Глюкоза
 Скелетная формула D -Glucose
| |
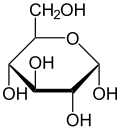 Haworth проекция α- D -гукопираноза
| |
 Фишер Проекция D -Glucose
| |
| Имена | |
|---|---|
| Произношение | / Ɡ l uː k oʊ z / , / ɡ l uː k oʊ s / |
| Имя IUPAC
Разрешены тривиальные названия: [ 1 ]
| |
| Предпочтительное имя IUPAC
Контакты не идентифицированы для натуральных продуктов. | |
Систематическое имя IUPAC
| |
| Другие имена
Сахар в крови
Декстроза Кукурузный сахар D -Glucose Виноградный сахар | |
| Идентификаторы | |
3D model ( JSmol )
|
|
| 3dmet | |
| Сокращения | GLC |
| 1281604 | |
| Чеби | |
| Химический | |
| Chemspider | |
| ЕС номер |
|
| 83256 | |
| Кегг | |
| Сетка | Глюкоза |
PubChem CID
|
|
| Rtecs номер |
|
| НЕКОТОРЫЙ |
|
| Характеристики | |
| C 6 H 12 O 6 | |
| Молярная масса | 180.156 g/mol |
| Появление | Белый порошок |
| Плотность | 1,54 г/см 3 |
| Точка плавления | а- D -Glucose: 146 ° C (295 ° F; 419 K) β- D -Glucose: 150 ° C (302 ° F; 423 K) |
| 909 г/л (25 ° C (77 ° F)) | |
| −101.5×10 −6 см 3 /мол | |
| 10.5674 | |
| Термохимия | |
Теплоемкость ( C )
|
218,6 J/(K · моль) [ 2 ] |
Std Molar
энтропия ( с ⦵ 298 ) |
209,2 J/(K · моль) [ 2 ] |
Энтальпия STD
образование (Δ f h ⦵ 298 ) |
-1271 кДж / раз [ 3 ] |
| 2805 кДж/моль (670 ккал/моль) | |
| Фармакология | |
| B05CX01 ( WHO ) V04CA02 ( WHO ), V06DC01 ( ВОЗ ) | |
| Опасности | |
| NFPA 704 (Огненная бриллиант) | |
| Лист данных безопасности (SDS) | ICSC 08655 |
За исключением случаев, когда отмечены, данные приведены для материалов в их стандартном состоянии (при 25 ° C [77 ° F], 100 кПа).
| |

Глюкоза - это сахар с молекулярной формулой C 6 H 12 O 6 . Глюкоза в целом является наиболее распространенным моносахаридом , [ 4 ] подкатегория углеводов . Глюкоза в основном производится растениями и большинством водорослей во время фотосинтеза из воды и углекислого газа, используя энергию от солнечного света. Глюкоза используется растениями для изготовления целлюлозы - наиболее распространенного углевода в мире - для использования в клеточных стенках и всех живых организмов для создания аденозинтрифосфата (АТФ), который используется клеткой в качестве энергии. [ 5 ] [ 6 ] [ 7 ]
В энергетическом метаболизме глюкоза является наиболее важным источником энергии во всех организмах . Глюкоза для метаболизма хранится как полимер , в растениях в основном как амилоза и амилопектин , а у животных в качестве гликогена . Глюкоза циркулирует в крови животных как сахара в крови . [ 5 ] [ 7 ] Природная форма глюкозы d -глюкоза, в то время как его стереоизомер L -глюкоза производится синтетически в сравнительно небольших количествах и менее биологически активна. [ 7 ] Глюкоза представляет собой моносахарид, содержащий шесть атомов углерода и альдегидную группу, и, следовательно, является альдогексозой . Молекула глюкозы может существовать в форме с открытой цепью (ациклической), а также в кольцевой (циклической) форме. Глюкоза встречается в природе и встречается в его свободном состоянии в фруктах и других частях растений. У животных глюкоза высвобождается от разрушения гликогена в процессе, известном как гликогенолиз .
Глюкоза, как внутривенный сахарный раствор , находится в списке основных лекарств Всемирной организации здравоохранения . [ 8 ] Это также в списке в сочетании с хлоридом натрия (табличная соль). [ 8 ]
Название глюкозы получено из древнегреческого глеков ( Glykýs вино, должно» от ' . ) « Sweeet' [ 9 ] [ 10 ] Суффикс -это химический классификатор, обозначающий сахар.
История
[ редактировать ]Глюкоза была сначала выделена из изюма в 1747 году немецким химиком Андреасом Маргграфом . [ 11 ] [ 12 ] Глюкоза была обнаружена в винограде другого немецкого химика - Иоганн Тобиас Лоудц - в 1792 году, и отличался как отличный от тростникового сахара ( сахароза ). Глюкоза - это термин, придуманный Джин Батист Дюма в 1838 году, который преобладал в химической литературе. Фридрих Август Кекуле предложил термин декстроза (из латинского декстера , что означает «справа»), потому что в водном растворе глюкозы плоскость линейно поляризованного света поворачивается вправо. Напротив, L-фруктоза (обычно называется как D -Fructose) (кетогексоза) и L -глюкоза ( L -Glucose) Поверните линейно поляризованный свет влево. Более ранние обозначения в соответствии с вращением плоскости линейно поляризованного света ( D и L -nomenclatore) были позже отброшены в пользу D - и L -Notation , которое относится к абсолютной конфигурации асимметричного центра самого дальнего от карбонильной группы и в соответствии с конфигурацией D - или L -Glyceraldehyde. [ 13 ] [ 14 ]
Поскольку глюкоза является основной необходимостью многих организмов, правильное понимание его химического макияжа и структуры в значительной степени способствовало общему прогрессу в органической химии . Это понимание произошло в значительной степени в результате исследований Эмиля Фишера , немецкого химика, получившего Нобелевскую премию 1902 года по химии за его выводы. [ 15 ] Синтез глюкозы установил структуру органического материала и, следовательно, образовал первую окончательную валидацию Якобуса Хенрикуса Ван-Т . теорий химической кинетики [ 16 ] Между 1891 и 1894 годами Фишер установил стереохимическую конфигурацию всех известных сахаров и правильно предсказал возможные изомеры , применяя теорию асимметричных углерода Ван -Т -Хоффа. Первоначально имена ссылались на естественные вещества. Их энантиомерам получило то же имя с введением систематических номенклатур с учетом абсолютной стереохимии (например, номенклатура Фишера, D / номенклатура ).
Для открытия метаболизма глюкозы Отто Мейерхоф получил Нобелевскую премию по физиологии или медицине в 1922 году. [ 17 ] Ханс фон Эйлер-Челпин был удостоен Нобелевской премии по химии вместе с Артуром Харденом в 1929 году за «Исследование ферментации сахара и их долю ферментов в этом процессе». [ 18 ] [ 19 ] В 1947 году Бернардо Houssay (за его открытие роли гипофиза в метаболизме глюкозы и полученных углеводов), а также Карл и Герти Кори (за их открытие обращения гликогена от глюкозы) получили Нобелевскую премию в Физиология или медицина. [ 20 ] [ 21 ] [ 22 ] В 1970 году Луис Лелуар был удостоен Нобелевской премии по химии за открытие нуклеотидов сахара, полученных из глюкозы, в биосинтезе углеводов. [ 23 ]
Химические и физические свойства
[ редактировать ]Глюкоза образует белые или бесцветные твердые тела, которые очень растворимы в воде и уксусной кислоте, но плохо растворимы в метаноле и этаноле . Они таяют при 146 ° C (295 ° F) ( α ) и 150 ° C (302 ° F) ( бета ), разлагаясь на 188 ° C (370 ° F) с высвобождением различных летучих продуктов, в конечном итоге оставив остаток углерод [ 24 ] Глюкоза имеет значение PKA 12,16 при 25 ° C (77 ° F) в воде. [ 25 ]
С шестью атомами углерода он классифицируется как гексоза , подкатегория моносахаридов . D -Glucose -один из шестнадцати альдогексозы стереоизомеров . А D - Изомер , D -глюкоза, также известная как декстроза, широко встречается в природе, но L -изомер, l -glucose , нет. Глюкоза может быть получена с помощью гидролиза углеводов, таких как молочный сахар ( лактоза ), тростник (сахароза), мальтоза , целлюлоза , гликоген и т. Д. Декстроза обычно коммерчески производится из крахмалов , таких как кукурузный крахмал в США и Японии, из картофеля, из картофеля, из кукурузного крахма. и пшеничный крахмал в Европе, а также от крахмала тапиоки в тропических районах. [ 26 ] Процесс производства использует гидролиз посредством паряка под давлением при контролируемом рН в струи, за которой следует дополнительная ферментативная деполимеризация. [ 27 ] Беспонированная глюкоза является одним из основных ингредиентов меда . [ 28 ] [ 29 ] [ 30 ] [ 31 ] [ 32 ]
Термин декстроза часто используется в клиническом (связанном с состоянием здоровья пациента) или в контексте питания (связанный с потреблением питания, такими как метки пищевых продуктов или рекомендации по пище молекулярные взаимодействия), [ 33 ] [ 34 ] [ 35 ] [ 36 ] Но оба термина относятся к одной и той же молекуле, в частности, D-глюкоза. [ 35 ] [ 37 ]
Моногидрат декстрозы является гидратированной формой D-глюкозы, что означает, что это молекула глюкозы с прикрепленной дополнительной молекулой воды. [ 38 ] Его химическая формула C 6 H 12 O 6 · H 2 o . [ 38 ] [ 39 ] Моногидрат декстрозы также называется гидратированной D-глюкозой и обычно производится из крахмалов. [ 38 ] [ 40 ] Моногидрат декстрозы используется в качестве преобладающего типа декстрозы в пищевых приложениях, таких как смеси напитков, - это распространенная форма глюкозы, широко используемой в качестве питания при производстве продуктов питания. Моногидрат декстрозы в основном потребляется в Северной Америке в качестве кукурузного сиропа или кукурузного сиропа с высоким содержанием фруктузы . [ 35 ]
Безводная декстроза , с другой стороны, представляет собой глюкозу, которая не имеет никаких молекул воды, прикрепленных к ней. [ 40 ] [ 41 ] Безводные химические вещества обычно продуцируются путем устранения воды из гидратированного вещества с помощью таких методов, как нагрев или высыхание (высыхание). [ 42 ] [ 43 ] [ 44 ] Моногидрат декстрозы может быть обезвожен до безводной декстрозы в промышленных условиях. [ 45 ] [ 46 ] Моногидрат декстрозы состоит из примерно 9,5% воды по массе; Благодаря процессу обезвоживания это содержание воды устраняется с учетом безводной (сухой) декстрозы. [ 40 ]
Безводная декстроза имеет химическую формулу C 6 H 12 O 6 , без прикрепленной молекулы воды, которая совпадает с глюкозой. [ 38 ] Безводная декстроза на открытом воздухе имеет тенденцию поглощать влагу и трансформироваться в моногидрат, и его более дороже для производства. [ 40 ] Безводная декстроза (безводная D-глюкоза) имеет повышенную стабильность и увеличение срока годности, [ 43 ] Имеет медицинское применение, например, в пероральном тесте на толерантность к глюкозе . [ 47 ]
Тогда как молекулярная масса (молярная масса) для моногидрата D-глюкозы составляет 198,17 г/моль, [ 48 ] [ 49 ] что для безводной D-глюкозы составляет 180,16 г/моль [ 50 ] [ 51 ] [ 52 ] Плотность этих двух форм глюкозы также отличается. [ указать ]
С точки зрения химической структуры, глюкоза является моносахаридом, то есть простым сахаром. Глюкоза содержит шесть атомов углерода и альдегидную группу и, следовательно, является альдогесозной . Молекула глюкозы может существовать в форме с открытой цепью присутствии спиртового и альдегида или кетоновых функциональных групп, форма, имеющая прямую цепь (ациклической), а также кольцевой (циклической) -в Кольцевая структура обычно встречается в углеводах. [ 53 ]
Структура и номенклатура
[ редактировать ]
Глюкоза присутствует в твердой форме в виде моногидрата с замкнутым пиранским кольцом (моногидрат α-глюкопиранозы, иногда точно известный декстрозой гидрат). В водном растворе, с другой стороны, это открытая цепь в небольшой степени и присутствует преимущественно в виде α- или β- пиранозы , которая взаимосвязана. Из водных растворов могут быть кристаллизованы три известные формы: α-глюкопираноза, β-глюкопираноза и моногидрат α-глюкопиранозы. [ 54 ] Глюкоза представляет собой строительный блок дисахаридов лактозы и сахарозы (тростника или свеклы), олигосахаридов , таких как рафиноза и полисахаридов , таких как крахмал , амилопектин , гликоген и целлюлоза . [ 7 ] [ 55 ] Температура стеклянного перехода глюкозы составляет 31 ° C (88 ° F), а константа Гордона - Тейлора (экспериментально определенная постоянная для прогнозирования температуры стекла для различных массовых фракций смеси двух веществ) [ 55 ] 4,5. [ 56 ]
| Формы и прогнозы D -Glucose по сравнению | ||
|---|---|---|
| Проекция NATTA | Haworth Projection | |

|
 а- D -глюкофураноза а- D -глюкофураноза
|
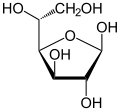 β- D -глюкофураноза β- D -глюкофураноза
|
 а- D -гукопираноза а- D -гукопираноза
|
 β- D -гукопираноза β- D -гукопираноза
| |
| а- D -глюкопираноза в (1) Tollens/Fischer (2) проекция Haworth (3) Конформация стула (4) Проекция мельницы | ||

| ||
Форма открытой цепи
[ редактировать ]
Форма с открытой цепью глюкозы составляет менее 0,02% молекул глюкозы в водном растворе в равновесии. [ 57 ] Остальное является одной из двух циклических гемиацетальных форм. В форме с открытой цепью молекула глюкозы имеет открытую (в отличие от циклической ) неразветвленной основы из шести атомов углерода, где C-1 является частью группы альдегида H (C = O) - . Следовательно, глюкоза также классифицируется как альдоза или альдогексоза . Группа альдегида делает глюкозу восстанавливающимся сахаром, давая положительную реакцию с тестом Fehling .
Циклические формы
[ редактировать ]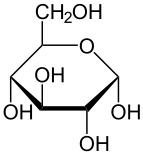
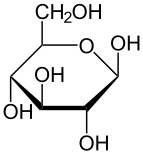
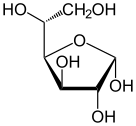
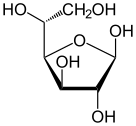
форма глюкозы (либо « D -», либо « L В растворах в равновесии существует . -») , каждая из которых содержит кольцо углеродов, закрытое одним атом кислорода Однако в водном растворе более 99% молекул глюкозы существуют в качестве пиранозных форм. Форма с открытой цепью ограничена около 0,25%, а формы фураноза существуют в незначительных количествах. Термины «глюкоза» и « d -глюкоза» обычно используются и для этих циклических форм. Кольцо возникает из формы с открытой цепью с помощью внутримолекулярной реакции нуклеофильного добавления между группой альдегида (AT C-1) и гидроксильной группой C-4 или C-5, образуя гемиацетальную связь, −c (OH) H -O- .
Реакция между C-1 и C-5 дает шестичленную гетероциклическую систему, называемую пиранозой, которая представляет собой моносахаридный сахар (следовательно, «-зо»), содержащий дериватизированный пиран- скелет. (Гораздо более редкая) реакция между C-1 и C-4 дает пятичленное фуранозное кольцо, названное в честь циклического эфирного фурана . В любом случае, каждый углерод в кольце имеет один водород и один гидроксил, за исключением последнего углерода (C-4 или C-5), где гидроксил заменяется оставшейся частью открытой молекулы (которая является - (C (Ch 2 OH) HOH) −H или - (Чо) −h соответственно).
Реакция закрытия кольца может дать два продукта, обозначенные «α-» и «β-». Когда молекула глюкопиранозы вводится в проекции Haworth , обозначение «α-» означает, что гидроксильная группа, прикрепленная к C-1 и Группа −ch 2 OH в C-5 лежит на противоположных сторонах плоскости кольца ( транс- расположение), в то время как «β-» означает, что они находятся на одной и той же стороне плоскости ( расположение CIS ). Следовательно, изомер D -глюкоза с открытой цепью дает четырех различных циклических изомеров: α -D -гукопираноза, β -D -гукопираноза, α -глюкофураноза и β -d -глюкофураноза. Эти пять структур существуют в равновесии и взаимосвязи, и взаимосвязь намного быстрее с кислотным катализом .
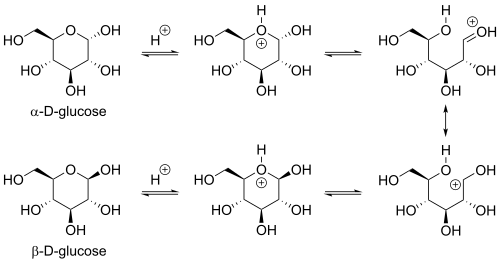

Другой изомер с открытой цепью L -глюкозой аналогично приводит к четырем различным циклическим формам L -глюкозы, каждый из которых зеркальное изображение соответствующей D -глюкозы.
Кольцо глюкопиранозы (α или β) может предполагать несколько непланарных форм, аналогичных конформациям циклогексана «стула» и «лодки» . Точно так же кольцо глюкофуранозы может принять несколько форм, аналогичных конформациям циклопентана «оболочки» .
В твердом состоянии наблюдаются только формы глюкопиранозы.
Некоторые производные глюкофуранозы, такие как 1,2 -O -изопропилиден -D -глюкофураноза, стабильны и могут быть получены чистыми в виде кристаллических твердых веществ. [ 58 ] [ 59 ] Например, реакция α-D-глюкозы с пара- толилбороновой кислотой H 3 C- (C 6 H 4 ) -B (OH) 2 реформирует нормальное пиранозное кольцо, чтобы получить 4-кратный эфир α-D-глюкофуранозы-1,2: 3,5-бис ( P- Талилбонат). [ 60 ]
Мутаротация
[ редактировать ]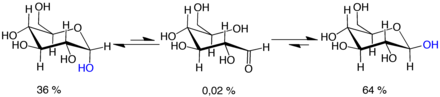
Мутаротация состоит из временного изменения реакции формирования кольца, что приводит к форме с открытой цепью с последующей реформированием кольца. Шаг закрытия кольца может использовать другой -OH Группа, чем тот, который воссоздан на стадии открытия (таким образом переключение между пиранозными и фуранозными формами), или новой гемиацетальной группы, созданной на C-1, может иметь такую же или противоположную руку, что и исходное (таким образом, переключение между α и β формы). Таким образом, хотя форма с открытой цепью едва обнаруживается в растворе, она является важным компонентом равновесия.
Форма с открытой цепью является термодинамически нестабильной , и она спонтанно изомеризуется циклическим формам. (Хотя реакция закрытия кольца может теоретически создавать четырех или трех атомных колец, они будут сильно натянуты и не наблюдаются на практике.) В растворах при комнатной температуре четыре циклические изомеры взаимосвязаны в течение временной шкалы часов, в процессе, называемом мутаротацией . [ 61 ] Начиная с любых пропорций, смесь сходится к стабильному соотношению α: β 36:64. Соотношение было бы α: β 11:89, если бы не влияние аномерного эффекта . [ 62 ] Мутаротация значительно медленнее при температурах, близких к 0 ° C (32 ° F).
Оптическая активность
[ редактировать ]Будь то в воде или в твердой форме, D -(+) -глюкоза является декстроротационной , а это означает, что она повернет направление поляризованного света по часовой стрелке, как видно, что смотрит на источник света. Эффект обусловлен хиральностью молекул и действительно изомером зеркального изображения, L -( -) -глюкоза, является леворотационной (вращается поляризованным светом против часовой стрелки) на то же количество. Сила эффекта различна для каждого из пяти таутомеров .
А D - Префикс не ссылается непосредственно к оптическим свойствам соединения. Это указывает на то, что хиральный центр C-5 имеет такую же руку, как и у D -Glyceraldehyde (который был так маркирован, потому что он декстроротатор). Тот факт, что D -глюкоза является декстроротационным, является комбинированным эффектом его четырех хиральных центров, а не только C -5; некоторые другие D -Ealdohexoses являются леворотационными.
Конверсия между двумя аномерами может наблюдаться в поляриметре , поскольку чистый α- D -Glucose имеет удельный угол вращения +112,2 ° мл/(дм · г), чистый β- d -глюкоза +17,5 ° мл/(дм · г). [ 63 ] Когда через определенное время достигнуто равновесие из -за мутаротации, угол вращения составляет +52,7 ° мл/(дм · г). [ 63 ] Добавляя кислоту или основание, это трансформация сильно ускоряется. Уравновешивание происходит с помощью формы альдегида с открытой цепью.
Изомеризация
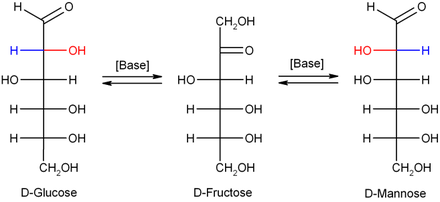
[ редактировать ]В разбавленных гидроксиде натрия или других разбавленных основаниях моносахариды манноза , глюкоза и фруктоза взаимодействуют (через преобразование Lobry de Bruyn -Alberda -Van Ekenstein ), так что формируется баланс между этими изомерами. Эта реакция проходит через энедиол :
Биохимические свойства
[ редактировать ]| Метаболизм общих моносахаридов и некоторых биохимических реакций глюкозы |
|---|

Глюкоза является наиболее распространенным моносахаридом. Глюкоза также является наиболее широко используемой альдогексозой в большинстве живых организмов. Одним из возможных объяснений этого является то, что глюкоза имеет более низкую тенденцию, чем другие альдогексозы, чтобы неспецифично реагировать на аминные группы белков . [ 64 ] Эта реакция - гликирование - переживание или разрушает функцию многих белков, [ 64 ] например, в гликированном гемоглобине . Низкая скорость гликации глюкозы может быть связана с тем, что она имеет более стабильную циклическую форму по сравнению с другими альдогексозами, что означает, что он тратит меньше времени, чем в своей реактивной форме с открытой цепью . [ 64 ] Причиной глюкозы, имеющей наиболее стабильную циклическую форму всех альдогексозов, является то, что ее гидрокси группы (за исключением гидрокси группы на аномерном углероде D -Glucose) находятся в экваториальном положении . Предположительно, глюкоза является наиболее распространенным природным моносахаридом, потому что она менее гликирован белками, чем другие моносахариды. [ 64 ] [ 65 ] Другая гипотеза заключается в том, что глюкоза, являющаяся единственной D -Ealdohexose, которая имеет все пять заместителей гидрокси в экваториальном положении в форме β- D -Glucose, более легко доступен для химических реакций, [ 66 ] : 194, 199 Например, для этерификации [ 67 ] : 363 или ацетальная формация. [ 68 ] По этой причине, D -Glucose также является очень предпочтительным строительным блоком в природных полисахаридах (гликаны). Полисахариды, которые составляются исключительно из глюкозы, называются глюканами .
Глюкоза производится растениями с помощью фотосинтеза с использованием солнечного света, [ 69 ] [ 70 ] Вода и углекислый газ и могут использоваться всеми живыми организмами в качестве источника энергии и углерода. Тем не менее, большая часть глюкозы происходит не в свободной форме, а в форме его полимеров, то есть лактоза, сахароза, крахмал и другие, которые являются энергетическими веществами, а также целлюлоза и хитин , которые являются компонентами клеточной стенки в растениях или грибах и членистоногие соответственно. Эти полимеры, когда они потребляются животными, грибами и бактериями, деградируют до глюкозы с использованием ферментов. Все животные также способны производить глюкозу от определенных предшественников по мере возникновения необходимости. Нейроны , клетки почечного мозгового мозга и эритроцитов зависят от глюкозы для их производства энергии. [ 70 ] У взрослых людей около 18 г (0,63 унции) глюкозы, [ 71 ] из которых около 4 г (0,14 унции) присутствует в крови. [ 72 ] Приблизительно 180–220 г (6,3–7,8 унции) глюкозы продуцируется в печени взрослого за 24 часа. [ 71 ]
Многие из долгосрочных осложнений диабета (например, слепоты , почечной недостаточности и периферической невропатии ), вероятно, связаны с гликированием белков или липидов . [ 73 ] Напротив, фермент -регулируемый добавление сахаров к белке называется гликозилированием и имеет важное значение для функции многих белков. [ 74 ]
Поглощение
[ редактировать ]Принтное глюкозу первоначально связывается с рецептором для сладкого вкуса на языке у людей. Этот комплекс белков T1R2 и T1R3 позволяет идентифицировать источники пищи, содержащие глюкозу. [ 75 ] [ 76 ] Глюкоза в основном поступает из еды - около 300 г (11 унций) в день производится путем преобразования пищи, [ 76 ] Но это также синтезируется от других метаболитов в клетках организма. У людей распад глюкозосодержащих полисахариды частично происходит во время жевания посредством амилазы , которая содержится в слюне , а также с малтазой , лактазой и саразой на границе кисти тонкой кишки . Глюкоза - это строительный блок многих углеводов и может быть отделена от них, используя определенные ферменты. Глюкозидазы , подгруппа гликозидаз, сначала катализируют гидролиз длинноцепочечных глюкозосодержащих полисахаридов, удаляя терминальную глюкозу. В свою очередь, дисахариды в основном деградируют специфическими гликозидазами до глюкозы. Названия деградирующих ферментов часто получены из конкретного поли- и дисахарида; В частности, для деградации полисахаридных цепей существуют амилазы (названные в честь амилозы, компонент крахмала), целлюлазы (названные в честь целлюлозы), хитиназы (названные в честь хитина) и многое другое. Кроме того, для расщепления дисахаридов существуют малтаза, лактаза, сахаза, Трегалаза и другие. У людей около 70 генов известно, что код для гликозидаз. Они имеют функции в расщеплении и деградации гликогена, сфинголипидов , мукополисахаридов и поли ( ADP-рибозы ). Люди не продуцируют целлюлазы, хитиназы или трегалазы, но бактерии в кишечной микробиоте .
Чтобы попасть в клеточные мембраны клеток и мембран клеточных компартментов, глюкоза требует специальных транспортных белков из основного суперсемейства . В тонкой кишке (точнее, в тощей степени ), [ 77 ] глюкоза поднимается в кишечный эпителий с помощью переносчиков глюкозы [ 78 ] Через вторичный механизм активного транспорта, называемый ионо-глюкозой симпорт на натрие/глюкозе котранспортер 1 (SGLT1). [ 79 ] Дальнейший перенос происходит на базолатеральной стороне кишечных эпителиальных клеток через глют2 глюкозы Glut2 , [ 79 ] Помимо поглощения клеток печени , клеток почек, клеток островков Лангерханса , нейронов , астроцитов и таницитов . [ 80 ] Глюкоза попадает в печень через портальную вену и хранится там как клеточный гликоген. [ 81 ] В клетках печени он фосфорилируется глюкокиназой в положении 6 с образованием 6-фосфата глюкозы , который не может покинуть клетку. Глюкоза 6-фосфатаза может преобразовать глюкозу 6-фосфат обратно в глюкозу исключительно в печени, поэтому организм может поддерживать достаточную концентрацию глюкозы в крови. В других клетках поглощение происходит пассивным транспортом через один из 14 белков Glut. [ 79 ] В других типах клеток фосфорилирование происходит через гексокиназу , после чего глюкоза больше не может диффундировать из клетки.
Glut1 глюкозы Glut1 продуцируется большинством типов клеток и имеет особое значение для нервных клеток и β-клеток поджелудочной железы . [ 79 ] GLUT3 высоко экспрессируется в нервных клетках. [ 79 ] Глюкоза из кровотока поглощена GLUT4 из мышечных клеток ( скелетных мышц [ 82 ] и сердечная мышца ) и жировые клетки . [ 83 ] GLUT14 выражается исключительно в яичках . [ 84 ] Избыточная глюкоза разбивается и превращается в жирные кислоты, которые хранятся в виде триглицеридов . В почках глюкоза в моче поглощается через SGLT1 и SGLT2 в апикальных клеточных мембранах и передается через GLUT2 в базолатеральных клеточных мембранах. [ 85 ] Около 90% реабсорбции глюкозы почек проходит через SGLT2 и около 3% через SGLT1. [ 86 ]
Биосинтез
[ редактировать ]У растений и некоторых прокариот глюкоза является продуктом фотосинтеза . [ 69 ] Глюкоза также образуется путем разрушения полимерных форм глюкозы, таких как гликоген (у животных и грибов ) или крахмала (у растений). Расщепление гликогена называется гликогенолизом, расщепление крахмала называется деградацией крахмала. [ 87 ]
Метаболический путь, который начинается с молекул, содержащих от двух до четырех атомов углерода (C) и заканчивается в молекуле глюкозы, содержащей шесть атомов углерода, называется глюконеогенез и встречается во всех живых организмах. Меньшие стартовые материалы являются результатом других метаболических путей. В конечном итоге почти все биомолекулы происходят из ассимиляции углекислого газа у растений и микробов во время фотосинтеза. [ 67 ] : 359 Свободная энергия образования α- D -Glucose составляет 917,2 килоджоульс на родинок. [ 67 ] : 59 У людей глюконеогенез происходит в печени и почках, [ 88 ] но и в других типах клеток. В печени около 150 г (5,3 унции) хранятся в скелетных мышцах около 250 г (8,8 унции). [ 89 ] Однако глюкоза, выделяемая в мышечных клетках при расщеплении гликогена, не может быть доставлена в циркуляцию, поскольку глюкоза фосфорилируется гексокиназой, а глюкозо-6-фосфатаза не экспрессируется для удаления фосфатной группы. В отличие от глюкозы, не существует транспортного белка для глюкозы-6-фосфата . Глюконеогенез позволяет организму создавать глюкозу из других метаболитов, включая лактат или определенные аминокислоты , одновременно потребляя энергию. Почечные трубчатые клетки также могут продуцировать глюкозу.
Глюкоза также можно найти вне живых организмов в окружающей среде. Концентрации глюкозы в атмосфере обнаруживаются с помощью сбора образцов самолетами и, как известно, различаются от местоположения к месту. Например, концентрации глюкозы в атмосферном воздухе из внутреннего Китая варьируются от 0,8 до 20,1 пг/л, тогда как концентрации глюкозы в восточном прибрежном прибрежном китайском языке варьируются от 10,3 до 142 пг/л. [ 90 ]
Деградация глюкозы
[ редактировать ]
У людей глюкоза метаболизируется гликолизом [ 91 ] и пентозофосфатный путь. [ 92 ] Гликолиз используется всеми живыми организмами, [ 66 ] : 551 [ 93 ] с небольшими вариациями, и все организмы генерируют энергию от разрушения моносахаридов. [ 93 ] В дальнейшем метаболизме он может быть полностью ухудшен за счет окислительного декарбоксилирования , цикла лимонной кислоты (синоним Krebs Cycle ) и дыхательной цепи до воды и диоксида углерода. Если для этого недостаточно кислорода, деградация глюкозы у животных происходит анаэробным к лактату посредством ферментации молочной кислоты и высвобождает гораздо меньшую энергию. Мышечный лактат попадает в печень через кровоток у млекопитающих, где происходит глюконеогенез ( цикл CORI ). При высоком запасе глюкозы ацетил-КоА метаболита из цикла Krebs также может использоваться для синтеза жирных кислот . [ 94 ] Глюкоза также используется для пополнения запасов гликогена организма, которые в основном содержатся в печени и скелетных мышцах. Эти процессы регулируются гормонально .
В других живых организмах могут возникнуть другие формы ферментации. Бактерия Escherichia coli может расти на питательной среде, содержащей глюкозу в качестве единственного источника углерода. [ 67 ] : 59 В некоторых бактериях и, в модифицированной форме, также в археи глюкоза разлагается через путь Эннер-Дадорофф . [ 95 ] При глюкозе был обнаружен механизм регуляции генов в E. coli , репрессии катаболита (ранее известная как эффект глюкозы ). [ 96 ]
Использование глюкозы в качестве источника энергии в клетках - это аэробное дыхание, анаэробное дыхание или ферментацию. [ 97 ] Первым этапом гликолиза является фосфорилирование глюкозы гексокиназой с образованием 6-фосфата глюкозы . Основная причина непосредственного фосфорилирования глюкозы заключается в предотвращении его диффузии из клетки, поскольку заряженная фосфатная глюкозы группа предотвращает легко пересекать клеточную мембрану . [ 97 ] Кроме того, добавление высокоэнергетической фосфатной группы активирует глюкозу для последующего разрушения на более поздних этапах гликолиза. [ 98 ]
В анаэробном дыхании одна молекула глюкозы производит чистое усиление двух молекул АТФ (четыре молекулы АТФ продуцируются во время гликолиза с помощью фосфорилирования на уровне субстрата, но два требуются ферментами, используемыми во время процесса). [ 99 ] При аэробном дыхании молекула глюкозы гораздо более прибыльной в том, что генерируется максимальная чистая продукция 30 или 32 молекул АТФ (в зависимости от организма). [ 100 ]
Нажмите на гены, белки и метаболиты ниже, чтобы связать соответствующие статьи. [ § 1 ]

- ^ Интерактивная карта пути может быть отредактирована в Wikipathways: "Glycolisisgluconeogenesis_WP534" .
Опухолевые клетки часто растут сравнительно быстро и потребляют увеличение глюкозы выше среднего гликолиза, [ 101 ] что приводит к образованию лактата, конечного продукта ферментации у млекопитающих, даже в присутствии кислорода. Это называется эффектом Варбурга . Для повышенного поглощения глюкозы в опухолях различные SGLT и GLUT чрезмерно продуцируются. [ 102 ] [ 103 ]
У дрожжей этанол ферментируется при высоких концентрациях глюкозы, даже в присутствии кислорода (что обычно приводит к дыханию, а не к ферментации). Это называется эффектом Crabtree .
Глюкоза также может ухудшаться, образуя диоксид углерода с помощью абиотических средств. Было продемонстрировано, что это происходит экспериментально посредством окисления и гидролиза при 22 ° C и pH 2,5. [ 104 ]
Источник энергии
[ редактировать ]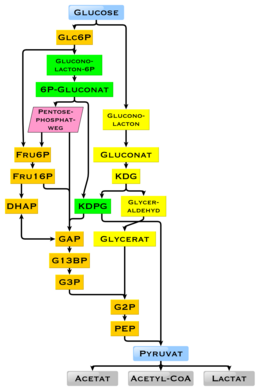
Глюкоза - это повсеместное топливо в биологии . Он используется в качестве источника энергии у организмов, от бактерий до людей, либо посредством аэробного дыхания , анаэробного дыхания (в бактериях) или ферментации . Глюкоза является ключевым источником энергии человеческого организма посредством аэробного дыхания, обеспечивающего около 3,75 килокалорий (16 килоджоул ) пищевой энергии на грамм. [ 105 ] Разрыв углеводов (например, крахмал) дает моно- и дисахариды , большая часть которых представляет собой глюкозу. Благодаря гликолизу , а затем в реакциях цикла лимонной кислоты и окислительного фосфорилирования глюкоза окисляется , чтобы в конечном итоге образовывать углекислый газ и воду, давая энергию, в основном в форме аденозинтрифосфата (АТФ). Реакция инсулина и другие механизмы регулируют концентрацию глюкозы в крови. Физиологическая калорийная ценность глюкозы, в зависимости от источника, составляет 16,2 килоджоульса на грамм [ 106 ] или 15,7 кДж/г (3,74 ккал/г). [ 107 ] Высокая доступность углеводов из биомассы растений привела к различным методам во время эволюции, особенно в микроорганизмах, для использования глюкозы для хранения энергии и углерода. Существуют различия, в которых конечный продукт больше не может использоваться для производства энергии. Присутствие отдельных генов и их генных продуктов, ферментов, определяют, какие реакции возможны. Метаболический путь гликолиза используется почти всеми живыми существами. Основным различием в использовании гликолиза является восстановление NADPH в качестве редуктаря для анаболизма , который в противном случае должен был бы быть создан косвенно. [ 108 ]
Глюкоза и кислород поставляют практически всю энергию для мозга , [ 109 ] Таким образом, его доступность влияет на психологические процессы. Когда глюкоза является низким , психологические процессы, требующие психических усилий (например, самоконтроль , применение решений), нарушаются. [ 110 ] [ 111 ] [ 112 ] [ 113 ] В мозге, который зависит от глюкозы и кислорода в качестве основного источника энергии, концентрация глюкозы обычно составляет от 4 до 6 мм (5 мм равна 90 мг/дл), [ 71 ] но уменьшается до 2-3 мм при голоже. [ 114 ] Путаница возникает ниже 1 мм, а кома на более низких уровнях. [ 114 ]
Глюкоза в крови называется сахаром в крови . Уровень сахара в крови регулируется глюкозо-связывающими нервными клетками в гипоталамусе . [ 115 ] Кроме того, глюкоза в мозге связывается с рецепторами глюкозы системы вознаграждения в прилегающем ядре . [ 115 ] Связывание глюкозы с сладким рецептором на языке вызывает высвобождение различных гормонов энергетического метаболизма, либо через глюкозу, либо через другие сахара, что приводит к повышению клеточного поглощения и снижению уровня сахара в крови. [ 116 ] Искусственные подсластители не снижают уровень сахара в крови. [ 116 ]
Содержание сахара в крови у здорового человека в коротком состоянии поста, например, после ночного поста, составляет от 70 до 100 мг/дл крови (от 4 до 5,5 мм). В плазме крови измеренные значения на 10–15% выше. Кроме того, значения в артериальной крови выше, чем концентрации в венозной крови, поскольку глюкоза поглощается в ткани во время прохождения капиллярного слоя . Также в капиллярной крови, которая часто используется для определения сахара в крови, значения иногда выше, чем в венозной крови. Содержание глюкозы в крови регулируется гормонным инсулином , инкрустином и глюкагоном . [ 115 ] [ 117 ] Инсулин понижает уровень глюкозы, глюкагон увеличивает его. [ 71 ] Кроме того, гормоны адреналин , тироксин , глюкокортикоиды , соматотропин и адренокортикотропин приводят к повышению уровня глюкозы. [ 71 ] Существует также гормон-независимая регуляция, которая называется ауторегуляцией глюкозы . [ 118 ] После потребления пищи концентрация сахара в крови увеличивается. Значения более 180 мг/дл в венозной цельной крови являются патологическими и называются гипергликемией , значения ниже 40 мг/дл называют гипогликемией . [ 119 ] Когда это необходимо, глюкоза высвобождается в кровоток глюкозо-6-фосфатазой из глюкозо-6-фосфата, происходящего из гликогена печени и почек, тем самым регулируя гомеостаз концентрации глюкозы в крови. [ 88 ] [ 70 ] У жвачных животных концентрация глюкозы в крови ниже (60 мг/дл у крупного рогатого скота и 40 мг/дл у овец ), потому что углеводы преобразуются больше с помощью кишечной микробиоты в жирные кислоты с короткой цепью . [ 120 ]
Некоторая глюкоза превращается в молочную кислоту астроцитами , которая затем используется в качестве источника энергии клетками мозга ; Некоторая глюкоза используется кишечными клетками и эритроцитами , в то время как остальные достигают печени , жировой ткани и мышечных клеток, где она поглощается и сохраняется как гликоген (под влиянием инсулина ). Гликоген клеток печени может быть преобразован в глюкозу и возвращен в кровь, когда инсулин низкий или отсутствует; Гликоген мышечных клеток не возвращается в кровь из -за отсутствия ферментов. В жировых клетках глюкоза используется для силовых реакций, которые синтезируют некоторые типы жиров и имеют другие цели. Гликоген - это механизм хранения энергии глюкозы, потому что он гораздо более «космический» и менее реактивный, чем сама глюкоза.
В результате его важности в здоровье человека глюкоза является аналитом в тестах на глюкозу , которые являются общими медицинскими анализом крови . [ 121 ] Еда или пост до получения образца крови влияет на анализ глюкозы в крови; Высокий уровень уровня сахара в крови натощак может быть признаком предстиата или сахарного диабета . [ 122 ]
Гликемический индекс является индикатором скорости резорбции и конверсии в уровни глюкозы в крови из проглощенных углеводов, измеренной как площадь под кривой уровней глюкозы в крови после потребления по сравнению с глюкозой (глюкоза определяется как 100). [ 123 ] Клиническая важность гликемического индекса является противоречивой, [ 123 ] [ 124 ] Поскольку продукты с высоким содержанием жира замедляют резорбцию углеводов и снижают гликемический индекс, например, мороженое. [ 124 ] Альтернативный индикатор - индекс инсулина , [ 125 ] измеряется как влияние потребления углеводов на уровень инсулина в крови. Гликемическая нагрузка является индикатором количества глюкозы, добавляемого к уровням глюкозы в крови после потребления, на основе гликемического индекса и количества потребляемой пищи.
Предшественник
[ редактировать ]Организмы используют глюкозу в качестве предшественника для синтеза нескольких важных веществ. Крахмал, целлюлоза и гликоген («животный крахмал») являются распространенными полимерами глюкозы (полисахариды). Некоторые из этих полимеров (крахмал или гликоген) служат запасами энергии, в то время как другие (целлюлоза и хитин , который изготовлен из производной глюкозы) играют структурные роли. Олигосахариды глюкозы в сочетании с другими сахарами служат важными запасами энергии. К ним относятся лактоза, преобладающий сахар в молоке, который представляет собой дисахарид глюкозы галактозы и сахароза, еще один дисахарид, который состоит из глюкозы и фруктозы. Глюкоза также добавляется в определенные белки и липиды в процессе, называемом гликозилированием . Это часто важно для их функционирования. Ферменты, которые соединяют глюкозу к другим молекулам, обычно используют фосфорилированную глюкозу для питания формирования новой связи путем сочетания ее с разрывом связей с фосфатом глюкозы.
Помимо прямого использования в качестве мономера, глюкоза может быть разбита, чтобы синтезировать широкий спектр других биомолекул. Это важно, поскольку глюкоза служит как основным запасом энергии, так и в качестве источника органического углерода. Глюкоза может быть разбита и преобразована в липиды. Это также является предшественником синтеза других важных молекул, таких как витамин С (аскорбиновая кислота). У живых организмов глюкоза превращается в несколько других химических соединений, которые являются начальным материалом для различных метаболических путей . Среди них все остальные моносахариды [ 126 ] такие как фруктоза (через путь полиола ), [ 79 ] Манноза (эпимер глюкозы в положении 2), галактоза (эпимер в положении 4), фукоза, различные уронические кислоты и аминохарные сахара производятся из глюкозы. [ 81 ] В дополнение к фосфорилированию в глюкозо-6-фосфат, который является частью гликолиза, глюкоза может окисляться во время ее деградации до глюконо-1,5-лактона . Глюкоза используется в некоторых бактериях в качестве строительного блока в трегалозе или биосинтез декстрана и у животных в качестве строительного блока гликогена. Глюкоза также может быть преобразована из бактериальной ксилозной изомеразы в фруктозу. Кроме того, метаболиты глюкозы продуцируют все несущественные аминокислоты, сахарные спирты , такие как маннитол и сорбит , жирные кислоты , холестерин и нуклеиновые кислоты . [ 126 ] Наконец, глюкоза используется в качестве строительного блока в гликозилировании белков к гликопротеинам , гликолипидам , пептидогликанам , гликозидам и другим веществам (катализируемым гликозилтрансферазами ) и может быть расщеплена из них гликозидазами .
Патология
[ редактировать ]Диабет
[ редактировать ]Диабет - это метаболическое расстройство, при котором организм не может регулировать уровни глюкозы в крови либо из -за отсутствия инсулина в организме, либо из -за неспособности к клеткам в организме, чтобы правильно реагировать на инсулин. Каждая из этих ситуаций может быть вызвана постоянно высоким уровнем уровня глюкозы в крови посредством выгорания поджелудочной железы и резистентности к инсулину . Поджелудочная железа является органом, ответственным за секрецию гормонного инсулина и глюкагона. [ 127 ] Инсулин - это гормон, который регулирует уровни глюкозы, позволяя клеткам организма поглощать и использовать глюкозу. Без этого глюкоза не может войти в ячейку и, следовательно, не может использоваться в качестве топлива для функций организма. [ 128 ] Если поджелудочная железа подвергается постоянному высоким уровням уровня глюкозы в крови, инсулино-продуцирующие клетки в поджелудочной железе могут быть повреждены, вызывая отсутствие инсулина в организме. Резистентность к инсулину возникает, когда поджелудочная железа пытается вырабатывать все больше и больше инсулина в ответ на постоянно повышенные уровни глюкозы в крови. В конце концов, остальная часть тела становится устойчивой к инсулину, который продуцирует поджелудочная железа, тем самым требуя большего инсулина для достижения того же эффекта снижения глюкозы в крови и вынуждения поджелудочной железы производить еще больше инсулина для конкурирования с сопротивлением. Эта негативная спираль способствует выгоранию поджелудочной железы и прогрессированию болезни диабета.
Чтобы отслеживать реакцию организма на терапию снижением уровня глюкозы в крови, можно измерить уровень глюкозы. Мониторинг глюкозы в крови может быть выполнен несколькими методами, такими как тест на глюкозу натощак, который измеряет уровень глюкозы в крови после 8 часов натощак. Другим тестом является 2-часовой тест на толерантность к глюкозе (GTT)-для этого теста у человека проходит тест на глюкозу натощак, а затем пьет 75-граммовый напиток глюкозы и проверяется. Этот тест измеряет способность тела человека обрабатывать глюкозу. Со временем уровни глюкозы в крови должны уменьшаться, поскольку инсулин позволяет его поднимать клетки и выходить из крови.
Управление гипогликемией
[ редактировать ]
Люди с диабетом или другими состояниями, которые приводят к низкому сахару в крови, часто несут небольшое количество сахара в различных формах. Одним из обычно используемых сахаров является глюкоза, часто в форме таблеток глюкозы (глюкоза, прижатая в форме таблетки, иногда с одним или несколькими другими ингредиентами в качестве связующего), твердой конфеты или пакета сахара .
Источники
[ редактировать ]
Большинство диетических углеводов содержат глюкозу, либо в качестве единственного строительного блока (как в крахмале полисахаридов и гликоген), либо вместе с другим моносахаридом (как в гетерополисахаридах сахарозы и лактозы). [ 129 ] Несвязанная глюкоза является одним из основных ингредиентов меда. Глюкоза чрезвычайно распространена и была изолирована из различных природных источников по всему миру, включая мужские конусы хвойного дерева Wollemia Nobilis в Риме, [ 130 ] Корни растений Ilex asprella в Китае, [ 131 ] и солома из риса в Калифорнии. [ 132 ]
| Еда элемент |
Углеводный общий, [ А ] включая диетическое волокно |
Общий сахар |
Бесплатно фруктоза |
Бесплатно глюкоза |
Сахароза | Соотношение фруктоза/ глюкоза |
Сахароза как доля общее количество сахаров (%) |
|---|---|---|---|---|---|---|---|
| Фрукты | |||||||
| Яблоко | 13.8 | 10.4 | 5.9 | 2.4 | 2.1 | 2.0 | 19.9 |
| Абрикос | 11.1 | 9.2 | 0.9 | 2.4 | 5.9 | 0.7 | 63.5 |
| Банан | 22.8 | 12.2 | 4.9 | 5.0 | 2.4 | 1.0 | 20.0 |
| Рис , высушен | 63.9 | 47.9 | 22.9 | 24.8 | 0.9 | 0.93 | 0.15 |
| Виноград | 18.1 | 15.5 | 8.1 | 7.2 | 0.2 | 1.1 | 1 |
| Пупок апельсин | 12.5 | 8.5 | 2.25 | 2.0 | 4.3 | 1.1 | 50.4 |
| Персик | 9.5 | 8.4 | 1.5 | 2.0 | 4.8 | 0.9 | 56.7 |
| Груша | 15.5 | 9.8 | 6.2 | 2.8 | 0.8 | 2.1 | 8.0 |
| Ананас | 13.1 | 9.9 | 2.1 | 1.7 | 6.0 | 1.1 | 60.8 |
| слива | 11.4 | 9.9 | 3.1 | 5.1 | 1.6 | 0.66 | 16.2 |
| Овощи | |||||||
| Свекла , красный | 9.6 | 6.8 | 0.1 | 0.1 | 6.5 | 1.0 | 96.2 |
| Морковь | 9.6 | 4.7 | 0.6 | 0.6 | 3.6 | 1.0 | 77 |
| Красный перец , сладкий | 6.0 | 4.2 | 2.3 | 1.9 | 0.0 | 1.2 | 0.0 |
| Лук , сладкий | 7.6 | 5.0 | 2.0 | 2.3 | 0.7 | 0.9 | 14.3 |
| Сладкий картофель | 20.1 | 4.2 | 0.7 | 1.0 | 2.5 | 0.9 | 60.3 |
| Элемент | 27.9 | 0.5 | Следы | Следы | Следы | — | Следы |
| Сахарный тростник | 13–18 | 0.2–1.0 | 0.2–1.0 | 11–16 | 1.0 | высокий | |
| Сахарная свекла | 17–18 | 0.1–0.5 | 0.1–0.5 | 16–17 | 1.0 | высокий | |
| Зерна | |||||||
| Кукуруза , сладкая | 19.0 | 6.2 | 1.9 | 3.4 | 0.9 | 0.61 | 15.0 |
- ^ Значение углеводов рассчитывается в базе данных USDA и не всегда соответствует сумме сахаров, крахмала и «диетического волокна».
Коммерческое производство
[ редактировать ]Глюкоза производится в промышленности из крахмала с помощью ферментативного гидролиза с использованием глюкозы амилазы или с помощью кислот . Ферментативный гидролиз в значительной степени смещенные кислотные реакции гидролиза, катализируемые кислотой. [ 134 ] Результатом является глюкозовый сироп (ферментативно с более чем 90% глюкозой в сухой веществе) [ 134 ] с ежегодным мировым объемом производства в 20 миллионов тонн (по состоянию на 2011 год). [ 135 ] Это является причиной бывшего общего названия «сахар крахмала». Амилазы чаще всего происходят из Bacillus licheniformis [ 136 ] или Bacillus subtilis (штамм MN-385), [ 136 ] которые являются более термостальными, чем первоначально используемые ферменты. [ 136 ] [ 137 ] Начиная с 1982 года, пуллуланазы из Aspergillus niger использовались для производства глюкозного сиропа для преобразования амилопектина в крахмал (амилозу), тем самым увеличивая выход глюкозы. [ 138 ] Реакция проводится при pH = 4,6–5,2 и температуре 55–60 ° C. [ 11 ] Кукурузный сироп имеет от 20% до 95% глюкозы в сухом веществе. [ 139 ] [ 140 ] Японская форма сиропа глюкозы, Mizuame , изготовлена из сладкого картофеля или рисового крахмала. [ 141 ] Мальтодекстрин содержит около 20% глюкозы.
Многие культуры можно использовать в качестве источника крахмала. Кукуруза , [ 134 ] рис, [ 134 ] пшеница , [ 134 ] Маниога , [ 134 ] картофель , [ 134 ] ячмень , [ 134 ] сладкий картофель, [ 142 ] Кукурузная шелуха и саго используются в различных частях света. В Соединенных Штатах кукурузный крахмал (от кукурузы) используется почти исключительно. Некоторая коммерческая глюкоза встречается в качестве компонента инвертированного сахара , примерно 1: 1 смесь глюкозы и фруктозы, которая производится из сахарозы. В принципе, целлюлоза может быть гидролизована до глюкозы, но этот процесс еще не является коммерчески практичным. [ 54 ]
Превращение в фруктозу
[ редактировать ]В США почти исключительно кукуруза (точнее, кукурузный сироп) используется в качестве источника глюкозы для производства изоглюкозы , которая представляет собой смесь глюкозы и фруктозы, поскольку фруктоза обладает более высокой силой - с такой же физиологической калорийностью 374 килокалорий за 100 г. Ежегодное мировое производство изоглукозы составляет 8 миллионов тонн (по состоянию на 2011 год). [ 135 ] При создании из кукурузного сиропа, конечным продуктом является кукурузный сироп с высоким содержанием фруктозы (HFC).
Коммерческое использование
[ редактировать ]
Глюкоза в основном используется для производства фруктозы и продуктов, содержащих глюкозу. В продуктах он используется в качестве подсластителя, увлажнителя , чтобы увеличить объем и создать более мягкий во рту . [ 134 ] Различные источники глюкозы, такие как виноградный сок (для вина) или солод (для пива), используются для ферментации в этанол во время производства алкогольных напитков . Большинство безалкогольных напитков в США используют HFCS-55 (с содержанием фруктозы 55% в сухой массе), в то время как в большинстве других подслащенных HFCS продуктов в США используют HFCS-42 (с содержанием фруктозы 42% в сухой массе ) [ 144 ] В Мексике, с другой стороны, безалкогольные напитки подслащены тростниковым сахаром, который обладает более высокой подслащенной силой. [ 145 ] Кроме того, используется глюкозный сироп, в частности, в производстве кондитерских изделий , таких как конфеты , ириска и помадка . [ 146 ] Типичными химическими реакциями глюкозы при нагревании в условиях без воды являются карамелизация и в присутствии аминокислот реакции Майяра .
Кроме того, различные органические кислоты могут быть биотехнологически продуцированы из глюкозы, например, путем ферментации с помощью Clostridium Thermoceticum для получения уксусной кислоты , с пенициллий Notatum для производства арабуаскорбиновой кислоты с делимаром Rhizopus для производства кислоты с aspergillus для для Rhizopus для производства кислоты Aspergillus для Rhizopus для производства кислоты с aspergillus для Rhizopus для производства кислоты с Aspergillus для Rhizopus для производства кислоты с Aspergillus для Rhizopus для производства кислоты с Aspergillus для Rhizopus для производства кислоты с Aspergillus для Rhizopus для производства кислоты с Aspergillus для Rhizopus для производства кислоты с Aspergillus для Производство кислоты с кандида глюконовой Aspergillus terreus для производства итаконовой кислоты , с псевдомонасом флуоресцентами для производства 2-кетоглюконовой кислоты , с глюконобактерными субоксиданами для производства 5-кетоглуконовой кислоты с Aspergillus oryzae для выработки коадзиновой кислоты с Lactobacillus delbreckii для производства для производства для производства для производства для производства молочная кислота с лактобациллусом Бревис для производства яблочной кислоты , с пропионибактер Шермани для производства пропионовой кислоты , с Pseudomonas aeruginosa для производства пировиновой кислоты и глюконобактерными субоксиданами для выработки татарной кислоты . [ 147 ] [ Дополнительные цитаты (ы) необходимы ] Мощные биологически активные натуральные продукты, такие как триптолид, которые ингибируют транскрипцию млекопитающих посредством ингибирования субъединицы XPB общего фактора транскрипции TFIIH, недавно сообщалось в качестве конъюгата глюкозы для нацеливания гипоксических раковых клеток с повышенной экспрессией транспортера глюкозы. [ 148 ] Недавно глюкоза получает коммерческое использование в качестве ключевого компонента «наборов», содержащих молочную кислоту и инсулин, предназначенные для индукции гипогликемии и гиперлактатемии для борьбы с различными раковыми заболеваниями и инфекциями. [ 149 ]
Анализ
[ редактировать ]Когда молекула глюкозы должна быть обнаружена в определенном положении в более крупной молекуле, ядерная магнитно-резонансная спектроскопия , рентгеновской кристаллографии анализ лектина или иммуноокрашивание выполняется с конъюгатом конканавалина , который связывает только глюкозу или маннозу.
Классические качественные реакции обнаружения
[ редактировать ]Эти реакции имеют только историческое значение:
Фехлинг тест
[ редактировать ]Тест Fehling - это классический метод обнаружения Aldoses. [ 150 ] Из-за мутаротации глюкоза всегда присутствует в небольшой степени в качестве альдегида с открытой цепью. Добавляя реагенты Fehling (раствор Fehling (i) и раствор Fehling (ii)), группа альдегида окисляется до карбоновой кислоты , в то время как Cu 2+ Тартратный комплекс уменьшается до Cu + и образует кирпичный красный осадок (Cu 2 O).
Сдав тест
[ редактировать ]В тесте Tollens после добавления аммиачного Agno 3 в раствор образца глюкоза уменьшает Ag + к элементарному серебра . [ 151 ]
Barfoed Test
[ редактировать ]В тесте Барфода , [ 152 ] Раствор растворенного медного ацетата , ацетата натрия и уксусной кислоты добавляется в раствор сахара, который должен быть протестирован и впоследствии нагревать в водяной бане в течение нескольких минут. Глюкоза и другие моносахариды быстро продуцируют красноватый цвет и красновато -коричневую меди (I) оксид (Cu 2 O).
Тест Ниландера
[ редактировать ]В качестве уменьшающегося сахара глюкоза реагирует в тесте Nylander . [ 153 ]
Другие тесты
[ редактировать ]После нагрева разбавленного раствора гидроксида калия с глюкозой до 100 ° C развивается сильное красновато-карамельное запах. [ 154 ] Концентрированная серная кислота растворяет сухую глюкозу без чинеления при комнатной температуре, образуя сахарную серу кислоту. [ 154 ] [ Проверка необходима ] В растворе дрожжей алкогольная ферментация продуцирует углекислый газ в соотношении 2,0454 молекул глюкозы к одной молекуле CO 2 . [ 154 ] Глюкоза образует черную массу с олово -хлоридом . [ 154 ] В растворе аммиака серебра глюкоза (а также лактоза и декстрин) приводит к осаждению серебра. В растворе аммиачного ацетата свинца белый гликозид свинца образуется в присутствии глюкозы, что становится менее растворимым при приготовлении пищи и становится коричневым. [ 154 ] В аммиачном растворе меди желтый гидрат оксида меди образуется с глюкозой при комнатной температуре, в то время как оксид красной меди образуется во время кипения (то же самое с декстрином, за исключением аммиачного раствора ацетата медиа -ацетата). [ 154 ] С реагентом Хагера глюкоза образует оксид ртуть во время кипения. [ 154 ] Руководство из щелочного висмута используется для осаждения элементарного, черно-коричневого висмута с глюкозой. [ 154 ] Глюкоза вареная в растворе молибдата аммония превращает раствор синим. Раствор с карбонатом индиго и карбонатом натрия ограничивается при кипячении с глюкозой. [ 154 ]
Инструментальная количественная оценка
[ редактировать ]Рефрактометрия и поляриметрия
[ редактировать ]В концентрированных растворах глюкозы с низкой доли других углеводов ее концентрация может быть определена с помощью поляриметра. Для сахарных смесей концентрация может быть определена с помощью рефрактометра , например, в определении Oechsle в ходе производства вина.
Фотометрические ферментативные методы в растворе
[ редактировать ]Фермент глюкозе оксидаза (GOX) превращает глюкозу в глюконовую кислоту и перекись водорода при потреблении кислорода. Другой фермент, пероксидаза, катализирует хромогенную реакцию (реакция Триндера) [ 155 ] фенола 4 - с аминоантипирином до фиолетового красителя. [ 156 ]
Фотометрический метод испытательной полосы
[ редактировать ]Метод испытательной полосы использует вышеупомянутое ферментативное преобразование глюкозы в глюконовую кислоту с образованием перекиси водорода. Реагенты иммобилизованы на полимерной матрице, так называемой испытательной полосе, которая предполагает более или менее интенсивный цвет. Это может быть измерено рефлекторнометрически при 510 нм с помощью портативного фотометра на основе светодиодов. Это обеспечивает рутинное определение сахара в крови со стороны не наук. В дополнение к реакции фенола с 4-аминоантипирином были разработаны новые хромогенные реакции, которые позволяют фотометрию на более высоких длинах волн (550 нм, 750 нм). [ 156 ] [ 157 ]
Амперометрический датчик глюкозы
[ редактировать ]Электроанализ глюкозы также основан на ферментативной реакции, упомянутой выше. Производимый перекись водорода может быть амперометрически количественно определена анодным окислением при потенциале 600 мВ. [ 158 ] Гокс иммобилизуется на поверхности электрода или в мембране, расположенной рядом с электродом. Драгоценные металлы, такие как платина или золото, используются в электродах, а также электроды углеродных нанотрубок, которые, например, легируют бором. [ 159 ] Нанопроволки Cu-Cuo также используются в качестве не содержащих ферментов амперометрических электродов, достигая предела обнаружения 50 мкмоль/л. [ 160 ] Особенно многообещающим методом является так называемая «проводка ферментов», где электрон, текущий во время окисления, переносится через молекулярную проволоку непосредственно от фермента в электрод. [ 161 ]
Другие сенсорные методы
[ редактировать ]Существует множество других химических датчиков для измерения глюкозы. [ 162 ] [ 163 ] Учитывая важность анализа глюкозы в науках о жизни, для сахаридов также были разработаны многочисленные оптические зонды на основе использования бориновых кислот, [ 164 ] которые особенно полезны для внутриклеточных сенсорных приложений, где другие (оптические) методы не являются или только условно используются. В дополнение к производным органической боиновой кислоты, которые часто тесно связаны с 1,2-диоль Полем Кроме того, были разработаны методы, которые косвенно обнаруживают концентрацию глюкозы посредством концентрации метаболизированных продуктов, например, по потреблению кислорода с использованием флуоресцентных-оптических датчиков. [ 165 ] Наконец, существуют концепции на основе ферментов, которые используют внутреннюю поглощение или флуоресценцию (меченных флуоресценцией) ферментами в качестве репортеров. [ 162 ]
Медная йодометрия
[ редактировать ]Глюкоза может быть количественно определено с помощью медной йодометрии. [ 166 ]
Хроматографические методы
[ редактировать ]В частности, для анализа сложных смесей, содержащих глюкозу, например, в меде, хроматографические методы, такие как высокоэффективная жидкая хроматография и газовая хроматография [ 166 ] часто используются в сочетании с масс -спектрометрией . [ 167 ] [ 168 ] Принимая во внимание соотношение изотопов, также возможно надежно обнаружить фальсификацию меда с помощью добавленных сахаров с помощью этих методов. [ 169 ] Дериватизация с использованием реагентов силилирования обычно используется. [ 170 ] Кроме того, пропорции ди- и трисахаридов могут быть определены количественно.
Анализ in vivo
[ редактировать ]Поглощение глюкозы в клетках организмов измеряется с помощью 2-дезокси-D-глюкозы или фтордезоксиглюкозы . [ 114 ] ( 18 F) Флородезоксиглюкоза используется в качестве индикатора в позитронной эмиссионной томографии в онкологии и неврологии, [ 171 ] где это, безусловно, наиболее часто используемый диагностический агент. [ 172 ]
Ссылки
[ редактировать ]- ^ Номенклатура углеводов (рекомендации 1996 г.) | 2-CARB-2 Архивировано 27 августа 2023 года на машине Wayback . iupac.qmul.ac.uk .
- ^ Jump up to: а беременный Boerio-Goates J (1991), «Измерения тепловой способности и термодинамические функции кристаллической α-D-глюкозы при температурах от 10K до 340K», J. Chem. Термодин. , 23 (5): 403–09, bibcode : 1991jchth..23..403b , doi : 10.1016/s0021-9614 (05) 80128-4
- ^ Пономарев В.В., Мигарскайя Л.Б. (1960), «Тепло сгорания некоторых аминокидов», Русс. J. Phys Химический (Engl. Перевод.) , 34 : 1182–83
- ^ Domb AJ, Kost J, Wiseman D (4 февраля 1998 г.). Справочник биоразлагаемых полимеров . CRC Press. п. 275. ISBN 978-1-4200-4936-7 .
- ^ Jump up to: а беременный «NCATS Inxight Nucriation - Dexstrose, неопределенная форма» . Архивировано из оригинала 11 декабря 2023 года . Получено 18 марта 2024 года .
- ^ Камид К (2005). Продукты целлюлозы и производные целлюлозы: молекулярная характеристика и ее применение (1 -е изд.). Амстердам: Elsevier. п. 1. ISBN 978-0-08-045444-3 Полем Получено 13 мая 2021 года .
- ^ Jump up to: а беременный в дюймовый "L-Glucose" . Биологические статьи, учебные пособия и словарь онлайн . 7 октября 2019 года. Архивировано с оригинала 25 мая 2022 года . Получено 6 мая 2022 года .
- ^ Jump up to: а беременный Всемирная организация здравоохранения (2019). Всемирная организация здравоохранения Список основных лекарств: 21 -й список 2019 . Женева: Всемирная организация здравоохранения. HDL : 10665/325771 . WHO/MVP/EMP/IAU/2019.06. Лицензия: CC BY-NC-SA 3.0 IGO.
- ^ «Онлайн этимологический словарь» . Etymonline.com . Архивировано с оригинала 26 ноября 2016 года . Получено 25 ноября 2016 года .
- ^ Lussac, Biot и Dumas (1838) . Thénard, Gay - Архивировано 6 декабря 2015 года в The Wayback Machine (отчет о мемуарах г -на Пелигиота под названием: Исследования о природе и химических свойствах сахара), отчеты , 7 : 106–113. Со страницы 109 . Архивировано 6 декабря 2015 года на машине Wayback : «Это происходит из -за сравнений, проведенных г -ном Пелиготом, этого винограда, сахара, крахмала, диабета и меда, имеют одинаковую композицию и одинаковые свойства, и составляют единственное тело, что и тела Мы предлагаем назвать глюкозу (1). , сделанных MR. Это следует из сравнений 1) ... (1) γλευχος, должен, Сладкое вино.
- ^ Jump up to: а беременный Энциклопедия пищи и здоровья . Академическая пресса. 2015. с. 239. ISBN 978-0-12-384953-3 Полем Архивировано из оригинала 23 февраля 2018 года.
- ^ Marggraf (1747) Химические переживания, созданные в целях привлечения настоящего сахара из различных растений, которые растут в наших регионах» июня 2016 « 24 В наших землях], История Королевской академии наук и Беллеса-Леттр. де Берлин , с. 79–90. С стр. 90: Архив 27 октября 2014 года в машине Wayback "изюм, навязываемый с небольшим количеством воды, их молита может быть забит, и сок, который выражает, подвергается подчиненному и утолщенному, обеспечит вид сахара. (Виноград, будучи маптиной с небольшим количеством воды, таким образом, что он может быть, может быть прессой, и сок, который выжимается, [после] очищенного и утолщенного, обеспечит своего рода сахар).)
- ^ Джон Ф. Робит: Основы углеводной химии. Springer Science & Business Media, 2012, 978-1-461-216222-3 . п. 7
- ^ Розанофф М.А. (1906). «О классификации стерео-изомеров Фишера.1» . Журнал Американского химического общества . 28 : 114–121. doi : 10.1021/ja01967a014 . Архивировано с оригинала 17 декабря 2019 года . Получено 1 июля 2019 года .
- ^ Эмиль Фишер , Нобелевский фонд, архивировав с оригинала 3 сентября 2009 года , извлечен 2 сентября 2009 г.
- ^ Фрейзер-Рейд B, "Van't Hoff's Glucose", Chem. Англ. Новости , 77 (39): 8
- ^ "Отто Мейерхоф - Факты - nobelprize.org" Аархивировано 15 июля 2018 года на машине Wayback . Nobelprize.org . Получено 5 сентября 2018 года.
- ^ "Ханс фон Эйлер -Челпин - Факты - Nobelprize.org" Аархивировано 3 сентября 2018 года на машине Wayback . Nobelprize.org . Получено 5 сентября 2018 года.
- ^ "Артур Харден - Факты - Нобелппция.org" Аархивировано 3 сентября 2018 года на машине Wayback . Nobelprize.org . Получено 5 сентября 2018 года.
- ^ "Bernardo Houssay - Факты - Nobelprize.org" Аархивировали 15 июля 2018 года на машине Wayback . Nobelprize.org . Получено 5 сентября 2018 года.
- ^ "Карл Кори - Факты - Nobelprize.org" Аархивировано 15 июля 2018 года на машине Wayback . Nobelprize.org . Получено 5 сентября 2018 года.
- ^ "Герти Кори - Факты - Nobelprize.org" Аархивировано 15 июля 2018 года на машине Wayback . Nobelprize.org . Получено 5 сентября 2018 года.
- ^ "Луис Лелуар - Факты - Nobelprize.org" Аархивировал 15 июля 2018 года на машине Wayback . Nobelprize.org . Получено 5 сентября 2018 года.
- ^ Wenyue Kang и Zhijun Zhang (2020): «Селективная продукция уксусной кислоты с помощью каталитического быстрого пиролиза гексозов над солями калия», катализаторы , том 10, страницы 502–515. Два : 10.3390/catal10050502
- ^ Bosch L, Fyles T, James T (ноябрь 2004 г.). «Бинарные и тройные комплексы фенилбороновой кислоты со сахаридами и основаниями Льюиса». Тетраэдр . 60 (49): 11175–11190. doi : 10.1016/j.tet.2004.08.046 .
- ^ Yebra-Burrun M (2005), «Подсластители», Энциклопедия аналитической науки , Elsevier, pp. 562–572, doi : 10.1016/b0-12-369397-7/00610-5 , ISBN 978-0-12-369397-6
- ^ "Глюкоза". Колумбия энциклопедия, 6 -е изд. 2015. Энциклопедия.com. ноября г. 2015 17
- ^ Ага М.Б., Шарма В., Дар А.Х., Дэш К.К., Сингх А., Шамс Р. и др. (2023). «Комплексный обзор функциональных и нутрицевтических свойств меда» . Эфуд . 4 (2). doi : 10.1002/efd2.71 .
- ^ Bobiş O, Dezmirean DS, Moise AR (2018). «Мед и диабет: важность натуральных простых сахаров в рационе для профилактики и лечения диабета различного типа» . Окислительная медицина и клеточная долговечность . 2018 : 1–12. doi : 10.1155/2018/4757893 . PMC 5817209 . PMID 29507651 .
- ^ Wani Ha, Majid S, Khan MS, Bhat AA, Wani RA, Bhat SA, et al. (2020). «Объем меда при диабете и метаболических расстройствах». Терапевтические применения меда и его фитохимических веществ . С. 195–217. doi : 10.1007/978-981-15-7305-7_9 . ISBN 978-981-15-7304-0 .
- ^ Alvarez-Surez JM, Somit S, Bertoli E, Battino M (2010). «Вклад меда в питание и здоровье человека: обзор». Средиземноморье 3 : 15–2 doi : 10.1007/ s12349-0051-005-6
- ^ Ишайек Джи, Керн М (август 2006 г.). «У нас жидники, различающиеся по содержанию глюкозы и фруктозы, вызывают сходные гликемические индексы». Журнал Американской диетической ассоциации . 106 (8): 1260–1262. doi : 10.1016/j.jada.2006.05.003 . PMID 16863724 .
- ^ «Потенциально важный вклад декстрозы, используемый в качестве разбавителя в гипергликемию у госпитализированных пациентов | Уход за диабетом | Американская диабетическая ассоциация» . Архивировано из оригинала 29 мая 2022 года . Получено 18 марта 2024 года .
- ^ "Декстроза: Почему это в еде и медицине?" Полем 24 июня 2018 года. Архивировано с оригинала 13 февраля 2024 года . Получено 18 марта 2024 года .
- ^ Jump up to: а беременный в «Что такое декстроза, как он используется, и это здорово? - инсайдер для питания» . 27 октября 2023 года. Архивировано из оригинала 14 февраля 2024 года . Получено 18 марта 2024 года .
- ^ "Декстроза против глюкозы: эти сахара равны?" Полем Архивировано с оригинала 29 сентября 2023 года . Получено 18 марта 2024 года .
- ^ Барон Д.Н., Макинтайр Н. (1976). «Письмо: глюкоза - это декстроза - глюкоза» . Британский медицинский журнал . 2 (6026): 41–42. doi : 10.1136/bmj.2.6026.41-c . PMC 1687736 . PMID 938892 .
- ^ Jump up to: а беременный в дюймовый «Prakash Chemicals International» . Архивировано из оригинала 6 июня 2023 года . Получено 18 марта 2024 года .
- ^ «API | Моногидрат глюкозы» . Архивировано из оригинала 24 марта 2023 года . Получено 18 марта 2024 года .
- ^ Jump up to: а беременный в дюймовый «Разница между безводом декстрозы и моногидратом декстрозы» . 28 декабря 2022 года. Архивировано с оригинала 18 марта 2024 года . Получено 18 марта 2024 года .
- ^ "Декстроза безводный" . Архивировано из оригинала 18 марта 2024 года . Получено 18 марта 2024 года .
- ^ Хворова Л.С., Андрев Н.Р., Лукин Н.Д. (январь 2020 г.). «Изучение условий для применения поверхностно -активных веществ при производстве кристаллической глюкозы». Российские сельскохозяйственные науки . 46 (1): 90–93. Bibcode : 2020ruags..46 ... 90K . doi : 10.3103/s1068367420010048 .
- ^ Jump up to: а беременный «В чем разница между безводной глюкозой и глюкозой» . Архивировано из оригинала 18 марта 2024 года . Получено 18 марта 2024 года .
- ^ "Безводная против моногидрата - в чем разница?" Полем Архивировано из оригинала 18 марта 2024 года . Получено 18 марта 2024 года .
- ^ Trasi NS, Boerrigter SX, Byrn SR, Carvajal TM (15 марта 2011 г.). «Исследование влияния условий обезвоживания на компактируемость глюкозы». Международный журнал фармацевтики . 406 (1–2): 55–61. doi : 10.1016/j.ijpharm.2010.12.042 . PMID 21232587 .
- ^ Mitra B, Wolfe C, Wu SJ (4 мая 2018 г.). «Декстроза моногидрат как альтернативный разбавитель, не являющийся животным, в составах с мокрым грануляцией с высоким сдвигом». Разработка лекарств и промышленная аптека . 44 (5): 817–828. doi : 10.1080/03639045.2017.1414231 . PMID 29300107 .
- ^ «Тесты диабета и предиабет - NIDDK» . Архивировано из оригинала 16 декабря 2023 года . Получено 18 марта 2024 года .
- ^ «Декстроза моногидрат» . Архивировано из оригинала 2 декабря 2023 года . Получено 18 марта 2024 года .
- ^ «D-(+)-моногидрат глюкозы» . Архивировано из оригинала 18 марта 2024 года . Получено 18 марта 2024 года .
- ^ "D-Glucose" . Архивировано из оригинала 15 декабря 2023 года . Получено 18 марта 2024 года .
- ^ "D-(+)-глюкоза" . Архивировано из оригинала 18 марта 2024 года . Получено 18 марта 2024 года .
- ^ "D-(+)-глюкоза" . Архивировано из оригинала 18 марта 2024 года . Получено 18 марта 2024 года .
- ^ «Глюкоза (декстроза)» . 2 октября 2013 года. Архивировано с оригинала 21 декабря 2023 года . Получено 18 марта 2024 года .
- ^ Jump up to: а беременный Schenck FW (2006). «Глюкоза и глюкоза, содержащие сиропы». Энциклопедия промышленной химии Уллмана . doi : 10.1002/14356007.a12_457.pub2 . ISBN 978-3-527-30673-2 .
- ^ Jump up to: а беременный Патрик Ф. Фокс: Advanced Dairy Chemistry Comemistry Том 3: Лактоза, вода, соли и витамины , Springer, 1992. Том 3, ISBN 9780412630200 . п. 316
- ^ Бенджамин Кабалеро, Пол Финглас, Фидель Толра: Энциклопедия пищи и здоровья . Академическая пресса (2016). ISBN 9780123849533 , том 1, P. 76
- ^ «16.4: Циклические структуры моносахаридов» . Химия Либретлекты . 18 июля 2014 года. Архивировано с оригинала 17 апреля 2023 года . Получено 17 апреля 2023 года .
- ^ Такаги С., Джеффри Г.А. (1979). «1,2-о-изопропилиден-D-глюкофураноза». Acta Crystallographica Раздел б . B35 (6): 1522–1525. Bibcode : 1979ccrb..35.1522t . doi : 10.1107/s0567740879006968 .
- ^ Bilecki M, Eggert H, Christian Norrild J (1999). «Флуоресцентный датчик глюкозы, связывающий ковалентно со всеми пятью гидрокси-группами α-D-глюкофуранозы. Реинвестирование». Журнал Химического общества, Перекин Транзакции . 2 (3): 449–456. doi : 10.1039/a808896i .
- ^ Чандран С.К., Нанья А. (2006). «Модулированная кристаллическая структура (z ′ = 2) α-D-глюкофуранозы-1,2: 3,5-бис (P-толил) борината». Crystengcomm . 8 (8): 581–585. doi : 10.1039/b608029d .
- ^ McMurry JE (1988), Органическая химия (2 -е изд.), Брукс/Коул, с. 866, ISBN 0534079687 .
- ^ Juaristi E, Cuevas G (1995), Аномерный эффект , CRC Press, стр. 9–10, ISBN 978-0-8493-8941-2
- ^ Jump up to: а беременный Манфред Хессе, Герберт Мейер, Бернд Зих, Стефан Биенц, Лоран Биглер, Томас Фокс: Спектроскопические методы в органической химии . 8 -е пересмотренное издание. Georg Thieme, 2011, ISBN 978-3-13-160038-7 , с. 34 (на немецком языке).
- ^ Jump up to: а беременный в дюймовый Bunn HF, Higgins PJ (1981). «Реакция моносахаридов с белками: возможная эволюционная значимость». Наука . 213 (4504): 222–24. Bibcode : 1981sci ... 213..222b . doi : 10.1126/science.12192669 . PMID 12192669 .
- ^ Джереми М. Берг: Страйер Биохимия. Springer-Lower, 2017, 978-3-662–54620-8 , с. 531. (Немецкий)
- ^ Jump up to: а беременный Гаррет Р.Х. (2013). Биохимия (5 -е изд.). Белмонт, Калифорния: Брукс/Коул, Cengage Learning. ISBN 978-1-133-10629-6 .
- ^ Jump up to: а беременный в дюймовый Voet D, Voet JG (2011). Биохимия (4 -е изд.). Хобокен, Нью -Джерси: Джон Вили и сыновья. ISBN 978-0-470-57095-1 .
- ^ Альберт Л. Лехнингер, Biochemistry, 6th Printing , Worth Publishers Inc. 1972, ISBN 0-87901-009-6 P. 228
- ^ Jump up to: а беременный «Химия для биологов: фотосинтез» . www.rsc.org . Архивировано с оригинала 4 августа 2016 года . Получено 5 февраля 2018 года .
- ^ Jump up to: а беременный в Питер С. Генрих: Биохимия и патобиохимия Лёффлер/Петридес. Springer-Verlag, 2014, 978-3-642-17972-3 , с. 195. (немецкий)
- ^ Jump up to: а беременный в дюймовый и U. Satyanarayana: Биохимия. Elsevier Health Sciences, 2014, 978-8-131-23713-7 . п. 674.
- ^ Вассерман Д.Х. (2009). «Четыре грамма глюкозы» . Американский журнал физиологии. Эндокринология и метаболизм . 296 (1): E11–21. doi : 10.1152/ajpendo.90563.2008 . PMC 2636990 . PMID 18840763 .
- ^ «Высокие осложнения глюкозы и диабета в крови: накопление молекул, известных как AGES, может быть ключевым звеном» , прогноз диабета , Американская диабетическая ассоциация, 2010, ISSN 0095-8301 , архивировано с первоначального 14 октября 2013 года , извлечен 20 мая 2010 г.
- ^ Варки А., Каммингс Р.Д., Эско Д.Д., Фриз Х.Х., Стэнли П., Бертоцци К.Р. и др. (2009). Варки А (ред.). Основы гликобиологии (2 -е изд.). Cold Spring Harbor Laboratories Press. ISBN 978-0-87969-770-9 Полем PMID 20301239 . Архивировано с оригинала 6 декабря 2016 года.
- ^ «Показывание соединения D -глюкозы (FDB012530) - FoodB» . Архивировано из оригинала 6 декабря 2022 года . Получено 18 марта 2024 года .
- ^ Jump up to: а беременный Питер С. Генрих: Биохимия и патобиохимия Лёффлер/Петридес. Springer-Verlag, 2014, 978-3-642-17972-3 , с. 404
- ^ Гарольд А. Харпер: Медицинская биохимия. Springer-Verlag, 2013, ISBN 978-3-662-22150-1 , с. 641. (на немецком языке)
- ^ Navale AM, Paranjape AN (2016). «Транспортеры глюкозы: физиологические и патологические роли» . Биофизические обзоры . 8 (1): 5–9. doi : 10.1007/s12551-015-0186-2 . PMC 5425736 . PMID 28510148 .
- ^ Jump up to: а беременный в дюймовый и фон Питер С. Генрих: Биохимия и патобиохимия Лёффлер/Петридес. Springer-Verlag, 2014, ISBN 978-3-642-17972-3 , с. 199, 200. (на немецком языке)
- ^ Thorens B (2015). «Glut2, глюкозо -зондирование и гомеостаз глюкозы» (PDF) . Диабетология . 58 (2): 221–32. doi : 10.1007/s00125-014-3451-1 . PMID 25421524 . Архивировано (PDF) из оригинала 2 декабря 2023 года . Получено 18 марта 2024 года .
- ^ Jump up to: а беременный Питер С. Генрих: Биохимия и патобиохимия Лёффлер/Петридес. Springer-Verlag, 2014, ISBN 978-3-642-17972-3 , с. 214. (на немецком языке)
- ^ Huang S, Czech MP (2007). «Glut4 Glucose Transsporter» . Клеточный метаболизм . 5 (4): 237–52. doi : 10.1016/j.cmet.2007.03.006 . PMID 17403369 .
- ^ Управления Р. (2014). Клеточная регуляция поглощения глюкозы с помощью переносчика глюкозы GLUT4 . Достижения в области клинической химии. Тол. 66. С. 173–240. doi : 10.1016/b978-0-12-801401-1.00006-2 . ISBN 978-0-12-801401-1 Полем PMID 25344989 .
- ^ Wu X, Freeze HH (декабрь 2002 г.). «GLUT14, дубликан GLUT3, специально экспрессируется в яичке как альтернативные формы сплайсинга». Геномика . 80 (6): 553–7. doi : 10.1006/geno.2002.7010 . PMID 12504846 .
- ^ Ghezzi C, Loo Ddf, Wright EM (2018). «Физиология обработки почечной глюкозы через SGLT1, SGLT2 и GLUT2» . Диабетология . 61 (10): 2087–2097. doi : 10.1007/s00125-018-4656-5 . PMC 6133168 . PMID 30132032 .
- ^ Поулсен С.Б., Фентон Р.А., Риг Т (2015). "Котранспорта на натрие-глюкозе" . Текущее мнение о нефрологии и гипертонии . 24 (5): 463–9. doi : 10.1097/mnh.0000000000000152 . PMC 5364028 . PMID 26125647 .
- ^ Смит, Элисон М., Зиман, Сэмюэль С., Смит, Стивен М. (2005). "Утолщение крахмала". Анну. Преподобный Плант Биол . 56 : 73–98. doi : 10.1146/annurev.arplant.56.032604.144257 . PMID 15862090 .
- ^ Jump up to: а беременный Leszek Szablewski: гомеостаз глюкозы и резистентность к инсулину. Bentham Science Publishers, 2011, 978-1-608-05189-2 , p. 46
- ^ Питер С. Генрих: Löffler/Petrides Biochemistry and Pathobiochemistry. Springer-Verlag, 2014, ISBN 978-3-642-17972-3 , с. 389. (на немецком языке)
- ^ Ван Г., Кавамура К., Хатакеяма С., Таками А., Ли Х, Ван В. (май 2007 г.). «Измерение самолетов органических аэрозолей над Китаем». Экологическая наука и технология . 41 (9): 3115–3120. Bibcode : 2007enst ... 41.3115w . doi : 10.1021/es062601h . PMID 17539513 .
- ^ Адева-Анани М.М., Перес-Фельпет Н., Фернандес-Фернандес С., Донапетрит-Гарсия С., Пазос-Гарсия С. (2016). «Метаболизм глюкозы в печени у людей» . Отчеты о биологии . 36 (6): E00416. Doi : 10.1042/bsr20160385 . PMC 5293555 . PMID 27707936 .
- ^ Х. Роберт Хортон, Лоуренс А. Моран, К. Грей Скримгур, Марк Д. Перри, Дж. Дэвид Роун: Биохимия . Пирсонские исследования; 4. Обновленное издание 2008; 978-3-8273-7312-0 ; п. 490–496. (Немецкий)
- ^ Jump up to: а беременный Брайан К. Холл: Эволюция Стрикбергера. Jones & Bartlett Publishers, 2013, 978-1-449–61484-3 , с. 164.
- ^ Джонс JG (2016). «Печеночная глюкоза и липидный метаболизм» . Диабетология . 59 (6): 1098–103. doi : 10.1007/s00125-016-3940-5 . PMID 27048250 .
- ^ Entner N, Doudoroff M (1952). «Окисление глюкозы и глюконовой кислоты Pseudomonas saccharophila» . J Biol Chem . 196 (2): 853–862. doi : 10.1016/s0021-9258 (19) 52415-2 . PMID 12981024 .
- ^ Ammar EM, Wang X, Rao CV (январь 2018 г.). «Регуляция метаболизма в Escherichia coli во время роста смесей не глюкозных сахаров: арабиноза, лактоза и ксилоза» . Научные отчеты . 8 (1): 609. Bibcode : 2018natsr ... 8..609a . doi : 10.1038/s41598-017-18704-0 . PMC 5766520 . PMID 29330542 .
- ^ Jump up to: а беременный Bonadonna RC, Bonora E, Del Prato S, Saccomani M, Cobelli C, Natali A, et al. (Июль 1996 г.). «Роли транспорта глюкозы и фосфорилирования глюкозы в мышечной инсулинорезистентности NIDDM» (PDF) . Диабет . 45 (7): 915–25. doi : 10.2337/diab.45.7.915 . PMID 8666143 . S2CID 219249555 . Архивировано (PDF) из оригинала 6 марта 2017 года . Получено 5 марта 2017 года .
- ^ "Глюкоза" . Архивировано из оригинала 5 декабря 2023 года . Получено 18 марта 2024 года .
- ^ Медицинская биохимия с первого взгляда @google Books , Blackwell Publishing, 2006, p. 52, ISBN 978-1-4051-1322-9 , архивировано с оригинала 23 февраля 2018 года
- ^ Медицинская биохимия с первого взгляда @google Books , Blackwell Publishing, 2006, p. 50, ISBN 978-1-4051-1322-9 , архивировано с оригинала 23 февраля 2018 года
- ^ Annibaldi A, Widmann C (2010). «Метаболизм глюкозы в раковых клетках». Современное мнение клинического питания и метаболической помощи . 13 (4): 466–70. doi : 10.1097/mco.0b013e32833a5577 . PMID 20473153 . S2CID 205782021 .
- ^ Szablewski L (2013). «Экспрессия переносчиков глюкозы при раке». Biochimica et Biophysica Acta (BBA) - Обзоры рака . 1835 (2): 164–9. doi : 10.1016/j.bbcan.2012.12.004 . PMID 23266512 .
- ^ Adekola K, Rosen St, Shanmugam M (2012). «Переносчики глюкозы в метаболизме рака» . Текущее мнение в онкологии . 24 (6): 650–4. doi : 10.1097/cco.0b013e328356da72 . PMC 6392426 . PMID 22913968 .
- ^ Schümann U, Gründler P (сентябрь 1998 г.). «Электрохимическая деградация органических веществ в анодах PBO2: мониторинг с непрерывными измерениями CO2». Водные исследования . 32 (9): 2835–2842. doi : 10.1016/s0043-1354 (98) 00046-3 .
- ^ «Глава 3: Расчет содержания энергии в пищевых продуктах - факторы преобразования энергии» , Пищевая энергия - Методы анализа и коэффициентов конверсии , FAO Food and Nutrition Paper 77, Рим: Организация по пищевой и сельскому хозяйству, 2003, ISBN 978-92-5-105014-9 , архивировано из оригинала 24 мая 2010 г.
- ^ Георг Шведт: сахарная сладкая химия. John Wiley & Sons, 2012, ISBN 978-3-527-66001-8 , с. 100 (на немецком языке) .
- ^ Шмидт, Ланг: Физиология человека , 30 -е издание. Springer Verlag, 2007, p. 907 (на немецком языке) .
- ^ Dandekar T, Schuster S, Snel B, Huynen M, Bork P (1999). «Выравнивание пути: применение к сравнительному анализу гликолитических ферментов» . Биохимический журнал . 343 (1): 115–124. doi : 10.1042/bj3430115 . PMC 1220531 . PMID 10493919 .
- ^ Dash P. «Кровавый мозг и метаболизм головного мозга (раздел 4, глава 11)» . Neuroscience Online: электронный учебник для нейронаук . Кафедра нейробиологии и анатомии - Медицинская школа Техасского университета в Хьюстоне. Архивировано с оригинала 17 ноября 2016 года.
- ^ Fairclough SH, Houston K (2004), «Метаболическая мера умственных усилий», Biol. Психол. , 66 (2): 177–190, doi : 10.1016/j.biopsycho.2003.10.001 , PMID 15041139 , S2CID 44500072
- ^ Gailliot MT, Baumeister RF, Dewall CN, Plant EA, Brewer LE, Schmeichel BJ, et al. (2007), «Самоконтроль полагается на глюкозу в качестве ограниченного источника энергии: сила воли-это больше, чем метафора» (PDF) , J. Pers. Соц Психол. , 92 (2): 325–336, Citeseerx 10.1.1.337.3766 , doi : 10.1037/0022-3514.92.2.325 , pmid 17279852 , s2cid 7496171 , архивированный (pdf) от оригинала 18 августа 2017
- ^ Gailliot MT, Baumeister RF (2007), «Физиология силы воли: связывание глюкозы в крови с самоконтролем», личная. Соц Психол. Rev. , 11 (4): 303–327, Citeseerx 10.1.1.475.9484 , doi : 10.1177/1088868307303030 , PMID 18453466 , S2CID 14380313
- ^ Masicampo EJ, Baumeister RF (2008), «На пути к физиологии рассуждений и суждений с двойным процессом: лимонада, силы воли и дорогостоящего анализа, основанного на правилах», Psychol. Наука , 19 (3): 255–60, doi : 10.1111/j.1467-9280.2008.02077.x , PMID 18315798 , S2CID 38596025
- ^ Jump up to: а беременный в Донард Дуайер: метаболизм глюкозы в мозге. Academic Press, 2002, ISBN 978-0-123-66852-3 , p. Xiii.
- ^ Jump up to: а беременный в Koekkoek LL, Mul JD, La Fleur SE (2017). «Оснащение глюкозы в системе вознаграждения» . Границы в нейробиологии . 11 : 716. doi : 10.3389/fnins.2017.00716 . PMC 5742113 . PMID 29311793 .
- ^ Jump up to: а беременный Такер Р.М., Тан Си (2017). «Влияют ли не питательные подсластители, влияют на острый гомеостаз глюкозы у людей? Систематический обзор» . Физиология и поведение . 182 : 17–26. doi : 10.1016/j.physbeh.2017.09.016 . PMID 28939430 . S2CID 38764657 . Архивировано из оригинала 29 июля 2020 года . Получено 7 июня 2020 года .
- ^ La Fleur SE, Fliers E, Kalsbeek A (2014). «Нейронаука гомеостаза глюкозы». Диабет и нервная система . Справочник по клинической неврологии. Тол. 126. С. 341–351. doi : 10.1016/b978-0-444-53480-4.00026-6 . ISBN 978-0-444-53480-4 Полем PMID 25410233 . Полем
- ^ Bisschop PH, Fliers E, Kalsbeek A (2015). «Автономная регуляция производства глюкозы в печени». Комплексная физиология . 5 (1): 147–165. doi : 10.1002/cphy.c140009 . PMID 25589267 .
- ^ Wa Scherbaum, BM Lobnig, In: Hans-Peter Wolff, Thomas R. Weihrauch: Hignist Therapy 2006, 2007 . 16 -е издание. Elsevier, 2006, ISBN 3-437-23182-0 , с. 927, 985 (на немецком языке) .
- ^ Гарольд А. Харпер: Медицинская биохимия . Springer-Verlag, 2013, 978-3-662-22150-1 , с. 294
- ^ Кларк С.Ф., Фостер -младший (2012). «История глюкозеров в анамнезе и их роль в самоконтроле сахарного диабета». Британский журнал биомедицинской науки . 69 (2): 83–93. Citeseerx 10.1.1.468.2196 . doi : 10.1080/09674845.2012.12002443 . PMID 22872934 . S2CID 34263228 .
- ^ «Диагностика диабета и изучение предиабета» . Американская диабетическая ассоциация . Архивировано из оригинала 28 июля 2017 года . Получено 20 февраля 2018 года .
- ^ Jump up to: а беременный Ричард А. Харви, Дениз Р. Ферер: Биохимия . 5 -е издание, Lippincott Williams & Wilkins, 2011, 978-1-608–31412-6 , с. 366
- ^ Jump up to: а беременный U Satyanarayana: Биохимия . Elsevier Health Sciences, 2014, 978-8-131-23713-7 , p. 508
- ^ Холт Ш.Х., Миллер Дж.С., Petocz P (1997). «Индекс инсулина в пищевых продуктах: потребность в инсулине, вызванная 1000-километровыми порциями общих продуктов» . Американский журнал клинического питания . 66 (5): 1264–1276. doi : 10.1093/ajcn/66.5.1264 . PMID 9356547 .
- ^ Jump up to: а беременный Питер С. Генрих: Биохимия и патобиохимия Лёффлер/Петридес. Springer-Verlag, 2014, ISBN 978-3-642-17972-3 , с. 27. (на немецком языке)
- ^ Röder PV, Wu B, Liu Y, Han W (2016). «Регуляция поджелудочной железы гомеостаза глюкозы» . Эксплуат Мол Медик 48 (3 марта): E219 -. doi : 10.1038/emm.2016.6 . PMC 4892884 . PMID 26964835 .
- ^ Estela, Carlos (2011) «Уровни глюкозы в крови», бакалавриат журнала математического моделирования: один + два: вып. 3: ISS. 2, статья 12.
- ^ «Углеводы и сахар в крови» . Источник питания . 5 августа 2013 года. Архивировано с оригинала 30 января 2017 года . Получено 30 января 2017 года - через Гарвардскую школу общественного здравоохранения.
- ^ Venditti A, Frezza C, Viencenti F, Brodella A, Scivobba F, Montesanno C, et al. (Февраль 2019 г.). «Производное синн-лабдадиен с редкой спиро-β-лактоном из мужских конусов Wolemia nobilis» Фитохимия 158 : 91–9 Bibcode : 2019pchem.158 ... 91V Doi : /j.phytochem.2018.11.0 10.1016 30481664PMID
- ^ Lei Y, Shi SP, Song YL, Bi D, Tu PF (май 2014). «Тритерпен Сапонины из корней Ilex Asprella». Химия и биоразнообразие . 11 (5): 767–775. doi : 10.1002/cbdv.201300155 . PMID 24827686 .
- ^ Balan V, Bals B, Chundawat SP, Marshall D, Dale BE (2009). «Предварительная обработка лигноцеллюлозной биомассы с использованием AFEX». Биотопливо . Методы в молекулярной биологии. Тол. 581. С. 61–77. doi : 10.1007/978-1-60761-214-8_5 . ISBN 978-1-60761-213-1 Полем PMID 19768616 .
- ^ "FoodData Central" . fdc.nal.usda.gov . Архивировано с оригинала 3 декабря 2019 года . Получено 18 марта 2024 года .
- ^ Jump up to: а беременный в дюймовый и фон глин час я PJ Fellows: Технология пищевой промышленности. Woodhead Publishing , 2016, 978-08-081-00523-1 , с. 197
- ^ Jump up to: а беременный Томас Беккер, Дитмар Брейтаупт, Хорст Вернер Доль, Армин Фихтер, Гюнтер Шлегель, Сакаю Шимидзу, Хидиаки Ямада: Биотехнология , в: Энциклопедия промышленной химии , 7th Edition, Wiley-VCH, 2011. ISBN 978-327-3294-4 . Том 6, с. 48
- ^ Jump up to: а беременный в Амилазное исследовательское общество Японии: Справочник по амилазам и связанных с ним ферментам. Elsevier, 2014, 978-1-483-29939-6 , с. 195.
- ^ Madsen GB, Norman BE, Slott S (1973). «Новая, тепловая стабильная бактериальная амилаза и ее использование в высокотемпературной разжижении». Крахмал - Стэрке . 25 (9): 304–308. doi : 10.1002/star.19730250906 .
- ^ Норман Бэй (1982). «Новый фермент обезболивания для применения в индустрии глюкозного сиропа». Крахмал - Стэрке . 34 (10): 340–346. doi : 10.1002/star.19820341005 .
- ^ Джеймс Н. Бемиллер, Рой Л. Уистлер (2009). Крахмал: химия и технология . Наука о продуктах питания и технике (3 -е изд.). Нью -Йорк: Академическая пресса. ISBN 978-0-08-092655-1 .
- ^ Bemiller, James N., Whistler, Roy L., eds. (2009). Крахмал: химия и технология . Наука о продуктах питания и технике (3 -е изд.). Нью -Йорк: Академическая пресса. ISBN 978-0-08-092655-1 Полем Получено 25 ноября 2016 года .
- ^ Алан Дэвидсон: Оксфордский компаньон To Food (1999). «Мизуам», с. 510 ISBN 0-19-211579-0 .
- ^ Алан Дэвидсон: Оксфордский компаньон до еды . OUP Oxford, 2014, 978-0-191-04072–6 , с. 527.
- ^ "Сахар" . Обучение, продовольственные ресурсы. Food.oregonstate.edu . Орегонский государственный университет , Корваллис, или. 23 мая 2012 года. Архивировано из оригинала 18 июля 2011 года . Получено 28 июня 2018 года .
- ^ «Кукурузный сироп с высоким содержанием фруктозы: вопросы и ответы» . Управление по контролю за продуктами и лекарствами США. 5 ноября 2014 года. Архивировано с оригинала 25 января 2018 года . Получено 18 декабря 2017 года .
- ^ Кевин Панг: Мексиканский кока -кола Хит в нас в: Seattle Times , 29 октября 2004 года.
- ^ Стив Т. Беккет: Промышленное шоколадное производство Беккета . John Wiley & Sons, 2017, 978-118-78014-5 , p. 82
- ^ Джеймс А. Кент: Руководство Ригеля по промышленной химии . Springer Science & Business Media, 2013, 978-1-475-76431-4 , с. 938
- ^ Datan E, Minn I, Peng X, He Ql, Ahn H, Yu B, et al. (2020). «Конъюгат глюкозы-триптолида избирательно нацелен на раковые клетки при гипоксии» . ISCAING . 23 (9): 101536. Bibcode : 202020SICI ... 23J1536D . doi : 10.1016/j.isci.2020.101536 . PMC 7509213 . PMID 33083765 .
- ^ Goodwin ML, Gladden LB, Nijsten MW (3 сентября 2020 г.). «Гипогликемия, защищенная лактатом (LPH)» . Границы в нейробиологии . 14 : 920. DOI : 10.3389/fnins.2020.00920 . ISSN 1662-453x . PMC 7497796 . PMID 33013305 .
- ^ H. Fehling: Количественное определение сахара в моче . В: Архив для физиологической медицины (1848), том 7, с. 64–73 (на немецком языке).
- ^ B. Tollens: О растворе с серебра Аммон-Алкали в качестве реагенов на архивировании альдегида 19 февраля 2022 года на машине Wayback . В сообщениях Германского химического общества (1882), том 15, с. 1635–1639 (на немецком языке).
- ^ Barfoed C (1873). «О инструкциях глюкозы рядом с декстрином и связанными телами» . Журнал аналитической химии (на немецком языке). 12 : 27–32. Doi : 10.1007/bf01462957 . S2CID 95749674 . Архивировано из оригинала 29 июля 2020 года . Получено 1 июля 2019 года .
- ^ Эмиль Ниландер: о щелочном растворе висмута как реагента на глюкозу в жестком журнале для физиологической химии . Том 8, выпуск 3, 1884, с. 175–185 Аннотация . Архивировано 23 сентября 2015 года на машине Wayback (на немецком языке).
- ^ Jump up to: а беременный в дюймовый и фон глин час я Георг Шведт: сахарная сладкая химия . John Wiley & Sons, 2012, ISBN 978-3-527-66001-8 , с. 102 (на немецком языке).
- ^ Триндер П. (1969). «Определение глюкозы в крови с использованием глюкозосидазы с альтернативным акцептором кислорода» . Анналы клинической биохимии . 6 : 24–27. doi : 10.1177/000456326900600108 . S2CID 58131350 .
- ^ Jump up to: а беременный Zhang Q, Zhao G, Yang N, Zhang L (2019). «Уровень глюкозы в крови натощак у пациентов с различными типами заболеваний». Гликаны и гликозаминогликаны как клинические биомаркеры и терапии - часть а . Прогресс в молекулярной биологии и трансляционной науке. Тол. 162. С. 277–292. doi : 10.1016/bs.pmbts.2019.01.004 . ISBN 978-0-12-817738-9 Полем PMID 30905457 .
- ^ Мизогучи М., Ишияма М., Шига М. (1998). «Водорастворимый хромогенный реагент для колориметрического обнаружения перекиси водорода-альтернатива 4-аминоантипирину, работающему на длинной длине волны». Аналитическая связь . 35 (2): 71–74. doi : 10.1039/a709038b .
- ^ Ван Дж. (2008). «Электрохимические биосенсоры глюкозы». Химические обзоры . 108 (2): 814–825. doi : 10.1021/cr068123a . PMID 18154363 . Полем
- ^ Chen X, Chen J, Deng C, Xiao C, Yang Y, Nie Z, et al. (2008). «Амперометрический биосенсор глюкозы на основе легированных бором углеродные нанотрубки, модифицированные электродом». Таланта . 76 (4): 763–767. doi : 10.1016/j.talanta.2008.04.023 . PMID 18656655 .
- ^ Wang G, Wei Y, Zhang W, Zhang X, Fang B, Wang L (2010). «Без ферментов амперометрическое восприятие глюкозы с использованием композитов нанопроволоков Cu-Cuo». Microchimica Acta . 168 (1–2): 87–92. doi : 10.1007/s00604-009-0260-1 . S2CID 98567636 .
- ^ Ohara TJ, Rajagopalan R, Heller A (1994). « Проводные» ферментные электроды для амперометрического определения глюкозы или лактата в присутствии мешающих веществ ». Аналитическая химия . 66 (15): 2451–2457. doi : 10.1021/ac00087a008 . PMID 8092486 .
- ^ Jump up to: а беременный Borisov SM, Wolfbeis OS (2008). «Оптические биосенсоры». Химические обзоры . 108 (2): 423–461. doi : 10.1021/cr068105t . PMID 18229952 .
- ^ Ferri S, Kojima K, Sode K (2011). «Обзор глюкозы оксидаз и глюкозойдегидрогеназы: вид птичьего глаза на глюкозу -чувствительные ферменты» . Журнал науки о диабете и технологии . 5 (5): 1068–76. doi : 10.1177/193229681100500507 . PMC 3208862 . PMID 22027299 .
- ^ Mader HS, Wolfbeis OS (2008). «Основанные на болоновой кислоте зонды для микроопределения сахаридов и гликозилированных биомолекул». Microchimica Acta . 162 (1–2): 1–34. doi : 10.1007/s00604-008-0947-8 . S2CID 96768832 .
- ^ Wolfbeis OS, Oehme I, Papkovskaya N, Klimant I (2000). «Биосенсоры глюкозы на основе Sol -Gel с использованием оптических преобразователей кислорода, и метод компенсации переменного кислородного фона». Биосенсоры и биоэлектроника . 15 (1–2): 69–76. doi : 10.1016/s0956-5663 (99) 00073-1 . PMID 10826645 .
- ^ Jump up to: а беременный Galant AL, Kaufman RC, Wilson JD (2015). «Глюкоза: обнаружение и анализ». Пищевая химия . 188 : 149–160. doi : 10.1016/j.foodchem.2015.04.071 . PMID 26041177 .
- ^ Sanz ML, Sanz J, Martínez-Castro I (2004). «Спектрометрический метод газовой хроматографической массы для качественного и количественного определения дисахаридов и трисахаридов в медах». Журнал хроматографии а . 1059 (1–2): 143–148. doi : 10.1016/j.chroma.2004.09.095 . PMID 15628134 .
- ^ Макс Планк Институт физиологии молекулярных растений в базе данных GOLM (19 июля 2007 г.). «Массовый спектр глюкозы» . GOLM METABOLOME DATABASE . Архивировано с оригинала 9 сентября 2018 года . Получено 4 июня 2018 года .
- ^ Cabañero AI, Recio JL, Rupérez M (2006). «Жидкая хроматография в сочетании с изотопной масс -спектрометрией: новая перспектива обнаружения фальсификации меда». J Agric Food Chem . 54 (26): 9719–9727. doi : 10.1021/jf062067x . PMID 17177492 .
- ^ Беккер М., Либнер Ф., Розенау Т., Поттхаст А (2013). «Подход к моно- и дисахаридным анализу и характеристике их параметров идентификации GC/MS». Таланта . 115 : 642–51. doi : 10.1016/j.talanta.2013.05.052 . PMID 24054643 .
- ^ Общество немецких химиков : Wayback = 20100331071121 Растения для позиционного документа « Архивированная нуклеархими» 31 марта 2010 года на машине Wayback , февраль 2000 года.
- ^ Maschauer S, Prante O (2014). «Подслащивание фармацевтической радиохимии по (18) F-Fluoroglycosylation: короткий обзор» . Biomed Research International . 2014 : 1–16. doi : 10.1155/2014/214748 . PMC 4058687 . PMID 24991541 .










Типы углеводов |
|---|
Сахар как продукт питания |
|---|