Калиевый канал

Калиевые каналы являются наиболее широко распределенным типом ионного канала, обнаруженного практически во всех организмах. [ 1 ] Они образуют калий -селективные поры , которые охватывают клеточные мембраны . Калиевые каналы обнаруживаются в большинстве типов клеток и контролируют широкий спектр функций клеток. [ 2 ] [ 3 ]
Функция
[ редактировать ]Калиевые каналы функционируют для проведения ионов калия вниз по электрохимическому градиенту , и оба быстро (вплоть до скорости диффузии k + ионы в объемной воде) и избирательно (исключая, в частности, натрий, несмотря на разницу в субсанге в ионном радиусе). [ 4 ] Биологически эти каналы действуют, чтобы установить или сбросить потенциал покоя во многих клетках. В возбудимых клетках, таких как нейроны , отсроченное противоположное значение ионам калия формирует потенциал действия .
Внося вклад в регуляцию продолжительности потенциала сердечного действия в сердечной мышцах , неисправность калия может вызвать опасные аритмии . Калиевые каналы также могут быть вовлечены в поддержание сосудистого тона .
Они также регулируют клеточные процессы, такие как секреция гормонов ( например , высвобождение инсулина из бета-клеток в поджелудочной железе ), поэтому их неисправность может привести к заболеваниям (таким как диабет ).
Некоторые токсины, такие как дендротоксин , являются мощными, потому что они блокируют калиевые каналы. [ 5 ]
Типы
[ редактировать ]Есть четыре основных класса калиевых каналов:
- Активированный кальцием калиевый канал - открыт в ответ на присутствие ионов кальция или других сигнальных молекул.
- Внутреннее выпрямление калиевого канала - проходит ток (положительный заряд) легче во внутреннем направлении (в ячейку).
- Канал калиевого домена тандемного домена - конститутивно открыт или обладает высокой базальной активацией, такой как «отдыхающие калиевые каналы» или «каналы утечки», которые устанавливают негативную мембранную потенциал нейронов.
- Управляемый напряжением калиевый канал - ионные ионные каналы, управляемые напряжением , которые открываются или близки в ответ на изменения трансмембранного напряжения .
Следующая таблица содержит сравнение основных классов каналов калия с репрезентативными примерами (для полного списка каналов в каждом классе см. В соответствующих страницах классов).
Для получения дополнительных примеров фармакологических модуляторов калиевых каналов см. Блокер -блокатор калия и открытие калия .
| Сорт | Подклассы | Функция | Блокировщики | Активаторы |
|---|---|---|---|---|
| Активируется кальцием 6 T & 1 P |
|
[ Цитация необходима ]
| ||
| Внутренне выпрямляющий 2 T & 1 P |
|
|
||
|
|
|||
|
|
|||
|
[ Цитация необходима ] | |||
| Тандемный домен 4 T & 2 P |
|
|
[ Цитация необходима ] | |
| Управляется напряжением 6 T & 1 P |
|
|
|
Структура
[ редактировать ]
Калиевые каналы имеют тетрамерную структуру, в которой четыре идентичные белковые субъединицы связываются с образованием четырехкратного симметричного ( C 4 ) комплекса, расположенного вокруг центрального иона, проводящего пор (то есть гомотетрамер). В качестве альтернативы четыре связанных, но не идентичных белковых субъединиц могут ассоциироваться с образованием гетеротетрамерных комплексов с псевдо C 4 симметрией. Все субъединицы канала калия имеют отличительную структуру пор, которая выстраивает верхнюю часть пор и отвечает за проницаемость селективной калиевой калиевой калие.
Существует более 80 млекопитающих генов канала калия , которые кодируют субъединицы . Однако калиевые каналы, обнаруженные в бактериях, являются одними из наиболее изученных из ионных каналов с точки зрения их молекулярной структуры. Использование рентгеновской кристаллографии , [ 55 ] [ 56 ] Глубокое понимание было получено в том, как ионы калия проходят через эти каналы и почему (меньшие) ионы натрия нет. [ 57 ] Нобелевская премия 2003 года за химию была присуждена Роду Маккиннону за его новаторскую работу в этой области. [ 58 ]
Селективный фильтр
[ редактировать ]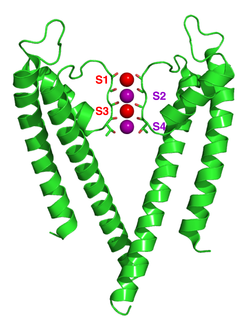
Ионные каналы калия удаляют гидратационную оболочку из иона, когда он входит в фильтр избирательности. Фильтр селективности сформируется последовательности из пяти остатков TVGYG, называется последовательности подписи, в каждой из четырех субъединиц. Эта последовательность подписи находится внутри петли между Pore Helix и TM2/6, исторически называемой P-петли. Эта последовательность подписи высоко консервативна, за исключением того, что валиновый остаток в прокариотических каналах калия часто заменяется изолейциновым остатком в эукариотических каналах. Эта последовательность принимает уникальную основную цепную структуру, структурно аналогичную структурному мотиву белка гнезда . Четыре набора электроотрицательных атомов карбонильного кислорода выровнены к центру пор фильтра и образуют квадратный антипризму, похожий на воду, оборудовающую оболочку вокруг каждого сайта связывания калия. Расстояние между карбонильными оксигенами и ионами калия в сайтах связывания фильтра селективности такое же, как и между водяными оксигенами в первой гидратационной оболочке и ионом калия в водном растворе, обеспечивая энергетически авансовый путь для дефекта. Сольвация ионов. Ионы натрия, однако, слишком малы, чтобы заполнять пространство между атомами карбонильного кислорода. Таким образом, для ионов натрия энергично благоприятно оставаться связанными молекулами воды во внеклеточном пространстве, а не проходить через ионную ионную пор калий. [ 60 ] Эта ширина, по -видимому, поддерживается водородной связью и силами Ван -дер -Ваальса в листе ароматических аминокислотных остатков, окружающих фильтр избирательности. [ 55 ] [ 61 ] Фильтр селективности открывается в сторону внеклеточного раствора, обнажая четыре карбонильные оксигены в остатках глицина (Gly79 в KCSA ). Следующим остатком в направлении внеклеточной стороны белка является отрицательно заряженный ASP80 (KCSA). Этот остаток вместе с пятью остатками фильтра образуют пор, которая соединяет заполненную водой полость в центре белка с внеклеточным раствором. [ 62 ]
Механизм избирательности
[ редактировать ]Механизм селективности канала калия остается под постоянными дебатами. Карбонильные оксигены сильно электроотрицательные и катионные. Фильтр может вместить ионы калия на 4 участка, обычно помеченные от S1 до S4, начинающимися с внеклеточной стороны. Кроме того, один ион может связываться в полости на участке, называемом SC или одним или несколькими ионами на внеклеточной стороне на более или менее четко определенных участках, называемых S0 или SEXT. Несколько различных занятий этих сайтов возможны. Поскольку рентгеновские структуры являются средними по многим молекулам, однако, невозможно вывести фактические занятия непосредственно из такой структуры. В целом, существует некоторый недостаток из -за электростатического отталкивания, чтобы иметь два соседних участка, занятых ионами. Предложения по механизму селективности были сделаны на основе моделирования молекулярной динамики , [ 63 ] игрушечные модели ионного переплета, [ 64 ] термодинамические расчеты, [ 65 ] топологические соображения, [ 66 ] [ 67 ] и структурные различия [ 68 ] между селективными и не селективными каналами.
Механизм транслокации ионов в KCSA широко изучался с помощью теоретических расчетов и моделирования. [ 62 ] [ 69 ] Прогнозирование механизма ионной проводимости, при котором два двойных занятых состояния (S1, S3) и (S2, S4) играют важную роль, был подтвержден обеими методами. Моделирование молекулярной динамики (MD) предполагает, что два внеклеточных состояния, S EXT и S 0 , отражающие ионы, входящие и покидающие фильтр, также являются важными субъектами в ионной проводимости.
Гидрофобная область
[ редактировать ]Этот регион нейтрализует окружающую среду вокруг иона калия, так что она не привлекает никаких обвинений. В свою очередь, это ускоряет реакцию.
Центральная полость
[ редактировать ]Центральная пор, шириной 10 Å, расположена рядом с центром трансмембранного канала, где энергетический барьер является самым высоким для обработки иона из -за гидрофобии стенки канала. Заполненная водой полость и полярный C-конец порра-спиралей ослабляют энергетический барьер для иона. Считается, что отталкивание предшествующим множественным ионам калия помогает пропускной способности ионов. Наличие полости может быть интуитивно понимать как один из механизмов канала для преодоления диэлектрического барьера или отталкивания низкой диэлектрической мембраной, сохраняя k + Ион в водянистой, высокой диэлектрической среде.
Регулирование
[ редактировать ]
Поток ионов через пору калия регулируется двумя родственными процессами, называемыми стробированием и инактивацией. Руководство - это открытие или закрытие канала в ответ на стимулы, в то время как инактивация - это быстрое прекращение тока из открытого калия и подавление способности канала возобновить проведение. В то время как оба процесса служат для регулирования проводимости канала, каждый процесс может быть опосредован рядом механизмов.
Как правило, считается, что стробирование опосредовано дополнительными структурными доменами, которые ощущают стимулы и, в свою очередь, открывают пор -канал. Эти домены включают домены RCK каналов BK, [ 70 ] [ 71 ] [ 72 ] и домены датчика напряжения напряжения, закрытого k + каналы. Считается, что эти домены реагируют на стимулы, физически открывая внутриклеточные ворота домена пор, что позволяет ионам калия пересекать мембрану. Некоторые каналы имеют несколько регуляторных доменов или вспомогательных белков, которые могут действовать для модуляции реакции на стимул. В то время как механизмы продолжают обсуждаться, существуют известные структуры ряда этих регуляторных доменов, включая домены RCK прокариотика [ 73 ] [ 74 ] [ 75 ] и эукариотический [ 70 ] [ 71 ] [ 72 ] каналы, ph gating domain kcsa, [ 76 ] Циклические домены стробирования нуклеотидов, [ 77 ] и напряжение, закрытые калиевые каналы. [ 78 ] [ 79 ]
Инактивация n-типа, как правило, является более быстрым механизмом инактивации и называется моделью «шарик и цепь» . [ 80 ] Инактивация n-типа включает взаимодействие N-конце канала или связанного белка, который взаимодействует с доменом пор и закупает путь ионной проводимости, как «мяч». В качестве альтернативы, как полагают, инактивация C-типа происходит в самом фильтре селективности, где структурные изменения в фильтре делают ее не проводящим. Существует ряд структурных моделей инактивированного C-типа C-типа + фильтры канала, [ 81 ] [ 82 ] [ 83 ] Хотя точный механизм остается неясным.
Фармакология
[ редактировать ]Блокировщики
[ редактировать ]Блокаторы калиевых каналов ингибируют поток ионов калия через канал. Они либо конкурируют с связыванием калия в фильтре селективности, либо связывают за пределами фильтра с ионной проводимостью. Примером одного из этих конкурентов являются четвертичные ионы аммония, которые связываются на внеклеточном лице [ 84 ] [ 85 ] или центральная полость канала. [ 86 ] Для блокировки из центральной полости ионов аммония в центральной полости также известны как блокаторы открытых каналов, поскольку связывание классически требует предыдущего открытия цитоплазматического затвора. [ 87 ]
Ионы бария также могут блокировать токи калиевого канала, [ 88 ] [ 89 ] Связываясь с высокой аффинностью в фильтре селективности. [ 90 ] [ 91 ] [ 92 ] [ 93 ] Считается, что это плотное связывание лежит в основе токсичности бария , ингибируя активность калия в возбудимых клетках.
С медицинской точки зрения блокаторы калия , такие как 4-аминопиридин и 3,4-диаминопиридин , были исследованы на предмет лечения таких состояний, как рассеянный склероз . [ 49 ] Выкл. Заключенные лекарственные эффекты могут привести к индуцированному препаратом синдрома QT , потенциально опасного для жизни состояния. Чаще всего это происходит из -за действия на канале калия Herg в сердце. Соответственно, все новые препараты протестированы на безопасность сердца.
Активаторы
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к этому . ( Май 2019 ) |
Мускариновый калиевый канал
[ редактировать ]
Некоторые типы калийных каналов активируются мускариновыми рецепторами , и они называются мускариновыми калиевыми каналами (i kach ). Эти каналы представляют собой гетеротетрамер, состоящий из двух субъединиц Girk1 и двух субъединиц Girk4 . [ 94 ] [ 95 ] Примерами являются калиевые каналы в сердце, которые, когда активируются парасимпатическими сигналами через мускариновые рецепторы M2 , вызывают внешний ток калия, который замедляет частоту сердечных сокращений . [ 96 ] [ 97 ]
В изобразительном искусстве
[ редактировать ]Родерик Маккиннон заказал рождение идеи , 5-футовой (1,5 м) скульптуры на основе канала калия KCSA. [ 98 ] Работа содержит проволочный объект, представляющий внутреннюю часть канала с выдувным стеклянным объектом, представляющим основную полость структуры канала.
Смотрите также
[ редактировать ]- Кальциевый канал - комплекс ионо -канала, через который проходят ионы кальция
- Внутренний ионный канал калия-калия -группа трансмембранных белков, которые пассивно переносят
- Калий в биологии - использование калия организмами
- Семейство переносчика калия (TRK) - семейство транспортных белков
- Поглощение калия Permease - Семейные
- Ионовый канал натрия - трансмембранное белок, позволяющий ионам натрия на страницах и выходе,
Ссылки
[ редактировать ]- ^ Littleton JT, Ganetzky B (апрель 2000 г.). «Ионные каналы и синаптическая организация: анализ генома Drosophila» . Нейрон . 26 (1): 35–43. doi : 10.1016/s0896-6273 (00) 81135-6 . PMID 10798390 . S2CID 5694563 .
- ^ Hille, Bertil (2001). «Глава 5: Калиевые каналы и хлоридные каналы». Ионные каналы возбудимых мембран . Сандерленд, Месса: Синауэр. С. 131–168. ISBN 978-0-87893-321-1 .
- ^ Джесселл Т.М., Кандель Э.Р. , Шварц Дж.Х. (2000). «Глава 6: Ионные каналы». Принципы нейронной науки (4 -е изд.). Нью-Йорк: МакГроу-Хилл. С. 105–124 . ISBN 978-0-8385-7701-1 .
- ^ Lim C, Dudev T (2016). Sigel A, Sigel H, Sigel RK (Eds.). «Глава 10. Селективность калия в зависимости от селективности натрия в фильтрах моновалентных ионных каналов». Ионы щелочных металлов: их роль в жизни . Металлические ионы в науках о жизни. 16 Springer: 325–347. doi : 10.1007/978-3-319-21756-7_9 . ISBN 978-3-319-21755-0 Полем PMID 26860305 .
- ^ косвенно цитируется из справочного числа 3,4,5,6 в Rehm H, Lazdunski M (июль 1988 г.). «Очистка и субъединица структуры предполагаемого K+-канального белка, идентифицированного его связывающими свойствами для дендротоксина I» . Труды Национальной академии наук Соединенных Штатов Америки . 85 (13): 4919–4923. Bibcode : 1988pnas ... 85.4919r . doi : 10.1073/pnas.85.13.4919 . PMC 280549 . PMID 2455300 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не Rang, HP (2015). Фармакология (8 изд.). Эдинбург: Черчилль Ливингстон. п. 59. ISBN 978-0-443-07145-4 .
- ^ Томпсон Дж, Бегенисич Т (май 2000). «Электростатическое взаимодействие между харибдотоксином и тетрамерным мутантом шейкеров K (+) каналов» . Биофизический журнал . 78 (5): 2382–2391. Bibcode : 2000bpj .... 78.2382t . doi : 10.1016/s0006-3495 (00) 76782-8 . PMC 1300827 . PMID 10777734 .
- ^ Наранджо Д., Миллер С (январь 1996 г.). «Сильно взаимодействующая пара остатков на контактной поверхности харибдотоксина и шейкера K+ -канала» . Нейрон . 16 (1): 123–130. doi : 10.1016/s0896-6273 (00) 80029-x . PMID 8562075 . S2CID 16794677 .
- ^ Yu M, Liu SL, Sun PB, Pan H, Tian CL, Zhang LH (январь 2016 г.). «Пептидные токсины и небольшие молекулярные блокаторы каналов BK» . Acta Pharmacologica Sinica . 37 (1): 56–66. doi : 10.1038/aps.2015.139 . PMC 4722972 . PMID 26725735 .
- ^ Candia S, Garcia ML, Latorre R (август 1992 г.). «Режим действия ибератоксина, мощный блокатор большой проводимости Ca (2+)-активированный K+-канал» . Биофизический журнал . 63 (2): 583–590. Bibcode : 1992bpj .... 63..583c . doi : 10.1016/s0006-3495 (92) 81630-2 . PMC 1262182 . PMID 1384740 .
- ^ Стокер М., Краузе М, Педарзани П (апрель 1999 г.). «Апамин чувствительный к Ca2+ -активированному K+ ток в гиппокампальных пирамидальных нейронах» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (8): 4662–4667. Bibcode : 1999pnas ... 96.4662s . doi : 10.1073/pnas.96.8.4662 . PMC 16389 . PMID 10200319 .
- ^ McLeod JF, Leempoels JM, Peng SX, Dax SL, Myers LJ, Golder FJ (ноябрь 2014 г.). «GAL-021, новый внутривенный блокатор BKCA-канала, хорошо переносится и стимулирует вентиляцию у здоровых добровольцев» . Британский журнал анестезии . 113 (5): 875–883. doi : 10.1093/bja/aeu182 . PMID 24989775 .
- ^ Dopico AM, Bukiya AN, Kuntamallappanavar G, Liu J (2016). «Модуляция каналов BK этанолом» . Международный обзор нейробиологии . 128 : 239–279. doi : 10.1016/bs.irn.2016.03.019 . ISBN 9780128036198 Полем PMC 5257281 . PMID 27238266 .
- ^ Jump up to: а беременный Patnaik, Pradyot (2003). Справочник неорганических химикатов . МакГроу-Хилл. С. 77–78 . ISBN 978-0-07-049439-8 .
- ^ Sackin H, Syn S, Palmer LG, Choe H, Walters de (февраль 2001 г.). «Регуляция РОМК внеклеточными катионами» . Биофизический журнал . 80 (2): 683–697. Bibcode : 2001bpj .... 80..683s . doi : 10.1016/s0006-3495 (01) 76048-1 . PMC 1301267 . PMID 11159436 .
- ^ Dhamoon AS, Jalife J (март 2005 г.). «Внутренний ток выпрямителя (IK1) контролирует возбудимость сердца и участвует в аритмогенезе». Сердечный ритм . 2 (3): 316–324. doi : 10.1016/j.hrthm.2004.11.012 . PMID 15851327 .
- ^ Jump up to: а беременный в дюймовый и фон Swale DR, Kharade SV, Denton JS (апрель 2014 г.). «Фармакология сердечного и почечного выпрямителя калиевого канала: новые инструменты для интегративной физиологии и терапии» . Современное мнение о фармакологии . 15 : 7–15. doi : 10.1016/j.coph.2013.11.002 . PMC 4097192 . PMID 24721648 .
- ^ Xynogalos P, Seyler C, Scherer D, Koepple C, Scholz EP, Thomas D, et al. (Декабрь 2014). «Антиаритмический препарат класса III класса Дроедарон ингибирует сердечную внутреннюю выпрямление каналов KIR2.1 посредством связывания в остатках E224». Архив фармакологии Наунин-Шмидеберга . 387 (12): 1153–1161. doi : 10.1007/s00210-014-1045-6 . PMID 25182566 . S2CID 10575229 .
- ^ Koepple C, Scherer D, Seyler C, Scholz E, Thomas D, Katus HA, Zitron E (май 2017 г.). «Двойной механизм ингибирования внутреннего выпрямления каналов KIR2.x хинидином, включающим прямой блок пор и PIP 2 -Interference» . Журнал фармакологии и экспериментальной терапии . 361 (2): 209–218. doi : 10.1124/jpet.116.238287 . PMID 28188270 . S2CID 206502631 .
- ^ Caballero R, Dolz-Gaitón P, Gómez R, Amorós I, Barana A, González de La Fuente M, et al. (Август 2010 г.). «Flecainide увеличивает токи Kir2.1, взаимодействуя с цистеином 311, уменьшая выпрямление, вызванное полиамином» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (35): 15631–15636. BIBCODE : 2010PNAS..10715631C . doi : 10.1073/pnas.1004021107 . PMC 2932566 . PMID 20713726 .
- ^ Kobayashi T, Washiyama K, Ikeda K (март 2006 г.). «Ингибирование G-белка, активируемого внутренним выпрямляющим K+-каналами с помощью IfenProdil» . Нейропсихофармакология . 31 (3): 516–524. doi : 10.1038/sj.npp.1300844 . PMID 16123769 .
- ^ Soeda F, Fujieda Y, Kinoshita M, Shirasaki T, Takahama K (май 2016 г.). «Центрально действующие ненаркотические противоопубликанцы предотвращают гиперактивность у мышей: вовлечение каналов Girk». Фармакология, биохимия и поведение . 144 : 26–32. doi : 10.1016/j.pbb.2016.02.006 . PMID 26892760 . S2CID 30118634 .
- ^ Yamoto G, Soeda F, Shirasaki T, Takahama K (апрель 2011 г.). «[Дорожая ли мишень для девочек, который является целевым в разработке нового терапевтического препарата нарушения мочи? ] Якугаку Засши 131 (4): 523–5 Doi : 10.1248/ srople.131.5 PMID 214677791
- ^ Kawaura K, Honda S, Soeda F, Shirasaki T, Takahama K (май 2010 г.). «[Новое антидепрессант-подобное действие препаратов, обладающих действием блокировки канала Girk у крыс]» . Якугаку Засши . 130 (5): 699–705. doi : 10.1248/yakushi.130.699 . PMID 20460867 .
- ^ Jin W, Lu Z (сентябрь 1998 г.). «Новый высокоаффинный ингибитор для K+ каналов k+-кантри». Биохимия . 37 (38): 13291–13299. doi : 10.1021/bi981178p . PMID 9748337 .
- ^ Kawaura K, Ogata Y, Inoue M, Honda S, Soeda F, Shirasaki T, Takahama K (декабрь 2009 г.). «Центрально действующий не наркотический противотуиный типепидин оказывает антидепрессант, подобный эффекту в испытании на принудительное плавание у крыс» (PDF) . Поведенческое исследование мозга . 205 (1): 315–318. doi : 10.1016/j.bbr.2009.07.004 . PMID 19616036 . S2CID 29236491 .
- ^ Kaufmann K, Romaine I, Days E, Pascual C, Malik A, Yang L, et al. (Сентябрь 2013 г.). «ML297 (VU0456810), первый мощный и селективный активатор канала калия Гирка, демонстрирует противоэпилептические свойства у мышей» . ACS Химическая нейробиология . 4 (9): 1278–1286. doi : 10.1021/cn400062a . PMC 3778424 . PMID 23730969 .
- ^ Serrano-Martín X, Payares G, Mendoza-León A (декабрь 2006 г.). «Глибенкламид, блокатор каналов K+(АТФ), демонстрирует антилейшманическую активность в экспериментальном кожном лейшманиозе мыши» . Антимикробные агенты и химиотерапия . 50 (12): 4214–4216. doi : 10.1128/aac.00617-06 . PMC 1693980 . PMID 17015627 .
- ^ Лоуренс К.Л., Прокс П., Родриго Г.С., Джонс П., Хаябучи Ю, Стенден Н.Б., Эшкрофт Ф.М. (август 2001 г.). «Gliclazide продуцирует высокоаффинный блок каналов KATP в изолированных бета-клетках поджелудочной железы мыши, но не сердца крысы или клеток гладких мышц артерий» . Диабетология . 44 (8): 1019–1025. doi : 10.1007/s001250100595 . PMID 11484080 .
- ^ Jump up to: а беременный в дюймовый и фон Enyedi P, Czirják G (апрель 2010 г.). «Молекулярный фон утечки K+ токов: двухпорсионные калиевые каналы» . Физиологические обзоры . 90 (2): 559–605. doi : 10.1152/physrev.00029.2009 . PMID 20393194 . S2CID 9358238 .
- ^ Jump up to: а беременный в дюймовый и фон Lotshaw DP (2007). «Биофизические, фармакологические и функциональные характеристики клонированных и нативных K+ каналов домена млекопитающих». Клеточная биохимия и биофизика . 47 (2): 209–256. doi : 10.1007/s12013-007-0007-8 . PMID 17652773 . S2CID 12759521 .
- ^ Fink M, Lesage F, Duprat F, Heurteaux C, Reyes R, Fosset M, Lazdunski M (июнь 1998 г.). «Нейрональный домен два P -домена K+, стимулированный арахидоновой кислотой и полиненасыщенными жирными кислотами» . Embo Journal . 17 (12): 3297–3308. doi : 10.1093/emboj/17.12.3297 . PMC 1170668 . PMID 9628867 .
- ^ Goldstein SA, Bockenhauer D, O'Kelly I, Zilberberg N (март 2001 г.). «Каналы утечки калия и семейство KCNK двух-P-доменных субъединиц» . Природные обзоры. Нейробиология . 2 (3): 175–184. doi : 10.1038/35058574 . PMID 11256078 . S2CID 9682396 .
- ^ Sano Y, Inamura K, Miyake A, Mochizuki S, Kitada C, Yokoi H, et al. (Июль 2003 г.). «Новый двухсконный домен K+-канал, Tresk, локализован в спинном мозге» . Журнал биологической химии . 278 (30): 27406–27412. doi : 10.1074/jbc.m206810200 . PMID 12754259 .
- ^ Czirják G, Tóth Ze, Enyedi P (апрель 2004 г.). «Канал K+ двухпоров K+, TRESK, активируется цитоплазматическим сигналом кальция через кальциневрин» . Журнал биологической химии . 279 (18): 18550–18558. doi : 10.1074/jbc.m312229200 . PMID 14981085 .
- ^ Kindler CH, Yost CS, Grey At (апрель 1999 г.). «Местное анестетическое ингибирование базовых калиевых каналов с двумя пор -доменами в тандеме» . Анестезиология . 90 (4): 1092–1102. doi : 10.1097/00000542-199904000-00024 . PMID 10201682 .
- ^ Jump up to: а беременный в Meadows HJ, Randall AD (март 2001 г.). «Функциональная характеристика человеческой задачи-3, чувствительного к кислоте двухско-доменного калиевого канала». Нейрофармакология . 40 (4): 551–559. doi : 10.1016/s0028-3908 (00) 00189-1 . PMID 11249964 . S2CID 20181576 .
- ^ Kindler CH, Paul M, Zou H, Liu C, Winegar BD, Grey AT, Yost CS (июль 2003 г.). «Амидные локальные анестетики активно ингибируют фон домена домена тандема человека K+ Task-2 Task-2 (KCNK5)». Журнал фармакологии и экспериментальной терапии . 306 (1): 84–92. doi : 10.1124/jpet.103.049809 . PMID 12660311 . S2CID 1621972 .
- ^ Punke MA, Licher T, Pongs O, Friederich P (июнь 2003 г.). «Ингибирование человеческих каналов Trek-1 Bupivacaine» . Анестезия и анальгезия . 96 (6): 1665–1673. doi : 10.1213/01.ane.0000062524.90936.1f . PMID 12760993 . S2CID 39630495 .
- ^ Lesage F, Guillemare E, Fink M, Duprat F, Lazdunski M, Romey G, Barhanin J (март 1996 г.). «TWIK-1, вездесущий человек слабо внутренний выпрямляющий K+-канал с новой структурой» . Embo Journal . 15 (5): 1004–1011. doi : 10.1002/j.1460-2075.1996.tb00437.x . PMC 449995 . PMID 8605869 .
- ^ Duprat F, Lesage F, Fink M, Reyes R, Heurteaux C, Lazdunski M (сентябрь 1997 г.). «Задача, человеческий фон K+ -канал, чтобы почувствовать внешние изменения pH вблизи физиологического рН» . Embo Journal . 16 (17): 5464–5471. doi : 10.1093/emboj/16.17.5464 . PMC 1170177 . PMID 9312005 .
- ^ Рейес Р., Дюпрат Ф., Лесадж Ф., Финк М., Салинас М., Фарман Н., Лаздунски М (ноябрь 1998). «Клонирование и экспрессия нового чувствительного к рН двух пор-домен K+ из почек человека» . Журнал биологической химии . 273 (47): 30863–30869. doi : 10.1074/jbc.273.47.30863 . PMID 9812978 .
- ^ Meadows HJ, Benham CD, Cairns W, Gloger I, Jennings C, Medhurst AD, et al. (Апрель 2000). «Клонирование, локализация и функциональная экспрессия человеческого ортолога канала калия Trek-1». Pflügers Archiv . 439 (6): 714–722. doi : 10.1007/s004240050997 . PMID 10784345 .
- ^ "Uniprotkb - q9npc2 (kcnk9_human)" . Uniprot . Получено 2019-05-29 .
- ^ Jump up to: а беременный Кеннард Ле, Чумлей -младший, Ранатунга К.М., Армстронг С.Дж., Вил Эл, Мати А (март 2005 г.). «Ингибирование человеческого двухпорного домена калиевого канала Trek-1, флуоксетином и его метаболитом норфуксетина» . Британский журнал фармакологии . 144 (6): 821–829. doi : 10.1038/sj.bjp.0706068 . PMC 1576064 . PMID 15685212 .
- ^ Patel AJ, Honoré E, Lesage F, Fink M, Romey G, Lazdunski M (май 1999 г.). «Ингаляционные анестетики активируют двухпоровные фон K+ каналы». Nature Neuroscience . 2 (5): 422–426. doi : 10.1038/8084 . PMID 10321245 . S2CID 23092576 .
- ^ Серый AT, Zhao BB, Kindler CH, Winegar BD, Mazurek MJ, Xu J, et al. (Июнь 2000 г.). «Излегальные анестетики активируют базовый домен Tandem Tandem Tandem Tandem Pore Kcnk5». Анестезиология . 92 (6): 1722–1730. doi : 10.1097/00000542-200006000-00032 . PMID 10839924 . S2CID 45487917 .
- ^ Кирш Г.Е., Нарахаши Т (июнь 1978 г.). «3,4-диаминопиридин. Мощный новый блокатор калиевого канала» . Биофизический журнал . 22 (3): 507–512. Bibcode : 1978bpj .... 22..507k . doi : 10.1016/s0006-3495 (78) 85503-9 . PMC 1473482 . PMID 667299 .
- ^ Jump up to: а беременный Судья SI, Bever CT (июль 2006 г.). «Блокаторы калия при рассеянном склерозе: нейрональные KV -каналы и эффекты симптоматического лечения». Фармакология и терапия . 111 (1): 224–259. doi : 10.1016/j.pharmthera.2005.10.006 . PMID 16472864 .
- ^ Tiku PE, Nowell Pt (декабрь 1991 г.). «Селективное ингибирование k (+)-стимуляция Na, K-АТФазы бретилием» . Британский журнал фармакологии . 104 (4): 895–900. doi : 10.1111/j.1476-5381.1991.tb12523.x . PMC 1908819 . PMID 1667290 .
- ^ Хилль Б (май 1967 г.). «Селективное ингибирование отсроченных токов калия в нерве с помощью иона тетраэтиламмония» . Журнал общей физиологии . 50 (5): 1287–1302. doi : 10.1085/jgp.50.5.1287 . PMC 2225709 . PMID 6033586 .
- ^ Армстронг КМ (октябрь 1971). «Взаимодействие производных ионов тетраэтиламмония с калиевыми каналами гигантских аксонов» . Журнал общей физиологии . 58 (4): 413–437. doi : 10.1085/jgp.58.4.413 . PMC 2226036 . PMID 5112659 .
- ^ "Амиодарон" . Наркоман . Получено 2019-05-28 .
- ^ Rogawski MA, Bazil CW (июль 2008 г.). «Новые молекулярные мишени для противоэпилептических препаратов: Alpha (2) Delta, SV2A и K (V) 7/KCNQ/M калиевые каналы» . Текущая неврология и неврологические отчеты . 8 (4): 345–352. doi : 10.1007/s11910-008-0053-7 . PMC 2587091 . PMID 18590620 .
- ^ Jump up to: а беременный Дойл Д.А., Морайс Кабрал Дж., Пфуэцнер Р.А., Куо А., Гулбис Дж. М., Коэн С.Л. и др. (Апрель 1998 г.). «Структура калиевого канала: молекулярная база проводимости и селективности K+». Наука . 280 (5360): 69–77. Bibcode : 1998sci ... 280 ... 69d . doi : 10.1126/science.280.5360.69 . PMID 9525859 .
- ^ Mackinnon R, Cohen SL, Kuo A, Lee A, Chait BT (апрель 1998 г.). «Структурное сохранение в прокариотических и эукариотических каналах калия». Наука . 280 (5360): 106–109. Bibcode : 1998sci ... 280..106M . doi : 10.1126/science.280.5360.106 . PMID 9525854 . S2CID 33907550 .
- ^ Армстронг С (апрель 1998 г.). «Видение пор». Наука . 280 (5360): 56–57. doi : 10.1126/science.280.5360.56 . PMID 9556453 . S2CID 35339674 .
- ^ «Нобелевская премия по химии 2003 года» . Нобелевский фонд . Получено 2007-11-16 .
- ^ Zhou Y, Morais-Cabral JH, Kaufman A, Mackinnon R (ноябрь 2001 г.). «Химия координации и увлажнения ионов, выявленная комплексом K+ канала в 2,0 A разрешения». Природа . 414 (6859): 43–48. Bibcode : 2001natur.414 ... 43Z . doi : 10.1038/35102009 . PMID 11689936 . S2CID 205022645 .
- ^ Lodish H, Berk A, Kaiser C, Krieger M, Bretscher A, Ploegh H, et al. (2016). Молекулярная клеточная биология (8 -е изд.). Нью -Йорк, Нью -Йорк: WH Freeman and Company. п. 499. ISBN 978-1-4641-8339-3 .
- ^ Sauer DB, Zeng W, Raghunathan S, Jiang Y (октябрь 2011 г.). «Белковые взаимодействия центральные для стабилизации фильтра селективности канала K+ в четырехместной конфигурации для селективного проникновения K+» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (40): 16634–16639. Bibcode : 2011pnas..10816634S . doi : 10.1073/pnas.1111688108 . PMC 3189067 . PMID 21933962 .
- ^ Jump up to: а беременный Hellgren M, Sandberg L, Edholm O (март 2006 г.). «Сравнение двух прокариотических каналов калия (Kirbac1.1 и KCSA) в имитационном исследовании молекулярной динамики (MD)». Биофизическая химия . 120 (1): 1–9. doi : 10.1016/j.bpc.2005.10.002 . PMID 16253415 .
- ^ Noskov SY, Roux B (февраль 2007 г.). «Важность гидратации и динамики в селективности каналов KCSA и NAK» . Журнал общей физиологии . 129 (2): 135–143. doi : 10.1085/jgp.200609633 . PMC 2154357 . PMID 17227917 .
- ^ Noskov SY, Bernèche S, Roux B (октябрь 2004 г.). «Контроль ионной селективности в калиевых каналах с помощью электростатических и динамических свойств карбонильных лигандов». Природа . 431 (7010): 830–834. Bibcode : 2004natur.431..830n . doi : 10.1038/nature02943 . PMID 15483608 . S2CID 4414885 .
- ^ Varma S, Rempe SB (август 2007 г.). «Настройка ионных координационных архитектур, чтобы обеспечить селективное разделение» . Биофизический журнал . 93 (4): 1093–1099. ARXIV : физика/0608180 . Bibcode : 2007bpj .... 93.1093V . doi : 10.1529/biophysj.107.107482 . PMC 1929028 . PMID 17513348 .
- ^ Томас М., Джаятилака Д., Корри Б. (октябрь 2007 г.). «Преобладающая роль координационного числа в селективности канала калия» . Биофизический журнал . 93 (8): 2635–2643. Bibcode : 2007bpj .... 93.2635t . doi : 10.1529/biophysj.107.108167 . PMC 1989715 . PMID 17573427 .
- ^ Bostick DL, Brooks CL (май 2007 г.). «Селективность в K+ каналах обусловлена топологическим контролем координированного состояния пермианта» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (22): 9260–9265. Bibcode : 2007pnas..104.9260b . doi : 10.1073/pnas.0700554104 . PMC 1890482 . PMID 17519335 .
- ^ Дереб М.Г., Сауэр Д.Б., Зенг В., Алам А., Ши Н., Цзян Ю (январь 2011 г.). «Настройка ионной селективности тетрамерных катионных каналов путем изменения количества сайтов связывания ион» » . Труды Национальной академии наук Соединенных Штатов Америки . 108 (2): 598–602. Bibcode : 2011pnas..108..598d . doi : 10.1073/pnas.1013636108 . PMC 3021048 . PMID 21187421 .
- ^ Morais-Cabral JH, Zhou Y, Mackinnon R (ноябрь 2001 г.). «Энергетическая оптимизация скорости ионной проводимости по фильтру K+ селективность». Природа . 414 (6859): 37–42. Bibcode : 2001natur.414 ... 37M . doi : 10.1038/35102000 . PMID 11689935 . S2CID 4429890 .
- ^ Jump up to: а беременный Yuan P, Leonetti MD, Pico AR, Hsiung Y, Mackinnon R (июль 2010 г.). «Структура устройства Ca2+-активации человека Ca2+-Activation при разрешении 3.0» . Наука . 329 (5988): 182–186. Bibcode : 2010sci ... 329..182y . doi : 10.1126/science.1190414 . PMC 3022345 . PMID 20508092 .
- ^ Jump up to: а беременный Wu Y, Yang Y, Ye S, Jiang Y (июль 2010 г.). «Структура стробильного кольца из человеческой большой проводимости Ca (2+)-Грубковое k (+) канал» . Природа . 466 (7304): 393–397. Bibcode : 2010natur.466..393W . doi : 10.1038/nature09252 . PMC 2910425 . PMID 20574420 .
- ^ Jump up to: а беременный Цзян Y, Пико А., Каден М., Чейт Б.Т., Макиннон Р. (март 2001 г.). «Структура домена RCK из канала E. coli K+ и демонстрация его присутствия в человеческом канале BK» . Нейрон . 29 (3): 593–601. doi : 10.1016/s0896-6273 (01) 00236-7 . PMID 11301020 . S2CID 17880955 .
- ^ Цзян Ю., Ли А., Чен Дж., Каден М., Чайт Б.Т., Макиннон Р (май 2002). «Кристаллическая структура и механизм калиевого канала, управляемого кальцием». Природа . 417 (6888): 515–522. Bibcode : 2002natur.417..515j . doi : 10.1038/417515a . PMID 12037559 . S2CID 205029269 .
- ^ Kong C, Zeng W, Ye S, Chen L, Sauer DB, Lam Y, et al. (Декабрь 2012 г.). «Отдельные механизмы стробирования, выявленные структурами многолигандного закрытого K (+) канала» . элиф . 1 : E00184. doi : 10.7554/elife.00184 . PMC 3510474 . PMID 23240087 .
- ^ Cao Y, Jin X, Huang H, Derebe MG, Levin EJ. (Март 2011 г.). Trkh " Природа 471 (738): 336–3 Bibcode : 2011 Natur . doi : 10.1038/ природа PMC 3077569 . PMID 21317882 .
- ^ Uysal S, Cuello LG, Cortes DM, Koide S, Kossiakoff AA, Perozo E (июль 2011 г.). «Механизм активации стробирования в полноразмерном канале KCSA K+» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (29): 11896–11899. Bibcode : 2011pnas..10811896U . doi : 10.1073/pnas.1105112108 . PMC 3141920 . PMID 21730186 .
- ^ Клейтон Г.М., Сильверман В.Р., Хегинботам Л., Морайс-Кабрал Дж.Х. (ноябрь 2004 г.). «Структурная основа активации лиганда в циклическом нуклеотидном регулируемом канале калия» . Клетка . 119 (5): 615–627. doi : 10.1016/j.cell.2004.10.030 . PMID 15550244 . S2CID 14149494 .
- ^ Цзян Ю., Ли А., Чен Дж., Рута В., Каден М., Чейт Б.Т., Макиннон Р. (май 2003). «Рентгеновская структура зависимого от напряжения K+-канала». Природа . 423 (6935): 33–41. Bibcode : 2003natur.423 ... 33J . doi : 10.1038/nature01580 . PMID 12721618 . S2CID 4347957 .
- ^ Long SB, Campbell EB, Mackinnon R (август 2005 г.). «Кристаллическая структура из-за зависимого от напряжения млекопитающего шейкера K+ канал» . Наука . 309 (5736): 897–903. Bibcode : 2005sci ... 309..897L . doi : 10.1126/science.1116269 . PMID 16002581 . S2CID 6072007 .
- ^ ANTZ C, Fakler B (август 1998 г.). «Быстрая инактивация каналов K (+), управляемых напряжением: от мультфильма к структуре». Новости в физиологических науках . 13 (4): 177–182. doi : 10.1152/physiologyonline.1998.13.4.177 . PMID 11390785 . S2CID 38134756 .
- ^ Cheng WW, McCoy JG, Thompson AN, Nichols CG , Nimigean CM (март 2011 г.). «Механизм для связи с инактивацией селективности в калиевых каналах KCSA» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (13): 5272–5277. BIBCODE : 2011PNAS..108.5272C . doi : 10.1073/pnas.1014186108 . PMC 3069191 . PMID 21402935 .
- ^ Cuello LG, Jogini V, Cortes DM, Perozo E (июль 2010 г.). «Структурный механизм инактивации C-типа в k (+) каналах» . Природа . 466 (7303): 203–208. Bibcode : 2010natur.466..203c . doi : 10.1038/nature09153 . PMC 3033749 . PMID 20613835 .
- ^ Cuello LG, Jogini V, Cortes DM, Pan AC, Gagnon DG, Dalmas O, et al. (Июль 2010). «Структурная основа для связи между воротами активации и инактивации в каналах k (+)» . Природа . 466 (7303): 272–275. BIBCODE : 2010NATR.466..272C . doi : 10.1038/nature09136 . PMC 3033755 . PMID 20613845 .
- ^ Luzhkov VB, Aqvist J (февраль 2005 г.). «Ионы и блокаторы в калиевых каналах: понимание моделирования свободной энергии». Biochimica et Biophysica Acta (BBA) - белки и протеомика . 1747 (1): 109–120. doi : 10.1016/j.bbapap.2004.10.006 . PMID 15680245 .
- ^ Luzhkov VB, Osterberg F, Aqvist J (ноябрь 2003 г.). «Структурно-активность взаимосвязь для внеклеточного блока K+ каналов с помощью ионов TetraAlcyLammonium» . Письма Febs . 554 (1–2): 159–164. Bibcode : 2003 Phebsl.554..159L . doi : 10.1016/s0014-5793 (03) 01117-7 . PMID 14596932 . S2CID 32031835 .
- ^ Поссон DJ, McCoy JG, Nimigean CM (февраль 2013 г.). «Зависимые от напряжения ворота в каналах калия MTHK расположены в фильтре селективности» . Природа структурная и молекулярная биология . 20 (2): 159–166. doi : 10.1038/nsmb.2473 . PMC 3565016 . PMID 23262489 .
- ^ Choi KL, Mossman C, Aubé J, Yellen G (март 1993 г.). «Внутренний четвертый рецептор аммония в шейкере калий -каналов». Нейрон . 10 (3): 533–541. doi : 10.1016/0896-6273 (93) 90340-W . PMID 8461140 . S2CID 33361945 .
- ^ Piasta KN, Theobald DL, Miller C (октябрь 2011 г.). «Калиевый селективный блок проникновения бария через отдельные каналы KCSA» . Журнал общей физиологии . 138 (4): 421–436. doi : 10.1085/jgp.201110684 . PMC 3182450 . PMID 21911483 .
- ^ Нейтон Дж., Миллер С (ноябрь 1988 г.). «Калиевые блокируют проницаемость бария через калийный калиевый канал» . Журнал общей физиологии . 92 (5): 549–567. doi : 10.1085/jgp.92.5.549 . PMC 2228918 . PMID 3235973 .
- ^ Бесплатный SW, Zhou M, Mackinnon R (май 2007 г.). «Структурные и термодинамические свойства селективного ионного связывания в канале K+» . PLOS Биология . 5 (5): E121. doi : 10.1371/journal.pbio.0050121 . PMC 1858713 . PMID 17472437 .
- ^ Цзян Й, Макиннон Р. (март 2000 г.). «Сайт бария в канале калия от рентгеновской кристаллографии» . Журнал общей физиологии . 115 (3): 269–272. doi : 10.1085/jgp.115.3.269 . PMC 2217209 . PMID 10694255 .
- ^ Lam YL, Zeng W, Sauer DB, Jiang Y (август 2014 г.). «Консервативный фильтр калиевого канала может иметь различные профили ионного связывания: структурный анализ связывания рубидия, цезия и бария в NAK2K» . Журнал общей физиологии . 144 (2): 181–192. doi : 10.1085/jgp.201411191 . PMC 4113894 . PMID 25024267 .
- ^ Guo R, Zeng W, Cui H, Chen L, Ye S (август 2014 г.). «Ионные взаимодействия блокад BA2+ в канале MTHK K+» . Журнал общей физиологии . 144 (2): 193–200. doi : 10.1085/jgp.201411192 . PMC 4113901 . PMID 25024268 .
- ^ Крапивинский Г., Гордон Э.А., Викман К., Велимирович Б., Крапивинский Л., Клэпхем де (март 1995 г.). «G-белок-предсердный K+-канал IKACH является гетеромультимером двух внутренних выпрямляющих белков K (+)-канала». Природа . 374 (6518): 135–141. Bibcode : 1995natur.374..135K . doi : 10.1038/374135A0 . PMID 7877685 . S2CID 4334467 .
- ^ Кори С., Крапивинский Г., Крапивинский Л., Клэпхэм де (февраль 1998 г.). «Число и стехиометрия субъединиц в нативном G-белковом канале, управляемом предсердием K+, Ikach» . Журнал биологической химии . 273 (9): 5271–5278. doi : 10.1074/jbc.273.9.5271 . PMID 9478984 .
- ^ Kunkel MT, Peralta EG (ноябрь 1995 г.). «Идентификация доменов, предоставляющих регуляцию G -белка на каналах калия внутреннего выпрямителя» . Клетка . 83 (3): 443–449. doi : 10.1016/0092-8674 (95) 90122-1 . PMID 8521474 . S2CID 14720432 .
- ^ Викман К., Крапивинский Г., Кори С., Кеннеди М., Немек Дж., Медина I, Клапхэм де (апрель 1999 г.). «Структура, активация белка G и функциональная значимость сердечного G-белка, управляемого K+-канала, Ikach» . Анналы нью -йоркской академии наук . 868 (1): 386–398. Bibcode : 1999nyasa.868..386w . doi : 10.1111/j.1749-6632.1999.tb11300.x . PMID 10414308 . S2CID 25949938 . Архивировано из оригинала 2006-01-29.
- ^ Ball P (март 2008 г.). «Крусибл: искусство, вдохновленное наукой, должно быть больше, чем просто симпатичная картина» . Мир химии . 5 (3): 42–43 . Получено 2009-01-12 .
Внешние ссылки
[ редактировать ]- Proteopedia channel Potassium channel in 3D
- Калиум+каналы в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)
- Центр нервно-мышечных заболеваний (2008-03-04). «Калиевые каналы» . Вашингтонский университет в Сент -Луисе . Получено 2008-03-10 .