другие препараты действуют путем связывания с рецептором и имитируют нормальный нейромедиатор. Такие препараты называются агонистами рецепторов . Примером агониста рецептора является морфин , опиат, который имитирует действие эндогенного нейромедиатора β-эндорфина и облегчает боль. Другие препараты препятствуют дезактивации нейромедиатора после его высвобождения, тем самым продлевая действие нейромедиатора. Этого можно достичь путем блокирования обратного захвата или ингибирования деградационных ферментов. Наконец, лекарства также могут предотвращать возникновение потенциала действия, блокируя активность нейронов в центральной и периферической нервной системе . Такие препараты, как тетродотоксин , которые блокируют нервную активность, обычно смертельны.
Лекарства, нацеленные на нейротрансмиттеры основных систем, влияют на всю систему, что может объяснить сложность действия некоторых лекарств. Кокаин , например, блокирует обратный захват дофамина обратно в пресинаптический нейрон, оставляя молекулы нейромедиатора в синаптической щели на длительный период времени. Поскольку дофамин дольше остается в синапсе, нейромедиатор продолжает связываться с рецепторами постсинаптического нейрона , вызывая приятную эмоциональную реакцию. Физическая зависимость от кокаина может возникнуть в результате длительного воздействия избытка дофамина в синапсах, что приводит к подавлению некоторых постсинаптических рецепторов. После того, как действие препарата проходит, у человека может возникнуть депрессия из-за снижения вероятности связывания нейромедиатора с рецептором. Флуоксетин представляет собой селективный ингибитор обратного захвата серотонина (СИОЗС), который блокирует обратный захват серотонина пресинаптической клеткой, что увеличивает количество серотонина, присутствующего в синапсе, и, кроме того, позволяет ему оставаться там дольше, обеспечивая потенциал для эффекта естественного высвободился серотонин. [56] AMPT предотвращает превращение тирозина в L-ДОФА , предшественник дофамина; резерпин предотвращает накопление дофамина в везикулах ; а депренил ингибирует моноаминоксидазу (МАО)-B и, таким образом, повышает уровень дофамина.
| Лекарство | Взаимодействует с: | Рецепторное взаимодействие: | Тип | Эффекты |
|---|---|---|---|---|
| Ботулотоксин (ботокс) | Ацетилхолин | – | Антагонист | Блокирует высвобождение ацетилхолина в ПНС Предотвращает мышечные сокращения |
| Яд паука Черной Вдовы | Ацетилхолин | – | Агонист | Способствует высвобождению ацетилхолина в ПНС. Стимулирует мышечные сокращения |
| Неостигмин | Ацетилхолин | – | – | Влияет на активность ацетилхолинеразы Усиливает действие АХ на рецепторы Используется для лечения миастении. |
| Никотин | Ацетилхолин | Никотиновый (скелетные мышцы) | Агонист | Повышает активность АХ Повышает внимание Усиливающие эффекты |
| d-тубокурарин | Ацетилхолин | Никотиновый (скелетные мышцы) | Антагонист | Снижает активность рецепторного участка |
| Излечивать | Ацетилхолин | Никотиновый (скелетные мышцы) | Антагонист | Снижает активность АХ Предотвращает мышечные сокращения |
| Мускарин | Ацетилхолин | Мускариновые (сердце и гладкие мышцы) | Агонист | Повышает активность АХ Токсичный |
| Атропин | Ацетилхолин | Мускариновые (сердце и гладкие мышцы) | Антагонист | Блокирует сужение зрачков Блокирует выработку слюны |
| Скополамин ( Гиосцин ) | Ацетилхолин | Мускариновые (сердце и гладкие мышцы) | Антагонист | Лечит укачивание, послеоперационную тошноту и рвоту. |
| АМРТ | Дофамин/норадреналин | – | – | Инактивирует тирозингидроксилазу и подавляет выработку дофамина. |
| Резерпин | Дофамин | – | – | Предотвращает накопление дофамина и других моноаминов в синаптических везикулах. Вызывает седацию и депрессию |
| Апоморфин | Дофамин | Рецептор D2 (пресинаптические ауторецепторы/постсинаптические рецепторы) | Антагонист (низкая доза)/Прямой агонист (высокая доза) | Низкая доза: блокирует ауторецепторы. Высокая доза: стимулирует постсинаптические рецепторы. |
| Амфетамин | Дофамин/норадреналин | – | Непрямой агонист | Высвобождает дофамин, норадреналин и серотонин. |
| Метамфетамин | Дофамин/норадреналин | – | – | Высвобождает дофамин и норадреналин. Блокирует обратный захват |
| Метилфенидат | Дофамин | – | – | Блокирует обратный захват Улучшает внимание и контроль импульсивности при СДВГ. |
| Кокаин | Дофамин | – | Непрямой агонист | Блокирует обратный захват в пресинапс Блокирует потенциалзависимые натриевые каналы Может использоваться в качестве местного анестетика (глазные капли). |
| Депренил | Дофамин | – | Агонист | Ингибирует МАО-Б Предотвращает разрушение дофамина |
| хлорпромазин | Дофамин | D2-рецепторы | Антагонист | Блокирует D2-рецепторы Облегчает галлюцинации |
| МПТП | Дофамин | – | – | Приводит к появлению паркинсоноподобных симптомов |
| ПКПА | Серотонин (5-НТ) | – | Антагонист | Нарушает синтез серотонина, блокируя активность триптофангидроксилазы. |
| Ондансетрон | Серотонин (5-НТ) | 5-HT3 - рецепторы | Антагонист | Уменьшает побочные эффекты химиотерапии и радиации. Уменьшает тошноту и рвоту |
| Буспирон | Серотонин (5-НТ) | 5-НТ 1А рецепторы | Частичный агонист | Лечит симптомы тревоги и депрессии |
| Флуоксетин | Серотонин (5-НТ) | поддерживает 5-HT обратный захват | СИОЗС | Тормозит обратный захват серотонина Лечит депрессию, некоторые тревожные расстройства и ОКР. [56] Типичные примеры: Прозак и Сарафем. |
| Фенфлурамин | Серотонин (5-НТ) | – | – | Вызывает выброс серотонина Тормозит обратный захват серотонина Используется как средство для подавления аппетита. |
| Диэтиламид лизергиновой кислоты | Серотонин (5-НТ) | Постсинаптические 5-НТ 2А рецепторы | Прямой агонист | Вызывает искажения зрительного восприятия. Стимулирует рецепторы 5-НТ 2А в переднем мозге. |
| Метилендиоксиметамфетамин ( МДМА ) | Серотонин (5-НТ)/норадреналин | – | – | Стимулирует высвобождение серотонина и норадреналина и подавляет обратный захват. Вызывает возбуждающие и галлюциногенные эффекты. |
| Стрихнин | Глицин | – | Антагонист | Вызывает сильные мышечные спазмы [58] |
| Димедрол | Гистамин | Пересекает гематоэнцефалический барьер, вызывая сонливость | ||
| Тетрагидроканнабинол (ТГК) | Эндоканнабиноиды | Каннабиноидные (CB) рецепторы | Агонист | Оказывает обезболивающее и седативное действие Повышает аппетит Когнитивные эффекты |
| Римонабант | Эндоканнабиноиды | Каннабиноидные (CB) рецепторы | Антагонист | Подавляет аппетит Используется при отказе от курения |
| МАФП | Эндоканнабиноиды | – | – | Подавляет ФААХ Используется в исследованиях для повышения активности каннабиноидной системы. |
| АМ1172 | Эндоканнабиноиды | – | – | Блокирует обратный захват каннабиноидов Используется в исследованиях для повышения активности каннабиноидной системы. |
| Анандамид (эндогенный) | – | Каннабиноидные (CB) рецепторы; 5-HT3 - рецепторы | – | Уменьшить тошноту и рвоту |
| Кофеин | Аденозин | Аденозиновые рецепторы | Антагонист | Блокирует аденозиновые рецепторы Увеличивает бодрствование |
| PCP | Глутамат | NMDA-рецептор | Косвенный антагонист | Блокирует сайт связывания PCP Предотвращает попадание ионов кальция в нейроны. Ухудшает обучение |
| АП5 | Глутамат | NMDA-рецептор | Антагонист | Блокирует сайт связывания глутамата на рецепторе NMDA. Нарушает синаптическую пластичность и некоторые формы обучения. |
| Кетамин | Глутамат | NMDA-рецептор | Антагонист | Используется в качестве анестезии Вызывает трансоподобное состояние, помогает облегчить боль и успокоить. |
| НМДА | Глутамат | NMDA-рецептор | Агонист | Используется в исследованиях по изучению рецептора NMDA. Ионотропный рецептор |
| АМПА | Глутамат | АМРА-рецептор | Агонист | Используется в исследованиях по изучению рецептора AMPA. Ионотропный рецептор |
| Аллиглицин | ПЕРЕДНИЙ | – | – | Ингибирует синтез ГАМК Вызывает судороги |
| Мусцимол | ПЕРЕДНИЙ | ГАМК-рецептор | Агонист | Вызывает седативный эффект |
| бикукулин | ПЕРЕДНИЙ | ГАМК-рецептор | Антагонист | Вызывает судороги |
| Бензодиазепины | ПЕРЕДНИЙ | ГАМК А- рецептор | Непрямые агонисты | Анксиолитик, седативный эффект, ухудшение памяти, мышечная релаксация |
| Барбитураты | ПЕРЕДНИЙ | ГАМК А- рецептор | Непрямые агонисты | Седация, ухудшение памяти, мышечная релаксация |
| Алкоголь | ПЕРЕДНИЙ | ГАМК-рецептор | Непрямой агонист | Седация, ухудшение памяти, мышечная релаксация |
| Пикротоксин | ПЕРЕДНИЙ | ГАМК А- рецептор | Косвенный антагонист | Высокие дозы вызывают судороги |
| Тиагабин | ПЕРЕДНИЙ | – | Антагонист | антагонист транспортера ГАМК Увеличение доступности ГАМК Снижает вероятность судорог |
| Моклобемид | Норадреналин | – | Агонист | Блокирует МАО-А для лечения депрессии |
| Идазоксан | Норадреналин | альфа-2-адренергические ауторецепторы | Агонист | Блокирует ауторецепторы альфа-2. Используется для изучения норадреналиновой системы. |
| Фузаровая кислота | Норадреналин | – | – | Подавляет активность дофамин-бета-гидроксилазы, которая блокирует выработку норадреналина. Используется для изучения системы норадреналина без воздействия на дофаминовую систему. |
| Опиаты ( опиум , морфин , героин и оксикодон ) | Опиоиды | Опиоидный рецептор [59] | Агонисты | Анальгезия, седативный эффект и усиливающий эффект. |
| Налоксон | Опиоиды | – | Антагонист | Устраняет симптомы опиатной интоксикации или передозировки (т. е. проблемы с дыханием) |
Агонисты
[ редактировать ]Этот раздел необходимо расширить , включив в него описание полных агонистов и их отличие от частичных и обратных агонистов. Вы можете помочь, добавив в него . ( август 2015 г. ) |
Агонист представляет собой химическое вещество, способное связываться с рецептором, таким как рецептор нейромедиатора, и инициировать ту же реакцию, которая обычно возникает при связывании эндогенного вещества. [60] Таким образом, агонист нейротрансмиттера инициирует тот же ответ рецептора, что и передатчик. В нейронах агонист может активировать рецепторы нейромедиаторов прямо или косвенно. Агонисты прямого связывания могут быть дополнительно охарактеризованы как полные агонисты , частичные агонисты и обратные агонисты . [61] [62]
Прямые агонисты действуют аналогично нейротрансмиттеру, связываясь непосредственно с связанными с ним рецепторными участками, которые могут располагаться на пресинаптическом нейроне или постсинаптическом нейроне, или на обоих. [63] Обычно рецепторы нейромедиаторов расположены на постсинаптическом нейроне, тогда как ауторецепторы нейромедиаторов расположены на пресинаптическом нейроне, как в случае с моноаминовыми нейромедиаторами ; [37] в некоторых случаях нейромедиатор использует ретроградную нейротрансмиссию — тип передачи сигналов обратной связи в нейронах, при котором нейромедиатор высвобождается постсинаптически и связывается с целевыми рецепторами, расположенными на пресинаптическом нейроне. [64] [примечание 1] Никотин , соединение, содержащееся в табаке , является прямым агонистом большинства никотиновых рецепторов ацетилхолина , главным образом расположенных в холинергических нейронах . [59] Опиаты , такие как морфин , героин , гидрокодон , оксикодон , кодеин и метадон , являются μ-опиоидных рецепторов агонистами ; это действие обеспечивает их эйфорические и обезболивающие свойства. [59]
Непрямые агонисты усиливают связывание нейротрансмиттеров с их рецепторами-мишенями, стимулируя высвобождение или предотвращая обратный захват нейротрансмиттеров. [63] Некоторые непрямые агонисты вызывают высвобождение нейромедиаторов и предотвращают обратный захват нейромедиаторов . Амфетамин , например, является непрямым агонистом постсинаптических рецепторов дофамина, норадреналина и серотонина в каждом соответствующем нейроне; [37] [38] он вызывает высвобождение нейротрансмиттеров в пресинаптический нейрон, а затем в синаптическую щель, и предотвращает их обратный захват из синаптической щели путем активации TAAR1 , пресинаптического рецептора, связанного с G-белком , и связывания с участком VMAT2 , типа переносчика моноаминов, расположенного в синапсе. везикулы внутри моноаминовых нейронов . [37] [38]
Антагонисты
[ редактировать ]Антагонист — это химическое вещество, которое действует внутри организма, снижая физиологическую активность другого химического вещества (например, опиата); особенно тот, который противодействует действию на нервную систему лекарства или вещества, естественным образом присутствующего в организме, путем объединения и блокирования его нервных рецепторов. [65]
Существует два основных типа антагонистов: антагонисты прямого действия и антагонисты непрямого действия:
- Антагонист прямого действия, который занимает место на рецепторах, которое в противном случае занято самими нейротрансмиттерами. Это приводит к тому, что нейротрансмиттеры блокируются от связывания с рецепторами. Примером одного из наиболее распространенных является Атропин.
- Антагонисты непрямого действия – препараты, подавляющие высвобождение/выработку нейротрансмиттеров (например, резерпин ).
Антагонисты наркотиков
[ редактировать ]Препарат-антагонист — это препарат, который прикрепляется (или связывается) к участку, называемому рецептором, без активации этого рецептора для возникновения биологического ответа. Поэтому говорят, что он не обладает внутренней активностью. Антагонист также можно назвать «блокатором» рецептора, поскольку он блокирует действие агониста на этот участок. Таким образом, фармакологические эффекты антагониста приводят к предотвращению связывания и активации соответствующего участка рецептора (например, лекарств, гормонов, нейротрансмиттеров). Антагонисты могут быть «конкурентными» или «необратимыми».
Конкурентный антагонист конкурирует с агонистом за связывание с рецептором. По мере увеличения концентрации антагониста связывание агониста постепенно ингибируется, что приводит к снижению физиологического ответа. Высокая концентрация антагониста может полностью подавить ответ. Однако это ингибирование можно обратить вспять путем увеличения концентрации агониста, поскольку агонист и антагонист конкурируют за связывание с рецептором. Таким образом, конкурентные антагонисты могут быть охарактеризованы как смещающие соотношение «доза-эффект» вправо для агониста. В присутствии конкурентного антагониста требуется повышенная концентрация агониста, чтобы вызвать тот же ответ, который наблюдается в отсутствие антагониста.
Необратимый антагонист настолько сильно связывается с рецептором, что делает рецептор недоступным для связывания с агонистом. Необратимые антагонисты могут даже образовывать ковалентные химические связи с рецептором. В любом случае, если концентрация необратимого антагониста достаточно высока, количество несвязанных рецепторов, остающихся для связывания агониста, может быть настолько низким, что даже высокие концентрации агониста не вызывают максимального биологического ответа. [66]
Прекурсоры
[ редактировать ]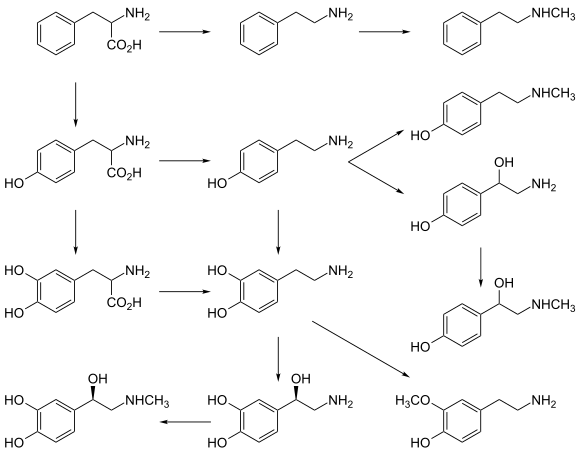
Хотя прием предшественников и активация постсинаптических рецепторов , неоднозначны ли высвобождение нейротрансмиттеров нейромедиаторов действительно увеличивает синтез нейромедиаторов, данные относительно того, увеличивается . Даже при повышенном высвобождении нейромедиаторов неясно, приведет ли это к долгосрочному увеличению силы сигнала нейромедиаторов, поскольку нервная система может адаптироваться к таким изменениям, как увеличение синтеза нейромедиаторов, и, следовательно, может поддерживать постоянную активность. [70] [ ненадежный медицинский источник? ] Некоторые нейротрансмиттеры могут играть роль в развитии депрессии, и есть некоторые данные, позволяющие предположить, что прием предшественников этих нейротрансмиттеров может быть полезен при лечении легкой и умеренной депрессии. [70] [ ненадежный медицинский источник? ] [71]
Катехоламины и предшественники следовых аминов
[ редактировать ]L -ДОФА , предшественник дофамина , проникающий через гематоэнцефалический барьер , используется при лечении болезни Паркинсона . Для пациентов с депрессией, у которых наблюдается низкая активность нейромедиатора норадреналина , имеется лишь мало доказательств пользы введения предшественников нейротрансмиттера. L-фенилаланин и L-тирозин являются предшественниками дофамина , норадреналина и адреналина . Для этих преобразований необходимы витамин B6 , витамин C и S-аденозилметионин . Несколько исследований предполагают потенциальные антидепрессивные эффекты L-фенилаланина и L-тирозина, но в этой области есть много возможностей для дальнейших исследований. [70] [ ненадежный медицинский источник? ]
Предшественники серотонина
[ редактировать ]Установлено, что введение L-триптофана , предшественника серотонина , удваивает выработку серотонина в мозге. Он значительно эффективнее плацебо при лечении легкой и умеренной депрессии. [70] [ ненадежный медицинский источник? ] Для этого преобразования требуется витамин С. [25] 5-гидрокситриптофан (5-HTP), также предшественник серотонина , более эффективен, чем плацебо. [70] [ ненадежный медицинский источник? ]
Болезни и расстройства
[ редактировать ]Заболевания и расстройства могут также влиять на определенные системы нейромедиаторов. Ниже приведены расстройства, связанные с увеличением, уменьшением или дисбалансом определенных нейротрансмиттеров.
Дофамин :
Например, проблемы с выработкой дофамина (в основном в черной субстанции ) могут привести к болезни Паркинсона — расстройству, которое влияет на способность человека двигаться так, как он хочет, что приводит к скованности, тремору или тряске и другим симптомам. Некоторые исследования показывают, что слишком мало или слишком много дофамина или проблемы с использованием дофамина в областях мышления и чувств мозга могут играть роль в таких расстройствах, как шизофрения или синдром дефицита внимания с гиперактивностью (СДВГ). Дофамин также участвует в зависимости и употреблении наркотиков, поскольку большинство рекреационных наркотиков вызывают приток дофамина в мозг (особенно опиоиды и метамфетамины ), что вызывает ощущение удовольствия, поэтому потребители постоянно жаждут наркотиков.
Серотонин :
Аналогичным образом, после того, как некоторые исследования показали, что препараты, блокирующие переработку или обратный захват серотонина, помогают некоторым людям с диагнозом депрессия, было высказано предположение, что у людей с депрессией уровень серотонина может быть ниже нормального. Несмотря на широкую популяризацию, эта теория не нашла подтверждения в последующих исследованиях. [72] Поэтому селективные ингибиторы обратного захвата серотонина (СИОЗС) используются для увеличения количества серотонина в синапсах.
Глутамат :
Кроме того, проблемы с выработкой или использованием глутамата предположительно связаны со многими психическими расстройствами, включая аутизм , обсессивно-компульсивное расстройство (ОКР), шизофрению и депрессию . [73] Избыток глютамата связан с неврологическими заболеваниями, такими как болезнь Паркинсона , рассеянный склероз , болезнь Альцгеймера , инсульт и БАС (боковой амиотрофический склероз). [74]
Дисбаланс нейромедиаторов
[ редактировать ]Как правило, не существует научно установленных «норм» для соответствующих уровней или «баланса» различных нейротрансмиттеров. В большинстве случаев практически невозможно даже измерить уровни нейротрансмиттеров в мозге или теле в какие-либо отдельные моменты времени. Нейротрансмиттеры регулируют высвобождение друг друга, и слабый постоянный дисбаланс в этой взаимной регуляции был связан с темпераментом у здоровых людей. [75] [76] [77] [78] [79] Сильный дисбаланс или нарушения нейромедиаторных систем связаны со многими заболеваниями и психическими расстройствами. К ним относятся болезнь Паркинсона, депрессия, бессонница, синдром дефицита внимания и гиперактивности (СДВГ), тревога, потеря памяти, резкие изменения веса и зависимости. Хронический физический или эмоциональный стресс может способствовать изменениям нейромедиаторной системы. Генетика также играет роль в активности нейромедиаторов. Помимо использования в рекреационных целях, лекарства, которые прямо или косвенно взаимодействуют с одним или несколькими передатчиками или его рецепторами, обычно назначаются при психиатрических и психологических проблемах. Примечательно, что лекарства, взаимодействующие с серотонином и норадреналином, назначают пациентам с такими проблемами, как депрессия и тревога, хотя представление о том, что существует много убедительных медицинских доказательств в поддержку таких вмешательств, подвергается широкой критике. [80] Исследования показали, что дисбаланс дофамина влияет на рассеянный склероз и другие неврологические расстройства. [81]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ В центральной нервной системе анандамид и другие эндоканнабиноиды используют ретроградную нейротрансмиссию, поскольку их высвобождение является постсинаптическим, а их целевой рецептор, каннабиноидный рецептор 1 (CB1), является пресинаптическим. [64] Растение каннабис содержит Δ 9 -тетрагидроканнабинол , который является прямым агонистом CB1. [64]
- ^ ГАМК — непротеиногенная аминокислота.
Ссылки
[ редактировать ]- ^ Смелзер, Нил Дж.; Балтес, Пол Б. (2001). Международная энциклопедия социальных и поведенческих наук (1-е изд.). Амстердам, Нью-Йорк: Эльзевир. ISBN 978-0-08-043076-8 .
- ^ Куэвас Дж. (1 января 2019 г.). «Нейромедиаторы и их жизненный цикл». Справочный модуль по биомедицинским наукам . Эльзевир. дои : 10.1016/b978-0-12-801238-3.11318-2 . ISBN 978-0-12-801238-3 .
- ^ Первс Д., Августин Г.Дж., Фитцпатрик Д., Кац Л.К., ЛаМантия А.С., Макнамара Д.О., Уильямс С.М. (2001). «Пептидные нейротрансмиттеры» . Нейронаука (2-е изд.). Синауэр Ассошиэйтс.
- ^ Сандерс К.М., Уорд С.М. (январь 2019 г.). «Оксид азота и его роль как неадренергического, нехолинергического тормозного нейромедиатора в желудочно-кишечном тракте» . Британский журнал фармакологии . 176 (2): 212–227. дои : 10.1111/bph.14459 . ПМК 6295421 . ПМИД 30063800 .
- ^ Элиас Л.Дж., Сосье Д.М. (2005). Нейропсихология: клинические и экспериментальные основы . Бостон: Пирсон.
- ^ Шерги К., Суо-Шаньи М.Ф., Гонон Ф. (октябрь 1994 г.). «Нелинейная связь между потоком импульсов, высвобождением дофамина и выведением дофамина в мозге крысы in vivo». Нейронаука . 62 (3): 641–645. дои : 10.1016/0306-4522(94)90465-0 . ПМИД 7870295 . S2CID 20465561 .
- ^ Мустафа, Асиф К.; Ким, Пол М.; Снайдер, Соломон Х. (август 2004 г.). «D-серин как предполагаемый глиальный нейромедиатор» . Биология нейронов глии . 1 (3): 275–281. дои : 10.1017/S1740925X05000141 . ISSN 1741-0533 . ПМК 1403160 . ПМИД 16543946 .
- ^ Волоскер, Герман; Думина, Елена; Балан, Ливия; Фолтын, Вероника Н. (июль 2008 г.). «D-Аминокислоты в мозге: d-серин в нейротрансмиссии и нейродегенерации: d-серин в нейротрансмиссии и нейродегенерации» . Журнал ФЭБС . 275 (14): 3514–3526. дои : 10.1111/j.1742-4658.2008.06515.x . ПМИД 18564180 . S2CID 25735605 .
- ^ Тапа С., Лев М., Сюй Х (30 ноября 2017 г.). «Ацетилхолинэстераза: основная мишень для лекарств и инсектицидов». Мини-обзоры по медицинской химии . 17 (17): 1665–1676. дои : 10.2174/1389557517666170120153930 . ПМИД 28117022 .
- ^ Васика Дж., Теннант CC (сентябрь 2002 г.). «Употребление кокаина и сердечно-сосудистые осложнения». Медицинский журнал Австралии . 177 (5): 260–262. дои : 10.5694/j.1326-5377.2002.tb04761.x . ПМИД 12197823 . S2CID 18572638 .
- ^ Саладин, Кеннет С. Анатомия и физиология: единство формы и функции . МакГроу Хилл. 2009 год ISBN 0-07-727620-5
- ^ Телеану, Ралука Иоана; Никулеску, Аделина-Габриэла; Роза, Евгения; Владаченко, Оана; Грумешеску, Александру Михай; Телеану, Даниэль Михай (25 мая 2022 г.). «Нейромедиаторы — ключевые факторы неврологических и нейродегенеративных заболеваний центральной нервной системы» . Международный журнал молекулярных наук . 23 (11): 5954. doi : 10.3390/ijms23115954 . ISSN 1422-0067 . ПМК 9180936 . ПМИД 35682631 .
- ^ Бридлав С.М. , Уотсон Н.В. (2013). Биологическая психология: введение в поведенческую, когнитивную и клиническую нейробиологию (Седьмое изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0878939275 .
- ^ Jump up to: а б Уишоу Б., Kolb IQ (2014). Введение в мозг и поведение (4-е изд.). Нью-Йорк, штат Нью-Йорк: Worth Publishers. стр. 150–151. ISBN 978-1429242288 .
- ^ Первс, Дейл; Августин, Джордж Дж.; Фитцпатрик, Дэвид; Кац, Лоуренс К.; ЛаМантия, Энтони-Самуэль; Макнамара, Джеймс О.; Уильямс, С. Марк (2001), «Возбуждающие и тормозящие постсинаптические потенциалы» , Неврология. 2-е издание , Sinauer Associates , получено 14 июля 2023 г.
- ^ Питерс А., Палай С.Л. (декабрь 1996 г.). «Морфология синапсов». Журнал нейроцитологии . 25 (12): 687–700. дои : 10.1007/BF02284835 . ПМИД 9023718 . S2CID 29365393 .
- ^ Шир Д., Батлер Дж., Льюис Р. (5 января 2015 г.). Анатомия и физиология человека Хоула (Четырнадцатое изд.). Нью-Йорк, штат Нью-Йорк. ISBN 978-0-07-802429-0 . OCLC 881146319 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Ди Кьяра Дж., Морелли М., Консоло С. (июнь 1994 г.). «Модуляторные функции нейротрансмиттеров в полосатом теле: взаимодействия АХ/дофамин/NMDA». Тенденции в нейронауках . 17 (6): 228–233. дои : 10.1016/0166-2236(94)90005-1 . ПМИД 7521083 . S2CID 32085555 .
- ^ Уишоу Б., Kolb IQ (2014). Введение в мозг и поведение (4-е изд.). Нью-Йорк, штат Нью-Йорк: Worth Publishers. ISBN 978-1429242288 .
- ^ Гросс Л (ноябрь 2006 г.). « Поддерживающие» игроки играют ведущую роль в защите чрезмерно возбужденного мозга» . ПЛОС Биология . 4 (11): е371. дои : 10.1371/journal.pbio.0040371 . ПМК 1609133 . ПМИД 20076484 .
- ^ Ян Дж.Л., Сикора П., Уилсон Д.М., Мэттсон член парламента, Бор В.А. (август 2011 г.). «Возбуждающий нейротрансмиттер глутамат стимулирует восстановление ДНК, повышая устойчивость нейронов» . Механизмы старения и развития . 132 (8–9): 405–11. дои : 10.1016/j.mad.2011.06.005 . ПМЦ 3367503 . ПМИД 21729715 .
- ^ Антагонисты рецепторов орексина — новый класс снотворных , Национальный фонд сна.
- ^ «Ацетилхолиновые рецепторы» . Ebi.ac.uk. Проверено 25 августа 2014 г.
- ^ Шактер, Гилберт и Вегер. Психология.Соединенные Штаты Америки.2009.Печать.
- ^ Jump up to: а б Бристольский университет. «Введение в серотонин» . Проверено 15 октября 2009 г.
- ^ Jump up to: а б Шеффлер, Закари М.; Редди, Вамси; Пилларисетти, Лила Шарат (2023), «Физиология, нейротрансмиттеры» , StatPearls , Остров сокровищ (Флорида): StatPearls Publishing, PMID 30969716 , получено 16 июля 2023 г.
- ^ Прасад Б.В. (2020). Изучение биологических основ человеческого поведения . Соединенные Штаты Америки: IGI Global. п. 81. ИСБН 978-1799-8286-17 .
- ^ Jump up to: а б Сапольский Р. (2005). «Биология и поведение человека: неврологические истоки индивидуальности, 2-е». Учебная компания.
см. страницы 13 и 14 путеводителя.
- ^ Снайдер С.Х., Иннис РБ (1979). «Пептидные нейротрансмиттеры». Ежегодный обзор биохимии . 48 : 755–82. дои : 10.1146/annurev.bi.48.070179.003543 . ПМИД 38738 .
- ^ Корбьер А., Водри Х., Чан П., Вале-Балье М.Л., Лекрок Т., Лефевр А. и др. (18 сентября 2019 г.). «Стратегии идентификации биоактивных нейропептидов у позвоночных» . Границы в неврологии . 13 : 948. дои : 10.3389/fnins.2019.00948 . ПМК 6759750 . ПМИД 31619945 .
- ^ Фрикер Л.Д., Деви Л.А. (май 2018 г.). «Орфанные нейропептиды и рецепторы: новые терапевтические мишени» . Фармакология и терапия . 185 : 26–33. doi : 10.1016/j.pharmthera.2017.11.006 . ПМК 5899030 . ПМИД 29174650 .
- ^ Кодиров С.А., Такидзава С., Джозеф Дж., Кандель Э.Р., Шумяцкий Г.П., Большаков В.Ю. (октябрь 2006 г.). «Синаптически высвобождаемый цинк обеспечивает долговременное усиление путей формирования страха» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (41): 15218–23. Бибкод : 2006PNAS..10315218K . дои : 10.1073/pnas.0607131103 . ПМЦ 1622803 . ПМИД 17005717 .
- ^ «Международный симпозиум по оксиду азота – доктор Джон Эндрюс – MaRS» . МаРС . Архивировано из оригинала 14 октября 2014 года.
- ^ «Дофамин: Биологическая активность» . Руководство IUPHAR/BPS по фармакологии . Международный союз фундаментальной и клинической фармакологии . Проверено 29 января 2016 г.
- ^ Гранди Д.К., Миллер ГМ, Ли Дж.С. (февраль 2016 г.). « TAAR Получение зависимости» - Аламо является свидетелем еще одной революции: обзор пленарного симпозиума Конференции по поведению, биологии и химии 2015 года» . Наркотическая и алкогольная зависимость . 159 : 9–16. doi : 10.1016/j.drugalcdep.2015.11.014 . ПМЦ 4724540 . ПМИД 26644139 .
TAAR1 представляет собой рецептор с высоким сродством к METH/AMPH и DA.
- ^ Лин Ю, Холл Р.А., Кухар М.Дж. (октябрь 2011 г.). «Стимуляция CART-пептидом передачи сигналов, опосредованной G-белком, в дифференцированных клетках PC12: идентификация PACAP 6-38 как антагониста рецептора CART» . Нейропептиды . 45 (5): 351–8. дои : 10.1016/j.npep.2011.07.006 . ПМК 3170513 . ПМИД 21855138 .
- ^ Jump up to: а б с д и Миллер GM (январь 2011 г.). «Новая роль рецептора 1, связанного с следами аминов, в функциональной регуляции переносчиков моноаминов и дофаминергической активности» . Журнал нейрохимии . 116 (2): 164–76. дои : 10.1111/j.1471-4159.2010.07109.x . ПМК 3005101 . ПМИД 21073468 .
- ^ Jump up to: а б с д Эйден Л.Е., Вэйхэ Э. (январь 2011 г.). «VMAT2: динамический регулятор функции моноаминергических нейронов мозга, взаимодействующий с наркотиками» . Анналы Нью-Йоркской академии наук . 1216 (1): 86–98. Бибкод : 2011NYASA1216...86E . дои : 10.1111/j.1749-6632.2010.05906.x . ПМЦ 4183197 . ПМИД 21272013 .
VMAT2 является везикулярным транспортером ЦНС не только для биогенных аминов DA, NE, EPI, 5-HT и HIS, но, вероятно, также для следовых аминов TYR, PEA и тиронамина (THYR). ЦНС млекопитающих можно идентифицировать как нейроны, экспрессирующие VMAT2 для хранения и биосинтетический фермент декарбоксилазу ароматических аминокислот (AADC).
- ^ Jump up to: а б с Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 6: Широко распространенные системы: моноамины, ацетилхолин и орексин». В Сидоре А., Брауне Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. п. 155. ИСБН 9780071481274 .
Различные субрегионы ВТА получают глутаматергические входы от префронтальной коры, орексинергические входы от латерального гипоталамуса, холинергические, а также глутаматергические и ГАМКергические входы от латеродорсального тегментального ядра и ядра ножки, норадренергические входы от голубого пятна, серотонинергические входы от ядер шва, и ГАМКергические входы от прилежащего ядра и вентрального паллидума.
- ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 6: Широко распространенные системы: моноамины, ацетилхолин и орексин». В Сидоре А., Брауне Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 145, 156–157. ISBN 9780071481274 .
Нисходящие волокна NE модулируют афферентные болевые сигналы. ... Голубое пятно (LC), расположенное на дне четвертого желудочка в ростральном мосту, содержит более 50% всех норадренергических нейронов головного мозга; он иннервирует как передний мозг (т. е. обеспечивает практически всю НЭ кору головного мозга), так и области ствола головного мозга и спинного мозга. ... Остальные норадренергические нейроны головного мозга встречаются в рыхлых скоплениях клеток в стволе мозга, включая латеральные покрышки. Эти нейроны проецируются в основном в стволе головного мозга и спинном мозге. НЭ, наряду с 5НТ, АХ, гистамином и орексином, является критическим регулятором цикла сна-бодрствования и уровня возбуждения. ... Запуск LC может также усилить тревогу ... Стимуляция β-адренергических рецепторов в миндалевидном теле приводит к усилению памяти на стимулы, закодированные под сильными негативными эмоциями ... Адреналин возникает лишь в небольшом количестве центральных нейронов, все они расположены в мозговое вещество. Адреналин участвует во висцеральных функциях, таких как контроль дыхания.
- ^ Jump up to: а б с д Позвонил HP (2003). Фармакология . Эдинбург: Черчилль Ливингстон. стр. 474 для норадреналиновой системы, стр. 476 для дофаминовой системы, стр. 480 для серотониновой системы и стр. 483 для холинергической системы. ISBN 978-0-443-07145-4 .
- ^ Jump up to: а б с д и Иванчук В., Гузничак П. (2015). «Нейрофизиологические основы сна, пробуждения, осознания и феноменов сознания. Часть 1» . Анестезиология Интенсивная терапия . 47 (2): 162–7. дои : 10.5603/AIT.2015.0015 . ПМИД 25940332 .
Восходящая ретикулярная активирующая система (ARAS) отвечает за длительное состояние бодрствования. ... В таламической проекции преобладают холинергические нейроны, происходящие из педункулопонтинного тегментального ядра моста и среднего мозга (ППТ) и латеродорсального тегментального ядра моста и среднего мозга (ЛДТ) [17, 18]. В гипоталамическую проекцию вовлекаются норадренергические нейроны голубого пятна (LC) и серотонинергические нейроны дорсального и срединного ядер шва (DR), которые проходят через латеральный гипоталамус и достигают аксонов гистаминергического туберомамиллярного ядра (TMN), образуя вместе Путь, идущий в передний мозг, кору и гиппокамп. Корковое возбуждение также использует преимущества дофаминергических нейронов черной субстанции (SN), вентральной области покрышки (VTA) и периакведуктальной серой зоны (PAG). Меньшее количество холинергических нейронов моста и среднего мозга посылают проекции в передний мозг по вентральному пути, минуя таламус [19, 20].
- ^ Jump up to: а б с д и Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 12: Сон и возбуждение». В Сидоре А., Брауне Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк, США: McGraw-Hill Medical. п. 295. ИСБН 9780071481274 .
ARAS представляет собой сложную структуру, состоящую из нескольких различных цепей, включая четыре моноаминергических пути... Путь норадреналина берет свое начало из голубого пятна (LC) и связанных с ним ядер ствола мозга; серотонинергические нейроны также происходят из ядер шва в стволе мозга; дофаминергические нейроны берут начало в вентральной покрышке (ВТА); а гистаминэргический путь начинается от нейронов туберомаммиллярного ядра (ТЯН) заднего гипоталамуса. Как обсуждалось в главе 6, эти нейроны широко разрастаются по всему мозгу из ограниченного набора тел клеток. Норадреналин, серотонин, дофамин и гистамин выполняют сложные модулирующие функции и в целом способствуют бодрствованию. ПТ ствола мозга также является важным компонентом ARAS. Активность PT-холинергических нейронов (клеток быстрого сна) способствует быстрому сну. Во время бодрствования клетки, активные для быстрого сна, ингибируются подмножеством нейронов норадреналина и серотонина ARAS, называемых клетками, выключенными для быстрого сна.
- ^ Ринаман Л. (февраль 2011 г.). «Норадренергические нейроны А2 заднего мозга: разнообразные роли в вегетативных, эндокринных, когнитивных и поведенческих функциях» . Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 300 (2): 222–35 рандов. дои : 10.1152/ajpregu.00556.2010 . ПМК 3043801 . ПМИД 20962208 .
- ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 6: Широко распространенные системы: моноамины, ацетилхолин и орексин». В Сидоре А., Брауне Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 147–148, 154–157. ISBN 9780071481274 .
Нейроны SNc плотно иннервируют дорсальное полосатое тело, где они играют решающую роль в обучении и выполнении двигательных программ. Нейроны ВТА иннервируют вентральное полосатое тело (прилежащее ядро), обонятельную луковицу, миндалевидное тело, гиппокамп, орбитальную и медиальную префронтальную кору и поясную кору. Нейроны VTA DA играют решающую роль в мотивации, поведении, связанном с вознаграждением, внимании и множественных формах памяти. ... Таким образом, действуя в различных терминальных полях, дофамин придает мотивационную значимость («желание») самому вознаграждению или связанным с ним сигналам (область оболочки прилежащего ядра), обновляет ценность, придаваемую различным целям в свете этого нового опыта (орбитальная префронтальная часть оболочки). кора), помогает консолидировать несколько форм памяти (миндалевидное тело и гиппокамп) и кодирует новые двигательные программы, которые облегчат получение этого вознаграждения в будущем (область ядра прилежащего ядра и дорсальное полосатое тело). ... ДА выполняет несколько действий в префронтальной коре. Он способствует «когнитивному контролю» поведения: выбору и успешному мониторингу поведения для облегчения достижения выбранных целей. Аспекты когнитивного контроля, в которых играет роль ДА, включают рабочую память, способность удерживать информацию «онлайн», чтобы направлять действия, подавление доминирующего поведения, которое конкурирует с целенаправленными действиями, а также контроль внимания и, следовательно, способность преодолевать отвлекающие факторы. ... Таким образом, норадренергические проекции LC взаимодействуют с дофаминергическими проекциями VTA, регулируя когнитивный контроль. ...
- ^ Калипари Э.С., Багот Р.К., Пурушотаман И., Дэвидсон Т.Дж., Йоргасон Дж.Т., Пенья С.Дж. и др. (март 2016 г.). «Визуализация in vivo идентифицирует временные характеристики средних шиповатых нейронов D1 и D2 в вознаграждении за кокаин» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (10): 2726–31. Бибкод : 2016PNAS..113.2726C . дои : 10.1073/pnas.1521238113 . ПМК 4791010 . ПМИД 26831103 .
Предыдущая работа продемонстрировала, что оптогенетическая стимуляция MSN D1 способствует вознаграждению, тогда как стимуляция MSN D2 вызывает отвращение.
- ^ Икемото С. (ноябрь 2010 г.). «Схема вознаграждения мозга за пределами мезолимбической дофаминовой системы: нейробиологическая теория» . Неврологические и биоповеденческие обзоры . 35 (2): 129–50. doi : 10.1016/j.neubiorev.2010.02.001 . ПМЦ 2894302 . ПМИД 20149820 .
Недавние исследования по внутричерепному самостоятельному введению нейрохимических веществ (лекарств) показали, что крысы учатся самостоятельно вводить различные лекарства в мезолимбические дофаминовые структуры — заднюю вентральную область покрышки, медиальное прилежащее ядро оболочки и медиальный обонятельный бугорок. ... В 1970-х годах было признано, что обонятельный бугорок содержит полосатый компонент, который заполнен ГАМКергическими средними шиповидными нейронами, получающими глутаматергические входы из кортикальных областей и дофаминергические входы из ВТА и проецирующимися на вентральное паллидум, как и прилежащее ядро.
Рисунок 3. Вентральное полосатое тело и самостоятельный прием амфетамина. - ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 6: Широко распространенные системы: моноамины, ацетилхолин и орексин». В Сидоре А., Брауне Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 175–176. ISBN 9780071481274 .
В головном мозге гистамин синтезируется исключительно нейронами, тела которых расположены в туберомаммиллярном ядре (TMN), расположенном в заднем гипоталамусе. У человека на каждой стороне имеется около 64 000 гистаминергических нейронов. Эти клетки проецируются по всему головному и спинному мозгу. Области, которые получают особенно плотные проекции, включают кору головного мозга, гиппокамп, неостриатум, прилежащее ядро, миндалевидное тело и гипоталамус. ... Хотя наиболее изученной функцией гистаминовой системы головного мозга является регуляция сна и пробуждения, гистамин также участвует в обучении и памяти ... Также оказывается, что гистамин участвует в регуляции питания и энергетического баланса.
- ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 6: Широко распространенные системы: моноамины, ацетилхолин и орексин». В Сидоре А., Брауне Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 158–160. ISBN 9780071481274 .
Дорсальный шов преимущественно иннервирует кору головного мозга, таламус, стриарные области (хвостатое ядро и прилежащее ядро) и дофаминергические ядра среднего мозга (например, черную субстанцию и вентральную область покрышки), в то время как срединный шов иннервирует гиппокамп, перегородка и другие структуры лимбической части переднего мозга. ... очевидно, что 5HT влияет на сон, возбуждение, внимание, обработку сенсорной информации в коре головного мозга, а также на важные аспекты эмоций (вероятно, включая агрессию) и регуляцию настроения. ...Ростральные ядра, включающие линейное ядро, дорсальный шов, медиальный шов и мостовой шов, иннервируют большую часть мозга, включая мозжечок. Каудальные ядра, включающие большой шов, бледный шов и неясный шов, имеют более ограниченные отростки, заканчивающиеся в мозжечке, стволе головного мозга и спинном мозге.
- ^ Нестлер Э.Дж. «Пути вознаграждения мозга» . Медицинская школа Икан на горе Синай . Лаборатория Нестлер . Проверено 16 августа 2014 г.
Дорсальный шов является основным местом расположения серотонинергических нейронов головного мозга, которые, как и норадренергические нейроны, повсеместно модулируют функцию мозга, регулируя состояние активации и настроение организма.
- ^ Марстон О.Дж., Гарфилд А.С., Хейслер Л.К. (июнь 2011 г.). «Роль центральных систем серотонина и меланокортина в контроле энергетического баланса». Европейский журнал фармакологии . 660 (1): 70–9. дои : 10.1016/j.ejphar.2010.12.024 . ПМИД 21216242 .
- ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 6: Широко распространенные системы: моноамины, ацетилхолин и орексин». В Сидоре А., Брауне Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 167–175. ISBN 9780071481274 .
Базальные холинергические ядра переднего мозга включают медиальное перегородочное ядро (Ch1), вертикальное ядро диагональной полосы (Ch2), горизонтальное крыло диагональной полосы (Ch3) и базальное ядро Мейнерта (Ch4). Холинергические ядра ствола мозга включают педункулопонтинное ядро (Ch5), латеродорсальное тегментальное ядро (Ch6), медиальную хабенулу (Ch7) и парабигеминальное ядро (Ch8).
- ^ Гайене П.Г., Сторнетта Р.Л., Бочоришвили Г., Депюи С.Д., Берк П.Г., Эбботт С.Б. (август 2013 г.). «Нейроны C1: ЕМТ организма» . Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 305 (3): Р187–204. дои : 10.1152/ajpregu.00054.2013 . ПМЦ 3743001 . ПМИД 23697799 .
- ^ Сторнетта Р.Л., Гайене П.Г. (март 2018 г.). «Нейроны C1: узловая точка стресса?» . Экспериментальная физиология . 103 (3): 332–336. дои : 10.1113/EP086435 . ПМЦ 5832554 . ПМИД 29080216 .
- ^ «Беседы нейронов: как общаются клетки мозга» . Brainfacts.org . Проверено 2 декабря 2014 г.
- ^ Jump up to: а б Ядав В.К., Рю Дж.Х., Суда Н., Танака К.Ф., Гингрич Дж.А., Шютц Г. и др. (ноябрь 2008 г.). «Lrp5 контролирует образование костной ткани, ингибируя синтез серотонина в двенадцатиперстной кишке» . Клетка . 135 (5): 825–37. дои : 10.1016/j.cell.2008.09.059 . ПМЦ 2614332 . ПМИД 19041748 .
- ^ Карлсон, Н.Р., и Биркетт, Массачусетс (2017). Физиология поведения (12-е изд.). Пирсон, стр. 100–115. ISBN 978-0134080918
- ^ «CDC Стрихнин | Факты о стрихнине | Готовность и реагирование общественного здравоохранения на чрезвычайные ситуации» . Emergency.cdc.gov . Проверено 7 мая 2018 г.
- ^ Jump up to: а б с «Таблица нейротрансмиттеров и лекарств» . Ocw.mit.edu . Проверено 25 августа 2014 г.
- ^ «Агонист – определение и многое другое из бесплатного словаря Мерриам-Вебстера» . Merriam-webster.com . Проверено 25 августа 2014 г.
- ^ Атак Дж., Лаврейсен Х. (2010) Агонист. В кн.: Столерман И.П. (ред.) Энциклопедия психофармакологии. Шпрингер, Берлин, Гейдельберг. https://doi.org/10.1007/978-3-540-68706-1_1565
- ^ Рот Б.Л. (февраль 2016 г.). «УЖАСЫ для нейробиологов» . Нейрон . 89 (4): 683–94. дои : 10.1016/j.neuron.2016.01.040 . ПМЦ 4759656 . ПМИД 26889809 .
- ^ Jump up to: а б Райс Р.К., Филлин Д.А., Миллер С.К. (2009). Принципы медицины зависимостей (4-е изд.). Филадельфия: Уолтерс Клювер/Липпинкотт Уильямс и Уилкинс. стр. 709–710. ISBN 9780781774772 . Проверено 16 августа 2015 г.
- ^ Jump up to: а б с Флорес А., Мальдонадо Р., Беррендеро Ф. (декабрь 2013 г.). «Перекрестные помехи каннабиноидов и гипокретина в центральной нервной системе: что мы знаем на данный момент» . Границы в неврологии . 7 : 256. дои : 10.3389/fnins.2013.00256 . ПМЦ 3868890 . ПМИД 24391536 .
• Рисунок 1: Схема экспрессии CB1 головного мозга и орексинергических нейронов, экспрессирующих OX1 или OX2.
• Рисунок 2: Механизмы синаптической передачи сигналов в системах каннабиноидов и орексина. - ^ «Антагонист» . Медицинское определение Антагониста . Проверено 5 ноября 2014 г.
- ^ Гедерс Н.Е. (2001). «Антагонист» . Энциклопедия наркотиков, алкоголя и аддиктивного поведения . Проверено 2 декабря 2014 г.
- ^ Бродли К.Дж. (март 2010 г.). «Сосудистые эффекты следовых аминов и амфетаминов». Фармакология и терапия . 125 (3): 363–375. doi : 10.1016/j.pharmthera.2009.11.005 . ПМИД 19948186 .
- ^ Линдеманн Л., Хонер MC (май 2005 г.). «Ренессанс следовых аминов, вдохновленный новым семейством GPCR». Тенденции в фармакологических науках . 26 (5): 274–281. дои : 10.1016/j.tips.2005.03.007 . ПМИД 15860375 .
- ^ Ван X, Ли Дж, Донг Дж, Юэ Дж (февраль 2014 г.). «Эндогенные субстраты CYP2D мозга». Европейский журнал фармакологии . 724 : 211–218. дои : 10.1016/j.ejphar.2013.12.025 . ПМИД 24374199 .
- ^ Jump up to: а б с д и Мейерс С. (февраль 2000 г.). «Использование предшественников нейромедиаторов для лечения депрессии» (PDF) . Обзор альтернативной медицины . 5 (1): 64–71. ПМИД 10696120 . Архивировано из оригинала (PDF) 5 августа 2004 года.
- ^ ван Прааг Х.М. (март 1981 г.). «Управление депрессией с помощью предшественников серотонина». Биологическая психиатрия . 16 (3): 291–310. ПМИД 6164407 .
- ^ Хили Д. (апрель 2015 г.). «Серотонин и депрессия». БМЖ . 350 :h1771. дои : 10.1136/bmj.h1771 . ПМИД 25900074 . S2CID 38726584 .
- ^ «Основы мозга NIMH» . Национальные институты здравоохранения США . Архивировано из оригинала 29 октября 2014 года . Проверено 29 октября 2014 г.
- ^ Биттигау П., Икономиду С (ноябрь 1997 г.). «Глутамат при неврологических заболеваниях». Журнал детской неврологии . 12 (8): 471–85. дои : 10.1177/088307389701200802 . ПМИД 9430311 . S2CID 1258390 .
- ^ Неттер, П. (1991) Биохимические переменные в изучении темперамента. В Стрелау, Дж. и Англейтнер, А. (ред.), Исследования темперамента: международные взгляды на теорию и измерения 147–161. Нью-Йорк: Пленум Пресс.
- ^ Трофимова И, Роббинс Т.В. (май 2016 г.). «Темперамент и системы возбуждения: новый синтез дифференциальной психологии и функциональной нейрохимии» . Неврологические и биоповеденческие обзоры . 64 : 382–402. doi : 10.1016/j.neubiorev.2016.03.008 . hdl : 11375/26202 . ПМИД 26969100 . S2CID 13937324 .
- ^ Клонингер Ч.Р., Свракич Д.М., Пшибек Т.Р. Психобиологическая модель темперамента и характера» Arch Gen Psychiatry 1993; 50:975-990.
- ^ Трофимова И.Н. (2016). «Взаимосвязь между функциональными аспектами деятельности и нейрохимической моделью темперамента взрослого человека». В Арнольде MC (ред.). Темпераменты: индивидуальные различия, социальные и экологические влияния и влияние на качество жизни . Нью-Йорк: Nova Science Publishers, Inc., стр. 77–147.
- ^ Депю Р.А., Морроне-Струпинский СП (июнь 2005 г.). «Нейроповеденческая модель аффилиативной связи: значение для концептуализации человеческой черты принадлежности». Поведенческие и мозговые науки . 28 (3): 313–50, обсуждение 350–95. дои : 10.1017/s0140525x05000063 . ПМИД 16209725 .
- ↑ Лео Дж. И Лакасс Дж. (10 октября 2007 г.). СМИ и теория химического дисбаланса депрессии. Получено 1 декабря 2014 г. с сайта http://psychrights.org/articles/TheMediaandChemicalImbalanceTheoryofDepression.pdf .
- ^ Добрякова Е., Дженова Х.М., ДеЛука Дж., Уайли Г.Р. (12 марта 2015 г.). «Гипотеза дисбаланса дофамина об усталости при рассеянном склерозе и других неврологических расстройствах» . Границы в неврологии . 6:52 . doi : 10.3389/fneur.2015.00052 . ПМЦ 4357260 . ПМИД 25814977 .
Внешние ссылки
[ редактировать ]- Первс, Дейл; Августин, Джордж Дж.; Фитцпатрик, Дэвид; Кац, Лоуренс К.; ЛаМантия, Энтони-Самуэль; Макнамара, Джеймс О.; Уильямс, С. Марк (2001). «Глава 6. Нейромедиаторы». Что определяет нейротрансмиттер? (2-е изд.). Сандерленд (Массачусетс): Sinauer Associates. ISBN 0-87893-742-0 .
{{cite book}}:|journal=игнорируется ( помогите ) - Хольц, Рональд В.; Фишер, Стивен К. (1999). «Глава 10. Синаптическая передача и сотовая сигнализация: обзор». В Сигеле, Джордж Дж; Агранов, Бернард В.; Альберс, Р. Уэйн; Фишер, Стивен К.; Улер, Майкл Д. (ред.). Синаптическая передача (6-е изд.). Филадельфия: Липпинкотт-Рэйвен. ISBN 0-397-51820-Х .
{{cite book}}:|journal=игнорируется ( помогите ) - Нейротрансмиттеры и нейроактивные пептиды на веб-сайте Neuroscience for Kids
