Актин
| Актин | |||
|---|---|---|---|
 Ленточная диаграмма G-Actin. ADP, актина связанный с активным участком (многоцветные палочки вблизи центра рисунка), а также комплексную дикацию кальция (зеленая сфера). Подчеркнут [ 1 ] | |||
| Идентификаторы | |||
| Символ | Актин | ||
| Pfam | PF00022 | ||
| InterPro | IPR004000 | ||
| PROSITE | PDOC00340 | ||
| Краткое содержание | 2btf / scope / supfam | ||
| |||
Актин представляет собой семейство глобулярных , многофункциональных белков которые образуют микрофиламенты в цитоскелете , и тонкие нити в мышечных фибриллах . Он обнаруживается практически во всех эукариотических клетках , где он может присутствовать в концентрации более 100 мкМ ; Его масса составляет примерно 42 кДа , с диаметром от 4 до 7 нм.
Актиновый белок является мономерной субъединицей двух типов филам в клетках: микрофиламентах , один из трех основных компонентов цитоскелета и тонких филаментов, часть сократительного аппарата в мышечных клетках. Он может присутствовать в виде свободного мономера , называемого G-актином (глобулярный), либо как часть линейного полимерного микрофиламента, называемого F-актина (ниткого), оба из которых необходимы для таких важных клеточных функций, как и сокращение клеток подвижность клеточная деление .
Актин участвует во многих важных клеточных процессах, включая сокращение мышц , подвижность клеток , деление клеток и цитокинез , пузырьков и органелл перемещение , передачу сигналов клеток , а также установление и поддержание клеточных соединений и формы клеток. Многие из этих процессов опосредованы обширным и интимным взаимодействием актина с клеточными мембранами . [ 2 ] У позвоночных три основные группы изоформ актина , альфа , бета и гамма были идентифицированы . Альфа -актины, обнаруженные в мышечных тканях, являются основным составляющим сократительного аппарата. Бета и гамма -актины сосуществуют в большинстве типов клеток в качестве компонентов цитоскелета и в качестве медиаторов внутренней клеток мощности . Считается, что разнообразный диапазон структур, образованных актином, позволяющим ему выполнять такой большой диапазон функций, регулируется посредством связывания тропомиозина вдоль филаментов. [ 3 ]
Способность ячейки динамически формировать микрофиламенты обеспечивает каркасы, которые позволяют ему быстро реконструировать себя в ответ на окружающую среду или на внутренние сигналы организма , например, увеличить абсорбцию клеточной мембраны или увеличить адгезию клеток с образованием клеточной ткани . Другие ферменты или органеллы , такие как реснички, могут быть прикреплены к этим лесам, чтобы контролировать деформацию внешней клеточной мембраны , которая позволяет эндоцитоз и цитокинез . Он также может создавать движение само по себе или с помощью молекулярных двигателей . Таким образом, актин способствует таким процессам, как внутриклеточный транспорт везикул и органеллов, а также мышечное сокращение и клеточная миграция . Поэтому он играет важную роль в эмбриогенезе , заживлении ран и инвазивности раковых клеток. Эволюционное происхождение актина можно проследить до прокариотических клеток , которые имеют эквивалентные белки. [ 4 ] Гомологи актина из прокариот и археи полимеризуются в различные спиральные или линейные филаменты, состоящие из одной или множественных цепей. Однако контакты в цепи и сайты связывания нуклеотидов сохраняются в прокариотах и в археи. [ 5 ] Наконец, актин играет важную роль в контроле экспрессии генов .
Большое количество заболеваний и заболеваний вызвано мутациями в аллелях генов , которые регулируют выработку актина или связанных с ним белков. Производство актина также является ключом к процессу инфекции некоторыми патогенными микроорганизмами . Мутации в различных генах, которые регулируют выработку актина у людей, могут вызывать мышечные заболевания , вариации размера и функции сердца , а также глухоты . Состав цитоскелета также связан с патогенностью внутриклеточных бактерий и вирусов , особенно в процессах, связанных с уклонением от действий иммунной системы . [ 6 ]
Функция
[ редактировать ]Основная роль актина в клетке состоит в том, чтобы образовать линейные полимеры, называемые микрофиламентами , которые выполняют различные функции в структуре клетки, сети транспорта, миграции и репликации. [ 7 ] Многогранная роль актина зависит от некоторых свойств микрофиламентов: во -первых, образование актиновых филаментов обратимо, и их функция часто включает в себя быструю полимеризацию и деполимеризацию. Во -вторых, микрофиламенты поляризованы - т.е. два конца филамента отличаются друг от друга. В -третьих, актиновые филаменты могут связываться со многими другими белками, которые вместе помогают модифицировать и организовать микрофиламенты для их разнообразных функций. [ 7 ]
В большинстве клеток актиновые филаменты образуют более масштабные сети, которые необходимы для многих ключевых функций: [ 8 ]
- Актиновые сети обеспечивают механическую поддержку клеткам и обеспечивают маршруты переноса через цитоплазму, чтобы помочь передаче сигнала.
- Быстрая сборка и разборка актиновой сети позволяют клеткам мигрировать ( миграция клеток ).
Актин чрезвычайно распространен в большинстве клеток, составляющий 1–5% от общей массы белка большинства клеток и 10% мышечных клеток. [ 7 ]
Актиновый белок обнаруживается как в цитоплазме , так и в ядре клеток . [ 9 ] Его местоположение регулируется путями трансдукции сигнала клеточной мембраны , которые интегрируют стимулы, которые ячейка получает, стимулирующую реструктуризацию актиновых сетей в ответ. [ 10 ]
Цитоскелет
[ редактировать ]
Существует ряд различных типов актина с немного разными структурами и функциями. α-актин обнаруживается исключительно в мышечных волокнах , в то время как β- и γ-актин обнаруживается в других клетках. Поскольку последние типы имеют высокий уровень оборота, большинство из них находятся за пределами постоянных структур. Микрофиламенты, обнаруженные в клетках, кроме мышечных клеток, присутствуют в трех формах: [ 11 ]
- Микрофиламентные сети - клетки животных обычно имеют клеточную кору под клеточной мембраной , которая содержит большое количество микрофиламентов, которые исключают присутствие органеллов . Эта сеть связана с многочисленными рецепторами , которые передают сигналы снаружи ячейки.
- Периодические актиновые кольца - периодическая структура, построенная из равномерно распределенных актиновых колец, обнаружена в аксонах . [ 12 ] В этой структуре актиновые кольца вместе со спектриновыми тетрамерами, которые связывают соседние кольца актина, образуют сплоченный цитоскелет , который поддерживает мембрану аксонов. Периодичность структуры может также регулировать ионные каналы натрия в аксонах.
Дрожжи
[ редактировать ]Цитоскелет актина является ключом к процессам эндоцитоза , цитокинеза , определения полярности клеток и морфогенеза в дрожжах . В дополнение к полаганию на актин, эти процессы включают 20 или 30 связанных белков, которые имеют высокую степень эволюционного сохранения, наряду со многими сигнальными молекулами. Вместе эти элементы позволяют пространственно и временно модулируемому сборке, которая определяет реакцию ячейки как на внутренние, так и на внешние стимулы. [ 13 ]
Дрожжи содержат три основных элемента, которые связаны с актином: пятнами, кабелями и кольцами. Несмотря на то, что эти структуры не присутствуют, эти структуры подвергаются динамическому равновесию из -за постоянной полимеризации и деполимеризации. У них есть ряд вспомогательных белков, включая ADF/кофилин, который имеет молекулярную массу 16 кДа и кодируется одним геном, называемым COF1 ; AIP1, кофлиновый кофактор, который способствует разборке микрофиламентов; SRV2/CAP, регулятор процесса, связанный с аденилатциклазы белками ; профилин с молекулярной массой приблизительно 14 кДа, который связан/связан с актиновыми мономерами; и Twinfilin, белок 40 кДа, участвующий в организации участков. [ 13 ]
Растения
[ редактировать ]растений Исследования генома выявили существование белковых изоваринов в семействе генов актина. В рамках Arabidopsis Thaliana , модельного организма , существует десять типов актина, шесть профилинов и десятки миозинов. Это разнообразие объясняется эволюционной необходимостью обладания вариантами, которые немного отличаются по их временному и пространственному выражению. [ 4 ] Большинство из этих белков были совместно экспрессированы в анализируемой ткани . Актиновые сети распределяются по всей цитоплазме клеток, которые культивированы in vitro . Существует концентрация сети вокруг ядра, которая подключена через спицы с клеточной корой, эта сеть очень динамична, с непрерывной полимеризацией и деполимеризацией. [ 14 ]

Несмотря на то, что у большинства растительных клеток есть клеточная стенка , которая определяет их морфологию, их микрофиламенты могут генерировать достаточную силу для достижения ряда клеточных активностей, таких как цитоплазматические токи, генерируемые микрофиламентами и миозином. Актин также участвует в движении органеллов и клеточного морфогенеза, который включает в себя деление клеток, а также удлинение и дифференцировку клетки. [ 16 ]
Наиболее заметные белки, связанные с актиновым цитоскелетом в растениях, включают: [ 16 ] Villin , который принадлежит к тому же семейству, что и Gelsolin /Severin и способен разрезать микрофиламенты и связывать мономеры актина в присутствии катионов кальция; фимбрин , который способен распознавать и объединять актиновые мономеры и который участвует в формировании сетей (в результате другого процесса регуляции от процесса животных и дрожжей); [ 17 ] Форманы , которые способны действовать как зарождающаяся полимеризация F-актина; миозин , типичный молекулярный мотор, специфичный для эукариот и который у Arabidopsis thaliana кодируется 17 генами в двух разных классах; CHUP1, который может связывать актин и участвует в пространственном распределении хлоропластов в клетке; KAM1/MUR3, который определяет морфологию аппарата Гольджи , а также состав ксилоглюканов в клеточной стенке; Ntwlim1, который облегчает появление структур клеток актина; и ERD10, который участвует в ассоциации органеллов в мембранах и микрофиламентах и которая, по -видимому, играет роль, которая участвует в реакции организма на стресс .
Ядерный актин
[ редактировать ]Ядерный актин был впервые замечен и описан в 1977 году Кларком и Мерриамом. [ 18 ] Авторы описывают белок, присутствующий в ядерной фракции, полученный из ооцитов Xenopus laevis , которые показывают те же особенности, что и актин скелетных мышц. С тех пор было много научных отчетов о структуре и функциях актина в ядре (для обзора см.: Hofmann 2009. [ 19 ] ) Контролируемый уровень актина в ядре, его взаимодействие с актин-связывающими белками (ABP) и наличие различных изоформ позволяют актину играть важную роль во многих важных ядерных процессах. [ 20 ]
Транспорт через ядерную мембрану
[ редактировать ]Последовательность актина не содержит сигнала ядерной локализации. Небольшой размер актина (около 43 кДа) позволяет ему войти в ядро путем пассивной диффузии. [ 21 ] Импорт актина в ядро (вероятно, в комплексе с кофилином) облегчается импортным белком Importin 9. [ 22 ]
Низкие уровни актина в ядре, по -видимому, важны, потому что актин имеет два ядерных экспортных сигнала (NES) в своей последовательности. Микроинъектированный актин быстро удаляется из ядра в цитоплазму. Актин экспортируется, по крайней мере, двумя способами, через экспортину 1 и экспорта 6 . [ 23 ] [ 24 ] Конкретные модификации, такие как сумоилирование, допускают удержание ядерного актина. Мутация, предотвращая сумоилирование, вызывает быстрый экспорт бета -актина из ядра. [ 25 ]
Организация
[ редактировать ]Ядерный актин существует в основном как мономер, но также может образовывать динамические олигомеры и короткие полимеры. [ 26 ] [ 27 ] [ 28 ] Организация ядерного актина варьируется в разных типах клеток. Например, в ооцитах Xenopus (с более высоким уровнем ядерного актина по сравнению с соматическими клетками) актин образует филаменты, которые стабилизируют архитектуру ядра. Эти филаменты можно наблюдать под микроскопом благодаря окрашиванию флуорофором фаллоидином. [ 18 ] [ 21 ]
Однако в ядрах соматических клеток актиновые филаменты не могут наблюдаться с использованием этой методики. [ 29 ] Анализ ингибирования ДНКазы I, единственный тест, который позволяет количественно определять полимеризованный актин непосредственно в биологических образцах, показал, что эндогенный ядерный актин действительно встречается в основном в мономерной форме. [ 28 ]
Точно контролируемый уровень актина в клеточном ядре, ниже, чем в цитоплазме, предотвращает образование филаментов. Полимеризация также уменьшается за счет ограниченного доступа к актиновым мономерам, которые связаны в комплексах с ABP, в основном кофилином. [ 30 ]
Актиновые изоформы
[ редактировать ]Различные изоформы актина присутствуют в клеточном ядре. Уровень изоформ актина может измениться в ответ на стимуляцию роста клеток или остановку пролиферации и транскрипционной активности. [ 31 ] Исследования по ядерному актину сосредоточены на бета -батах изоформы. [ 32 ] [ 33 ] [ 34 ] [ 35 ] Однако использование антител, направленных против различных изоформ актина, позволяет идентифицировать не только цитоплазматическую бета в клеточном ядре, но и альфа-и гамма-актин в определенных типах клеток. [ 28 ] [ 36 ] [ 37 ] Наличие различных изоформ актина может оказать существенное влияние на его функцию в ядерных процессах, поскольку уровень отдельных изоформ может контролироваться независимо. [ 28 ]
Функции
[ редактировать ]Функции актина в ядре связаны с его способностью полимеризировать и взаимодействовать с различными ABP и со структурными элементами ядра. Ядерный актин участвует в:
- Архитектура ядра - взаимодействие актина с альфа -II -спектрином и другими белками важна для поддержания правильной формы ядра. [ 38 ] [ 39 ]
- Транскрипция - актин участвует в реорганизации хроматина, [ 9 ] [ 32 ] [ 40 ] [ 41 ] инициация транскрипции и взаимодействие с комплексом транскрипции. [ 42 ] Актин принимает участие в регуляции структуры хроматина, [ 43 ] [ 44 ] [ 45 ] взаимодействие с РНК -полимеразой I, [ 35 ] II [ 33 ] и iii. [ 34 ] В Pol I транскрипции актин и миозин ( MYO1C , который связывает ДНК), действуют как молекулярный мотор . Для транскрипции Pol II β-актин необходим для образования предварительного комплекса. Pol III содержит β-актин в качестве субъединицы. Актитин также может быть компонентом комплексов ремоделирования хроматина, а также частиц Pre-MRNP (то есть предшественника Мессенджер РНК, связанной с белками) и участвует в ядерном экспорте РНК и белков. [ 46 ]
- Регуляция активности генов - актин связывается с регуляторными областями различных видов генов. [ 47 ] [ 48 ] [ 49 ] [ 50 ] Способность актина регулировать активность генов используется в методе молекулярного перепрограммирования, который позволяет дифференцированным клеткам возвращаться в свое эмбриональное состояние. [ 49 ] [ 51 ]
- Транслокация активированного фрагмента хромосомы из под мембранной области в эухроматин, где начинается транскрипция. Это движение требует взаимодействия актина и миозина. [ 52 ] [ 53 ]
- Интеграция различных клеточных компартментов . Актин - это молекула, которая интегрирует пути цитоплазматической и ядерной передачи сигнала. [ 54 ] Примером является активация транскрипции в ответ на стимуляцию сыворотки клеток in vitro . [ 55 ] [ 56 ] [ 57 ]
- Иммунный ответ - ядерный актин полимеризуется при стимуляции рецептора Т -клеток и необходим для экспрессии цитокинов и выработки антител in vivo . [ 58 ]
- Репарация ДНК - ядерный актин опосредует восстановление двойных разрывов ДНК . [ 59 ] В клеточном ядре нитевидное полимер актина (F-актин) действует как в пути репарации ДНК не гомологичного конца соединения, так и на пути гомологичного рекомбинационного восстановления . [ 59 ]
Благодаря своей способности преодолевать конформационные изменения и взаимодействие со многими белками, актин действует как регулятор образования и активности белковых комплексов, таких как транскрипционный комплекс. [ 42 ]
Движение клеток
[ редактировать ]Актин также участвует в движении клеток. Сетка актиновых филаментов отмечает передний край движущейся ячейки, а полимеризация новых актиновых филаментов подталкивает клеточную мембрану вперед в выступах, называемых Lamellipodia . [ 60 ] [ 61 ] [ 62 ] Эти мембранные выступы затем прикрепляются к субстрату, образуя структуры, известные как фокальные спайки , которые подключаются к сети актина. [ 62 ] После прикрепления задняя часть тела ячейки сокращается, сжимая свое содержимое вперед мимо точки адгезии. [ 62 ] Как только точка адгезии перенесена в заднюю часть ячейки, ячейка разбирает ее, позволяя задней клетке двигаться вперед. [ 62 ]

Движение актина/миозина
[ редактировать ]В дополнение к физической силе, генерируемой актиновой полимеризацией, микрофиламенты способствуют движению различных внутриклеточных компонентов, служа в качестве проезжей части, вдоль которого семейство моторных белков, называемых перемещением миозинов . [ 63 ]
Мышечное сокращение
[ редактировать ]Актин играет особенно заметную роль в мышечных клетках, которые в основном состоят из повторных пучков актина и миозина II . [ 64 ] Каждое повторное устройство-называемое саркомером -состоит из двух наборов противоположно ориентированных цепей F-актина («тонкие нити»), переплетенные с пучками миозина («толстые нити»). Два набора пряди актина ориентированы на их (+) концы, встроенные в любой конец саркомера в разграничивающие структуры, называемые z-дискими . [ 64 ] Миозиновые фибриллы находятся в середине между наборами актиновых филаментов, с нити, обращенными в обоих направлениях. Когда мышцы сокращаются, нити миозина движутся вдоль актиновых филаментов в направлении (+) конца, сталкивая концы саркомера и сокращая его примерно на 70% от его длины. [ 64 ] Чтобы двигаться по актиновой нити, миозин должен гидролизовать АТФ; Таким образом, АТФ служит источником энергии для сокращения мышц. [ 64 ]
Во времена отдыха белки тропомиозин и тропонин связываются с актиновыми филаментами, предотвращая прикрепление миозина. [ 64 ] Когда сигнал активации (то есть потенциал действия ) прибывает на мышечное волокно, он запускает высвобождение CA 2+ от саркоплазматического ретикулума в цитозоль. Полученный всплеск в цитозольном кальцие быстро высвобождает тропомиозин и тропонин из актиновой нити, позволяя связывать миозин, и начинается противоречие с мышц. [ 65 ]
Клеточная деление
[ редактировать ]На последних этапах клеточного деления многие ячейки образуют кольцо актина в средней точке клетки. Это кольцо, удачно называемое « сократительным кольцом », использует аналогичный механизм, как мышечные волокна, где миозин II тянет вдоль актинового кольца, заставляя его сжиматься. [ 66 ] Это сокращение расщепляет родительскую ячейку на две части, завершая цитокинез . [ 66 ] Сократительное кольцо состоит из актина, миозина, аниллина и α-актинина . [ 67 ] В делящихся дрожжах Schizosaccharomyces pombe актин активно образуется в сжимающем кольце с участием ARP3 , Formin CDC12, Profilin и WASP , а также предварительно сформированных микрофиламентов. После того, как кольцо было построено, структура поддерживается постоянной сборкой и разборкой, которая, помогая комплекс ARP2/3 и формами, является ключом к одному из центральных процессов цитокинеза. [ 68 ]
Внутриклеточная торговля
[ редактировать ]Пары актина-миозина также могут участвовать в переносе различных мембранных пузырьков и органеллов в клетке. Миозин V активируется путем связывания с различными грузовыми рецепторами на органелле, а затем движется вдоль актиновой нити в сторону (+) конец, тянуя его груз вместе с ним. [ 69 ]
Эти нетрадиционные миозины используют гидролиз АТФ для транспортировки груза, такого как везикулы и органелл, в направлении, намного быстрее, чем диффузия. Myosin V идет к колючим концу актиновых филаментов, в то время как миозин VI идет к островам конце. Большинство актиновых филаментов расположены с колючей концом в направлении клеточной мембраны и заостренным концом к клеточной внутренней части. Такое расположение позволяет миозину V быть эффективным двигателем для экспорта автомобилей, а миозин VI - это эффективный двигатель для импорта.
Другие биологические процессы
[ редактировать ]Традиционный образ функции актина связан с поддержанием цитоскелета и, следовательно, организации и движением органеллов, а также к определению формы клетки. [ 11 ] Тем не менее, актин играет более широкую роль в физиологии эукариотических клеток, в дополнение к аналогичным функциям в прокариотах .
- Апоптоз . Во время запрограммированной гибели клеток семейство протеаз ICE/CED-3 (один из протеаз межлейкин-1β-конвертер) разлагает актин в два фрагмента in vivo ; Один из фрагментов - 15 кДа, а другой - 31 кДа. Это представляет собой один из механизмов, участвующих в разрушении жизнеспособности клеток, которые формируют основу апоптоза. [ 70 ] Также было показано, что протеазовый кальпаин участвует в этом типе разрушения клеток; [ 71 ] Так же, как было показано, что использование ингибиторов кальпаина снижает протеолиз актина и деградацию ДНК (еще один характерный элемент апоптоза). [ 72 ] С другой стороны, вызванный стрессом запуск апоптоза вызывает реорганизацию актинового цитоскелета (который также включает его полимеризацию), что приводит к структурам, называемым стрессовыми волокнами ; Это активируется путем киназы карты . [ 73 ]

- Клеточная адгезия и развитие . Адгезия между клетками является характерной для многоклеточных организмов , которые обеспечивают специализацию тканей и, следовательно, увеличивают сложность клеток. Адгезия клеточной эпителии включает в себя актиновый цитоскелет в каждой из соединенных клеток, а также кадгерины, действующие в качестве внеклеточных элементов, с соединением между двумя опосредованными катеенинами . [ 74 ] Вмешательство в актиновую динамику имеет последствия для развития организма, на самом деле актин является таким важным элементом, что системы избыточных генов доступны . Например, если ген α-актининового или гелевого фактора был удален у индивидуумов диктиостелия , не показывает аномальный фенотип , возможно, из-за того, что каждый из белков может выполнять функцию другого. развитие двойных мутаций , в которых отсутствует оба типа генов. Тем не менее, затрагивает на [ 75 ]
- экспрессии генов Модуляция . Состояние полимеризации актина влияет на паттерн экспрессии генов . В 1997 году было обнаружено, что опосредованная цитокалазин D-опосредованная деполимеризация в клетках Шванна вызывает специфический характер экспрессии для генов, участвующих в миелинизации этого типа нервной клетки . [ 76 ] Было показано, что F-актин модифицирует транскриптом на некоторых этапах жизни одноклеточных организмов, таких как гриб- кандида-альбиканы . [ 77 ] Кроме того, белки, сходные с актином, играют регуляторную роль во время сперматогенеза у мышей [ 78 ] и в дрожжах, как полагают, актиноподобные белки играют роль в регуляции экспрессии генов . [ 79 ] Фактически, актин способен действовать в качестве инициатора транскрипции, когда он реагирует с типом ядерного миозина, который взаимодействует с РНК -полимеразами и другими ферментами, участвующими в процессе транскрипции. [ 9 ]
- Динамика стереоцилии . Некоторые ячейки развивают мелкие фирменные расстройства на их поверхности, которые имеют механо -енсорную функцию. Например, этот тип органелл присутствует в органе Корти , который расположен в ухе . Основная характеристика этих структур заключается в том, что их длина может быть изменена. [ 80 ] Молекулярная архитектура стереоцилии включает в себя паракристаллическое ядро актина в динамическом равновесии с мономерами, присутствующими в соседнем цитозоле. Миозины типа VI и VIIA присутствуют на протяжении всего этого ядра, в то время как миозин XVA присутствует в его конечностях в количествах, которые пропорциональны длине стереоцилии. [ 81 ]
- Внутренняя хиральность . Актомиозиновые сети участвуют в создании внутренней хиральности в отдельных клетках. [ 82 ] Клетки, выращенные на хиральных поверхностях, могут показывать направленное левое/правое смещение, которое зависит от актомиозина. [ 83 ] [ 84 ]
Структура
[ редактировать ]
Мономерный актин, или G-актин, имеет глобулярную структуру, состоящую из двух дол, разделенных глубокой расщелиной. [ 85 ] Дно расщелины представляет собой «складку АТФазы», структуру, консервированную среди АТФ и GTP-связывающих белков, которая связывается с ионом магния и молекулой АТФ. [ 85 ] Связывание АТФ или АДП необходимо для стабилизации каждого актинового мономера; Без одной из этих молекул актин быстро становится денатурированным . [ 85 ]
Рентгеновская кристаллографическая модель актина, которая была продуцирована Кабшем из полосатой мышечной ткани кроликов была , наиболее часто используется в структурных исследованиях, поскольку она была первой, которая очищена . G-Actin, кристаллизованный Kabsch, приблизительно 67 x 40 x 37 Å составляет , имеет молекулярную массу 41 785 DA и предполагаемую изоэлектрическую точку 4,8. Его чистый заряд при pH = 7 составляет -7. [ 86 ] [ 87 ]
- Первичная структура
Elzinga и коллеги сначала определили полную пептидную последовательность для этого типа актина в 1973 году, причем более поздняя работа того же автора добавила к модели дополнительные детали. Он содержит 374 аминокислотных остатков. Его N-конец очень кислый и начинается с в аспартата аминогруппе. В то время как его С-конце является щелочным и образуется фенилаланином, которому предшествовал цистеин , который имеет определенную степень функционального значения. Обе крайности находятся в непосредственной близости в I-субдомене. Аномальный n Т -Метелхистидин расположен в позиции 73. [ 87 ]
- Третичная структура - домены
Третичная структура образуется двумя доменами, известными как большие и малые, которые разделены расщелиной, сосредоточенной вокруг расположения связи с АТФ - ADP + P i . Ниже это более глубокая выемка, называемая «канавкой». В родном состоянии , несмотря на их имена, оба имеют сопоставимую глубину. [ 86 ]
Нормальное соглашение в топологических исследованиях означает, что белок показан с самой большой доменом на левой стороне и наименьшим доменом с правой стороны. В этом положении меньший домен, в свою очередь, разделен на два: поддомен I (нижнее положение, остатки 1–32, 70–144 и 338–374) и поддомен II (верхняя позиция, остатки 33–69). Большая домен также делится на два: поддомен III (нижний, остатки 145–180 и 270–337) и поддомен IV (выше, остатки 181–269). Разоблаченные участки поддоменов I и III называются «колючей» заканчиваются, в то время как открытые области доменов II и IV называются «заостренными» концами. Эта номенклатура относится к тому факту, что из -за небольшой массы субдомена II актин полярно; Важность этого будет обсуждаться ниже в обсуждении динамики сборки. Некоторые авторы называют субдомены IA, IB, IIA и IIB соответственно. [ 88 ]
- Другие важные структуры
Наиболее заметной супер-классовой структурой является пять цепных бета-листов , который состоит из β-среднего и β-β-β-единицы по часовой стрелке. Он присутствует в обоих доменах, предполагая, что белок возник из дупликации генов. [ 89 ]
- Сайт связывания нуклеотидов аденозина расположен между двумя конструкциями в форме бета -шпильки, относящихся к доменам I и III. Остатки, которые вовлечены, являются ASP11-LYS18 и ASP154-HIS161 соответственно.
- Двалентный сайт связывания катионов расположен чуть ниже, чтобы для аденозинового нуклеотида. In vivo это чаще всего образуется MG 2+ или 2+ В то время как in vitro это образуется хелатирующей структурой, состоящей из Lys18 и двух оксигенов нуклеотида из α-и β- фосфатов . Этот кальций координируется с шестью молекулами воды, которые сохраняются аминокислотами ASP11 , ASP154 и GLN137 . Они образуют комплекс с нуклеотидом, который ограничивает движения так называемой области «шарнир», расположенной между остатками 137 и 144. Это сохраняет нативную форму белка до тех пор, пока его отмену денаюры актинового мономера. Эта область также важна, потому что он определяет, находится ли расщелина белка в «открытой» или «закрытой» конформации. [ 1 ] [ 88 ]
- Весьма вероятно, что существует как минимум три других центра с меньшим сродством (промежуточным) и третьими с низким сродством к двухвалентным катионам. Было высказано предположение, что эти центры могут играть роль в полимеризации актина, действуя на стадии активации. [ 88 ]
- В поддомене 2 существует структура, которая называется «D-петли», потому что она связывает ДНКазу I , она расположена между остатками HIS40 и GLY48 . Он имеет появление беспорядочного элемента в большинстве кристаллов, но он выглядит как β-лист, когда он комплекс с ДНКазой I. Предположено, что ключевым событием в полимеризации, вероятно, является распространение конформационного изменения из Центр связи с нуклеотидом с этим доменом, который меняется от петли на спираль. [ 1 ] Однако эта гипотеза была опровергнута другими исследованиями. [ 90 ]
F-Actin
[ редактировать ]
В различных условиях молекулы G-актина полимеризуются в более длинные нити, называемые «нитими, или« f-Actin ». Эти F-актиновые нити обычно состоят из двух спиральных цепей актина, наводненных вокруг друг друга, образуя ширину от 7 до 9 нанометра , которая повторяется каждые 72 нанометров (или каждые 14 субъединиц G-актина). [ 92 ] В нитях F-актина все молекулы G-актина ориентированы в одном и том же направлении. Два конца нити F-актина отличаются друг от друга. На одном конце-обозначен, конец (-)-АТФ-связывающая расщелина терминальной молекулы актина сталкивается наружу. На противоположном конце-назначено (+)-АТФ-связывающая расщелина похоронена в филаменте, контактируя с соседней молекулой актина. [ 92 ] По мере роста F-актиновых нитей новые молекулы имеют тенденцию соединяться на (+) конце существующей прядь F-актина. И наоборот, нити имеют тенденцию к сокращению, выбросив мономеры актина с конца ( -). [ 92 ]
Некоторые белки, такие как кофилин , по -видимому, увеличивают угол поворота, но опять же это можно интерпретировать как установление различных структурных состояний. Это может быть важно в процессе полимеризации. [ 93 ]
Согласно измерениям радиуса поворота и толщины накаливания меньше: в то время как первые модели присваивали длину 25 Å, текущие рентгеновские дифракционные данные, подкрепленные криоэлектронной микроскопией, предполагают длину 23,7 Å. Эти исследования показали точные точки контакта между мономерами. Некоторые из них образуются с единицами одной и той же цепи, между «колючей» конец на одном мономере и «острым» конец следующего. В то время как мономеры в соседних цепях вступают в боковой контакт через проекции из поддомена IV, причем наиболее важными проекциями являются те, которые образованы С-концевой и гидрофобной связью, образованными тремя телами, включающими остатки 39–42, 201–203 и 286. Модель предполагает, что нить образуется мономерами в формировании «листа», в которой субдомены обращаются вокруг себя, эта форма также обнаруживается в бактериальном актиновом гомологе Mreb . [ 94 ]
Термины «заостренные» и «колючие», относящиеся к двум концам микрофиламентов, вытекают из их появления при просвечивающей электронной микроскопии , когда образцы исследуются с помощью метода подготовки, называемого «украшения». Этот метод состоит из добавления фрагментов миозина S1 в ткани, которые были зафиксированы с помощью дубильной кислоты . Этот миозин образует полярные связи с актиновыми мономерами, что приводит к конфигурации, которая выглядит как стрелки с перьями вдоль его вала, где вал - это актин, а Fletchings - миозин. Следуя этой логике, конец микрофиламента, который не имеет какого -либо выступающего миозина, называется точкой стрелки ( - Энд), а другой конец называется колючей конец (+ конец). [ 95 ] Фрагмент S1 состоит из доменов головы и шеи миозина II . В физиологических условиях G-актин ( мономерная форма) трансформируется в F-актин ( полимерная форма) с помощью АТФ, где важна роль АТФ. [ 96 ]
Спиральная F-актиновая нить, обнаруженная в мышцах, также содержит молекулу тропомиозина , которая представляет собой белок длиной 40 нанометра , который обернут вокруг спирали F-Actin. [ 97 ] Во время фазы покоя тропомиозин охватывает активные участки актина, так что взаимодействие актино-иозин не может происходить и создавать мышечное сокращение. Существуют другие белковые молекулы, связанные с нитью тропомиозина, это тропонины , которые имеют три полимера: тропонин I , тропонин T и тропонин c . [ 98 ]
F-Actin является одновременно сильным и динамичным. В отличие от других полимеров , таких как ДНК , составные элементы, связанные с ковалентными связями , мономеры актиновых филаментов собираются более слабыми связями. [ 99 ] Боковые связи с соседними мономерами разрешают эту аномалию, которая теоретически должна ослабить структуру, поскольку они могут быть нарушены тепловой перемешиванием. Кроме того, слабые связи дают преимущество, что концы филамента могут легко высвободить или включать мономеры. Это означает, что филаменты могут быть быстро реконструированы и могут изменять клеточную структуру в ответ на стимул окружающей среды. Который, наряду с биохимическим механизмом, с помощью которого он вызван, известен как «динамика сборки». [ 6 ]
Складывание
[ редактировать ]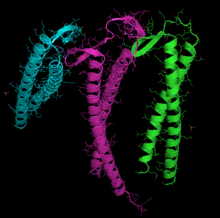
Актин может спонтанно приобретать большую часть своей третичной структуры . [ 101 ] Однако то, как он получает свою полностью функциональную форму от своей недавно синтезированной нативной формы, является особенным и почти уникальным в химии белка. Причиной этого особого маршрута может быть необходимость избежать наличия неправильно сложенных актиновых мономеров, которые могут быть токсичными, поскольку они могут действовать как неэффективные терминаторы полимеризации. Тем не менее, это является ключом к установлению стабильности цитоскелета, и, кроме того, это важный процесс для координации клеточного цикла . [ 102 ] [ 103 ]
CCT требуется, чтобы убедиться, что складывание происходит правильно. CCT является шаперонином группы II, большого белкового комплекса, который помогает в складывании других белков. CCT образуется из двойного кольца из восьми различных субъединиц (гетерооктамерных), и он отличается от шаперонинов I группы, таких как Groel , который встречается в Eubacteria и у эукариотических органелле, поскольку он не требует со-чаперона, чтобы действовать как крышка над центральной каталитической полостью. Субстраты связываются с CCT через определенные домены. Первоначально считалось, что он связан только с актином и тубулином , хотя недавние исследования иммунопреципитации показали, что он взаимодействует с большим количеством полипептидов , которые, возможно, функционируют как субстраты . Он действует через АТФ-зависимые конформационные изменения, которые иногда требуют нескольких раундов освобождения и катализа, чтобы завершить реакцию. [ 104 ]
Чтобы успешно завершить их складывание, как актин, так и тубулин должны взаимодействовать с другим белком, называемым Prefoldin молекулы , который является гетерогексамерным комплексом (образованным шестью различными субъединицами), во взаимодействии, которое настолько специфична, что [ Цитация необходима ] Полем Актиновые комплексы с префрогом, в то время как он все еще формируется, когда длиной около 145 аминокислот , в частности, на N-конце. [ 105 ]
Различные подзадушки распознавания используются для актина или тубулина, хотя есть некоторое совпадение. В актине субъединицы, которые связываются с префрофином, вероятно, являются PFD3 и PFD4, которые связываются в двух местах между остатками 60–79, а другой - между остатками 170–198. Актин распознается, загружается и доставляется в цитозольный шаперонин (CCT) в открытой конформации внутренним концом «щупальц» Prefoldin (см. Изображение и Примечание). [ 101 ] Контакт при доставке актина настолько кратко, что третичный комплекс не образуется, что сразу же освобождает дофото. [ 100 ]

Затем CCT вызывает последовательное складывание актина, образуя связи с его субъединицами, а не просто окладывая его в его полость. [ 106 ] Вот почему он обладает конкретными областями распознавания в своем апикальном β-домене. Первый этап в складывании состоит из распознавания остатков 245–249. Затем другие детерминанты устанавливают контакт. [ 107 ] И актин, и тубулин связываются с CCT в открытых конформациях в отсутствие АТФ. В случае актина две субъединицы связаны во время каждого конформационного изменения, тогда как для связывания тубулина происходит с четырьмя субъединицами. Актитин имеет специфические последовательности связывания, которые взаимодействуют с субъединицей Δ и β-CCT или с Δ-CCT и ε-CCT. После того, как AMP-PNP обязан для CCT, подложки движутся в полости шаперонина. Также кажется, что в случае актина белок CAP требуется в качестве возможного кофактора в последних состояниях складывания актина. [ 103 ]
Точный способ регулируется этот процесс, до сих пор не до конца понят, но известно, что белок PHLP3 (белок, сходное с фосдуцином ), ингибирует его активность посредством образования третичного комплекса. [ 104 ]
Каталитический механизм АТФазы
[ редактировать ]Актин является АТФазой , что означает, что это фермент , который гидролизует АТФ. Эта группа ферментов характеризуется их медленной скоростью реакции. Известно, что эта АТФаза «активна», то есть его скорость увеличивается примерно на 40 000 раз, когда актин содержит часть филамента. [ 93 ] Справочное значение для этой скорости гидролиза в идеальных условиях составляет около 0,3 с. −1 Полем Затем P I остается связанным с актином рядом с ADP в течение долгого времени, пока он не будет совместно освобожден от внутренней части нити. [ 108 ] [ 109 ]
Точные молекулярные детали каталитического механизма до сих пор не до конца понятны. Хотя в этом вопросе много споров, кажется, что для гидролиза АТФ требуется «закрытая» конформация, и считается, что остатки, которые участвуют в процессе, перемещаются на соответствующее расстояние. [ 93 ] Глютаминовая кислота Glu137 является одним из ключевых остатков, который расположен в поддомене 1. Его функция заключается в связывании молекулы воды, которая вызывает нуклеофильную атаку АТФ на γ-фосфатном связке , в то время как нуклеотид . Весьма вероятно, что конформационное изменение, вызванное вращением доменов между формами G и F, приближает Glu137, позволяя его гидролизу. Эта модель предполагает, что полимеризация и функция АТФазы будут развязаны сразу же. [ 94 ] [ 97 ] «Открытое» в «закрытое» преобразование между формами G и F и его последствиями для относительного движения нескольких ключевых остатков и образования водных проводов были охарактеризованы при молекулярной динамике и моделировании QM/MM . [ 110 ] [ 111 ]
Динамика сборки
[ редактировать ]
Актинские филаменты часто быстро собираются и разбираются, что позволяет им генерировать силу и поддерживать движение ячеек. [ 112 ] Сборка классически происходит в трех этапах. Во-первых, «фаза нуклеации», в которой от двух до трех молекул G-актина медленно соединяются, образуя небольшого олигомера, который зародит дальнейший рост. Во -вторых, «фаза удлинения», когда актиновая нить быстро растет путем добавления многих молекул актина к обоим концам. По мере роста нити, молекулы актина добавляются в (+) конец нити накаливания примерно в 10 раз быстрее, чем к конец ( -), и поэтому нити имеют тенденцию в первую очередь растут на конце (+). [ 113 ] В-третьих, «стационарная фаза», где равноправное равноправное, когда молекулы актина соединяются и оставляют нити с той же скоростью, поддерживая длину филамента. [ 112 ] В то время как длина нити остается постоянной в стационарной фазе, новые молекулы постоянно добавляются к (+) конец и падают с (-) конца, явление, называемое «беговой межсвязкой», как может быть, молекула актина, по-видимому, движется в прядь. [ 114 ] В изоляции, будет ли нить расти или сжиматься, и как быстро определяются концентрацией G-актина вокруг нити; [ 113 ] Однако в клетках динамика актиновых филаментов сильно влияет различные актин-связывающие белки .
Актин -связывающие белки
[ редактировать ]Актиновый цитоскелет in vivo не состоит исключительно из актина, другие белки необходимы для его образования, продолжения и функции. Эти белки называются актин-связывающими белками и участвуют в полимеризации, деполимеризации, стабильности и организации актина. [ 115 ] Разнообразие этих белков таково, что актин считается белком, который принимает участие в наибольшем количестве белковых взаимодействий . [ 116 ]

Зарождению новых актиновых филаментов- ограничивающей скорости стадии полимеризации актина-помогает актино-нуклеатирующие белки, такие как форманы (например, форман-2 ) и комплекс ARP2/3 . [ 118 ] Форманы помогают зародить длинные актиновые филаменты. Они связывают две свободные молекулы актина-ATP, объединяя их. Затем, когда нить начинает расти, форман движется вдоль (+) конца растущей нити, все время рекрутируя актин-связывающие белки, которые способствуют росту филаментов, и исключая белки, которые блокируют расширение нити. [ 118 ] Ветви в актиновых филаментах обычно зародываются комплексом ARP2/3 совместно с факторами, способствующими развитию нуклеации . Факторы, способствующие развитию нуклеации, связывают две свободные молекулы G-актина, затем рекрутируют и активируют комплекс ARP2/3. Комплекс активированного ARP2/3 прикрепляется к существующей актиновой нити и использует две связанные молекулы G-актина, чтобы зародить новую актиновую нить, разветвленную со старой под углом 70 °. [ 119 ]

По мере роста филаментов пул доступных молекул G-актина управляется G-актин-связывающими белками, такими как профилин и тимозин β-4 . Профилин обеспечивает поставку доступного актина-ATP путем связывания с ADP-связанным G-актином и способствуя обмену ADP на ATP. Связывание Profilin с молекулой актина физически блокирует свое дополнение к концу ( -) филамента, но позволяет ему присоединиться к конец (+). Как только актин-ATP присоединился к нити, профилин выпускает его. [ 114 ] Поскольку формины способствуют зарождению и расширению новых актиновых филаментов, они набирают профилин в область, увеличивая локальную концентрацию актина-ATP для повышения роста филаментов. [ 118 ] Напротив, тимозин β-4 связывает и секвестрирует актин-АТФ, предотвращая его соединение с микрофиламентом. [ 121 ]
Как только актиновое волокно установлено, динамика его роста или коллапса подвержена влиянию многочисленных белков. Существующие пряди могут быть прерваны белками, расщепляющими нити, такие как кофилин и гельсолин . Кофилин связывается вдоль двух молекул актина-ADP в филаменте, заставляя движение, которое дестабилизирует филаментацию и заставляет его ломаться. [ 122 ] Гельсолиновый вводит себя между молекулами актина в филаменте, нарушая нить. После разрыва нити Гельсолинь остается прикрепленным к новому (+) конец, предотвращая его растущее, тем самым вынуждая его разборку. [ 121 ]

Другие белки связываются с концами актиновых филаментов, стабилизируя их. Они называются «покрытыми белками» и включают в себя капц и тропомодулин . CAPZ связывает (+) конец нити, предотвращая дальнейшее добавление или потерю актина с этой цели. [ 121 ] Тропомодулин связывается с концом ( -) филамента, снова предотвращая добавление или потерю молекул на этой цели. Тропомодулин обычно обнаруживается в клетках, которые требуют чрезвычайно стабильных актиновых филаментов, таких как в мышцах и эритроцитах. [ 121 ]
Эти актин -связывающие белки обычно регулируются различными клеточными сигналами для контроля динамики сборки актина в различных клеточных местах. Форманы, например, обычно сложены в неактивной конформации до тех пор, пока они активируются связыванием небольшого GTPase Rho . [ 118 ] Разветвление актина в клеточной мембране важно для движения клеток, и поэтому плазматическая мембрана липид PIP 2 активирует нуклеацию, способствующее фактору, способствуя WASP и ингибирует CAPZ. [ 123 ] WASP также активируется небольшой GTPase CDC42 , в то время как еще одна волна, способствующего фактору, способствуя нуклеации, активируется GTPase RAC1 . [ 124 ]
Генетика
[ редактировать ]
Хотя большинство дрожжей имеют только один ген актина, более высокие эукариоты , в целом экспрессируют несколько изоформ актина, кодируемых семейством родственных генов. У млекопитающих есть как минимум шесть изоформ актина, закодированные отдельными генами, [ 125 ] которые разделены на три класса - альфа, бета и гамма - в соответствии с их изоэлектрическими точками . В целом, альфа-актины обнаружены в мышцах ( α-коркелет , α-аортальный гладкий , α-кардиак ), тогда как изоформы бета и гамма заметны в некмысленных клетках ( β-цитоплазматический, γ1-цитоплазматический, γ2-инвентарный гладкий) гладкий)) (β-цитоплазматический , γ1-цитоплазматический , γ2-инвентарный )) Полем Хотя аминокислотные последовательности и свойства in vitro изоформ очень похожи, эти изоформы не могут полностью заменить друг друга in vivo . [ 126 ] Растения содержат более 60 актиновых генов и псевдогенов . [ 85 ]
Типичный ген актина имеет приблизительно 100-нуклеотид 5 'UTR площадь 1200 нуклеотидов , переведенную и 200-нуко -леотид 3' UTR . Большинство генов актина прерываются интронами , и в любом из 19 хорошо охарактеризованных мест до шести интронов. Высокое сохранение семьи делает актин предпочтительной моделью для исследований, сравнивающих интронс-ра-ранние и интронские модели эволюции интрона.
Эволюция
[ редактировать ]Актин и тесно связанные белки присутствуют во всех организмах, что позволяет предположить, что общий предок всей жизни на земле имел актин. [ 127 ] Актин является одним из самых консервативных белков на протяжении всей эволюции эукариот. Последовательности актиновых белков от животных и амебы идентичны на 80%, несмотря на то, что они разделены примерно на один миллиард лет эволюции. [ 85 ] Многие одноклеточные эукариоты имеют единый ген актина, в то время как многоклеточные эукариоты часто имеют несколько тесно связанных генов, которые выполняют специализированные функции. У людей шесть; Растения имеют 10 или более. [ 127 ] В дополнение к актину, у эукариот есть большое семейство связанных с актином белков, или «ARP», которые имеют общего предка с актином и называются ARP1-ARP11, а ARP1 наиболее тесно связан с актином, а ARP11-наименьшим. [ 127 ]
Бактерии кодируют три типа актина: MREB влияет на форму клеток, деление клеток FTSA и разделение PARM крупных плазмид . [ 127 ] У некоторых археи есть ген Mreb, похожий на бактерии, в то время как у других есть ген актина, который более похож на эукариоте-актин. [ 127 ]
Эукариотический цитоскелет организмов среди всех таксономических групп имеет сходные компоненты для актина и тубулина. Например, белок, который кодируется геном ACTG2 у людей, полностью эквивалентен гомологам , присутствующим у крыс и мышей, хотя на уровне нуклеотидов сходство уменьшается до 92%. [ 128 ] Тем не менее, существуют серьезные различия с эквивалентами в прокариотах ( FTSZ и MREB ), где сходство между нуклеотидными последовательностями составляет от 40 до 50% среди различных бактерий и видов археи . Некоторые авторы предполагают, что наследственный белок, который дал модель эукариотического актина, напоминает белки, присутствующие в современных бактериальных цитоскелетах. [ 4 ] [ 129 ]

Некоторые авторы указывают, что поведение актина, тубулина и гистона , белка, участвующего в стабилизации и регуляции ДНК, сходны по своей способности связывать нуклеотиды и в их способности воспользоваться движением Браун . Также было высказано предположение, что у всех них есть общий предок. [ 130 ] Следовательно, эволюционные процессы привели к диверсификации наследственных белков в разновидности, присутствующие сегодня, сохранив, среди прочего, актины как эффективные молекулы, которые могли справиться с важными биологическими процессами предков, таких как эндоцитоз . [ 131 ]
Комплекс ARP2/3 широко встречается во всех эукариотических организмах. [ 132 ]
Эквиваленты у прокариотов
[ редактировать ]Бактериальный цитоскелет содержит белки, которые очень похожи на актиновые мономеры и полимеры. Бактериальный белок Mreb полимеризуется в тонкие негелические нити и иногда в спиральные структуры, сходные с F-актином. [ 94 ] [ 133 ] Кроме того, его кристаллическая структура очень похожа на структуру G-актина (с точки зрения его трехмерной конформации), существует даже сходство между протофиламентами MREB и F-актином. Бактериальный цитоскелет также содержит белки FTSZ , которые похожи на тубулин . [ 134 ]
Таким образом, бактерии обладают цитоскелетом с гомологичными элементами для актина (например, MREB, ALFA, PARM , FTSA и MAMK), хотя аминокислотная последовательность этих белков расходится из той, которая присутствует в клетках животных. Однако такие белки имеют высокую степень структурного сходства с эукариотическим актином. Высоко динамические микрофиламенты, образованные агрегацией MREB и PARM, необходимы для жизнеспособности клеток, и они участвуют в морфогенезе клеток, сегрегации хромосом и полярности клеток. PARM - это актиновый гомолог, который кодируется в плазмиде и участвует в регуляции плазмидной ДНК. [ 4 ] [ 135 ] Пармы из разных бактериальных плазмид могут образовывать удивительно разнообразные спиральные структуры, включающие две [ 136 ] [ 137 ] или четыре [ 138 ] пряди, чтобы поддерживать верную плазмиду наследование.
В археи гомолог TA0583 еще больше похож на эукариотические актины. [ 139 ]
Молекулярная патология
[ редактировать ]Большинство млекопитающих обладают шестью различными генами актина . Из них два кода для цитоскелета ( ACTB и ACTG1 ), в то время как остальные четыре участвуют в мышцах с полосатой скелета ( ACTA1 ), ткани гладких мышц ( ACTA2 ), кишечника мышцах ( ACTG2 ) и сердца ( ACTC1 ). Актин в цитоскелете участвует в патогенных механизмах многих инфекционных агентов , включая ВИЧ . Подавляющее большинство мутаций , которые влияют на актин, являются точечными мутациями, которые оказывают доминирующий эффект , за исключением шести мутаций, связанных с немалиновой миопатией . Это связано с тем, что во многих случаях мутант актинового мономера действует как «кепка», предотвращая удлинение F-актина. [ 88 ]
Патология, связанная с Acta1
[ редактировать ]Acta1- это ген, который кодирует α- изоформу актина, которая преобладает в мышцах с скелетными скелетами человека , хотя он также экспрессируется в сердечных мышцах и в щитовидной железе . [ 140 ] Его последовательность ДНК состоит из семи экзонов , которые производят пять известных транскриптов . [ 141 ] Большинство из них состоят из точечных мутаций, вызывающих замещение аминокислот . Мутации во многих случаях связаны с фенотипом , который определяет тяжесть и курс страданий. [ 88 ] [ 141 ]
Мутация изменяет структуру и функцию скелетных мышц, продуцирующих одну из трех форм миопатии : немалиновая миопатия типа 3 , врожденная миопатия с избытком тонких миофиламентов (см) и врожденной миопатией с диспропорцией типа волокна (CMFTD). Также были обнаружены мутации, которые дают основные миопатии . [ 143 ] Хотя их фенотипы похожи, в дополнение к типичной немалинской миопатии, некоторые специалисты различают другой тип миопатии, называемый актинической немалиновой миопатией. В первом, скопления актиновой формы вместо типичных стержней. Важно сказать, что пациент может показать более одного из этих фенотипов в биопсии . [ 144 ] Наиболее распространенные симптомы состоят из типичной морфологии лица (миопатическая фация ), мышечной слабости, задержки в моторном развитии и трудностях дыхания. Также обнаружены курс болезни, ее гравитации и возраста, в котором, по -видимому, все являются переменными и перекрывающимися формами миопатии. Симптом немалитной миопатии заключается в том, что «немалитные стержни» появляются в разных местах в мышечных волокнах 1 типа. Эти стержни представляют собой не патогномонические структуры, которые имеют сходную композицию с дисками z, обнаруженными в саркоме . [ 145 ]
Патогенез . этой миопатии очень разнообразен Многие мутации происходят в области углубления актина вблизи его сайтов связывания нуклеотидов , в то время как другие встречаются в домене 2 или в областях, где взаимодействие происходит с связанными белками. Это идет некоторым способом объяснить большое разнообразие скоплений, которые образуются в этих случаях, таких как немалитные или внутриядерные тела или тела зебры. [ 88 ] актина Изменения в складывании происходят при немалинской миопатии, а также изменениях в его агрегации, а также существуют изменения в экспрессии других связанных белков. В некоторых вариантах, где внутриядерные тела обнаруживаются, изменения в складывании маскируют сигнал экспорта белка ядра , так что накопление мутированной формы актина происходит в ядре клеток . [ 146 ] С другой стороны, кажется, что мутации к Acta1 , которые приводят к CFTDM, оказывают большее влияние на саркомерную функцию, чем на его структуру. [ 147 ] Недавние исследования попытались понять этот очевидный парадокс, который предполагает, что нет четкой корреляции между количеством стержней и мышечной слабостью. Похоже, что некоторые мутации способны вызывать большую скорость апоптоза в мышечных волокнах типа II. [ 102 ]
В гладких мышцах
[ редактировать ]Есть две изоформы, которые кодируют актины в ткани гладких мышц :
ACTG2 кодирует для крупнейшей изоформы актина, которая имеет девять экзонов , одна из которых, которая находится на 5 -дюймовом конце, не переведена . [ 128 ] Это γ-актин, который экспрессируется в кишечной гладкой мышце. Не было обнаружено никаких мутаций этого гена, которые соответствуют патологии, хотя микрочипы показали, что этот белок чаще экспрессируется в случаях, которые устойчивы к химиотерапии с использованием цисплатина . [ 148 ]
ACTA2 кодирует для α-актина, расположенного в гладких мышцах, а также в гладких мышцах сосудов. Было отмечено, что мутация MYH11 может быть ответственной по меньшей мере за 14% наследственных аневризмов торакальной аорты, особенно типа 6. Это связано с тем, что мутированный вариант вызывает неправильную нити и сниженную способность к сокращению сосудистых гладких мышц. Университет аорты была зарегистрирована у этих людей, с областями дезорганизации и гиперплазии, а также стенозом аорты Vasa Vasorum . [ 149 ] Количество страданий, в которые причастна ген, увеличивается. Это было связано с заболеванием Мойамуи , и кажется вероятным, что определенные мутации в гетерозигозе могут придать предрасположенность многим сосудистым патологиям, таким как аневризма грудной аорты и ишемическая болезнь сердца . [ 150 ] Α-актин, обнаруженный в гладких мышцах, также является интересным маркером для оценки прогресса цирроза печени . [ 151 ]
В сердечных мышцах
[ редактировать ]Ген ACTC1 кодирует изоформу α-актина, присутствующая в сердечных мышцах. Впервые он был секвенирован Хамадой и коллегами в 1982 году, когда было обнаружено, что он прерывается пятью интронами. [ 152 ] Это был первый из шести генов, где были обнаружены аллели, которые были вовлечены в патологические процессы. [ 153 ]
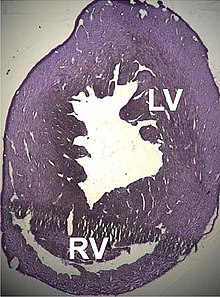
Был описан ряд структурных расстройств, связанных с точечными мутациями этого гена, которые вызывают неисправность сердца, такие как дилатационная кардиомиопатия типа 1R и гипертрофическая кардиомиопатия типа 11 . некоторые дефекты предсердной перегородки , которые также могут быть связаны с этими мутациями. Недавно были описаны [ 155 ] [ 156 ]
Были изучены два случая дилатационной кардиомиопатии с заменой высококонсервативных аминокислот, принадлежащих к белковым доменам которые связывают и впрыскивают с дисками Z. , Это привело к теории о том, что дилатация порождается дефектом в передаче сократительной силы в миоцитах . [ 157 ] [ 153 ]
Мутации в ACTC1 ответственны как минимум 5% гипертрофических кардиомиопатий. [ 158 ] Существование ряда точечных мутаций также было найдено: [ 159 ]
- Мутация E101K: Изменения чистого заряда и образование слабой электростатической связи в сайте связывания актомиозин.
- P166A: зона взаимодействия между актиновыми мономерами.
- A333P: зона взаимодействия актина-с-сед.
Патогенез, по -видимому, включает в себя компенсаторный механизм: мутированные белки действуют как токсины с доминирующим эффектом, снижая способность сердца сокращаться, вызывая аномальное механическое поведение, так что гипертрофия, которая обычно задерживается, является следствием нормального ответа мышцы сердца на стресс на стресс на стресс на стресс на стресс на стресс на стресс. Полем [ 160 ]
Недавние исследования обнаружили мутации ACTC1, которые участвуют в двух других патологических процессах: инфантильная идиопатическая ограничительная кардиомиопатия , [ 161 ] и некомпакция миокарда левого желудочка . [ 162 ]
В цитоплазматических актинах
[ редактировать ]ACTB - очень сложный локус . Существует ряд псевдогенов , которые распределены по всему геному , и его последовательность содержит шесть экзонов, которые могут привести к 21 различным транскрипциям альтернативным сплайсингом , которые известны как β-актины. В соответствии с этой сложностью, его продукты также обнаружены в ряде мест, и они составляют часть широкого спектра процессов ( цитоскелет , гистон комплекс -ацилтрансферазы Nua4, ядро клеток ), и, кроме того, они связаны с механизмами большого числа патологических процессов ( карциномы , ювенильная дистония , механизмы инфекции, пороки развития нервной системы и инвазия опухоли, среди прочего). [ 163 ] Была обнаружена новая форма актина, каппа актин, который, по-видимому, заменяет β-актин в процессах, связанных с опухолями . [ 164 ]

До сих пор были обнаружены три патологических процесса, которые вызваны прямым изменением в последовательности генов:
- Гемангиоперицитома с t (7; 12) (p22; Q13) -трансликации являются редким заболеванием, в котором трансационная мутация вызывает слияние гена ActB по сравнению с глизом в хромосоме 12 . [ 166 ]
- Ювенильная начала дистония является редким дегенеративным заболеванием , которое влияет на центральную нервную систему ; В частности, это влияет на области неокортекса и таламуса , включения , похожие на удилища эозинофильные где образуются . Пострадавшие люди представляют фенотип с деформациями на медианной линии, сенсорной потерей слуха и дистонией. Это вызвано точечной мутацией, в которой аминокислотный триптофан заменяет аргинин в положении 183. Это изменяет взаимодействие актина с системой ADF/ кофилина , которая регулирует динамику формирования цитоскелета нервных клеток . [ 167 ]
- Также была обнаружена доминирующая точечная мутация, которая вызывает дисфункцию нейтрофилов гранулоцитов и повторяющиеся инфекции . Похоже, что мутация изменяет домен, ответственный за связывание между профилином и другими регуляторными белками. Сродство Актина к профилину значительно снижена в этом аллеле. [ 168 ]
Локус ACTG1 кодирует цитозольный γ-актиновый белок , который отвечает за образование цитоскелетных микрофиламентов . Он содержит шесть экзонов , приводящих к 22 различным мРНК , которые производят четыре полных изоформе , форма экспрессии, вероятно, зависит от типа ткани, в которой они обнаружены. Он также имеет два различных промотора ДНК . [ 169 ] Было отмечено, что последовательности, переведенные из этого локуса и из-за β-актина, очень похожи на прогнозируемые, что свидетельствует об общей предков, которая перенесла дупликацию и генетическую конверсию. [ 170 ]
С точки зрения патологии, она была связана с такими процессами, как амилоидоз , пигментирование ретинита , механизмы инфекции, заболевания почек и различные типы врожденных потерь слуха. [ 169 ]
Было обнаружено, что шесть аутосомно-доминантных точечных мутаций в последовательности вызывают различные типы потери слуха, особенно сенсонирусную потерю слуха, связанную с локусом DFNA 20/26. Похоже, что они влияют на стереоцилии ресничных клеток, присутствующих в органе внутреннего уха Корти . β-актин является наиболее распространенным белком, обнаруженным в тканях человека, но он не очень распространен в ресничных клетках, что объясняет местоположение патологии. С другой стороны, представляется, что большинство этих мутаций влияют на области, связанные с связыванием с другими белками, особенно актомиозином. [ 88 ] Некоторые эксперименты показали, что патологический механизм для этого типа потери слуха связан с F-актином в мутациях, более чувствительных к кофилину, чем обычно. [ 171 ]
Однако, хотя нет никаких записей о каком-либо случае, известно, что γ-актин также экспрессируется в скелетных мышцах, и, хотя он присутствует в небольших количествах, модельные организмы показали, что его отсутствие может привести к миопатиям. [ 172 ]
Другие патологические механизмы
[ редактировать ]Некоторые инфекционные агенты используют актин, особенно цитоплазматический актин, в своем жизненном цикле . Две основные формы присутствуют в бактериях :
- Listeria monocytogenes , некоторые виды Rickettsia , Shigella Flexneri и других внутриклеточных микробов выходят из фагоцитарных вакуолей, покрывая себя капсулой актиновых филаментов. L. monocytogenes и S. flexneri генерируют хвост в форме «хвоста кометы», который дает им подвижность. Каждый вид демонстрирует небольшие различия в механизме молекулярной полимеризации их «хвостов кометы». Например, наблюдались различные скорости смещения, а Листерия и Шигелла признаны самыми быстрыми. [ 173 ] Многие эксперименты продемонстрировали этот механизм in vitro . Это указывает на то, что бактерии не используют миозиноподобный белковый двигатель, и, по-видимому, их движение получает давление, оказываемое полимеризацией, которая происходит рядом с клеточной стенкой микроорганизма. Бактерии ранее были окружены ABP от хозяина, и как минимум покрытие содержит комплекс ARP2/3 , белки ENA/VASP , кофилин, промоторы буферизации белка и нуклеации, такие как винкулина комплекс . Благодаря этим движениям они образуют выступы, которые достигают соседних клеток, а также заражая их, так что иммунная система могла бороться с инфекцией только с помощью клеточного иммунитета. Движение может быть вызвано модификацией кривой и разбором филаментов. [ 174 ] Другие виды, такие как Mycobacterium marinum и burkholderia pseudomallei , также способны локализовать полимеризацию клеточного актина, чтобы помочь их движению через механизм, который сосредоточен на комплексе ARP2/3. Кроме того, вируса вакциния против вакцины также использует элементы актинового цитоскелета для его распространения. [ 175 ]
- Pseudomonas aeruginosa способна образовывать защитную биопленку , чтобы избежать защиты организма хозяина , особенно лейкоцитов и антибиотиков . Биопленка строится с использованием ДНК и актиновых филаментов из организма хозяина. [ 176 ]
В дополнение к ранее приведенным примеру, актиновая полимеризация стимулируется на начальных этапах интернализации некоторых вирусов, особенно ВИЧ , например, инактивируя комплекс кофилина. [ 177 ]
Роль, которую актин играет в процессе инвазии раковых клеток, до сих пор не была определена. [ 178 ]
В условиях высокого липопероксидирования, было показано, что актин посттрансляционно модифицирован продуктом липопероксидирования 4-гидроксиноненал (4-HNE) [ 179 ] Полем Эта модификация предотвращает ремоделирование актинового цитоскелета, что необходимо для подвижности клеток. Кроме того, другой функциональный белок, коронин-1А, который стабилизирует филаменты F-актина, также ковалентно модифицируется 4-HNE. Эти модификации могут нарушать транс-эндотелиальную миграцию иммунных клеток или их фагоцитарную способность [ 179 ] потенциально приводит к снижению иммунного ответа при заболеваниях, характеризующихся высоким окислительным стрессом, таким как малярия, рак, метаболический синдром, атеросклероз, болезнь Альцгеймера, ревматоидный артрит, нейродегенеративные заболевания и преэклампсия. [ 180 ]
Приложения
[ редактировать ]Актин используется в научных и технологических лабораториях в качестве дорожки для молекулярных двигателей, таких как миозин (либо в мышечной ткани, либо за его пределами), и в качестве необходимого компонента для клеточного функционирования. Его также можно использовать в качестве диагностического инструмента, поскольку некоторые из его аномальных вариантов связаны с появлением специфических патологий.
- Нанотехнология . Актин-миозиновые системы действуют как молекулярные двигатели, которые позволяют транспортировать везикул и органелл по всей цитоплазме. Вполне возможно, что актин может применяться к нанотехнологиям , поскольку его динамическая способность была использована в ряде экспериментов, в том числе и проводимых в бесклеточных системах. Основная идея состоит в том, чтобы использовать микрофиламенты в качестве треков для направления молекулярных двигателей, которые могут переносить данную нагрузку. То есть актин может быть использован для определения схемы, по которой нагрузка может быть транспортирована более или менее контролируемым и направленным образом. С точки зрения общих приложений, он может быть использован для направленного транспортировки молекул для отложения в определенных местах, что позволило бы контролировать сборку наноструктур. [ 181 ] Эти атрибуты могут быть применены к лабораторным процессам, таким как на лаборатории на чипе , в механике нанокомпонентов и на нанотрансформаторах, которые преобразуют механическую энергию в электрическую энергию. [ 182 ]
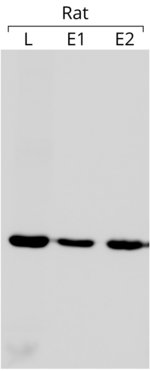
- Актин используется в качестве внутреннего контроля в вестерн -блотах, чтобы установить, что равные количества белка были загружены на каждую полосу геля. В примере блоттинга, показанном на левой стороне, 75 мкг общего белка загружали в каждую скважину. Блот реагировали с анти-β-актиновым антителом (другие детали блота см. Ссылку [ 183 ] )
Использование актина в качестве внутреннего контроля основано на предположении, что его экспрессия практически постоянно и не зависит от условий эксперимента. Сравнивая экспрессию интересующего гена с экспрессией актина, можно получить относительную величину, которую можно сравнить между различными экспериментами, [ 184 ] Всякий раз, когда выражение последнего является постоянным. Стоит отметить, что актин не всегда обладает желаемой стабильностью в экспрессии генов . [ 185 ]
- Здоровье. Некоторые аллели актина вызывают заболевания; По этой причине были разработаны методы их обнаружения. Кроме того, актин может использоваться в качестве косвенного маркера в хирургической патологии: можно использовать различия в структуре его распределения в тканях в качестве маркера инвазии при неоплазии , васкулите и других состояниях. [ 186 ] Кроме того, из -за тесной связи актина с аппаратом мышечного сокращения его уровни в скелетных мышцах уменьшается при атрофии этих тканей , поэтому его можно использовать в качестве маркера этого физиологического процесса. [ 187 ]
- Продовольственные технологии . Можно определить качество определенных обработанных продуктов, таких как колбасы , путем количественной оценки количества актина, присутствующего в составляющем мясе. Традиционно использовался метод, основанный на обнаружении 3-метилхистидина в гидролизованных образцах этих продуктов, поскольку это соединение присутствует в тяжелой цепи актина и F-миозина (оба являются основными компонентами мышц). Генерация этого соединения во плоти происходит от метилирования , остатков гистидина присутствующих в обоих белках. [ 188 ] [ 189 ]
История
[ редактировать ]
актин наблюдался экспериментально Впервые в 1887 году WD Halliburton , который извлекал белок из мышц, которые «коагулировали» препараты миозина , которые он назвал «миозином». [ 190 ] Тем не менее, Халлибертон не смог еще больше усовершенствовать свои результаты, и открытие актина вместо этого приписывается Брунору Ференку Штраубу , молодому биохимике, работающему в лаборатории Альберта Шент-Гиргей в Институте медицинской химии в Университете Зегла , Венгрия .
После обнаружения Илоны Банга и Сцент-Герги в 1941 году, что коагуляция происходит только в некоторых извлечениях миозина и была изменена при добавлении АТФ, [ 191 ] Штрауб идентифицировал и очищал актин из тех препаратов миозина, которые действительно коагулировали. Опираясь на оригинальный метод извлечения Банга, он разработал новую технику для извлечения мышечного белка, который позволил ему выделить значительные количества относительно чистого актина, опубликованного в 1942 году. [ 192 ] Метод Штрауба по сути такой же, как и в лабораториях сегодня. Поскольку белок Штрауба был необходим для активации коагуляции миозина, его дублировали актин . [ 191 ] [ 193 ] Понимая, что коагулирующие миозиновые препараты Банга также содержали актин, Szent-györgyi называется смесью обоих белков актомиозин . [ 194 ]
Военные действия Второй мировой войны означали, что Szent-Gyorgyi не смог опубликовать работу своей лаборатории в западных научных журналах . Поэтому актин стал хорошо известен на Западе только в 1945 году, когда их статья была опубликована в качестве дополнения к Scandinavica Acta Physiologica . [ 195 ] Штрауб продолжал работать над актином, и в 1950 году сообщил, что актин содержит связанный АТФ [ 196 ] и что во время гидролизуют до ADP и полимеризации белка в микрофиламенты нуклеотид неорганического фосфата связанными ( которые остаются с микрофиламентом). Штрауб предположил, что трансформация АТФ-связанного актина в ADP-связанный актин сыграла роль в мышечном сокращении. Фактически, это верно только в гладких мышцах и не поддерживалось через эксперименты до 2001 года. [ 196 ] [ 197 ]
Аминокислотное секвенирование актина было завершено М. Эльзингом и коллегами в 1973 году. [ 86 ] Кристаллическая структура G-актина была решена в 1990 году Кабшем и его коллегами. [ 89 ] В том же году Холмс и его коллеги предложили модель для F-Actin после экспериментов с использованием совместной кристаллизации с различными белками. [ 91 ] Процедура совместной кристаллизации с различными белками использовалась неоднократно в течение следующих лет, пока в 2001 году изолированный белок был кристаллизован вместе с ADP. Тем не менее, до сих пор нет рентгеновской структуры F-Actin высокого разрешения. Кристаллизация G-актина была возможна из-за использования конъюгата родамина , который препятствует полимеризации путем блокирования аминокислотной Cys-374 . [ 1 ] Кристина Ориол-Аудит умерла в том же году, когда актин был впервые кристаллизован, но она была исследователем, что в 1977 году впервые кристаллизовал актин в отсутствие актина, связывающих белки (ABP). Тем не менее, полученные кристаллы были слишком малы для доступной технологии того времени. [ 198 ]
Хотя в настоящее время в 2008 году команда Sawaya не существует, в 2008 году команда Sawaya не существует, в 2008 году команда Sawaya смогла создать более точную модель своей структуры на основе нескольких кристаллов димеров актина , которые связываются в разных местах. [ 199 ] Эта модель впоследствии была дополнительно усовершенствована Sawaya и Lorenz. Другие подходы, такие как использование криоэлектронной микроскопии и синхротроновое излучение , недавно позволили увеличить разрешение и лучшее понимание природы взаимодействий и конформационных изменений, связанных с формированием актиновых филаментов. [ 200 ] [ 94 ] [ 97 ]
Исследовать
[ редактировать ]Химические ингибиторы
[ редактировать ]
Ряд естественных токсинов , которые мешают динамике актина, широко используются в исследованиях для изучения роли актина в биологии. Латрункулин -токсин, продуцируемый губками -связывается с G-актином, предотвращая его соединение микрофиламентов. [ 201 ] Цитохалазин D - произведенный определенными грибами - служит фактором защиты, связывание с (+) конец нити и предотвращение дальнейшего добавления молекул актина. [ 201 ] Напротив, губчатый токсин ясплакинолид способствует зарождению новых актиновых филаментов путем связывания и стабильзируя пары молекул актина. [ 202 ] Phalloidin -от грибов «смертная крышка» amanita phalloides -связывается с соседними молекулами актина в нити F-актина, стабилизируя нити и предотвращая ее деполимеризацию. [ 202 ]
Фаллоидин часто помечается флуоресцентными красителями для визуализации актиновых филаментов с помощью флуоресцентной микроскопии . [ 202 ]
Смотрите также
[ редактировать ]- Ремоделирование актина - влияние на структуру и форму клеток
- Активная материя
- Филоподия
- Промежуточная нить
- Ламеллиподиум
- Моторный белок - превращает химическую энергию в механическую работу
- Нейрон
- Фаллотоксин
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый ВВП : 1J6Z ; Otterbein LR, Graceffa P, Dominguez R (июль 2001 г.). «Кристаллическая структура бессмысленного актина в состоянии ADP». Наука . 293 (5530): 708–711. doi : 10.1126/science.1059700 . PMID 11474115 . S2CID 12030018 .
- ^ Доэрти Г.Дж., МакМахон Х.Т. (2008). «Посредничество, модуляция и последствия взаимодействия мембран-цитоскелета». Ежегодный обзор биофизики . 37 (1): 65–95. doi : 10.1146/annurev.biophys.37.032807.125912 . PMID 18573073 . S2CID 17352662 .
- ^ Vindin H, Gunning P (август 2013 г.). «Цитоскелетные тропомиозины: хореографы функционального разнообразия актиновых филаментов» . Журнал исследований мышц и подвижности клеток . 34 (3–4): 261–274. doi : 10.1007/s10974-013-9355-8 . PMC 3843815 . PMID 23904035 .
- ^ Jump up to: а беременный в дюймовый Gunning PW, Ghoshdastider U, Whitaker S, Popp D, Robinson RC (Jun 2015). «Эволюция композиционно и функционально различных актиновых филаментов» . Журнал сотовой науки . 128 (11): 2009–2019. doi : 10.1242/jcs.165563 . PMID 25788699 .
- ^ Ghoshdastider U, Jiang S, Popp D, Robinson RC (июль 2015). «В поисках изначальной накаливания актина» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (30): 9150–9151. doi : 10.1073/pnas.1511568112 . PMC 4522752 . PMID 26178194 .
- ^ Jump up to: а беременный Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Глава 16: Цитоскелет» . Молекулярная биология клетки . Нью -Йорк: Гарлендская наука. С. 907–982. ISBN 978-0-8153-3218-3 .
- ^ Jump up to: а беременный в Lodish et al. 2016 , с. 778.
- ^ Хубер Ф., Шнаус Дж., Рёнике С., Раух П., Мюллер К., Фюттерер С., Кес Дж. (Январь 2013 г.). «Эффективная сложность цитоскелета: от отдельных нитей до ткани» . Достижения в области физики . 62 (1): 1–112. Bibcode : 2013 adphy..62 .... 1h . doi : 10.1080/00018732.2013.771509 . PMC 3985726 . PMID 24748680 .
- ^ Jump up to: а беременный в Граммт I (апрель 2006 г.). «Актин и миозин как факторы транскрипции». Текущее мнение в области генетики и развития . 16 (2): 191–196. doi : 10.1016/j.gde.2006.02.001 . PMID 16495046 .
- ^ Eckert R, Randall D, Burggren WW, French K (2002). Экерт Физиология животных: механизмы и адаптация . Нью -Йорк: WH Freeman и Co. Isbn 978-0-7167-3863-3 .
- ^ Jump up to: а беременный Paniagaua R, Nisal M, Sesma P, Alvarez-uría M, Album B, Anadón R, Hosé Saz F (2002). Цитология и гизология животных и животных (по -испански). Inter-American McGraw- SIN 978-84-486-0436-3 .
- ^ Сюй К, Чжун Г., Чжуан Х (январь 2013). «Актин, спектр и связанные белки образуют периодическую цитоскелетную структуру в аксонах» . Наука . 339 (6118): 452–456. Bibcode : 2013sci ... 339..452X . doi : 10.1126/science.12322251 . PMC 3815867 . PMID 23239625 .
- ^ Jump up to: а беременный Мозли Дж.Б., Гуд Б.Л. (сентябрь 2006 г.). «Дрожжевой актиновый цитоскелет: от клеточной функции до биохимического механизма» . Микробиология и молекулярная биология обзоры . 70 (3): 605–645. doi : 10.1128/mmbr.00013-06 . PMC 1594590 . PMID 16959963 .
- ^ Meagher RB, McKinney EC, Kandasamy MK (Jun 1999). «Изовариантная динамика расширяет и буферирует реакцию сложных систем: разнообразное семейство генов актина растений» . Растительная ячейка . 11 (6): 995–1006. doi : 10.1105/tpc.11.6.995 . PMC 1464670 . PMID 10368172 .
- ^ Pdb 1unc ; Vermeulen W, Vanhaesebrouck P, Van Troys M, Verschueren M, Fant F, Goethals M, Ampe C, Martins JC, Borremans FA (май 2004). «Структуры решений С-терминальных поддоменов головного ущерба человеческого виллина и администратора, оценка требований к связыванию F-актин головного костюма» . Белковая наука . 13 (5): 1276–1287. doi : 10.1110/ps.03518104 . PMC 2286768 . PMID 15096633 .
- ^ Jump up to: а беременный Higaki T, Sano T, Hasezawa S (Dec 2007). «Актиновая микрофиламентная динамика и актиновые боковые белки у растений». Современное мнение о биологии растений . 10 (6): 549–556. Bibcode : 2007copb ... 10..549h . doi : 10.1016/j.pbi.2007.08.012 . PMID 17936064 .
- ^ Kovar DR, Staiger CJ, Weaver EA, McCurdy DW (Dec 2000). «ATFIM1 - это актиновый филамент сшивающий белок из Arabidopsis thaliana» . Заводский журнал . 24 (5): 625–636. doi : 10.1046/j.1365-313x.2000.00907.x . PMID 11123801 .
- ^ Jump up to: а беременный Кларк Т.Г., Мерриам Р.В. (декабрь 1977 г.). «Диффузные и связанные ядра актина ооцитов Xenopus laevis». Клетка . 12 (4): 883–891. doi : 10.1016/0092-8674 (77) 90152-0 . PMID 563771 . S2CID 34708250 .
- ^ Hofmann WA (2009-01-01). Клеточная и молекулярная биология ядерного актина . Международный обзор клеточной и молекулярной биологии. Тол. 273. С. 219–263. doi : 10.1016/s1937-6448 (08) 01806-6 . ISBN 9780123748041 Полем PMID 19215906 .
- ^ Ulferts S, Prajapati B, Grosse R, Vartiainen Mk (февраль 2021 г.). «Новые свойства и функции актиновых и актиновых филаментов внутри ядра» . Перспективы Cold Spring Harbor в биологии . 13 (3): A040121. doi : 10.1101/cshperspect.a040121 . PMC 7919393 . PMID 33288541 .
- ^ Jump up to: а беременный Bohnsack MT, Stüven T, Kuhn C, Cordes VC, Görlich D (март 2006 г.). «Селективный блок экспорта ядерного актина стабилизирует гигантские ядра ооцитов Xenopus». Природная клеточная биология . 8 (3): 257–263. doi : 10.1038/ncb1357 . HDL : 11858/00-001M-0000-0012-E6EB-9 . PMID 16489345 . S2CID 16529470 .
- ^ Dopie J, Skarp KP, Rajakylä EK, Tanhuanpää K, Vartiainen MK (февраль 2012 г.). «Активное обслуживание ядерного актина Importin 9 поддерживает транскрипцию» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (9): E544–552. doi : 10.1073/pnas.1118880109 . PMC 3295300 . PMID 22323606 .
- ^ Wada A, Fukuda M, Mishima M, Nishida E (март 1998 г.). «Ядерный экспорт актина: новый механизм, регулирующий субклеточную локализацию основного цитоскелетного белка» . Embo Journal . 17 (6): 1635–1641. doi : 10.1093/emboj/17.6.1635 . PMC 1170511 . PMID 9501085 .
- ^ Stüven T, Hartmann E, Görlich D (ноябрь 2003 г.). «Экспортин 6: новый рецептор ядерного экспорта, специфичный для комплексов Profilin.Actin» . Embo Journal . 22 (21): 5928–5940. doi : 10.1093/emboj/cdg565 . PMC 275422 . PMID 14592989 .
- ^ Хофманн В.А., Ардуини А., Никол С.М., Камачо С.Дж., Лессард Дж.Л., Фуллер-Пейс Ф.В., Де Ланероль П (июль 2009). «Sumoylation ядерного актина» . Журнал клеточной биологии . 186 (2): 193–200. doi : 10.1083/jcb.200905016 . PMC 2717643 . PMID 19635839 .
- ^ McDonald D, Carrero G, Andrin C, De Vries G, Hendzel MJ (февраль 2006 г.). «Нуклеоплазматический бета-актин существует в динамическом равновесии между полимерными видами с низкой мобильностью и быстро диффундирующими популяциями» . Журнал клеточной биологии . 172 (4): 541–552. doi : 10.1083/jcb.200507101 . PMC 2063674 . PMID 16476775 .
- ^ Jockusch BM, Schoenenberger CA, Stetefeld J, Aebi U (август 2006 г.). «Отслеживание различных форм ядерного актина». Тенденции в клеточной биологии . 16 (8): 391–396. doi : 10.1016/j.tcb.2006.06.006 . PMID 16828286 .
- ^ Jump up to: а беременный в дюймовый Migocka-Patrzałek M, Makowiecka A, Nowak D, Mazur AJ, Hofmann WA, Malicka-Błszzkiewicz M (ноябрь 2015). «β- и γ-актины в ядре клеток меланомы A375 человека» . Гистохимия и клеточная биология . 144 (5): 417–428. doi : 10.1007/s00418-015-1349-8 . PMC 4628621 . PMID 26239425 .
- ^ Педерсон Т., Эби У (2002-12-01). «Актин в ядре: какая форма и для чего?». Журнал структурной биологии . 140 (1–3): 3–9. doi : 10.1016/s1047-8477 (02) 00528-2 . PMID 12490148 .
- ^ Chhabra D, Dos Remedios CG (сентябрь 2005 г.). «Кофилин, актин и их комплекс наблюдаются in vivo с использованием переноса энергии резонанса флуоресценции» . Биофизический журнал . 89 (3): 1902–1908. Bibcode : 2005bpj .... 89.1902c . doi : 10.1529/biophysj.105.062083 . PMC 1366693 . PMID 15994898 .
- ^ Спенсер В.А. (сентябрь 2011 г.). «Ядерный актин: ключевой игрок в общении внеклеточного матрикса-нуклеуса» . Коммуникативная и интегративная биология . 4 (5): 511–512. doi : 10.4161/cib.16256 . PMC 3204115 . PMID 22046450 .
- ^ Jump up to: а беременный Чжао К., Ван В., Рандо О.Дж., Сюэ Ю., Свейдерк К., Куо А., Крэбтри Г.Р. (ноябрь 1998). «Быстрое и фосфоинозитол-зависимое связывание комплекса BAF-подобного SWI/SNF с хроматином после передачи сигналов рецептора Т-лимфоцитов» . Клетка . 95 (5): 625–636. doi : 10.1016/s0092-8674 (00) 81633-5 . PMID 9845365 . S2CID 3184211 .
- ^ Jump up to: а беременный Hofmann WA, Stojiljkovic L, Fuchsova B, Vargas GM, Mavrommatis E, Philimonenko V, Kysela K, Goodrich JA, Lessard JL, Hope TJ, Hozak P, De Lanerolle P (ноябрь 2004 г.). «Актин является частью предварительных комплексов и необходим для транскрипции с помощью РНК-полимеразы II». Природная клеточная биология . 6 (11): 1094–1101. doi : 10.1038/ncb1182 . PMID 15502823 . S2CID 23909479 .
- ^ Jump up to: а беременный Hu P, Wu S, Hernandez N (Dec 2004). «Роль бета-актина в транскрипции РНК-полимеразы III» . Гены и развитие . 18 (24): 3010–3015. doi : 10.1101/gad.1250804 . PMC 535912 . PMID 15574586 .
- ^ Jump up to: а беременный Филимоненко В.В., Чжао Дж., Ибен С., Дингова Х., Кайсела К., Кале М., Зендграф Х., Хофманн В.А., Де Ланероль П., Хозак П., Граммт I (декабрь 2004). «Ядерный актин и миозин I необходимы для транскрипции РНК -полимеразы I». Природная клеточная биология . 6 (12): 1165–1172. doi : 10.1038/ncb1190 . PMID 15558034 . S2CID 6633625 .
- ^ Maraldi NM, Lattanzi G, Marmiroli S, Squarzoni S, Manzoli FA (2004-01-01). «Новые роли для ламинов, белков ядерной оболочки и актина в ядре». Достижения в регулировании ферментов . 44 : 155–172. doi : 10.1016/j.advenzreg.2003.11.005 . PMID 15581488 .
- ^ Summier D, Lambright A, Müller M, Jonckor V, Doll T, Vandame D, Bakkali K, Watershot D, Lemature M, Debeir O, Decasters A, Terms A, Term BA, AMPE C (ARD 2012). «Отсутствие β-актина в клетках генетически репродуктивна и условная миграционная способность» . Молекулярная и клеточная протувика . 11 (8): 255–271. doi : 10 1074/MC.M11115099 . PMC 3412960 . PMID 22448045 .
- ^ Holaska JM, Kowalski AK, Wilson KL (сентябрь 2004 г.). «Эмерин ограничивает заостренный конец актиновых филаментов: доказательства актиновой корковой сети на ядерной внутренней мембране» . PLOS Биология . 2 (9): E231. doi : 10.1371/journal.pbio.0020231 . PMC 509406 . PMID 15328537 .
- ^ Puckelwartz M, McNally EM (2011-01-01). «Эмери -девственная мышечная дистрофия». Мышечная дистрофия . Справочник по клинической неврологии. Тол. 101. С. 155–166. doi : 10.1016/b978-0-08-045031-5.00012-8 . ISBN 9780080450315 Полем PMID 21496632 .
- ^ Farrants AK (Jun 2008). «Реконструирование хроматина и организация актина» . Письма Febs . 582 (14): 2041–2050. BIBCODE : 2008 FEBSL.582.2041F . doi : 10.1016/j.febslet.2008.04.032 . PMID 18442483 . S2CID 23147656 .
- ^ Sjölinder M, Björk P, Söderberg E, Sabri N, Farrants AK, Visa N (август 2005 г.). «Растущая пре-мРНК рекрутирует актин и индивидуальные факторы, модифицирующие хроматин в транскрипционно активные гены» . Гены и развитие . 19 (16): 1871–1884. doi : 10.1101/gad.339405 . PMC 1186187 . PMID 16103215 .
- ^ Jump up to: а беременный Percipalle P, Visa N (март 2006 г.). «Молекулярные функции ядерного актина в транскрипции» . Журнал клеточной биологии . 172 (7): 967–971. doi : 10.1083/jcb.200512083 . PMC 2063754 . PMID 16549500 .
- ^ Fedorovo E, Zink D (новый 2008). "Неясная архитектура и регулирование " Клеточные исследования 1783 (11): 2174–2 два 10.1016/j.bbamcr.2008.07.018: 18718493PMID
- ^ Skarp KP, Vartiainen MK (август 2010 г.). «Актин на ДНК-древних и динамичных отношениях» . Цитоскелет . 67 (8): 487–495. doi : 10.1002/cm.20464 . PMID 20593452 . S2CID 37763449 .
- ^ Olave IA, Reck-Peterson SL, Crabtree GR (2002-01-01). «Ядерный актин и связанные с актином белки при ремоделировании хроматина». Ежегодный обзор биохимии . 71 : 755–781. doi : 10.1146/annurev.biochem.71.110601.135507 . PMID 12045110 .
- ^ Zheng B, Han M, Bernier M, Wen JK (май 2009 г.). «Ядерный актин и актин-связывающие белки в регуляции транскрипции и экспрессии генов» . Журнал FEBS . 276 (10): 2669–2685. doi : 10.1111/j.1742-4658.2009.06986.x . PMC 2978034 . PMID 19459931 .
- ^ Ferrai C, Naum-Onganía G, Longobardi E, Palazzolo M, Disanza A, Diaz VM, Crippa MP, Scita G, Blasi F (август 2009 г.). «Индукция транскрипции HOXB ретиноевой кислотой требует полимеризации актина» . Молекулярная биология клетки . 20 (15): 3543–3551. doi : 10.1091/mbc.e09-02-0114 . PMC 2719572 . PMID 19477923 .
- ^ Xu YZ, Thuraisingam T, Morais DA, Rola-Plesszczynski M, Radzioch D (Mar 2010). «Ядерная транслокация бета-актина участвует в регуляции транскрипции во время дифференцировки макрофагов клеток HL-60» . Молекулярная биология клетки . 21 (5): 811–820. doi : 10.1091/mbc.e09-06-0534 . PMC 2828967 . PMID 20053683 .
- ^ Jump up to: а беременный Miyamoto K, Pasque V, Jullien J, Gurdon JB (май 2011). «Полимеризация ядерного актина требуется для транскрипционного перепрограммирования OCT4 ооцитами» . Гены и развитие . 25 (9): 946–958. doi : 10.1101/gad.615211 . PMC 3084028 . PMID 21536734 .
- ^ Huang W, Ghisletti S, Saijo K, Gandhi M, Aouadi M, Tesz GJ, Zhang DX, Yao J, Czech MP, Goode BL, Rosenfeld MG, Glass CK (февраль 2011 г.). «Коронин 2A опосредует актин-зависимую депрессию генов воспалительного ответа» . Природа . 470 (7334): 414–418. Bibcode : 2011natur.470..414H . doi : 10.1038/nature09703 . PMC 3464905 . PMID 21331046 .
- ^ Miyamoto K, Gurdon JB (сентябрь 2011 г.). «Ядерный актин и активация транскрипции» . Коммуникативная и интегративная биология . 4 (5): 582–583. doi : 10.4161/cib.16491 . PMC 3204135 . PMID 22046469 .
- ^ Chuang CH, Carpenter AE, Fuchsova B, Johnson T, De Lanerolle P, Belmont As (Apr 2006). «Направленное движение на дальнем диапазоне межфазной хромосомной сайта» . Текущая биология . 16 (8): 825–831. Bibcode : 2006cbio ... 16..825c . doi : 10.1016/j.cub.2006.03.059 . PMID 16631592 . S2CID 1191289 .
- ^ Hofmann WA, Vargas GM, Ramchandran R, Stojiljkovic L, Goodrich JA, De Lanerolle P (ноябрь 2006 г.). «Ядерный миозин I необходим для образования первой фосфодиэфирной связи во время инициации транскрипции с помощью РНК -полимеразы II». Журнал сотовой биохимии . 99 (4): 1001–1009. doi : 10.1002/jcb.21035 . PMID 16960872 . S2CID 39237955 .
- ^ Олсон Эн, Нордхайм А (май 2010 г.). «Связание динамики актина и транскрипции генов с управлением клеточными подвижными функциями» . Природа обзор молекулярной клеточной биологии . 11 (5): 353–365. doi : 10.1038/nrm2890 . PMC 3073350 . PMID 20414257 .
- ^ Miralles F, Posern G, Zaromytidou Ai, Treisman R (май 2003). «Актин динамика контролирует активность SRF путем регуляции его коактиватора MAL». Клетка . 113 (3): 329–342. Citeseerx 10.1.1.327.7451 . doi : 10.1016/s0092-8674 (03) 00278-2 . PMID 12732141 . S2CID 17209744 .
- ^ Vartiainen MK (Jun 2008). «Динамика ядерного актина-от формы для функционирования». Письма Febs . 582 (14): 2033–2040. BIBCODE : 2008 FEBSL.582.2033V . doi : 10.1016/j.febslet.2008.04.010 . PMID 18423404 . S2CID 35474838 .
- ^ Knöll B (Jun 2010). «Актитин-опосредованная экспрессия генов в нейронах: соединение MRTF-SRF». Биологическая химия . 391 (6): 591–597. doi : 10.1515/bc.2010.061 . PMID 20370316 . S2CID 36373214 .
- ^ Цопулидис Н., Кау С., Лакета В., Кутшейдт С., Баарлинк С., Столп Б., Гросс Р., Факлер ОТ (январь 2019). «Т-клеточные рецептор, вызванные ядерной сетью ядерной сети, движутся функциями эффекторных клеток CD4+ T-клеток» . Научная иммунология . 4 (31): EAAV1987. doi : 10.1126/sciimmunol.aav1987 . PMID 30610013 .
- ^ Jump up to: а беременный Huang Y, Zhang S, Park Ji. Динамика ядерного актина в экспрессии генов, репарации ДНК и рака. Результаты Проблема ячейка отличается. 2022; 70: 625-663. doi: 10.1007/978-3-031-06573-6_23. PMID 36348125 ; PMCID: PMC9677682
- ^ Приятель, Дхиман Санкар; Bannerjee, Tatat; Лин, магазин; де Трогофф, Феликс; Борлейс, Джейн; Церковь, Пол А.; Devreetes, Peter N. (июль 2023 г.). «Приведение в действие отдельных нижестоящих узлов в сети фактора роста направляет миграцию иммунных клеток» . Развитие Cel 58 (13): 1170–1188.e7. doi : 10.1016/j.devel . ISSN 1534-5807 . PMC 1052437 . PMID 37220748 .
- ^ Лин, магазин; Приятель, Дхиман Санкар; Банерджи, Париджат; Bannerjee, Tatat; Цинь, держись; Град, Ю; Борлейс, Джейн; Церковь, Пол А.; Devreotes, Peter N. (2024-07-01). Дополнение и миграция Nature Cel Biology : 1–1 doi : 10.1038/ s4156-024-01453-4 ISSN 1476-4679 . PMID 38951708 .
- ^ Jump up to: а беременный в дюймовый Lodish et al. 2016 , с. 811–812.
- ^ Lodish et al. 2016 , с. 796.
- ^ Jump up to: а беременный в дюймовый и Lodish et al. 2016 , с. 803–805.
- ^ Lodish et al. 2016 , с. 805–806.
- ^ Jump up to: а беременный Lodish et al. 2016 , с. 807.
- ^ Fujiwara K, Porter ME, Pollard TD (октябрь 1978 г.). «Локализация альфа-актинина в борозде расщепления во время цитокинеза» . Журнал клеточной биологии . 79 (1): 268–275. doi : 10.1083/jcb.79.1.268 . PMC 2110217 . PMID 359574 .
- ^ Пелхэм Р.Дж., Чанг Ф. (сентябрь 2002). «Динамика актина в сократительном кольце во время цитокинеза в дрожжах делящихся». Природа . 419 (6902): 82–86. Bibcode : 2002natur.419 ... 82p . doi : 10.1038/nature00999 . PMID 12214236 . S2CID 4389564 .
- ^ Lodish et al. 2016 , с. 809.
- ^ Машима Т., Найто М., Ногучи К., Миллер Д.К., Николсон Д.В., Цуруо Т (март 1997 г.). «Расщепление актина CPP-32/Appopain во время развития апоптоза» . Онкоген . 14 (9): 1007–1012. doi : 10.1038/sj.onc.1200919 . PMID 9070648 .
- ^ Ван К.К. (январь 2000). «Калпайн и каспаза: Можете ли вы сказать разницу?». Тенденции в нейронауках . 23 (1): 20–26. doi : 10.1016/s0166-2236 (99) 01479-4 . PMID 10631785 . S2CID 17571984 .
- ^ Villa PG, Henzel WJ, Sensenbrenner M, Henderson CE, Pettmann B (Mar 1998). «Ингибиторы кальпаина, но не ингибиторы каспазы, предотвращают протеолиз актина и фрагментацию ДНК во время апоптоза». Журнал сотовой науки . 111 (Pt 6): 713–722. doi : 10.1242/jcs.111.6.713 . PMID 9472000 .
- ^ Huot J, Houle F, Rousseau S, Deschesnes RG, Shah GM, Landry J (ноябрь 1998). «SAPK2/P38-зависимая реорганизация F-актина регулирует раннюю мембранную блеббинг во время апоптоза, вызванного стрессом» . Журнал клеточной биологии . 143 (5): 1361–1373. doi : 10.1083/jcb.143.5.1361 . PMC 2133090 . PMID 9832563 .
- ^ Адамс К.Л., Нельсон В.Дж., Смит С.Дж. (декабрь 1996 г.). «Количественный анализ реорганизации кадгерина-катенина-актина во время развития клеточной адгезии» . Журнал клеточной биологии . 135 (6 Pt 2): 1899–1911. doi : 10.1083/jcb.135.6.1899 . PMC 2133977 . PMID 8991100 .
- ^ Witke W, Schleicher M, Noegel AA (январь 1992). «Избыточность в микрофиламентной системе: аномальное развитие клеток диктиостелия, в которых отсутствует два белка сшивания F-актина». Клетка . 68 (1): 53–62. doi : 10.1016/0092-8674 (92) 90205-q . PMID 1732064 . S2CID 37569656 .
- ^ Фернандес-Валле С., Горман Д., Гомес А.М., Банге М.Б. (январь 1997). «Актин играет роль как в изменениях в форме клеток, так и в экспрессии генов, связанных с миелинией клеток Шванна» . Журнал нейробиологии . 17 (1): 241–250. doi : 10.1523/jneurosci.17-01-00241.1997 . PMC 6793673 . PMID 8987752 .
- ^ Wolyniak MJ, Sundstrom P (октябрь 2007 г.). «Роль актиновой цитоскелетной динамики в активации циклического пути AMP и экспрессии гена HWP1 в Candida Albicans» . Эукариотическая клетка . 6 (10): 1824–1840. doi : 10.1128/ec.00188-07 . PMC 2043390 . PMID 17715368 .
- ^ Танака Х., Игути Н., Егидио де Карвалью С., Тадокоро Ю., Йомогида К., Нишимун Юн (август 2003 г.). «Новые актин-подобные белки T-актин 1 и T-актин 2 дифференциально экспрессируются в цитоплазме и ядре гаплоидных зародышевых клеток мыши» . Биология размножения . 69 (2): 475–482. doi : 10.1095/biolreprod.103.015867 . PMID 12672658 .
- ^ Цзян Ю.В., Стиллман -диджей (март 1996 г.). «Эпигенетическое влияние на транскрипцию дрожжей, вызванное мутациями в белке, связанном с актином, присутствующего в ядре» . Гены и развитие . 10 (5): 604–619. doi : 10.1101/gad.10.5.604 . PMID 8598290 .
- ^ Manor U, Kachar B (Dec 2008). «Динамическая регуляция сенсорной стереоцилии» . Семинары в биологии клеток и развития . 19 (6): 502–510. doi : 10.1016/j.semcdb.2008.07.006 . PMC 2650238 . PMID 18692583 .
- ^ Рзадзинска А.К., Шнайдер М.Е., Дэвис С., Риордан Г.П., Качар Б (март 2004 г.). «Молекулярная беговая дорожка и миозины актин поддерживают функциональную архитектуру и самообновление стереоцилии» . Журнал клеточной биологии . 164 (6): 887–897. doi : 10.1083/jcb.200310055 . PMC 2172292 . PMID 15024034 .
- ^ Сюй Дж., Ван Кеймюлен А., Вакида Н.М., Карлтон П., Бернс М.В., Борн Хр (май 2007 г.). «Полярность выявляет внутреннюю клеточную хиральность» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (22): 9296–9300. Bibcode : 2007pnas..104.9296x . doi : 10.1073/pnas.0703153104 . PMC 1890488 . PMID 17517645 .
- ^ Tamada A, Kawase S, Murakami F, Kamiguchi H (февраль 2010 г.). «Автономное вращение правого свинка роста конуса филоподий приводит к повороту нейритов» . Журнал клеточной биологии . 188 (3): 429–441. doi : 10.1083/jcb.200906043 . PMC 2819689 . PMID 20123994 .
- ^ Wan LQ, Ronaldson K, Park M, Taylor G, Zhang Y, Gimble JM, Vunjak-Novakovic G (июль 2011). «Микропаттерные клетки млекопитающих демонстрируют фенотипическую асимметрию, специфичную для фенотипа» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (30): 12295–12300. Bibcode : 2011pnas..10812295W . doi : 10.1073/pnas.1103834108 . PMC 3145729 . PMID 21709270 .
- ^ Jump up to: а беременный в дюймовый и Lodish et al. 2016 , с. 779.
- ^ Jump up to: а беременный в Elzinga M, Collins JH, Kuehl WM, Adelstein RS (сентябрь 1973 г.). «Полная аминокислотная последовательность актина кроличьей скелетной мышцы» . Труды Национальной академии наук Соединенных Штатов Америки . 70 (9): 2687–2691. Bibcode : 1973pnas ... 70.2687e . doi : 10.1073/pnas.70.9.2687 . PMC 427084 . PMID 4517681 .
- ^ Jump up to: а беременный Коллинз Дж. Х., Эльзинга М (август 1975 г.). «Первичная структура актина из кроличьей скелетной мышцы. Завершение и анализ аминокислотной последовательности» . Журнал биологической химии . 250 (15): 5915–5920. doi : 10.1016/s0021-9258 (19) 41139-3 . PMID 1150665 .
- ^ Jump up to: а беременный в дюймовый и фон глин DOS Remedios CG, Chhabra D (2008). Актитинсвязывающие белки и болезнь . Спрингер. ISBN 978-0-387-71747-0 .
- ^ Jump up to: а беременный Kabsch W, Mannherz HG, Suck D, Pai EF, Holmes KC (сентябрь 1990). «Атомная структура актина: DNASE I комплекса». Природа . 347 (6288): 37–44. Bibcode : 1990natur.347 ... 37K . doi : 10.1038/347037a0 . PMID 2395459 . S2CID 925337 .
- ^ Рулд М.А., Ван Q, Джоэл П.Б., Лоуи С., Трибус К.М. (октябрь 2006 г.). «Кристаллические структуры экспрессированных неполимеризуемых мономерного актина в состояниях ADP и ATP» . Журнал биологической химии . 281 (42): 31909–31919. doi : 10.1074/jbc.m601973200 . PMID 16920713 .
- ^ Jump up to: а беременный Holmes KC, Popp D, Gebhard W, Kabsch W (сентябрь 1990). «Атомная модель актиновой нити». Природа . 347 (6288): 44–49. Bibcode : 1990natur.347 ... 44H . doi : 10.1038/347044A0 . PMID 2395461 . S2CID 4317981 .
- ^ Jump up to: а беременный в Lodish et al. 2016 , с. 780.
- ^ Jump up to: а беременный в Рейслер Э., Эгельман Э.Х. (декабрь 2007 г.). «Структура и функция актина: то, чего мы до сих пор не понимаем» . Журнал биологической химии . 282 (50): 36133–36137. doi : 10.1074/jbc.r700030200 . PMID 17965017 .
- ^ Jump up to: а беременный в дюймовый Oda T, Iwasa M, Aihara T, Maéda Y, Narita A (январь 2009 г.). «Природа глобального перехода к фиброзному актину». Природа . 457 (7228): 441–445. Bibcode : 2009natur.457..441o . doi : 10.1038/nature07685 . PMID 19158791 . S2CID 4317892 .
- ^ Бегг Да, Родевальд Р., Ребхун Ли (декабрь 1978 г.). «Визуализация полярности актиновых филаментов в тонких участках. Доказательства равномерной полярности нити, ассоциированных с мембраной» . Журнал клеточной биологии . 79 (3): 846–852. doi : 10.1083/jcb.79.3.846 . PMC 2110270 . PMID 569662 .
- ^ Geneser F (1981). Гистология . Мунксгаард. П. 978-87-16-08418-7 .
- ^ Jump up to: а беременный в Von der Ecken J, Müller M, Lehman W, Manstein DJ, Penczek PA, Raunser S (май 2015). «Структура комплекса F-Actin-Tropomyosin» . Природа . 519 (7541): 114–117. Bibcode : 2015natur.519..114V . doi : 10.1038/nature14033 . PMC 4477711 . PMID 25470062 .
- ^ Hall Je, Guyton AC (2006). Учебник медицинской физиологии . Сент -Луис, Миссури: Elsevier Saunders. п. 76 ISBN 978-0-7216-0240-0 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К, Уолтер П., ред. (2002). «Самосборка и динамическая структура цитоскелетных нитей» . Молекулярная биология клетки (4 -е изд.). Гарлендская наука. ISBN 978-0-8153-3218-3 .
- ^ Jump up to: а беременный Simons CT, Staes A, Rommelaere H, Ampe C, Lewis SA, Cowan NJ (февраль 2004 г.). «Селективный вклад эукариотических преподжин -субъединиц в связывание актина и тубулина» . Журнал биологической химии . 279 (6): 4196–4203. doi : 10.1074/jbc.m306053200 . PMID 14634002 .
- ^ Jump up to: а беременный JL, Simons Ct. Полем Embo Journal 21 (23): 6377–6386. doi : 10.1093/ cboj/ cdf640 PMC 136944 . PMID 124566645 .
- ^ Jump up to: а беременный Vandamme D, Lambert E, Waterschot D, Cognard C, Vandakerchove J, Ampe C, Constantin B, Rommelaere H (июль 2009). «Актин-энемалиновая миопатия мутантов энемалиновой миопатии вызывают гибель клеток в культивируемых мышечных клетках» (PDF) . Biochemica et Biophysica Acta (BBA) - исследования молекулярных клеток . 1793 (7): 1259–1271. Doi : 10.1016/j.bbamcr.2009.04.004 . PMID 19393268 .
- ^ Jump up to: а беременный Брэкли Ки, Грантем Дж (январь 2009 г.). «Активность каперонина, содержащего TCP-1 (CCT): последствия для прогрессирования клеточного цикла и организации цитоскелета» . Клеточный стресс и шапероны . 14 (1): 23–31. doi : 10.1007/s12192-008-0057-x . PMC 2673901 . PMID 18595008 .
- ^ Jump up to: а беременный Стирлинг П.К., Куэллар Дж., Альфаро Г.А., Эль Хадали Ф., Бех К.Т., Вальпеста Дж.М., Мелки Р., Леру М.Р. (март 2006 г.). «PHLP3 модулирует CCT-опосредованную актин и складывание тубулина через тройные комплексы с субстратами» . Журнал биологической химии . 281 (11): 7012–7021. doi : 10.1074/jbc.m513235200 . PMID 16415341 .
- ^ Хансен В.Дж., Коуэн Н.Дж., Уэлч В.Дж. (апрель 1999 г.). «Прекрасфотолочные цепные комплексы в складывании цитоскелетных белков» . Журнал клеточной биологии . 145 (2): 265–277. doi : 10.1083/jcb.145.2.265 . PMC 2133115 . PMID 10209023 .
- ^ Martín-Benito J, Grantham J, Boskovic J, Brackley Ki, Carrascosa JL, Willison KR, Valpuesta JM (март 2007 г.). «Международное расположение цитозольного CCT CAPERONIN» . Embo сообщает . 8 (3): 252–257. doi : 10.1038/sj.embor.7400894 . PMC 1808031 . PMID 17304242 .
- ^ Neirynck K, Waterschoot D, Vandekerckhove J, Ampe C, Rommelaere H (январь 2006 г.). «Актин взаимодействует с CCT через дискретные сайты связывания: модель спуска перехода связывания для складывания актина, опосредованного CCT». Журнал молекулярной биологии . 355 (1): 124–138. doi : 10.1016/j.jmb.2005.10.051 . PMID 16300788 .
- ^ Vavylonis D, Yang Q, O'Shaughnessy B (Jun 2005). «Кинетика актина полимеризация, структура CAP и колебания» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (24): 8543–8548. arxiv : Q-bio/0404004 . Bibcode : 2005pnas..102.8543v . doi : 10.1073/pnas.0501435102 . PMC 1150824 . PMID 15939882 .
- ^ Каткар Х.Х., Давтин А., Дурумерный А.Е., Хоки Г.М., Шрамм А., Энрике М., Вот Г.А. (сентябрь 2018 г.). «Понимание кооперативного характера гидролиза АТФ в актиновых филаментах» . Биофизический журнал . 115 (8): 1589–1602. Bibcode : 2018bpj ... 115.1589K . doi : 10.1016/j.bpj.2018.08.034 . PMC 6260209 . PMID 30249402 .
- ^ McCullagh M, Saunders MG, Voth GA (сентябрь 2014 г.). «Раскрытие тайны гидролиза АТФ в актиновых филаментах» . Журнал Американского химического общества . 136 (37): 13053–13058. doi : 10.1021/ja507169f . PMC 4183606 . PMID 25181471 .
- ^ Saunders MG, Voth GA (октябрь 2011 г.). «Молекулы воды в нуклеотидном связывании расщелины актина: влияние на конформацию субъединиц и последствия для гидролиза АТФ». Журнал молекулярной биологии . 413 (1): 279–291. doi : 10.1016/j.jmb.2011.07.068 . PMID 21856312 .
- ^ Jump up to: а беременный Lodish et al. 2016 , с. 781.
- ^ Jump up to: а беременный Lodish et al. 2016 , с. 783.
- ^ Jump up to: а беременный Lodish et al. 2016 , с. 784.
- ^ Клеточная биология (на испанском). Elsevier Испания. 2002. с. 132. ISBN 978-84-458-1105-4 .
- ^ Домингес Р. (ноябрь 2004 г.). «Актитинсвязывающие белки-объединяющая гипотеза». Тенденции в биохимических науках . 29 (11): 572–578. doi : 10.1016/j.tibs.2004.09.004 . PMID 15501675 .
- ^ Робинсон Р.К., Турбедский К., Кайзер Д.А., Маршанд Дж. Б., Хиггс Х.Н., Чо С., Поллард Т.Д. (ноябрь 2001 г.). «Кристаллическая структура комплекса ARP2/3». Наука . 294 (5547): 1679–1684. Bibcode : 2001sci ... 294.1679R . doi : 10.1126/science.1066333 . PMID 11721045 . S2CID 18088124 .
- ^ Jump up to: а беременный в дюймовый Lodish et al. 2016 , с. 786.
- ^ Lodish et al. 2016 , с. 787–788.
- ^ ВВП : 2BTF ; Schutt CE, Myslik JC, Rozycki MD, Goonesekere NC, Lindberg U (октябрь 1993 г.). «Структура кристаллического профилин-бета-актина». Природа . 365 (6449): 810–816. Bibcode : 1993natur.365..810s . doi : 10.1038/365810A0 . PMID 8413665 . S2CID 4359724 .
- ^ Jump up to: а беременный в дюймовый Lodish et al. 2016 , с. 785.
- ^ Ladish et al. 2016 , стр. 784–785.
- ^ Ladish et al. 2016 , стр. 785, 788.
- ^ Ladish et al. 2016 , стр. 788–789.
- ^ Vandekerckhove J, Weber K (декабрь 1978 г.). «По меньшей мере шесть различных актинов экспрессируются в более высоком млекопитающем: анализ, основанный на аминокислотной последовательности аминоконцевого триптического пептика». Журнал молекулярной биологии . 126 (4): 783–802. doi : 10.1016/0022-2836 (78) 90020-7 . PMID 745245 .
- ^ Хейтлина С.А. (2001). Функциональная специфичность изоформ актина . Международный обзор цитологии. Тол. 202. С. 35–98. doi : 10.1016/s0074-7696 (01) 02003-4 . ISBN 9780123646064 Полем PMID 11061563 .
- ^ Jump up to: а беременный в дюймовый и Поллард 2016 , «Гены, сохранение последовательности, распределение и изобилие».
- ^ Jump up to: а беременный Мива Т., Манабе Ю., Курокава К., Камада С., Канда Н., Брунс Г., Уеяма Х, Какунага Т (июнь 1991 г.). «Структура, расположение хромосомы и экспрессия гена гамма-актина человека (энтеровой тип) человека: эволюция шести человеческих генов актина» . Молекулярная и клеточная биология . 11 (6): 3296–3306. doi : 10.1128/mcb.11.6.3296 . PMC 360182 . PMID 1710027 .
- ^ Эриксон HP (июль 2007). «Эволюция цитоскелета» . Биологии . 29 (7): 668–677. doi : 10.1002/bies.20601 . PMC 2630885 . PMID 17563102 .
- ^ Гардинер Дж., МакГи П., Общий Р., Марк Дж. (2008). «Гистоны, тубулин и актин, полученные из общего наследственного белка?». Протоплазма . 233 (1–2): 1–5. doi : 10.1007/s00709-008-0305-z . PMID 18615236 . S2CID 21765920 .
- ^ Галлетта BJ, Купер JA (февраль 2009 г.). «Актин и эндоцитоз: механизмы и филогения» . Современное мнение в клеточной биологии . 21 (1): 20–27. doi : 10.1016/j.ceb.2009.01.006 . PMC 2670849 . PMID 19186047 .
- ^ Маллинс Р.Д., Поллард Т.Д. (апрель 1999 г.). «Структура и функция комплекса ARP2/3». Современное мнение в структурной биологии . 9 (2): 244–249. doi : 10.1016/s0959-440x (99) 80034-7 . PMID 10322212 .
- ^ Попп Д., Нарита А., Маэда К., Фудзисава Т., Гошдастидер У, Иваса М., Маэда Ю., Робинсон Р.К. (май 2010). «Структура нити, организация и динамика в листах Mreb» . Журнал биологической химии . 285 (21): 15858–15865. doi : 10.1074/jbc.m109.095901 . PMC 2871453 . PMID 20223832 .
- ^ Van Den Ent F, Amos LA, Löwe J (сентябрь 2001 г.). «Прокариотическое происхождение актинового цитоскелета». Природа . 413 (6851): 39–44. Bibcode : 2001natur.413 ... 39V . doi : 10.1038/35092500 . PMID 11544518 . S2CID 4427828 .
- ^ Carballido-López R (декабрь 2006 г.). «Бактериальный актин-подобный цитоскелет» . Микробиология и молекулярная биология обзоры . 70 (4): 888–909. doi : 10.1128/mmbr.00014-06 . PMC 1698507 . PMID 17158703 .
- ^ Попп Д., Сюй В., Нарита А., Брзоска А.Дж., Скюррей Р.А., Ферт Н., Гошдастидер У., Гошдастидер У, Маеда Ю., Робинсон Р.К., Шумахер М.А. (март 2010 г.). «Структура и динамика филаментов актиноподобного белка PARM PSK41: последствия для сегрегации плазмидной ДНК» . Журнал биологической химии . 285 (13): 10130–10140. doi : 10.1074/jbc.m109.071613 . PMC 2843175 . PMID 20106979 .
- ^ Попп Д., Нарита А., Гошдастидер У, Маэда К., Маэда Й., Ода Т., Фудзисава Т., Одноши Х, Ито К., Робинсон Р.К. (апрель 2010). «Полимерные структуры и динамические свойства бактериального актина альфа». Журнал молекулярной биологии . 397 (4): 1031–1041. doi : 10.1016/j.jmb.2010.02.010 . PMID 20156449 .
- ^ Попп Д., Нарита А., Ли Л.Дж., Гошдастидер У, Сюэ Б., Сринивасан Р., Баласубраманян М.К., Танака Т., Робинсон Р.К. (июнь 2012 г.). «Новая актиноподобная структура нити от Clostridium tetani» . Журнал биологической химии . 287 (25): 21121–21129. doi : 10.1074/jbc.m112.341016 . PMC 3375535 . PMID 22514279 .
- ^ Хара Ф., Ясухиро К., Немото Н., Охта Ю., Ясанага Т., поцелуй А., Ямагиш А (март 2007 г.). Acido гомолог Журнал бактерологии 189 (5): 2039–2045. doi : 10.1128/ jb.01444-0 PMC 1855749 . PMID 17189356 .
- ^ Su Ai, Wiltshire T, Batalov S, Lapp H, Ching Ka, Block D, Zhang J, Soden R, Hayakawa M, Kreiman G, Cooke MP, Walker JR, Hogenesch JB (апрель 2004 г.). «Атлас генов транскриптомов кодирования белка мыши и человека» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (16): 6062–6067. Bibcode : 2004pnas..101.6062S . doi : 10.1073/pnas.0400782101 . PMC 395923 . PMID 15075390 .
- ^ Jump up to: а беременный "Acts_human" . P68133 . Консорциум Uniprot. Архивировано из оригинала 2012-11-05 . Получено 2013-01-21 .
- ^ Jump up to: а беременный Бата FS, Rommelaere H, Machesky LM (2007). «Фенотипы связанных с миопатией актин-мутантов в дифференцированных миотрубках C2C12» . BMC клеточная биология . 8 (1): 2. doi : 10.1186/1471-2121-8-2 . PMC 1779783 . PMID 17227580 .
- ^ Кайндл А.М., Рюшендорф Ф., Краузе С., Гебель Х.Х., Келер К., Беккер С., Понграц Д., Мюллер-Хокер Дж., Нюрнберг П., Штолтенбург-Дидингер Г., Лохмюллер Х, Хьюбнер А (ноябрь 2004 г.). «Миссенс -мутации Acta1 вызывают доминирующую врожденную миопатию с ядрами» . Журнал медицинской генетики . 41 (11): 842–848. Doi : 10.1136/jmg.2004.020271 . PMC 1735626 . PMID 15520409 .
- ^ Sparrow JC, Nowak KJ, Durling HJ, Beggs AH, Wallgren-Pettersson C, Romero N, Nonaka I, Laing NG (сентябрь 2003 г.). «Мышечная болезнь, вызванная мутациями в гене альфа-актина скелетных мышц (ACTA1)». Нервно -мышечные расстройства . 13 (7–8): 519–531. doi : 10.1016/s0960-8966 (03) 00101-9 . PMID 12921789 . S2CID 20716 .
- ^ North K, Ryan MM (2002). «Немалийная миопатия» . В Pagon RA, Bird TD, Dolan CR, Stephens K, Adam Mp (Eds.). GenereViews [Интернет] . Сиэтл (Вашингтон): Университет Вашингтона, Сиэтл. PMID 20301465 . Архивировано из оригинала 2017-01-18.
- ^ Илковский Б., Новак К.Дж., Домазетовский А. А., Максвелл А.Л., Климент С., Дэвис К.Е., Лейнг Нг, Северный Кн, Купер -стрит (август 2004 г.). «Свидетельство о доминантно-негативном эффекте при немалиновой миопатии ACTA1, вызванных аномальным складыванием, агрегацией и измененной полимеризацией изоформ мутантного актина» . Молекулярная генетика человека . 13 (16): 1727–1743. doi : 10.1093/hmg/ddh185 . PMID 15198992 .
- ^ Кларк Н.Ф., Илкавски Б., Купер С., Валова В.А., Робинсон П.Дж., Нонака И., Фенг Дж.Дж., Марстон С., Северный К (июнь 2007 г.). «Патогенез диспропорции врожденного волокна, связанного с ACTA1». Анналы неврологии . 61 (6): 552–561. doi : 10.1002/ana.21112 . PMID 17387733 . S2CID 11746835 .
- ^ Уотсон М.Б., Линд М.Дж., Смит Л., Дрю П.Дж., Коуквелл Л. (2007). «Анализ микрочипов экспрессии выявляет гены, связанные с устойчивостью in vitro к цисплатину в модели клеточной линии» . Acta oncologica . 46 (5): 651–658. doi : 10.1080/02841860601156157 . PMID 17562441 . S2CID 7163200 .
- ^ Guo DC, Pannu H, Tran-Fadulu V, Papke CL, Yu RK, Avidan N, Bourgeois S, Estrera AL, Safi HJ, Sparks E, Amor D, Ades L, McConnell V, Willoughby CE, Abuelo D, Willing M, Льюис Р.А., Ким Д.Х., Шерер С., Тунг П.П., Ан С., Буха Л.М., Раман С.С., Шете С.С., Милевич Д.М. (декабрь 2007 г.). «Мутации в альфа-актине гладких мышц (ACTA2) приводят к аневризмам и рассечениям в грудной аорте». Природа генетика . 39 (12): 1488–1493. doi : 10.1038/ng.2007.6 . PMID 17994018 . S2CID 62785801 .
- ^ Guo DC, Papke CL, Tran-Fadulu V, Regalado ES, Avidan N, Johnson RJ, Kim DH, Pannu H, Willing MC, Sparks E, Pyeritz RE, Singh MN, Dalman RL, Grotta JC, Marian AJ, Boerwinkle EA, Frazier LQ, Lemaire SA, Coselli JS, Estrera AL, Safi HJ, Veeraraghavan S, Muzny DM, Wheeler DA, Willerson JT, Yu RK, Shete SS, Scherer SE, Raman CS, Buja LM, Milewicz DM (май 2009 г.). «Мутации в альфа-актине гладких мышц (ACTA2) вызывают заболевание коронарной артерии, инсульт и болезнь Мойамуи, наряду с болезнью торакальной аорты» . Американский журнал человеческой генетики . 84 (5): 617–627. doi : 10.1016/j.ajhg.2009.04.007 . PMC 2680995 . PMID 19409525 .
- ^ Akpolat N, Yahsi S, Godekmerdan A, Yalniz M, Demirbag K (сентябрь 2005 г.). «Значение альфа-SMA в оценке тяжести фиброза печени при инфекции гепатита В и развития цирроза: гистопатологическое и иммуногистохимическое исследование». Гистопатология . 47 (3): 276–280. doi : 10.1111/j.1365-2559.2005.02226.x . PMID 16115228 . S2CID 23800095 .
- ^ Хамада Х, Петрино М.Г., Какунага Т (октябрь 1982). «Молекулярная структура и эволюционное происхождение гена актина сердца человека» . Труды Национальной академии наук Соединенных Штатов Америки . 79 (19): 5901–5905. Bibcode : 1982pnas ... 79.5901H . doi : 10.1073/pnas.79.19.5901 . PMC 347018 . PMID 6310553 .
- ^ Jump up to: а беременный Olson TM, Michels VV, Thibodeau SN, Tai YS, Keating Mt (май 1998). «Актиновые мутации в дилатационной кардиомиопатии, наследственная форма сердечной недостаточности». Наука . 280 (5364): 750–752. Bibcode : 1998sci ... 280..750o . doi : 10.1126/science.280.5364.750 . PMID 9563954 . S2CID 26971894 .
- ^ Xia XG, Zhou H, Samper E, Melov S, Xu Z (январь 2006 г.). «Pol II-экспрессированная ShRNA сбивает экспрессию гена SOD2 и вызывает фенотипы нокаута гена у мышей» . PLOS Genetics . 2 (1): E10. doi : 10.1371/journal.pgen.0020010 . PMC 1358942 . PMID 16450009 .
- ^ Онлайн -наследство Менделян в Человеке (Омим): актин, альфа, сердечная мышца; ACTC1 - 102540
- ^ Matsson H, Eason J, Bookwalter CS, Klar J, Gustavsson P, Sunnegårdh J, Enell H, Jonzon A, Vikkula M, Gutierrez I, Granados-Riveron J, Pope M, Bu'lock F, Cox J, Robinson Te, Song Song. F, Brook DJ, Marston S, Trybus KM, Dahl N (январь 2008 г.). «Альфа-кардиакс-актин-мутации вызывают дефекты перегородки предсердий» . Молекулярная генетика человека . 17 (2): 256–265. doi : 10.1093/hmg/ddm302 . PMID 17947298 .
- ^ Devlin TM (2006). Биохимия Барселона: Реверс. ISBN 978-84-291-7208-9 .
- ^ Кабаева З. (11 ноября 2002 г.). Генетический анализ в гипертрофической кардиомиопатии (тезис). doi : 10.18452/14800 .
- ^ Olson TM, Doan TP, Kishimoto NY, Whitby FG, Ackerman MJ, Fananapazir L (сентябрь 2000). «Унаследованные и мутации de novo в гене сердца актина вызывают гипертрофическую кардиомиопатию». Журнал молекулярной и клеточной кардиологии . 32 (9): 1687–1694. doi : 10.1006/jmcc.2000.1204 . PMID 10966831 .
- ^ Ramírez CD, Padrón R (2004). «Семейная гипертрофическая кардиомиопатия: гены, мутации и модели на животных. Обзор» [Гипертрофическая кардиомиопатия: гены, мутации и модели на животных. для просмотра]. Клинические исследования (на испанском). 45 (1): 69–100.
- ^ Каски Дж.П., Сиррис П., Берч М., Томе-Эстебан М.Т., Фентон М., Кристиансен М., Андерсен П.С., Себир Н., Эшворт М., Динфилд Дж., МакКенна В.Дж., Эллиотт Прем (ноябрь 2008). «Идиопатическая ограничительная кардиомиопатия у детей вызвана мутациями в генах белка сердечной саркоры». Сердце . 94 (11): 1478–1484. doi : 10.1136/hrt.2007.134684 . PMID 18467357 . S2CID 44257334 .
- ^ Пиготт Т.Дж., Джефферсон Д. (1991). «Идиопатический паралич из пернеального нерва-обзор тринадцати случаев». Британский журнал нейрохирургии . 5 (1): 7–11. doi : 10.3109/02688699108998440 . PMID 1850600 .
- ^ "Джин: ACTB" . Aceview . Национальный центр биотехнологии США (NCBI). Архивировано из оригинала 2013-06-18 . Получено 2013-01-21 .
- ^ Чанг К.В., Ян П.Ю., Лай Хи, Йех Т.С., Чен Т.С., Йе Кт (сентябрь 2006 г.). «Идентификация новой изоформы актина при гепатоцеллюлярной карциноме». Гепатологические исследования . 36 (1): 33–39. doi : 10.1016/j.hepres.2006.05.003 . PMID 16824795 .
- ^ Williams KL, Rahimtula M, Mearow KM (2005). «HSP27 и рост аксонов в сенсорных нейронах взрослых in vitro» . BMC Neuroscience . 6 (1): 24. doi : 10.1186/1471-2202-6-24 . PMC 1087488 . PMID 15819993 .
- ^ «Опухоли мягких тканей: перицитома с t (7; 12)» . Атлас генетики и цитогенетики в онкологии и гематологии . Университетская больница Пуайера. Архивировано с оригинала в 2008-12-30 . Получено 2013-01-21 .
- ^ Procaccio V, Salazar G, Ono S, Styers ML, Gearing M, Davila A, Jimenez R, Juncos J, Gutekunst CA, Meroni G, Fontanella B, Sontag E, Sontag JM, Faundez V, Wainer BH (Jun 2006). «Мутация бета -актина, которая изменяет динамику деполимеризации, связана с аутосомно -доминантными пороками развития, глухотой и дистонией» . Американский журнал человеческой генетики . 78 (6): 947–960. doi : 10.1086/504271 . PMC 1474101 . PMID 16685646 .
- ^ Nunoi H, Yamazaki T, Tsuchiya H, Kato S, Malech HL, Matsuda I, Kanegasaki S (июль 1999). «Гетерозиготная мутация бета-актина, связанная с дисфункцией нейтрофилов и рецидивирующей инфекцией» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (15): 8693–8698. Bibcode : 1999pnas ... 96.8693n . doi : 10.1073/pnas.96.15.8693 . PMC 17578 . PMID 10411937 .
- ^ Jump up to: а беременный "Джин: Actg1" . Aceview . Национальный центр биотехнологии США (NCBI). Архивировано из оригинала 2013-06-18 . Получено 2013-01-21 .
- ^ Erba HP, Gunning P, Kedes L (июль 1986). «Нуклеотидная последовательность мРНК с цитоскелетном актином человека человеком: аномальная эволюция генов нешебочных актина позвоночных» . Исследование нуклеиновых кислот . 14 (13): 5275–5294. doi : 10.1093/nar/14.13.5275 . PMC 311540 . PMID 3737401 .
- ^ Брайан К, Рубенштейн П.А. (июль 2009). «Аллеле-специфические эффекты мутаций гамма-актина человеческого глухоты (DFNA20/26) на взаимодействие актина/кофилина» . Журнал биологической химии . 284 (27): 18260–18269. doi : 10.1074/jbc.m109.015818 . PMC 2709362 . PMID 19419963 .
- ^ Sonnemann KJ, Fitzsimons DP, Patel JR, Liu Y, Schneider MF, Moss RL, Ervasti JM (сентябрь 2006 г.). «Цитоплазматический гамма-актин не требуется для развития скелетных мышц, но его отсутствие приводит к прогрессирующей миопатии» . Ячейка развития . 11 (3): 387–397. doi : 10.1016/j.devcel.2006.07.001 . PMID 16950128 .
- ^ Gouin E, Gantelet H, Egile C, Lasa I, Ohayon H, Villiers V, et al. (Июнь 1999 г.). «Сравнительное исследование основанных на актиновых подмогательствах патогенных бактерий Listeria Monocytogenes, Shigella Flexneri и Rickettsia conorii». Журнал сотовой науки . 112 (11): 1697–1708. doi : 10.1242/jcs.112.11.1697 . PMID 10318762 .
- ^ Lambrechts A, Gevaert K, Cossart P, Vandekerckhove J, Van Troys M (май 2008 г.). «Листерия Комета Хвост: Актиновая мощность на работе». Тенденции в клеточной биологии . 18 (5): 220–227. doi : 10.1016/j.tcb.2008.03.001 . PMID 18396046 .
- ^ Gouin E, Welch MD, Cossart P (февраль 2005 г.). «Актин мощность внутриклеточных патогенов». Текущее мнение о микробиологии . 8 (1): 35–45. doi : 10.1016/j.mib.2004.12.013 . PMID 15694855 .
- ^ Паркс QM, Young RL, Poch KR, Malcolm KC, Vasil ML, Nick Ja (апрель 2009 г.). «Повышение нейтрофилов Pseudomonas Aeruginosa BioFilm Development: F-актин и ДНК человека в качестве мишеней для терапии» . Журнал медицинской микробиологии . 58 (Pt 4): 492–502. doi : 10.1099/jmm.0.005728-0 . PMC 2677169 . PMID 19273646 .
- ^ Лю Y, Белкина Н.В., Шоу С. (2009). «ВИЧ-инфекция Т-клеток: актин-в-актиновый» . Наука сигнализация . 2 (66): PE23. doi : 10.1126/scisignal.266pe23 . PMID 19366992 . S2CID 30259258 .
- ^ Machesky LM, Tang HR (Jul 2009). «На основе актина выступы: промоутеры или ингибиторы инвазии рака?» Полем Раковая клетка . 16 (1): 5–7. doi : 10.1016/j.ccr.2009.06.009 . PMID 19573806 .
- ^ Jump up to: а беременный Skorokhod OA, Barrera V, Heller R, Carta F, Turrini F, Arese P, Schwarzer E (2014). «Малярийный пигмент гемозоин нарушает хемотаксическую подвижность и трансендотелиальную миграцию моноцитов через 4-гидроксиноненал» . Свободный Радик Биол Мед . 75 : 210–21. doi : 10.1016/j.freeradbiomed.2014.07.004 . PMID 25017964 .
- ^ Kıran TR, Otlu O, Karabulut AB (2023). «Окислительный стресс и антиоксиданты в здоровье и заболеваниях» . Журнал лабораторной медицины . 47 (1): 1–1. doi : 10.1515/labmed-2022-0108 .
- ^ Hess H, Clemmens J, Qin D, Howard J, Vogel V (2001). «Молекулярные шаттлы, контролируемые светом, изготовленные из моторных белков, несущих груз на спроектированных поверхностях». Нано буквы . 1 (5): 235–239. Bibcode : 2001nanol ... 1..235H . doi : 10.1021/nl015521e .
- ^ Мэнссон А., Сандберг М., Банк Р., Балаз М., Николс И.А., Омлинг П., Тегенфельдт Д.О., Тагеруд С., Монтелиус Л. (2005). «Молекулярные двигатели на основе актина для транспортировки груза в нанотехнологиях-потенциала и проблемы». IEEE транзакции на продвинутой упаковке . 28 (4): 547–555. doi : 10.1109/tadvp.2005.858309 . S2CID 33608087 .
- ^ Шарма С., Ханукоглу I (апрель 2019 г.). «Картирование сайтов локализации эпителиального натриевого канала (ENAC) и CFTR в сегментах эпидидимиса млекопитающих». Журнал молекулярной гистологии . 50 (2): 141–154. doi : 10.1007/s10735-019-09813-3 . PMID 30659401 . S2CID 58026884 .
- ^ Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, Speleman F (Jun 2002). «Точная нормализация количественных данных ОТ-ПЦР в реальном времени путем геометрического усреднения нескольких генов внутреннего контроля» . Биология генома . 3 (7): Research0034. doi : 10.1186/gb-2002-3-7-research0034 . PMC 126239 . PMID 12184808 .
- ^ Selvey S, Thompson EW, Matthaei K, Lea Ra, Irving MG, Griffiths LR (октябрь 2001 г.). «Бета-актин-неподходящий внутренний контроль для ОТ-ПЦР» . Молекулярные и клеточные зонды . 15 (5): 307–311. doi : 10.1006/mcpr.2001.0376 . PMID 11735303 .
- ^ Мукай К, Шоллмейер СП, Росай Дж (январь 1981). «Иммуногистохимическая локализация актина: применение в хирургической патологии». Американский журнал хирургической патологии . 5 (1): 91–97. doi : 10.1097/00000478-198101000-00013 . PMID 7018275 .
- ^ Haddad F, Roy RR, Zhong H, Edgerton VR, Baldwin KM (август 2003 г.). «Ответы атрофии на неактивность мышц. II. Молекулярные маркеры дефицита белка». Журнал прикладной физиологии . 95 (2): 791–802. doi : 10.1152/japplphysiol.01113.2002 . PMID 12716877 . S2CID 8268572 .
- ^ Hocquette JF, Lehnert S, Barendse W, Cassar-Malek I, Picard B (2006). «Текущие достижения в протеомном анализе и его использование для разрешения качества мяса птицы». Всемирный птичий журнал . 62 (1): 123–130. doi : 10.1079/wps200589 . S2CID 86189373 .
- ^ Nollet L (2004). «Методы и инструменты в анализе прикладной пищи». Справочник по анализу продуктов питания . Тол. 3 (2 изд.). Нью -Йорк, Нью -Йорк: Марсель Деккер. С. 1741–2226. ISBN 978-0-8247-5039-8 .
- ^ Halliburton WD (август 1887 г.). «На мышечной плазме» . Журнал физиологии . 8 (3–4): 133–202. doi : 10.1113/jphysiol.1887.sp000252 . PMC 1485127 . PMID 16991477 .
- ^ Jump up to: а беременный Банга I (1942). Szent-györgyi a (ed.). «Подготовка и свойства миозин А и Б.» Исследования Института медицинской химии Университет Сегеда. 1941-1942 . Я : 5–15.
- ^ Штрауб BF (1942). Szent-györgyi a (ed.). "Актин" . Исследования Института медицинской химии Университет Сегеда. 1942 . II : 3–15.
- ^ Bugyi B, Kellermayer M (март 2020 г.). «Открытие актина:« Чтобы увидеть то, что видели все остальные, и думать о том, что никто не думал » » . Журнал исследований мышц и подвижности клеток . 41 (1): 3–9. doi : 10.1007/s10974-019-09515-z . PMC 7109165 . PMID 31093826 .
- ^ Szent-györgyi A (1942). Szent-györgyi a (ed.). «Обсуждение» . Исследования Института медицинской химии Университет Сегеда. 1941-1942 . Я : 67-71.
- ^ Szent-Gyorgyi A (1945). «Исследования на мышцах». Acta Physiol Scandinav . 9 (приложение): 25.
- ^ Jump up to: а беременный Straub FB, Feuer G (1989). «Аденозинетрифосфат. Функциональная группа актина. 1950». Biochimica et Biophysica Acta . 1000 : 180–195. doi : 10.1016/0006-3002 (50) 90052-7 . PMID 2673365 .
- ^ Bárány M, Barron JT, Gu L, Bárány K (декабрь 2001 г.). «Обмен связанными с актиновыми нуклеотидом в неповрежденных артериальных гладких мышцах» . Журнал биологической химии . 276 (51): 48398–48403. doi : 10.1074/jbc.m106227200 . PMID 11602582 .
- ^ Ориол С, Дюборд С, Лэндон Ф. (январь 1977 г.). «Кристаллизация нативного поперечного мышечного актина» . Письма Febs . 73 (1): 89–91. Bibcode : 1977febsl..73 ... 89o . doi : 10.1016/0014-5793 (77) 80022-7 . PMID 320040 . S2CID 5142918 .
- ^ Sawaya MR, Kudryashov DS, Pashkov I, Adisetiyo H, Reisler E, Yeates to (Apr 2008). «Многочисленные кристаллические структуры димеров актина и их последствия для взаимодействия в актиновой нити» . Acta Crystallographica Раздел d . 64 (Pt 4): 454–465. Bibcode : 2008Ccrd..64..454S . doi : 10.1107/s0907444908003351 . PMC 2631129 . PMID 18391412 .
- ^ Нарита А., Такеда С., Ямашита А., Маеда Ю (ноябрь 2006 г.). «Структурная основа актиновых филаментов в колюче: исследование криоэлектронной микроскопии» . Embo Journal . 25 (23): 5626–5633. doi : 10.1038/sj.emboj.7601395 . PMC 1679762 . PMID 17110933 .
- ^ Jump up to: а беременный Ludish et al. 2016 , стр. 791–792.
- ^ Jump up to: а беременный в Ludish et al. 2016 , стр. 792.
Работы цитируются
[ редактировать ]- Lodish HF, Berk A, Kaiser C, Krieger M, Bretscher A, Ploegh H, et al. (2016). «Организация клеток и движение I: микрофиламенты». Молекулярная клеточная биология (восьмое изд.). Нью -Йорк: WH Freeman. ISBN 978-1-4641-8339-3 .
- Поллард ТД (август 2016 г.). «Актитин и актин-связывающие белки» . Холодный весна Harb Perspect Biol . 8 (8): A018226. doi : 10.1101/cshperspect.a018226 . PMC 4968159 . PMID 26988969 .
Внешние ссылки
[ редактировать ]- Методы окрашивания актина (живое и фиксированное окрашивание клеток)
- Eukaryotic Linear Motif Resource Motif Class LIG_ACTIN_RPEL_3
- Eukaryotic Linear Motif Resource Motif Class LIG_ACTIN_WH2_1
- Eukaryotic Linear Motif Resource Motif Class LIG_ACTIN_WH2_2
- 3D макромолекулярные структуры актиновых филаментов из Банка данных EM (EMDB)