Рецептор клеточной поверхности

Рецепторы клеточной поверхности ( мембранные рецепторы , трансмембранные рецепторы ) представляют собой рецепторы которые встроены в плазматическую мембрану клеток , . [ 1 ] Они действуют в передаче сигналов клеток путем получения (связывания с) внеклеточными молекулами . Они представляют собой специализированные интегральные мембранные белки , которые допускают связь между клеткой и внеклеточным пространством . Внеклеточными молекулами могут быть гормоны , нейротрансмиттеры , цитокины , факторы роста , молекулы клеточной адгезии или питательные вещества ; Они реагируют с рецептором, чтобы вызвать изменения в метаболизме и активности клетки. В процессе сигнала передачи связывание лиганда влияет на каскадное химическое изменение через клеточную мембрану.
Структура и механизм
[ редактировать ]Многие мембранные рецепторы являются трансмембранными белками . Есть различные виды, в том числе гликопротеины и липопротеины . [ 2 ] Сотни разных рецепторов известны, и многие другие еще не изучены. [ 3 ] [ 4 ] Трансмембранные рецепторы обычно классифицируются на основе их третичной (трехмерной) структуры. Если трехмерная структура неизвестна, их можно классифицировать на основе топологии мембраны . В самых простых рецепторах полипептидные цепи один раз пересекают липидный бислой , в то время как другие, такие как рецепторы, связанные с G-белком , пересекают до семи раз. Каждая клеточная мембрана может иметь несколько видов мембранных рецепторов с различными распределениями поверхности. Один рецептор также может быть по -разному распределен в разных положениях мембраны, в зависимости от вида мембранной и клеточной функции. Рецепторы часто кластерируются на поверхности мембраны, а не равномерно распределены. [ 5 ] [ 6 ]
Механизм
[ редактировать ]Были предложены две модели, чтобы объяснить механизм действия трансмембранных рецепторов.
- Димеризация : модель димеризации предполагает, что до связывания лиганда рецепторы существуют в мономерной форме. Когда происходит связывание агониста, мономеры объединяются с образованием активного димера .
- Вращение : связывание лиганда с внеклеточной частью рецептора индуцирует вращение (конформационное изменение) части трансмембранных спиралей рецептора. Вращение изменяет, какие части рецептора подвергаются воздействию на внутриклеточной стороне мембраны, изменяя то, как рецептор может взаимодействовать с другими белками внутри клетки. [ 7 ]
Домены
[ редактировать ]P = плазматическая мембрана
I = внутриклеточное пространство
Трансмембранные рецепторы в плазматической мембране обычно можно разделить на три части.
Внеклеточные домены
[ редактировать ]Внеклеточный домен просто внешне из клетки или органелл . Если полипептидная цепь несколько раз пересекает бислой, внешний домен включает петли, переплетенные через мембрану. По определению, основной функцией рецептора является распознавание и реагирование на тип лиганда. Например, нейротрансмиттер , гормон или атомные ионы могут связываться с внеклеточным доменом в виде лиганда, связанного с рецептором. Клото - это фермент, который влияет на рецептор для распознавания лиганда ( FGF23 ).
Трансмембранные домены
[ редактировать ]Два наиболее распространенных класса трансмембранных рецепторов-это GPCR и однопроходные трансмембранные белки . [ 8 ] [ 9 ] В некоторых рецепторах, таких как никотиновый ацетилхолиновый рецептор , трансмембранный домен образует белковую пор через мембрану или вокруг ионного канала . После активации внеклеточного домена путем связывания соответствующего лиганда, пор становится доступной для ионов, которые затем распространяются. У других рецепторов трансмембранные домены подвергаются конформационному изменению при связывании, которое влияет на внутриклеточные условия. В некоторых рецепторах, таких как члены суперсемейства 7TM , трансмембранная домен включает в себя лиганд -связывающий карман.
Внутриклеточные домены
[ редактировать ]Внутриклеточный (или цитоплазматический ) домен рецептора взаимодействует с внутренней частью клетки или органеллы, передавая сигнал. Для этого взаимодействия есть два фундаментальных пути:
- Внутриклеточный домен связывается с помощью белковых взаимодействий с эффекторными белками , которые, в свою очередь, передают сигнал в пункт назначения.
- С ферментными рецепторами внутриклеточный домен обладает ферментативной активностью . Часто это активность тирозинкиназы . Ферментативная активность также может быть связана с ферментом, связанным с внутриклеточным доменом.
Сигнальная трансдукция
[ редактировать ]
Процессы трансдукции сигнала через мембранные рецепторы включают внешние реакции, в которых лиганд связывается с мембранным рецептором и внутренними реакциями, в которых запускается внутриклеточный ответ. [ 10 ] [ 11 ]
Передача сигнала с помощью мембранных рецепторов требует четырех частей:
- Внеклеточная сигнальная молекула: внеклеточная сигнальная молекула продуцируется одной ячейкой и, по крайней мере, способна перемещаться в соседние клетки.
- Рецепторный белок: клетки должны иметь белки рецептора клеточной поверхности, которые связываются с сигнальной молекулой и передают внутрь в клетку.
- Внутриклеточные сигнальные белки: они передают сигнал в органеллы клетки. Связывание молекулы сигнала с рецепторным белком будет активировать внутриклеточные сигнальные белки, которые инициируют сигнальный каскад.
- Целевые белки: конформации или другие свойства целевых белков изменяются, когда сигнальный путь активен, и изменяет поведение клетки. [ 11 ]

Мембранные рецепторы в основном разделяются на структуру и функцию на 3 класса: рецептор связанного ионного канала ; Рецептор , связанный с ферментом ; и G-белок-рецептор .
- Связанные ионные рецепторы имеют ионные каналы для анионов и катионов и представляют собой большое семейство многопроседающих трансмембранных белков. Они участвуют в быстрых событиях передачи сигналов, обычно встречающихся в электрически активных клетках, таких как нейроны . Их также называют лиганд-управляемыми ионными каналами . Открытие и закрытие ионных каналов контролируется нейротрансмиттерами .
- Ферменты, связанные с рецепторами, являются самими ферментами, либо непосредственно активируют ассоциированные ферменты. Обычно это однопроходные трансмембранные рецепторы, а ферментативный компонент рецептора сохраняется внутриклеточным. Большинство ферментных рецепторов или ассоциируются с протеинкиназами.
- G-белковые рецепторы являются интегральными мембранными белками, которые обладают семь трансмембранных спиралей. Эти рецепторы активируют G-белок при связывании агониста , и G-белок опосредует воздействие рецептора на внутриклеточные сигнальные пути.
Ионно-канальный рецептор
[ редактировать ]Во время события передачи сигнала в нейроне нейротрансмиттер связывается с рецептором и изменяет конформацию белка. Это открывает ионный канал, позволяя внеклеточному ионам в клетку. Ионная проницаемость плазматической мембраны изменяется, и это превращает внеклеточный химический сигнал в внутриклеточный электрический сигнал, который изменяет возбудимость клеток . [ 12 ]
Рецептор ацетилхолина является рецептором, связанным с катионным каналом. Белок состоит из четырех субъединиц: субъединицы альфа (α), бета (β), гамма (γ) и дельта (Δ). Существует две α -субъединицы, с одним ацетилхолина сайтом связывания . Этот рецептор может существовать в трех конформациях. Закрытое и незанятое состояние является нативной конформацией белка. В качестве двух молекул ацетилхолина оба связываются с сайтами связывания на α -субъединицах, конформация рецептора изменяется и открывается затвор, что позволяет проникнуть во многих ионов и малых молекул. Тем не менее, это открытое и оккупированное состояние длится только в течение небольшой продолжительности, а затем ворота закрыты, становясь закрытым и оккупированным состоянием. Две молекулы ацетилхолина скоро будут диссоциация от рецептора, возвращая его в нативное закрытое и незанятое состояние. [ 13 ] [ 14 ]
Фермент, связанные с рецепторами
[ редактировать ]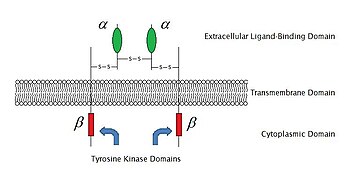
По состоянию на 2009 год существует 6 известных типов ферментных рецепторов : рецепторные тирозинкиназы ; Рецепторы, связанные с тирозинкиназой; Рецепторные тирозинфосфатазы ; Рецептор серин / треонинкиназы ; Рецептор гуанилилциклазы и связанные с гистидинкиназой рецепторы. Рецепторные тирозинкиназы имеют самое большое население и самое широкое применение. Большинство этих молекул являются рецепторами для факторов роста , таких как эпидермальный фактор роста (EGF), фактор роста, происходящий из тромбоцитов (PDGF), фактор роста фибробластов (FGF), фактор роста гепатоцитов (HGF), фактор роста нервов (NGF) и гормоны такие как инсулин . Большинство из этих рецепторов будут димеризировать после связывания с их лигандами, чтобы активировать дальнейшие передачи сигнала. Например, после рецептора эпидермального фактора роста (EGF) с его лигандом EGF, два рецептора димеризируют, а затем подвергаются фосфорилированию остатков тирозина связывается в ферментной части каждой молекулы рецептора. Это активирует тирозинкиназу и катализирует дальнейшие внутриклеточные реакции.
G-белковые рецепторы
[ редактировать ]G-белковые рецепторы составляют большие белковые семейства трансмембранных рецепторов. Они встречаются только у эукариот . [ 15 ] Лиганды , которые связывают и активируют эти рецепторы, включают в себя: фоточувствительные соединения, запахи , феромоны , гормоны и нейротрансмиттеры . Они различаются по размеру от мелких молекул до пептидов и больших белков . G-белковые рецепторы участвуют во многих заболеваниях и, следовательно, являются мишенями многих современных лекарственных препаратов. [ 16 ]
Существуют два основных пути передачи сигнала, включающих рецепторы, связанные с G-белком: сигнальный путь CAMP и сигнальный путь фосфатидилинозитола . [ 17 ] Оба опосредованы активацией G -белка . G-белок представляет собой тримерный белок, с тремя субъединицами, обозначенными как α, β и γ. В ответ на активацию рецептора α -субъединица высвобождает связанный гуанозин дифхосфат (GDP), который смещается с помощью гуанозин -трифосфата (GTP), тем самым активируя α -субъединицу, которая затем диссоциирует из субъединиц β и γ. Активированная α -субъединица может непосредственно влиять на внутриклеточные сигнальные белки или целевые функциональные белки.
Мембранная рецепторная болезнь
[ редактировать ]Если мембранные рецепторы денатурированы или дефицитны, передача сигнала может быть затруднена и вызвать заболевания. Некоторые заболевания вызваны нарушениями функции мембранных рецепторов. Это связано с дефицитом или деградацией рецептора посредством изменений в генах, которые кодируют и регулируют рецепторный белок. Мембранный рецептор TM4SF5 влияет на миграцию печени и гепатому . [ 18 ] Кроме того, рецептор кортикального NMDA влияет на текучесть мембраны и изменяется при болезни Альцгеймера. [ 19 ] Когда клетка инфицирует не развитым вирусом, вирус сначала связывается со специфическими мембранными рецепторами, а затем передает себя или субвирурный компонент с цитоплазматической стороной клеточной мембраны. В случае полиовируса известно, что взаимодействия с рецепторами вызывают конформационные перестройки, называемый VP4 , выделяют белок вириона которые . Предполагается, что конформационные изменения, вызванные рецепторным связыванием, приводят к прикреплению миристической кислоты на VP4 и образованию канала для РНК.
Структурный дизайн лекарств
[ редактировать ]
С помощью таких методов, как рентгеновская кристаллография и ЯМР-спектроскопия , информация о трехмерных структурах молекул-мишеней резко возросла, и поэтому имеет структурную информацию о лигандах. Это способствует быстрому разработке структурного дизайна лекарств . Некоторые из этих новых препаратов нацелены на мембранные рецепторы. Текущие подходы к структурной конструкции лекарств можно разделить на две категории. Первая категория посвящена определению лигандов для данного рецептора. Это обычно выполняется с помощью запросов базы данных, биофизического моделирования и построения химических библиотек. В каждом случае большое количество потенциальных молекул лиганда подвергается скринингу, чтобы найти тех, кто устанавливает связывающий карман рецептора. Этот подход обычно называют лигандным дизайном лекарств. Ключевым преимуществом поиска базы данных является то, что она экономит время и питание для получения новых эффективных соединений. Другой подход дизайна лекарств на основе структуры-это комбинаторное отображение лигандов, которое называется дизайном лекарств на основе рецепторов. В этом случае молекулы лиганда спроектированы в пределах ограничений связующего кармана путем сборки мелких кусочков поэтапно. Эти части могут быть атомами или молекулами. Ключевым преимуществом такого метода является то, что новые структуры могут быть обнаружены. [ 20 ] [ 21 ] [ 22 ]
Другие примеры
[ редактировать ]- Адренергический рецептор
- Обонятельные рецепторы
- Рецепторные тирозинкиназы
- Рецептор эпидермального фактора роста
- Рецептор инсулина
- Рецепторы фактора роста фибробластов ,
- Рецепторы с высокой аффинностью нейротрофина
- Рецепторы эфрина
- Интегрины
- Рецептор фактора роста низкого аффинного нерва
- Рецептор NMDA
- Несколько иммунных рецепторов
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ «9.3: сигнальные молекулы и клеточные рецепторы - типы рецепторов» . Биология либретекса . 12 июля 2018 года . Получено 24 июля 2023 года .
- ^ Cuatrecasas P. (1974). «Мембранные рецепторы». Ежегодный обзор биохимии . 43 : 169–214. doi : 10.1146/annurev.bi.43.070174.001125 . PMID 4368906 . S2CID 44727052 .
- ^ Dautzenberg FM, Hauger RL (февраль 2002 г.). «Семья пептидов CRF и их рецепторы: все же больше партнеров обнаружили». Тенденции Фармакол. Наука 23 (2): 71–7. doi : 10.1016/s0165-6147 (02) 01946-6 . PMID 11830263 .
- ^ Rivière S, Challet L, Fluegge D, Spehr M, Rodriguez I (май 2009 г.). «Формил-пептидные рецепторные белки-это новое семейство хемосенсоров из вомероназальных». Природа . 459 (7246): 574–7. doi : 10.1038/nature08029 . PMID 19387439 .
- ^ Ротберг Кг; Ying ys; Камен Ба; Андерсон Р.Г. (1990). «Холестерин контролирует кластеризацию мембранного рецептора, заправленного гликофосфолипидом, для 5-метилтетрагидрофолата» . Журнал клеточной биологии . 111 (6): 2931–2938. doi : 10.1083/jcb.111.6.2931 . PMC 2116385 . PMID 2148564 .
- ^ Джейкобсон С.; Côté PD; Росси С.Г.; Rotundo RL; Carbonetto S. (2001). «Комплекс дистрогликана необходим для стабилизации кластеров рецепторов ацетилхолина в нервно -мышечных соединениях и образования синаптической базарной мембраны» . Журнал клеточной биологии . 152 (3): 435–450. doi : 10.1083/jcb.152.3.435 . PMC 2195998 . PMID 11157973 .
- ^ Maruyama, Ichiro N. (2015-09-01). «Активация трансмембранных рецепторов поверхности клеток с помощью общего механизма?« Модель вращения » » . Биологии . 37 (9): 959–967. doi : 10.1002/bies.201500041 . ISSN 1521-1878 . PMC 5054922 . PMID 26241732 .
- ^ Суперсемейства однопроходных трансмембранных рецепторов в базе данных мембраномов
- ^ Суперсемейства трансмембранных белковых лигандов однопроходных белков и регуляторы рецепторов в базе данных мембраны
- ^ Ullricha A., Schlessingerb J.; Schlessinger, J (1990). «Сигнальная трансдукция рецепторами с активностью тирозинкиназы». Клетка . 61 (2): 203–212. doi : 10.1016/0092-8674 (90) 90801-K . PMID 2158859 .
- ^ Jump up to: а беременный Кеннет Б. Стори (1990). Функциональный метаболизм . Wiley-Ieee. С. 87–94. ISBN 978-0-471-41090-4 .
- ^ Hille B. (2001). Ионные каналы возбудимых мембран . Сандерленд, штат Массачусетс 978-0-87893-321-1 .
- ^ Miyazawa A.; Fujiyoshi y.; Unwin N. (2003). «Структура и стробирующий механизм пор -рецептора ацетилхолинового рецептора». Природа . 423 (6943): 949–955. doi : 10.1038/nature01748 . PMID 12827192 .
- ^ Акабас М.Х.; Стауффер да; Сюй М.; Карлин А. (1992). «Структура канала ацетилхолинового рецептора, исследованная у мутантов-проститирования цистеина». Наука . 258 (5080): 307–310. doi : 10.1126/science.13844130 . PMID 1384130 .
- ^ Кинг Н., Хиттингер К.Т., Кэрролл С.Б. (2003). «Эволюция ключевых клеточных сигналов и семейств адгезии белков предшествует происхождению животных». Наука . 301 (5631): 361–3. doi : 10.1126/science.1083853 . PMID 12869759 .
- ^ Filmore, David (2004). «Это мир GPCR» . Современное открытие наркотиков . 2004 (ноябрь): 24–28.
- ^ Гилман А.Г. (1987). «G-белки: преобразователи сигналов, сгенерированных рецептором». Ежегодный обзор биохимии . 56 : 615–649. doi : 10.1146/annurev.bi.56.070187.003151 . PMID 3113327 . S2CID 33992382 .
- ^ Müller-Pillascha F.; Wallrappa C.; Lachera U.; Friessb H.; Büchlerb M.; Adlera G.; Гресс Т.М. (1998). «Идентификация нового антигена TM4SF5, ассоциированного с опухолем, и его экспрессия при раке человека». Ген . 208 (1): 25–30. doi : 10.1016/s0378-1119 (97) 00633-1 . PMID 9479038 .
- ^ Scheeruer K.; Marasb A.; Gattazb wf; Cairnsc N.; Förstlb H.; Мюллер мы (1996). «Свойства кортикального рецептора NMDA и текучесть мембраны удерживаются при болезни Альцгеймера» Деменция 7 (4): 210–2 Doi : 10.1159/ 000106881 8835885PMID
- ^ Ван Р.; Gao y.; Лай Л. (2000). «Ligbuilder: многоцелевая программа для структурного дизайна лекарств». Журнал молекулярного моделирования . 6 (7–8): 498–516. doi : 10.1007/s0089400060498 .
- ^ Schneider G.; Фехнер У. (2005). «Компьютерный дизайн de novo молекул, подобные лекарствам». Природа Обзоры наркотиков . 4 (8): 649–663. doi : 10.1038/nrd1799 . PMID 16056391 .
- ^ Jorgensen WL (2004). «Многочисленные роли вычислений в обнаружении лекарств». Наука . 303 (5665): 1813–1818. doi : 10.1126/science.1096361 . PMID 15031495 . S2CID 1307935 .
Внешние ссылки
[ редактировать ]- базы данных Iuphar GPCR Архив 2019-03-23 на The Wayback Machine
- Cell+Surface+Рецепторы в Национальной библиотеке медицинской библиотеки Медицинской библиотеки США (Mesh)