Аденозинтрифосфат
 | |||
| |||
| Имена | |||
|---|---|---|---|
| Название ИЮПАК Аденозин 5'-(тетрагидротрифосфат) | |||
| Систематическое название ИЮПАК ТО 1 -{[(2R , 3S , 4R , 5R ) -5-(6-амино-9Н - пурин-9-ил)-3,4-дигидроксиоксолан-2-ил]метил}тетрагидротрифосфат | |||
| Идентификаторы | |||
| |||
3D model ( JSmol ) | |||
| ЧЭБИ | |||
| ЧЕМБЛ | |||
| ХимическийПаук | |||
| Лекарственный Банк | |||
| Информационная карта ECHA | 100.000.258 | ||
| КЕГГ | |||
ПабХим CID | |||
| НЕКОТОРЫЙ | |||
Панель управления CompTox ( EPA ) | |||
| Характеристики | |||
| С 10 Ч 16 Н 5 О 13 П 3 | |||
| Молярная масса | 507.18 g/mol | ||
| Плотность | 1,04 г/см 3 (динатриевая соль) | ||
| Температура плавления | 187 ° C (369 ° F; 460 К) динатриевая соль; разлагается | ||
| Кислотность ( pKa ) | 0.9, 1.4, 3.8, 6.5 | ||
| УФ-видимое излучение (λ макс .) | 259 нм [1] | ||
| Поглощение | ε 259 = 15,4 мМ −1 см −1 [1] | ||
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |||

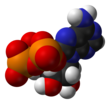

Аденозинтрифосфат ( АТФ ) — нуклеотид [2] который обеспечивает энергию для управления и поддержки многих процессов в живых клетках , таких как сокращение мышц , распространение нервных импульсов и химический синтез . Встречающийся во всех известных формах жизни , его часто называют «молекулярной денежной единицей » для внутриклеточной передачи энергии . [3]
При потреблении в метаболическом процессе АТФ превращается либо в аденозиндифосфат (АДФ), либо в аденозинмонофосфат (АМФ). Другие процессы восстанавливают АТФ. Он также является ДНК и и РНК предшественником используется в качестве кофермента . Среднестатистический взрослый человек ежедневно перерабатывает около 50 килограммов. [4]
С точки зрения биохимии АТФ классифицируется как нуклеозидтрифосфат , что указывает на то, что она состоит из трех компонентов: азотистого основания ( аденина ), сахарной рибозы и трифосфата .
Структура
[ редактировать ]АТФ состоит из аденина , присоединенного атомом азота #9 к 1'- углерода атому сахара ( рибозы ), который, в свою очередь, присоединен к 5'-атому углерода сахара к трифосфатной группе. Во многих реакциях, связанных с обменом веществ, адениновая и сахарная группы остаются неизменными, но трифосфат превращается в ди- и монофосфат, давая соответственно производные АДФ и АМФ . Три фосфорильные группы обозначены как альфа (α), бета (β) и, для терминального фосфата, гамма (γ). [5]
В нейтральном растворе ионизированный АТФ существует в основном в виде АТФ. 4− , с небольшой долей АТФ 3− . [6]
Связывание катионов металлов
[ редактировать ]Полианионный АТФ, обладающий потенциально хелатирующей полифосфатной группой, связывает катионы металлов с высоким сродством. Константа связывания Mg 2+
есть ( 9 554 ). [7] Связывание двухвалентного катиона , почти всегда магния , сильно влияет на взаимодействие АТФ с различными белками. Благодаря силе АТФ-Mg 2+ взаимодействия АТФ существует в клетке преимущественно в виде комплекса с Mg 2+
связаны с фосфатными кислородными центрами. [6] [8]
Второй ион магния имеет решающее значение для связывания АТФ в киназном домене. [9] Наличие магния 2+ регулирует активность киназы. [10] С точки зрения мира РНК интересно, что АТФ может переносить ион Mg, который катализирует полимеризацию РНК. [ нужна ссылка ]
Химические свойства
[ редактировать ]Соли АТФ можно выделить в виде бесцветных твердых веществ. [11]

АТФ стабилен в водных растворах при pH от 6,8 до 7,4 (в отсутствие катализаторов). При более экстремальных уровнях pH он быстро гидролизуется до АДФ и фосфата. Живые клетки поддерживают соотношение АТФ и АДФ на уровне десяти порядков от равновесия, при этом концентрации АТФ в пять раз превышают концентрацию АДФ. [12] [13] В контексте биохимических реакций связи POP часто называют высокоэнергетическими связями . [14]
Реактивные аспекты
[ редактировать ]Гидролиз АТФ до АДФ и неорганического фосфата.
- СПС 4- (вод) + Н 2 О (л) = АДФ 3- (водный раствор) + ГПО 2- (вод) + Н + (вода)
выделяет 20,5 килоджоулей на моль (4,9 ккал/моль) энтальпии . Это может отличаться в физиологических условиях, если реагент и продукты не находятся точно в этих состояниях ионизации. [15] Значения свободной энергии, высвобождаемой при отщеплении фосфатного (P i ) или пирофосфатного (PP i ) звена от АТФ при стандартных концентрациях 1 моль/л при pH 7, составляют: [16]
- АТФ + Н
2 O → АДФ + P i Δ G °' = -30,5 кДж/моль (-7,3 ккал/моль) - АТФ + Н
2 O → AMP + PP i Δ G °' = -45,6 кДж/моль (-10,9 ккал/моль)
Эти сокращенные уравнения при pH около 7 можно записать более явно (R = аденозил ):
- [РО-П(О) 2 -ОП(О) 2 -О-РО 3 ] 4− + Ч
2 О → [РО-П(О) 2 -О-РО 3 ] 3− + [ГПО 4 ] 2− + Ч + - [РО-П(О) 2 -ОП(О) 2 -О-РО 3 ] 4− + Ч
2 О → [РО-ПО 3 ] 2− + [НО 3 П-О-ПО 3 ] 3− + Ч +
В цитоплазматических условиях, когда соотношение АДФ/АТФ находится на 10 порядков от равновесия, ΔG составляет около -57 кДж/моль. [12]
Наряду с pH изменение свободной энергии гидролиза АТФ также связано с Mg. 2+ концентрации, от ΔG°' = -35,7 кДж/моль при Mg 2+ нулевой концентрации до ΔG°' = -31 кДж/моль при [Mg 2+ ] = 5 мМ. Более высокие концентрации Mg 2+ уменьшить свободную энергию, выделяющуюся в реакции за счет связывания Mg 2+ ионы к отрицательно заряженным атомам кислорода АТФ при рН 7. [17]
Производство из AMP и ADP
[ редактировать ]Производство, аэробные условия
[ редактировать ]Типичная внутриклеточная концентрация АТФ может составлять 1–10 мкмоль на грамм ткани у различных эукариот. [18] Дефосфорилирование АТФ и рефосфорилирование АДФ и АМФ происходят неоднократно в ходе аэробного метаболизма. [19]
АТФ может вырабатываться рядом различных клеточных процессов; Тремя основными путями у эукариот являются (1) гликолиз , (2) цикл лимонной кислоты / окислительное фосфорилирование и (3) бета-окисление . Общий процесс окисления глюкозы до углекислого газа , комбинация путей 1 и 2, известный как клеточное дыхание , производит около 30 эквивалентов АТФ из каждой молекулы глюкозы. [20]
аэробными Производство АТФ нефотосинтезирующими эукариотами происходит главным образом в митохондриях , которые составляют около 25% объема типичной клетки. [21]
Гликолиз
[ редактировать ]При гликолизе глюкоза и глицерин метаболизируются до пирувата . При гликолизе образуются два эквивалента АТФ посредством фосфорилирования субстрата , катализируемого двумя ферментами: фосфоглицераткиназой (PGK) и пируваткиназой . два эквивалента никотинамидадениндинуклеотида Также образуются (НАДН), которые могут окисляться через цепь переноса электронов и приводить к образованию дополнительного АТФ с помощью АТФ-синтазы . Пируват, образующийся в качестве конечного продукта гликолиза, является субстратом цикла Кребса . [22]
Гликолиз рассматривается как состоящий из двух фаз по пять стадий в каждой. На этапе 1, «подготовительной фазе», глюкоза превращается в 2-d-глицеральдегид-3-фосфат (g3p). Один АТФ вкладывается в этап 1, а другой АТФ вкладывается в этап 3. Этапы 1 и 3 гликолиза называются «этапами прайминга». На этапе 2 два эквивалента g3p превращаются в два пирувата. На этапе 7 производятся два АТФ. Кроме того, на этапе 10 производятся еще два эквивалента АТФ. На этапах 7 и 10 АТФ генерируется из АДФ. В цикле гликолиза образуется сеть из двух АТФ. Путь гликолиза позже связан с циклом лимонной кислоты, который производит дополнительные эквиваленты АТФ. [ нужна ссылка ]
Регулирование
[ редактировать ]При гликолизе гексокиназа непосредственно ингибируется ее продуктом, глюкозо-6-фосфатом, а пируваткиназа ингибируется самой АТФ. Основной контрольной точкой гликолитического пути является фосфофруктокиназа (ФФК), которая аллостерически ингибируется высокими концентрациями АТФ и активируется высокими концентрациями АМФ. Ингибирование ПФК АТФ необычно, поскольку АТФ также является субстратом реакции, катализируемой ПФК; активная форма фермента представляет собой тетрамер , существующий в двух конформациях, только одна из которых связывает второй субстрат фруктозо-6-фосфат (F6P). Белок имеет два сайта связывания АТФ: активный сайт доступен в любой конформации белка, но связывание АТФ с сайтом-ингибитором стабилизирует конформацию, которая плохо связывает F6P. [22] Ряд других небольших молекул могут компенсировать вызванный АТФ сдвиг равновесной конформации и реактивировать ПФК, включая циклический АМФ , ионы аммония , неорганический фосфат и фруктозо-1,6- и -2,6-бифосфат. [22]
Цикл лимонной кислоты
[ редактировать ]В митохондриях пируват окисляется пируватдегидрогеназным комплексом до ацетильной группы, которая полностью окисляется до углекислого газа в ходе цикла лимонной кислоты (также известного как цикл Кребса ). Каждый «виток» цикла лимонной кислоты производит две молекулы углекислого газа, один эквивалент АТФ- гуанозинтрифосфата (ГТФ) посредством фосфорилирования на уровне субстрата, катализируемого сукцинил-КоА-синтетазой , поскольку сукцинил-КоА превращается в сукцинат, три эквивалента НАДН. и один эквивалент FADH 2 . НАДН и ФАДН 2 перерабатываются (в НАД + и FAD соответственно) путем окислительного фосфорилирования , генерируя дополнительный АТФ. Окисление НАДН приводит к синтезу 2–3 эквивалентов АТФ, а окисление одного ФАДН 2 дает 1–2 эквивалента АТФ. [20] В результате этого процесса образуется большая часть клеточного АТФ. Хотя сам цикл лимонной кислоты не включает молекулярный кислород , это обязательно аэробный процесс, поскольку O 2 используется для переработки НАДН и ФАДН 2 . При отсутствии кислорода цикл лимонной кислоты прекращается. [21]
Генерация АТФ митохондриями из цитозольного НАДН осуществляется за счет малат-аспартатного челнока (и в меньшей степени глицерин-фосфатного челнока ), поскольку внутренняя мембрана митохондрий непроницаема для НАДН и НАД. + . Вместо переноса образовавшегося НАДН фермент малатдегидрогеназа превращает оксалоацетат в малат , который транспортируется в митохондриальный матрикс. Другая реакция, катализируемая малатдегидрогеназой, происходит в противоположном направлении, образуя оксалоацетат и НАДН из вновь транспортированного малата и внутренних запасов НАД в митохондриях. + . Трансаминаза аспартат превращает оксалоацетат в для транспортировки обратно через мембрану в межмембранное пространство. [21]
При окислительном фосфорилировании прохождение электронов от НАДН и ФАДН 2 через цепь переноса электронов высвобождает энергию для перекачки протонов из митохондриального матрикса в межмембранное пространство. Эта накачка генерирует движущую силу протонов , которая является итоговым эффектом градиента pH и градиента электрического потенциала на внутренней митохондриальной мембране. Поток протонов вниз по этому градиенту потенциала, то есть из межмембранного пространства в матрикс, дает АТФ с помощью АТФ-синтазы. [23] За ход производится три АТФ.
Хотя потребление кислорода представляется основополагающим для поддержания движущей силы протонов, в случае нехватки кислорода ( гипоксии ) внутриклеточный ацидоз (опосредованный повышенными скоростями гликолиза и гидролизом АТФ ) способствует увеличению мембранного потенциала митохондрий и непосредственно управляет синтезом АТФ. [24]
Большая часть АТФ, синтезируемая в митохондриях, будет использована для клеточных процессов в цитозоле; таким образом, он должен быть экспортирован из места синтеза в митохондриальном матриксе. Движению АТФ наружу благоприятствует электрохимический потенциал мембраны, поскольку цитозоль имеет относительно положительный заряд по сравнению с относительно отрицательным матриксом. Каждый вывезенный АТФ стоит 1 H. + . Производство одной АТФ обходится примерно в 3 часа. + . Следовательно, для производства и экспорта одного АТФ требуется 4 часа. +. Внутренняя мембрана содержит антипортер , транслоказу АДФ/АТФ, который представляет собой интегральный мембранный белок, используемый для обмена вновь синтезированного АТФ в матриксе на АДФ в межмембранном пространстве. [25]
Регулирование
[ редактировать ]Цикл лимонной кислоты регулируется главным образом наличием ключевых субстратов, в частности соотношением НАД + НАДН и концентрации кальция , неорганического фосфата, АТФ, АДФ и АМФ. Цитрат – ион, давший название циклу – является ингибитором цитратсинтазы по принципу обратной связи , а также ингибирует ПФК, обеспечивая прямую связь между регуляцией цикла лимонной кислоты и гликолизом. [22]
Бета-окисление
[ редактировать ]В присутствии воздуха и различных кофакторов и ферментов жирные кислоты превращаются в ацетил-КоА . Этот путь называется бета-окислением . Каждый цикл бета-окисления укорачивает цепь жирной кислоты на два атома углерода и производит по одному эквиваленту ацетил-КоА, НАДН и ФАДН 2 . Ацетил-КоА метаболизируется в цикле лимонной кислоты с образованием АТФ, тогда как НАДН и ФАДН 2 используются в результате окислительного фосфорилирования для образования АТФ. Десятки эквивалентов АТФ образуются в результате бета-окисления одной длинной ацильной цепи. [26]
Регулирование
[ редактировать ]При окислительном фосфорилировании ключевым контрольным моментом является реакция, катализируемая цитохром-с-оксидазой , которая регулируется наличием ее субстрата – восстановленной формы цитохрома-с . Количество доступного восстановленного цитохрома с напрямую связано с количеством других субстратов:

из которого непосредственно следует это уравнение:
![{\displaystyle {\frac {[\mathrm {cyt~c_{red}} ]}{[\mathrm {cyt~c_{ox}} ]}}=\left({\frac {[\mathrm {NADH} ] }{[\mathrm {NAD} ]^{+}}}\right)^{\frac {1}{2}}\left({\frac {[\mathrm {ADP} ][\mathrm {P_{i }} ]}{[\mathrm {ATP} ]}}\right)K_ {\mathrm {eq} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/baed008d658c507750599ae155a9a1c11da873e7)
Таким образом, высокое соотношение [НАДН] к [НАД + ] или высокое соотношение [ADP] [P i ] к [АТФ] подразумевают большое количество восстановленного цитохрома с и высокий уровень активности цитохром с оксидазы. [22] Дополнительный уровень регуляции обеспечивается скоростью транспорта АТФ и НАДН между митохондриальным матриксом и цитоплазмой. [25]
Кетоз
[ редактировать ]Кетоновые тела можно использовать в качестве топлива, образуя 22 молекулы АТФ и 2 молекулы ГТФ на молекулу ацетоацетата при окислении в митохондриях. Кетоновые тела транспортируются из печени в другие ткани, где ацетоацетат и бета -гидроксибутират могут быть повторно преобразованы в ацетил-КоА с образованием восстанавливающих эквивалентов (НАДН и ФАДН 2 ) посредством цикла лимонной кислоты. Кетоновые тела не могут использоваться печенью в качестве топлива, поскольку в печени отсутствует фермент β-кетоацил-КоА-трансфераза, также называемый тиолазой . Ацетоацетат в низких концентрациях поглощается печенью и подвергается детоксикации по метилглиоксальному пути, который заканчивается лактатом. Ацетоацетат в высоких концентрациях поглощается клетками, отличными от клеток печени, и поступает другим путем через 1,2-пропандиол . Хотя этот путь проходит через другую серию этапов, требующих АТФ, 1,2-пропандиол можно превратить в пируват. [27]
Производство, анаэробные условия
[ редактировать ]Брожение – это обмен органических соединений в отсутствие воздуха. Он включает фосфорилирование на уровне субстрата при отсутствии дыхательной цепи транспорта электронов . Уравнение реакции глюкозы с образованием молочной кислоты :
- С
66Ч
12 О
6 + 2 АДФ + 2 П и → 2 СН
3 СН(ОН)СООН + 2 АТФ + 2 Н
22О
Анаэробное дыхание – это дыхание в отсутствие О.
2 . Прокариоты могут использовать множество акцепторов электронов. К ним относятся нитраты , сульфаты и углекислый газ.
Пополнение АТФ нуклеозиддифосфаткиназами
[ редактировать ]АТФ также может быть синтезирован посредством нескольких так называемых реакций «пополнения», катализируемых семейством ферментов нуклеозиддифосфаткиназ (NDK), которые используют другие нуклеозидтрифосфаты в качестве высокоэнергетического донора фосфата, и семейством АТФ:гуанидо-фосфотрансферазы . [ нужна ссылка ]
Производство АТФ в процессе фотосинтеза
[ редактировать ]У растений АТФ синтезируется в мембране хлоропласта тилакоидной . Этот процесс называется фотофосфорилированием . «Машина» аналогична таковой в митохондриях, за исключением того, что энергия света используется для перекачки протонов через мембрану и создания протондвижущей силы. Затем происходит АТФ-синтаза точно так же, как при окислительном фосфорилировании. [28] Часть АТФ, вырабатываемого в хлоропластах, расходуется в цикле Кальвина , в результате которого образуются триозные сахара.
переработка АТФ
[ редактировать ]Общее количество АТФ в организме человека составляет около 0,1 моль/л . [29] Большая часть АТФ перерабатывается из АДФ в результате вышеупомянутых процессов. Таким образом, в любой момент времени общее количество АТФ + АДФ остается довольно постоянным.
Энергия, используемая клетками человека у взрослого человека, требует ежедневного гидролиза от 100 до 150 моль/л АТФ, а это означает, что человек обычно использует количество АТФ, соответствующее весу его тела, в течение дня. [30] Каждый эквивалент АТФ перерабатывается 1000–1500 раз в течение одного дня ( 150/0,1 = 1500 ), [29] примерно 9×10 20 молекул/с. [29]
Биохимические функции
[ редактировать ]Внутриклеточная передача сигналов
[ редактировать ]АТФ участвует в передаче сигнала , служа субстратом для киназ — ферментов, переносящих фосфатные группы. Киназы являются наиболее распространенными АТФ-связывающими белками. У них есть небольшое количество общих складок. [31] Фосфорилирование белка киназой может активировать каскад, такой как митоген-активируемый протеинкиназный каскад. [32]
АТФ также является субстратом аденилатциклазы , чаще всего в путях передачи сигнала рецептора, связанного с G-белком , и трансформируется во второй мессенджер , циклический АМФ, который участвует в запуске сигналов кальция путем высвобождения кальция из внутриклеточных запасов. [33] Эта форма передачи сигнала особенно важна для функции мозга, хотя она участвует в регуляции множества других клеточных процессов. [34]
Синтез ДНК и РНК
[ редактировать ]АТФ — один из четырех мономеров, необходимых для синтеза РНК . Процессу способствуют РНК-полимеразы . [35] Аналогичный процесс происходит при образовании ДНК, за исключением того, что АТФ сначала превращается в дезоксирибонуклеотид дАТФ. Как и многие реакции конденсации в природе, репликация и транскрипция ДНК также потребляют АТФ.
Активация аминокислот в синтезе белка
[ редактировать ]Ферменты аминоацил-тРНК-синтетазы потребляют АТФ в месте прикрепления тРНК к аминокислотам, образуя комплексы аминоацил-тРНК. Аминоацилтрансфераза связывает АМФ-аминокислоту с тРНК. Реакция сочетания протекает в две стадии:
- аа + АТФ ⟶ аа-АМФ + ПП и
- аа-АМФ + тРНК ⟶ аа-тРНК + АМФ
Аминокислота связана с предпоследним нуклеотидом на 3'-конце тРНК (буква А в последовательности CCA) посредством сложноэфирной связи (перевернутая на иллюстрации).
Транспортер кассет, связывающих АТФ
[ редактировать ]Транспортировка химических веществ из клетки против градиента часто связана с гидролизом АТФ. Транспорт осуществляется АТФ-связывающими кассетными переносчиками . Геном человека кодирует 48 транспортеров ABC, которые используются для экспорта лекарств, липидов и других соединений. [36]
Внеклеточная передача сигналов и нейротрансмиссия
[ редактировать ]Клетки секретируют АТФ для связи с другими клетками в процессе, называемом пуринергической передачей сигналов . АТФ служит нейротрансмиттером во многих частях нервной системы, модулирует биение ресничек, влияет на снабжение сосудов кислородом и т. д. АТФ либо секретируется непосредственно через клеточную мембрану через канальные белки. [37] [38] или закачивается в везикулы [39] которые затем сливаются с мембраной. Клетки обнаруживают АТФ с помощью пуринергических рецепторов белков- P2X и P2Y . [40] Было показано, что АТФ является критически важной сигнальной молекулой для взаимодействия микроглии и нейронов во взрослом мозге. [41] а также во время развития мозга. [42] Более того, АТФ-сигнализация, индуцированная повреждением тканей, является основным фактором быстрых изменений фенотипа микроглии. [43]
Сокращение мышц
[ редактировать ]АТФ питает мышечные сокращения . [44] Мышечные сокращения регулируются сигнальными путями, хотя разные типы мышц регулируются специфическими путями и стимулами, основанными на их конкретной функции. Однако во всех типах мышц сокращение осуществляется белками актином и миозином . [45]
АТФ изначально связан с миозином. Когда АТФаза гидролизует связанный АТФ до АДФ и неорганического фосфата , миозин располагается таким образом, что может связываться с актином. Миозин, связанный АДФ и Pi , образует поперечные мостики с актином, и последующее высвобождение АДФ и Pi высвобождает энергию в виде силового удара. Силовой удар заставляет актиновую нить скользить мимо миозиновой нити, укорачивая мышцу и вызывая сокращение. Затем другая молекула АТФ может связываться с миозином, освобождая его от актина и позволяя этому процессу повториться. [45] [46]
Растворимость белка
[ редактировать ]Недавно было предложено действовать как биологический гидротроп АТФ. [47] и было показано, что он влияет на растворимость во всем протеоме. [48]
Абиогенное происхождение
[ редактировать ]Ацетилфосфат (AcP), предшественник АТФ, может быть легко синтезирован с умеренными выходами из тиоацетата при pH 7 и 20 °C и pH 8 и 50 °C, хотя ацетилфосфат менее стабилен при более высоких температурах и щелочных условиях, чем в более холодных условиях. и от кислой до нейтральной среды. Он не способен способствовать полимеризации рибонуклеотидов и аминокислот и способен только к фосфорилированию органических соединений. Было показано, что он может способствовать агрегации и стабилизации АМФ в присутствии Na. + , агрегация нуклеотидов может способствовать полимеризации при температуре выше 75 ° C в отсутствие Na. + . Возможно, что полимеризация, стимулируемая AcP, может происходить на минеральных поверхностях. [49] Было показано, что АДФ может фосфорилироваться до АТФ только с помощью AcP, а другие нуклеозидтрифосфаты не фосфорилируются с помощью AcP. Это может объяснить, почему все формы жизни используют АТФ для запуска биохимических реакций. [50]
Аналоги АТФ
[ редактировать ]Биохимические лаборатории часто используют исследования in vitro для изучения АТФ-зависимых молекулярных процессов. Аналоги АТФ также используются в рентгеновской кристаллографии для определения структуры белка в комплексе с АТФ, часто вместе с другими субстратами. [ нужна ссылка ]
Ингибиторы АТФ-зависимых ферментов, такие как киназы, необходимы для изучения сайтов связывания и переходных состояний, участвующих в АТФ-зависимых реакциях. [ нужна ссылка ]
Большинство полезных аналогов АТФ не могут подвергаться гидролизу, как это происходит с АТФ; вместо этого они удерживают фермент в структуре, тесно связанной с АТФ-связанным состоянием. Аденозин-5'-(γ-тиотрифосфат) представляет собой чрезвычайно распространенный аналог АТФ, в котором один из атомов кислорода гамма-фосфата заменен атомом серы ; этот анион гидролизуется значительно медленнее, чем сам АТФ, и действует как ингибитор АТФ-зависимых процессов. В кристаллографических исследованиях переходные состояния гидролиза моделируются связанным ионом ванадата .
Следует с осторожностью интерпретировать результаты экспериментов с аналогами АТФ, поскольку некоторые ферменты могут гидролизовать их с заметной скоростью при высоких концентрациях. [51]
Медицинское использование
[ редактировать ]АТФ используется внутривенно при некоторых заболеваниях сердца. [52]
История
[ редактировать ]АТФ был открыт в 1929 году Карлом Ломанном [53] и Джендрассик [54] и независимо Сайрусом Фиске и Йеллапрагадой Субба Рао из Гарвардской медицинской школы , [55] обе команды соревнуются друг с другом, чтобы найти анализ на фосфор.
в 1941 году предположил, что он является посредником между энерговыделяющими и энергозатратными реакциями в клетках Фриц Альберт Липманн . [56]
Впервые он был синтезирован в лаборатории Александром Тоддом в 1948 году. [57] и частично за эту работу он был удостоен Нобелевской премии по химии в 1957 году.
1978 года Нобелевская премия по химии была присуждена Питеру Деннису Митчеллу за открытие хемиосмотического механизма синтеза АТФ.
Нобелевская премия по химии 1997 года была разделена: половина совместно Пола Д. Бойера и Джона Э. Уокера «за объяснение ферментативного механизма, лежащего в основе синтеза аденозинтрифосфата (АТФ)», а другая половина — Йенса К. Скоу . за первое открытие фермента, переносящего ионы, Na + , К + -АТФаза». [58]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б «Информация о продукте динатриевая соль аденозин-5'-трифосфата» (PDF) . Сигма. Архивировано (PDF) из оригинала 23 марта 2019 г. Проверено 22 марта 2019 г.
- ^ Данн, Джейкоб; Грайдер, Майкл Х. (2023). «Физиология аденозинтрифосфата» . СтатПерлс . Издательство StatPearls. ПМИД 31985968 . Проверено 13 ноября 2023 г.
- ^ Ноулз, младший (1980). «Реакции переноса фосфорила, катализируемые ферментами». Анну. Преподобный Биохим . 49 : 877–919. дои : 10.1146/annurev.bi.49.070180.004305 . ПМИД 6250450 .
- ^ «Средний человек с ежедневным рационом 8000 кДж и 30% эффективностью превращения пищевых продуктов в химическую энергию будет синтезировать (и гидролизовать) около 50 кг АТФ в течение 1 дня». Уилкенс, Стефан (2008). «Синтез АТФ, химия». Энциклопедия химической биологии Wiley . дои : 10.1002/9780470048672.wecb648 . ISBN 9780471754770 .
- ^ Данн, Джейкоб; Грайдер, Майкл Х. (2023), «Физиология, аденозинтрифосфат» , StatPearls , Остров сокровищ (Флорида): StatPearls Publishing, PMID 31985968 , получено 28 сентября 2023 г.
- ^ Перейти обратно: а б Сторер, А.; Корниш-Боуден, А. (1976). «Концентрация MgATP 2− и другие ионы в растворе. Расчет истинных концентраций видов, присутствующих в смесях ассоциированных ионов» . Biochem. J. 159 ( 1): 1–5. : 10.1042 /bj1590001 . PMC 1164030. . PMID 11772 doi
- ^ Уилсон, Дж.; Чин, А. (1991). «Хелирование двухвалентных катионов АТФ, изученное методом титрационной калориметрии». Анальный. Биохим . 193 (1): 16–19. дои : 10.1016/0003-2697(91)90036-S . ПМИД 1645933 .
- ^ Гарфинкель, Л.; Альтшульд, Р.; Гарфинкель, Д. (1986). «Магний в сердечном энергетическом обмене». Дж. Мол. Клетка. Кардиол . 18 (10): 1003–1013. дои : 10.1016/S0022-2828(86)80289-9 . ПМИД 3537318 .
- ^ Сэйлор, П.; Ван, К.; Хираи, Т.; Адамс, Дж. (1998). «Второй ион магния имеет решающее значение для связывания АТФ в киназном домене онкопротеина v-Fps». Биохимия . 37 (36): 12624–12630. дои : 10.1021/bi9812672 . ПМИД 9730835 .
- ^ Лин, X.; Айрапетов М; Сан, Г. (2005). «Характеристика взаимодействий между активным центром протеинтирозинкиназы и активатором двухвалентного металла» . БМК Биохим . 6:25 . дои : 10.1186/1471-2091-6-25 . ПМК 1316873 . ПМИД 16305747 .
- ^ Будавари, Сьюзен, изд. (2001), Индекс Merck: Энциклопедия химических веществ, лекарств и биологических препаратов (13-е изд.), Merck, ISBN 0911910131
- ^ Перейти обратно: а б Фергюсон, С.Дж.; Николлс, Дэвид; Фергюсон, Стюарт (2002). Биоэнергетика 3 (3-е изд.). Сан-Диего, Калифорния: Академический. ISBN 978-0-12-518121-1 .
- ^ Берг, Дж. М.; Тимочко, Дж. Л.; Страйер, Л. (2003). Биохимия . Нью-Йорк, штат Нью-Йорк: WH Freeman. п. 376 . ISBN 978-0-7167-4684-3 .
- ^ Шанс, Б.; Лиз, Х.; Постгейт, Дж. Г. (1972). «Значение «обратного потока электронов» и «электрона высокой энергии» в биохимии». Природа . 238 (5363): 330–331. Бибкод : 1972Natur.238..330C . дои : 10.1038/238330a0 . ПМИД 4561837 . S2CID 4298762 .
- ^ Гаевский, Э.; Стеклер, Д.; Голдберг, Р. (1986). «Термодинамика гидролиза аденозин-5'-трифосфата до аденозин-5'-дифосфата» . Ж. Биол. Хим . 261 (27): 12733–12737. дои : 10.1016/S0021-9258(18)67153-4 . ПМИД 3528161 .
- ^ Берг, Джереми М.; Тимочко, Джон Л.; Страйер, Люберт (2007). Биохимия (6-е изд.). Нью-Йорк, штат Нью-Йорк: WH Freeman. п. 413. ИСБН 978-0-7167-8724-2 .
- ^ Гарретт, Реджинальд Х.; Гришэм, Чарльз М. (2016). Биохимия (6-е изд.). Cengage Обучение. п. 68. ИСБН 978-1305577206 .
- ^ Бейс, И.; Ньюхолм, EA (1 октября 1975 г.). «Содержание адениновых нуклеотидов, фосфагенов и некоторых гликолитических промежуточных продуктов в покоящихся мышцах позвоночных и беспозвоночных» . Биохим. Дж . 152 (1): 23–32. дои : 10.1042/bj1520023 . ПМЦ 1172435 . ПМИД 1212224 .
- ^ «Аденозинтрифосфат» . Британника. 11 ноября 2023 г. Проверено 1 декабря 2023 г.
- ^ Перейти обратно: а б Рич, PR (2003). «Молекулярный механизм дыхательной цепи Кейлина». Биохим. Соц. Транс . 31 (6): 1095–1105. дои : 10.1042/BST0311095 . ПМИД 14641005 .
- ^ Перейти обратно: а б с Лодиш, Х.; Берк, А.; Мацудайра, П.; Кайзер, Калифорния; Кригер, М.; Скотт, член парламента; Зипурский, С.Л.; Дарнелл, Дж. (2004). Молекулярно-клеточная биология (5-е изд.). Нью-Йорк, штат Нью-Йорк: WH Freeman. ISBN 978-0-7167-4366-8 .
- ^ Перейти обратно: а б с д и Фут, Д.; Воэт, JG (2004). Биохимия . Том. 1 (3-е изд.). Хобокен, Нью-Джерси: Уайли. ISBN 978-0-471-19350-0 .
- ^ Абрахамс, Дж.; Лесли, А.; Луттер, Р.; Уокер, Дж. (1994). «Структура F1-АТФазы из митохондрий бычьего сердца с разрешением 2,8 Å». Природа . 370 (6491): 621–628. Бибкод : 1994Natur.370..621A . дои : 10.1038/370621a0 . ПМИД 8065448 . S2CID 4275221 .
- ^ Дево, JBL; Хеджес, CP; Хики, AJR (январь 2019 г.). «Ацидоз поддерживает функцию митохондрий головного мозга у толерантных к гипоксии трехплефиновых рыб: стратегия выживания при остром гипоксическом воздействии?» . Фронт Физиол . 9, 1914: 1941. doi : 10.3389/fphys.2018.01941 . ПМК 6346031 . ПМИД 30713504 .
- ^ Перейти обратно: а б Даут-Гонсалес, К.; Нури, Х.; Трезеге, В.; Локен, Г.; Пебай-Пейрула, Э.; Брандолин, Г. (2006). «Молекулярные, функциональные и патологические аспекты митохондриального носителя АДФ/АТФ». Физиология . 21 (4): 242–249. дои : 10.1152/физиол.00005.2006 . ПМИД 16868313 .
- ^ Роннетт, Г.; Ким, Э.; Ландри, Л.; Ту, Ю. (2005). «Метаболизм жирных кислот как цель лечения ожирения». Физиол. Поведение . 85 (1): 25–35. дои : 10.1016/j.physbeh.2005.04.014 . ПМИД 15878185 . S2CID 24865576 .
- ^ «Интегрированная система информации о рисках» (PDF) . 15 марта 2013 г. Архивировано (PDF) из оригинала 24 сентября 2015 г. Проверено 1 февраля 2019 г.
- ^ Аллен, Дж. (2002). «Фотосинтез АТФ-электронов, протонных насосов, роторов и уравновешивания» . Клетка . 110 (3): 273–276. дои : 10.1016/S0092-8674(02)00870-X . ПМИД 12176312 . S2CID 1754660 .
- ^ Перейти обратно: а б с Фурман, Брэдли П.; Циммерман, Джерри Дж. (2011). Детская реанимационная помощь . Эльзевир. п. 1061. ИСБН 978-0-323-07307-3 . Проверено 16 мая 2020 г.
- ^ Фурман, Брэдли П.; Циммерман, Джерри Дж. (2011). Детская реанимационная помощь . Эльзевир. стр. 1058–1072. ISBN 978-0-323-07307-3 . Проверено 16 мая 2020 г.
- ^ Шифф, Э.; Борн, П. (2005). «Структурная эволюция суперсемейства протеинкиназоподобных» . ПЛОС Компьютер. Биол . 1 (5): е49. Бибкод : 2005PLSCB...1...49S . дои : 10.1371/journal.pcbi.0010049 . ПМЦ 1261164 . ПМИД 16244704 .
- ^ Мишра, Н.; Тутея, Р.; Тутея, Н. (2006). «Передача сигналов через сети MAP-киназы у растений». Арх. Биохим. Биофиз . 452 (1): 55–68. дои : 10.1016/j.abb.2006.05.001 . ПМИД 16806044 .
- ^ Каменецкий, М.; Миддельхауфе, С.; Банк, Э.; Левин, Л.; Бак, Дж.; Стигборн, К. (2006). «Молекулярные детали генерации цАМФ в клетках млекопитающих: история двух систем» . Дж. Мол. Биол . 362 (4): 623–639. дои : 10.1016/j.jmb.2006.07.045 . ПМЦ 3662476 . ПМИД 16934836 .
- ^ Ханун, Дж.; Дефер, Н. (2001). «Регуляция и роль изоформ аденилатциклазы». Анну. Преподобный Фармакол. Токсикол . 41 (1): 145–174. doi : 10.1146/annurev.pharmtox.41.1.145 . ПМИД 11264454 .
- ^ Джойс, CM; Стейтц, Т.А. (1995). «Структуры и функции полимеразы: вариации на тему?» . Дж. Бактериол . 177 (22): 6321–6329. дои : 10.1128/jb.177.22.6321-6329.1995 . ПМК 177480 . ПМИД 7592405 .
- ^ Борст, П.; Эльферинк, Р. Ауд (2002). «Переносчики ABC млекопитающих в здоровье и болезни» (PDF) . Ежегодный обзор биохимии . 71 : 537–592. doi : 10.1146/annurev.biochem.71.102301.093055 . ПМИД 12045106 . S2CID 34707074 . Архивировано (PDF) из оригинала 21 апреля 2018 г. Проверено 20 апреля 2018 г.
- ^ Романов Роман А.; Лэшер, Роберт С.; Высоко, Бриджит; Сэвидж, Логан Э.; Лоусон, Адам; Рогачевская Ольга Александровна; Чжао, Гаитян; Рогачевский Вадим Владимирович; Быстрова Марина Ф.; Чурбанов Глеб Д.; Адамейко Игорь; Харкани, Тибор; Ян, Жуйбяо; Кидд, Грэм Дж.; Марамбо, Филипп; Киннамон, Джон К.; Колесников Станислав С.; Фингер, Томас Э. (2018). «Химические синапсы без синаптических везикул: пуринергическая нейротрансмиссия через канал CALHM1-митохондриальный сигнальный комплекс» . Научная сигнализация . 11 (529): eaao1815. дои : 10.1126/scisignal.aao1815 . ISSN 1945-0877 . ПМК 5966022 . ПМИД 29739879 .
- ^ Даль, Герхард (2015). «Выброс АТФ через каналы Паннексона» . Философские труды Королевского общества B: Биологические науки . 370 (1672): 20140191. doi : 10.1098/rstb.2014.0191 . ISSN 0962-8436 . ПМЦ 4455760 . ПМИД 26009770 .
- ^ Ларссон, Макс; Савада, Кейсуке; Морланд, Сесилия; Хиаса, Мики; Ормель, Лассе; Морияма, Ёсинори; Гундерсен, Видар (2012). «Функциональная и анатомическая идентификация везикулярного транспортера, опосредующего высвобождение АТФ в нейронах» . Кора головного мозга . 22 (5): 1203–1214. дои : 10.1093/cercor/bhr203 . ISSN 1460-2199 . ПМИД 21810784 .
- ^ Пухалович, Камила; Тарновский, Мацей; Барановская-Босяцкая, Ирена; Хлубек, Дариуш; Дзедейко, Виолетта (18 декабря 2014 г.). «Рецепторы P2X и P2Y — роль в патофизиологии нервной системы» . Международный журнал молекулярных наук . 15 (12): 23672–23704. дои : 10.3390/ijms151223672 . ISSN 1422-0067 . ПМЦ 4284787 . ПМИД 25530618 .
- ^ Чаба, Череп; Балаж, Посфаи (2020). «Микроглия контролирует и защищает функции нейронов посредством специализированных соматических пуринергических соединений» . Наука . doi : 10.1126/science.aax6752 . ПМИД 31831638 .
- ^ Чаба, Череп; Анетт, Шварц Д. (2022). «Микроглиальный контроль развития нейронов через соматические пуринергические соединения» . Отчеты по ячейкам . дои : 10.1016/j.celrep.2022.111369 . ПМИД 36130488 .
- ^ Питер, Берки; Чаба, Череп; Жужанна, Кёрней (2024). «Микроглия способствует нейрональной синхронизации, несмотря на фенотипическую трансформацию, связанную с эндогенной АТФ, в острых срезах мозга мышей» . Природные коммуникации . дои : 10.1038/s41467-024-49773-1 . ПМЦ 11208608 . ПМИД 38926390 .
- ^ Хультман, Э.; Гринхафф, Польша (1991). «Энергетический обмен скелетных мышц и утомляемость при интенсивных физических нагрузках у человека» . Научный прогресс . 75 (298, ч. 3–4): 361–370. ISSN 0036-8504 . ПМИД 1842855 .
- ^ Перейти обратно: а б Куо, Ивана Ю.; Эрлих, Барбара Э. (февраль 2015 г.). «Сигнализация при сокращении мышц» . Перспективы Колд-Спринг-Харбор в биологии . 7 (2): а006023. doi : 10.1101/cshperspect.a006023 . ISSN 1943-0264 . ПМЦ 4315934 . ПМИД 25646377 .
- ^ «38.17: Мышечное сокращение и передвижение — АТФ и мышечное сокращение» . Свободные тексты по биологии . 16 июля 2018 г. Проверено 1 мая 2024 г.
- ^ Хайман, Энтони А.; Кришнан, Ямуна; Альберти, Саймон; Ван, Цзе; Саха, Шамбадитья; Малиновская, Лилиана; Патель, Авинаш (19 мая 2017 г.). «АТФ как биологический гидротроп». Наука . 356 (6339): 753–756. Бибкод : 2017Sci...356..753P . doi : 10.1126/science.aaf6846 . ISSN 0036-8075 . ПМИД 28522535 . S2CID 24622983 .
- ^ Савицкий Михаил М.; Банщефф, Маркус; Хубер, Вольфганг; Доминик Хелм; Гюнтнер, Ина; Вернер, Тило; Курзава, Нильс; Шридхаран, Синдхуджа (11 марта 2019 г.). «Профилирование растворимости и термостабильности в масштабе всего протеома показывает четкую регуляторную роль АТФ» . Природные коммуникации . 10 (1): 1155. Бибкод : 2019NatCo..10.1155S . дои : 10.1038/s41467-019-09107-y . ISSN 2041-1723 . ПМК 6411743 . ПМИД 30858367 .
- ^ Уичер, Александра; Кампруби, Элои; Пинна, Сильвана; Херши, Барри; Лейн, Ник (01 июня 2018 г.). «Ацетилфосфат как первичная энергетическая валюта в зарождении жизни» . Происхождение жизни и эволюция биосфер . 48 (2): 159–179. Бибкод : 2018OLEB...48..159W . дои : 10.1007/s11084-018-9555-8 . ISSN 1573-0875 . ПМК 6061221 . ПМИД 29502283 .
- ^ «Древняя химия может объяснить, почему живые существа используют АТФ в качестве универсальной энергетической валюты: ранний шаг метаболической эволюции подготовил почву для появления АТФ в качестве универсального носителя энергии» . ScienceDaily . Проверено 27 августа 2023 г.
- ^ Реретар, AM; Чалович, Дж. М. (1995). «Аденозин-5'-(гамма-тиотрифосфат): аналог АТФ, который следует использовать с осторожностью в исследованиях мышечных сокращений». Биохимия . 34 (49): 16039–16045. дои : 10.1021/bi00049a018 . ПМИД 8519760 .
- ^ Пеллег, Амир; Куталек, Стивен П.; Фламманг, Дэниел; Бендит, Дэвид (февраль 2012 г.). «ATPace: инъекционный аденозин-5'-трифосфат» . Пуринергическая сигнализация . 8 (Приложение 1): 57–60. дои : 10.1007/s11302-011-9268-1 . ISSN 1573-9538 . ПМК 3265710 . ПМИД 22057692 .
- ^ Ломанн, К. (август 1929 г.). «О пирофосфатной фракции в мышцах». Естественные науки (на немецком языке). 17 (31): 624–625. Бибкод : 1929NW.....17..624. . дои : 10.1007/BF01506215 . S2CID 20328411 .
- ^ Воан, Марта; Хилл, Роберт Л.; Симони, Роберт Д. (2002). «Определение фосфора и открытие фосфокреатина и АТФ: работа Фиске и СуббаРоу» . Журнал биологической химии . 277 (32): е21. ПМИД 12161449 . Архивировано из оригинала 8 августа 2017 г. Проверено 24 октября 2017 г.
- ^ Маруяма, К. (март 1991 г.). «Открытие аденозинтрифосфата и установление его структуры». Дж. Хист. Биол . 24 (1): 145–154. дои : 10.1007/BF00130477 . S2CID 87425890 .
- ^ Липманн, Ф. (1941). «Метаболическое образование и использование энергии фосфатных связей». Адв. Энзимол . 1 : 99–162. ISSN 0196-7398 .
- ^ «История: АТФ впервые обнаружен в 1929 году» . Нобелевская премия по химии 1997 года . Нобелевский фонд . Архивировано из оригинала 23 января 2010 г. Проверено 26 мая 2010 г.
- ^ «Нобелевская премия по химии 1997 года» . Нобелевская премия . Архивировано из оригинала 24 октября 2017 года . Проверено 21 января 2018 г.
Внешние ссылки
[ редактировать ]- АТФ связан с белками в PDB
- ScienceAid: Энергетическая АТФ и упражнения
- Запись PubChem об аденозинтрифосфате
- Запись KEGG об аденозинтрифосфате
метаболизма Карта |
|---|
