Фас-лиганд
| ФАСЛГ | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | FASLG , ALPS1B, APT1LG1, APTL, CD178, CD95-L, CD95L, FASL, TNFSF6, TNLG1A, лиганд Fas | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 134638 ; МГИ : 99255 ; Гомологен : 533 ; GeneCards : FASLG ; ОМА : FASLG – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





Лиганд Fas ( FASL или CD95 типа II, L) представляет собой трансмембранный белок экспрессируемый на различных типах клеток, включая цитотоксические Т-лимфоциты , моноциты, нейтрофилы , эпителиальные клетки сосудов клетки молочной железы, эндотелиальные и естественные клетки-киллеры (NK) . Он связывается со своим рецептором , называемым рецептором FAS (также называемым CD95), и играет решающую роль в регуляции иммунной системы и индукции апоптоза — запрограммированной гибели клеток. [ 5 ]
Конструктивные особенности
[ редактировать ]Лиганд Fas или FasL представляет собой трансмембранный белок типа II, принадлежащий к суперсемейству факторов некроза опухоли (TNFSF). Он гомотримерен , что означает, что он состоит из трех идентичных полипептидов. Он имеет длинный цитоплазматический домен, область стебля, трансмембранный домен (TM), домен гомологии TNF (THD), ответственный за гомотримеризацию. Включая С-концевую область, участвующую в связывании с CD95, также известную как рецептор fas. [ 6 ] [ 7 ]
FasL связывается с fas, что приводит к образованию сборки fas:FasL. Это взаимодействие инициирует образование сигнального комплекса, индуцирующего смерть, что приводит к апоптозу. [ 6 ]
FasL экспрессируется на различных типах клеток, включая Т-клетки, естественные клетки-киллеры, моноциты, нейтрофилы и клетки эндотелия сосудов. FasL существует как в мембраноскрепленной, так и в растворимой формах. [ 5 ]
Рецепторы
[ редактировать ]- FasR : Рецептор Fas (FasR), или CD95 , является наиболее изученным членом семейства рецепторов смерти. Ген расположен на 10-й хромосоме у человека и 19-й у мышей. Предыдущие отчеты идентифицировали целых восемь вариантов сплайсинга, которые транслируются в семь изоформ белка. Многие из этих изоформ связаны с редкими гаплотипами , которые обычно связаны с болезненным состоянием. Рецептор Fas, индуцирующий апоптоз, получил название изоформы 1 и представляет собой трансмембранный белок 1 типа . Он состоит из трех богатых цистеином псевдоповторов, трансмембранного домена и внутриклеточного домена смерти. [ 8 ]
- DcR3 : Рецептор-ловушка 3 (DcR3) представляет собой недавно открытый рецептор-ловушку из суперсемейства факторов некроза опухоли , который связывается с FasL, LIGHT и TL1A . DcR3 представляет собой растворимый рецептор, который не обладает способностью передавать сигнал (следовательно, является «приманкой») и действует для предотвращения взаимодействий FasR-FasL путем конкурентного связывания с мембраносвязанными лигандами Fas и делает их неактивными. [ 9 ]
Клеточная сигнализация и механизм
[ редактировать ]Сигнальный путь Fas включает активацию апоптоза (запрограммированной гибели клеток). Это происходит за счет взаимодействия рецептора Fas и лиганда Fas. Как уже упоминалось, лиганд Fas/FasL представляет собой трансмембранный белок типа II, который может существовать как в мембранозакрепленной, так и в растворимой формах. Взаимодействие между FasR на соседней клетке и закрепленным на мембране FasL приводит к тримеризации, образуя сигнальный комплекс, индуцирующий смерть (DISC). [ 10 ]
После последующей агрегации домена смерти (DD) рецепторный комплекс интернализуется через клеточный эндосомальный механизм. Это позволяет адапторной молекуле, связанной с Fas-доменом смерти (FADD), связываться с доменом смерти (DD) Fas через свой собственный домен смерти (DD). FADD также содержит эффекторный домен смерти (DED) рядом с его аминоконцом, который облегчает связывание с DED FADD -подобного ICE ( FLICE ), чаще называемого каспазой-8 . Затем FLICE может самоактивироваться посредством протеолитического расщепления на субъединицы p10 и p18, две из которых образуют активный фермент гетеротетрамер. Активная каспаза-8 затем высвобождается из DISC в цитозоль, где она расщепляет другие эффекторные каспазы, что в конечном итоге приводит к деградации ДНК, образованию мембранных пузырей и другим признакам апоптоза. [ 11 ] [ 10 ]
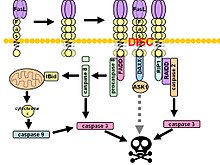
В некоторых сообщениях предполагается, что внешнего пути Fas достаточно, чтобы вызвать полный апоптоз в определенных типах клеток посредством сборки сигнального комплекса, индуцирующего смерть (DISC), и последующей активации каспазы-8. [ 10 ] Эти клетки получили название клеток типа 1 и характеризуются неспособностью антиапоптотических членов семейства Bcl-2 (а именно Bcl-2 и Bcl-xL ) защищать от Fas-опосредованного апоптоза. Охарактеризованные клетки типа 1 включают H9, CH1, SKW6.4 и SW480, все из которых представляют собой линии лимфоцитов, за исключением SW480, который относится к линии аденокарциномы толстой кишки. [ 10 ]
Более того, пути в сигнальном каскаде Fas демонстрируют перекрестные помехи. В большинстве типов клеток каспаза-8 катализирует расщепление проапоптотического BH3 белка , содержащего только BH3, до его усеченной формы tBid. Только BH-3 члены семейства Bcl-2 взаимодействуют исключительно с антиапоптотическими членами семейства (Bcl-2, Bcl-xL), позволяя Bak и Bax перемещаться на внешнюю митохондриальную мембрану, тем самым обеспечивая ее проницаемость и облегчая высвобождение про -апоптотические белки, такие как цитохром с и Smac/DIABLO , антагонист ингибиторов белков апоптоза ( IAP ). [ 10 ]
Кроме того, белок c-FLIP, структурно напоминающий каспазу-8, но лишенный ферментативной активности, играет двойную роль в Fas-индуцированном апоптозе. Считается, что в низких концентрациях c-FLIP способствует активации каспазы-8. Возможно, это связано с тем, что каспаза-8 связывается с c-FLIP с более высоким сродством, чем с самой собой (гомодимеризация каспазы-8). Однако в высоких концентрациях c-FLIP снижает протеолитическую активность каспазы-8, потенциально конкурируя за связывание с FADD. Эта двойная роль подчеркивает сложность передачи сигналов Fas и ее регуляции с помощью c-FLIP в различных концентрациях. [ 10 ]
Функция апоптоза в иммунной системе
[ редактировать ]Апоптоз , запускаемый связыванием лиганда FasR-Fas, играет фундаментальную роль в регуляции иммунной системы . В его функции входят:
- Т-клеток Гомеостаз : активация Т-клеток приводит к экспрессии ими лиганда Fas. Т-клетки изначально устойчивы к Fas-опосредованному апоптозу во время клональной экспансии, но становятся все более чувствительными, чем дольше они активируются, что в конечном итоге приводит к гибели клеток, индуцированной активацией (AICD). Этот процесс необходим для предотвращения чрезмерного иммунного ответа и устранения аутореактивных Т-клеток. У людей и мышей с вредными мутациями Fas или Fas-лиганда развивается накопление аберрантных Т-клеток, что приводит к лимфаденопатии , спленомегалии и красной волчанке . [ 12 ]
- Цитотоксическая активность Т-клеток . Fas-индуцированный апоптоз и перфориновый путь являются двумя основными механизмами, с помощью которых цитотоксические Т-лимфоциты вызывают гибель клеток в клетках, экспрессирующих чужеродные антигены. [ 13 ]
- Иммунная привилегия : Клетки в иммунопривилегированных областях, таких как роговица или семенники, экспрессируют лиганд Fas и вызывают апоптоз инфильтрирующих лимфоцитов . Это один из многих механизмов, которые организм использует для установления и поддержания иммунных привилегий. [ 14 ]
- Материнская толерантность : Лиганд Fas может играть важную роль в предотвращении торговли лейкоцитами между матерью и плодом, хотя дефекты беременности еще не были связаны с дефектами системы лигандов Fas-Fas. [ 14 ]
- Контратака опухоли : Опухоли могут сверхэкспрессировать лиганд Fas и индуцировать апоптоз инфильтрирующих лимфоцитов , позволяя опухоли избежать воздействия иммунного ответа . [ 15 ] Повышение регуляции лиганда Fas часто происходит после химиотерапии , в результате которой опухолевые клетки приобретают устойчивость к апоптозу . [ 16 ]
Роль в болезни
[ редактировать ]Дефектный Fas-опосредованный апоптоз может привести к онкогенезу , а также к лекарственной устойчивости существующих опухолей. Зародышевая мутация Fas связана с аутоиммунным лимфопролиферативным синдромом (АЛПС), детским заболеванием апоптоза. [ 17 ]
Увеличение Fas-опосредованной передачи сигналов связано с патологией миелодиспластических синдромов низкого риска (МДС). [ 18 ] и глиобластома . [ 19 ]
Совсем недавно FasL-опосредованный апоптоз Т-клеток также был предложен как механизм, уклоняющийся от иммунитета, с помощью которого опухоли могут подавлять инфильтрацию Т-клеток, аналогично ингибирующим иммунным контрольным точкам, таким как PD-1 и CTLA-4 . [ 20 ] [ 21 ] [ 22 ]
Взаимодействия
[ редактировать ]Было показано, что лиганд Fas взаимодействует с:
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Ensembl выпуск 89: ENSG00000117560 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000000817 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б Криппнер-Хайденрайх А, Шойрих П (2006). «FasL и Fas. Типичные члены семейства лигандов и рецепторов TNF». Сигнализация Фас. Отдел медицинской разведки . Бостон, Массачусетс: Спрингер. дои : 10.1007/0-387-34573-6_1 . ISBN 0-387-34573-6 .
- ^ Jump up to: а б Левуан Н., Жан М., Легембр П. (2020). «Структура CD95, агрегация и передача сигналов в клетках» . Границы клеточной биологии и биологии развития . 8 : 314. дои : 10.3389/fcell.2020.00314 . ПМК 7214685 . ПМИД 32432115 .
- ^ Орлиник-младший, Вайшнав А.К., Элкон К.Б. (1999). «Структура и функция Fas/Fas-лиганда». Международные обзоры иммунологии . 18 (4): 293–308. дои : 10.3109/08830189909088485 . ПМИД 10626245 .
- ^ Лю В., Рамагопал У., Ченг Х., Бонанно Дж.Б., Торо Р., Бхосле Р. и др. (ноябрь 2016 г.). «Кристаллическая структура комплекса FasL человека и его рецептора-ловушки DcR3» . Структура . 24 (11): 2016–2023. doi : 10.1016/j.str.2016.09.009 . ПМИД 27806260 .
- ^ Шейх М.С., Форнас А.Дж. (август 2000 г.). «Рецепторы смерти и ловушки и p53-опосредованный апоптоз». Лейкемия . 14 (8): 1509–1513. дои : 10.1038/sj.leu.2401865 . ПМИД 10942251 . S2CID 12572810 .
- ^ Jump up to: а б с д и ж Штрассер А., Йост П.Дж., Нагата С. (февраль 2009 г.). «Множество ролей передачи сигналов рецептора ФАС в иммунной системе» . Иммунитет . 30 (2): 180–192. doi : 10.1016/j.immuni.2009.01.001 . ПМК 2956119 . ПМИД 19239902 .
- ^ Ёлку Э.С., Ширван Х., Аскенасы Н. (27 марта 2017 г.). «Взаимодействие Fas/Fas-лиганда как механизм иммунного гомеостаза и цитотоксичности β-клеток: усиление, а не нейтрализация для лечения диабета 1 типа» . Границы в иммунологии . 8 : 342. дои : 10.3389/fimmu.2017.00342 . ПМК 5366321 . ПМИД 28396667 .
- ^ Бойман О., Пертон Дж. Ф., Сур К. Д., Срент Дж. (июнь 2007 г.). «Цитокины и гомеостаз Т-клеток». Современное мнение в иммунологии . Активация лимфоцитов/Эффекторные функции лимфоцитов. 19 (3): 320–326. дои : 10.1016/j.coi.2007.04.015 . ПМИД 17433869 .
- ^ Андерсен М.Х., Шрама Д., Тор Стратен П., Беккер Дж.К. (январь 2006 г.). «Цитотоксические Т-клетки» . Журнал исследовательской дерматологии . 126 (1): 32–41. дои : 10.1038/sj.jid.5700001 . ПМИД 16417215 .
- ^ Jump up to: а б Ежак М., Бишоф П. (январь 2002 г.). «Апоптоз в плаценте человека в первом триместре беременности: роль в поддержании иммунных привилегий на границе между матерью и плодом и в ремоделировании трофобласта». Европейский журнал акушерства, гинекологии и репродуктивной биологии . 100 (2): 138–142. дои : 10.1016/S0301-2115(01)00431-6 . ПМИД 11750952 .
- ^ Игни Ф.Х., Краммер П.Х. (ноябрь 2005 г.). «Контратака опухоли: факт или вымысел?» . Иммунология рака, иммунотерапия . 54 (11): 1127–1136. дои : 10.1007/s00262-005-0680-7 . ПМЦ 11034178 . ПМИД 15889255 . S2CID 19331352 .
- ^ Пистритто Дж., Тришульо Д., Сечи С., Гаруфи А., Д'Орази Дж. (апрель 2016 г.). «Апоптоз как противораковый механизм: функция и дисфункция его модуляторов и целевые терапевтические стратегии» . Старение . 8 (4): 603–619. дои : 10.18632/aging.100934 . ПМЦ 4925817 . ПМИД 27019364 .
- ^ Лламби Ф, Грин ДР (февраль 2011 г.). «Апоптоз и онкогенез: давать и брать в семье BCL-2» . Текущее мнение в области генетики и развития . 21 (1): 12–20. дои : 10.1016/j.gde.2010.12.001 . ПМК 3040981 . ПМИД 21236661 .
- ^ Классенс Ю., Бускари Д., Дюпон Ж.М., Пикард Ф., Мелле Дж., Гиссельбрехт С. и др. (март 2002 г.). «Пролиферация и дифференциация эритроидных предшественников in vitro у пациентов с миелодиспластическими синдромами: доказательства Fas-зависимого апоптоза» . Кровь . 99 (5): 1594–1601. дои : 10.1182/blood.V99.5.1594 . ПМИД 11861273 .
- ^ Татибана О, Наказава Х, Лампе Дж, Ватанабэ К, Клейхуэс П, Огаки Х (декабрь 1995 г.). «Экспрессия Fas/APO-1 во время прогрессирования астроцитомы». Исследования рака . 55 (23): 5528–5530. ПМИД 7585627 .
- ^ Моц Г.Т., Санторо С.П., Ван Л.П., Гаррабрант Т., Ластра Р.Р., Хагеманн И.С. и др. (июнь 2014 г.). «FasL эндотелия опухоли устанавливает селективный иммунный барьер, способствующий толерантности в опухолях» . Природная медицина . 20 (6): 607–615. дои : 10.1038/нм.3541 . ПМК 4060245 . ПМИД 24793239 .
- ^ Чжу Дж., Повис де Тенбосше К.Г., Кане С., Колау Д., ван Барен Н., Луркин С. и др. (ноябрь 2017 г.). «Устойчивость к иммунотерапии рака, опосредованная апоптозом инфильтрирующих опухоль лимфоцитов» . Природные коммуникации . 8 (1): 1404. Бибкод : 2017NatCo...8.1404Z . дои : 10.1038/s41467-017-00784-1 . ПМК 5680273 . ПМИД 29123081 .
- ^ Лакинс М.А., Горани Э., Мунир Х., Мартинс К.П., Шилдс Дж.Д. (март 2018 г.). «Связанные с раком фибробласты индуцируют антигенспецифическую делецию CD8. + Т-клетки для защиты опухолевых клеток» . Nature Communications . 9 (1): 948. Бибкод : 2018NatCo...9..948L . doi : /s41467-018-03347-0 . PMC 5838096. 10.1038 PMID 29507342 .
- ^ Jump up to: а б с д Гаджате К., Моллинедо Ф (март 2005 г.). «Цитоскелет-опосредованный рецептор смерти и концентрация лигандов в липидных рафтах образуют кластеры, способствующие апоптозу при химиотерапии рака» . Журнал биологической химии . 280 (12): 11641–11647. дои : 10.1074/jbc.M411781200 . ПМИД 15659383 .
- ^ Jump up to: а б с Мишо О, Чопп Дж (июль 2003 г.). «Индукция апоптоза, опосредованного рецептором TNF I, посредством двух последовательных сигнальных комплексов». Клетка . 114 (2): 181–190. дои : 10.1016/s0092-8674(03)00521-x . ПМИД 12887920 . S2CID 17145731 .
- ^ Парлато С., Джаммариоли А.М., Логоцци М., Лозупоне Ф., Матаррезе П., Лучани Ф. и др. (октябрь 2000 г.). «Связь CD95 (APO-1/Fas) с актиновым цитоскелетом через эзрин в Т-лимфоцитах человека: новый регуляторный механизм апоптотического пути CD95» . Журнал ЭМБО . 19 (19): 5123–5134. дои : 10.1093/emboj/19.19.5123 . ПМК 302100 . ПМИД 11013215 .
- ^ Jump up to: а б с Гадими М.П., Санценбахер Р., Тиде Б., Венцель Дж., Цзин К., Пломанн М. и др. (май 2002 г.). «Идентификация партнеров взаимодействия цитозольной полипролиновой области лиганда CD95 (CD178)». Письма ФЭБС . 519 (1–3): 50–58. дои : 10.1016/s0014-5793(02)02709-6 . ПМИД 12023017 . S2CID 26765451 .
- ^ Jump up to: а б Венцель Дж., Санценбахер Р., Гадими М., Левицки М., Чжоу К., Каплан Д.Р. и др. (декабрь 2001 г.). «Множественные взаимодействия цитозольной полипролиновой области лиганда CD95: намеки на способность обратной передачи сигнала фактора смерти». Письма ФЭБС . 509 (2): 255–262. дои : 10.1016/s0014-5793(01)03174-x . ПМИД 11741599 . S2CID 33084576 .
- ^ Хане М., Лоуин Б., Пайч М., Беккер К., Чопп Дж. (октябрь 1995 г.). «Взаимодействие пептидов, полученных из лиганда Fas, с доменом Fyn-SH3» . Письма ФЭБС . 373 (3): 265–268. дои : 10.1016/0014-5793(95)01051-ф . ПМИД 7589480 . S2CID 24130275 .
- ^ Старлинг Г.К., Баджорат Дж., Эмсвилер Дж., Ледбеттер Дж.А., Аруффо А., Кинер П.А. (апрель 1997 г.). «Идентификация аминокислотных остатков, важных для связывания лиганда с Fas» . Журнал экспериментальной медицины . 185 (8): 1487–1492. дои : 10.1084/jem.185.8.1487 . ПМК 2196280 . ПМИД 9126929 .
- ^ Шнайдер П., Бодмер Дж.Л., Холлер Н., Маттманн С., Скудери П., Терских А. и др. (июль 1997 г.). «Характеристика взаимодействия Fas (Apo-1, CD95)-Fas-лиганда» . Журнал биологической химии . 272 (30): 18827–18833. дои : 10.1074/jbc.272.30.18827 . ПМИД 9228058 .
- ^ Ю К.Ю., Квон Б., Ни Дж., Чжай Ю., Эбнер Р., Квон Б.С. (май 1999 г.). «Недавно идентифицированный член суперсемейства рецепторов фактора некроза опухоли (TR6) подавляет LIGHT-опосредованный апоптоз» . Журнал биологической химии . 274 (20): 13733–13736. дои : 10.1074/jbc.274.20.13733 . ПМИД 10318773 .
- ^ Сюй Т.Л., Чанг Ю.К., Чен С.Дж., Лю Ю.Дж., Чиу А.В., Чио CC и др. (май 2002 г.). «Модуляция дифференцировки и созревания дендритных клеток с помощью рецептора-ловушки 3» . Журнал иммунологии . 168 (10): 4846–4853. дои : 10.4049/jimmunol.168.10.4846 . ПМИД 11994433 .
- ^ Питти Р.М., Марстерс С.А., Лоуренс Д.А., Рой М., Кишкель Ф.К., Дауд П. и др. (декабрь 1998 г.). «Геномная амплификация рецептора-ловушки для лиганда Fas при раке легких и толстой кишки». Природа . 396 (6712): 699–703. Бибкод : 1998Natur.396..699P . дои : 10.1038/25387 . ПМИД 9872321 . S2CID 4427455 .
Дальнейшее чтение
[ редактировать ]- Чой С., Бенвенист Э.Н. (январь 2004 г.). «Fas-лиганд/Fas-система в головном мозге: регулятор иммунных и апоптотических ответов». Исследования мозга. Обзоры исследований мозга . 44 (1): 65–81. дои : 10.1016/j.brainresrev.2003.08.007 . ПМИД 14739003 . S2CID 46587211 .
- Толструп М., Остергаард Л., Лаурсен А.Л., Педерсен С.Ф., Дах М. (апрель 2004 г.). «ВИЧ/ВИV ускользают от иммунного надзора: сосредоточьтесь на Нефе». Текущие исследования ВИЧ . 2 (2): 141–151. дои : 10.2174/1570162043484924 . ПМИД 15078178 .
Внешние ссылки
[ редактировать ]- Запись GeneReviews/NCBI/NIH/UW об аутоиммунном лимфопролиферативном синдроме
- Интернет-менделевское наследование у человека (OMIM): 601859
- Fas + Ligand + Protein Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
- Обзор всей структурной информации, доступной в PDB для UniProt : P48023 (член суперсемейства лигандов фактора некроза опухоли 6) на PDBe-KB .