Натриевый канал

Натриевые каналы представляют собой интегральные мембранные белки , образующие ионные каналы , проводящие ионы натрия (Na + ) через клеточную мембрану . [1] [2] Они принадлежат к суперсемейству катионных каналов .
Классификация
[ редактировать ]Их разделяют на 2 типа:
| Тип натриевого канала | Синонимы | Курок (фактор, стимулирующий канал) |
|---|---|---|
| Потенциал-управляемые натриевые каналы |
| изменение мембранного потенциала , которое также называют изменением напряжения |
| лиганд-управляемые натриевые каналы | LGSC | связывание веществ, таких как лиганды, с каналом |
| Утечка натриевого канала | НАЛКН | Без ворот, всегда открыт |
Функция
[ редактировать ]В возбудимых клетках, таких как нейроны , миоциты и некоторые типы глии , натриевые каналы ответственны за фазу роста действия потенциалов . Эти каналы проходят три различных состояния: состояние покоя, активное и неактивное. Несмотря на то, что состояния покоя и неактивности не позволяют ионам проходить через каналы, существует разница в их структурной конформации.
Селективность
[ редактировать ]Натриевые каналы обладают высокой избирательностью в транспортировке ионов через клеточные мембраны. Высокая селективность по отношению к ионам натрия достигается множеством различных способов. Все они подразумевают инкапсуляцию иона натрия в полости определенного размера внутри более крупной молекулы. [3]
Потенциал-управляемые натриевые каналы
[ редактировать ]Структура
[ редактировать ]
Натриевые каналы состоят из крупных альфа-субъединиц , которые связаны с вспомогательными белками, такими как бета-субъединицы . Альфа-субъединица образует ядро канала и функционирует сама по себе. Когда белок альфа-субъединицы экспрессируется клеткой, он способен образовывать поры в клеточной мембране, которые проводят Na. + потенциалзависимым образом, даже если бета-субъединицы или другие известные модулирующие белки не экспрессируются. Когда вспомогательные белки собираются с α-субъединицами, образующийся комплекс может демонстрировать измененную зависимость от напряжения и клеточную локализацию.
Альфа-субъединица состоит из четырех повторяющихся доменов, обозначенных от I до IV, каждый из которых содержит шесть трансмембранных сегментов, обозначенных от S1 до S6. Высококонсервативный сегмент S4 действует как датчик напряжения канала. Чувствительность этого канала к напряжению обусловлена положительными аминокислотами, расположенными в каждой третьей позиции. [5] При стимуляции изменением трансмембранного напряжения этот сегмент перемещается к внеклеточной стороне клеточной мембраны, позволяя каналу стать проницаемым для ионов. Ионы проводятся через центральную полость поры, которая состоит из двух основных областей. Более внешняя (т.е. более внеклеточная) часть поры образована «P-петлями» (область между S5 и S6) четырех доменов. Эта область является самой узкой частью поры и отвечает за ее ионную селективность. Внутренняя часть (т.е. более цитоплазматическая) поры представляет собой ворота поры и образована объединенными сегментами S5 и S6 четырех доменов. Поровый домен также имеет боковые туннели или фенестрации, которые проходят перпендикулярно оси поры. Предполагается, что эти фенестрации, соединяющие центральную полость с мембраной, важны для доступности лекарств. [6] [7] [8]
В натриевых каналах млекопитающих область, связывающая домены III и IV, также важна для функции канала. Этот линкер DIII-IV отвечает за закрытие ворот поры после открытия канала, инактивируя их. [9]
ворота
[ редактировать ]Напряжение-управляемый Na + каналы имеют три основных конформационных состояния: закрытое, открытое и инактивированное. Переходы вперед/назад между этими состояниями соответственно называются активацией/деактивацией (между открытым и закрытым соответственно), инактивацией/реактивацией (между неактивным и открытым соответственно) и восстановлением из инактивации/инактивацией закрытого состояния (между неактивным и закрытым состоянием). , соответственно). Закрытые и инактивированные состояния непроницаемы для ионов.
До возникновения потенциала действия аксональная мембрана находится в нормальном потенциале покоя , около -70 мВ в большинстве нейронов человека, и Na + каналы находятся в деактивированном состоянии и заблокированы на внеклеточной стороне воротами активации . В ответ на повышение мембранного потенциала примерно до -55 мВ (в данном случае вызванное потенциалом действия) активационные ворота открываются, позволяя положительно заряженному Na + ионы поступают в нейрон через каналы, вызывая повышение напряжения на мембране нейрона до +30 мВ в нейронах человека. Поскольку напряжение на мембране изначально отрицательное, а по мере того, как его напряжение увеличивается до нуля и выше (от -70 мВ в состоянии покоя до максимума +30 мВ), говорят, что оно деполяризуется. Это увеличение напряжения представляет собой фазу нарастания потенциала действия.
| Потенциал действия | Мембранный потенциал | Целевой потенциал | Целевое состояние ворот | Целевое состояние нейрона |
|---|---|---|---|---|
| Отдых | −70 мВ | −55 мВ | Деактивировано → Активировано | поляризованный |
| Восходящий | −55 мВ | 0 мВ | Активировано | Поляризованный → Деполяризованный |
| Восходящий | 0 мВ | +30 мВ | Активировано → Деактивировано | деполяризованный |
| Падение | +30 мВ | 0 мВ | Деактивирован | Деполяризованный → Реполяризованный |
| Падение | 0 мВ | −70 мВ | Деактивирован | реполяризованный |
| Перекус | −70 мВ | −75 мВ | Деактивировано → Деактивировано | Реполяризованный → Гиперполяризованный |
| Отскок | −75 мВ | −70 мВ | Деактивирован | Гиперполяризованный → Поляризованный |
На пике потенциала действия, когда достаточно Na + проник в нейрон и потенциал мембраны стал достаточно высоким, Na + каналы инактивируются, закрывая свои ворота инактивации . Ворота инактивации можно рассматривать как «пробку», привязанную к доменам III и IV внутриклеточной альфа-субъединицы канала. Закрытие ворот инактивации вызывает Na + поток через канал прекращается, что, в свою очередь, приводит к прекращению роста мембранного потенциала. Закрытие ворот инактивации создает рефрактерный период внутри каждого отдельного Na. + канал. Этот рефрактерный период исключает возможность движения потенциала действия в противоположном направлении обратно к соме. Когда ворота инактивации закрыты, канал считается инактивированным. С На + Канал больше не вносит вклад в мембранный потенциал, потенциал снижается обратно до потенциала покоя по мере того, как нейрон реполяризуется и впоследствии гиперполяризуется, и это составляет фазу спада потенциала действия. Таким образом, рефрактерный период каждого канала жизненно важен для однонаправленного распространения потенциала действия вниз по аксону для правильной связи между нейронами.
Когда напряжение мембраны становится достаточно низким, ворота инактивации снова открываются, а ворота активации закрываются в процессе, называемом деинактивацией . Когда ворота активации закрыты и ворота инактивации открыты, Na + канал снова находится в деактивированном состоянии и готов участвовать в другом потенциале действия.
Когда какой-либо ионный канал не инактивируется, о нем говорят, что он постоянно (или тонически) активен. Некоторые виды ионных каналов по своей природе постоянно активны. Однако генетические мутации, вызывающие постоянную активность в других каналах, могут вызвать заболевание, вызывая чрезмерную активность определенных типов нейронов. Мутации, мешающие Na + инактивация каналов может способствовать сердечно-сосудистым заболеваниям или эпилептическим припадкам из-за оконных токов , что может привести к перевозбуждению мышечных и/или нервных клеток.
Моделирование поведения ворот
[ редактировать ]Временное поведение Na + каналы можно моделировать с помощью марковской схемы или формализма типа Ходжкина – Хаксли . В первой схеме каждый канал занимает отдельное состояние с дифференциальными уравнениями, описывающими переходы между состояниями; во втором случае каналы рассматриваются как совокупность , на которую влияют три независимые переменные стробирования. Каждая из этих переменных может принимать значения от 1 (полная проницаемость для ионов) до 0 (полная непроницаемость), произведение этих переменных дает процент проводящих каналов. Можно показать, что модель Ходжкина – Хаксли эквивалентна марковской модели. [ нужны дальнейшие объяснения ]
Непроницаемость для других ионов
[ редактировать ]Поры натриевых каналов содержат селективный фильтр, состоящий из отрицательно заряженных аминокислотных остатков, которые притягивают положительные Na. + ион и не допускайте отрицательно заряженных ионов, таких как хлорид . Катионы текут в более суженную часть поры шириной 0,3 на 0,5 нм , что достаточно велико, чтобы позволить одному Na + воды, ион с молекулой связанной с прохождением. Чем больше К + ион не может пройти через эту область. Ионы разных размеров также не могут хорошо взаимодействовать с отрицательно заряженными остатками глутаминовой кислоты , выстилающими поры. [ нужна ссылка ]
Разнообразие
[ редактировать ]Потенциал-управляемые натриевые каналы обычно состоят из альфа-субъединицы, которая образует пору ионной проводимости, и одной-двух бета-субъединиц, которые выполняют несколько функций, включая модуляцию шлюзования каналов. [10] Экспрессии одной альфа-субъединицы достаточно для создания функционального канала.
Альфа-субъединицы
[ редактировать ]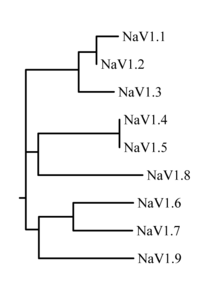
Семейство натриевых каналов включает 9 известных членов с идентичностью аминокислот >50% в трансмембранных сегментах и внеклеточных петлевых областях. В настоящее время используется стандартизированная номенклатура натриевых каналов, которая поддерживается IUPHAR . [11]
Белки этих каналов получили названия от Na v 1.1 до Na v 1.9. Названия генов обозначаются как от SCN1A до SCN5A, затем от SCN8A до SCN11A. [11] «Десятый член», Na x , не действует потенциалзависимым образом. Он имеет схожую общую структуру. О его реальной функции известно немногое, кроме того, что он также связан с бета-субъединицами. [12]
Вероятная эволюционная связь между этими каналами, основанная на сходстве их аминокислотных последовательностей, показана на рисунке 1. Отдельные натриевые каналы отличаются не только различиями в их последовательностях, но также по их кинетике и профилям экспрессии. Некоторые из этих данных обобщены в таблице 1 ниже.
| Название белка | Ген | Профиль выражения | Сопутствующие каналопатии человека |
|---|---|---|---|
| В версии 1.1 | SCN1A | Центральные нейроны , [периферические нейроны] и сердечные миоциты | фебрильная эпилепсия , GEFS+ , синдром Драве (также известный как тяжелая миклоническая эпилепсия младенчества или SMEI), пограничный SMEI (SMEB), синдром Веста (также известный как инфантильные спазмы ), синдром Дуза (также известный как миоклоническая астатическая эпилепсия ), трудноизлечимая детская эпилепсия с генерализованными тонико-клоническими припадками (ICEGTC), синдромом Панайотопулоса, семейной гемиплегической мигренью (СГМ), семейным аутизмом, энцефалитом Расмуссена и синдромом Леннокса-Гасто [13] |
| На версии 1.2 | SCN2A | Центральные нейроны, периферические нейроны | наследственные фебрильные судороги , эпилепсия и расстройства аутистического спектра. |
| На версии 1.3 | SCN3A | Центральные нейроны, периферические нейроны и кардиомиоциты | эпилепсия, боль, пороки развития головного мозга [14] [15] |
| На версии 1.4 | SCN4A | Скелетные мышцы | гиперкалиемический периодический паралич , врожденная парамиотония и миотония, отягощенная калием. |
| На версии 1.5 | SCN5A | Сердечные миоциты, неиннервированные скелетные мышцы, центральные нейроны, гладкомышечные клетки желудочно-кишечного тракта и интерстициальные клетки Кахаля. | Со стороны сердца: синдром удлиненного интервала QT 3 типа, синдром Бругада , прогрессирующее заболевание сердечной проводимости , семейная фибрилляция предсердий и идиопатическая фибрилляция желудочков ; [16] Желудочно-кишечный тракт: синдром раздраженного кишечника ; [17] |
| На версии 1.6 | SCN8A | Центральные нейроны, ганглии дорсальных корешков , периферические нейроны , сердце, глиальные клетки. | Эпилепсия , [18] атаксия , дистония , тремор [19] |
| На версии 1.7 | SCN9A | Ганглии дорсальных корешков , симпатические нейроны, шванновские клетки и нейроэндокринные клетки. | эритромелалгия , ПЭПД , нечувствительность к боли, связанная с каналопатией [14] и недавно обнаружил инвалидизирующую форму фибромиалгии (полиморфизм rs6754031). [20] |
| На версии 1.8 | SCN10A | Дорсальные корешковые ганглии | боль, [14] нервно-психические расстройства |
| В версии 1.9 | SCN11A | Дорсальные корешковые ганглии | боль [14] |
| На х | SCN7A | сердце, матка, скелетные мышцы, астроциты, ганглиозные клетки дорсальных корешков | никто не известен |
Бета-субъединицы
[ редактировать ]Бета-субъединицы натриевых каналов представляют собой трансмембранные гликопротеины типа 1 с внеклеточным N-концом и цитоплазматическим С-концом. Будучи членами суперсемейства Ig, бета-субъединицы содержат прототипную петлю V-set Ig во внеклеточном домене. Они не имеют никакой гомологии со своими аналогами кальциевых и калиевых каналов. [21] Вместо этого они гомологичны молекулам адгезии нервных клеток (CAM) и большому семейству CAM L1. Существует четыре различных бета-версии, названных в порядке открытия: SCN1B, SCN2B, SCN3B, SCN4B (таблица 2). Бета-1 и бета-3 взаимодействуют с альфа-субъединицей нековалентно, тогда как бета-2 и бета-4 связываются с альфа-субъединицей через дисульфидную связь. [22] Натриевые каналы с большей вероятностью остаются открытыми на подпороговом мембранном потенциале при взаимодействии с бета-токсинами, что, в свою очередь, вызывает немедленное ощущение боли. [23]
Роль бета-субъединиц как молекул клеточной адгезии
[ редактировать ]Помимо регуляции открытия каналов, бета-субъединицы натриевых каналов также модулируют экспрессию каналов и образуют связи с внутриклеточным цитоскелетом через анкирин и спектрин . [10] [24] [25] Потенциал-управляемые натриевые каналы также собираются с множеством других белков, таких как белки FHF (гомологичный фактор фактора роста фибробластов), кальмодулин, цитоскелет или регуляторные киназы. [26] [10] [27] [28] [29] которые образуют комплекс с натриевыми каналами, влияя на его экспрессию и/или функцию. Несколько бета-субъединиц взаимодействуют с одной или несколькими молекулами внеклеточного матрикса (ECM). Контактин, также известный как F3 или F11, связывается с бета-1, как показано посредством коиммунопреципитации. [30] Фибронектин- подобные (FN-подобные) повторы тенасцина -C и тенасцина -R связываются с бета-2, в отличие от повторов, подобных эпидермальному фактору роста (EGF-подобных), которые отталкивают бета2. [31] Дезинтегрин и металлопротеиназа (ADAM) 10 отбрасывает эктодомен бета 2, возможно, вызывая рост нейритов. [32] Бета-3 и бета-1 связываются с нейрофасцином в узлах Ранвье в развивающихся нейронах. [33]
| Название белка | Генная ссылка | Собирается с | Профиль выражения | Сопутствующие каналопатии человека |
|---|---|---|---|---|
| На v β1 | SCN1B | от Na v 1,1 до Na v 1,7 | Центральные нейроны, периферические нейроны, скелетные мышцы, сердце, глия. | эпилепсия (GEFS+), синдром Бругада [34] |
| Na v β2 | СКН2Б | от Na v 1,1, от Na v 1,2, от Na v 1,5 до Na v 1,7 | Центральные нейроны, периферические нейроны, сердце, глия | синдром Бругада [34] |
| Na v β3 | SCN3B | от Na v 1,1 до Na v 1,3, Na v 1,5 | центральные нейроны, надпочечники, почки, периферические нейроны | синдром Бругада [34] |
| Na v β4 | SCN4B | Na v 1,1, Na v 1,2, Na v 1,5 | сердце, скелетные мышцы, центральные и периферические нейроны | никто не известен |
Лиганд-управляемые натриевые каналы
[ редактировать ]Лиганд-управляемые натриевые каналы активируются путем связывания лиганда, а не изменения мембранного потенциала.
Они обнаруживаются, например, в нервно-мышечных соединениях в виде никотиновых рецепторов , где лигандами являются ацетилхолина молекулы . Большинство каналов этого типа в той или иной степени проницаемы для калия, а также для натрия.
Роль в потенциале действия
[ редактировать ]Потенциал-управляемые натриевые каналы играют важную роль в потенциалах действия . Если при изменении мембранного потенциала клетки открывается достаточное количество каналов , небольшое, но значительное количество Na + ионы будут перемещаться в клетку по электрохимическому градиенту , еще больше деполяризуя клетку. Таким образом, чем больше Na + Каналы, локализованные в области клеточной мембраны, тем быстрее будет распространяться потенциал действия и тем более возбудимой будет эта область клетки. Это пример положительной обратной связи . Способность этих каналов принимать закрыто-инактивированное состояние вызывает рефрактерный период и имеет решающее значение для распространения потенциалов действия вниз по аксону .
Уже + каналы открываются и закрываются быстрее, чем K + каналы , производящие приток положительного заряда (Na + ) к началу потенциала действия и истечению (K + ) ближе к концу.
С другой стороны, лиганд-управляемые натриевые каналы создают изменение мембранного потенциала в первую очередь в ответ на связывание с ним лиганда. Каналы утечки натрия дополнительно способствуют регуляции потенциала действия путем модуляции потенциала покоя (и, в свою очередь, возбудимости) клетки. [35]
Фармакологическая модуляция
[ редактировать ]Блокаторы
[ редактировать ]Активаторы
[ редактировать ]Следующие природные вещества стойко активируют (открывают) натриевые каналы:
- алкалоидов Токсины на основе
- аконитин
- батрахотоксин
- бреветоксин
- сигуатоксин
- дельфинин
- некоторые граанотоксины , например, граанотоксин I (другие гранотоксины неактивны или закрывают натриевые каналы)
- правдиво
Модификаторы стробирования
[ редактировать ]Следующие токсины изменяют работу натриевых каналов:
- пептидов Токсины на основе
- μ-конотоксин
- δ-атракотоксин [36]
- яда скорпиона Токсины [37]
Канал утечки натрия (NALCN)
[ редактировать ]В каналах утечки натрия не наблюдается никакого напряжения или лигандного стробирования. Вместо этого они всегда открыты или «протекают» небольшой фоновый ток, регулирующий мембранный потенциал покоя нейрона. [35] У большинства животных один ген кодирует белок NALCN (канал утечки натрия, неселективный). [38]
Структурные и функциональные различия
[ редактировать ]Несмотря на ту же базовую структуру, что и другие натриевые каналы, NALCN не чувствителен к изменениям напряжения. Потенциально-чувствительный трансмембранный домен S4 NALCN имеет меньше положительно заряженных аминокислот (13 вместо 21 в потенциалзависимом канале), что, возможно, объясняет его нечувствительность к напряжению. [35] NALCN также гораздо менее селективен к Na. + ионов и проницаем для Ca 2+ и К + ионы. Аминокислотный мотив EEKE в домене порового фильтра NALCN подобен как мотиву EEEE потенциал -управляемого кальциевого канала , так и мотиву DEKA потенциал-управляемого натриевого канала, что, возможно, объясняет его недостаточную селективность. [38]

NALCN не блокируется многими распространенными блокаторами натриевых каналов, включая тетродотоксин . NALCN неспецифически блокируется как Gd 3+ и верапамил . [39] Вещество P и нейротензин активируют киназы семейства Src через соответствующие GPCR (независимо от связанных G-белков ), которые, в свою очередь, увеличивают проницаемость NALCN посредством активации UNC80. [40] Ацетилхолин также может повышать активность NALCN через М 3 мускариновые ацетилхолиновые рецепторы . [41] Более высокие уровни внеклеточного кальция 2+ уменьшают проницаемость NALCN путем активации CaSR , который ингибирует UNC80. [42]
Белковый комплекс
[ редактировать ]NALCN образует комплексы с белками UNC79, UNC80 и FAM155A. [43] [44] [45] UNC79, по-видимому, связан с мембранной стабильностью NALCN и связью с UNC 80. [44] UNC80 опосредует химическую модуляцию NALCN несколькими путями. [35] [42] [41] [40] FAM155A помогает сворачивать белки в эндоплазматическом ретикулуме, транспортировать шапероны к аксону и способствует стабильности мембраны. [45]
Биологическая функция
[ редактировать ]Мембранный потенциал покоя нейрона обычно составляет от -60 мВ до -80 мВ и обусловлен главным образом K + потенциал при -90мВ. Деполяризация от K + потенциал обусловлен в первую очередь небольшим содержанием Na + ток утечки. Около 70% этого тока проходит через NALCN. [39] Увеличение проницаемости NALCN снижает мембранный потенциал покоя, приближая его к триггеру потенциала действия (-55 мВ), тем самым увеличивая возбудимость нейрона.
Роль в патологии
[ редактировать ]Мутации NALCN приводят к серьезным нарушениям дыхательного ритма у мышей [39] и измененное циркадное передвижение у мух. [46] Мутации NALCN также связаны с множественными тяжелыми нарушениями развития. [47] и цервикальная дистония. [48] Шизофрения и биполярное расстройство также связаны с мутациями NALCN. [49]
Модуляция pH
[ редактировать ]Изменения рН крови и тканей сопровождают физиологические и патофизиологические состояния, такие как физические упражнения, ишемия сердца, ишемический инсульт и прием кокаина. Известно, что эти состояния вызывают симптомы электрических заболеваний у пациентов с мутациями натриевых каналов. Протоны вызывают разнообразный набор изменений в воротах натриевых каналов, которые обычно приводят к уменьшению амплитуды переходного натриевого тока и увеличению доли неинактивирующих каналов, пропускающих постоянные токи. Эти эффекты характерны для болезнетворных мутантов в нейронах, скелетных мышцах и сердечной ткани и могут усугубляться у мутантов, которые придают большую чувствительность к протонам натриевым каналам, что указывает на роль протонов в запуске острых симптомов электрического заболевания. [50]
Молекулярные механизмы протонной блокировки
[ редактировать ]Данные одного канала кардиомиоцитов показали, что протоны могут снижать проводимость отдельных натриевых каналов. [51] Фильтр селективности натриевых каналов состоит из одного остатка в каждой из четырех петель пор четырех функциональных доменов. Эти четыре остатка известны как мотив DEKA. [52] Скорость проникновения натрия через натриевый канал определяется четырьмя карбоксилатными остатками, мотивом EEDD, которые составляют внешнее заряженное кольцо. [52] Протонирование этих карбоксилатов является одним из основных факторов блокады протонов в натриевых каналах, хотя существуют и другие остатки, которые также способствуют чувствительности к pH. [53] Одним из таких остатков является C373 в натриевых каналах сердца , что делает его наиболее чувствительным к pH натриевым каналом среди изученных на сегодняшний день натриевых каналов. [54]
Модуляция pH воротами натриевых каналов
[ редактировать ]Поскольку сердечный натриевый канал является наиболее чувствительным к pH натриевым каналом, большая часть того, что известно, основана на этом канале. Было показано, что снижение внеклеточного pH деполяризует зависимость активации и инактивации от напряжения до более положительных потенциалов. Это указывает на то, что во время действий, которые снижают pH крови, таких как физические упражнения, вероятность активации и инактивации каналов выше, чем более положительные мембранные потенциалы, что может привести к потенциальным неблагоприятным последствиям. [55] Натриевые каналы, экспрессируемые в волокнах скелетных мышц, превратились в относительно pH-нечувствительные каналы. Было высказано предположение, что это защитный механизм против потенциальной чрезмерной или недостаточной возбудимости скелетных мышц, поскольку уровень pH крови очень чувствителен к изменению во время движения. [56] [57] Недавно было показано, что мутация смешанного синдрома, вызывающая периодический паралич и миотонию в скелетных натриевых каналах, придает этому каналу чувствительность к pH, что делает ворота этого канала сходными с таковыми при сердечном подтипе. [58]
Модуляция pH в изученных на данный момент подтипах
[ редактировать ]Эффекты протонирования охарактеризованы в Na v 1,1–Na v 1,5. Среди этих каналов Na v 1,1–Na v 1,3 и Na v 1,5 демонстрируют деполяризованную зависимость активации от напряжения, в то время как активация Na v 1,4 остается нечувствительной к ацидозу. Зависимость стационарной быстрой инактивации от напряжения не меняется в Na v 1,1–Na v 1,4, но стационарная быстрая инактивация в Na v 1,5 деполяризуется. Следовательно, среди изученных на сегодняшний день натриевых каналов Na v 1,4 является наименее чувствительным, а Na v 1,5 — наиболее протон-чувствительным подтипом. [59]
См. также
[ редактировать ]- Кальциевый канал - комплекс ионных каналов, через который проходят ионы кальция.
- Хронаксия - электрофизиологический показатель.
- Эпителиальный натриевый канал - группа мембранных белков.
- Ионный канал – мембранный белок, образующий поры.
- Канал покоящихся ионов - класс транспортных белков.
- Натрий в биологии - Использование натрия организмами
Ссылки
[ редактировать ]- ^ Джесселл ТМ, Кэндл ЭР , Шварц Дж.Х. (2000). Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. стр. 100-1 154–69 . ISBN 978-0-8385-7701-1 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бертиль Гилель (2001). Ионные каналы возбудимых мембран (3-е изд.). Сандерленд, Массачусетс: Синауэр. стр. 73–7. ISBN 978-0-87893-321-1 .
- ^ Лим С., Дудев Т. (2016). «Глава 10. Селективность по калию и натрию в фильтрах с селективностью моновалентных ионных каналов». В Астрид С., Хельмут С., Роланд К.О. С. (ред.). Ионы щелочных металлов: их роль в жизни . Ионы металлов в науках о жизни. Том. 16. Спрингер. стр. 325–347. дои : 10.1007/978-3-319-21756-7_9 . ISBN 978-3-319-21755-0 . ПМИД 26860305 .
- ^ Ю Ф.Х., Каттералл, Вашингтон (2003). «Обзор семейства потенциал-управляемых натриевых каналов» . Геномная биология . 4 (3): 207. doi : 10.1186/gb-2003-4-3-207 . ПМК 153452 . ПМИД 12620097 .
- ^ Николлс, Мартин, Фукс, Браун, Даймонд, Вайсблат. (2012) «От нейрона к мозгу», 5-е изд. стр. 86
- ^ Хилле, Б. (1977) Местные анестетики: гидрофильные и гидрофобные пути реакции лекарственного средства на рецептор. Журнал общей физиологии, 69, 497–515. http://dx.doi.org/10.1085/jgp.69.4.497
- ^ Гамаль Эль-Дин, Тамер М. и др. «Фенестрации контролируют блокировку потенциалзависимого натриевого канала в состоянии покоя». Труды Национальной академии наук 115.51 (2018): 13111-13116. https://doi.org/10.1073/pnas.1814928115
- ^ Тао, Элейн и Бен Корри. «Характеристика размера фенестрации подтипов натриевых каналов и их доступности для ингибиторов». Биофизический журнал 121.2 (2022): 193-206. https://doi.org/10.1016/j.bpj.2021.12.025
- ^ Ян, Чжэнь и др. «Структура комплекса Nav1.4-β1 электрического угря». Ячейка 170.3 (2017): 470–482. https://doi.org/10.1016/j.cell.2017.06.039
- ^ Jump up to: а б с Исом Л.Л. (февраль 2001 г.). «Бета-субъединицы натриевых каналов: что угодно, кроме вспомогательных». Нейробиолог . 7 (1): 42–54. дои : 10.1177/107385840100700108 . ПМИД 11486343 . S2CID 86422657 .
- ^ Jump up to: а б Каттералл В.А., Голдин А.Л., Ваксман С.Г. (декабрь 2005 г.). «Международный союз фармакологии. XLVII. Номенклатура и структурно-функциональные связи потенциалзависимых натриевых каналов» . Фармакологические обзоры . 57 (4): 397–409. дои : 10.1124/пр.57.4.4 . ПМИД 16382098 . S2CID 7332624 .
- ^ Ноланд, Кэмерон Л.; Чуа, Хан Чоу; Кшонсак, Марк; Хойссер, Стефани Андреа; Браун, Нина; Чанг, Тимоти; Тэм, Кристина; Тан, Цзя; Артур, Кристофер П.; Чиферри, Клаудио; Плесс, Стефан Александр; Паянде, Цзян (17 марта 2022 г.). «Структурно-ориентированное разблокирование NaX обнаруживает неселективный, чувствительный к тетродотоксину катионный канал» . Природные коммуникации . 13 (1): 1416. doi : 10.1038/s41467-022-28984-4 . ПМЦ 8931054 . ПМИД 35301303 .
- ^ Лоссин С. «Информационная база SCN1A» . Архивировано из оригинала 21 июля 2011 г. Проверено 30 октября 2009 г.
компиляция генетических вариаций гена SCN1A, которые изменяют экспрессию или функцию Nav1.1
- ^ Jump up to: а б с д Беннетт Д.Л., Кларк А.Дж., Хуан Дж., Ваксман С.Г., Диб-Хадж С.Д. (апрель 2019 г.). «Роль потенциалзависимых натриевых каналов в передаче сигналов боли» . Физиологические обзоры . 99 (2): 1079–1151. дои : 10.1152/physrev.00052.2017 . ПМИД 30672368 .
- ^ Смит Р.С., Кенни С.Дж., Ганеш В., Джанг А., Борхес-Монрой Р., Партлоу Дж.Н. и др. (сентябрь 2018 г.). «V1.3) Регуляция складок коры головного мозга человека и развитие моторики полости рта» . Нейрон . 99 (5): 905–913.e7. дои : 10.1016/j.neuron.2018.07.052 . ПМК 6226006 . ПМИД 30146301 .
- ^ Чокалингам П., Уайлд А. (сентябрь 2012 г.). «Многогранный сердечный натриевый канал и его клиническое значение». Сердце . 98 (17): 1318–24. doi : 10.1136/heartjnl-2012-301784 . ПМИД 22875823 . S2CID 44433455 .
- ^ Бейдер А., Маццоне А., Стреге П.Р., Тестер DJ, Сайто Я.А., Бернард CE, Эндерс Ф.Т., Эк ВЕ, Шмидт П.Т., Длугош А., Линдберг Г., Карлинг П., Олссон Б., Газули М., Нардоне Г., Куомо Р., Усай- Сатта П., Галеацци Ф., Нери М., Портинкаса П., Беллини М., Барбара Дж., Камиллери М., Локк Г.Р., Талли Нью-Джерси, Д'Амато М., Акерман М.Дж., Фарруджа Дж. (июнь 2014 г.). «Потеря функции потенциалзависимого натриевого канала NaV1,5 (каналопатии) у больных с синдромом раздраженного кишечника» . Гастроэнтерология . 146 (7): 1659–1668. дои : 10.1053/j.gastro.2014.02.054 . ПМК 4096335 . ПМИД 24613995 .
- ^ Батлер К.М., да Силва С., Шафир Ю., Вайсфельд-Адамс Дж.Д., Александр Дж.Дж., Хегде М., Эскайг А. (январь 2017 г.). «De novo и унаследованные мутации эпилепсии SCN8A, обнаруженные с помощью анализа генной панели» . Исследования эпилепсии . 129 : 17–25. doi : 10.1016/j.eplepsyres.2016.11.002 . ПМК 5321682 . ПМИД 27875746 .
- ^ Мейслер М.Х., Кирни Дж.А. (август 2005 г.). «Мутации натриевых каналов при эпилепсии и других неврологических расстройствах» . Журнал клинических исследований . 115 (8): 2010–7. дои : 10.1172/JCI25466 . ПМК 1180547 . ПМИД 16075041 .
- ^ Варгас-Аларкон Дж., Альварес-Леон Э., Фрагосо Х.М., Варгас А., Мартинес А., Вальехо М., Мартинес-Лавин М. (февраль 2012 г.). «Полиморфизм натриевых каналов дорсальных корешковых ганглиев, кодируемый геном SCN9A, связанный с тяжелой фибромиалгией» . BMC Заболевания опорно-двигательного аппарата . 13:23 . дои : 10.1186/1471-2474-13-23 . ПМК 3310736 . ПМИД 22348792 .
- ^ Каттералл, Вашингтон (апрель 2000 г.). «От ионных токов к молекулярным механизмам: структура и функции потенциалзависимых натриевых каналов» . Нейрон . 26 (1): 13–25. дои : 10.1016/S0896-6273(00)81133-2 . ПМИД 10798388 . S2CID 17928749 .
- ^ Исом Л.Л., Де Йонг К.С., Паттон Д.Е., Ребер Б.Ф., Оффорд Дж., Шарбонно Х., Уолш К., Голдин А.Л., Каттералл В.А. (май 1992 г.). «Первичная структура и функциональное выражение субъединицы бета 1 натриевого канала мозга крысы». Наука . 256 (5058): 839–42. Бибкод : 1992Sci...256..839I . дои : 10.1126/science.1375395 . ПМИД 1375395 .
- ^ «Сервер Blackboard недоступен» (PDF) . blackboard.jhu.edu . Проверено 16 июля 2020 г.
- ^ Малхотра Дж.Д., Казен-Гиллеспи К., Хорч М., Исом Л.Л. (апрель 2000 г.). «Бета-субъединицы натриевых каналов опосредуют гомофильную клеточную адгезию и рекрутируют анкирин в точки межклеточного контакта» . Журнал биологической химии . 275 (15): 11383–8. дои : 10.1074/jbc.275.15.11383 . ПМИД 10753953 .
- ^ Малхотра Дж.Д., Купманн М.К., Казен-Гиллеспи К.А., Феттман Н., Хорч М., Исом Л.Л. (июль 2002 г.). «Структурные требования для взаимодействия субъединиц бета 1 натриевого канала с анкирином» . Журнал биологической химии . 277 (29): 26681–8. дои : 10.1074/jbc.M202354200 . ПМИД 11997395 .
- ^ Кантрелл А.Р., Каттералл Вашингтон (июнь 2001 г.). «Нейромодуляция каналов Na+: неожиданная форма клеточной пластичности». Обзоры природы. Нейронаука . 2 (6): 397–407. дои : 10.1038/35077553 . ПМИД 11389473 . S2CID 22885909 .
- ^ Шах Б.С., Раш А.М., Лю С., Тиррел Л., Блэк Дж.А., Диб-Хадж С.Д., Ваксман С.Г. (август 2004 г.). «Контактин связывается с натриевым каналом Nav1.3 в нативных тканях и увеличивает плотность каналов на поверхности клетки» . Журнал неврологии . 24 (33): 7387–99. doi : 10.1523/JNEUROSCI.0322-04.2004 . ПМК 6729770 . ПМИД 15317864 .
- ^ Виттмак Э.К., Раш А.М., Крэнер М.Дж., Гольдфарб М., Ваксман С.Г., Диб-Хадж С.Д. (июль 2004 г.). «Гомологический фактор 2B фактора роста фибробластов: ассоциация с Nav1.6 и селективная колокализация в узлах Ранвье аксонов дорсальных корешков» . Журнал неврологии . 24 (30): 6765–75. doi : 10.1523/JNEUROSCI.1628-04.2004 . ПМК 6729706 . ПМИД 15282281 .
- ^ Раш А.М., Виттмак Э.К., Тиррелл Л., Блэк Дж.А., Диб-Хадж С.Д., Ваксман С.Г. (май 2006 г.). «Дифференциальная модуляция натриевого канала Na (v) 1,6 двумя членами подсемейства гомологичного фактора 2 фактора роста фибробластов». Европейский журнал неврологии . 23 (10): 2551–62. дои : 10.1111/j.1460-9568.2006.04789.x . ПМИД 16817858 . S2CID 21411801 .
- ^ Казаринова-Нойес К., Малхотра Дж.Д., МакИвен Д.П., Маттей Л.Н., Берглунд Э.О., Раншт Б., Левинсон С.Р., Шахнер М., Шрагер П., Исом Л.Л., Сяо З.К. (октябрь 2001 г.). «Контактин связывается с каналами Na+ и увеличивает их функциональную экспрессию» . Журнал неврологии . 21 (19): 7517–25. doi : 10.1523/JNEUROSCI.21-19-07517.2001 . ПМК 6762905 . ПМИД 11567041 .
- ^ Шринивасан Дж., Шахнер М., Каттералл В.А. (декабрь 1998 г.). «Взаимодействие потенциалзависимых натриевых каналов с молекулами внеклеточного матрикса тенасцин-С и тенасцин-R» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (26): 15753–7. Бибкод : 1998PNAS...9515753S . дои : 10.1073/pnas.95.26.15753 . ПМК 28116 . ПМИД 9861042 .
- ^ Ким Д.Ю., Ингано Л.А., Кэри Б.В., Петтингелл В.Х., Ковач Д.М. (июнь 2005 г.). «Пресенилин/гамма-секретаза-опосредованное расщепление бета2-субъединицы потенциалзависимого натриевого канала регулирует клеточную адгезию и миграцию» . Журнал биологической химии . 280 (24): 23251–61. дои : 10.1074/jbc.M412938200 . ПМИД 15833746 .
- ^ Рэтклифф К.Ф., Вестенбрук Р.Э., Кертис Р., Каттералл В.А. (июль 2001 г.). «Субъединицы бета1 и бета3 натриевых каналов связываются с нейрофасцином через свой внеклеточный иммуноглобулинподобный домен» . Журнал клеточной биологии . 154 (2): 427–34. дои : 10.1083/jcb.200102086 . ПМК 2150779 . ПМИД 11470829 .
- ^ Jump up to: а б с Анцелевич С, Паточкай Б (январь 2016 г.). «Синдром Бругада: клинические, генетические, молекулярные, клеточные и ионные аспекты» . Современные проблемы кардиологии . 41 (1): 7–57. doi : 10.1016/j.cpcardiol.2015.06.002 . ПМЦ 4737702 . ПМИД 26671757 .
- ^ Jump up to: а б с д Рен, Децзян (22 декабря 2011 г.). «Каналы утечки натрия в возбудимости нейронов и ритмическом поведении» . Нейрон . 72 (6): 899–911. дои : 10.1016/j.neuron.2011.12.007 . ISSN 1097-4199 . ПМК 3247702 . ПМИД 22196327 .
- ^ Гролло Ф., Станкевич М., Бириньи-Страхан Л., Ван К.С., Николсон Г.М., Пелхате М., Лапьед Б. (февраль 2001 г.). «Электрофизиологический анализ нейротоксического действия токсина воронкообразного паука, дельта-атракотоксина-HV1a, на потенциалзависимые Na+-каналы насекомых». Журнал экспериментальной биологии . 204 (Часть 4): 711–21. дои : 10.1242/jeb.204.4.711 . ПМИД 11171353 .
- ^ Поссани Л.Д., Бесерриль Б., Делепьер М., Титгат Дж. (сентябрь 1999 г.). «Токсины скорпиона, специфичные для Na+-каналов» . Европейский журнал биохимии . 264 (2): 287–300. дои : 10.1046/j.1432-1327.1999.00625.x . ПМИД 10491073 .
- ^ Jump up to: а б Ли, Юнг-Ха; Криббс, Лиэнн Л.; Перес-Рейес, Эдвард (26 февраля 1999 г.). «Клонирование нового белка с четырьмя повторами, связанного с потенциал-управляемыми натриевыми и кальциевыми каналами» . Письма ФЭБС . 445 (2–3): 231–236. дои : 10.1016/S0014-5793(99)00082-4 . ISSN 0014-5793 . ПМИД 10094463 . S2CID 26183219 .
- ^ Jump up to: а б с Лу, Боксун; Су, Яньхуа; Дас, Судипто; Лю, Цзинь; Ся, Цзиншэн; Рен, Децзян (20 апреля 2007 г.). «Нейрональный канал NALCN способствует проницаемости натрия в состоянии покоя и необходим для нормального дыхательного ритма» . Клетка . 129 (2): 371–383. дои : 10.1016/j.cell.2007.02.041 . ISSN 0092-8674 . ПМИД 17448995 . S2CID 17165089 .
- ^ Jump up to: а б Лу, Боксун; Су, Яньхуа; Дас, Судипто; Ван, Хайкунь; Ван, Ян; Лю, Цзинь; Рен, Децзян (5 февраля 2009 г.). «Пептидные нейромедиаторы активируют комплекс катионных каналов NALCN и UNC-80» . Природа . 457 (7230): 741–744. Бибкод : 2009Natur.457..741L . дои : 10.1038/nature07579 . ISSN 1476-4687 . ПМК 2810458 . ПМИД 19092807 .
- ^ Jump up to: а б Суэйн, Ли Энн; Мезграни, Александр; Варро, Энни; Путь, Джон; Бертран, Гислен; Далле, Стефан; Бурине, Эммануэль; Лори, Филипп; Миллер, Ричард Дж.; Наргеот, Джоэл; Монтей, Арно (3 июля 2009 г.). «Ионный канал NALCN активируется мускариновыми рецепторами M3 в линии бета-клеток поджелудочной железы» . Отчеты ЭМБО . 10 (8): 873–880. дои : 10.1038/embor.2009.125 . ISSN 1469-3178 . ПМК 2710536 . ПМИД 19575010 .
- ^ Jump up to: а б Лу, Боксун; Чжан, Ци; Ван, Хайкунь; Ван, Ян; Накаяма, Манабу; Рен, Децзян (04 ноября 2010 г.). «Внеклеточный кальций контролирует фоновый ток и возбудимость нейронов через комплекс катионных каналов UNC79-UNC80-NALCN» . Нейрон . 68 (3): 488–499. дои : 10.1016/j.neuron.2010.09.014 . ISSN 1097-4199 . ПМК 2987630 . ПМИД 21040849 .
- ^ Да, Эдвард; Нг, Шэрон; Чжан, Ми; Бухур, Магали; Ван, Ин; Ван, Мин; Хунг, Уэсли; Аояги, Киота; Мельник-Мартинес, Катя; Ли, Мишель; Лю, Фанг; Шафер, Уильям Р.; Чжэнь, Мэй (11 марта 2008 г.). «Предполагаемый катионный канал NCA-1 и новый белок UNC-80 передают активность нейронов у C. elegans» . ПЛОС Биология . 6 (3): е55. doi : 10.1371/journal.pbio.0060055 . ISSN 1545-7885 . ПМК 2265767 . ПМИД 18336069 .
- ^ Jump up to: а б Хамфри, Джон А.; Хэмминг, Кевин С.; Такер, Колин М.; Скотт, Роберт Л.; Седенски, Маргарет М.; Снутч, Терренс П.; Морган, Фил Г.; Нэш, Ховард А. (3 апреля 2007 г.). «Предполагаемый катионный канал и его новый регулятор: межвидовое сохранение воздействия на общую анестезию» . Современная биология . 17 (7): 624–629. дои : 10.1016/j.cub.2007.02.037 . ISSN 0960-9822 . ПМИД 17350263 . S2CID 9395896 .
- ^ Jump up to: а б Се, Цзюнфан; Кэ, Мэн; Линь, Шии; Чжан, Цзябэй; Ву, Цзяньпин; «Структура утечки натрия у человека» . канал NALCN в комплексе с FAM155A" . Nature Communications . 11 (1): 5831. Бибкод : 2020NatCo..11.5831X . doi : /s41467-020-19667-z . ISSN 2041-1723 . PMC 7672056. 10.1038 PMID 33 203 861 .
- ^ Лир, Бриджит К.; Линь, Цзюй-Мин; Кит, Дж. Рассел; МакГилл, Джермейн Дж.; Раман, Индира М.; Аллада, Рави (2 декабря 2005 г.). «Узкая брюшная полость с ионным каналом имеет решающее значение для работы нейронов циркадного кардиостимулятора дрозофилы» . Нейрон . 48 (6): 965–976. дои : 10.1016/j.neuron.2005.10.030 . ISSN 0896-6273 . ПМИД 16364900 . S2CID 14910947 .
- ^ Аль-Сайед, Моиналдин Д.; Аль-Зайдан, Хамад; Альбахит, Албандари; Хаками, Ханна; Кенана, Розан; Аль-Яфи, Юсра; Аль-Досари, майор; Кари, Аля; Аль-Шедди, Тарфа; Аль-Мухейза, Мухаммед; Аль-Куббай, Вафа; Лакмаче, Ямина; Аль-хинди, хинди; Газиуддин, Мухаммед; Чолак, Дилек (3 октября 2013 г.). «Мутации в NALCN вызывают аутосомно-рецессивный синдром с выраженной гипотонией, нарушениями речи и задержкой когнитивных функций» . Американский журнал генетики человека . 93 (4): 721–726. дои : 10.1016/j.ajhg.2013.08.001 . ISSN 1537-6605 . ПМЦ 3791267 . ПМИД 24075186 .
- ^ Мок, Кин Ю.; Шнайдер, Сюзанна А.; Трабзуни, Дания; Стамелу, Мария; Эдвардс, Марк; Касперавичюте, Даля; Пикеринг-Браун, Стюарт; Сильвердейл, Монти; Харди, Джон; Бхатия, Кайлаш П. (1 февраля 2014 г.). «Общегеномное исследование ассоциации при цервикальной дистонии демонстрирует возможную связь с каналом утечки натрия» . Двигательные расстройства . 29 (2): 245–251. дои : 10.1002/mds.25732 . ISSN 1531-8257 . ПМК 4208301 . ПМИД 24227479 .
- ^ Ван, Кэ-Шэн; Лю, Сюэ-Фэн; Арагам, Нагеш (1 декабря 2010 г.). «Полногеномный метаанализ идентифицирует новые локусы, связанные с шизофренией и биполярным расстройством» . Исследования шизофрении . 124 (1): 192–199. doi : 10.1016/j.schres.2010.09.002 . ISSN 0920-9964 . ПМИД 20889312 . S2CID 21402201 .
- ^ Петерс Ч., Гованлоо М.Р., Гершоме К., Рубен ПК (февраль 2018 г.). «Модуляция pH потенциалзависимых натриевых каналов». Потенциал-управляемые натриевые каналы: структура, функции и каналопатии . Справочник по экспериментальной фармакологии. Том. 246. стр. 147–160. дои : 10.1007/164_2018_99 . ISBN 978-3-319-90283-8 . ПМИД 29460150 .
- ^ Чжан Дж. Ф., Сигельбаум С.А. (декабрь 1991 г.). «Влияние внешних протонов на одиночные сердечные натриевые каналы из миоцитов желудочков морских свинок» . Журнал общей физиологии . 98 (6): 1065–83. дои : 10.1085/jgp.98.6.1065 . ПМК 2229074 . ПМИД 1664454 .
- ^ Jump up to: а б Сунь Ю.М., Фавр И., Шильд Л., Мочидловски Э. (декабрь 1997 г.). «О структурных основах селективного по размеру проникновения органических катионов через потенциал-управляемый натриевый канал. Влияние мутаций аланина в локусе DEKA на селективность, ингибирование Ca2+ и H+ и молекулярное просеивание» . Журнал общей физиологии . 110 (6): 693–715. дои : 10.1085/jgp.110.6.693 . ПМК 2229404 . ПМИД 9382897 .
- ^ Хан А., Романцева Л., Лам А., Липкинд Г., Фоззард Х.А. (август 2002 г.). «Роль карбоксилатов внешнего кольца пор натриевых каналов скелетных мышц крысы в блоке протонов» . Журнал физиологии . 543 (Часть 1): 71–84. дои : 10.1113/jphysicalol.2002.021014 . ПМК 2290475 . ПМИД 12181282 .
- ^ Вилин Ю.Ю., Петерс Ч., Рубен П.С. (2012). «Ацидоз по-разному модулирует инактивацию каналов na(v)1.2, na(v)1.4 и na(v)1.5» . Границы в фармакологии . 3 : 109. дои : 10.3389/fphar.2012.00109 . ПМК 3372088 . ПМИД 22701426 .
- ^ Джонс Д.К., Питерс CH, Аллард CR, Клейдон Т.В., Рубен ПК (февраль 2013 г.). «Протонные сенсоры в поровой области сердечного потенциалзависимого натриевого канала» . Журнал биологической химии . 288 (7): 4782–91. дои : 10.1074/jbc.M112.434266 . ПМК 3576083 . ПМИД 23283979 .
- ^ Хан А., Кайл Дж.В., Хэнк Д.А., Липкинд Г.М., Фоззард Х.А. (октябрь 2006 г.). «Изоформ-зависимое взаимодействие потенциалзависимых натриевых каналов с протонами» . Журнал физиологии . 576 (Часть 2): 493–501. дои : 10.1113/jphysicalol.2006.115659 . ПМК 1890365 . ПМИД 16873405 .
- ^ Хермансен Л., Оснес Дж.Б. (март 1972 г.). «РН крови и мышц после максимальной тренировки у человека». Журнал прикладной физиологии . 32 (3): 304–8. дои : 10.1152/яп.1972.32.3.304 . ПМИД 5010039 .
- ^ Гованлоо М.Р., Абдельсайед М., Петерс Ч., Рубен ПК (апрель 2018 г.). «Мутант P1158S со смешанным периодическим параличом и миотонией придает pH-чувствительность в потенциалзависимых натриевых каналах скелетных мышц» . Научные отчеты . 8 (1): 6304. Бибкод : 2018НатСР...8.6304Г . дои : 10.1038/s41598-018-24719-y . ПМЦ 5908869 . ПМИД 29674667 .
- ^ Гованлоо М.Р., Петерс CH, Рубен ПК (октябрь 2018 г.). «Влияние ацидоза на потенциалзависимые натриевые каналы нейронов: Nav1.1 и Nav1.3» . Каналы . 12 (1): 367–377. дои : 10.1080/19336950.2018.1539611 . ПМК 6284583 . ПМИД 30362397 .
Внешние ссылки
[ редактировать ]- Натрий+каналы в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- «Натриевые каналы, управляемые напряжением» . База данных IUPHAR по рецепторам и ионным каналам . Международный союз фундаментальной и клинической фармакологии. Архивировано из оригинала 9 декабря 2021 г. Проверено 18 декабря 2008 г.