Одомашненные растения и животные Австронезии.
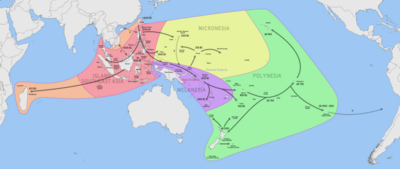
Одним из крупнейших событий миграции людей было морское заселение островов региона Индо -Тихоокеанского австронезийскими народами , которое, как полагают, началось по крайней мере с 5500 до 4000 лет назад (3500–2000 лет до нашей эры). Эти миграции сопровождались набором одомашненных, полуодомашненных и комменсальных растений и животных, транспортированных на кораблях и катамаранах с выносными опорами , что позволило ранним австронезийцам процветать на островах Приморской Юго-Восточной Азии (также известных как «Острова Юго-Восточной Азии»). например: Филиппины , Индонезия ), Ближняя Океания ( Меланезия ), Отдаленная Океания ( Микронезия и Полинезия ), Мадагаскар и Коморские острова . [2] [3]
К ним относятся сельскохозяйственные культуры и животные, которые, как полагают, произошли из культур Хемуду и Маджиабанг на гипотетических доавстронезийских родинах в материковом Китае . [4] а также другие растения и животные, которые, как полагают, были впервые одомашнены на Тайване , в Приморской Юго-Восточной Азии и Новой Гвинее . [5] [6] Некоторые из этих растений иногда также называют « растениями каноэ », особенно в контексте полинезийских миграций. [7] [8] [9] Домашние животные и растения, завезенные в исторические времена , не включены.
Растения
[ редактировать ]К одомашненным, полуодомашненным и комменсальным растениям, привезенным австронезийскими путешественниками, относятся следующие:
Aleurites moluccanus (свечник)
[ редактировать ]
Свечной орех ( Aleurites moluccanus ) был впервые одомашнен на островах Юго-Восточной Азии. Остатки собранных свечных орехов были обнаружены на археологических раскопках на Тиморе и Моротаи в восточной Индонезии и датируются примерно 13 000 и 11 000 лет назад соответственно. [10] Археологические свидетельства выращивания свечных орехов также обнаружены на неолитической стоянках культуры Тоали на юге Сулавеси, датируемые примерно 3700–2300 годами до нашей эры . [11] [12] Свечной орех был широко завезен на острова Тихого океана первыми австронезийскими путешественниками и натурализовался на высоких вулканических островах. [13] [14] [15]

Свечной орех имеет очень широкий спектр применения, и каждую часть дерева можно собирать. В первую очередь их выращивали из-за высокого содержания масла в орехов ядрах . Они широко использовались для освещения до появления других источников света, отсюда и название «свечной орех». Ядра нанизывали на сердцевины кокосовых орехов, а затем поджигали. Для горения каждого ядра требуется около трех минут, поэтому серия может действовать как факел. Эта традиция изготовления факелов из свечей существует как в Юго-Восточной Азии, так и в Океании. Масло свечного ореха, полученное из орехов, также можно использовать непосредственно в лампах. Их также можно использовать в производстве мыла, мазей и в качестве консервантов для рыболовных снастей. Другие традиционные применения включают использование древесины для изготовления небольших каноэ и резьбы по дереву; сок для лака и смол; скорлупа орехов для украшения (особенно леев ), рыболовные крючки, игрушки и производство черных красок; кора для лекарств и клетчатки; и так далее. Некоторые нетоксичные сорта также используются в качестве приправ или ингредиентов в кухнях Юго-Восточной Азии и Тихоокеанского региона. [16]
Прото -малайско-полинезийское слово, обозначающее свечной орех, реконструировано как *kamiri , с современными родственными словами , включая Hanunó'o , Iban и сунданский kamiri ; яванский и малайский кемири ; и Тетун камии . Однако считается, что океанийские слова, обозначающие «свечный орех», произошли от протоавстронезийского *CuSuR , который стал прото-малайско-полинезийским *tuhuR , что первоначально означало «нанизывать вместе, как бусины», что относится к конструкции факелов из свечных орехов. Он стал протовосточно-малайско-полинезийским и протоокеанским *tuRi, который затем дублировался . Современные родственные слова, включая фиджийский , тонганский , раротонганский и ниуэский туи-туи ; и гавайский куй-куй или кукуи . [17]
Алоказия крупноризозная (таро гигантское)
[ редактировать ]
Гигантское таро ( Alocasia macrorhizos ) первоначально было одомашнено на Филиппинах , но известно от диких экземпляров до ранних австронезийцев на Тайване . С Филиппин они распространились на остальную часть островов Юго-Восточной Азии и на восток до Океании , где стали одной из основных сельскохозяйственных культур жителей островов Тихого океана . [18] [19] Это один из четырех основных видов ароидных (тарос), культивируемых австронезийцами в первую очередь как источник крахмала , остальные — Amorphophallus paeoniifolius , Colocasia esculenta и Cyrtosperma merkusii , каждый из которых имеет несколько культивируемых разновидностей. Их листья и стебли также съедобны, если их тщательно приготовить, хотя с гигантским таро это делают редко, поскольку оно содержит большее количество рафидов , вызывающих зуд. [20] [21]
Реконструированное слово гигантского таро на протоавстронезийском языке — *biRaq , которое стало протоокеанским *piRaq . Современные родственники этого слова на островах Юго-Восточной Азии и Микронезии включают Рукай vi'a или bi'a ; Ифугао место ; Илокано , Себуано и Бикол ; Тирурай пиво ; Нгаджу плохой ; Малагасийский через ; малайский и ачехский бирах ; Монгондоу бига ; Палау бисə ; Чаморро- пига ; Бима Тиа ; Роти и Тетун Король ; Асилулу хила ; и Ковиай означает родственникам В Океании к его относятся Wuvulu и Aua pia ; Моту и Аре — хира ; Киливила и Фиджия через ; и гавайская пиа Обратите внимание, что в некоторых случаях родственные слова изменились и стали обозначать другие типы таро. [17] [18]
Amorphophallus paeoniifolius (elephant foot yam)
[edit]
The elephant foot yam (Amorphophallus paeoniifolius) is used as food in Island Southeast Asia, Mainland Southeast Asia, and South Asia. Its origin and center of domestication was formerly considered to be India, where it is most widely utilized as a food resource in recent times. But a genetic study in 2017 has shown that Indian populations of elephant foot yams have lower genetic diversity than those in Island Southeast Asia, therefore it is now believed that elephant foot yams originated from Island Southeast Asia and spread westwards into Thailand and India, resulting in three independent domestication events. From Island Southeast Asia, they were also spread even further west into Madagascar, and eastwards to coastal New Guinea and Oceania by Austronesians. Though they may have spread south into Australia without human intervention.[22][23][24]
The elephant foot yam is one of the four main species of aroids (taros) cultivated by Austronesians primarily as a source of starch, the others being Alocasia macrorrhizos, Colocasia esculenta, and Cyrtosperma merkusii, each with multiple cultivated varieties. Elephant foot yam, however, is the least important among the four and was likely only eaten as a famine crop, since it contains more raphides that cause irritation if not cooked thoroughly.[23][21][25]
Areca catechu (areca palm)
[edit]The practice of chewing areca nuts originated in Island Southeast Asia, where the areca palm is native. The oldest known evidence of areca nut chewing was found in a burial pit in the Duyong Cave site in the Philippines (to which areca palms are native), which dates to around 4,630±250 BP. Its diffusion is closely tied to the Neolithic expansion of the Austronesian peoples. It was spread to the Indo-Pacific during prehistoric times, reaching Micronesia at 3,500 to 3,000 BP, Near Oceania at 3,400 to 3,000 BP; South India and Sri Lanka by 3,500 BP; Mainland Southeast Asia by 3,000 to 2,500 BP; Northern India by 1500 BP; and Madagascar by 600 BP. From India, it was also spread westwards to Persia and the Mediterranean. It was also previously present in the Lapita culture, based on archaeological remains dated from 3,600 to 2,500 BP, but it was not carried into Polynesia.[26]
Artocarpus
[edit]Numerous species of Artocarpus are traditionally cultivated or harvested from semi-domesticated or wild populations in Island Southeast Asia and Micronesia for food, timber, traditional medicine, and other uses. They include Artocarpus anisophyllus (entawak),[27] Artocarpus heterophyllus (jackfruit or nangka),[28] Artocarpus integer (cempedak),[29] Artocarpus lacucha (lakuch),[30] Artocarpus mariannensis (Marianas breadfruit),[31] Artocarpus odoratissimus (tarap or marang),[32] and Artocarpus treculianus (tipuho),[33] among many others. The most important species pertaining to the Austronesian expansion however, are Artocarpus camansi (breadnut or seeded breadfruit) and Artocarpus altilis (breadfruit).[34]

Artocarpus altilis (breadfruit)
[edit]
According to DNA fingerprinting studies, the wild seeded ancestor of Artocarpus altilis is the Artocarpus camansi, which is native to New Guinea, the Maluku Islands, and the Philippines. A. camansi was domesticated and selectively bred in Polynesia, giving rise to the mostly seedless Artocarpus altilis. Micronesian breadfruit also show evidence of hybridization with the native Artocarpus mariannensis, while most Polynesian and Melanesian cultivars do not. This indicates that Micronesia was initially colonized separately from Polynesia and Melanesia through two different migration events which later came into contact with each other in eastern Micronesia.[34][35][36]

The reconstructed Proto-Malayo-Polynesian word for breadfruit is *kuluʀ, which became Proto-Oceanic *kulur and Proto-Polynesian *kulu. Modern cognates include Sundanese and Malay kulur or kelur; Acehnese kulu; Iban kurur; Cebuano kulo or kolo; Muna kula; Mussau ulu; Kapingamarangi gulu; Wayan Fijian kulu; Emae kuro; Tuamotuan, Takuu, and Rarotongan kuru; Tahitian ʻuru; Samoan and Hawaiian ʻulu; and Māori kuru. Note that in Māori, kuru is only mentioned in tradition, but does not refer to the plant because breadfruit did not survive into Aotearoa.[18][37][25][38][39] Also note that it is believed that breadfruit only reached western Island Southeast Asia (Java, Sumatra, Malay Peninsula) during the recent centuries, as a result of trade with the Maluku Islands.[40]
Another notable reconstructed word for breadfruit is Proto-Oceanic *maRi or *mai. It is a common root for words for breadfruit in Micronesia, northern and western New Guinea, the Solomon Islands, the Admiralty Islands, St Matthias Islands, New Caledonia, and parts of the Central Pacific. The term itself may have originally been for Artocarpus mariannensis instead of Artocarpus altilis. Cognates include Pohnpeian, Mokil, and Ngatik māi; Palauan, Satawal, and Tuvaluan mai; Puluwat mais; Yapese maiyah; and Tongan, Niuean, and Marquesan mei.[18][37][25]
Artocarpus heterophyllus (jackfruit)
[edit]
The jackfruit (Artocarpus heterophyllus) was domesticated independently in South Asia and Southeast Asia, as evidenced by the fact that the Southeast Asian names for the fruit are not derived from the Sanskrit roots. It was probably first domesticated by Austronesians in Java or the Malay Peninsula. The word for jackfruit in Proto-Western-Malayo-Polynesian is reconstructed as *laŋkaq. Modern cognates include Sundanese, Javanese, Malay, Balinese, and Cebuano nangka; Tagalog, Pangasinan, Bikol and Ilocano langka; Chamorro lanka or nanka; Kelabit nakan; Wolio nangke; Ibaloi dangka; and Lun Dayeh laka. Note, however, that the fruit was only recently introduced to Guam via Filipino settlers when both were part of the Spanish Empire.[40][17]
Bambusoideae (bamboos)
[edit]
Various species of bamboo (subfamily Bambusoideae) are found throughout Island Southeast Asia, Mainland Southeast Asia, East Asia, and South Asia. In Austronesian regions, different types of bamboos have different names, as well as the products made from them. They are used variously as building materials, fishing gear, musical instruments, knives, water and food vessels, and so on. Bamboo shoots are also a food source in Southeast Asia. A few species of bamboo were carried by Austronesian settlers as they colonized the Pacific islands. They include the ʻohe (Schizostachyum glaucifolium), the common bamboo (Bambusa vulgaris), and the thorny bamboo (Bambusa bambos).[41][42]
Reconstructed Proto-Austronesian words that referred to bamboo include *qauR, *kawayan, *buluq, and *betung. The latter entered Proto-Malayo-Polynesian and Proto-Oceanic as *bitung, with cognates including Sundanese awi bitung; Fijian bitu; and Tongan pitu. Most terms for bamboo in Polynesia, however, originated from Proto-South-Central-Pacific *kofe (originally from Proto-Polynesian *kofe, "root"). Modern cognates include Tongan and Niue kofe; Tokelau, Marquesan, Tuamotuan, and Māori kohe; Rarotongan koʻe; Samoan and Tahitian ʻofe; and Hawaiian ʻohe. Some names have also shifted to refer to bamboo-like plants; especially in islands where they were not introduced into or did not survive, like in Aotearoa.[17][41][43]

This section needs expansion. You can help by adding to it. (March 2022) |
Broussonetia papyrifera (paper mulberry)
[edit]
Paper mulberry (Broussonetia papyrifera), better known as "tapa cloth tree" in the Pacific, originates from subtropical regions in mainland Asia and is one of the best evidence for the mainstream "Out of Taiwan" hypothesis of the Austronesian expansion. Various genetic studies have traced the origins of paper mulberry populations in the Remote Pacific all the way to Taiwan via New Guinea and Sulawesi. In the Philippines, which was along the expansion path, paper mulberry are mostly descendants of modern introductions in 1935. It is presumed that ancient introductions of paper mulberry went extinct in prehistory due to its replacement with hand-woven fabrics, given that paper mulberry generally only survives under human cultivation. However, its absence in the Philippines further underlines its origins in Taiwan, and not within Island Southeast Asia. Additionally paper mulberry populations in New Guinea also show genetic inflow from another expansion out of Indochina and South China.[44][45][46]

It is believed to be the most widely transported fiber crop in prehistory, having been transported along with the full range of the Austronesian expansion, as opposed to most of the other commensal crops in Oceania. Paper mullbery is present in almost every island or island group in Polynesia, including Rapa Nui and Aotearoa. Some populations have gone recently extinct after they stopped being cultivated, like in the Cook Islands and Mangareva, although accounts and prepared barkcloth and herbarium specimens of them exist in museum collections gathered by Europeans during the Colonial Period. They were spread by Polynesians primarily through vegetative propagation with cuttings and root shoots. They were rarely cultivated from seeds as most plants were harvested prior to flowering, when the stems reach around 1 in (2.5 cm) in diameter, as described by 18th century European accounts. It is also unknown if the feral plants reproduced sexually as the plants are dioecious and require both male and female specimens to be present in one island.[44][45]



Paper mulberry is primarily used in the Pacific Islands to make barkcloth (tapa in most Polynesian languages).[44][45] Barkcloth, can also be made from other members of the mulberry family (Moraceae), including Ficus (figs) and Artocarpus. Barkcloth was also occasionally made from Pipturus nettles, especially in Hawaii. However the highest quality of barkcloth was from paper mulberry.[18]
Barkcloth was mainly used for clothing among ancient Austronesians and is traditionally made using characteristic stone or wooden beaters which are among the most common artifacts found in Austronesian archaeological sites. Numerous archaeological remains of barkcloth beaters in southern China has been regarded as evidence that the pre-Taiwan Austronesian homelands were located in the region prior to the southward expansion of the Han Dynasty, particularly around the Pearl River Delta. The oldest such remains is from the Dingmo Site in Guangxi, dated to around 7,900 BP.[47] Barkcloth remained an important source of clothing fabrics in pre-colonial Melanesia, Polynesia, and parts of Indonesia. However, it has been mostly replaced by woven fiber clothing in most of Island Southeast Asia and Micronesia.[18]
There are numerous names for paper mulberry throughout Austronesia, the most general can be reconstructed to Proto-Central Eastern Malayo-Polynesian *malaw, which also refers to the loincloth and other items of clothing made from paper mulberry bark. Its cognates including Selaru mal; Asilulu mala ai; Buli māl; Numfor mār; Tanga, Tolai, and Gedaged mal; Rennellese mago; Kairiru myal; Lusi, Kove, Manam, Gitua, Mota, Niue, Futunan, Samoan, Tuvaluan, Nukuoro, Anuta, and Hawaiian malo; and Arosi, Rarotongan, and Māori maro.[17]
In Eastern Polynesia, terms for paper mulberry can also be reconstructed to Proto-Central Eastern-Polynesian *aute, with cognates including Tahitian and Rarotongan ʻaute; Marquesan ute; Hawaiian wauke; Rapa and Māori aute.[48]
In most of Polynesia, the term for barkcloth can also be reconstructed from Proto-Nuclear-Polynesian *taba, meaning "bark", with cognates including Wayan taba; Tongan, Samoan, Mangareva, and Rarotongan tapa; and Hawaiian kapa. Other terms widely used for barkcloth and paper mulberry are derived from the Proto-Polynesian reconstructed word *siapo, with cognates including Niue, Tongan, and Marquesan hiapo; and Samoan and East Futunan siapo.[18] The term for barkcloth beater, however, can be reconstructed more extensively back to Proto-Malayo-Polynesian *ike, with cognates including Uma ike; Sa'a iki; Bauan, Tongan, and East Futunan ike; and Samoan and Hawaiian iʻe.[18]
Calophyllum inophyllum (mastwood)
[edit]
Mastwood (Calophyllum inophyllum) is a widespread timber tree native to tropical Asia. It is notable for its ability to grow to massive sizes in sandy or rocky beaches of island and coastal habitats, as well as its habit of sending out arching large trunks over the water where its seeds are dispersed via the currents.[49][50] Due to these characters, mastwood are of particular importance to traditional shipbuilding of the larger Austronesian outrigger ships and were carried with them as they migrated to Oceania and Madagascar.[50]
Other species of the genus Calophyllum were also used similarly, like Calophyllum soulattri, Calophyllum peekelii, and Calophyllum goniocarpum. The wood grain of the members of the genus are characteristically interlocked, which make them harder to work with but also makes them stronger as well as being more suitable for carving intricate shapes. They were comparable in importance to how oaks were in European shipbuilding and timber industries.[50] In many parts of Polynesia, mastwood groves planted in marae were considered sacred and abodes of spirits. Mastwood were also carved into religious objects like tiki.[51][52] They are also commonly mentioned in the chants and folklore of Polynesia.[53]

Various parts of the mastwood were integral to the manufacture of outrigger canoes. The large curving limbs were commonly carved into the dugout canoes that formed the keel of the Austronesian outriggers ships. The strakes, which are attached to the keel by the uniquely Austronesian technique of "sewing" them with a combination of dowels and lashed lugs instead of nails, can also be made from mastwood, but it is more commonly made from other "softer" timber species like Artocarpus. Other pieces became masts, outrigger floats, and outrigger spars. Smaller curving limbs can also be carved into the ribs of the boat.[50]
Aside from shipbuilding, tamanu oil extracted from the fruit kernels were important in Polynesian culture. The oils, as well as poultices made from leaves and flowers, are also commonly used for traditional medicine.[49][51] The leaves contain compounds that are poisonous to fish and can be used as fish poison.[49]
The reconstructed Proto-Austronesian word for mastwood is *bitaquR, with modern cognates including Ilocano bittáug; Ifugao bitául; Bikol, Cebuano, Maranao, Mansaka and Manobo bitáʻog or bitaʻug; Nias bito; Palauan btáʻəs; Wetan witora; and Asilulu hataul.[17] The Western Malayo-Polynesian words for mastwood is derived from the doublet Proto-Austronesian *bintaŋuR, with cognates including Iban, Malay and Toba Batak bintangur or bentangur; Tontemboan wintangor; and Malagasy vintáno.[17] In Proto-Oceanic, the reconstructed word is pitaquR, with cognates including Nauna pitɨ; Loniu pitow; Nali pirow; Seimat hita; Aua piʻaw; Pohnpeian isou; Rotuman hefau; Fijian vetau, Tongan fetaʻu; Niue, Samoan, and Tuvaluan fetau; Nukuoro hedau; and Rennellese hetaʻu.[17] In most of these languages, the name specifically refers to C. inophyllum, although in Ifugao, Maranao, Nias, Wetan, and Fijian, the name has become more generalized to large timber trees.[17][54]
Another set of cognates for C. inophyllum in Proto-Oceanic can be reconstructed as *tamanu. Its difference from *pitaquR is unclear, but given the distinction between the terms in the Mussau reflex, *tamanu probably originally referred to specimens of the tree that grow in island interiors and not on the coastlines. Modern cognates include Mussau, Tongan, Niue, Samoan, and Rarotongan tamanu; Fijian damanu; and Hawaiian kamani.[17]
Cananga odorata (ylang-ylang)
[edit]
Cananga odorata, with its large, aromatic flowers, is used for ornamentation. The species originated from the Philippines.[55] It is not known whether it is native to Polynesia and Melanesia or introduced.[56]
Citrus
[edit]
Numerous species of Citrus are native to Island Southeast Asia, Mainland Southeast Asia, East Asia, South Asia and Near Oceania. The Austronesians cultivated and gathered a variety of citrus for food, medicine and washing with the thorns being used as piercing implements for tattooing. Citrus hystrix, Citrus macroptera, and Citrus maxima were also among the canoe plants carried by Austronesian voyagers eastwards into Micronesia and Polynesia.[57] Most Polynesian names describing citruses are named moli ultimately from Proto-Oceanic *molis;[58][59] whether a link to PMP *limaw is present (albeit indirect) or otherwise is uncertain.
Cocos nucifera (coconut)
[edit]
The region between Southwest Asia and Melanesia is the center of origin for coconuts (Cocos nucifera), where it shows greatest genetic diversity.[63][64][65][60] A study in 2011 identified two highly genetically differentiated subpopulations of coconuts, one originating from Island Southeast Asia (the Pacific group) and the other from the southern margins of the Indian subcontinent (the Indo-Atlantic group). The Pacific group is the only one to display clear genetic and phenotypic indications that they were domesticated; including dwarf habit, self-pollination, and the round "niu vai" fruit morphology with larger endosperm-to-husk ratios. The distribution of the Pacific coconuts correspond to the regions settled by Austronesian voyagers indicating that its spread was largely the result of human introductions.[60][66]

It is most strikingly displayed in Madagascar, an island settled by Austronesian sailors at around 2,000 to 1,500 BP. The coconut populations in the island show genetic admixture between the two subpopulations indicating that Pacific coconuts were brought by the Austronesian settlers that later interbred with the local Indo-Atlantic coconuts.[60][66]
Most words for "coconut" in Austronesian languages are derived from proto-Malayo-Polynesian *niuʀ. Modern cognates include Tagalog niyog; Chamorro niyok; Malay nyiur or nyior; Tetum nuu; Drehu nu; Gilbertese nii; Hawaiian, Samoan, Tongan, Fijian, and Rapa Nui niu; and Malagasy nio.[18][67][68][69][70]

Genetic studies of coconuts have also confirmed pre-Columbian populations of coconuts in Panama in South America. However, it is not native and display a genetic bottleneck resulting from a founder effect. A study in 2008 showed that the coconuts in the Americas are genetically closest related to coconuts in the Philippines, and not to any other nearby coconut populations (including Polynesia). Such an origin indicates that the coconuts were not introduced naturally, such as by sea currents. The researchers concluded that it was brought by early Austronesian sailors to the Americas from at least 2,250 BP, and may be proof of pre-Columbian contact between Austronesian cultures and South American cultures, albeit in the opposite direction than what early hypotheses like Heyerdahl's had proposed. It is further strengthened by other similar botanical evidence of contact, like the pre-colonial presence of sweet potato in Oceanian cultures.[65][62][71] During the colonial era, Pacific coconuts were further introduced to Mexico from the Spanish East Indies via the Manila galleons.[60]
In contrast to the Pacific coconuts, Indo-Atlantic coconuts were largely spread by Arab and Persian traders into the East African coast. Indo-Atlantic coconuts were also introduced into the Atlantic Ocean by Portuguese ships from their colonies in coastal India and Sri Lanka; first being introduced to coastal West Africa, then onwards into the Caribbean and the east coast of Brazil. All of these introductions are within the last few centuries, relatively recent in comparison to the spread of Pacific coconuts.[60]
Coix lacryma-jobi (Job's tears)
[edit]
The size and nutrition of Job's tears set it apart from other long-used ancient grains native to this area of Asia, and the well established and recorded use in culinary practices indicate a history at least proportional to that of millet and rice. Dating of a site in Shizitan has presented dates as early as 28,000 to 18,000 years ago.[72] Archaeologists have found evidence supporting Job's tears as among the earliest domesticates in Asia, accompanied by millet.[73]
This section needs expansion. You can help by adding to it. (January 2024) |
Colocasia esculenta (taro)
[edit]
The taro (Colocasia esculenta), sometimes referred to as the "true taro", is one of the most ancient cultivated crops and pre-dated the Austronesian expansion.[6] Taro is found widely in tropical and subtropical regions of South Asia, East Asia, Southeast Asia, Papua New Guinea, and northern Australia and is highly polymorphic, making taxonomy and distinction between wild and cultivated types difficult. It is believed that they were domesticated independently multiple times, with authors giving possible locations as New Guinea, Mainland Southeast Asia, and northeastern India, based largely on the assumed native range of the wild plants.[74][75][76] However, more recent studies have pointed out that wild taro may have a much larger native distribution than previously believed, and wild breeding types may also likely be indigenous to other parts of Island Southeast Asia.[77][78]


Archaeological traces of taro exploitation have been recovered from numerous sites pre-dating the Austronesian expansion, though whether these were cultivated or wild types can not be ascertained. They include the Niah Caves of Borneo, dating to <40,000 BP;[79] Ille Cave of Palawan, dated to at least c. 11,000 BP;[79][80] Kuk Swamp of New Guinea, dated to 10,200 to 9,910 cal BP;[81][82] and Kilu Cave in the Solomon Islands dated to around c. 28,000 to 20,000 BP.[83] In the case of Kuk Swamp, there is evidence of formalized agriculture emerging by about c. 10,000 BP, with evidence of cultivated plots, though which plant was cultivated remains unknown.[84]

Regardless, taro were definitely among the cultivated plants of Austronesians as well as preceding populations in Island Southeast Asia. However, their importance in Island Southeast Asia had largely been replaced by rice, although they are still planted at the margins of rice paddies in some communities. They remained a staple in the islands of Melanesia and Polynesia where rice wasn't introduced. They are one of the four species of aroids (taros) cultivated by Austronesians primarily as a source of starchy corms, the others being Alocasia macrorrhizos, Amorphophallus paeoniifolius, and Cyrtosperma merkusii. They are the most important and the most preferred among the four, because they were less likely to contain the irritating raphides present in the other plants.[21][85]
Taro is also identified as one of the staples of Micronesia, from archaeological evidence dating back to the pre-colonial Latte Period (c. 900 – 1521 AD), indicating that it was also carried by Micronesians when they colonized the islands.[86][87] Due to the unsuitability of the low-lying atoll islands of most of Micronesia, Micronesians innovated by digging pits that could then be filled up with compost suitable for taro cultivation.[25] Taro pollen and starch residue have also been identified in earlier Lapita sites, dated to around c. 3,050 – 2,500 cal BP.[25]
There are numerous terms for taro in the Austronesian languages, both specific and generalized. The reconstructed Proto-Austronesian term for taro is *cali, with cognates in Formosan languages including Seediq sali, Thao lhari; Bunun tai; and Amis tali.[17]
It became *taləs in Proto-Malayo-Polynesian, which in turn became *talos or *talo in Proto-Oceanic. Modern cognates include Hanunó'o tálus; Aborlan Tagbanwa talis; Palawan Batak täläs; Nias talõ; Malay talas; Minangkabau taleh; Rejang and Sundanese taleus; Javanese tales; Palauan dáit; Rotinese tale; and Tetun talas.[17][88][89] In Polynesian languages, the cognates include Motu, Marovo, Tongan, Samoan, Niue, Futunan, Tuvaluan talo; Kwaio, Lau (Malaita), and Toqabaqita alo; 'Āre'āre, Arosi, and Bauro aro; Nakanamanga na-tale; Sye tal or nal; Fijian and Nukuoro dalo; Rennellese tago; Anuta, Rarotongan, and Māori taro; and Hawaiian kalo. The English name for the plant is itself derived from the Polynesian names.[17] A red variety of taro also has names derived from reconstructed Proto-Polynesian *pongi, with cognates including Niue pongi; Marquesan poki; Hawaiian poni; and Māori pongi.[90]
In Proto-Western-Malayo-Polynesian, another reconstructed term is *kaladi, with cognates including Agutaynen, Sabah Bisaya, Iban, Tae', and Wolio kaladi; Balinese and Malay keladi; and Mongondow koladi.[17]
Cordia subcordata (beach cordia)
[edit]
The beach cordia (Cordia subcordata) is an important timber tree with light, finely textured, and somewhat soft wood ideal for carving. It has no taste and thus was most commonly used for carving utensils, cups, bowls, and other containers; as well as ornamental carvings and musical instruments throughout Austronesia. The wood is flammable and is commonly used in New Guinea as firewood. In some cultures, the wood may also be used to build paddles and the keels of the boats.[91][92] The seeds can also be eaten, though only as famine food. Other parts can also be used for traditional medicine and for the extraction of dyes. Like Calophyllum inophyllum, beach cordia were commonly planted in marae. They have cultural and religious significance in some cultures like in Kiribati and the Karimunjawa Islands of Indonesia. In Hawaii, it was traditional to plant beach cordia around houses and use their bright orange flowers as leis.[91][93][94][95]
Beach cordia, like most trees favored by Austronesians, grow well in sandy, clay, and rocky soil and are a common component in coastal forests and mangrove forests. Beach cordia was once thought to be an introduced species, but it is now known to be indigenous to most of the islands and coastlines of the Indo-Pacific, propagated naturally by their buoyant seeds. Nevertheless, they were still deliberately introduced in some islands, with artificial introductions usually found growing with other common trees cultivated by Austronesians. Especially in the atolls of Micronesia.[91][96][97]
Terms for beach cordia is reconstructed to Proto-Malayo-Polynesian *kanawa, with cognates including Iban kenawa; Makasarese kanawa; Palauan kəláu; Gilbertese kanawa; Tokelau kanava; and Nukuoro ganava.
Another set of cognates can be reconstructed to Proto-Oceanic *toRu, with cognates including Nehan to-tor; Petats to-tol; Fijian, Tongan, and Rarotongan tou; and Hawaiian kou.[17][98]
An older reconstructed term is Proto-Austronesian *qaNuNaŋ, however it is not specific to beach cordia and can refer to other members of the genus with sticky fruits, especially the glue berry (Cordia dichotoma) and the lasura (Cordia myxa). It also did not reach the Oceanic languages. Cognates include Tsou həhngə; Isneg anúnang; Hanunó'o and Cebuano anúnang; Maranao nonang; Manobo enunang; Mansaka anonang; Malay, Minangkabau, Sasak, Manggarai, and Rembong nunang; and Mongondow onunang.[17]
Cordyline fruticosa (ti)
[edit]
Ti (Cordyline fruticosa) is a palm-like plant growing up to 3 to 4 m (9.8 to 13.1 ft) tall with an attractive fan-like and spirally arranged cluster of broadly elongated leaves at the tip of the slender trunk. It has numerous color variations, ranging from plants with red leaves to green, yellow, and variegated cultivars. Its original native distribution is unknown, but it is believed to be native to the region from Bangladesh, to Mainland Southeast Asia, South China, Taiwan, Island Southeast Asia, New Guinea, and Northern Australia. It has the highest morphological diversity in New Guinea and is believed to have been extensively cultivated there. It is commonly misidentified as a "Dracaena", along with members of the genus Cordyline, due to past classification systems.[99][100]

It was carried throughout Oceania by Austronesians, reaching as far as Hawaii, New Zealand, and Easter Island at their furthest extent. A particularly important type of ti in Polynesia is a large green-leafed cultivar grown for their enlarged edible rhizomes. Unlike the ti populations in Southeast Asia and Near Oceania, this cultivar is almost entirely sterile in the further islands of eastern Polynesia. It can only be propagated by cuttings from the stalks or the rhizomes. It is speculated that this was the result of deliberate artificial selection, probably because they produce larger and less fibrous rhizomes more suitable for use as food.[99][101][102]
Ti has many uses but it is most notable as one of the most important plants related to the indigenous animist religions of Austronesians, along with fig trees (Ficus spp.). It is very widely regarded as having mystical or spiritual powers in various Austronesian (as well as Papuan) cultures. Among a lot of ethnic groups in Austronesia it is regarded as sacred. Common features include the belief that they can hold souls and thus are useful in healing "soul loss" illnesses and in exorcising against malevolent spirits, their use in ritual attire and ornamentation, and their use as boundary markers. Red and green cultivars also commonly represented dualistic aspects of culture and religion and are used differently in rituals. Red ti plants commonly symbolize blood, war, and the ties between the living and the dead; while green ti plants commonly symbolize peace and healing.[103][104][105][101] They are also widely used for traditional medicine, dye, and ornamentation throughout Austronesia and New Guinea.[106] Their ritual uses in Island Southeast Asia have largely been obscured by the introduction of Hinduism, Buddhism, Islamic, and Christian religions, but they still persist in certain areas or are coopted for the rituals of the new religions.[103]
In Polynesia, the leaves of the green-leafed form are used to wrap food, line earth ovens and fermentation pits of breadfruit, and their rhizomes harvested and processed into a sweet molasses-like pulp eaten like candy or used to produce a honey-like liquid used in various sweet treats. In Hawaii, the roots are also mixed with water and fermented into an alcoholic beverage known as okolehao.[99][107][108][109] Fibers extracted from leaves are also used in cordage and in making bird traps.[107] The consumption of ti as food, regarded as a sacred plant and thus was originally taboo, is believed to have been a daring innovation of Polynesian cultures as a response to famine conditions. The lifting of the taboo is believed to be tied to the development of the firewalking ritual.[103]

In Philippine anitism, ti were commonly used by babaylan (female shamans) when conducting mediumship or healing rituals. A common belief in Filipino cultures is that the plant has the innate ability to host spirits. Among the Ifugao people of Northern Luzon, it is planted around terraces and communities to drive away evil spirits as well as mark boundaries of cultivated fields. The red leaves are believed to be attractive to spirits and is worn during important rituals as part of the headdresses and tucked into armbands. In the past, it was also worn during ceremonial dances called bangibang, which was performed by both men and women for warriors who died in battle or through violent means. They are also used to decorate ritual objects.[110][111][112][113] Among the Palaw'an people, it is planted in burial grounds to prevent the dead from becoming malevolent spirits.[114]


In Indonesia, red ti are used similarly as in the Philippines. Among the Dayak, Sundanese, Kayan, Kenyah, Berawan, Iban and Mongondow people, red ti are used as wards against evil spirits and as boundary markers. They are also used in rituals like in healing and funerals and are very commonly planted in sacred groves and around shrines.[103][115] The Dayak also extract a natural green dye from ti.[116] During healing rituals of the Mentawai people, the life-giving spirit are enticed with songs and offerings to enter ti stems which are then reconciled with the sick person.[117] Among the Sasak people, green ti leaves are used as part of the offerings to spirits by the belian shamans.[116] Among the Baduy people, green ti represent the body, while red ti represent the soul. Both are used in rice planting rituals. They are also planted on burial grounds.[118][119] Among the Balinese and Karo people, ti plants are planted near village or family shrines in a sacred grove.[120][121] Among the Toraja people, red ti plants are used in rituals and as decorations of ritual objects. They are believed to occur in both the material and the spirit worlds (a common belief in Austronesian animism). In the spirit world, they exist as fins and tails of spirits. In the material world, they are most useful as guides used to attract the attentions of spirits. The red leaves are also symbolic of blood and thus of life and vitality.[122][123][124] Among the Ngaju people, ti plants were symbolic of the sacred groves of ancestors. They were also important in ritual promises dedicated to high gods. They were regarded as symbolic of the masculine "Tree of Life", in a dichotomy against Ficus species which symbolize the feminine "Tree of the Dead".[103]

In New Guinea, ti are commonly planted to indicate land ownership for cultivation and are also planted around ceremonial men's houses. They are also used in various rituals and are commonly associated with blood and warfare.[125][126][127] Among the Tsembaga Maring people, they are believed to house "red spirits" (spirits of men who died in battle). Prior to a highly ritualized (but lethal) warfare over land ownership, they are uprooted and pigs are sacrificed to the spirits. After the hostilities, they are re-planted in the new land boundaries depending on the outcome of the fight. The men involved ritually place their souls into the plants. The ritual warfare have been suppressed by the Papua New Guinea government, but parts of the rituals still survive.[105][128] Among the Ankave people, red ti is part of their creation myth, believed as having arisen from the site of the first murder.[129] Among the Mendi and Sulka people they are made into dyes used as body paint, and their leaves are used for body adornments and purification rituals.[130] Among the Nikgini people, the leaves have magical abilities to bring good luck and are used in divination and in decorating ritual objects.[131] Among the Kapauku people, ti plants are regarded as magical plants and are believed to be spiritual beings themselves. Unlike other magical plants which are controlled by other spirits, ti plants had their own spirits and are powerful enough to command other spiritual beings. Red plants are used in white magic rituals, while green plants are used in black magic rituals. They are also commonly used in protection and warding rituals. Among the Baktaman people, red plants are used for initiation rites, while green plants are used for healing. The Ok-speaking peoples also regard ti plants as their collective totem.[103]
In Island Melanesia, ti are regarded as sacred by various Austronesian-speaking peoples and are used in rituals for protection, divination, and fertility.[103] Among the Kwaio people, red ti are associated with feuding and vengeance, while green ti are associated with ancestor spirits, markers of sacred groves, and wards against evil. The Kwaio cultivate these varieties around their communities.[132] Among the Maenge people of New Britain, ti leaves are worn as everyday skirts by women. The color and size of leaves can vary by personal preference and fashion. New cultivars with different colors are traded regularly and strands of ti are grown near the village. Red leaves can only worn by women past puberty. Ti is also the most important plant in magic and healing rituals of the Maenge. Some ti cultivars are associated with supernatural spirits and have names and folklore around them.[133] In Vanuatu, Cordyline leaves, known locally by the Bislama name nanggaria, are worn tucked into a belt in traditional dances like Māʻuluʻulu, with different varieties having particular symbolic meanings. Cordylines are often planted outside nakamal buildings.[134] In Fiji, red ti leaves are used as skirts for dancers and are used in rituals dedicated to the spirits of the dead. They are also planted around ceremonial buildings used for initiation rituals.[103]

In Micronesia, ti leaves are buried under newly built houses in Pohnpei to ward off malign sorcery.[125] In instances of an unknown death, shamans in Micronesia communicate with the dead spirit through ti plants, naming various causes of death until the plant trembles.[101] There is also archaeological evidence that the rhizomes of the plants were eaten in the past in Guam prior to the Latte Period.[135]

In Polynesia, green ti were cultivated widely for food and religious purposes. They are commonly planted around homes, in sacred places (including marae and heiau), and in grave sites. The leaves are also carried as a charm when traveling and the leaves are used in rituals that communicate with the species. Like in Southeast Asia, they are widely believed to protect against evil spirits and bad luck; as well as having the ability to host spirits of dead people, as well as nature spirits.[99][101][107]

In ancient Hawaiʻi the plant was thought to have great spiritual power; only kahuna (shamans) and aliʻi (chiefs) were able to wear leaves around their necks during certain ritual activities. Ti was sacred to the god of fertility and agriculture Lono, and the goddess of the forest and the hula dance, Laka. Ti leaves were also used to make lei, and to outline borders between properties. It was also planted at the corners of the home to keep evil spirits away. To this day some Hawaiians plant ti near their houses to bring good luck. The leaves are also used for lava sledding. A number of leaves are lashed together and people ride down hills on them. The leaves were also used to make items of clothing including skirts worn in dance performances. The Hawaiian hula skirt is a dense skirt with an opaque layer of at least fifty green leaves and the bottom (top of the leaves) shaved flat. The Tongan dance dress, the sisi, is an apron of about 20 leaves, worn over a tupenu, and decorated with some yellow or red leaves.[136][137][138]
In Aotearoa, certain place names are derived from the use and folklore of ti, like Puketī Forest and Temuka. The ti plants in Kaingaroa are known as nga tī whakāwe o Kaingaroa ("the phantom trees of Kaingaroa"), based on the legend of two women who were turned into ti plants and seemingly follow people traveling through the area.[107]
The reconstructed Proto-Malayo-Polynesian word for ti is *siRi. Cognates include Malagasy síly; Palauan sis; Ere and Kuruti siy; Araki jihi; Arosi diri; Chuukese tii-n; Wuvulu si or ti; Tongan sī; Samoan, Tahitian, and Māori tī; and Hawaiian kī. The names in some languages have also been applied to the garden crotons (Codiaeum variegatum), which similarly have red or yellow leaves. The cognates of Proto-Western-Malayo-Polynesian *sabaqaŋ, similarly, have been applied to both garden crotons and ti plants.[17][139]
In the Philippines, they are also known by names derived from the Proto-Austronesian *kilala, "to know", due to its use in divination rituals. Cognates derived from that usage include Tagalog sagilala; and Visayan and Bikol kilála or kilaa. In New Zealand, the terms for ti were also transferred to the native and closely related cabbage tree (Cordyline australis), as tī kōuka.[17][139]
Cucumis melo (melon)
[edit]
This section needs expansion. You can help by adding to it. (January 2024) |
Cyrtosperma merkusii (giant swamp taro)
[edit]
This section needs expansion. You can help by adding to it. (January 2024) |
Dioscorea (yams)
[edit]Yams (Dioscorea spp.) is a very large group of plants native throughout tropical and warm temperate regions of the world. Various species of yams were domesticated and cultivated independently within Island Southeast Asia and New Guinea for their starchy tubers, including the ube (Dioscorea alata), round yam (Dioscorea bulbifera), intoxicating yam (Dioscorea hispida), lesser yam (Dioscorea esculenta), Pacific yam (Dioscorea nummularia), fiveleaf yam (Dioscorea pentaphylla), and pencil yam (Dioscorea transversa).[140] Among these, D. alata and D. esculenta were the only ones regularly cultivated and eaten, while the rest were usually considered as famine food due to their higher levels of the toxin dioscorine which requires that they be prepared correctly before consumption.[141]
D. alata and D. esculenta were the most suitable for long transport in Austronesian ships and were carried through all or most of the range of the Austronesian expansion. D. alata in particular, were introduced into the Pacific Islands and New Zealand. They were also carried by Austronesian voyagers into Madagascar and the Comoros.[66][142][143]
Dioscorea alata (ube)
[edit]
The ube (Dioscorea alata), also known as the greater yam or water yam, is one of the most important staple crops in Austronesian cultures. It is the main species cultivated among Dioscorea, largely because of its much larger tubers and its ease of processing.[144] Its center of origin is unknown, but archaeological evidence suggests that it was exploited in Island Southeast Asia and New Guinea before the Austronesian expansion. Ube is believed to be a true cultigen, only known from its cultivated forms. It is a polyploid and is sterile, and thus can not cross bodies of water. This restricts its introduction into islands purely by human agency, making them a good indicator of human movement. Some authors have proposed an origin in Mainland Southeast Asia without evidence, but it shows the greatest phenotypic variability in the Philippines and New Guinea.[145][146][147]

Based on archaeological evidence of early farming plots and plant remains in the Kuk Swamp site, authors have suggested that it was first domesticated in the highlands of New Guinea from around 10,000 BP and spread into Island Southeast Asia via the Lapita culture at around c. 4,000 BP, along with D. nummularia and D. bulbifera. In turn, D. esculenta is believed to have been introduced by the Lapita culture into New Guinea. There is also evidence of an agricultural revolution during this period brought by innovations from contact with Austronesians, including the development of wet cultivation.[76][148]

However, much older remains identified as being probably D. alata have also been recovered from the Niah Caves of Borneo (Late Pleistocene, <40,000 BP) and the Ille Cave of Palawan (c. 11,000 BP), along with remains of the toxic ubi gadong (D. hispida) which requires processing before it can be edible. Although it doesn't prove cultivation, it does show that humans already had the knowledge to exploit starchy plants and that D. alata were native to Island Southeast Asia. Furthermore, it opens the question on whether D. alata is a true species or cultivated much older than believed.[140][79][80][149][150][151]
Ube remains an important crop in Southeast Asia. Particularly in the Philippines where the vividly purple variety is widely used in various traditional and modern desserts. It also remains important in Melanesia, where it is also grown for ceremonial purposes tied to the size of the tubers at harvest time. Its importance in eastern Polynesia and New Zealand, however, has waned after the introduction of other crops, most notably the sweet potato.[144]
The reconstructed Proto-Austronesian word for ube is *qubi, which became Proto-Malayo-Polynesian *qubi, and Proto-Oceanic *qupi. It has some of the most recognizable and widespread reflexes in Austronesian languages. Modern cognates include Yami uvi; Itbayaten ovi; Bontoc and Hanunó'o ʻúbi; Ilocano, Tagalog, Cebuano, Pangasinan, Aklanon, Itneg, and Itawis úbi or úbe; Kalamian Tagbanwa kubi; Maranao obi; Tiruray ʻubi; Manobo uvi; Kenyah, Malay, Iban, Balinese, Sasak, Mongondow, and Toba Batak ubi; Javanese uwi; Kelabit ubih; Melanau ubey; Ngaju Dayak owi; Malagasy óvy; Tsat phai; Jarai hebey; Moken koboi; Sundanese huwi; Tontemboan, Bimanese, and Manggarai uwi; Ngadha uvi; Rotinese ufi; Erai uhi; Selaru uh or uhi-re; Watubela kuwi; Buruese ubi-t; Koiwai uf; Buli up; and Waropen uwi.[144][17]
Among Oceanic languages, cognates include Nauna kuh; Penchal kup; Leipon uh; Tolai up; Lakalai la-huvi; Gapapaiwa and Kilivila kuvi; Papapana na-uvi; Simbo, Bugotu, and Nggela, and Fijian uvi; Kwaio, Niue, and Samoan ufi; Sa'a, Arosi, Tuamotuan, Hawaiian and Rapa Nui uhi; Marquesan puauhi; Haununu a-uhi; Avava ''o-ovi; Rennellese ʻuhi; Tongan ʻufi; Anuta upi; Rarotongan uʻi; and Māori uwhi or uhi.[144][17]
In some ethnic groups, the word has been generalized or shifted to mean other types of yams, as well as the sweet potato and cassava. Other words for ube are also derived from the ancestral names of other species of yam.[144][17]
Dioscorea bulbifera (air yam)
[edit]
The air yam (Dioscorea bulbifera), also known as the bitter yam, is one of the lesser cultivated species of yam. It is usually only eaten as famine food in Island Southeast Asia, Melanesia, and Polynesia, because of the toxicity of some wild or feral plants when not cooked correctly. However it is one of only three yams that were carried by Austronesians into Remote Oceania, the others being D. alata and D. nummularia. The part of the plant harvested are the aerial tubers, as it does not usually produce large underground tubers.[23][152]
It can be reconstructed to Proto-Oceanic as *pwatika or *pʷatik, with cognates including Lou puet; Lamusong patik; Boanaki posika; and Kwara'ae fasia. However, in Lamusong its meaning has shifted to the lesser yam, while in Boanaki, the meaning has shifted to a more generalized term for yams. It can also be reconstructed to the more generalized Proto-Oceanic *balai, meaning "wild yam", which became Proto-Micronesian *palai, with cognates including Rotuman parai; Tongan, Niue, and Samoan palai; and Rennellese pagai.[17]
Dioscorea esculenta (lesser yam)
[edit]
The lesser yam (Dioscorea esculenta) is the second most important yam crop among Austronesians. Like D. alata, it requires minimal processing, unlike the other more bitter yam species. However, it has smaller tubers than D. alata and is usually spiny.[153] Like D. alata it was introduced to Madagascar and the Comoros by Austronesians, where it spread to the East African coast.[154][142][155] They are also a dominant crop in Near Oceania, However, it did not reach to the furthest islands in Polynesia, being absent in Hawaii and New Zealand.[156][157][158]
Starch grains identified to be from the lesser yam have been recovered from archaeological sites of the Lapita culture in Viti Levu, Fiji, dated to around 3,050 to 2,500 cal BP.[159] Traces of D. esculenta (along with D. alata, D. bulbifera, D. nummularia and D. pentaphylla) yams have also been identified from the Mé Auré Cave site in Moindou, New Caledonia, dated to around 2,700 to 1,800 BP.[160] Remains of D. esculenta have also been recovered from archaeological sites in Guam, dated to around 1031 CE.[135] D. esculenta is believed to have been introduced by the Lapita culture into New Guinea at around 4,000 BP, along with agricultural innovations like wet cultivation as well as swidden farming.[18][76][148] In archaeological sites in New Guinea, it is associated with the appearance of high-density populations in the coastal areas.[161]
Terms for lesser yam in Austronesian languages are mostly affixed or two-word forms derived from the *qubi root for D. alata, like Samoan ufi lei, Sundanese ubi aung, Malay ubi torak, and Javanese ubi gemblii. A term for lesser yam can be reconstructed in Proto-Philippine as *tugiq, but its cognates are limited to the island of Luzon, including Ivatan togi; Ilocano and Kankana-ey tugí; Bontoc and Ifugao tugi; and Tagalog tugiʻ.[17] No Proto-Oceanic term can be reconstructed for the lesser yam because it is absent in Remote Oceania. However, it can be reconstructed in Proto-Western-Oceanic as *kamisa, *qamisa, or *mamisa.[162]
Dioscorea hispida (intoxicating yam)
[edit]The intoxicating yam (Dioscorea hispida), is native to tropical Asia and New Guinea. It is only cultivated minimally in parts of Java. Elsewhere it is harvested from the wild. Like D. bulbifera it has toxic tubers that need to be prepared correctly before they can be eaten, and thus were only suitable for famine food.[163] However, it is one of the Dioscorea species identified from the Niah Caves archaeological site dating to <40,000 BP.[79][164] Its names can be reconstructed to Proto-Western-Malayo-Polynesian *gaduŋ. Its modern cognates in most Western Malayo-Polynesian languages is gadung or gadong (also ubi gadung or ubi gadong). The names are also applied to the similarly toxic introduced cassava.[17]
Ficus (fig trees)
[edit]Ficus is a genus of about 850 species of woody trees, shrubs, vines, epiphytes and hemiepiphytes of the family Moraceae, which are collectively known as fig trees or figs. These plants are native to the tropics, with some species extending into the semi-warm temperate zone. Despite not being a genus exclusive to the Austronesian regions, several species such as ficus dammaropsis, ficus fistulosa, ficus hispida, ficus nota, ficus pseudopalma, ficus septica, ficus variegata, ficus aspera, ficus fraseri, ficus tinctoria, ficus ulmifolia, ficus wassa, ficus mutabilis, ficus deltoidea, ficus nota and ficus pseudopalma are endemic to these regions, and played an important role in Austronesian cultures.[165][166][167][168][169][170][171][172][173][174][175][176]
Ficus aspera
[edit]
Ficus aspera, also known as mosaic fig, is a plant native to Vanuatu, in the South Pacific region. The fruits of this plant are cauliflowerous (fruits that form from their main stems or woody trunks instead of new shoots). The mosaic fig is used as an ornamental plant.[173][177]
Ficus dammaropsis
[edit]Ficus dammaropsis, known as kapiak in Tok pisin, is a tropical fig with huge 60 cm pleated leaves native to the highlands and highland fringe of New Guinea . Its fruit is edible, but it is rarely eaten except as an emergency food. When consumed, the young leaves are pickled or boiled and eaten as a salad with pork.[178]
Ficus fraseri
[edit]Ficus fraseru, also known as white sandpaper fig and bright sandpaper fig is one of several species of figs known as paper figs' sandpaper . This fig is native to New South Wales, Queensland, and Northern New Caledonia and Vanuatu. This fig grows as a bush or as a tree, with a height that varies between 6 and 15 meters. Its leaves are 6 to 14 cm long and 2.5 to 6.5 cm wide on petioles 1 to 2 cm long. The rounded figs are 1 to 1.5 cm long and start out yellow, maturing to orange-red between May and February in the species' native range. These are edible but tasteless.[179][174][180][181] Although rarely seen in cultivation, it is a fast growing ornamental species. It can be easily propagated from seeds.
Ficus nota
[edit]
Ficus nota, is a species of flowering plant known as tibig, found near water at low altitudes. The tibig is native to the Philippines. They are also found in parts of northern Borneo, in Malaysia. The tree can grow up to 9 meters high.
The fruits are also edible for humans, although they are quite tasteless. They are usually eaten with sugar and cream in the Philippines. The raw leaves are also eaten as a vegetable.[182][183][184]
Ficus pseudopalma
[edit]Ficus pseudopalma, is a species of fig commonly known as Philippine fig, Philippine fig, dracaena fig or palm leaf fig' . This is an endemic species of Philippines, especially the island of Luzon.[185][186]
This is a bush that grows erect with a bare, branchless stem topped with a clump of leaves that give it the appearance of a palm tree (hence the term pseudopalma in its name, meaning "false palm"). The fruit is a dark green fig that grows in pairs, each fruit just over an inch long.[187]
In Luzon, this plant is found in grassland and forest habitats, where it is considered common. The shoots of this plant are consumed as a type of vegetable, and there are several traditional uses; among these is its use as a remedy for kidney stones, which is obtained from the leaves. In the Bicol region, the plant is known as Lubi-lubi, and the ojas are cooked in coconut milk. This shrub has also been used as a landscaping plant in Hawaii, but it never escaped cultivation or became established in the wild, because the wasp species that pollinates it never reached the islands.[188][189]
Ficus tinctoria
[edit]
Ficus tinctoria, also known as coloring fig or hunchback fig, is a tree belonging to one of the species known as strangler figs. This is found in Malaysia, northern Australia and the islands of the South Pacific.
Palms are favorable host species. The root systems of the coloring fig can join to be self-supporting, but the epiphyte usually drops if the host tree dies or rots.The small rust-brown fruit of the dye fig is the source of a red dye used in traditional fabric making in parts of Oceania and Indonesia. The fruit is also edible and an important food source in the low-lying atolls of Micronesia and Polynesia.[190][191]
Ficus variegata
[edit]Ficus variegata, is a species of tropical fig found in various parts of Asia, Pacific islands and southeastern Australia. There are several names for this species, such as common red-stemmed fig, green-fruited fig and variegated fig.[172][171]
Ipomoea batatas (sweet potato)
[edit]Lagenaria siceraria (bottle gourd)
[edit]Morinda citrifolia (noni)
[edit]
Noni (Morinda citrifolia) is native to Southeast Asia extending to New Guinea and northern Australia. It grows readily in beach and rocky environments. It has been introduced widely into the Pacific. All parts of the plant were used by Austronesians for traditional medicine and timber, but its most common traditional use is for the extraction of red or yellow dyes. The odor of the plant and the fruit was also traditionally believed to repel evil spirits. The fruit is also edible, but is usually only eaten as famine food.[192]
There are several terms for noni that can be reconstructed. The most widespread is Proto-Central-Eastern Malayo-Polynesian *ñəñu. Cognates include Kapampangan lino; Tagalog and Bikol níno; Cebuano ninú; Gedaged nanom or nonom; Takia nom; Bimanese nonu; Tetun nenu-k; Leti and Asilulu nenu; Leti (Moa) nienu; Wetan neni. It became Proto-Oceanic *ñoñu, with cognates including Nali non; Leipon and Wogeo ñoñ; Bipi ñoy; Gitua and Rarotongan nono; Gilbertese non; Motu, Tongan, Niue, Futunan, Samoan, Tuvaluan, Kapingamarangi, Nukuoro, and Anuta nonu; and Hawaiian noni (from which the English name is derived from). In some languages the meaning has shifted to mean "small tree" or "shrub" or to the closely related Morinda umbellata and Morinda bracteata.[17]
In Western Malayo-Polynesian, another term that can be reconstructed is Proto-Western Malayo-Polynesian *baŋkudu, which may have referred to a different species of Morinda originally. Its cognates including Tagalog and Cebuano bangkúro; Agutaynen bangkoro; Tausug, Toba Batak, and Balinese bangkudu; Sundanese cangkudu; Sasak bengkudu; Mongondow bongkudu; and Malay mengkudu.[17]
There are also smaller cognate sets, like Proto-Philippine *apatut for the tree and Proto-Oceanic *gurat and *kurat for the red dye produced from the tree.[17]
Musa (bananas)
[edit]
The earliest domestication of bananas (Musa spp.) were initially from naturally occurring parthenocarpic (seedless) individuals of Musa acuminata banksii in New Guinea, before the arrival of Austronesian-speakers. Numerous phytoliths of bananas have been recovered from the Kuk Swamp archaeological site and dated to around 10,000 to 6,500 BP. From New Guinea, cultivated bananas spread westward into Island Southeast Asia through proximity (not migrations). They hybridized with other (possibly independently domesticated) subspecies of Musa acuminata as well as Musa balbisiana in the Philippines, northern New Guinea, and possibly Halmahera. These hybridization events produced the triploid cultivars of bananas commonly grown today. From Island Southeast Asia, they became part of the staple crops of Austronesian peoples and were spread during their voyages and ancient maritime trading routes into Oceania, East Africa, South Asia, and Indochina.[6][193][194]

These ancient introductions resulted in the banana subgroup now known as the "true" plantains, which include the East African Highland bananas and the Pacific plantains (the Iholena and Maoli-Popo'ulu subgroups). East African Highland bananas originated from banana populations introduced to Madagascar probably from the region between Java, Borneo, and New Guinea; while Pacific plantains were introduced to the Pacific Islands from either eastern New Guinea or the Bismarck Archipelago.[6][193]
A second wave of introductions later spread bananas to other parts of tropical Asia, particularly Indochina and the Indian Subcontinent.[6][193]
Musa abaca (abacá)
[edit]Abacá (Musa textilis), also known as Manila Hemp, is grown traditionally for its fiber in the Philippines. It was once one of the world's premier fibers, valued for its use in soft, lustrous, and silky fabrics. It was a major luxury export of the Philippines during the Colonial Era, and was introduced to Hawaii and Central America by Europeans. It has since been replaced by synthetic fibers like rayon and nylon.[195]
Musa × troglodytarum (fe'i banana)
[edit]Fe'i bananas (Musa × troglodytarum), also spelled Fehi or Féi, are banana cultivars unique to Melanesia, the Maluku Islands, and Polynesia. Unlike other domesticated banana cultivars which are derived from Musa acuminata and Musa balbisiana, fe'i bananas are believed to be hybrids derived from entirely different species. Proposed progenitors of fe'i bananas include Musa jackeyi, Musa lolodensis, Musa maclayi, and Musa peekelii, all of which are native to New Guinea and surrounding islands. Like other bananas, they were spread eastwards to Polynesia for use as food. However, they are absent in Island Southeast Asia, reaching only as far as the Maluku Islands.[195]
Myristica fragrans (nutmeg)
[edit]The earliest evidence of use of nutmeg (Myristica fragrans ) comes in the form of 3,500-year-old potsherd residues from the island of Pulau Ai, one of the Banda Islands in eastern Indonesia.[196] The Banda Islands consist of eleven small volcanic islands, and are part of the larger Maluku Islands group. These islands were the only source of nutmeg and mace production until the mid-19th century.[197] It was one of the spices traded over the Austronesian maritime spice trade network since at least 1500 BCE.[26]
In the sixth century AD, nutmeg use spread to India, then further west to Constantinople.[198] By the 13th century, Arab traders had pinpointed the origin of nutmeg to the Banda Islands, but kept this location a secret from European traders.[198]
Oryza sativa (rice)
[edit]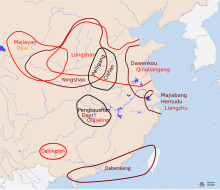
(8,500 to 1,500 BCE)
Rice (Oryza sativa) is one of the most ancient Austronesian staples, and is likely to have been originally domesticated by their ancestors long before the Austronesian expansion. It remains the main crop plant cultivated in Island Southeast Asia.[199]

There are two most likely centers of domestication for rice as well as the development of the wetland agriculture technology. The first, and most likely, is in the lower Yangtze River, believed to be the homelands of early Austronesian speakers and associated with the Kauhuqiao, Hemudu, Majiabang, and Songze cultures. It is characterized by typical Austronesian innovations, including stilt houses, jade carving, and boat technologies. Their diet were also supplemented by acorns, water chestnuts, foxnuts, and pig domestication.[199][201][202][203][204][200][205]

The second is in the middle Yangtze River, believed to be the homelands of the early Hmong-Mien-speakers and associated with the Pengtoushan and Daxi cultures. Both of these regions were heavily populated and had regular trade contacts with each other, as well as with early Austroasiatic speakers to the west, and early Kra-Dai speakers to the south, facilitating the spread of rice cultivation throughout southern China.[200][199]
The spread of japonica rice cultivation to Southeast Asia started with the migrations of the Austronesian Dapenkeng culture into Taiwan between 5,500 and 4,000 BP. The Nanguanli site in Taiwan, dated to ca. 4,800 BP, has yielded numerous carbonized remains of both rice and millet in waterlogged conditions, indicating intensive wetland rice cultivation and dryland millet cultivation.[199]

From about 4,000 to 2,500 BP, the Austronesian expansion began, with settlers from Taiwan moving south to colonize Luzon in the Philippines, bringing rice cultivation technologies with them. From Luzon, Austronesians rapidly colonized the rest of Island Southeast Asia, moving westwards to Borneo, the Malay Peninsula and Sumatra; and southwards to Sulawesi and Java. By 2,500 BP, there is evidence of intensive wetland rice agriculture already established in Java and Bali, especially near very fertile volcanic islands.[199]
However, rice (as well as dogs and pigs) did not survive the first Austronesian voyages into Micronesia due to the sheer distance of ocean they were crossing. These voyagers became the ancestors of the Lapita culture. By the time they migrated southwards to the Bismarck Archipelago, they had already lost the technology of rice farming, as well as pigs and dogs. However, knowledge of rice cultivation is still evident in the way they adapted the wetland agriculture techniques to taro cultivation. The Lapita culture in Bismarck reestablished trade connections with other Austronesian branches in Island Southeast Asia.[199]
The Lapita culture also came into contact with the non-Austronesian (Papuan) early agriculturists of New Guinea and introduced wetland farming techniques to them. In turn, they assimilated their range of indigenous cultivated fruits and tubers, as well as reacquiring domesticated dogs and pigs, before spreading further eastward to Island Melanesia and Polynesia.[199]
Rice, along with other Southeast Asian food plants, were also later introduced to Madagascar, the Comoros, and the coast of East Africa by around the 1st millennium CE by Austronesian sailors from the Greater Sunda Islands.[142]
Much later Austronesian voyages from Island Southeast Asia succeeded in bringing rice to Guam during the Latte Period (1,100 to 300 BP). Guam is the only island in Oceania where rice was grown in pre-colonial times.[206][207]
Pandanus (pandan)
[edit]
Pandanus (Pandanus spp.) are very important cultivated plants in the Pacific, second only in importance and pervasiveness to coconuts. Every part of the plant is utilized, including for food, building materials, traditional medicine, and fiber and weaving materials in various cultures in Austronesia. The plants (particularly the fragrant flowers) also had spiritual significance among the native animist Austronesian religions.[208][209]

Pandanus were also profoundly crucial in enabling the Austronesian expansion. Their leaves were traditionally woven into mats used in the sails for Austronesian outrigger ships. Sails allowed Austronesians to embark on long-distance voyaging. In some cases, however, they were one-way voyages. The failure of pandanus to establish populations in Rapa Nui and Aotearoa is believed to have isolated their settlements from the rest of Polynesia.[210][209]
The word for pandanus in Austronesian languages is derived from Proto-Austronesian *paŋudaN, which became Proto-Oceanic *padran and Proto-Polynesian *fara, the latter two usually referring specifically to Pandanus tectorius. Cognates in modern Austronesian languages include Kanakanavu pangətanə; Thao and Bunun panadan; Tagalog pandan; Chamorro pahong; Ratahan pondang; Malay pandan (from which the English name is derived from); Manggarai pandang; Malagasy fandrana; Lau fada-da; Fijian vadra; Samoan fala; Tongan fā; Tahitian fara; Hawaiian hala; and Māori whara or hara. Note that among the Formosan languages of Indigenous Taiwanese, the meaning of the words have largely shifted to mean "pineapple", a physically similar non-native European-introduced plant. In Māori, as well, the meaning has shifted to Astelia spp. and Phormium tenax (harakeke), similar plants used for weaving, since pandanus did not survive the voyage into Aotearoa.[211][212]

Pandanus grow well in island habitats, being very salt-tolerant and easy to propagate, making them ideal plants for early Austronesian sailors. Like coconuts, they grow predominantly along strandlines, mangrove forests, and other coastal ecosystems. They can also be found in the understory of forests in larger islands. Others may also be found in highland groves, likely planted by humans. Both pandanus and coconuts are adapted to withstand the strong winds of the frequent typhoons of the Indo-Pacific. The greatest center of diversity of Pandanus is the western Pacific and Island Southeast Asia. The genus has around 600 species, but the most important and the most widespread group of species in Austronesian cultures and is the Pandanus tectorius complex.[213]

Pandanus tectorius in Oceania show evidence of long cultivations, with hundreds of different selectively bred cultivars which are primarily propagated through cuttings. These varieties often have different names in local languages and have different physical characteristics. The varieties are predominantly distinguished by the color and edibility of their fruit, but they may also be differentiated based on other criteria like the color and shape of their leaves used for weaving.[213]
Very old fossils of Pandanus tectorius have been recovered from Hawaii, dated to more than 1.2 million years old. This indicates that the plants once colonized Hawaii (and likely the rest of the Pacific islands) naturally through their buoyant fruits. However, useful domesticated varieties were carried by Austronesians from island to island. Especially since wild pandanus have calcium oxalate crystals (raphides) in their fruit tissue. They cause itchiness and irritation when eaten raw and thus need to be cooked. Domesticated varieties which have less raphides (which are also usually less fibrous and more nutritious), were therefore valued . It is thus considered both native and introduced.[213][209] There are also fossil evidence of pandanus fruits being harvested for food in New Guinea from archaeological sites dated to around 34,000 to 36,000 BP.[209]
Other important species of pandanus utilized by Austronesians include Pandanus amaryllifolius, Pandanus odorifer, Pandanus furcatus, Pandanus julianettii, Pandanus simplex, Pandanus utilis, Pandanus dubius, and Pandanus whitmeeanus, among many others. Pandanus odorifer is widespread in the region from western Micronesia, to Island Southeast Asia and South Asia. It is possibly a subspecies of Pandanus tectorius and they hybridize readily.[213] Pandanus amaryllifolius, the pandan, is another important species widely used as a spice in the cuisines of Southeast Asia for their vanilla-like fragrant leaves.[214]
Piper (peppers)
[edit]Peppers (Piper) ancestrally cultivated by Austronesians include the betel (Piper betle), cubeb pepper (Piper cubeba), kava (Piper methysticum), and the Javanese long pepper (Piper retrofractum). Many others were also harvested from the wild for medicinal or religious purposes, including Piper caducibracteum, Piper excelsum, Piper ornatum, and Piper sarmentosum.[215] Black pepper (Piper nigrum) and long pepper (Piper longum) were also extensively cultivated in Island Southeast Asia after early contact by Austronesian traders with South India and Sri Lanka.[216]
Piper betle (betel)
[edit]
The betel (Piper betle) is one of the two plants that comprise the main ingredients of betel chewing, the other being the areca nut (Areca catechu). It is one of the most ubiquitous practices of the Austronesians. It is consumed by taking a leaf of betel, wrapping it around an areca nut and some lime (obtained from grinding seashells), and then chewing it for some time. It is a stimulant, inducing slight dizziness followed by euphoria and alertness. It is also highly addictive, damages the teeth and gums, and stains the teeth red.[26][25]

Based on archaeological, linguistic, and botanical evidence, betel chewing is most strongly associated with Austronesian cultures, despite its widespread adoption by neighboring cultures in prehistoric and historic times. The original range of betel is unknown, but Areca catechu is known to be originally native to the Philippines, where it has the greatest morphological diversity as well as the most number of closely related endemic species. It is unknown when the two were combined, as areca nut alone can be chewed for its narcotic properties.[26] In eastern Indonesia, however, leaves from the wild Piper caducibracteum (known as sirih hutan) are also harvested and used in place of betel leaves.[217]
The oldest unequivocal evidence of betel chewing is from the Philippines. Specifically that of several individuals found in a burial pit in the Duyong Cave site of Palawan island dated to around 4,630±250 BP . The dentition of the skeletons are stained, typical of betel chewers. The grave also includes Anadara shells used as containers of lime, one of which still contained lime. Burial sites in Bohol dated to the first millennium CE also show the distinctive reddish stains characteristic of betel chewing. Based on linguistic evidence of how the reconstructed Proto-Austronesian term *buaq originally meaning "fruit" came to refer to "areca nut" in Proto-Malayo-Polynesian, it is believed that betel chewing originally developed somewhere within the Philippines shortly after the beginning of the Austronesian expansion (~5,000 BP). From the Philippines, it spread back to Taiwan, as well as onwards to the rest of Austronesia.[26]

It reached Micronesia at around 3,500 to 3,000 BP with the Austronesian voyagers, based on both linguistic and archaeological evidence.[218] It was also previously present in the Lapita culture, based on archaeological remains from Mussau dated to around 3,600 to 2,500 BP. But it did not reach Polynesia further east. It is believed that it stopped in the Solomon Islands due to the replacement of betel chewing with the tradition of kava drinking prepared from the related Piper methysticum.[219][17] It was also diffused into East Africa via the Austronesian settlement of Madagascar and the Comoros by around the 7th century.[26]
The practice also diffused to the cultures the Austronesians had historical contact with. It reached South Asia by 3,500 BP, through early contact of Austronesian traders from Sumatra, Java, and the Malay Peninsula with the Dravidian-speakers of Sri Lanka and southern India. This also coincides with the introduction of Southeast Asian plants like Santalum album and Cocos nucifera, as well as the adoption of the Austronesian outrigger ship and crab-claw sail technologies by Dravidian-speakers. It Mainland Southeast Asia by 3,000 to 2,500 BP through trade with Borneo, as well as the settlement of the Champa polities in southern Vietnam. From there, it was spread northwards into China. Lastly, it reached Northern India by 500 BP through trade in the Bay of Bengal. From there it was spread westwards into Persia and the Mediterranean.[26][220][221]
There are very old claims of betel chewing dating to at least 13,000 BP at the Kuk Swamp site in New Guinea, based on probable Areca sp. recovered. However, it is now known that these might have been due to modern contamination of sample materials. Similar claims have also been made at other older sites with Areca sp. remains, but none can be conclusively identified as A. carechu and their association with betel peppers is tenuous or nonexistent.[26]
There are numerous cognate sets reconstructible in Austronesian languages relating to various aspects of betel chewing. Ranging from chewing something without swallowing to equipment used to climb areca nut palms to the betel spittle. One cognate set that can be reconstructed for betel pepper is Proto-Western Malayo-Polynesian *Rawed which became Proto-Philippine *gawed, with cognates including Yami gaod, Itbayaten gawed; Ilocano gawéd; Isneg khawád; Casiguran Dumagat gawə́d; and Ibaloy kawed; Balangaw lawɨ'd; Kalagan lawód; and Kenyah auat or awet.[17]
Two other cognate sets reached into Oceania. The first is Proto-Malayo-Polynesian *pu-pulu, which became Proto-Oceanic *[pu-]pulu. Cognates include Mussau ulo; Loniu pun; Bipi pun or puepun; Lukep ul; Takia ful; Gedaged fu; Manam ulusalaga; and Bugotu vu-vulu. The other is Proto-Meso-Melanesian *siqa(r,R)(a), with cognates including Kara and Lihir sie; Tabar sia; Patpatar sier; Tolai ier; Nehan hiara; Petats sil; Teop hia(kuru); Tinputz (ta)sian; Banoni siɣana; and Marovo hirata.[25][222]
Piper cubeba (cubeb pepper)
[edit]The cubeb pepper (Piper cubeba) are native to Island Southeast Asia. Like Piper retrofractum, however, it was only cultivated extensively in the Greater Sunda Islands for the spice trade. The Javanese protected the monopoly of the trade by sterilizing the seeds before trading them. It has a pungent smell, often compared to allspice, quite unlike that of the other culinary peppers. It also has a slightly bitter taste. It is notable as having reached as far as Greece during ancient times via the Silk Road. It was a valuable rare spice in Medieval Europe and the Middle East, reputed to have medicinal and magical properties. Medieval Arab physicians commonly used it for a range of treatments, ranging from treating infertility to poison antidotes. It is mentioned in The Book of One Thousand and One Nights as well as in the travelogues of Marco Polo. Its trade waned during the Colonial Era when the Portuguese Empire banned its importation to promote the black pepper produced by its own colonies.[223][224][225][226]
Piper excelsum (kawakawa)
[edit]
Kawakawa (Piper excelsum) is a small tree or shrub endemic to New Zealand and nearby Norfolk Island and Lord Howe Island. It was exploited by Austronesian settlers based on previous knowledge of the kava, as the latter could not survive in the colder climates of Aotearoa. The Māori name for the plant, kawakawa, is derived from the same etymon as kava, but reduplicated. It is a sacred tree among the Māori people. It is seen as a symbol of death, corresponding to the rangiora (Brachyglottis repanda) which is the symbol of life. Boughs of kawakawa are often used in purification rituals.[227]
However, kawakawa's resemblance to true kava is only superficial. Kawakawa roots do not have psychoactive properties. Instead, kawakawa's primary use is for traditional medicine.[227]
Piper methysticum (kava)
[edit]Kava (Piper methysticum) is a small tree or shrub believed to have been domesticated in either New Guinea or Vanuatu by Papuans. It is believed to be a domesticated variety of Piper subbullatum which is native to New Guinea and the Philippines.[222]


It was spread by Austronesians after contact into the rest of Polynesia. It is endemic to Oceania and is not found in other Austronesian groups. Kava has great cultural and religious significance among Polynesians. The roots are pounded and mixed with water then strained through fibers. The resulting cloudy gray liquid is bitter with mildly psychoactive and narcotic properties, with a common effect being numbness around the lips and mouth. However, it is not hallucinogenic nor addictive. The potency of the root depends on the age of the plants. The leaves and roots can also be chewed directly resulting in a numbing effect and relaxation. It is traditionally consumed both in everyday social interactions and in religious rituals. Kava reached Hawaii, but it is absent in Aotearoa where it can not grow.[222][219][228]
Consumption of kava is also believed to be the reason why betel chewing, ubiquitous elsewhere, was lost for Austronesians in Oceania.[17]
According to Lynch (2002), the reconstructed Proto-Polynesian term for the plant, *kava, was derived from the Proto-Oceanic term *kawaRi in the sense of a "bitter root" or "potent root [used as fish poison]". It originally referred to Zingiber zerumbet, which was used to make a similar mildly psychoactive bitter drink in Austronesian rituals. Cognates for *kava include Pohnpeian sa-kau; Tongan, Niue, Rapa Nui, Tuamotuan, and Rarotongan kava; Samoan and Marquesan ʻava; and Hawaiian ʻawa. In some languages, most notably Māori kawa, the cognates have come to mean "bitter", "sour", or "acrid" to the taste.[222][229][218][25]
In the Cook Islands, the reduplicated forms of kawakawa or kavakava are also applied to the unrelated members of the genus Pittosporum. And in other languages like in Futunan, compound terms like kavakava atua refer to other species belonging to the genus Piper. The reduplication of the base form is indicative of falsehood or likeness, in the sense of "false kava".[227][228]
Piper retrofractum (Javanese long pepper)
[edit]The Javanese long pepper (Piper retrofractum) is native to Island Southeast Asia from the Philippines to Sumatra. Its northern range also extends to southern China, mainland Southeast Asia, Taiwan, and the Ryukyu Islands. However it was historically only cultivated in any great extent in the islands of Java and Bali, and surrounding islands, for the spice trade. Elsewhere it is mostly grown informally in the backyards of houses. It is very similar to the Indian long pepper (Piper longum) and is used in the same way in Southeast Asian cuisine.[27][155]
Saccharum (sugarcane)
[edit]
There are two centers of domestication for sugarcane (Saccharum spp.): one for Saccharum officinarum by Papuans in New Guinea and another for Saccharum sinense by Austronesians in Taiwan and southern China. Papuans and Austronesians originally primarily used sugarcane as food for domesticated pigs. The spread of both S. officinarum and S. sinense is closely linked to the migrations of the Austronesian peoples.[231][232]

Saccharum officinarum was first domesticated in New Guinea and the islands east of the Wallace Line by Papuans, where it is the modern center of diversity. Beginning at around 6,000 BP they were selectively bred from the native Saccharum robustum. From New Guinea it spread westwards to Island Southeast Asia after contact with Austronesians, where it hybridized with Saccharum spontaneum.[232]
The second domestication center is mainland southern China and Taiwan where S. sinense (though other authors identify it as S. spontaneum) was one of the original major crops of the Austronesian peoples from at least 5,500 BP. Introduction of the sweeter S. officinarum may have gradually replaced it throughout its cultivated range in Island Southeast Asia.[18][230][233][234] From Island Southeast Asia, S. officinarum was spread eastward into Polynesia and Micronesia by Austronesian voyagers as a canoe plant by around 3,500 BP. It was also spread westward and northward by around 3,000 BP to China and India by Austronesian traders, where it further hybridized with Saccharum sinense and Saccharum barberi. From there it spread further into western Eurasia and the Mediterranean.[232][230]
The reconstructed word for "sugarcane" in Proto-Austronesian is *CəbuS or *təbuS, which became Proto-Malayo-Polynesian *təbuh, Proto-Oceanic *topu, and Proto-Polynesian *to.[25][235][236] Modern cognates include Hoanya and Bunun sibus; Rukai cobosə or tibóso; Tagalog tubó; Chamorro tupu; Murik Kayan tebu; Malay tebu; Old Sundanese teuwu; Modern Sundanese tiwu; Ansus tobu; Malmariv tov; Fijian dovu; Mele-Fila and Takuu toro; Samoan tolo; Tagula ro; Pohnpeian cheu; Tahitian to; Pukapukan, Rarotongan, and Tongan tō; Hawaiian kō; and Rapa Nui to or ta.[25][17] In Malagasy, however, the word for "sugarcane" is fary, which is instead derived from Proto-Austronesian *pajey, meaning "rice".[237]
Santalum album (sandalwood)
[edit]Santalum album is originally native to dry areas in Indonesia (Java and the Lesser Sunda Islands), the Philippines, and Western Australia, where it is found with close congeners.[238] It was introduced very early (c. 1500 BCE) into Dravidian regions of South Asia via the Austronesian maritime spice trade, along with other Austronesian domesticates like areca nut and coconuts. It first appears in archaeological records in South Asia in the southern Deccan by 1300 BCE. It became naturalized in these regions where dry sandy soils are common.[239] [26]
Solanum
[edit]
Several species of Solanum have been utilised as food and medicine by the Austronesian people. Species cultivated include Kangaroo apple (Solanum aviculare), poroporo (Solanum laciniatum), Indian nightshade (Solanum lasiocarpum), pacific tomato (Solanum repandum) and cannibals tomato (Solanum viride).
Syzygium
[edit]Trees in the genus Syzygium contain some of the most important fruit trees among Austronesian peoples. Species cultivated or harvested for their edible fruit include the Java plum (Syzygium cumini), jambos (Syzygium jambos), lubeg (Syzygium lineatum), swamp maire (Syzygium maire), mountain apple (Syzygium malaccense), lipote (Syzygium polycephaloides), and the Java apple (Syzygium samarangense), among others. Two species are also important sources of spice: the clove tree (Syzygium aromaticum) and Indonesian bay leaf (Syzygium polyanthum).[18]
Syzygium aromaticum (cloves)
[edit]Until the colonial era, cloves only grew on a few islands in the Moluccas (historically called the Spice Islands), including Bacan, Makian, Moti, Ternate, and Tidore.[240] One clove tree named Afo that experts believe is the oldest in the world on Ternate may be 350–400 years old.[241]
Cloves were first traded by the Austronesian peoples in the Austronesian maritime trade network (which began around 1500 BC, later becoming the Maritime Silk Road and part of the Spice Trade). The first notable example of modern clove farming developed on the east coast of Madagascar, and is cultivated in three separate ways, a monoculture, agricultural parklands, and agroforestry systems.[242]
Syzygium malaccense (mountain apple)
[edit]
The mountain apple (Syzygium malaccense) along with the closely related species like the water apple (Syzygium aqueum) and the Java apple (Syzygium samarangense), are native throughout Island Southeast Asia and were cultivated since prehistory. They were all carried by Austronesians into the Pacific and planted deliberately.[10][243]

They were valued primarily for their abundant edible fruits. It is also used for timber (usually for building houses) and parts of the trees are used in traditional medicine. The attractive flowers are also worn as personal hair adornments and in making leis. They were primarily propagated through cuttings by Melanesians and Polynesians. The groves of mountain apples found in the Pacific are often remnants of ancient plantings, as the seeds of the fruits are too large to be dispersed by the native birds. Related species endemic to the Pacific Islands were also utilized similarly, like Syzygium corynocarpum and Syzygium neurocalyx.[10][244][243]
There numerous names for mountain apples in Austronesian languages. In the Philippines, the terms can be reconstructed to Proto-Philippine *makúpa, with cognates including Ilocano, Aklanon, and Cebuano makúpa; and Tagalog and Bikol makópa.[17]
In Oceania, there are several cognate sets reconstructible for mountain apples and related species. Four of which are *pokaq, *marisapa, *sakau and *cay, with limited reflexes and may have originally referred to other species. The most widespread cognate set, however, can be reconstructed to Proto-Oceanic *kapika. Its cognates include Mussau kaviu; Seimat ahi, Lou keik; Maenge and Nakanai gaiva; Tami kapig; Yabem àing; Motu gavika; Bola kavika; Babatana kapika; Gela gaviga; Kwara'ae ʻafiʻo; Paamese ahie; Wayan, Niue, East Uvean, and Bauan kavika; Tongan fekika; Anutan kapika; Marquesan kehika; Mangarevan keʻika; Tahitian ʻahiʻa; Hawaiian ʻoohiaʻai; Rarotongan kaʻika; and Māori kahika. In Māori, the names have shifted to Metrosideros fulgens, which have similar-looking flowers, as Malay apples did not survive into Aotearoa.[17][222]
Tacca leontopetaloides (Polynesian arrowroot)
[edit]Polynesian arrowroot (Tacca leontopetaloides) is another ancient Austronesian root crop closely related to yams. It is originally native to Island Southeast Asia. It was introduced throughout the entire range of the Austronesian expansion, including Micronesia, Polynesia, and Madagascar. Polynesian arrowroot have been identified as among the cultivated crops in Lapita sites in Palau, dating back to 3,000 to 2,000 BP.[245] It was also introduced to Sri Lanka, southern India, and possibly also Australia through trade and contact.[246]

Polynesian arrowroot was a minor staple among Austronesians. The roots are bitter if not prepared properly, thus it was only cultivated as a secondary crop to staples like Dioscorea alata and Colocasia esculenta. Its importance increased for settlers in the Pacific Islands, where food plants were scarcer, and it was introduced to virtually all the inhabited islands. They were valued for their ability to grow in low islands and atolls, and were often the staple crops in islands with these conditions. In larger islands, they were usually allowed to grow feral and were useful only as famine food. Several cultivars have been developed in Polynesia due to the thousands of years of artificial selection. The starch extracted from the root with traditional methods can last for a very long time, and thus can be stored or traded.[245] The starch can be cooked in leaves to make starchy puddings.[247] Due to the introduction of modern crops, it is rarely cultivated today.[245]
The names for Polynesian arrowroot in Austronesian languages reflect its secondary importance as a crop. They are often reassignments from names of other starch crops, rather than specifically being for Polynesian arrowroot. Usually, the names of Polynesian arrowroot are transferred from the names of the sago palms (Metroxylon sagu), giant swamp taros (Cyrtosperma merkusii), and fermented breadfruit (Artocarpus altilis).[222]
Derivations from Proto-Malayo-Polynesian *sagu ("sago palm"), include Chamorro and Toba Batak sagu. Derivations from Proto-Polynesian *mā ("fermented breadfruit"), included Tongan māhoaʻa; Tokelauan mahoā; Anutan maoa; East Futunan māsoʻā; Samoan māsoā; and Tuvaluan māsoa. Derivations from Proto-Polynesian *bulaka (giant swamp taro) include Patpatar and Tolai pulaka. Derivations from Proto-Austronesian *biRaq (giant taro) include Äiwoo (to)piya. And finally, derivations from Proto-Oceanic Rabia (sago) include Bauan yabia; and Pileni, Rarotongan, and Hawaiian pia.[222]
Talipariti tiliaceum (sea hibiscus)
[edit]
Sea hibiscus (Talipariti tiliaceum) is a common tree in beaches in the tropical Indo-Pacific. It is widely used by Austronesian peoples for timber and fiber. It has several subspecies, two of which are endemic to the Philippines and Sulawesi, with the rest widespread throughout its range or native to large regions of Southeast Asia and the Pacific. The seeds remain viable for months after floating in the sea. However, no remains of beach hibiscus have been recovered from Polynesia prior to the Austronesian arrival, making it clear that they were introduced by Austronesian voyagers.[248][249]
The wood is soft and not very durable, so it is mostly only used for products like carvings, spears, bowls, and bracelets. However, it is also resistant to saltwater and thus can be used to make small canoes and outriggers. The wood is also preferred for fire making by friction. The fiber extracted from the bark is widely used to make cordage and for caulking. The bark is also used in the production of tempeh in Southeast Asia, and kava drinks in Polynesia. The attractive flowers are commonly made into leis in Hawaii.[250]
The terms for beach hibiscus can be reconstructed to Proto-Malayo-Polynesian *baRu, which became Proto-Oceanic *paRu and Proto-Micronesian *kili-fau. Modern cognates include Itbayaten vayu; Ilocano bagó; Kankana-ey bágo; Chamorro pagu; Tagalog balibago; Cebuano malabago or maribago; Maranao bago; Ngaju Dayak baro; Malagasy baro or varo; Malay baru; Javanese, Rembong, and Kambera waru; Sangir and Soboyo bahu; Makasar baru; Erai hau; Leti paru; Paulohi haru; Buruese fahu; Gitua paru; Mailu waru; Mota var or varu; Sye nau or vau; Anejom n-hau; Fijian vau; Tongan and Samoan fau; Rotuman, Rennellese, and Hawaiian hau; and Māori whau.[17]
In addition, there are numerous terms relating to the use of sea hibiscus for cordage and fiber in various Austronesian languages which can be traced back to Proto-Malayo-Polynesian or Proto-Austronesian, like *Calis, "rope".[17]
Thespesia populnea (Pacific rosewood)
[edit]
The Pacific rosewood (Thespesia populnea) is closely related to beach hibiscus. They are similar in appearance and grow in the same habitats, thus they are commonly confused with each other. It is also used similarly among Austronesian cultures, being one of the main sources of bast fibers for the production of cordage and wood for Austronesian outrigger ships and carving. Pacific rosewood is native to the Old World tropics. Like beach hibiscus, the seeds remain viable for months after floating in the sea but no remains of T. populnea have been recovered from Polynesia prior to the Austronesian expansion. Thus it is regarded as deliberate introductions by Austronesian settlers.[248][249]
The trees were regarded as sacred in Polynesian culture, and were commonly planted in marae sites along with trees like Ficus, Fagraea berteroana, Casuarina equisetifolia and Calophyllum inophyllum.[248][249]
The terms for Pacific rosewood can be reconstructed to Proto-Malayo-Polynesian *balu, with cognates including Itbayaten valu; Malagasy válo; Simeulue falu; Ngela valu; Arosi haru; and Lonwolwol bal.[17] Another term which extends to Oceanic is Proto-Malayo-Polynesian *banaRu with cognates including Hanunó'o banagu; Tolai banar; Patpatar banaro; Mota vanau; and Pohnpeian pana.[17]
In Eastern Polynesia, most modern names can be reconstructed back to Proto-Eastern Oceanic *milo, with cognates including Tongan, Niue, Samoan, and Hawaiian milo; Rapa Nui, Tahitan, Tuamotuan, and Māori miro; and Marquesan miʻo. In some islands, the names have shifted to refer to trees that are used similarly, like Prumnopitys ferruginea in Aotearoa and Sophora toromiro in Rapa Nui.[251]
Zingiberaceae (ginger family)
[edit]Gingers (family Zingiberaceae) were cultivated extensively by Austronesians for food, medicine, weaving materials, and for religious purposes. The most commonly cultivated species include the lengkuas (Alpinia galanga), fingerroot (Boesenbergia rotunda), turmeric (Curcuma longa), torch ginger (Etlingera elatior), and ginger (Zingiber officinale). Other species were also exploited at a smaller scale or harvested from the wild, including dwarf cardamom (Alpinia nutans), panasa cardamom (Amomum acre), white turmeric (Curcuma zedoaria), jiddo (Hornstedtia scottiana), white ginger lily (Hedychium coronarium), and bitter ginger (Zingiber zerumbet).[222][17][252]
Alpinia galanga (lengkuas)
[edit]
The lengkuas (Alpinia galanga) is native to Southeast Asia. Its original center of cultivation during the spice trade was Java, and today it is still cultivated extensively in Island Southeast Asia, most notably in the Greater Sunda Islands and the Philippines. It is valued for its use in food and for traditional medicine and is regarded as being superior to ginger. It has a pungent smell reminiscent of black pepper. The red and white cultivars are often used differently, with the red cultivars being primarily medicinal, and the white cultivars being primarily a spice.[155][253] Lengkuas is also the source of the leaves used to make nanel among the Kavalan people of Taiwan, a rolled leaf instrument used as a traditional children's toy common among Austronesian cultures.[254]
Lengkuas can be reconstructed to Proto-Western Malayo-Polynesian *laŋkuas, with cognates including Ilokano langkuás; Tagalog, Bikol, Kapampangan, Visayan, and Manobo langkáuas or langkáwas; Aklanon eangkawás; Kadazan Dusun hongkuas; Ida'an lengkuas; Ngaju Dayak langkuas; Iban engkuas; and Malay lengkuas (from which the English name is derived from). Some of the names have become generalized and are also applied to other species of Alpinia as well as for Curcuma zedoaria.[17]
Curcuma longa (turmeric)
[edit]
There is strong evidence that turmeric (Curcuma longa) as well as the related white turmeric (Curcuma zedoaria) were independently domesticated by Austronesians. Turmeric has a very widespread distribution and names that pre-date contact with India, being found among all Austronesian regions with the exception of Taiwan. However, it was seemingly originally domesticated for the production of dyes, eventually contributing to the words for "yellow" and "red" in various Austronesian languages.[255]

The plant is important in the Philippines and Indonesia as a traditional dye for clothing and food coloring. It was particularly valued for coloring food offerings to spirits as well as body painting in religious rituals or social ceremonies. It is also used as a spice, as medicine and as food. Similar uses are also found in the other islands settled by Austronesians, including Madagascar and the Comoros in East Africa. In Micronesia, it was a valuable trade item acquired from Yap. In Polynesia and Melanesia, they are primarily used as body paint in rituals or as a cosmetic. The latter regions have been isolated for centuries from the rest of Island Southeast Asia prior to European contact.[255][256]

There are two main cognate sets for C. longa and C. zedoaria (both of which produce yellow dye) in Austronesian languages. The first is reconstructed as Proto-Malayo-Polynesian *kunij which originally referred to turmeric. Its cognates include Ilocano, Kankana-ey, and Isneg kúnig; Bontoc kúnəg; Ifugao ūnig; Casiguran Dumagat kuneg; Iban and Malay kunyit; Toba Batak hunik; Javanese kunir; Sangir and Tae' kuniʻ; Uma kuni; Rembong kunis; Ngadha wuné; and Manggarai wunis. In Malagasy and Betsimisaraka, the cognates hónitra and húnitra have shifted meaning to a different plant used to make red dye. Other cognates like Ilocano kimmúnig; Uma mo-kuni, and Tae' pakuniran all mean "yellow" or "to dye something yellow".[255][17]
The other cognate set is derived from reconstructed Proto-Western-Malayo-Polynesian *temu, and originally meant C. zedoaria which was used primarily as a spice. It also sometimes shifted to ginger and other ginger-like plants used for cooking (rather than dye production). Its cognates include Kapampangan and Balinese tamu; Tagalog támo; Visayan tamangyan; Bukidnon tamohilang; Bikol tamahilan or tamaylan; Malay, Javanese, and Sasak temu; Makasarese tammu; and Malagasy tamutamu. In other Austronesian languages in East Africa, however, the other cognates mean "yellow", including Comorian Shibushi and Antemoro tamutamu; and Antambahoaka and Antankarana manamutamu.[255][17]
In Proto-Oceanic, there are two main cognate sets derived from reconstructed *aŋo and *deŋ(w)a, both are unrelated to the Proto-Malayo-Polynesian etymons. The latter probably originally applied to the dye produced from turmeric, while the former referred originally to the plant itself. Cognates include Fijian cango; and Tongan and Rennellese ango. Cognates that mean "yellow" also exist in numerous other languages in Near Oceania.[255][17]

Zingiber officinale (ginger)
[edit]Ginger (Zingiber officinale) is native to Island Southeast Asia and was probably originally domesticated by Austronesians. It is an ancient and ubiquitous crop among Austronesians, reaching all the way to Remote Oceania and Madagascar.[225][257][258] Aside from being used for cuisine, ginger appears to have significant religious and medicinal roles in early Austronesian cultures, based on the glosses it acquired. Ginger were chewed by shamans and spat out intermittently in rituals for healing, warding, and blessing ships.[17]
In Proto-Austronesian, the terms for ginger can be reconstructed to *dukduk. With cognates including Pazeh dukuduk; Thao suksuk; Tsou cucʻu; and Saaroa suusuku. This was replaced by *laqia in languages south of Taiwan.[17]
The terms for ginger beyond Taiwan can be reconstructed to Proto-Malayo-Polynesian *laqia, which became Proto-Oceanic *laqia and Proto-Central Polynesian *laya. Cognates include Ilocano, Agta, Isneg, Itawis, Kankana-ey, and Casiguran Dumagat layá; Bontoc, Ifugao and Kapampangan láya; Batad Ifugao lāya; Tagalog luya; Bikol láʻya; Visayan luyʻa; Tboli leʻiye; Kadazan Dusun hazo; Tombonuwo and Abai Sembuak layo; Ida'an Begak lejo; Basap, Long Anap Kenyah, Sangir, and Tontemboan lia; Lun Dayeh and Kelabit lieh; Berawan and Miriʼ lejeh; Narum lejieh; Kenyah (Òma Lóngh) lezó; Murik and Iban liaʻ; Kelai and Wahau Kenyah jeʻ; Segai aljoʻ; Modang lejao̯ʻ; Kiput lecih; Bintulu leza; Iban liaʻ; Dayak roiʻi; Jarai reya; Malay halia; Tialo loía; Balaesang láia; Bare'e leʻia; Tae laia or laya; Makasarese laia; Muna longhia; Bimanese rea; Manggarai, Roti, Erai, Leti, Wetan. and Lamaholot lia; and Sika and Ngadha lea; Kowiai and Kei leii. In Oceanic languages, cognates include Lou and Kairiru lei; Penchal lai; Ahus and Kurti liy; Drehet lip; Lindrow ley; Mussau and Wuvulu, Neham laia; Tanga lae; Lakalai la lahia; Gitua laea; Wedau naia; 'Āre'āre and Arosi ria; Sa'a lie; and Fijian cango laya.[17]
Zingiber zerumbet (bitter ginger)
[edit]
Bitter ginger (Zingiber zerumbet) is native to tropical Asia and Australasia. Like the ginger, was carried by Austronesian settlers all the way to Remote Oceania during prehistoric times. Thus it is likely that it was originally domesticated by Austronesians. Remains of bitter ginger have also been identified from the Kuk Swamp archaeological site in New Guinea at the Phase 1 layers dated to 10,220 to 9,910 BP. However, whether they were cultivated or simply exploited from the wild is unknown.[259][229]
Bitter ginger is primarily used for traditional medicine. It also has mildly psychoactive properties when consumed, and thus had ritual importance among early Austronesian cultures. According to Lynch (2002), terms for bitter ginger in the sense of "bitter root" or "potent root [used as fish poison]", reconstructed as Proto-Oceanic *kawaRi, is believed to have been transferred to the kava (Piper methysticum), which has similar properties and is also bitter-tasting, when Austronesians of the Lapita culture first encountered it among the indigenous non-Austronesian peoples in Melanesia.[222][229][218]
Some reflexes of it still refer to bitter ginger, including Sissano (una)kaw; Gapapaiwa kaware; Tikopia, Anutan, and Wallisian kava-pui; Samoan ʻava-pui; Tahitian ava-puhi; and Hawaiian ʻawa-puhi.[222][229][218] Other reflexes also refer to other members of the genus Piper, to fish poison, or as words to describe bitter, sour, or acrid flavors.[25]
In non-Oceanic languages, terms for bitter ginger can be reconstructed to Proto-Western Malayo-Polynesian *lampuyaŋ, with cognates including Cebuano and Ngaju Dayak lampuyang; Javanese lempuyang; and Malay lempoyang.[17]
Animals
[edit]Domesticated, semi-domesticated, and commensal animals carried by Austronesian voyagers include the following:
Bubalus bubalis (water buffalo)
[edit]
Water buffaloes are essential work animals in Austronesian paddy field agriculture and were carried along with rice to Island Southeast Asia from mainland Asia. Early introductions were specifically of the swamp-type water buffaloes (like the carabao), although they are increasingly being replaced by river-type water buffaloes imported from South Asia in recent times.[260]
The earliest remains of water buffaloes in Island Southeast Asia with signs of domestication comes from multiple fragmentary skeletal remains recovered from the upper layers of the Neolithic Nagsabaran site, part of the Lal-lo and Gattaran Shell Middens (~2200 BCE to 400 CE) of northern Luzon. Most of the remains consisted of skull fragments, almost all of which have cut marks indicating they were butchered. The remains are associated with red slipped pottery, spindle whorls, stone adzes, and jade bracelets; which have strong affinities to similar artifacts from Neolithic Austronesian archeological sites in Taiwan. Based on the radiocarbon date of the layer in which the oldest fragments were found, water buffaloes were first introduced to the Philippines by at least 500 BCE.[261][262]
The English term "carabao" is borrowed from the Spanish word carabao, which is derived from Eastern Visayan (likely Waray) karabàw.[263][264] Cognates include Cebuano kábaw, Tagalog kalabáw, Kavalan qabaw, Minangkabau kabau, Malay/Indonesian kerbau, and Javanese kebo.[265] These Austronesian terms appear to be loanwords from the Austroasiatic languages and likely derives from a secondary pre-colonial introduction of water buffaloes into Island Southeast Asia via western Indonesia.[266][267]
However, it is also clear that Austronesians already had ancient terms for the carabao, reconstructed as Proto-Austronesian *qaNuaŋ. Cognates include Papora loan, Thao qnuwan, Siraya luang, Rukai nwange, Ilocano nuang, Tagalog anwang or anowang, Kankanaey nuang, Isneg nuang, Itawis nwang, Bontoc nuwang, Ifugao nuwang, and Aklanon anwang. Cognates survive into Sulawesi, but the terms there apply to the related anoa. Similarly, Hanunó'o anwang also refers to the tamaraw of Mindoro, rather than the carabao. These terms spread southwards from Taiwan, indicating that domesticated carabaos were carried partially into the Philippines during the Austronesian expansion, but didn't move further south into the rest of Island Southeast Asia until the second introduction from Mainland Southeast Asia. The reason for this is unknown.[266][264][268]
Other native names for carabaos include damulag in Tagalog, Bikol, and Kapampangan; dueg in Pangasinan; and pagad in Ivatan.[269]
Bos javanicus domesticus (Bali cattle)
[edit]Bali cattle were domesticated in Bali from the wild banteng around 3500 BCE.
Canis lupus familiaris (dog)
[edit]
Dogs were primarily valued for their social functions in various Austronesian cultures, acting as companions and pets. They were also trained to be hunting or guard dogs. Ornaments made from dog fur, teeth, and bones are found in archaeological sites throughout Austronesia. These could be traded as commodities, along with dog pups. Dogs were also sometimes eaten, but this varies by culture, with most groups refusing to eat dogs, while in others they were apparently a main food source.[270][271][272][273][274]

In 2020, an mDNA study of ancient dog fossils from the Yellow River and Yangtze River basins of southern China showed that most of the ancient dogs fell within haplogroup A1b, as do the Australian dingoes and the pre-colonial dogs of the Pacific, but in low frequency in China today. The specimen from the Tianluoshan archaeological site (Hemudu culture, pre-Austronesians), Zhejiang province dates to 7,000 YBP and is basal to the entire lineage. The dogs belonging to this haplogroup were once widely distributed in southern China, then dispersed through Southeast Asia into New Guinea and Oceania, but were replaced in China 2,000 YBP by dogs of other lineages.[275]
The origins of the dog (Canis lupus familiaris) population in Island Southeast Asia, Australia, and New Guinea are contentious, with various authors proposing origins from either Mainland Southeast Asia, Taiwan, or both at different times. These introduction events have been linked to the origin of the Australian dingoes and the New Guinea singing dogs, both of which are clearly descended from domesticated dogs. The specifics of which population they are derived from, who introduced them, and whether they come from a common ancestor, however, still do not have a consensus.[270][276][271]
Regardless, most authors agree that there were at least two introduction events. One arriving with Paleolithic maritime hunter-gatherers by at least 10,000 to 5,000 BP, and another arriving with later Neolithic seafaring migrations of farming and trading Austronesian peoples (via Taiwan) by at least by at least 5,000 BP. The Neolithic dogs are differentiated from previous populations in having the ability to digest starch, indicating that they accompanied humans who cultivated cereal crops.[270][276] The Neolithic introductions are believed to have partially replaced the original introductions and became the ancestors of the modern village dogs of Southeast Asia.[276][273] Austronesian dogs like the Taiwan dog were deeply valued as hunting companions (particularly for wild boar).[277][278][279] In the Philippines, dog remains have been found buried near or beside human graves in archaeological sites.[280]


The oldest archaeological remains of dogs in Island Southeast Asia and Oceania is a dog burial in Timor and dingo remains in Australia, both of which are dated to around 3,500 BP. The former are believed to have been part of the second wave and the latter from the first wave.[272][281][274]
From Island Southeast Asia, they were carried by Austronesian voyagers into Near Oceania.[271][272][274] However, unlike in Island Southeast Asia, dogs lost their economic importance as hunting animals among Austronesians that reached the smaller islands in Melanesia and Polynesia, which had no populations of wild boar or other large mammals that could be hunted. They became a competitor for limited food resources and thus were themselves eaten. The Austronesian domesticated dogs originally carried by the Lapita Culture migrations were eaten to extinction in many islands since ancient times.[279][282]
Thus Austronesian dogs were "lost" during the early colonization of Near Oceania. This caused a marked discontinuity in the genes of domesticated dogs, as well as the terms for "dog", among Austronesians in the Pacific Islands and Island Melanesia, in comparison to other Austronesian regions in Island Southeast Asia.[279][282]
However, dogs were later reintroduced from neighboring Papuan groups and were subsequently carried eastward into Polynesia by post-Lapita Austronesian migrations, reaching as far as Hawaii and Aotearoa. Though these dogs were treated as food animals, rather than hunting companions.[279][282] Genetic studies have confirmed this, showing that Polynesian dogs are descended from the first wave of dog introductions and are not related to the dogs originating from Taiwan and the Philippines.[271][272][274] Dogs were historically eaten in Tahiti and other islands of Polynesia, including Hawaii[283][284] at the time of first European contact. James Cook, when first visiting Tahiti in 1769, recorded in his journal, "few were there of us but what allow'd that a South Sea Dog was next to an English Lamb, one thing in their favour is that they live entirely upon Vegetables".[285] Calvin Schwabe reported in 1979 that dog was widely eaten in Hawaii and considered to be of higher quality than pork or chicken. When Hawaiians first encountered early British and American explorers, they were at a loss to explain the visitors' attitudes about dog meat. The Hawaiians raised both dogs and pigs as pets and for food. They could not understand why their British and American visitors only found the pig suitable for consumption.[286] This practice seems to have died out, along with the native Hawaiian breed of dog, the unique Hawaiian Poi Dog, which was primarily used for this purpose.[287]
On certain Pacific islands, dogs died out after original settlement or were never brought by the settlers in the first place, notably: the Mariana Islands,[288] Palau,[289] Marshall Islands,[290] Gilbert Islands,[290] New Caledonia,[291] Vanuatu,[291][292] Tonga,[292] Marquesas,[292] Mangaia in the Cook Islands, Rapa Iti in French Polynesia, Easter Island,[292] Chatham Islands,[293] and Pitcairn Island (settled by the Bounty mutineers, who killed off their dogs in order to escape discovery by passing ships).[294]
Dogs were not introduced to Madagascar by Austronesians. A genetic study by Ardalan et al. (2015) revealed that the dog population in Madagascar were all derived solely from African dog populations and did not come from Southeast Asian dog populations. This aberrant origin is also reflected in the Malagasy languages, where the terms for "dog" originate entirely from African Bantu languages.[295][296] Given the inferred importance of dogs to Austronesian voyagers, the authors proposed that the Austronesian settlers in Madagascar may have initially brought dogs, but they either died or were used as food sources during the journey. Another possibility is that the limited initial number of Austronesian dogs may have simply resulted in their genes getting swamped by the influx of a far larger population of dogs from Africa.[295][296]
Gallus gallus (chicken)
[edit]
Junglefowl were one of the three main animals (along with domesticated pigs and dogs) carried by early Austronesian peoples from Island Southeast Asia in their voyages to the islands of Oceania.[298][297]
Sus scrofa domesticus (pig)
[edit]Pigs were one of the three main animals (along with domesticated chickens and dogs) carried by early Austronesian peoples from Island Southeast Asia in their voyages to the islands of Oceania.[298]
Rodentia (rodents)
[edit]The following rodent species are common in mainland Southeast Asia, but are restricted to areas of wet rice cultivation in western Indonesia (Sumatra and Java).[299]
The following two rodents are native to South Asia and also present in western Indonesian rice fields, so their presence in Island Southeast Asia cannot easily be explained by Austronesian expansions, but perhaps instead by the Indian Ocean trade.[299]
Mus terricolor (also known as Mus dunni)
[edit]Indigenous to northwestern India
Rattus nitidus (indigenous to Nepal)
[edit]This rat originated on the island of Flores in Indonesia.[300] Polynesians accidentally or deliberately introduced it to the islands they settled. This rat has been implicated in many of the extinctions of native birds and insects in the Pacific; these species had evolved in the absence of mammals and could not cope with predation by the rat.
See also
[edit]- Agriculture in Papua New Guinea
- Catamaran
- Crab claw sail
- Domesticated plants of Mesoamerica
- Genomics of domestication
- History of agriculture
- List of food origins
- Outrigger canoe
- Proto-Oceanic language § Lexicon
- Tanja sail
References
[edit]- ^ Chambers, Geoff (2013). "Genetics and the Origins of the Polynesians". eLS. John Wiley & Sons, Inc. doi:10.1002/9780470015902.a0020808.pub2. ISBN 978-0470016176.
- ^ Bellwood, Peter (2004). "The origins and dispersals of agricultural communities in Southeast Asia" (PDF). In Glover, Ian; Bellwood, Peter (eds.). Southeast Asia: From Prehistory to History. RoutledgeCurzon. pp. 21–40. ISBN 9780415297776.
- ^ Reilly, Kevin (2012). Volume I: Prehistory to 1450. The Human Journey: A Concise Introduction to World History. Vol. 1. Rowman & Littlefield Publishers, Inc. pp. 207–209. ISBN 9781442213869.
- ^ Liu, Li; Chen, Xingcan (2012). "Emergence of social inequality – The middle Neolithic (5000–3000 BC)". The Archaeology of China: From the Late Paleolithic to the Early Bronze Age. Cambridge World Archaeology. Cambridge University Press. p. 204. doi:10.1017/CBO9781139015301.007. ISBN 9780521644327.
- ^ Bourke, Richard Michael (2009). "History of agriculture in Papua New Guinea" (PDF). In Bourke, Richard Michael; Harwood, Tracy (eds.). Food and Agriculture in Papua New Guinea. ANU E Press. pp. 10–26. doi:10.22459/FAPNG.08.2009. ISBN 9781921536618.
- ^ Jump up to: a b c d e Denham, Tim (October 2011). "Early Agriculture and Plant Domestication in New Guinea and Island Southeast Asia". Current Anthropology. 52 (S4): S379–S395. doi:10.1086/658682. hdl:1885/75070. S2CID 36818517.
- ^ W. Arthur Whistler (1991). "Islands, Plants, and Polynesians—An Introduction to Polynesian Ethnobotany." National Tropical Botanical Garden c/o Botany Department University of Hawaii Honolulu, Hawaii 96822. Retrieved March 2022.
- ^ Kitalong, Ann Hillmann; Ballick, MichaelJ.; Rehuher, Faustina; Besebes, Meked; Hanser, Sholeh; Soaladaob, Kiblas; Ngirchobong, Gemma; Wasisang, Flora; Law, Wayne; Lee, Roberta; Tadeo, Van Ray; Kitalong, Clarence; Kitalong, Christopher (2011). "Plants, people and culture in the villages of Oikull and Ibobang, Republic of Palau". In Liston, Jolie; Clark, Geoffrey; Alexander, Dwight (eds.). Pacific Island Heritage: Archaeology, Identity & Community. Terra Australis. Vol. 35. ANU E Press. pp. 63–84. ISBN 9781921862489.
- ^ Theroux, Paul (December 2002). "The Hawaiians". National Geographic. 202 (6): 2–41.
- ^ Jump up to: a b c Blench, Roger (2004). "Fruits and arboriculture in the Indo-Pacific region". Bulletin of the Indo-Pacific Prehistory Association. 24 (The Taipei Papers (Volume 2)): 31–50.
- ^ Simanjuntak, Truman (2006). "Advancement of Research on the Austronesian in Sulawesi". In Simanjuntak, Truman; Hisyam, M.; Prasetyo, Bagyo; Nastiti, Titi Surti (eds.). Archaeology: Indonesian Perspective : R.P. Soejono's Festschrift. Indonesian Institute of Sciences (LIPI). pp. 223–231. ISBN 9789792624991.
- ^ Hasanuddin (2018). "Prehistoric sites in Kabupaten Enrekang, South Sulawesi". In O'Connor, Sue; Bulbeck, David; Meyer, Juliet (eds.). The Archaeology of Sulawesi: Current Research on the Pleistocene to the Historic Period. terra australis. Vol. 48. ANU Press. pp. 171–189. doi:10.22459/TA48.11.2018.11. ISBN 9781760462574. S2CID 134786275.
- ^ Larrue, Sébastien; Meyer, Jean-Yves; Chiron, Thomas (2010). "Anthropogenic Vegetation Contributions to Polynesia's Social Heritage: The Legacy of Candlenut Tree (Aleurites moluccana) Forests and Bamboo (Schizostachyum glaucifolium) Groves on the Island of Tahiti". Economic Botany. 64 (4): 329–339. doi:10.1007/s12231-010-9130-3. S2CID 28192073.
- ^ Weisler, Marshall I.; Mendes, Walter P.; Hua, Quan (2015). "A prehistoric quarry/habitation site on Moloka'i and a discussion of an anomalous early date on the Polynesian introduced candlenut (kukui, Aleurites moluccana)". Journal of Pacific Archaeology. 6 (1): 37–57.
- ^ Kirch, Patrick V. (1989). "Second Millennium B.C. Arboriculture in Melanesia: Archaeological Evidence from the Mussau Islands". Economic Botany. 43 (2): 225–240. doi:10.1007/BF02859865. S2CID 29664192.
- ^ Elevitch, Craig R.; Manner, Harley I. (2006). "Aleurites moluccana (kukui)". In Elevitch, Craig R. (ed.). Traditional Trees of Pacific Islands: Their Culture, Environment, and Use. Permanent Agricultural Resources (PAR). pp. 41–56. ISBN 9780970254450.
- ^ Jump up to: a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at Blust, Robert; Trussel, Stephen (2013). "The Austronesian Comparative Dictionary: A Work in Progress". Oceanic Linguistics. 52 (2): 493–523. doi:10.1353/ol.2013.0016. S2CID 146739541.
- ^ Jump up to: a b c d e f g h i j k l Osmond, Meredith (1998). "Horticultural practices" (PDF). In Ross, Malcolm; Pawley, Andrew; Osmond, Meredith (eds.). The lexicon of Proto Oceanic : The culture and environment of ancestral Oceanic society. Vol. 1. Pacific Linguistics. pp. 115–142. doi:10.15144/PL-C152.115.
- ^ Nauheimer, Lars; Boyce, Peter C.; Renner, Susanne S. (April 2012). "Giant taro and its relatives: A phylogeny of the large genus Alocasia (Araceae) sheds light on Miocene floristic exchange in the Malesian region". Molecular Phylogenetics and Evolution. 63 (1): 43–51. doi:10.1016/j.ympev.2011.12.011. PMID 22209857.
- ^ Manner, Harley I. (2006). "Farm and Forestry Production Marketing Profile for Giant Tao (Alocasia macrorrhiza)" (PDF). In Elevitch, Craig R. (ed.). Specialty Crops for Pacific Island Agroforestry. Permanent Agricultural Resource (PAR).
- ^ Jump up to: a b c Matthews, Peter J. (1995). "Aroids and the Austronesians". Tropics. 4 (2/3): 105–126. doi:10.3759/tropics.4.105. S2CID 84341068.
- ^ Сантоса, Эди; Лиан, Чун Лан; Сугияма, Нобуо; Мисра, Радж Шекхар; Бункоркаев, Патчария; Таномчит, Канокван; Чан, Цзэнь-Ю (28 июня 2017 г.). «Популяционная структура ямса слоновьей лапки (Amorphophallus paeoniifolius (Dennst.) Nicolson) в Азии» . ПЛОС Один . 12 (6): e0180000. Бибкод : 2017PLoSO..1280000S . дои : 10.1371/journal.pone.0180000 . ПМК 5489206 . ПМИД 28658282 .
- ^ Jump up to: а б с МакКлатчи, Уилл К. (2012). «Дикие пищевые растения Отдаленной Океании» . Журнал Польского ботанического общества . 81 (4): 371–380. дои : 10.5586/asbp.2012.034 .
- ^ Хоррокс, М; Ньювудт, МК; Кинастон, Р; Бакли, Х; Бедфорд, С. (13 ноября 2013 г.). «Инфракрасный анализ микрофоссилий и преобразования Фурье человеческого зубного камня лапита и постлапита из Вануату, юго-западная часть Тихого океана». Журнал Королевского общества Новой Зеландии . 44 (1): 17–33. дои : 10.1080/03036758.2013.842177 . S2CID 128880569 .
- ^ Jump up to: а б с д и ж г час я дж к Маклин, Мервин (2014). Музыка, лапита и проблема полинезийского происхождения . Полинезийское происхождение. ISBN 9780473288730 .
- ^ Jump up to: а б с д и ж г час я Зумбройх, Томас Дж. (2007–2008 гг.). «Происхождение и распространение жевания бетеля: синтез данных из Южной Азии, Юго-Восточной Азии и за ее пределами» . Электронный журнал индийской медицины . 1 : 87–140.
- ^ Jump up to: а б Лим, ТК (2012). Съедобные лекарственные и нелекарственные растения . Спрингер. ISBN 9789048186617 .
- ^ Орва, К.; Мутуа, А.; Киндт, Р.; Джамнадасс, Р.; Энтони, С. (2009). Artocarpus гетерофиллус (PDF) . База данных Agroforestree: справочник по деревьям и руководство по выбору, версия 4.0. Мировое агролесомелиорирование.
- ^ Моника М., де Алмейда-Лопес; де Соуза, Келлина О.; Эбенезер, де Оливейра-Сильва (2018). «Цемпедак — Artocarpus champeden ». В Родригесе, Суэли; де Оливейра-Сильва, Эбенезер; де Брито, Эди Соуза (ред.). Экзотические фрукты: Справочное руководство . Академическая пресса. стр. 121–127. дои : 10.1016/B978-0-12-803138-4.00017-4 . ISBN 9780128031384 .
- ^ Орва, К.; Мутуа, А.; Киндт, Р.; Джамнадасс, Р.; Энтони, С. (2009). Artocarpus lakoocha (PDF) . База данных Agroforestree: справочник по деревьям и руководство по выбору, версия 4.0. Мировое агролесомелиорирование.
- ^ Орва, К.; Мутуа, А.; Киндт, Р.; Джамнадасс, Р.; Энтони, С. (2009). Artocarpus mariannensis (PDF) . База данных Agroforestree: справочник по деревьям и руководство по выбору, версия 4.0. Мировое агролесомелиорирование.
- ^ Абу Бакар, Фазлин Иззани; Абу Бакар, Мохд Фадзелли (2018). «Тарап — Artocarpus odoratissimus ». В Родригесе, Суэли; де Оливейра-Сильва, Эбенезер; де Брито, Эди Соуза (ред.). Экзотические фрукты: Справочное руководство . Академическая пресса. стр. 413–418. дои : 10.1016/B978-0-12-803138-4.00041-1 . ISBN 9780128031384 .
- ^ Мадулид, Доминго А. (6 августа 2011 г.). « Артокарпус трекулианус » . Филиппинские местные растения . Филиппинская звезда . Проверено 14 января 2019 г.
- ^ Jump up to: а б Рагон, Дайан (2006). « Artocarpus camansi (хлебный орех)» (PDF) . В Элевиче, Крейг Р. (ред.). Профили видов для агролесомелиорации островов Тихого океана . Постоянные сельскохозяйственные ресурсы (PAR).
- ^ Зерега, Нью-Джерси; Рагон Д. и Мотли Т. Дж. (2004). «Сложное происхождение плодов хлебного дерева ( Artocarpus altilis , Moraceae): последствия для миграции людей в Океании». Американский журнал ботаники . 91 (5): 760–766. дои : 10.3732/ajb.91.5.760 . ПМИД 21653430 .
- ^ Матисоо-Смит, Элизабет А. (3 ноября 2015 г.). «Отслеживание австронезийской экспансии в Тихий океан через завод по производству бумажной шелковицы» . Труды Национальной академии наук . 112 (44): 13432–13433. Бибкод : 2015PNAS..11213432M . дои : 10.1073/pnas.1518576112 . ПМК 4640783 . ПМИД 26499243 .
- ^ Jump up to: а б «*Кулу» . Те Мара Рео: Языковой сад . Семейный фонд Бентонов . Проверено 15 января 2019 г.
- ^ Бласт, Роберт (1989). «Австронезийская этимология: IV». Океаническая лингвистика . 28 (2): 111–180. дои : 10.2307/3623057 . JSTOR 3623057 .
- ^ Халл, Джеффри (2000). «Историческая фонология Тетума» . Исследования языков и культур Восточного Тимора . 3 (2000): 158–212.
- ^ Jump up to: а б Бленч, Роджер (2008). «История фруктов на материковой части Юго-Восточной Азии» (PDF) . В Осаде Тошики; Уэсуги, Акинори (ред.). Периодический доклад 4: Лингвистика, археология и прошлое человечества . Индский проект. стр. 115–137. ISBN 9784902325331 .
- ^ Jump up to: а б "*Кофе" . Те Мара Рео: Языковой сад . Семейный фонд Бентонов . Проверено 15 января 2019 г.
- ^ Беллвуд, Питер (2009). «Археология и происхождение языковых семей» . В Бентли Р. Александр; Машнер, Герберт Д.Г.; Чиппиндейл, Кристофер (ред.). Справочник археологических теорий . АльтаМира Пресс. стр. 225–244. ISBN 9780759100336 .
- ^ Фаулер, Синтия (2005). «Почему кукуруза является священным растением? Социальная история и аграрные изменения в Сумбе» . Журнал этнобиологии . 25 (1): 39–57. doi : 10.2993/0278-0771(2005)25[39:WIMASP]2.0.CO;2 . S2CID 37057768 .
- ^ Jump up to: а б с Чанг, Чи-Шань; Лю, Сяо-Лэй; Монкада, Химена; Зееленфройнд, Андреа; Зееленфройнд, Даниэла; Чунг, Го-Фанг (2015). «Целостная картина австронезийских миграций, выявленная с помощью филогеографии тихоокеанской бумажной шелковицы» . Труды Национальной академии наук . 112 (44): 13537–13542. Бибкод : 2015PNAS..11213537C . дои : 10.1073/pnas.1503205112 . ПМЦ 4640734 . ПМИД 26438853 .
- ^ Jump up to: а б с Зееленфройнд, Даниэла; Кларк, Эндрю С.; Оянедель-Джаверини, Нария Фактина; Пинья-Муньос, Рикардо; Лобос, Серхио; Матису-Смит, Лиза; Зееленфройнд, А. (сентябрь 2010 г.). «Бумажная шелковица ( Broussonetia papyrifera ) как комменсальная модель передвижения человека в Океании: антропологические, ботанические и генетические соображения». Новозеландский журнал ботаники . 48 (3–4): 231–247. Бибкод : 2010NZJB...48..231S . дои : 10.1080/0028825X.2010.520323 . hdl : 10533/143279 . S2CID 83993320 .
- ^ Гонсалес-Лорка, Дж.; Ривера-Хутинель, А.; Монкада, X.; Лобос, С.; Зееленфройнд, Д.; Зееленфройнд, А. (2 апреля 2015 г.). «Древняя и современная интродукция Broussonetia papyrifera ([L.] Vent.; Moraceae) в Тихий океан: генетические, географические и исторические свидетельства» . Новозеландский журнал ботаники . 53 (2): 75–89. Бибкод : 2015NZJB...53...75G . дои : 10.1080/0028825X.2015.1010546 . S2CID 54664583 .
- ^ Ли, Давэй; Ван, Вэй; Тянь, Фэн; Ляо, Вэй; Бэ, Кристофер Дж. (декабрь 2014 г.). «Старейшая выбивалка для ткани из коры на юге Китая (Динмо, бассейн Бубин, Гуанси)». Четвертичный интернационал . 354 : 184–189. Бибкод : 2014QuInt.354..184L . дои : 10.1016/j.quaint.2014.06.062 .
- ^ "*Ауте" . Те Мара Рео: Языковой сад . Семейный фонд Бентонов . Проверено 15 января 2019 г.
- ^ Jump up to: а б с Орва, К.; Мутуа, А.; Киндт, Р.; Энтони, С. (2009). « Калофиллум инофиллум » (PDF) . База данных Agroforestree: справочник по деревьям и руководство по выбору, версия 4.0 . Мировое агролесомелиорирование.
- ^ Jump up to: а б с д Дэймон, Фредерик Х. (2016). «История калофиллума : от экологических к социальным фактам» . Деревья, узлы и опоры: экологические знания на северо-востоке кольца Кула . Книги Бергана. стр. 180–246. ISBN 9781785332333 .
- ^ Jump up to: а б Двек, AC; Медоуз, Т. (2002). «Таману ( Calophyllum inophyllum ) – африканская, азиатская, полинезийская и тихоокеанская панацея» (PDF) . Международный журнал косметической науки . 24 (6): 341–348. дои : 10.1046/j.1467-2494.2002.00160.x . ПМИД 18494888 . S2CID 21847865 . Архивировано из оригинала (PDF) 19 января 2019 года . Проверено 17 января 2019 г.
- ^ Хальперн, Жорж М.; Веверка, Питер (2002). Тропа исцеления: эфирные масла Мадагаскара . Basic Health Publications, Inc., стр. 43–50. ISBN 9781591200161 .
- ^ Аллен, Джеймс А. (2002). « Калофиллум инофиллум » (PDF) . В Воззо, Дж. А. (ред.). Руководство по семенам тропических деревьев . Справочник по сельскому хозяйству. Том. 721. Лесная служба Министерства сельского хозяйства США. стр. 357–359.
- ^ Зорк, Р. Дэвид (1995). «Глоссарий австронезийских реконструкций» . В Трайоне, Даррелл Т.; Росс, Малкольм Д.; Граймс, Чарльз Э.; Клайнс, Адриан; Аделаар, К.А. (ред.). Сравнительный австронезийский словарь: Введение в австронезийские исследования . Тенденции в лингвистической документации. Том. 10. Мутон де Грюйтер. ISBN 9783110884012 .
- ^ « Cananga odorata (Lam.) Hook.f. & Thomson» . Растения мира онлайн . Королевский ботанический сад, Кью . Проверено 11 февраля 2019 г.
- ^ Уистлер, В. Артур (2009). «Этноботаника». В Гиллеспи, Розмари; Клэг, Дэвид (ред.). Энциклопедия островов . Беркли: Издательство Калифорнийского университета. ISBN 978-0-520-25649-1 .
- ^ Бленч, РМ (2005). «Фрукты и лесоводство в Индо-Тихоокеанском регионе» . Бюллетень Индо-Тихоокеанской ассоциации предыстории . 24 :31–50.
- ^ «Цитрус (цитрусовые) и Фортунелла (кумкват)» . Профили видов для агролесомелиорации островов Тихого океана. Апрель 2006. с. 4.
- ^ Росс, Малкольм Д .; Эндрю Поли ; Мередит Осмонд (2008). Лексикон протоокеанского языка: Том 3, Растения (PDF) . Канберра: Pacific Linguistics 599. стр. 338–9. ISBN 9780858835894 . OCLC 1075805883 .
- ^ Jump up to: а б с д и ж Ганн, Би Ф.; Бодуэн, Люк; Олсен, Кеннет М.; Ингварссон, Пер К. (22 июня 2011 г.). «Независимое происхождение культивируемого кокоса ( Cocos nucifera L.) в тропиках Старого Света» . ПЛОС ОДИН . 6 (6): e21143. Бибкод : 2011PLoSO...621143G . дои : 10.1371/journal.pone.0021143 . ПМК 3120816 . ПМИД 21731660 .
- ^ Лутц, Диана (24 июня 2011 г.). «Расшифрована глубокая история кокосов» . Источник . Проверено 10 января 2019 г.
- ^ Jump up to: а б Брауэрс, Лукас (1 августа 2011 г.). «Кокосы: не местные, но тем не менее вполне домашние» . Научный американец . Проверено 10 января 2019 г.
- ^ Лалит, Перера; Сурия ACN, Перера; Чампа К., Бандаранаяке; Хью К., Харрис (2009). "Кокос" . В Вольманн, Иоганн; Раджчан, Иштван (ред.). Масличные культуры . Спрингер. стр. 370–372. ISBN 9780387775937 . Архивировано из оригинала 20 апреля 2016 года.
- ^ Чан, Эдвард; Элевич, Крейг Р. (2006). « Cocos nucifera (кокос)» (PDF) . В Элевиче, Крейг Р. (ред.). Профили видов для агролесомелиорации островов Тихого океана . Постоянные сельскохозяйственные ресурсы (PAR).
- ^ Jump up to: а б Бодуэн, Люк; Лебрен, Патрисия (26 июля 2008 г.). «Исследования ДНК кокоса ( Cocos nucifera L.) подтверждают гипотезу древней миграции австронезийцев из Юго-Восточной Азии в Америку». Генетические ресурсы и эволюция сельскохозяйственных культур . 56 (2): 257–262. дои : 10.1007/s10722-008-9362-6 . S2CID 19529408 .
- ^ Jump up to: а б с Кроутер, Элисон; Лукас, Лейлани; Хелм, Ричард; Хортон, Марк; Шиптон, Кери; Райт, Генри Т.; Уолшоу, Сара; Павлович, Мэтью; Радимилахи, Шанталь; Дука, Катерина; Пикорнелл-Гелаберт, Льоренс; Фуллер, Дориан К.; Бойвен, Николь Л. (14 июня 2016 г.). «Древние посевы являются первым археологическим свидетельством австронезийской экспансии на запад» . Труды Национальной академии наук . 113 (24): 6635–6640. Бибкод : 2016PNAS..113.6635C . дои : 10.1073/pnas.1522714113 . ПМЦ 4914162 . ПМИД 27247383 .
- ^ Бласт, Роберт (1988). «Австронезийская родина: лингвистическая перспектива» (PDF) . Азиатские перспективы . 26 (1): 45–67.
- ^ Элберт, Сэмюэл Х. (1964). «Гавайские рефлексы прото-малайо-полинезийских и прото-полинезийских реконструированных форм» . Журнал Полинезийского общества . 73 (4): 399–410. Архивировано из оригинала 27 сентября 2021 года . Проверено 15 января 2019 г.
- ^ Бласт, Роберт (2014). «Некоторые недавние предложения относительно классификации австронезийских языков» . Океаническая лингвистика . 53 (2): 300–391. дои : 10.1353/ол.2014.0025 . S2CID 144931249 .
- ^ Ахуджа, Южная Каролина; Ахуджа, Сиддхарт; Ахуджа, Ума (2014). «Кокос – история, использование и фольклор» (PDF) . Азиатская агроистория . 18 (3): 221–248. Архивировано из оригинала (PDF) 10 августа 2017 года.
- ^ Уорд, Р.Г.; Брукфилд, М. (1992). «Специальная бумага: распространение кокоса: он плавал или его увезли в Панаму?». Журнал биогеографии . 19 (5): 467–480. Бибкод : 1992JBiog..19..467W . дои : 10.2307/2845766 . JSTOR 2845766 .
- ^ Лю, Ли; Дункан, Нил А.; Чен, Синцань; Цуй, Цзяньсинь (сентябрь 2019 г.). «Эксплуатация слез Иова в палеолите и неолите Китая: методологические проблемы и решения» . Четвертичный интернационал . 529 : 25–37. Бибкод : 2019QuInt.529...25L . дои : 10.1016/j.quaint.2018.11.019 . ISSN 1040-6182 – через Science Direct.
- ^ Лю, Ли; Дункан, Нил А; Чен, Синцань; Лю, Госян; Чжао, Хао (11 августа 2015 г.). «Одомашнивание, выращивание и добыча растений первыми фермерами раннего неолита Северо-Восточного Китая: данные микроботанических остатков» . Голоцен . 25 (12): 1965–1978. Бибкод : 2015Holoc..25.1965L . дои : 10.1177/0959683615596830 . ISSN 0959-6836 .
- ^ Крайке, CM; Ван Эк, HJ; Лебот, В. (20 мая 2004 г.). «Генетическое разнообразие таро, Colocasia esculenta (L.) Schott, в Юго-Восточной Азии и Тихоокеанском регионе». Теоретическая и прикладная генетика . 109 (4): 761–768. дои : 10.1007/s00122-004-1691-z . ПМИД 15156282 . S2CID 21917510 .
- ^ Лебот, Винсент (2009). Тропические корнеплоды и клубнеплоды: маниока, сладкий картофель, ямс и ароиды . Наука растениеводства в садоводстве. Том. 17. КАБИ. стр. 279–280. ISBN 9781845936211 .
- ^ Jump up to: а б с Председатель, Х.; Траоре, RE; Дюваль, МФ; Риваллан, Р.; Мукерджи, А.; Абоагье, LM; Ван Ренсбург, WJ; Андрианавалона, В.; Пиньейру де Карвалью, MAA; Саборио, Ф.; Шри Прана, М.; Комолонг, Б.; Лавач, Ф.; Лебот, В.; Чан, Цзэнь-Ю (17 июня 2016 г.). «Генетическая диверсификация и распространение таро ( Colocasia esculenta (L.) Schott)» . ПЛОС ОДИН . 11 (6): e0157712. Бибкод : 2016PLoSO..1157712C . дои : 10.1371/journal.pone.0157712 . ПМЦ 4912093 . ПМИД 27314588 .
- ^ Мэтьюз, Питер Дж.; Нгуен, Ван Дзу; Танданг, Дэниел; Агу, Э. Марибель; Мадулид, Доминго А. (2015). «Таксономия и этноботаника Colocasia esculenta и C. formosana (Araceae): значение для эволюции, естественного ареала и одомашнивания таро» . Ароидеана . 38Е (1). Архивировано из оригинала 19 января 2019 года . Проверено 18 января 2019 г.
- ^ Мэтьюз, П.Дж.; Агу, ЭМГ; Танданг, DN; Мадулид, Д.А. (2012). «Этноботаника и экология дикого таро ( Colocasia esculenta ) на Филиппинах: последствия одомашнивания и расселения» (PDF) . В Сприггсе, Мэтью; Аддисон, Дэвид; Мэтьюз, Питер Дж. (ред.). Орошаемый таро ( Colocasia esculenta ) в Индо-Тихоокеанском регионе: биологические, социальные и исторические перспективы . Сенри-этнологические исследования (SES). Том. 78. Национальный музей этнологии, Осака. стр. 307–340. ISBN 9784901906937 . Архивировано из оригинала (PDF) 19 января 2019 года . Проверено 18 января 2019 г.
- ^ Jump up to: а б с д Баркер, Грэм; Ллойд-Смит, Линдси; Бартон, Хью; Коул, Франка; Хант, Крис; Пайпер, Филип Дж.; Рабетт, Райан; Пас, Виктор; Сабо, Кэтрин (2011). «Переход к собирательству и земледелию в пещерах Ниах, Саравак, Борнео» (PDF) . Античность . 85 (328): 492–509. дои : 10.1017/S0003598X00067909 . S2CID 131187696 . Архивировано из оригинала (PDF) 19 января 2019 года . Проверено 18 января 2019 г.
- ^ Jump up to: а б Бальбалиго, Иветт (15 ноября 2007 г.). «Краткая заметка о раскопках 2007 года в пещере Иль, Палаван, Филиппины» . Материалы Института археологии . 18 (2007): 161. doi : 10.5334/pia.308 .
- ^ Фуллагар, Ричард; Филд, Джудит; Денхэм, Тим; Лентфер, Кэрол (май 2006 г.). «Использование орудий и обработка таро ( Colocasia esculenta ), ямса ( Dioscorea sp.) и других растений в начале и середине голоцена в болоте Кук в высокогорье Папуа-Новой Гвинеи». Журнал археологической науки . 33 (5): 595–614. Бибкод : 2006JArSc..33..595F . дои : 10.1016/j.jas.2005.07.020 .
- ^ Денхэм, Тим П.; Голсон, Дж.; Хьюз, Филип Дж. (2004). «Чтение раннего земледелия на болоте Кук, долина Вахги, Папуа-Новая Гвинея: археологические особенности (этапы 1–3)». Труды Доисторического общества . 70 : 259–297. дои : 10.1017/S0079497X00001195 . S2CID 128780288 .
- ^ Лой, Томас Х.; Сприггс, Мэтью; Уиклер, Стивен (1992). «Прямое свидетельство использования растений человеком 28 000 лет назад: остатки крахмала на каменных артефактах с северных Соломоновых островов» . Античность . 66 (253): 898–912. дои : 10.1017/S0003598X00044811 . S2CID 162663964 .
- ^ Баркер, Грэм; Хант, Крис; Карлос, Джейн (2011). «Переход к сельскому хозяйству на островах Юго-Восточной Азии: археологические, биомолекулярные и палеоэкологические перспективы» (PDF) . В Баркере, Грэм; Яновский, Моника (ред.). Зачем культивировать? Антропологические и археологические подходы к переходу от собирательства к земледелию в Юго-Восточной Азии . Институт археологических исследований Макдональда. стр. 61–74. ISBN 9781902937588 .
- ^ Бартон, Хью (2012). «Перевернутая судьба саго и риса Oryza sativa в тропических лесах Саравака, Борнео» (PDF) . Четвертичный интернационал . 249 : 96–104. Бибкод : 2012QuInt.249...96B . дои : 10.1016/j.quaint.2011.03.037 .
- ^ «Древние сельскохозяйственные практики чаморро» . Гуампедия . 10 февраля 2011 года . Проверено 18 января 2019 г.
- ^ Диксон, Бойд; Уокер, Сэмюэл; Голаби, Мохаммед Х.; Маннер, Харли (2012). «Два, вероятно, сельскохозяйственных объекта периода латте на севере Гуама: их растения, почвы и интерпретации» (PDF) . Микронесика . 42 (1/2): 209–257.
- ^ «талас» . Малайский литературный справочный центр . Куала-Лумпур: Деван Бахаса дан Пустака. Архивировано из оригинала 3 июня 2021 года . Проверено 22 мая 2022 г.
- ^ Шьямсарул (2013). Индонезийско-минангкабауский словарь [ Индонезийско-минангкабауский словарь ] (PDF) (на индонезийском языке) (пересмотренная редакция). Паданг: Языковой центр провинции Западная Суматра, Агентство развития и развития языков, Министерство образования и культуры. п. 201. ИСБН 9789790691254 . Архивировано из оригинала (PDF) 19 августа 2021 года.
- ^ «*Тало/Таро» . Те Мара Рео: Языковой сад . Семейный фонд Бентонов . Проверено 18 января 2019 г.
- ^ Jump up to: а б с Пятница, Джей Би; Окано, Дана (2006). « Cordia subcordata (коу)» (PDF) . В Элевиче, Крейг Р. (ред.). Профили видов для агролесомелиорации островов Тихого океана . Постоянные сельскохозяйственные ресурсы (PAR).
- ^ Оно, Ринтаро; Аддисон, Дэвид Дж. (2013). «Историческая экология и 600 лет использования рыбы на атолле Атафу, Токелау» . В Оно, Ринтаро; Моррисон, Алекс; Аддисон, Дэвид Дж. (ред.). Использование доисторических морских ресурсов в Индо-Тихоокеанском регионе . Южная земля. Том. 39. АНУ Э Пресс. ISBN 9781925021264 .
- ^ Судармин, М.Си; Пуджиастути, Шри Энданг (2015). «Культура, основанная на научных знаниях, и местная мудрость в Каримунджаве для развития и сохранения социальных навыков» (PDF) . Международный журнал науки и исследований . 4 (9): 598–604.
- ^ Бургер, Бренда Х. (2009). «Деревья острова Санта-Крус и их метафоры» (PDF) . Материалы семнадцатого ежегодного симпозиума о языке и обществе, Остин . Техасский лингвистический форум. Том. 53. С. 100–109. Архивировано из оригинала (PDF) 21 сентября 2021 года . Проверено 18 января 2019 г.
- ^ Кан, Дженнифер Г.; Катушка, Джеймс Х. (2006). «Что домашние сообщения говорят нам о различиях в статусах в доисторическом обществе: интерпретация анализа древесного угля, священных лесов и изменчивости между местами» (PDF) . Журнал Полинезийского общества . 115 (4): 319–352. Архивировано из оригинала (PDF) 23 октября 2023 года . Проверено 18 января 2019 г.
- ^ Галька, Мэтью (2008). «На этом прекрасном берегу нет фруктов: какие растения были завезены на субтропические полинезийские острова до контакта с европейцами?» (PDF) . В Кларке, Джеффри Ричард; О'Коннор, Сью; Лич, Брайан Фосс (ред.). Острова исследования: колонизация, мореплавание и археология морских ландшафтов . Южная земля. Том. 29. АНУ Э Пресс. стр. 227–251. ISBN 9781921313905 .
- ^ Миллерстрем, Сидсель; Катушка, Джеймс Х. (2008). «Доконтактное лесоводство и растительность на Маркизских островах, Французская Полинезия: идентификация по древесному углю и радиоуглеродный анализ из долины Хатихеу, Нуку-Хива» (PDF) . Азиатские перспективы . 47 (2): 330–351. дои : 10.1353/asi.0.0005 . hdl : 10125/17292 . S2CID 161160463 .
- ^ Грин, Валери Дж.; Грин, Роджер К. (2007). «Акцент на атоллах в подходах к истории населения отдаленной Океании». В Кирхе, Патрик В.; Раллю, Жан-Луи (ред.). Рост и распад обществ тихоокеанских островов: археологические и демографические перспективы . Гавайский университет Press. стр. 232–256 . ISBN 9780824831349 .
- ^ Jump up to: а б с д Хинкль, Аня Э. (май 2007 г.). «Структура населения тихоокеанской Cordyline fruticosa (Laxmanniaceae) с последствиями для населенных пунктов Полинезии». Американский журнал ботаники . 94 (5): 828–839. дои : 10.3732/ajb.94.5.828 . ПМИД 21636452 .
- ^ Филип, Симпсон (9 сентября 2012 г.). « Кордилина fruticosa (ти растение)» . Справочник инвазивных видов . Международный центр сельского хозяйства и биологических наук . Проверено 19 января 2019 г.
- ^ Jump up to: а б с д Хинкль, Аня Э. (2004). «Распространение мужской стерильной формы ти ( Cordyline fruticosa ) в Полинезии: случай человеческого отбора?» (PDF) . Журнал Полинезийского общества . 113 (3): 263–290. Архивировано из оригинала (PDF) 5 июля 2022 года . Проверено 19 января 2019 г.
- ^ Триша, Борланд (2009). « Кордилина fruticosa : распространение и преемственность священного растения» (PDF) . Класс UCB Муреа: Биология и геоморфология тропических островов . Студенческие исследовательские работы, осень 2009 г. Калифорнийский университет.
- ^ Jump up to: а б с д и ж г час Эрлих, Селия (2000). « От несъедобного к съедобному: хождение по огню и растение ти [ Cordyline fruticosa (L.) A. Chev.]» . Журнал Полинезийского общества . 109 (4): 371–400. Архивировано из оригинала 1 декабря 2019 года . Проверено 21 января 2019 г.
- ^ Эрлих, Селия (1989). «Специальные проблемы поиска этноботанической литературы: Cordyline Terminalis (L.) Kunth, «гавайское растение ти» » (PDF) . Журнал этнобиологии . 9 (1): 51–63.
- ^ Jump up to: а б Раппапорт, Рой А. (1989). Свиньи для предков: Ритуал в экологии народа Новой Гвинеи, второе издание . Уэйвленд Пресс. стр. 19, 125, 231. ISBN. 9781478610021 .
- ^ Ленсе, Обед (1 апреля 2012 г.). «Дикие растения, используемые коренными жителями Маноквари, Западное Папуа, в качестве традиционных лекарств» . Биоразнообразие . 13 (2): 98–106. дои : 10.13057/biodiv/d130208 .
- ^ Jump up to: а б с д Симпсон, Филип (1 января 1997 г.). «Стоят ли чего-нибудь капустные деревья? Связь экологических и человеческих ценностей в капустном дереве, tī kōuka» . Журнал новозеландских исследований . 7 (1). дои : 10.26686/jnzs.v7i1.395 .
- ^ Ланье, Клинт (10 ноября 2016 г.). «Околехао, сладкий гавайский самогон с отвратительным прошлым» . Пожиратель . Проверено 21 января 2019 г.
- ^ Гриффит, Роберт Эглсфельд (1847). Медицинская ботаника . Леа и Бланшар. п. 655.
- ^ ван Шуневелд, Инге (18 июля 2018 г.). «Ритуал сбора риса Пуннук прославляет изобилие Земли» . Синчи . Проверено 19 января 2019 г.
- ^ де Гузман, Даниэль (6 июля 2018 г.). «Флора де Филиппинас: краткий обзор филиппинских растений» . Проект Асванг . Проверено 19 января 2019 г.
- ^ Ритуалы сбора урожая в Хапао (PDF) . Социальные практики, ритуалы и праздничные мероприятия. Международный информационно-сетевой центр нематериального культурного наследия в Азиатско-Тихоокеанском регионе под эгидой ЮНЕСКО (ICHCAP). Архивировано из оригинала (PDF) 9 сентября 2018 года . Проверено 19 января 2019 г.
- ^ Балангкод, ТД; Балангкод, АК (январь 2009 г.). «Недостаточно используемые растительные ресурсы в Тиноке, Ифугао, административный район Кордильеры, остров Лусон, Филиппины» Acta Horticulturae (806): 647–654. doi : 10.17660/ActaHortic.2009.806.80 .
- ^ Новеллино, Дарио (2001). «Отношение Палавана к болезням» (PDF) . Филиппинские исследования . 49 (1): 78–93.
- ^ Гунаван (2013). «Изучение этноботаники растений андонг рода Cordyline в даякнесе деревни Мератус Лок Лахунг, Локсадо, Южное Борнео» . 4-я Международная конференция по сохранению глобальных ресурсов и 10-й Конгресс Индонезийского общества таксономии растений . Университет Бравиджая. Архивировано из оригинала 26 января 2020 года . Проверено 19 января 2019 г.
- ^ Jump up to: а б Рахаю, Муляти; Рустиами, Химма; Ругая (2016). «Этноботаническое исследование сасакской народности, Восточный Ломбок, Западная Нуса-Тенгара» (PDF) . Журнал тропической биологии и охраны природы : 85–99.
- ^ Леб, Э.М. (1929). «Шаман и Провидец». Американский антрополог . 31 (1): 60–84. дои : 10.1525/aa.1929.31.1.02a00050 .
- ^ Хаким, Нурул (2006). Культурные и духовные ценности лесов региона Бади, Бантен, Индонезия . Магистр лесной и природоохранной политики. Вагенингенский университет.
- ^ Искандар, Йохан; Искандар, Будиавати С. (15 марта 2017 г.). «Различные растения традиционной обрядности: этноботанические исследования среди бадуйской общины» . Биосаинтификация: Журнал биологии и биологического образования . 9 (1): 114. doi : 10.15294/biosaintifika.v9i1.8117 .
- ^ Домениг, Гауденц (2014). Хофте, Розмари; Нордхольт, Хенк Шульте (ред.). Религия и архитектура в домодерной Индонезии: исследования пространственной антропологии . Документы Королевского института лингвистики, географии и этнологии. Том. 294. БРИЛЛ. п. 74. ИСБН 9789004274075 .
- ^ Сумантера, И. Вайан (1999). «Растения, переплетенные с культурой» . Корни . 1 (19). Архивировано из оригинала 19 января 2019 года . Проверено 19 января 2019 г.
- ^ Нуй-Палм, Хетти (1979). Садан-Тораджа: исследование их общественной жизни и религии . Документы Королевского института лингвистики, географии и этнологии. Том. 87. Спрингер. п. 225. ИСБН 9789401771504 .
- ^ Волкман, Тоби Элис (1985). Праздники почета: ритуалы и перемены в нагорье Тораджа . Исследования Иллинойса в области антропологии. Том. 16. Издательство Университета Иллинойса. ISBN 9780252011832 .
- ^ Фокс, Джеймс Дж. (2006). Внутри австронезийских домов: перспективы внутреннего дизайна для жизни . АНУ Э Пресс. п. 21. ISBN 9781920942847 .
- ^ Jump up to: а б Шеридан, Майкл (1 января 2016 г.). «Пограничные растения, социальное производство пространства и вегетативная деятельность в аграрных обществах». Окружающая среда и общество . 7 (1): 29–49. дои : 10.3167/ares.2016.070103 .
- ^ Уэст, Пейдж (2006). Сохранение природы теперь наше правительство: экологическая политика в Папуа-Новой Гвинее . Издательство Университета Дьюка. п. 22. ISBN 9780822388067 .
- ^ Моррис, Брайан (2006). Религия и антропология: критическое введение . Издательство Кембриджского университета. п. 249. ИСБН 9780521852418 .
- ^ Раппапорт, Рой А. (1967). «Ритуальное регулирование экологических отношений среди жителей Новой Гвинеи». Этнология . 6 (1): 17–30. CiteSeerX 10.1.1.460.3366 . дои : 10.2307/3772735 . JSTOR 3772735 .
- ^ Боннемер, Паскаль (март 2018 г.). «Действия, отношения и трансформации: жизненный цикл согласно Анкаве Папуа-Новой Гвинеи» (PDF) . Океания . 88 (1): 41–54. дои : 10.1002/ocea.5180 .
- ^ Хилл, Ровена Кэтрин (2011). Цвет и церемония: роль красок у народов менди и сулка в Папуа-Новой Гвинее (магистр наук). Даремский университет.
- ^ Номбо, Порер; Лич, Джеймс (2010). «Растения реите: этноботаническое исследование на ток-писинском и английском языках» (PDF) . Монография по окружающей среде Азиатско-Тихоокеанского региона . Том. 4. АНУ Э Пресс. ISBN 9781921666018 .
- ^ Кизинг, Роджер М. (сентябрь 2012 г.). «О непонимании символов». HAU: Журнал этнографической теории . 2 (2): 406–430. дои : 10.14318/hau2.2.023 . S2CID 143871873 .
- ^ Панофф, Франсуаза (2018). Барбира-Фридман, Франсуаза (ред.). Сады Мэнге: исследование отношений Мэнге с домашними животными . Публикации pacific-credo. дои : 10.4000/books.pacific.690 . ISBN 9782956398189 .
- ^ Грей, Эндрю (2012). Языки острова Пятидесятницы . Издательство Manples (BFoV). ISBN 9780956098542 .
- ^ Jump up to: а б Мур, Дарлин Р. (2005). «Археологические свидетельства доисторической техники земледелия на Гуаме» (PDF) . Микронесика . 38 (1): 93–120.
- ^ Кавате, М.; Учида, Дж.; Кофлин, Дж.; Мельцер, М.; Кадука, К.; Кам, Дж.; Сугано, Дж.; Фукуда, С. (2014). «Болезни листьев Ti ( Cordyline Terminalis или Fruticosa ) в коммерческих садах Гавайев» (PDF) . Hānai'Ai/Поставщик еды : 1–11. Архивировано из оригинала (PDF) 10 декабря 2015 года . Проверено 21 января 2019 г.
- ^ Гамайо, Дарде (4 августа 2016 г.). «Лиф Ти: растение каноэ древних Гавайев» . Большой остров сейчас . Проверено 21 января 2019 г.
- ^ Да Силва, Александр (24 июля 2005 г.). «Азартная поездка» . Звездный вестник . Проверено 21 января 2019 г.
- ^ Jump up to: а б "*Ти" . Те Мара Рео: Языковой сад . Семейный фонд Бентонов . Проверено 15 января 2019 г.
- ^ Jump up to: а б Баркер, Грэм; Хант, Крис; Бартон, Хью; Госден, Крис; Джонс, Сэм; Ллойд-Смит, Линдси; Фарр, Люси; Ньири, Борбала; О'Доннелл, Шон (август 2017 г.). «Культурные тропические леса Борнео» (PDF) . Четвертичный интернационал . 448 : 44–61. Бибкод : 2017QuInt.448...44B . дои : 10.1016/j.quaint.2016.08.018 .
- ^ Беваква, Роберт Ф. (1994). «Происхождение садоводства в Юго-Восточной Азии и распространение одомашненных растений на острова Тихого океана полинезийскими путешественниками: пример Гавайских островов» (PDF) . ХортСайенс . 29 (11): 1226–1229. дои : 10.21273/HORTSCI.29.11.1226 .
- ^ Jump up to: а б с Божар, Филипп (август 2011 г.). «Первые мигранты на Мадагаскар и завоз растений: лингвистические и этнологические данные» (PDF) . Азания: Археологические исследования в Африке . 46 (2): 169–189. дои : 10.1080/0067270X.2011.580142 . S2CID 55763047 .
- ^ Уолтер, Энни; Лебот, Винсент (2007). Сады Океании . IRD Éditions-CIRAD. ISBN 9781863204705 .
- ^ Jump up to: а б с д и «*Цюфи ~ Прикрыть, прикрыть» . Языковой сад: Языковой сад . Семейный фонд Бентонов . Проверено 21 января 2019 г.
- ^ Малапа, Р.; Арнау, Г.; Нойер, Дж.Л.; Лебот, В. (ноябрь 2005 г.). «Генетическое разнообразие ямса большого (Dioscorea alata L.) и родство с D. nummularia Lam. и D. transversa Br., выявленное с помощью маркеров AFLP». Генетические ресурсы и эволюция сельскохозяйственных культур . 52 (7): 919–929. дои : 10.1007/s10722-003-6122-5 . S2CID 35381460 .
- ^ Круз, ВМВ; Альтоверос, Северная Каролина; Мендиоро, MS; Рамирес, Д.А. (1999). «Географические закономерности разнообразия филиппинской коллекции съедобного ямса». Информационный бюллетень о генетических ресурсах растений . 119 : 7–11.
- ^ Пас, Виктор Дж. (1999). «Передвижение человека в эпоху неолита на острова Юго-Восточной Азии: поиск археоботанических свидетельств» . Бюллетень Индо-Тихоокеанской ассоциации предыстории . 18 (Документы Малакки, том 2): 151–158. doi : 10.7152/bippa.v18i0.11710 (неактивен 7 февраля 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на февраль 2024 г. ( ссылка ) - ^ Jump up to: а б Бэйлисс-Смит, Тим; Голсон, Джек; Хьюз, Филип (2017). «Фаза 4: Основные каналы утилизации, щелевидные канавы и сетчатые поля» . В Голсоне, Джек; Денхэм, Тим; Хьюз, Филип; Пеленание, Памела; Мьюк, Джон (ред.). Десять тысяч лет культивирования на болоте Кук в высокогорье Папуа-Новой Гвинеи . Южная земля. Том. 46. АНУ Пресс. стр. 239–268. ISBN 9781760461164 .
- ^ Бартон, Хью (2005). «Дело о собирателях тропических лесов: запись о крахмале в пещере Ниах, Саравак» (PDF) . Азиатские перспективы . 44 (1): 56–72. дои : 10.1353/asi.2005.0005 . hdl : 10125/17222 . S2CID 11596542 .
- ^ Бартон, Хью; Денхэм, Тимоти (2011). «Доисторическая растительность и общественная жизнь на островах Юго-Восточной Азии и Меланезии» (PDF) . В Баркере, Грэм; Яновский, Моника (ред.). Зачем культивировать? Антропологические и археологические подходы к переходу от собирательства к земледелию в Юго-Восточной Азии . Институт археологических исследований Макдональда. стр. 61–74. ISBN 9781902937588 . Архивировано из оригинала (PDF) 28 марта 2019 года . Проверено 21 января 2019 г.
- ^ Рейнольдс, Тим; Баркер, Грэм; Бартон, Хью; Крэнбрук, Гаторн; Хант, Крис; Килхофер, Лиза; Пас, Виктор; Пайк, Аласдер; Пайпер, Филип; Рабетт, Райан; Рашворт, Гэри; Стимпсон, Кристофер; Сабо, Кэтрин (2013). «Первые современные люди в Ние, ок. 50 000–35 000 лет назад» (PDF) . В Баркере, Грэм (ред.). Собирательство и земледелие в тропических лесах на островах Юго-Восточной Азии . Институт археологических исследований Макдональда. стр. 133–170. ISBN 9781902937540 .
- ^ « Dioscorea Bulbifera – Air Yam» . Гавайские растения и тропические цветы . 5 ноября 2012 года . Проверено 21 января 2019 г.
- ^ Андрес, К.; АдеОлува, ОО; Бхуллар, GS (2016). «Ямс ( Dioscorea spp.)» . В Томасе, Брайане; Мерфи, Денис Дж.; Мюррей, Брайан Г. (ред.). Энциклопедия прикладных наук о растениях (2-е изд.). Академическая пресса. стр. 435–441. ISBN 9780123948076 .
- ^ Бленч, Роджер (2010). «Свидетельства австронезийских путешествий в Индийский океан» (PDF) . В Андерсоне, Атолл ; Барретт, Джеймс Х.; Бойл, Кэтрин В. (ред.). Глобальные истоки и развитие мореплавания . Институт археологических исследований Макдональда. стр. 239–248. ISBN 9781902937526 .
- ^ Jump up to: а б с Хугерворст, Том (2013). «Если бы только растения могли говорить...: Реконструкция досовременных биологических транслокаций в Индийском океане» (PDF) . В Чандре — Сатиш; Прабха Рэй, Химаншу (ред.). Море, идентичность и история: от Бенгальского залива до Южно-Китайского моря . Манохар. стр. 67–92. ISBN 9788173049866 .
- ^ Кирч, Патрик Винтон; Грин, Роджер К. (2001). Гавайки, Наследственная Полинезия: очерк исторической антропологии . Издательство Кембриджского университета. п. 267. ИСБН 9780521788793 .
- ^ Сайкс, WR (декабрь 2003 г.). «Диоскореевые, новые для адвентивной флоры Новой Зеландии» . Новозеландский журнал ботаники . 41 (4): 727–730. Бибкод : 2003NZJB...41..727S . дои : 10.1080/0028825X.2003.9512884 . S2CID 85828982 .
- ^ Уайт, Линтон Дав. «Ухи» . На Меакану о Ваа о Гавайи Кахико: «Растения каноэ» древних Гавайев . Проверено 21 января 2019 г.
- ^ Хоррокс, Марк; Нанн, Патрик Д. (май 2007 г.). «Свидетельства интродукции таро ( Colocasia esculenta ) и малого батата ( Dioscorea esculenta ) в отложениях эпохи Лапита (около 3050–2500 кал. лет назад) из Бурева, юго-запад острова Вити-Леву, Фиджи». Журнал археологической науки . 34 (5): 739–748. Бибкод : 2007JArSc..34..739H . дои : 10.1016/j.jas.2006.07.011 .
- ^ Хоррокс, М.; Грант-Маки, Дж.; Матису-Смит, Э. (январь 2008 г.). «Интродуцированы таро ( Colocasia esculenta ) и ямс ( Dioscorea spp.) в подтанских (2700–1800 лет назад) отложениях из пещеры Ме Ауре (WMD007), Муинду, Новая Каледония». Журнал археологической науки . 35 (1): 169–180. Бибкод : 2008JArSc..35..169H . дои : 10.1016/j.jas.2007.03.001 .
- ^ Пеленание, Памела; Хидэ, Робин (2005). «Изменение ландшафта и социального взаимодействия: взгляд на историю сельского хозяйства с точки зрения Сепика-Раму» (PDF) . В Поли, Эндрю; Аттенборо, Роберт; Голсон, Джек; Хидэ, Робин (ред.). Папуасское прошлое: культурная, языковая и биологическая история папуасоязычных народов . Тихоокеанская лингвистика. стр. 289–327. ISBN 9780858835627 .
- ^ Эндрю, Поли (2017). «Были ли первые колонизаторы лапита фермерами отдаленной Океании, а также собирателями?» . В Пайпер, Филип Дж.; Мацумура, Хирофуми; Булбек, Дэвид (ред.). Новые перспективы в предыстории Юго-Восточной Азии и Тихого океана . Южная земля. Том. 45. АНУ Пресс. стр. 293–310. ISBN 9781760460952 .
- ^ «Корнеплоды: Ямс» . Проверено 25 июля 2021 г.
- ^ Оливейра, Нуно Васко (2012). «Восстановление, анализ и идентификация видов Colocasia esculenta и Dioscorea в археологических условиях Тимора-Лешти». Сенри Этнологические исследования . 78 : 265–284. дои : 10.15021/00002521 .
- ^ Эзедин З.; Вайблен, Германия (27 сентября 2019 г.). «Дополнения и изменения к фикусу (Moraceae) в Новой Гвинее с комментариями к самому большому в мире инжиру» . Бюллетень садов Сингапур . 71 (приложение 2): 197–216. doi : 10.26492/gbs71(дополнение 2).2019-15 . ISSN 0374-7859 . S2CID 208557447 .
- ^ «Растения мира онлайн» . Проверено 8 апреля 2019 г.
- ^ «Фикус гипсидский» . Почтовый индексЗоопарк . Архивировано из оригинала 30 августа 2010 года . Проверено 17 апреля 2012 г.
- ^ «Биотик.орг» . Фикус хиспида . Архивировано из оригинала 25 июля 2010 года . Проверено 17 апреля 2012 г.
- ^ «Фикус нота (Бланко) Мерр» . Растения мира онлайн . Попечители Королевского ботанического сада Кью. нд . Проверено 23 августа 2020 г. .
- ^ « Одомашненные растения и животные Австронезии ». Информационная сеть по ресурсам зародышевой плазмы . Служба сельскохозяйственных исследований Министерства сельского хозяйства США .
- ^ Jump up to: а б Берг, CC; Уголок EJH (2005). «Моровые». Флора Малезиана . Том. 17. ISBN 1-930723-40-7 .
- ^ Jump up to: а б Корнер, EJH (1965). «Контрольный список фикусов Азии и Австралазии с ключами к идентификации» . Бюллетень садов Сингапура . (оцифровано, онлайн, на сайте biodiversitylibrary.org). 21 (1): 1–186 . Проверено 5 февраля 2014 г.
- ^ Jump up to: а б «Фикус Аспера» . Международный указатель названий растений (IPNI) . Проверено 14 февраля 2020 г.
- ^ Jump up to: а б « Фикус фразери » . Эндемик – фауна и флора Новой Каледонии . Архивировано из оригинала 16 июля 2011 года . Проверено 12 июля 2008 г.
- ^ Полезные тропические растения
- ^ Хасан, МЫ (2006). Лечебные травы Малайзии . Куала-Лумпур: Федеральное агентство землеустройства. ISBN 9789839954425 .
- ^ «Фикус Аспера Г.Форст» . Список растений. Рабочий список всех видов растений . Проверено 14 февраля 2020 г.
- ^ Эзедин З.; Вайблен, Германия (27 сентября 2019 г.). «Дополнения и изменения к фикусу (Moraceae) в Новой Гвинее с комментариями к самому большому в мире инжиру» . Бюллетень садов Сингапур . 71 (приложение 2): 197–216. doi : 10.26492/gbs71(дополнение 2).2019-15 . ISSN 0374-7859 . S2CID 208557447 .
- ^ « Фикус фразери » . Австралийский индекс названий растений (APNI), база данных IBIS . Центр исследований биоразнообразия растений, правительство Австралии.
- ^ Николсон, Нэн и Хью (1985). Растения тропических лесов Австралии . Новый Южный Уэльс: Питомник тропических лесов Терании. ISBN 0958943605 .
- ^ Лоу, Т. (1991). Дикие пищевые растения Австралии . Австралия: Ангус и Робертсон. ISBN 0207169306 .
- ^ «Фикус нота» . Экосистемы тихоокеанских островов под угрозой . Проект «Гавайские экосистемы под угрозой». 1 января 1999 года . Проверено 5 мая 2009 г.
- ^ Ферн, Кен. «Фикус нота» . База данных тропических растений . Проверено 18 мая 2019 г.
- ^ «Тибиг» . Расширенная национальная программа экологизации . Департамент окружающей среды и природных ресурсов Республики Филиппины. Архивировано из оригинала 8 декабря 2022 года . Проверено 18 мая 2019 г.
- ^ <!- -Не указано--> (б. д.). «Фикус псевдопальма Бланко» . Растения мира онлайн . Попечители Королевского ботанического сада Кью . Проверено 23 августа 2020 г. .
- ^ Старр, Ф. и др. (2003). Информационный бюллетень о фикусе псевдопальме . Гавайские экосистемы под угрозой.
- ^ «Фрукты и овощи из тропического леса» (PDF) . Технологический центр продуктов питания и удобрений . Архивировано из оригинала (PDF) 24 июля 2011 года . Проверено 17 февраля 2022 г.
- ^ Виллегас, КЛ и Ф.А. Поллиско-младший (2008). Цветочное обследование субводораздела Лайбан в горном хребте Сьерра-Мадре на Филиппинах. Архивировано 7 марта 2012 г. в Wayback Machine журнале тропической биологии и охраны природы 4 (1) 1-14.
- ^ «Терпеноиды и стеролы из эндемичных и находящихся под угрозой исчезновения филиппинских деревьев Ficus pseudopalma и Ficus ulmifolia » (PDF) . Филиппинский научный журнал 138(2) 205 . Архивировано из оригинала (PDF) 30 апреля 2015 года . Проверено 17 февраля 2022 г.
- ^ Лю В., Ван П., Ли Дж., Лю Вэньяо и Ли Хунмей (2014), Пластичность поглощения исходной воды в эпифитных, переходных и наземных фазах роста Ficus tinctoria , Ecoгидрол. , 7; страницы 1524–1533, doi:10.1002/eco.1475
- ^ «Мати – Языковой сад» . termarareo.org . Проверено 30 сентября 2020 г.
- ^ Нельсон, Скот К. (2006). « Pandanus tectorius (панданус)» (PDF) . В Элевиче, Крейг Р. (ред.). Профили видов для агролесомелиорации островов Тихого океана . Постоянные сельскохозяйственные ресурсы (PAR). Архивировано из оригинала (PDF) 31 августа 2021 года . Проверено 22 января 2019 г.
- ^ Jump up to: а б с Перье, Ксавье; Бакри, Фредерик; Каррель, Франсуаза; Дженни, Кристоф; Хорри, Жан-Пьер; Лебот, Винсент; Ипполит, Изабель (2009). «Объединение биологических подходов, чтобы пролить свет на эволюцию съедобных бананов» . Этноботаника: исследования и приложения . 7 : 199–216. дои : 10.17348/era.7.0.199-216 . hdl : 10125/12515 .
- ^ Фуллер, Дориан Кью; Бойвен, Николь; Хугерворст, Том; Аллаби, Робин (2 января 2015 г.). «Через Индийский океан: доисторическое перемещение растений и животных» . Античность . 85 (328): 544–558. дои : 10.1017/S0003598X00067934 . S2CID 15456546 .
- ^ Jump up to: а б Плетц, Рэнди К.; Кеплер, Анджела Кей; Дэниелс, Джефф; Нельсон, Скотт К. (2007). «Банан и подорожник — обзор с акцентом на сорта тихоокеанских островов» (PDF) . В Элевиче, Крейг Р. (ред.). Профили видов для агролесомелиорации островов Тихого океана . Постоянные сельскохозяйственные ресурсы (PAR).
- ^ Питер Лэйп; Эмили Петерсон; Дауд Танудирджо; Чунг-Чинг Шиунг; Гён-А Ли; Джудит Филд; Адель Костер (2018). «Новые данные с открытой неолитической стоянки в Восточной Индонезии». Азиатские перспективы . 57 (2): 222–243. дои : 10.1353/asi.2018.0015 . hdl : 10125/72091 . S2CID 165484454 .
- ^ «Мриристицин — Молекула месяца — август 2014 г. (HTML-версия)» . www.chm.bris.ac.uk. Архивировано из оригинала 6 октября 2022 года . Проверено 28 апреля 2022 г.
- ^ Jump up to: а б Пикерсгилл, Барбара (2005). Пранс, Гиллин; Несбитт, Марк (ред.). Культурная история растений . Рутледж. п. 166. ИСБН 0415927463 .
- ^ Jump up to: а б с д и ж г час Беллвуд, Питер (9 декабря 2011 г.). «Клетчатая предыстория движения риса на юг как одомашненного злака - от Янцзы до экватора» (PDF) . Рис . 4 (3–4): 93–103. Бибкод : 2011Рис....4...93B . дои : 10.1007/s12284-011-9068-9 . S2CID 44675525 .
- ^ Jump up to: а б с Он, Кейанг; Лу, Хоуюань; Чжан, Цзяньпин; Ван, Джан; Хуан, Сюцзя (7 июня 2017 г.). «Доисторическая эволюция дуалистической структуры смешанного выращивания риса и проса в Китае» . Голоцен . 27 (12): 1885–1898. Бибкод : 2017Holoc..27.1885H . дои : 10.1177/0959683617708455 . S2CID 133660098 .
- ^ Нормил, Деннис (1997). «Янцзы считается самым ранним местом выращивания риса». Наука . 275 (5298): 309–310. дои : 10.1126/science.275.5298.309 . S2CID 140691699 .
- ^ Воган, округ Колумбия; Лу, Б; Томука, Н. (2008). «Развивающаяся история эволюции риса» . Наука о растениях . 174 (4): 394–408. doi : 10.1016/j.plantsci.2008.01.016 .
- ^ Харрис, Дэвид Р. (1996). Истоки и распространение земледелия и скотоводства в Евразии . Психология Пресс. п. 565. ИСБН 978-1-85728-538-3 .
- ^ Чжан, Цзяньпин; Лу, Хоуюань; Гу, Ванфа; Ву, Найцин; Чжоу, Куньшу; Ху, Яйи; Синь, Инцзюнь; Ван, Джан; Кашкуш, Халил (17 декабря 2012 г.). «Раннее смешанное земледелие проса и риса 7800 лет назад в районе средней реки Хуанхэ, Китай» . ПЛОС ОДИН . 7 (12): е52146. Бибкод : 2012PLoSO...752146Z . дои : 10.1371/journal.pone.0052146 . ПМЦ 3524165 . ПМИД 23284907 .
- ^ Се, Джау-шу; Хсинг, Юэ-и Кэролайн; Сюй, Цзы-фу; Ли, Пол Джен-куэй; Ли, Куанг-ти; Цанг, Ченг-хва (24 декабря 2011 г.). «Исследования древнего риса — место встречи ботаников, агрономов, археологов, лингвистов и этнологов» . Рис . 4 (3–4): 178–183. Бибкод : 2011Рис....4..178H . дои : 10.1007/s12284-011-9075-x . S2CID 37711686 .
- ^ Карсон, Майк Т. (2012). «Обзор археологии периода латте» (PDF) . Микронесика . 42 (1/2): 1–79.
- ^ Петерсон, Джон А. (2012). «Деревни латте на Гуаме и Марианских островах: монументальность или монументальность?» (PDF) . Микронесика . 42 (1/2): 183–08.
- ^ Баба, Сигэюки; Чан, Хунг; Кезука, Мио; Иноуэ, Томоми; Чан, Эрик (2016). « Artocarpus altilis и Pandanus tectorius : два важных плода Океании, имеющих лекарственную ценность» . Эмиратский журнал продовольствия и сельского хозяйства . 28 (8): 531. doi : 10.9755/ejfa.2016-02-207 .
- ^ Jump up to: а б с д Галлахер, Тимоти (2014). «Прошлое и будущее Халы ( Pandanus tectorius ) на Гавайях». В Киаве, Лия О'Нил, Массачусетс; Макдауэлл, Марша; Дьюхерст, К. Курт (ред.). `Айк Улана Лау Хала: жизнеспособность и яркость ткацких традиций Лау Хала на Гавайях . Гавайская школа гавайских знаний; Гавайский университет Press. дои : 10.13140/RG.2.1.2571.4648 . ISBN 9780824840938 .
- ^ Кирч, Патрик Винтон (2012). Акула, идущая вглубь суши, — мой вождь: островная цивилизация древних Гавайев . Издательство Калифорнийского университета. стр. 25–26. ISBN 9780520953833 .
- ^ Вольф, Джон У. (2018). Протоавстронезийская фонология с глоссарием . Том. II. Публикации Корнеллской программы Юго-Восточной Азии. п. 927. ИСБН 9781501735998 .
- ^ «Эволюция названий растений» . Те Мара Рео: Языковой сад . Семейный фонд Бентонов . Проверено 15 января 2019 г.
- ^ Jump up to: а б с д Томсон, Лекс Эй.Дж.; Энглбергер, Лоис; Гуарино, Луиджи; Таман, РР; Элевич, Крейг Р. (2006). « Pandanus tectorius (панданус)» (PDF) . В Элевиче, Крейг Р. (ред.). Профили видов для агролесомелиорации островов Тихого океана . Постоянные сельскохозяйственные ресурсы (PAR).
- ^ Кастро, Ирма Ремо (2006). Путеводитель по семействам распространенных цветковых растений на Филиппинах . Издательство Филиппинского университета. стр. 144–145. ISBN 9789715425254 .
- ^ Виерсема, Джон Х.; Леон, Бланка (2016). Мировые экономические предприятия: стандартный справочник (2-е изд.). ЦРК Пресс. ISBN 9781466576810 .
- ^ Кришнамурти, Канзас; Партасарати, Вирджиния; Саджи, КВ; Кришнамурти, Б. (2010). «Идеотипическая концепция черного перца ( Piper nigrum L.)» . Журнал специй и ароматических культур . 19 (1 и 2): 1–13.
- ^ Каннингем, AB; Ингрэм, В.; Даос Кадати, В.; Хау, Дж.; Судятмоко, С.; Рефли, Р.; Лием, СП; Тари, А.; Марук, Т.; Робианто, Н.; Синлае, А.; Ндун, Ю.; Мадуарта, И.; Сулистьохарди, Д.; Коеслутат, Э. (2011). Скрытая экономика, варианты будущего: торговля недревесными лесными товарами в восточной Индонезии . Австралийский центр международных сельскохозяйственных исследований (ACIAR). ISBN 9781921738685 .
- ^ Jump up to: а б с д Хиткот, Гэри М.; Диего, Винсент П.; Исида, Хадзиме; Сава, Винсент Дж. (2012). «Остебиография замечательного доисторического человека чаморро из Таги, Тиниан». Микронесика . 43 (2): 131–213.
- ^ Jump up to: а б Лебот, В.; Левеск, Ж. (1989). «Происхождение и распространение кавы ( Piper methysticum Forst. F., Piperaceae): фитохимический подход». Аллертония . 5 (2): 223–281.
- ^ Махди, Варуно (1999). «Распространение австронезийских лодочных форм в Индийском океане». В Бленче, Роджер; Сприггс, Мэтью (ред.). Археология и язык III: Языки и тексты артефактов . Единая мировая археология. Том. 34. Рутледж. стр. 144–179. ISBN 978-0415100540 .
- ^ Гильбоа, Айелет; Намдар, Дворы (9 февраля 2016 г.). «О начале торговли специями из Южной Азии со Средиземноморским регионом: обзор» . Радиоуглерод . 57 (2): 265–283. дои : 10.2458/azu_rc.57.18562 . S2CID 55719842 .
- ^ Jump up to: а б с д и ж г час я дж Росс, Малкольм (2008). «Прочие культурные растения» . В Россе, Малкольм; Поли, Эндрю; Осмонд, Мередит (ред.). Лексикон протоокеанского общества: культура и окружающая среда древнего океанического общества . Том. 3. Тихоокеанская лингвистика. стр. 389–426. ISBN 9780858835894 .
- ^ Равиндран, ПН (2017). Энциклопедия трав и специй . КАБИ. ISBN 9781780643151 .
- ^ Виарт, Кристоф (2006). Лекарственные растения Азиатско-Тихоокеанского региона: лекарства будущего? . Всемирная научная. ISBN 9789814480338 .
- ^ Jump up to: а б Далби, Эндрю (2002). Опасные вкусы: история специй . Издательство Калифорнийского университета. ISBN 9780520236745 .
- ^ Снодграсс, Мэри Эллен (2013). Мировая еда: энциклопедия истории, культуры и социального влияния от охотников-собирателей до эпохи глобализации: энциклопедия истории, культуры и социального влияния от охотников-собирателей до эпохи глобализации . Справочник Шарпа. ISBN 9781317451600 .
- ^ Jump up to: а б с «Горько ~ Горько» . Языковой сад: Языковой сад . Семейный фонд Бентонов . Проверено 15 января 2019 г.
- ^ Jump up to: а б «*Кава ~ *Кавакава» . Те Мара Рео: Языковой сад . Семейный фонд Бентонов . Проверено 15 января 2019 г.
- ^ Jump up to: а б с д Линч, Джон (2002). «Мощные корни и происхождение кавы». Океаническая лингвистика . 41 (2): 493–513. дои : 10.1353/ол.2002.0010 . S2CID 145424062 .
- ^ Jump up to: а б с Дэниелс, Кристиан; Мензис, Николас К. (1996). Нидхэм, Джозеф (ред.). Наука и цивилизация в Китае: Том 6, Биология и биологические технологии, Часть 3, Агропромышленность и лесное хозяйство . Издательство Кембриджского университета. стр. 177–185. ISBN 9780521419994 .
- ^ Дэниелс, Джон; Дэниелс, Кристиан (апрель 1993 г.). «Сахарный тростник в предыстории». Археология в Океании . 28 (1): 1–7. дои : 10.1002/j.1834-4453.1993.tb00309.x .
- ^ Jump up to: а б с Патерсон, Эндрю Х.; Мур, Пол Х.; Том Л., Тью (2012). «Генофонд видов Saccharum и его улучшение» . В Патерсоне, Эндрю Х. (ред.). Геномика сахарин . Springer Science & Business Media. стр. 43–72. ISBN 9781441959478 .
- ^ Альджанаби, Салах М. (1998). «Генетика, филогенетика и сравнительная генетика Saccharum L., полисомного полиплоида Poales: Andropogoneae» . В Эль-Гевели, М. Раафат (ред.). Ежегодный обзор биотехнологии . Том. 4. Elsevier Science BV, стр. 285–320. ISBN 9780444829719 .
- ^ Болдик, Джулиан (2013). Древние религии австронезийского мира: от Австралазии до Тайваня . ИБТаурис. п. 2. ISBN 9780857733573 .
- ^ Бласт, Роберт (1984–1985). «Австронезийская родина: лингвистическая перспектива» . Азиатские перспективы . 26 (1): 44–67. hdl : 10125/16918 .
- ^ Сприггс, Мэтью (2 января 2015 г.). «Археология и австронезийская экспансия: где мы сейчас?». Античность . 85 (328): 510–528. дои : 10.1017/S0003598X00067910 . S2CID 162491927 .
- ^ Аделаар, Александр (2016). «Австронезийцы на Мадагаскаре: критическая оценка работ Поля Оттино и Филиппа Божара» . В Кэмпбелле, Гвин (ред.). Ранний обмен между Африкой и большим миром Индийского океана . Серия Пэлгрейва по исследованиям мира Индийского океана. Пэлгрейв Макмиллан. стр. 77–112. ISBN 9783319338224 .
- ^ "Санталумовый альбом Л." Растения мира онлайн . Королевский ботанический сад Кью . Проверено 19 июня 2024 г.
- ^ Фуллер, Дориан К.; Бойвен, Николь; Кастильо, Кристина Кобо; Хугерворст, Том; Аллаби, Робин Г. (2015). «Археобиология перемещений в Индийском океане: современные очертания культурных обменов между протоисторическими мореплавателями». В Трипати, Сила (ред.). Морские контакты прошлого: расшифровка связей между сообществами . Дели: Книги Кавери. стр. 1–23. ISBN 9788192624433 .
- ^ Тернер, Джек (2004). Спайс: История искушения . Винтажные книги. стр. xxvii – xxviii. ISBN 978-0-375-70705-6 .
- ^ Уорролл, Саймон (23 июня 2012 г.). «Самое старое гвоздичное дерево в мире» . Журнал BBC News . Проверено 24 июня 2012 г.
- ^ Арималала, Наташа; Пено, Эрик; Мишельс, Тьерри; Ракотоариманана, Воджисон; Мишель, Изабель; Раваоманалина, Харизоа; Роджер, Эдмонд; Джахиэль, Мишель; Леонг Пок Цы, Жан-Мишель; Данту, Паскаль (август 2019 г.). «Системы земледелия на основе гвоздики на восточном побережье Мадагаскара: как история оставляет свой след в ландшафте» . Агролесомелиорационные системы . 93 (4): 1577–1592. Бибкод : 2019AgrSy..93.1577A . дои : 10.1007/s10457-018-0268-9 . ISSN 0167-4366 . S2CID 49583653 .
- ^ Jump up to: а б Уистлер, В. Артур; Элевич, Крейг Р. (2006). « Syzygiummalaccense (малайское яблоко) (пляжный гибискус)» (PDF) . В Элевиче, Крейг Р. (ред.). Традиционные деревья островов Тихого океана: их культура, окружающая среда и использование . Постоянные сельскохозяйственные ресурсы (ПАР). стр. 41–56. ISBN 9780970254450 .
- ^ Дотт-Сарут, Эмили (2016). «Свидетельства лесопользования и лесоводства на основе данных о древесном угле: антракологическое исследование на двух доколониальных участках канаков Новой Каледонии». История растительности и археоботаника . 26 (2): 195–211. дои : 10.1007/s00334-016-0580-0 . S2CID 132637794 .
- ^ Jump up to: а б с Фарли, Джина; Шнайдер, Лариса; Кларк, Джеффри; Хаберле, Саймон Г. (декабрь 2018 г.). «Позднеголоценовая палеоэкологическая реконструкция острова Улонг, Палау, на основе крахмального зерна, древесного угля и геохимического анализа» . Журнал археологической науки: отчеты . 22 : 248–256. Бибкод : 2018JArSR..22..248F . дои : 10.1016/j.jasrep.2018.09.024 . hdl : 1885/159550 . S2CID 135051639 .
- ^ Спеннеманн, Дирк Х.Р. (1994). «Традиционное производство и использование аррорута на Маршалловых островах». Журнал этнобиологии . 14 (2): 211–234.
- ^ Таман, Р.Р. (1994). «Этноботаника прибрежных равнин тихоокеанских островов» . В Моррисоне, Джон; Джерати, Пол; Кроул, Линда (ред.). Фауна, флора, еда и медицина: наука народов островов Тихого океана . Том. 3. Институт тихоокеанских исследований. стр. 147–184. ISBN 9789820201064 .
- ^ Jump up to: а б с Преббл, Матиу; Андерсон, Атолл (2012). «Археоботаника камнезащитных отложений Рапана» (PDF) . В Андерсоне, Атолл ; Кеннетт, Дуглас Дж. (ред.). Взятие на возвышенность: археология Рапы, укрепленного острова в отдаленной Восточной Полинезии . Южная земля. Том. 37. АНУ Э Пресс. стр. 77–95. ISBN 9781922144256 .
- ^ Jump up to: а б с Дотт-Сарут, Эмили; Кан, Дженнифер Г. (ноябрь 2017 г.). «Древние леса Полинезии: пилотное антракологическое исследование на острове Маупити, Французская Полинезия». Четвертичный интернационал . 457 : 6–28. Бибкод : 2017QuInt.457....6D . дои : 10.1016/j.quaint.2016.10.032 .
- ^ Элевич, Крейг Р.; Томсон, Лекс Эй Джей (2006). « Hibiscus tiliaceus (гибискус пляжный)» (PDF) . В Элевиче, Крейг Р. (ред.). Традиционные деревья островов Тихого океана: их культура, окружающая среда и использование . Постоянные сельскохозяйственные ресурсы (ПАР). стр. 41–56. ISBN 9780970254450 .
- ^ «*Мило» . Те Мара Рео: Языковой сад . Семейный фонд Бентонов . Проверено 22 января 2019 г.
- ^ Уджанг, Занария; Нордин, Нурул Изза; Субраманиам, Таваманитеви (декабрь 2015 г.). «Виды имбиря и их традиционное использование в современных целях». Журнал промышленных технологий . 23 (1): 59–70. doi : 10.21908/jit.2015.4 (неактивен 7 февраля 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на февраль 2024 г. ( ссылка ) - ^ Равиндран, Пенсильвания; Пиллаи, Гита С.; Бабу, К. Нирмал (2004). «Недостаточно используемые травы и специи». В Питере К.В. (ред.). Справочник трав и специй . Том. 2. Издательство Вудхед. ISBN 9781855737211 .
- ^ Ченг, Ланчини Джен-Хао (2014). Таксономия музыкальных инструментов тайваньских аборигенов (PDF) (доктор философии). Университет Отаго.
- ^ Jump up to: а б с д и Кикусава, Рицуко; Рид, Лоуренс А. (2007). «Прото, кто использовал куркуму и как?» (PDF) . В Сигеле, Джефф; Линч, Джон; Идс, Диана (ред.). Описание языка, история и развитие: лингвистическое послабление памяти Терри Кроули . Издательство Джона Бенджамина. стр. 339–352. ISBN 9789027292940 . Архивировано из оригинала (PDF) 25 ноября 2021 года . Проверено 18 января 2019 г.
- ^ МакКлатчи, В. (1993). «Традиционное использование Curcuma longa (Zingiberaceae) в Ротуме». Экономическая ботаника . 47 (3): 291–296. дои : 10.1007/bf02862297 . S2CID 20513984 .
- ^ Виестад, Андреас (2007). Где родился вкус: рецепты и кулинарные путешествия по маршруту специй Индийского океана . Книги летописи. п. 89. ИСБН 9780811849654 .
- ^ Равиндран, Пенсильвания; Нирмал Бабу, К. (2016). Имбирь: род Zingiber . ЦРК Пресс. п. 7. ISBN 9781420023367 .
- ^ Аллаби, Робин (2007). «Истоки эксплуатации растений в странах Ближней Океании: обзор» . Во Фридлендере, Джонатан С. (ред.). Гены, язык и история культуры в юго-западной части Тихого океана . Серия «Эволюция человека». Издательство Оксфордского университета. п. 191. ИСБН 9780198041085 .
- ^ Гроувс, Колин П. (2006). «Одомашненные и комменсальные млекопитающие Австронезии и их истории». В Беллвуде, Питер; Фокс, Джеймс Дж.; Трайон, Даррелл (ред.). Австронезийцы: исторические и сравнительные перспективы . АНУ Э Пресс. стр. 161–174. ISBN 1920942858 . JSTOR j.ctt2jbjx1.11 .
- ^ Амано, Ноэль; Пайпер, Филип Дж.; Хун, Сяо-чунь; Беллвуд, Питер (2013). «Интродуцированные домашние животные в эпоху неолита и металла на Филиппинах: данные из Нагсабарана, Северный Лусон». Журнал островной и прибрежной археологии . 8 (3): 317–335. дои : 10.1080/15564894.2013.781084 . S2CID 131368496 .
- ^ Михарес, Арманд Сальвадор Б. (2006). «Ранняя миграция австронезийцев на Лусон: перспективы пещер Пеньябланка». Бюллетень Индо-Тихоокеанской ассоциации предыстории . 26 : 72–78. CiteSeerX 10.1.1.403.7868 . doi : 10.7152/bippa.v26i0.11995 (неактивен 7 февраля 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на февраль 2024 г. ( ссылка ) - ^ Робертс, Э.А. (2014). Большой этимологический словарь испанского языка с семействами слов, основанными на индоевропейских корнях . Том. Я (А–Г). ООО «Экслибрис». п. 311. ИСБН 9781493191109 .
- ^ Jump up to: а б Уильям Генри Скотт (1990). «Висайская еда и сельское хозяйство шестнадцатого века». Филиппинский ежеквартальный журнал культуры и общества . 18 (4): 291–311. JSTOR 29792029 .
- ^ Кокрилл, WR, изд. (1977). Водяной буйвол (PDF) . Серия «Животноводство и здоровье». Том. 4. Продовольственная и сельскохозяйственная организация Объединенных Наций. ISBN 978-9251001080 . Архивировано из оригинала (PDF) 16 июня 2013 года . Проверено 23 февраля 2013 г.
- ^ Jump up to: а б Бласт, Роберт; Трассел, Стивен. «*qaNuaŋ» . Австронезийский сравнительный словарь . Проверено 7 января 2024 г.
- ^ "Корова" . Бинисая – кебуанский словарь и тезаурус . Приходить . Проверено 10 августа 2017 г.
- ^ Зорк, Р. Дэвид Пол (1982). Основной этимологический словарь филиппинского языка (PDF) . Том. фаск. 3. Дарвинский общественный колледж, Консорциум PNC-La Salle-Ateneo, Лингвистическое общество Филиппин, Исследование национального языка.
- ^ Эстрада-Вильегас, Валенте (1952). Животноводство Карабао . Манила: ДП Перес. п. 24.
- ^ Jump up to: а б с д Филлиос, Мелани А.; Тасон, Пол СК (2016). «Кто впустил собак? Обзор недавних генетических данных о завозе динго в Австралию и последствиях для передвижения людей» . Журнал археологической науки: отчеты . 7 : 782–792. Бибкод : 2016JArSR...7..782F . дои : 10.1016/j.jasrep.2016.03.001 .
- ^ Jump up to: а б с д Балмер, Сьюзен (2001). «Собаки лапита и поющие собаки и история собак Новой Гвинеи» . В Кларке, Джеффри Ричард; Андерсон, Атолл ; Вунидило, Тариси (ред.). Археология распространения лапита в Океании: материалы четвертой конференции лапита, июнь 2000 г., Канберра, Австралия . Книги Пандануса. стр. 183–202. ISBN 9781740760102 .
- ^ Jump up to: а б с д Грейг, Карен; Букок, Джеймс; Прост, Стефан; Хорсбург, К. Энн; Джейкомб, Крис; Уолтер, Ричард; Матису-Смит, Элизабет; Кайзер, Манфред (2015). «Полные митохондриальные геномы первых собак Новой Зеландии» . ПЛОС ОДИН . 10 (10): e0138536. Бибкод : 2015PLoSO..1038536G . дои : 10.1371/journal.pone.0138536 . ПМЦ 4596854 . ПМИД 26444283 .
- ^ Jump up to: а б О'Коннор, Сью; Махирта; Танудирджо, Дауд; Риримасс, Марлон; Хусни, Мухаммед; Кили, Симона; Хокинс, Стюарт; Алифа (4 декабря 2017 г.). «Идеология, ритуальное исполнение и его проявления в наскальном искусстве Тимора-Лешти и острова Кисар, острова Юго-Восточной Азии». Кембриджский археологический журнал . 28 (2): 225–241. дои : 10.1017/S0959774317000816 . S2CID 165822029 .
- ^ Jump up to: а б с д Грейг, Карен; Букок, Джеймс; Аллен, Мелинда С.; Матису-Смит, Элизабет; Уолтер, Ричард (2018). «Древние ДНК-доказательства интродукции и расселения собак ( Canis familis ) в Новой Зеландии» . Журнал тихоокеанской археологии . 9 (1): 1–10.
- ^ Чжан, Мин, Жэнь, Леле; Чжан, Личжао; Цао, Пэн; Ко, Альберт Ян, Мелинда А.; , Го-Дун, Цяомей (2020). распространении тихоокеанских собак» . Китая свидетельствуют о « 37 (5): 1462–1469. : 10.1093 /molbev/msz311 . doi Древние данные ДНК из . PMID 31913480
- ^ Jump up to: а б с Сакс, Бенджамин Н.; Браун, Сара К.; Стивенс, Даниэль; Педерсен, Нильс К.; Ву, Джуй-Те; Берри, Оливер (2013). «Анализ Y-хромосомы динго и деревенских собак Юго-Восточной Азии предполагает неолитическую континентальную экспансию из Юго-Восточной Азии с последующим множественным расселением австронезийцев» . Молекулярная биология и эволюция . 30 (5): 1103–1118. дои : 10.1093/molbev/mst027 . ПМИД 23408799 .
- ^ Райт, Ник (2020). «Коренные народы и животные Тайваня» . Проверено 20 марта 2024 г.
- ^ Кук, Крис; Стивенсон, Джом (2005). Путеводитель Рутледжа по всемирной истории с 1914 года . Нью-Йорк: Рутледж. п. 376. ИСБН 0415345847 .
- ^ Jump up to: а б с д Бласт, Роберт; Трассел, Стивен. «*асу₁» . Австронезийский сравнительный словарь . Проверено 20 марта 2024 г.
- ^ Очоа, Джанин Терез (2005). «В упорном преследовании: переоценка приручения и социальной интеграции собаки». Хукай . 8 : 37–66.
- ^ Саволайнен, Питер; Лейтнер, Томас; Уилтон, Алан Н.; Матису-Смит, Элизабет; Лундеберг, Лундеберг (6 августа 2004 г.). «Подробная картина происхождения австралийского динго, полученная в результате изучения митохондриальной ДНК» . Труды Национальной академии наук . 101 (33): 12387–12390. Бибкод : 2004PNAS..10112387S . дои : 10.1073/pnas.0401814101 . ПМК 514485 . ПМИД 15299143 .
- ^ Jump up to: а б с Грейг, К.; Гослинг, А.; Коллинз, CJ; Букок, Дж.; Макдональд, К.; Эддисон, диджей; Аллен, MS; Дэвид, Б.; Гиббс, М.; Хайэм, CFW; Лю, Ф.; МакНивен, Ай-Джей; О'Коннор, С.; Цанг, Швейцария; Уолтер, Р.; Матисоо-Смит, Э. (14 июня 2018 г.). «Сложная история происхождения и транслокаций собак (Canis familis) в Тихом океане, выявленная древними митогеномами». Научные отчеты . 8 (1). дои : 10.1038/s41598-018-27363-8 . hdl : 1885/265530 .
- ^ Титкомб, М (1969). Собака и человек в Древнем Тихом океане . Гонолулу: Специальное издание 59 музея Бернис П. Бишоп . ISBN 978-0-910240-10-9 .
- ^ Эллис, W (1839 г.). Полинезийские исследования . Том. 4. Лондон: Фишер, Джексон. ISBN 978-1-4325-4966-4 .
- ^ Мамфорд, Дэвид (1971). Исследования капитана Джеймса Кука в Тихом океане . Нью-Йорк: Dover Publications. ISBN 978-0-486-22766-5 .
- ^ Швабе, Кэлвин В. (1979). Бесподобная кухня . Университет Вирджинии Пресс. п. 168 . ISBN 978-0-8139-1162-5 .
- ^ Фредерик Вуд-Джонс (февраль 1931 г.). «Черепные признаки гавайской собаки». Журнал маммологии . 12 (1): 39–41. дои : 10.2307/1373802 . JSTOR 1373802 .
- ^ Хунг, Х.; Карсон, Майк Т.; Беллвуд, Питер; и др. (2011). «Первое поселение Отдаленной Океании: от Филиппин до Марианских островов» . Античность . 85 (329): 909–926. дои : 10.1017/S0003598X00068393 . S2CID 163099583 .
- ^ Осборн, Дуглас (1966). Археология островов Палау . Бюллетень музея Бернис П. Бишоп. Том. 230. с. 29. ISBN 978-0-910240-58-1 .
- ^ Jump up to: а б Урбан, Манфред (1961). Домашние животные полинезийцев . Геттинген: Ханцшель.
- ^ Jump up to: а б Матису-Смит, Элизабет (февраль 2015 г.). «Древняя ДНК и населенные пункты Тихого океана: обзор». Журнал эволюции человека . 79 : 93–104. дои : 10.1016/j.jhevol.2014.10.017 . ПМИД 25556846 .
- ^ Jump up to: а б с д Форстер, Иоганн Рейнхольд (1778). Наблюдения, сделанные во время кругосветного путешествия . п. 188.
- ^ Шарп, Эндрю (1964). Древние путешественники в Полинезии . Беркли: Издательство Калифорнийского университета. п. 120.
- ^ «Остров Питкэрн» . Азиатский журнал и ежемесячный реестр для британской и зарубежной Индии, Китая и Австралии . 10:38 . 1820.
- ^ Jump up to: а б Ардалан, Арман; Оскарссон, Маттиас ЧР; ван Аш, Барбара; Рабаконандриания, Элизабет Рабаконандриания; Саволайнен, Питер (2015). «Африканское происхождение мадагаскарских собак выявлено с помощью анализа мтДНК» . Королевское общество открытой науки . 2 (5): 140552. Бибкод : 2015RSOS....240552A . дои : 10.1098/rsos.140552 . ПМЦ 4453261 . ПМИД 26064658 .
- ^ Jump up to: а б ван Аш, Барбара (12 июня 2015 г.). «Мадагаскарские собаки родом из Африки, в отличие от своих человеческих собратьев» . Разговор . ООО «Разговор Медиа Групп» . Проверено 15 января 2019 г.
- ^ Jump up to: а б Томсон, Вики А.; Лебрассер, Офелия; Остин, Джереми Дж.; Хант, Терри Л.; Берни, Дэвид А.; Денхэм, Тим; Роуленс, Николас Дж.; Вуд, Джейми Р.; Гонгора, Хайме; Гирдланд Флинк, Лайнус; Линдерхольм, Анна; Добни, Кейт; Ларсон, Грегер; Купер, Алан (апрель 2014 г.). «Использование древней ДНК для изучения происхождения и расселения предков полинезийских кур по Тихому океану» . Труды Национальной академии наук . 111 (13): 4826–4831. Бибкод : 2014PNAS..111.4826T . дои : 10.1073/pnas.1320412111 . ПМЦ 3977275 . ПМИД 24639505 .
- ^ Jump up to: а б Пайпер, Филип Дж. (2017). «Происхождение и появление первых домашних животных на материковой и островной Юго-Восточной Азии: сложная и развивающаяся история» . В Пайпер, Филип Дж.; Мацумура, Хирофуми; Булбек, Дэвид (ред.). Новые перспективы в предыстории Юго-Восточной Азии и Тихого океана . Южная земля. Том. 45. АНУ Пресс. ISBN 9781760460945 .
- ^ Jump up to: а б Гровс, Колин П. 2006. Одомашненные и комменсальные млекопитающие Австронезии и их истории . В Беллвуд, Питер; Фокс, Джеймс Дж; Трайон, Даррелл, ред. (2006). Австронезийцы: исторические и сравнительные перспективы . АНУ Пресс. дои : 10.22459/А.09.2006 . hdl : 2027/mdp.39015051647942 . ISBN 9780731521326 .
- ^ Томсон, Вики; Аплин, Кен П.; Купер, Алан; Хише, Сьюзен; Сузуки, Хитоши; Марианто, Ибну; Да, Грейс; Доннеллан, Стивен К. (17 марта 2014 г.). Лалуэса-Фокс, Карлес (ред.). «Молекулярно-генетические данные о месте происхождения тихоокеанской крысы Rattus exulans» . ПЛОС ОДИН . 9 (3): е91356. Бибкод : 2014PLoSO...991356T . дои : 10.1371/journal.pone.0091356 . ISSN 1932-6203 . ПМЦ 3956674 . ПМИД 24637896 .
- Одомашнивание отдельных видов
- Сельское хозяйство в Юго-Восточной Азии
- Сельское хозяйство в Океании
- Сельское хозяйство на Мадагаскаре
- Сельское хозяйство на Коморских островах
- Списки растений по местоположению
- Списки животных по местонахождению
- Австронезийские народы
- Австронезийская культура
- Культуры из Азии
- Культуры из Австралазии
- Культуры, происходящие из Тихого океана
- История сельского хозяйства
- Доисторическое земледелие
- Флора Полинезии
- Флора Малайзии
- Флора Меланезии
- Флора Юго-Восточной Азии
- Флора Океании
- Флора Микронезии
- Флора Тайваня
- Флора Мадагаскара
- Флора Коморских островов
- Фауна Океании
- Фауна Юго-Восточной Азии
- Фауна Мадагаскара
- Фауна Коморских островов
- Фауна Тайваня
- Флора Новой Гвинеи
- Фауна Новой Гвинеи
- Биота Новой Зеландии
- Австронезийское сельское хозяйство