CCR5
| CCR5 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CCR5 , CC-CKR-5, CCCKR5, CCR-5, CD195, CKR-5, CKR5, CMKBR5, IDDM22, CC Motif Chemokine Рецептор 5 (ген/псевдоген), CC Motif Chemokine Рецептор 5 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Омим : 601373 ; MGI : 107182 ; Гомологен : 37325 ; GeneCards : CCR5 ; OMA : CCR5 - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




CC -хемокиновый рецептор типа 5 , также известный как CCR5 или CD195 , представляет собой белок на поверхности лейкоцитов , который участвует в иммунной системе, поскольку он действует как рецептор для хемокинов . [ 5 ]
У людей ген CCR5 , который кодирует белок CCR5, расположен на коротком (P) руке в положении 21 в хромосоме 3 . Некоторые популяции унаследовали мутацию Delta 32 , что привело к генетической делеции части гена CCR5. Гомозиготные носители этой мутации устойчивы к инфекции макрофаг-тропическими (M-триропическими) штаммами ВИЧ-1 . [ 6 ] [ 7 ] [ 8 ] [ 9 ] [ 10 ] [ 11 ]
Функция
[ редактировать ]Белок CCR5 принадлежит к бета -хемокинов семейству интегральных мембранных рецепторов . [ 12 ] [ 13 ] Это G -белок -рецептор [ 12 ] который функционирует как рецептор хемокинов в группе хемокинов CC .
CCR5 Сознательные лиганды включают CCL3 , CCL4 (также известный как MIP 1 α и 1 β , соответственно), и CCL3L1 . [ 14 ] [ 15 ] Кроме того, CCR5 взаимодействует с CCL5 ( хемотаксический цитокиновый белок, также известный как Rantes). [ 14 ] [ 16 ] [ 17 ]
CCR5 преимущественно экспрессируется на Т -клетках , макрофагах , дендритных клетках , эозинофилах , микроглии и субпопуляции клеток рака молочной железы или простаты. [ 18 ] [ 19 ] Экспрессия CCR5 селективно индуцируется во время процесса трансформации рака и не экспрессируется в нормальных эпителиальных клетках молочной железы или простаты. Приблизительно 50% рака молочной железы человека экспрессировали CCR5, в первую очередь при тройном негативном раке молочной железы . [ 18 ] Ингибиторы CCR5 блокировали миграцию и метастазирование клеток рака молочной железы и рака простаты, которые экспрессировали CCR5, что позволяет предположить, что CCR5 может функционировать как новая терапевтическая мишень. [ 18 ] [ 19 ] [ 20 ] Недавние исследования показывают, что CCR5 экспрессируется в подмножестве раковых клеток с характеристиками раковых стволовых клеток, которые, как известно, управляют устойчивостью к терапии, и что ингибиторы CCR5 усиливали количество клеток, убитых в результате современной химиотерапии. [ 21 ] Вполне вероятно, что CCR5 играет роль в воспалительных реакциях на инфекцию, хотя его точная роль в нормальной иммунной функции неясна. Области этого белка также имеют решающее значение для связывания хемокиновых лигандов, функционального ответа рецептора и активности со-рецептора ВИЧ. [ 22 ]
Модуляция активности CCR5 способствует непатогенному курсу инфекции вирусом обезживания иммунодефицита (SIV) у нескольких африканских нечеловеческих видов приматов, которые являются долгосрочными природными хозяевами SIV и избегают иммунодефицита при инфекции. [ 23 ] Эти регуляторные механизмы включают в себя: генетические делеции, которые отменяют экспрессию клеточной поверхности CCR5, [ 24 ] подавление CCR5 на поверхности CD4+ T -клеток, в частности на клетках памяти, [ 25 ] и отсроченное начало экспрессии CCR5 на CD4+ T -клетках во время развития. [ 26 ] [ 27 ]

ВИЧ
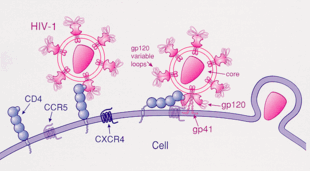
[ редактировать ]ВИЧ-1 чаще всего использует рецепторы хемокинов CCR5 и/или CXCR4 в качестве со-рецепторов для введения иммунологических клеток-мишеней. [ 28 ] Эти рецепторы расположены на поверхности иммунных клеток-хозяев, в результате чего они обеспечивают метод входа для вируса ВИЧ-1 для заражения клетки. [ 29 ] Структура гликопротеина ВИЧ-1 огибала необходима для обеспечения вирусной проникновения ВИЧ-1 в клетку-мишени. [ 29 ] Структура гликопротеина оболочки состоит из двух белковых субъединиц, расщепленных из предшественника белка GP160, кодируемого для гена ENV-1 : внешняя субъединица GP120 и трансмембранная субъединица GP41. [ 29 ] Эта структура гликопротеинов конверта расположена в скайно-подобную структуру, расположенную на поверхности вириона и состоит из тримера гетеродимеров GP120-GP41. [ 29 ] Белок оболочки GP120 является имитиром хемокинов. [ 28 ] Хотя ему не хватает уникальной структуры хемокина, он все еще способен связываться с рецепторами хемокинов CCR5 и CXCR4. [ 28 ] Во время ВИЧ-1 субъединица гликопротеина GP120 гликопротеина связывается с гликопротеином CD4 и со-рецептором ВИЧ-1, экспрессируемого на клетке-мишени, образуя гетеротримерный комплекс. [ 28 ] Образование этого комплекса стимулирует высвобождение фузогенного пептида, вызывая слиться с вирусной мембраной с мембраной клетки -мишени. [ 28 ] Поскольку связывание только с CD4 иногда может привести к выбросу GP120, GP120 должен затем связываться с CCR5 с помощью корецептора, чтобы слияние продолжалось. Тирозин-сольфы-амино-концерн этого со-рецептора является «основным детерминантом» связывания с гликопротеином GP120. [ 30 ] Совместный рецептор также распознает область V1-V2 GP120 и мостовой лист (антипараллельный 4-цепорный β-лист, который соединяет внутренние и внешние домены GP120). Ствол V1-V2 может влиять на «использование совместного рецептора через его пептидную композицию, а также степень N-связанного гликозилирования». Однако, в отличие от V1-V2, петля V3 сильно варьируется и, следовательно, является наиболее важным фактором, определяющим специфичность корецептора. [ 30 ] Нормальные лиганды для этого рецептора, Rantes , MIP-1β и MIP-1α способны подавлять ВИЧ-1 инфекцию in vitro . У людей, инфицированных ВИЧ, вирусы, использующие CCR5, являются преобладающими видами, выделенными на ранних стадиях вирусной инфекции, [ 31 ] предполагая, что эти вирусы могут иметь селективное преимущество во время передачи или острой фазы заболевания. Более того, по крайней мере половина всех инфицированных людей содержит только вирусы, использующие CCR5 в течение всего инфекции.
CCR5 является основным корецептором, используемым GP120, последовательно с CD4. Это связывание приводит к GP41, другому белковому продукту GP160, высвобождаемому из его метастабильной конформации и вставленного в мембрану клетки -хозяина. Хотя это не было подтверждено, связывание GP120-CCR5 включает в себя два важных этапа: 1) Амино-концерн, сорфалированный тирозин, является «основным детерминантом» связывания с GP120 (как указывалось ранее) 2) после шага 1 ., Должно быть взаимное действие (синергия, взаимосвязь) между GP120 и трансмембранными доменами CCR5. [ 30 ]
CCR5 необходим для распространения R5-штамма вируса ВИЧ-1. [ 32 ] Знание механизма, с помощью которого этот штамм ВИЧ-1 опосредует инфекцию, вызвало исследование развития терапевтических вмешательств для блокировки функции CCR5. [ 33 ] Ряд новых экспериментальных лекарств от ВИЧ, называемых антагонистами рецептора CCR5 , были разработаны для межответствующего связывания между белком оболочки GP120 и CCR5 CCR5 COR-рецептора GP120. [ 32 ] Эти экспериментальные препараты включают Pro140 ( Cytodyn ), Vicriviroc (исследования фазы III были отменены в июле 2010 года) ( Schering Plow ), Aplaviroc (GW-873140) ( Glaxosmithkline ) и Maraviroc (UK-427857) ( Pfizer ). Maraviroc был утвержден для использования FDA в августе 2007 года. [ 32 ] Это единственный, что давно одобрен FDA для клинического использования, тем самым становясь первым ингибитором CCR5. [ 30 ] Проблема такого подхода заключается в том, что, хотя CCR5 является основным со-рецептором, с помощью которого ВИЧ-инфекции клетки, это не единственный такой совместный рецептор. Вполне возможно, что под селективным давлением ВИЧ будет развиваться, чтобы использовать другой совместный рецептор. Тем не менее, исследование устойчивости к вирусам к AD101, молекулярному антагонисту CCR5, показало, что устойчивые вирусы не переключались на другой корецептор (CXCR4), но сохранялись при использовании CCR5: они либо связаны с альтернативными доменами CCR5, либо с рецептором в Более высокая аффинность. Однако, поскольку есть еще один доступный совместный рецептор, вероятно, что отсутствие гена CCR5 не делает одного невосприимчивым к вирусу; Для человека было бы просто сложнее заключить его. Кроме того, вирус все еще имеет доступ к CD4. В отличие от CCR5, который не требуется (о чем свидетельствует тех, кто живет здоровой жизнью, даже если у него нет гена в результате мутации Delta32), CD4 имеет решающее значение в системе иммунной защиты организма. [ 34 ] Даже без наличия любого корецептора (даже CCR5) вирус все еще может вторгаться в клетки, если GP41 должен был пройти через изменение (включая его цитоплазматический хвост), что привело к независимости CD4 без необходимости CCR5 и/или CXCR4 как дверной проем. [ 35 ]
Рак
[ редактировать ]Экспрессия CCR5 индуцируется в эпителиальных клетках молочной железы и простаты при трансформации. [ 18 ] [ 19 ] Индукция экспрессии CCR5 способствует клеточной инвазии, миграции и метастазированию. [ 5 ] [ 18 ] [ 21 ] Индукция метастазирования включает в себя домашнее животное в метастатическое место. Было показано, что ингибиторы CCR5, включая Maraviroc и Leronlimab, блокируют метастазирование легких клеточных линий рака молочной железы человека. [ 18 ] [ 36 ] В доклинических исследованиях ингибиторов CCR5 CCR5 CCR5 иммунных мышей блокировали метастазирование в кости и мозг. [ 19 ] Ингибиторы CCR5 также уменьшают инфильтрацию связанных с опухолью макрофагов. [ 37 ] Клиническое исследование фазы 1 ингибитора CCR5 у пациентов с метастатическим раком толстой кишки, сильно предварительно обработанной метастатическим раком толстой кишки, продемонстрировало объективный клинический ответ и снижение метастатической нагрузки опухоли. [ 38 ]
Мозг
[ редактировать ]Повышенные уровни CCR5 являются частью воспалительного ответа на инсульт и смерть. Блокирование CCR5 с помощью Maraviroc (препарат, утвержденный для ВИЧ), может усилить восстановление после инсульта. [ 39 ] [ 40 ]
В развивающемся мозге рецепторы хемокинов, такие как CCR5, влияют на миграцию и связь нейронов. После инсульта они, кажется, уменьшают количество участков соединения на нейронах вблизи повреждения. [ 39 ]
CCR5-Δ32
[ редактировать ]CCR5-Δ32 (или CCR5-D32 или CCR5 Delta 32) является аллелем CCR5. [ 41 ] [ 42 ]
CCR5 Δ32-это делеция с 32 базой, которая вводит преждевременный стоп-кодон в локус рецептора CCR5, что приводит к нефункциональному рецептору. [ 43 ] [ 44 ] CCR5 необходим для ввода вируса ВИЧ-1 М-тропии. [ 45 ] Особощие люди, гомозиготные (обозначенные Δ32/Δ32) для CCR5 Δ32, не экспрессируют функциональные рецепторы CCR5 на своих клеточных поверхностях и устойчивы к инфекции ВИЧ-1 , несмотря на множественные воздействия высокого риска. [ 45 ] У индивидуумов, гетерозиготных (+/Δ32) для мутантного аллеля более 50% снижения функциональных рецепторов CCR5 на их клеточных поверхностях из-за димеризации между мутантными и рецепторами дикого типа, которые мешают транспортировке CCR5 к клеточной поверхности. [ 46 ] Гетерозиготные носители устойчивы к инфекции ВИЧ-1 по сравнению с дикими типами, и при заражении гетерозиготы демонстрируют снижение вирусных нагрузок и прогрессирование на 2-3 года к СПИДу относительно диких типов. [ 43 ] [ 45 ] [ 47 ] Гетерозиготность для этого мутантного аллеля также показала, что улучшает свой вирусологический ответ на антиретровирусное лечение. [ 48 ] CCR5 Δ32 имеет частоту аллеля (гетерозигот) в 9% в Европе, а гомозиготная частота - 1%. [ 49 ]
Недавнее исследование показывает, что CCR5 Δ32 усиливает познание и память. В 2016 году исследователи показали, что удаление гена CCR5 от мышей значительно улучшило их память. [ 50 ] CCR5 является мощным подавителем нейрональной пластичности, обучения и памяти; Чрезмерная активация CCR5 вирусными белками может способствовать когнитивному дефициту ВИЧ. [ 51 ]
Эволюционная история и возраст аллеля
[ редактировать ]Аллель CCR5 Δ32 известен его недавним происхождением, неожиданно высокой частотой и различным географическим распределением, [ 52 ] которые вместе предполагают, что (а) это возникло из одной мутации, и (б) исторически подвергался положительному отбору.
В двух исследованиях использовался анализ связей для оценки возраста делеции CCR5 Δ32, предполагая, что количество рекомбинации и мутации, наблюдаемых в геномных областях, окружающих делецию CCR5 Δ32, будут пропорциональными возрасту удаления. [ 42 ] [ 53 ] Используя выборку из 4000 человек из 38 этнических популяций, Stephens et al. По оценкам, делеция CCR5-Δ32 произошла 700 лет назад (275–1875, 95% доверительный интервал). Другая группа, Libert et al. (1998), использовали микросателлитные мутации для оценки возраста мутации CCR5 Δ32, которая составляет 2100 лет (700–4800, 95% доверительный интервал). На основании наблюдаемых событий рекомбинации они оценили возраст мутации 2250 лет (900–4700, 95% доверительный интервал). [ 53 ] Третья гипотеза зависит от градиента частоты аллелей в Европе с севера на юг, которая показывает, что самая высокая частота аллелей произошла в скандинавских странах и наименьшей частоте аллелей в Южной Европе. Поскольку викинги исторически занимали эти страны, возможно, что аллель распространился по всей Европе из -за рассеивания викингов в 8-10 веках. [ 54 ] Викинги были позже заменены варангянами в России, которые, возможно, способствовали наблюдаемому клину с восток-запад частоты аллеля. [ 52 ] [ 54 ]
ВИЧ-1 первоначально передавался от шимпанзе ( пан-троглодитов ) на человека в начале 1900-х годов на юго-востоке Камеруна, Африка, [ 55 ] Посредством воздействия инфицированных крови и жидкостей для тела во время битья по бушке. [ 56 ] Однако ВИЧ-1 эффективно отсутствовал в Европе до 1980-х годов. [ 57 ] Следовательно, учитывая средний возраст примерно 1000 лет для аллеля CCR5-Δ32, можно установить, что ВИЧ-1 не оказывал давления отбора на человеческую популяцию достаточно долго для достижения текущих частот. [ 52 ] Следовательно, другие патогенные микроорганизмы были предложены в качестве агентов положительного отбора для CCR5 Δ32, включая бубонскую чуму ( Yersinia pestis ) и оспа ( Variola мажор ). Другие данные свидетельствуют о том, что частота аллелей испытывала негативное давление отбора в результате патогенов, которые стали более распространенными во время римского расширения. [ 58 ] Идея о том, что негативный отбор сыграл роль в низкой частоте аллеля, также подтверждается экспериментами с использованием нокаутированных мышей и гриппа А, что продемонстрировало, что присутствие рецептора CCR5 важно для эффективного ответа на патоген. [ 59 ] [ 60 ]
Доказательства одной мутации
[ редактировать ]Несколько линий доказательств предполагают, что аллель CCR5 Δ32 развивался только один раз. [ 52 ] Во -первых, CCR5 Δ32 имеет относительно высокую частоту в нескольких различных европейских популяциях, но сравнительно отсутствует в азиатских, ближневосточных и американских индейских населениях, [ 42 ] предполагая, что одна мутация произошла после расхождения европейцев от их африканского предка. [ 42 ] [ 43 ] [ 61 ] Во -вторых, анализ генетического сцепления указывает на то, что мутация происходит на гомогенном генетическом фоне, подразумевая, что наследование мутации происходит от общего предка. [ 53 ] Это было продемонстрировано, показывая, что аллель CCR5 Δ32 находится в сильной неравновесии сцепления с высокополиморфными микросателлитами. Более 95% хромосом CCR5 Δ32 также несли аллель IRI3,1-0, в то время как 88% несли аллель IRI3,2. Напротив, микросателлитные маркеры IRI3.1-0 и IRI3.2-0 были обнаружены только в 2 или 1,5% хромосом, несущих аллель CCR5 дикого типа. [ 53 ] Это свидетельство неравновесного сцепления подтверждает гипотезу о том, что большинство, если не все, аллели CCR5 Δ32 возникли из одного мутационного события. Наконец, аллель CCR5 Δ32 имеет уникальное географическое распределение, указывающее на одно северное происхождение, за которым следует миграция. Исследование, измеряющее частоты аллелей в 18 европейских популяциях, обнаружило градиент с севера на юг, с самыми высокими частотами аллелей в финских и морвинианских популяциях (16%) и самыми низкими в сардинии (4%). [ 53 ]
Положительный выбор
[ редактировать ]В отсутствие отбора одна мутация потребуется около 127 500 лет, чтобы увеличить частоту населения 10%. [ 42 ] Оценки, основанные на уровне генетической рекомбинации и уровня мутаций, устанавливают возраст аллеля от 1000 до 2000 лет. Это расхождение является подписью положительного отбора.
По оценкам, ВИЧ-1 вошел в население в Африке в начале 1900-х годов, [ 55 ] Но симптоматические инфекции не сообщались до 1980 -х годов. Следовательно, эпидемия ВИЧ-1 слишком молода, чтобы быть источником положительного отбора, который привел к частоте CCR5 Δ32 от нуля до 10% в 2000 году.
Защита от бубонной чумы . Стивенс и др. (1998), предполагают, что бубонная чума ( Yersinia pestis ) оказывала положительное селективное давление на CCR5 Δ32. [ 42 ] Эта гипотеза была основана на сроках и серьезности пандемии черной смерти, в результате которой погибли 30% европейского населения всех возрастов в возрасте от 1346 до 1352 года. [ 62 ] После черной смерти были менее тяжелые, прерывистые эпидемии. Отдельные города испытывали высокую смертность, но общая смертность в Европе составляла всего несколько процентов. [ 62 ] [ 63 ] [ 64 ] В 1655-1656 гг. Вторая пандемия под названием «Великая чума» убила 15-20% населения Лондона. [ 65 ] [ 66 ] [ сомнительно - обсудить ] Важно отметить, что эпидемии чумы были прерывистыми. Бубонная чума - это зоонозная болезнь, в первую очередь инфицирующую грызуны, распространяется от блох и лишь изредка инфицирует людей. [ 67 ] От человеческой инфекции бубонной чумы не возникает, хотя это может возникнуть в пневмонической чуме, которая заражает легкие. [ 68 ] Только когда плотность грызунов низкая, инфицированные блохи вынуждены питаться альтернативными хозяевами, такими как люди, и при этих обстоятельствах может возникнуть эпидемия человека. [ 67 ] Основываясь на популяционных генетических моделях, Galvani и Slatkin (2003) утверждают, что прерывистый характер эпидемий чумы не генерировал достаточно сильную селективную силу для управления частотой аллелей CCR5 Δ32 до 10% в Европе. [ 41 ] Чтобы проверить эту гипотезу, Galvani и Slatkin (2003) смоделировали историческое давление отбора, создаваемое чумой и оспой. [ 41 ]
Защита от оспы . Чума была смоделирована в соответствии с историческими отчетами, [ 69 ] [ 70 ] В то время как возрастная смертность от оспы была получена из возрастного распределения захоронения в слабой оспе в Йорке (Англия) между 1770 и 1812 годами. [ 63 ] Смолл оспа преимущественно заражает молодых, допродуктивных представителей населения, поскольку они являются единственными людьми, которые не иммунизированы или не мертвы от прошлой инфекции. Поскольку малая оспа преимущественно убивает допродуктивные члены населения, она создает более сильное селективное давление, чем чума. [ 41 ] В отличие от чумы, оспа не имеет резервуара животных и передается только от человека к человеку. [ 71 ] [ 72 ] Авторы подсчитали, что если бы чума выбирала для CCR5 Δ32, частота аллеля по -прежнему составила бы менее 1%, в то время как оспа оказывает селективную силу, достаточную для достижения 10%.
Гипотеза о том, что оспа проявляла положительный отбор для CCR5 Δ32, также является биологически правдоподобной, поскольку паксвирусы, такие как ВИЧ, входят в лейкоциты с использованием рецепторов хемокинов. [ 73 ] Напротив, Yersinia pestis - бактерия с совершенно другой биологией.
Хотя европейцы являются единственной группой, имеющей субпопуляции с высокой частотой CCR5 Δ32, они не единственная популяция, которая подвергалась отбору с помощью оспы, которая имела мировое распределение до того, как оно было объявлено, уничтожено в 1980 году. Самые ранние безошибочные описания оспа появляется в 5 веке нашей эры в Китае, в 7 веке нашей эры в Индии и Средиземноморье, а также в 10 веке нашей эры на юго -западе Азия. [ 72 ] Напротив, мутация CCR5 Δ 32 обнаружена только в европейских, западной азиатской и североафриканской популяции. [ 74 ] Аномально высокая частота CCR5 Δ32 в этих популяциях, по -видимому, требует как уникального происхождения в Северной Европе, так и последующего отбора по оспе.
Потенциальные затраты
[ редактировать ]CCR5 Δ32 может быть полезен для хозяина при некоторых инфекциях (например, ВИЧ-1, возможно, оспа), но в других в других (например, клещевой энцефалит , вирус Западного Нила ). Независимо от того, является ли функция CCR5 полезной или вредной в контексте данной инфекции, зависит от сложного взаимодействия между иммунной системой и патогеном. [ 75 ]
В целом, исследования показывают, что мутация CCR5 Δ32 может играть вредную роль в воспалительных процессах после инфекции, которые могут повредить ткани и создавать дополнительную патологию. [ 76 ] Лучшее доказательство этой предложенной антагонистической плейотропии обнаружено при флавивирусных инфекциях. В целом многие вирусные инфекции являются бессимптомными или вызывают только легкие симптомы в подавляющем большинстве населения. Тем не менее, некоторые несчастные люди испытывают особенно разрушительный клинический курс, который в противном случае необъяснимо, но, по -видимому, генетически опосредован. Было обнаружено, что пациенты, гомозиготные по CCR5 Δ32, подвержены более высокому риску нейроинвазивной формы энцефалита клещей, вызванного флавивирусом ). [ 77 ] Кроме того, функциональный CCR5 может потребоваться для предотвращения симптоматических заболеваний после заражения вирусом Западного Нила, еще одним флавивирусом; CCR5 Δ32 был связан с ранним развитием симптомов и более выраженными клиническими проявлениями после заражения вирусом Западного Нила. [ 78 ]
Это открытие у людей подтвердило ранее наблюдаемый эксперимент на животной модели гомозиготности CCR5 Δ32. После заражения вирусом Западного Нила мыши CCR5 Δ32 значительно увеличили вирусные титры в центральной нервной системе и имели повышенную смертность [ 79 ] По сравнению с экспрессией мышей дикого типа, тем самым предполагая, что экспрессия CCR5 была необходима для укрепления сильной защиты хозяина от вируса Западного Нила.
Медицинские заявки
[ редактировать ]Генетический подход с участием внутрибодий, который блокирует экспрессию CCR5, был предложен в качестве лечения ВИЧ-1 . инфицированных [ 80 ] Когда Т-клетки модифицированы таким образом, чтобы они больше не экспрессировали CCR5, смешанные с немодифицированными Т-клетками, экспрессирующими CCR5, а затем бросали вызов инфекции ВИЧ-1, модифицированные Т-клетки, которые не выражают CCR5 в конечном итоге захватывают культуру, как ВИЧ-1 Убивает не модифицированные Т-клетки. Этот же метод может быть использован in vivo для создания устойчивого к вирус-клеточным пулам у инфицированных людей. [ 80 ]
Эта гипотеза была протестирована у пациента СПИДа, у которого также развился миелоидный лейкоз , и получала химиотерапию , чтобы подавить рак. Трансплантат костного мозга, содержащий стволовые клетки от соответствующего донора, затем использовалась для восстановления иммунной системы. Тем не менее, трансплантация была выполнена из донора с 2 копиями мутационного гена CCR5-Δ32. Через 600 дней пациент был здоровым и имел неопределяемые уровни ВИЧ в крови и в исследованных тканях мозга и прямой кишки. [ 7 ] [ 81 ] Перед пересадкой также были обнаружены низкие уровни ВИЧ X4 , который не использует рецептор CCR5. Однако после пересадки этот тип ВИЧ также не был обнаружен. [ 7 ] Тем не менее, этот результат согласуется с наблюдением, что клетки, экспрессирующие вариант CCR5-Δ32, имеют рецепторы CCR5 и CXCR4 на их поверхностях, тем самым обеспечивая устойчивость к широкому диапазону вариантов ВИЧ, включая ВИЧК4. [ 82 ] Через более шести лет пациент поддерживал устойчивость к ВИЧ и был произносится из лечения ВИЧ -инфекции. [ 8 ]
В 2009 году было начато регистрация ВИЧ-позитивных пациентов в клиническое исследование, в котором клетки пациентов были генетически модифицированы нуклеазой цинкового пальца для переноса признака CCR5-Δ32, а затем реинтродузированы в организм в качестве потенциального лечения ВИЧ. [ 83 ] [ 84 ] Результаты, о которых сообщалось в 2014 году, были многообещающими. [ 11 ]
Вдохновленный первым человеком, когда -либо вылеченным от ВИЧ, пациента с Берлином , Стемцит начал сотрудничество с банками пуповинной крови по всему миру, чтобы систематически скринировать образцы пуповинной крови для мутации CCR5, начиная с 2011 года. [ 85 ] [ 86 ] [ 87 ]
В ноябре 2018 года Цзянкуи объявил, что отредактировал два человека -эмбриона, чтобы попытаться отключить ген для CCR5, который кодирует рецептор, который ВИЧ использует для входа в ячейки. Он сказал, что девочки -близнецы, Лулу и Нана , родились несколькими неделями ранее, и что девочки все еще несли функциональные копии CCR5 вместе с инвалидом CCR5 ( мозаицизм ), поэтому все еще были уязвимы для ВИЧ. Работа была широко осуждена как неэтичная, опасная и преждевременная. [ 88 ] [ 89 ]
Смотрите также
[ редактировать ]- Открытие и развитие антагонистов рецепторов CCR5
- Ингибитор входа
- ВИЧ Тропизм
- Стивен Крон
- ВИЧ -иммунитет
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в GRCH38: Ensembl Release 89: ENSG00000160791 - ENSEMBL , май 2017 г.
- ^ Jump up to: а беременный в GRCM38: Ensembl Release 89: Ensmusg00000079227 - Ensembl , май 2017 г.
- ^ «Человеческая PubMed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «Мышь Pubmed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ Jump up to: а беременный Jiao X, Nawab O, Patel T, Kossenkov AV, Halama N, Jaeger D, et al. (Октябрь 2019). «Недавние достижения, нацеленные на CCR5 для рака и его роль в иммуноонкологии» . РАНКА . 79 (19): 4801–4807. doi : 10.1158/0008-5472.can-19-1167 . PMC 6810651 . PMID 31292161 .
- ^ Де Сильва Е, Stumpf MP (декабрь 2004 г.). «ВИЧ и аллель сопротивления CCR5-Delta32» . Письма микробиологии FEMS . 241 (1): 1–12. doi : 10.1016/j.femsle.2004.09.040 . PMID 15556703 .
- ^ Jump up to: а беременный в Hütter G, Nowak D, Mossner M, Ganepola S, Müssig A, Allers K, et al. (Февраль 2009 г.). «Долгосрочный контроль ВИЧ с помощью трансплантации стволовых клеток CCR5 Delta32/Delta32» . Новая Англия Журнал медицины . 360 (7): 692–698. doi : 10.1056/nejmoa0802905 . PMID 19213682 .
- ^ Jump up to: а беременный Allers K, Hütter G, Hofmann J, Loddenkemper C, Rieger K, Thiel E, et al. (Март 2011 г.). «Свидетельство о лечении ВИЧ -инфекции с помощью трансплантации стволовых клеток CCR5Δ32/Δ32» . Кровь . 117 (10): 2791–2799. doi : 10.1182/blood-2010-09-309591 . PMID 21148083 .
- ^ Zhen A, Kitchen S (декабрь 2013 г.). «Генная терапия на основе стволовых клеток при ВИЧ-инфекции» . Вирусы . 6 (1): 1–12. doi : 10.3390/v6010001 . PMC 3917429 . PMID 24368413 .
- ^ Кей Ма, Уокер Б.Д. (март 2014 г.). «Инженерная клеточная устойчивость к ВИЧ». Новая Англия Журнал медицины . 370 (10): 968–969. doi : 10.1056/nejme1400593 . PMID 24597871 .
- ^ Jump up to: а беременный Tebas P, Stein D, Tang WW, Frank I, Wang SQ, Lee G, et al. (Март 2014 г.). «Редактирование генов CCR5 в аутологичных CD4 T -клетках лиц, инфицированных ВИЧ» . Новая Англия Журнал медицины . 370 (10): 901–910. doi : 10.1056/nejmoa1300662 . PMC 4084652 . PMID 24597865 .
- ^ Jump up to: а беременный «CCR5 - хемокин (CC Motif) Рецептор 5 (ген/псевдоген)» . Генетика дома ссылка . Архивировано с оригинала 24 сентября 2009 года . Получено 30 августа 2009 года .
- ^ Samson M, Labbe O, Mollereau C, Vassart G, Parmentier M (март 1996 г.). «Молекулярное клонирование и функциональная экспрессия нового гена рецептора CC-хемокина человека». Биохимия . 35 (11): 3362–3367. doi : 10.1021/bi952950g . PMID 8639485 .
- ^ Jump up to: а беременный Struyf S, Menten P, Lenaerts JP, Pot W, D'Haese A, De Clercq E, et al. (Июль 2001 г.). «Расходящая связывающая способность природных изоформ LD78Beta воспалительного белка-1-млэфа макрофагов к рецепторам хемокинов CC CC 7 и 5 влияют на их активность против HIV-1 и хемотаксические потенции для нейтрофилов и эозинофилов» . Европейский журнал иммунологии . 31 (7): 2170–2178. doi : 10.1002/1521-4141 (200107) 31: 7 <2170 :: AID-IMMU2170> 3.0.CO; 2-D . PMID 11449371 .
- ^ Миякава Т., Обару К., Маеда К., Харада С., Мицуя Х (февраль 2002 г.). «Идентификация аминокислотных остатков, критических для LD78-бета, вариант воспалительного белка макрофага человека, связывания с CCR5 и ингибирования репликации вируса иммунодефицита R5 типа 1 типа» . Журнал биологической химии . 277 (7): 4649–4655. doi : 10.1074/jbc.m109198200 . PMID 11734558 .
- ^ Slimani H, Charnaux N, Memba E, Saffar L, Vassy R, Vita C, et al. (Октябрь 2003 г.). «Взаимодействие Rantes с Syndecan-1 и Syndecan-4 экспрессируется первичными макрофагами человека » Biochimica et Biophysica Acta (BB) - Biomemmbranes 1617 (1–2): 80–8 Doi : 10.1016/ j.beem.2003.09.0 14637022PMID
- ^ Proudfoot AE, Fritchley S, Borlat F, Shaw JP, Vilbois F, Zwahlen C, et al. (Апрель 2001 г.). «Мотив BBXB RANTES является основным сайтом для связывания гепарина и контроля селективности рецепторов» . Журнал биологической химии . 276 (14): 10620–10626. doi : 10.1074/jbc.m010867200 . PMID 11116158 .
- ^ Jump up to: а беременный в дюймовый и фон Velasco-Velázquez M, Jiao X, De la Fuente M, Pestell TG, Ettel A, Lisanti MP, et al. (Август 2012 г.). «Антагонист CCR5 блокирует метастазирование базальных клеток рака молочной железы» . РАНКА . 72 (15): 3839–3850. Doi : 10.1158/0008-5472.can-1-3917 . PMID 22637726 .
- ^ Jump up to: а беременный в дюймовый Sicoli D, Jiao X, Ju X, Velasc-velazquez M, Ertel A, Addya S, et al. (Декабрь 2014). «Антагонисты рецепторов CCR5 блокируют метастазирование в кость V-Src, трансформированные метастатическими клеточными линиями раковой раковой простаты,» . РАНКА . 74 (23): 7103–7114. doi : 10.1158/0008-5472.can-14-0612 . PMC 4294544 . PMID 25452256 .
- ^ Velasco-Velázquez M, Xolalpa W, Pestell RG (ноябрь 2014). «Потенциал для нацеливания CCL5/CCR5 при раке молочной железы». Экспертное мнение о терапевтических целях . 18 (11): 1265–1275. doi : 10.1517/147282222.2014.949238 . PMID 25256399 . S2CID 7976259 .
- ^ Jump up to: а беременный Jiao X, Velasco-Velázquez MA, Wang M, Li Z, Rui H, Peck AR, et al. (Апрель 2018). «CCR5 регулирует восстановление повреждения ДНК и расширение стволовых клеток рака молочной железы» . РАНКА . 78 (7): 1657–1671. doi : 10.1158/0008-5472.can-17-0915 . PMC 6331183 . PMID 29358169 .
- ^ Barmania F, Pepper MS (декабрь 2013 г.). «CC -хемокиновый рецептор типа пятый (CCR5): новая мишень для контроля ВИЧ -инфекции» . Прикладная и трансляционная геномика . 2 (а): 3–16. doi : 10.1016/j.atg.2013.05.004 . PMC 5133339 . PMID 27942440 .
- ^ Jasinska AJ, Pandrea I, Apetrei C (27 января 2022 г.). «CCR5 как корецептор вируса иммунодефицита человека и вирусов иммунодефицита обезьян: прототипический роман любви-ненависти» . Границы в иммунологии . 13 : 835994. DOI : 10.3389/fimmu.2022.835994 . ISSN 1664-3224 . PMC 8829453 . PMID 35154162 .
- ^ Chen Z, Kwon D, Jin Z, Monard S, Telfer P, Jones MS, et al. (Декабрь 1998). «Естественная инфекция гомозиготной дельты 24 CCR5 Mangabey с вирусом R2B-тропического иммунодефицита R2B-тропика» . Журнал экспериментальной медицины . 188 (11): 2057–2065. doi : 10.1084/jem.188.11.2057 . PMC 2212380 . PMID 9841919 .
- ^ Paiardini M, Cervasi B, Reyes-Aviles E, Micci L, Ortiz AM, Chahroudi A, et al. (Июнь 2011 г.). «Низкие уровни инфекции SIV в CD⁴⁺ CD -клетках центральной памяти Mangabey, связанные с ограниченной экспрессией CCR5» . Природная медицина . 17 (7): 830–836. doi : 10.1038/nm.2395 . PMC 3253129 . PMID 21706028 .
- ^ Ma D, Jasinska AJ, Feyertag F, Wijewardana V, Kristoff J, He T, et al. (Май 2014). «Факторы, связанные с передачей вируса иммунодефицита Симана в естественном африканском нечеловеческом примате в дикой природе» . Журнал вирусологии . 88 (10): 5687–5705. doi : 10.1128/jvi.03606-13 . PMC 4019088 . PMID 24623416 .
- ^ Pandrea I, Onanga R, Soukiere S, Mouinga-Oondéme A, Bourry O, Makuwa M, et al. (Июнь 2008 г.). «Недостаток CD4+ CCR5+ T-клеток может предотвратить передачу вируса иммунодефицита SIMIIAN у естественных нечеловеческих приматов с помощью Breest-Feeling » Журнал вирусологии 82 (11): 5501–5 Doi : 10.1128/ jvi.025555-0 2395173PMC 18385229PMID
- ^ Jump up to: а беременный в дюймовый и Murphy PM (февраль 2001 г.). «Вирусная эксплуатация и подрыв иммунной системы посредством мимики хемокинов» . Природа иммунология . 2 (2): 116–122. doi : 10.1038/84214 . PMID 11175803 . S2CID 29364407 .
- ^ Jump up to: а беременный в дюймовый Алхатиб G (март 2009 г.). «Биология CCR5 и CXCR4» . Текущее мнение в ВИЧ и СПИДе . 4 (2): 96–103. doi : 10.1097/coh.0b013e328324bbec . PMC 2718543 . PMID 19339947 .
- ^ Jump up to: а беременный в дюймовый «Ингибиторы CCR5 и ВИЧ» .
- ^ Андерсон Дж., Аккина Р (сентябрь 2007 г.). «Полный нокдаун CCR5 с помощью лентивирусных векторов-экспрессированных миРНК и защита трансгенных макрофагов от инфекции ВИЧ-1» . Генная терапия . 14 (17): 1287–1297. doi : 10.1038/sj.gt.3302958 . PMID 17597795 .
- ^ Jump up to: а беременный в Lieberman-Blum SS, Fung HB, Bandres JC (июль 2008 г.). «Маравирок: антагонист CCR5-рецептора для лечения ВИЧ-1 инфекции». Клиническая терапия . 30 (7): 1228–1250. doi : 10.1016/s0149-2918 (08) 80048-3 . PMID 18691983 .
- ^ Назари Р., Джоши С (август 2008 г.). «CCR5 как мишень для генной терапии ВИЧ-1». Современная генная терапия . 8 (4): 264–272. doi : 10.2174/156652308785160674 . PMID 18691022 .
- ^ Starr DB (19 ноября 2009 г.). «Любые люди генетически предрасположены к тому, чтобы быть невосприимчивыми к ВИЧ?» Полем Технологический интерактивный . Спросите генетика . Получено 5 августа 2024 года .
- ^ Тейлор Б.М., Фулке Дж.С., Флинко Р., Едиа А., Девико А., Рейц М (июнь 2008 г.). «Изменение вируса иммунодефицита человека GP41 приводит к снижению зависимости CCR5 и независимости CD4» . Журнал вирусологии . 82 (11): 5460–5471. doi : 10.1128/jvi.01049-07 . PMC 2395218 . PMID 18353949 .
- ^ Jiao X, Wang M, Zhang Z, Li Z, Ni D, Ashton AW, et al. (Январь 2021 г.). «Леронлимаб, гуманизированное моноклональное антитело к CCR5, блокирует метастазирование клеток рака рака молочной железы и усиливает гибель клеток, вызванную химиотерапией ДНК, повреждающей химиотерапию» . Исследование рака молочной железы . 23 (1): 11. doi : 10.1186/s13058-021-01391-1 . PMC 7825185 . PMID 33485378 .
- ^ Франкенбергер С., Рабе Д., Бейнер Р., Санкарашарма Д., Чада К., Крауш Т. и др. (Октябрь 2015). «Супрессоры метастазирования регулируют микроокружение опухоли, блокируя рекрутирование макрофагов, ассоциированных с опухолью» . РАНКА . 75 (19): 4063–4073. doi : 10.1158/0008-5472.can-14-3394 . PMC 4592465 . PMID 26238785 .
- ^ Halama N, Zoernig I, Berthel A, Kahlert C, Klupp F, Suarez-Carmona M, et al. (Апрель 2016 г.). «Эксплуатация опухолевых иммунных клеток при метастазах с колоректальным раком может эффективно подвергаться терапии против CCR5 у больных раком» . Раковая клетка . 29 (4): 587–601. doi : 10.1016/j.ccell.2016.03.005 . PMID 27070705 .
- ^ Jump up to: а беременный «Препарат ВИЧ может улучшить выздоровление после инсульта» . Sciencemag . 21 февраля 2019 года . Получено 22 февраля 2019 года .
- ^ Joy Mt, Ben Assayag E, Shabashov-Stone D, Liraz-Zaltsman S, Mazzitelli J, Arenas M, et al. (Февраль 2019 г.). «CCR5 является терапевтической мишенью для восстановления после инсульта и черепно -мозговой травмы» . Клетка . 176 (5): 1143–1157.e13. doi : 10.1016/j.cell.2019.01.044 . PMC 7259116 . PMID 30794775 .
- ^ Jump up to: а беременный в дюймовый Galvani AP, Slatkin M (декабрь 2003 г.). «Оценка чумы и оспы в качестве исторического селективного давления для аллеля CCR5-Delta 32 ВИЧ-устойчивости» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (25): 15276–15279. Bibcode : 2003pnas..10015276G . doi : 10.1073/pnas.2435085100 . PMC 299980 . PMID 14645720 .
- ^ Jump up to: а беременный в дюймовый и фон Стивенс Дж.С., Рейх Д.Е., Гольдштейн Д.Б., Шин Х.Д., Смит М.В., Каррингтон М. и др. (Июнь 1998 г.). «Датирование происхождением аллеля по борьбе с СПИД-дистанцией CCR5-DELTA32 путем коалесценции гаплотипов» . Американский журнал человеческой генетики . 62 (6): 1507–1515. doi : 10.1086/301867 . PMC 1377146 . PMID 9585595 .
- ^ Jump up to: а беременный в Дин М., Каррингтон М., Винклер С., Хаттли Г.А., Смит М.В., Алликметс Р. и др. (Сентябрь 1996). «Генетическое ограничение инфекции и прогрессирования ВИЧ-1 в СПИДе с помощью аллеля делеционного аллеля структурного гена CKR5. Исследование роста и развития гемофилии, многоцентровое когортное исследование СПИД, многоцентровое когортное исследование гемофилии, городское когорта Сан-Франциско, живое исследование» . Наука . 273 (5283): 1856–1862. Bibcode : 1996sci ... 273.1856d . doi : 10.1126/science.273.5283.1856 . PMID 8791590 . S2CID 21160580 .
- ^ Самсон М., Либерт Ф., Доранц Б.Дж., Ракер Дж., Лиснард С., Фарбер С.М. и др. (Август 1996). «Устойчивость к инфекции ВИЧ-1 у кавказских индивидуумов, несущих мутантные аллели гена рецептора хемокина CCR-5». Природа . 382 (6593): 722–725. Bibcode : 1996natur.382..722S . doi : 10.1038/3827222A0 . PMID 8751444 . S2CID 4370818 .
- ^ Jump up to: а беременный в Liu R, Paxton WA, Choe S, Ceradini D, Martin SR, Horuk R, et al. (Август 1996). «Гомозиготный дефект в корецепторе ВИЧ-1 объясняет устойчивость некоторых людей, подвергшихся множеству, к инфекции ВИЧ-1» . Клетка . 86 (3): 367–377. doi : 10.1016/s0092-8674 (00) 80110-5 . PMID 8756719 . S2CID 16965624 .
- ^ Бенкиране М., Джин Да, Чун Р.Ф., Куп Р.А., Жанг К.Т. (декабрь 1997 г.). «Механизм трансдоминанта ингибирования CCR5-опосредованной ВИЧ-1 инфекции CCR5DELTA32» . Журнал биологической химии . 272 (49): 30603–30606. doi : 10.1074/jbc.272.49.30603 . PMID 9388191 .
- ^ Майкл Н.Л., Луи Л.Г., Рорбол А.Л., Шульц К.А., Дейхофф Д.Е., Ван К.Е. и др. (Октябрь 1997). «Роль полиморфизмов CCR5 и CCR2 в передаче ВИЧ-1 и прогрессировании заболевания». Природная медицина . 3 (10): 1160–1162. doi : 10.1038/nm1097-1160 . PMID 9334732 . S2CID 23196048 .
- ^ Laurichesse JJ, Persoz A, Theodorou I, Rouzioux C, Delfraissy JF, Meyer L (май 2007 г.). «Улучшенный вирусологический ответ на высоко активную антиретровирусную терапию у пациентов с ВИЧ-1, несущими делецию CCR5 Delta32» . ВИЧ Медицина . 8 (4): 213–219. doi : 10.1111/j.1468-1293.2007.00455.x . PMID 17461848 . S2CID 24900064 .
- ^ Мерфи К., Уивер С. (2016). Иммунобиология Джейнвея 9 -е издание . Гарлендская наука. п. 582. ISBN 978-0-8153-4550-3 .
- ^ «Близнецы Китая CRISPR могли бы непреднамеренно улучшить их мозг» . Технологический обзор . Получено 22 февраля 2019 года .
- ^ Zhou M, Greenhill S, Huang S, Silva TK, Sano Y, Wu S, et al. (Декабрь 2016 г.). «CCR5 является подавителем кортикальной пластичности, обучения и памяти гиппокампа» . элиф . 5 doi : 10.7554/elife.20985 . PMC 5213777 . PMID 27996938 .
- ^ Jump up to: а беременный в дюймовый Galvani AP, Novembre J (февраль 2005 г.). «Эволюционная история мутации ВИЧ-устойчивости CCR5-Delta32» . Микробы и инфекция . 7 (2): 302–309. doi : 10.1016/j.micinf.2004.12.006 . PMID 15715976 .
- ^ Jump up to: а беременный в дюймовый и Libert F, Cochaux P, Beckman G, Samson M, Aksenova M, Cao A, et al. (Март 1998 г.). «Мутация Deltaccr5, обеспечивающая защиту от ВИЧ-1 в кавказских популяциях, имеет одно и недавнее происхождение в северо-восточной Европе» . Молекулярная генетика человека . 7 (3): 399–406. doi : 10.1093/hmg/7.3.399 . PMID 9466996 .
- ^ Jump up to: а беременный Lucotte G (сентябрь 2001 г.). «Распределение делеции гена CCR5 32-BasePair в Западной Европе. Гипотеза о возможной рассеивании мутации викингами в исторические времена». Человеческая иммунология . 62 (9): 933–936. doi : 10.1016/s0198-8859 (01) 00292-0 . PMID 11543895 .
- ^ Jump up to: а беременный Keele Bf, Van Heuverswyn F, Li Y, Bailes E, Takehisa J, Santiago ML, et al. (Июль 2006 г.). «Шимпанзе водохранилища пандемии и непенемического ВИЧ-1» . Наука . 313 (5786): 523–526. Bibcode : 2006sci ... 313..523K . doi : 10.1126/science.1126531 . PMC 2442710 . PMID 16728595 .
- ^ Hahn BH, Shaw Gm, de Cock KM, Sharp PM (январь 2000 г.). «СПИД как зооноз: научные последствия и последствия для общественного здравоохранения». Наука . 287 (5453): 607–614. Bibcode : 2000sci ... 287..607h . doi : 10.1126/science.287.5453.607 . PMID 10649986 .
- ^ Matic S. «ВИЧ/СПИД в Европе: переход от смертного приговора к лечению хронических заболеваний» (PDF) . Всемирная организация здравоохранения.
- ^ Faure E, Royer-Carenzi M (декабрь 2008 г.). «Является ли европейское пространственное распределение аллеля CCR5-Delta32, устойчивого к ВИЧ-1, образуется путем разбивки патоценоза из-за исторической римской экспансии?» (PDF) . Инфекция, генетика и эволюция . 8 (6): 864–874. Bibcode : 2008infge ... 8..864f . doi : 10.1016/j.meegid.2008.08.007 . PMID 18790087 . S2CID 32881202 .
- ^ Доусон Т.К., Бек М.А., Кузиэль В.А., Хендерсон Ф., Маеда Н. (июнь 2000 г.). «Контрастные эффекты дефицита CCR5 и CCR2 в легочном воспалительном ответе на вирус гриппа А» . Американский журнал патологии . 156 (6): 1951–1959. doi : 10.1016/s0002-9440 (10) 65068-7 . PMC 1850091 . PMID 10854218 .
- ^ Vargas AE, Cechim G, Correa JF, Gomes PA, Macedo GS, De Medeiros RM, et al. (Июль 2009 г.). «Плюсы и минусы отсутствующего рецептора хемокинов-на« Комментарии », является ли европейское пространственное распределение аллеля, устойчивого к ВИЧ-1 CCR5-D32. Ройер-Карензи (2008) ». Инфекция, генетика и эволюция . 9 (4): 387–389. doi : 10.1016/j.meegid.2009.01.001 . PMID 19472441 .
- ^ О'Брайен С.Дж., Дин М (сентябрь 1997 г.). «В поисках генов устойчивости к СПИД». Scientific American . 277 (3): 44–51. Bibcode : 1997sciam.277c..44o . doi : 10.1038/scientificamerican0997-44 . PMID 9274039 .
- ^ Jump up to: а беременный McEedy C (февраль 1988 г.). «Бубонная чума». Scientific American . 258 (2): 118–123. Bibcode : 1988sciam.258b.118m . doi : 10.1038/Scientificamerican0288-118 . PMID 3055286 .
- ^ Jump up to: а беременный Galley C (1998). Демография ранних современных городов: Йорк в шестнадцатом и семнадцатом веках . Лондон: Макмиллан.
- ^ Хэтчер Дж. (1977). Чума, население и английская экономика 1348-1530 . Лондон: Макмиллан.
- ^ «Великая чума Лондона, 1665» . Инфекция, исторические взгляды на болезни и эпидемии . Гарвардский университет . Получено 2 марта 2015 года .
- ^ «ДНК в лондонской могиле может помочь решить загадки великой чумы» . 8 сентября 2016 года. Архивировано с оригинала 9 сентября 2016 года . Получено 18 сентября 2016 года .
- ^ Jump up to: а беременный Килинг М.Дж., Гиллиган Калифорния (ноябрь 2000 г.). «Бубонная чума: модель метапопуляции зооноза» . Разбирательство. Биологические науки . 267 (1458): 2219–2230. doi : 10.1098/rspb.2000.1272 . PMC 1690796 . PMID 11413636 .
- ^ Appleby AB (1980). «Исчезновение чумы: продолжающаяся головоломка». Обзор экономической истории . 33 (2): 161–173. doi : 10.2307/2595837 . JSTOR 2595837 . PMID 11614424 .
- ^ Рассел JS (1948). Британское средневековое население . Альбукерке: Университет Нью -Мексико Пресс.
- ^ Twigg G (1984). Черная смерть: биологическая переоценка . Лондон: Batsfod.
- ^ Чапин CV (1913). «Различия в типе инфекционных заболеваний, как показано в истории оспы в Соединенных Штатах, 1815-1912» . J Infect Dis . 13 (2): 171–196. doi : 10.1093/infdis/13.2.171 .
- ^ Jump up to: а беременный Fenner F (1998). «Историческая виньетка: жизнь с оспами и издателями». Достижения в области исследования вируса . 51 : 1–33. doi : 10.1016/s0065-3527 (08) 60782-4 . ISBN 978-0-12-039851-5 Полем PMID 9891584 .
- ^ Lalani AS, Masters J, Zeng W, Barrett J, Pannu R, Everett H, et al. (Декабрь 1999). «Использование рецепторов хемокинов оспами». Наука . 286 (5446): 1968–1971. doi : 10.1126/science.286.5446.1968 . PMID 10583963 .
- ^ Novembre J, Galvani AP, Slatkin M (ноябрь 2005 г.). «Географическое распространение аллеля CCR5 Delta32 ВИЧ-устойчивости» . PLOS Биология . 3 (11): E339. doi : 10.1371/journal.pbio.0030339 . PMC 1255740 . PMID 16216086 .
- ^ Обриен С.Дж. (март 2024 г.). «Наследие магического гена- CCR5 -∆ 32 : от открытия до клинической выгоды в поколении» . Труды Национальной академии наук Соединенных Штатов Америки . 121 (12): E2321907121. Bibcode : 2024pnas..12121907O . doi : 10.1073/pnas.2321907121 . PMC 10962972 . PMID 38457490 .
- ^ Кляйн Р.С. (январь 2008 г.). «Движущаяся цель: множественная роль CCR5 в инфекционных заболеваниях» . Журнал инфекционных заболеваний . 197 (2): 183–186. doi : 10.1086/524692 . PMID 18179384 .
- ^ Киндберг Е., Микиен А., Акс С., Акерлинд Б., Вене С., Линдквист Л. и др. (Январь 2008 г.). «Удаление в гене рецептора хемокинов 5 (CCR5) связана с клещерожденным энцефалитом» . Журнал инфекционных заболеваний . 197 (2): 266–269. doi : 10.1086/524709 . PMID 18179389 .
- ^ Лим Дж.К., Луи Сай, Глейзер С., Джин С., Джонсон Б., Джонсон Х. и др. (Январь 2008 г.). «Генетический дефицит хемокинового рецептора CCR5 является сильным фактором риска симптоматической инфекции вируса Западного Нила: метаанализ 4 когортов в эпидемии США» . Журнал инфекционных заболеваний . 197 (2): 262–265. doi : 10.1086/524691 . PMID 18179388 .
- ^ Glass WG, Lim JK, Cholera R, Pletnev AG, Gao JL, Murphy PM (октябрь 2005 г.). «Рецептор хемокинов CCR5 способствует переносу лейкоцитов в мозг и выживание при инфекции вируса Западного Нила» . Журнал экспериментальной медицины . 202 (8): 1087–1098. doi : 10.1084/jem.20042530 . PMC 2213214 . PMID 16230476 .
- ^ Jump up to: а беременный Steinberger P, Andris-Widhopf J, Bühler B, Torbett BE, Barbas CF (январь 2000 г.). «Функциональная делеция рецептора CCR5 с помощью внутриклеточной иммунизации продуцирует клетки, которые рефрактерны к CCR5-зависимой инфекции ВИЧ-1 и слияния клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (2): 805–810. Bibcode : 2000pnas ... 97..805s . doi : 10.1073/pnas.97.2.805 . PMC 15412 . PMID 10639161 .
- ^ Schoofs M (7 ноября 2008 г.). «Доктор, мутация и потенциальное лекарство от СПИДа» . Wall Street Journal . Получено 15 декабря 2010 года .
- ^ Agrawal L, Lu X, Qingwen J, Vanhorn-Ali Z, Nicolescu IV, McDermott DH, et al. (Март 2004 г.). «Роль белка CCR5DELTA32 в устойчивости к R5, R5X4 и X4 вируса иммунодефицита человека типа 1 в первичных клетках CD4+» . Журнал вирусологии . 78 (5): 2277–2287. doi : 10.1128/JVI.78.5.2277-2287.2004 . PMC 369216 . PMID 14963124 .
- ^ «Аутологичные Т-клетки, генетически модифицированные в гене CCR5 с помощью нуклеаз цинковых пальцев SB-728 для ВИЧ (цинковый пальцев)» . Национальные институты здравоохранения США. 9 декабря 2009 г. Получено 30 декабря 2009 года .
- ^ WADE N (28 декабря 2009 г.). «Цинковые пальцы могут быть ключом к возрождению генной терапии» . New York Times . Получено 30 декабря 2009 года .
- ^ Corbyn Z (июль 2012 г.). «План запущен, чтобы найти лечение от ВИЧ» . Лансет . 380 (9838): 203–204. doi : 10.1016/s0140-6736 (12) 61199-4 . PMID 22826833 . S2CID 34773481 .
- ^ Гонсалес Г., Парк С., Чен Д., Армитаж С., Шпалл Е., Бехрингер Р. (сентябрь 2011 г.). «Идентификация и частота CCR5Δ32/Δ32-устойчивых к ВИЧ-резистентным подразделениям из больниц Хьюстонского района» . ВИЧ Медицина . 12 (8): 481–486. doi : 10.1111/j.1468-1293.2010.00911.x . PMC 4021858 . PMID 21375684 .
- ^ Petz LD, Burnett JC, Li H, Li S, Tonai R, Bakalinskaya M, et al. (2015). «Прогресс в направлении лечения ВИЧ -инфекции с трансплантацией гематопоэтических клеток» . Стволовые ячейки и клонирование: достижения и применение . 8 : 109–116. doi : 10.2147/sccaa.s56050 . PMC 4524463 . PMID 26251620 .
- ^ Begley S (28 ноября 2018 г.). «Среди взрыва китайский ученый защищает детей, отредактированных по генам» . Статистика
- ^ Коллинз Ф.С. (28 ноября 2018 г.). «Заявление о претензии первых детей, отредактированных по генам китайским исследователем» . Национальные институты здравоохранения (NIH) . Министерство здравоохранения и социальных служб США.
Дальнейшее чтение
[ редактировать ]- Уилкинсон Д. (сентябрь 1996 г.). «Кофакторы предоставляют клавиши входа. ВИЧ-1» . Текущая биология . 6 (9): 1051–1053. doi : 10.1016/s0960-9822 (02) 70661-1 . PMID 8805353 . S2CID 18710567 .
- Broder CC, Dimitrov DS (1996). «ВИЧ и 7-трансмембранные доменные рецепторы» . Патобиология . 64 (4): 171–179. doi : 10.1159/000164032 . PMID 9031325 .
- Чо Х., Мартин К.А., Фарзан М., Содерски Дж., Джерард Н.П., Джерард С (июнь 1998 г.). «Структурные взаимодействия между рецепторами хемокинов, gp120 env и cd4». Семинары по иммунологии . 10 (3): 249–257. doi : 10.1006/smim.1998.0127 . PMID 9653051 .
- Sheppard HW, Celum C, Michael NL, O'Brien S, Dean M, Carrington M, et al. (Март 2002 г.). «Инфекция ВИЧ-1 у людей с генотипом CCR5-DELTA32/Delta32: приобретение вируса, индуцирующего синцития при сероконверсии». Журнал приобретенных синдромов иммунной дефицита . 29 (3): 307–313. doi : 10.1097/00042560-200203010-00013 . PMID 11873082 .
- Freedman BD, Liu QH, Del Corno M, Collman RG (2003). «ВИЧ-1 GP120-рецептор-рецептор-опосредованная передача сигналов в макрофагах человека». Иммунологические исследования . 27 (2–3): 261–276. doi : 10.1385/ir: 27: 2-3: 261 . PMID 12857973 . S2CID 32006625 .
- Esté Ja (сентябрь 2003 г.). «Вход вируса в качестве мишени для вмешательства против ВИЧ». Текущая лекарственная химия . 10 (17): 1617–1632. doi : 10.2174/0929867033457098 . PMID 12871111 .
- Gallo SA, Finnegan CM, Viard M, Raviv Y, Dimitrov A, Rawat SS, et al. (Июль 2003 г.). «Опосредованная ВИЧ ENV-реакция слияния» . Biochemica et Biophysica Acta (BBA) - Biomembranes . 1614 (1): 36–50. Doi : 10.1016/s0005-2736 (03) 00161-5 . PMID 12873764 .
- Zaitseva M, Peden K, Golding H (июль 2003 г.). «Корецепторы ВИЧ: роль структуры, посттрансляционных модификаций и интернализация в слиянии вирусных клеток и в качестве мишеней для ингибиторов проникновения» . Biochimica et Biophysica Acta (BBA) - Biomembranes . 1614 (1): 51–61. doi : 10.1016/s0005-2736 (03) 00162-7 . PMID 12873765 .
- Lee C, Liu QH, Tomkowicz B, Yi Y, Freedman BD, Collman RG (ноябрь 2003 г.). «Активация макрофагов через CCR5- и CXCR4-опосредованные сигнальные пути, вызванные GP120» . Журнал биологии лейкоцитов . 74 (5): 676–682. doi : 10.1189/jlb.0503206 . PMID 12960231 . S2CID 11362623 .
- Йи Й., Ли С., Лю К.Х., Фридман Б.Д., Колман Р.Г. (2004). «Утилизация рецепторов хемокинов и передача сигналов макрофагов с помощью вируса иммунодефицита человека GP120 типа 1: последствия для невропатогенеза». Журнал нейровирологии . 10 (Suppl 1): 91–96. doi : 10.1080/753312758 . PMID 14982745 . S2CID 9065929 .
- Seibert C, Sakmar TP (2004). «Антагонисты мелкомолекулярных антагонистов CCR5 и CXCR4: многообещающий новый класс лекарств против HIV-1». Текущий фармацевтический дизайн . 10 (17): 2041–2062. doi : 10.2174/1381612043384312 . PMID 15279544 .
- Cutler CW, Jotwani R (апрель 2006 г.). «Экспрессия слизистой оболочки полости рта рецепторов ВИЧ-1, корецепторов и альфа-дефенсинов: таблица резистентности или восприимчивости к ВИЧ-инфекции?» Полем Достижения в области стоматологических исследований . 19 (1): 49–51. doi : 10.1177/154407370601900110 . PMC 3750741 . PMID 16672549 .
- Ajuebor MN, Carey JA, Swain MG (август 2006 г.). "CCR5 при заболеваниях печени, опосредованных Т-клетками: что происходит?" Полем Журнал иммунологии . 177 (4): 2039–2045. doi : 10.4049/jimmunol.177.4.2039 . PMID 16887960 .
- Липп М., Мюллер Г. (2003). «Формирование адаптивного иммунитета: влияние CCR7 и CXCR5 на обороты лимфоцитов». Переговоры немецкого патологического общества . 87 : 90-101. PMID 16888899 .
- Balisteri CR, Caruso C, Grimaldi MP, Listì F, Vasto S, Orlando V, et al. (Апрель 2007 г.). «Рецептор CCR5: биологические и генетические последствия при возрастных заболеваниях». Анналы нью -йоркской академии наук . 1100 (1): 162–172. Bibcode : 2007nyasa1100..162b . doi : 10.1196/annals.1395.014 . PMID 17460174 . S2CID 8437349 .
- Мэдсен Хо, Поулсен К., Даль О., Кларк Б.Ф., Хьорт Дж.П. (март 1990 г.). «Retropseudogenes составляют основную часть семейства Alpha Gene Factor 1 человека» . Исследование нуклеиновых кислот . 18 (6): 1513–1516. doi : 10.1093/nar/18.6.1513 . PMC 330519 . PMID 2183196 .
- Uetsuki T, Naito A, Nagata S, Kaziro Y (апрель 1989 г.). «Выделение и характеристика хромосомного гена человека для полипептидной цепи удлинения Фактора 1 Альфа-1» . Журнал биологической химии . 264 (10): 5791–5798. doi : 10.1016/s0021-9258 (18) 83619-5 . PMID 2564392 .
- WhiteHeart SW, Shenbagamurthi P, Chen L, Cotter RJ, Hart GW (август 1989 г.). «Фактор удлинения мыши 1 Альфа (EF-1 альфа) посттрансляционно модифицируется новыми амид-связанными этаноламино-фосфоглицериновыми фрагментами. Добавление этаноламина-фосфоглицерина к специфическим остаткам глутаминовой кислоты на алфа-1 AF-1» » . Журнал биологической химии . 264 (24): 14334–14341. doi : 10.1016/s0021-9258 (18) 71682-7 . PMID 2569467 .
- Энн Д.К., Ву М.М., Хуан Т., Карлсон Д.М., Ву Р (март 1988 г.). «Ретинол-регулируемая экспрессия генов в эпителиальных эпителиальных клетках человека. Увеличенная экспрессия фактора удлинения EF-1 альфа» . Журнал биологической химии . 263 (8): 3546–3549. doi : 10.1016/s0021-9258 (18) 68958-6 . PMID 3346208 .
- Брэндс Дж. Х., Массен Дж.А., Ван Хемерт Ф.Дж., Амонс Р., Мёллер В. (февраль 1986 г.). «Основная структура альфа-субъединицы фактора удлинения человека 1. Структурные аспекты гуанино-нуклеотидсвязывающих сайтов» . Европейский журнал биохимии . 155 (1): 167–171. doi : 10.1111/j.1432-1033.1986.tb09472.x . PMID 3512269 .
Внешние ссылки
[ редактировать ]- Видео и текст из документального фильма PBS о обнаружении CCR5
- «Рецепторы хемокинов: CCR5» . База данных IUPHAR рецепторов и ионных каналов . Международный союз базовой и клинической фармакологии. Архивировано из оригинала 18 января 2021 года . Получено 21 июля 2006 года .
- ГИВКОПРЕТ СЕРВЕР ДЛЯ ПРЕДИСЛОЖЕНИЯ ИСПОЛЬЗОВАНИЯ КОРИЗАТОРА ВИЧ (CCR5). Plos one 8 (4): e61437
- человека CCR5 Расположение генома CCR5 и страница гена в браузере UCSC Genome .
- Обзор всей структурной информации, доступной в PDB для Uniprot : P51681 (CC-рецептор хемокинового рецептора 5) в PDBE-KB .