Некоторые виды дрожжей обладают способностью развивать многоклеточные характеристики, образуя цепочки связанных почкующихся клеток, известных как псевдогифы или ложные гифы , или быстро эволюционировать в многоклеточный кластер со специализированными функциями клеточных органелл . [5] [6] Размеры дрожжей сильно различаются в зависимости от вида и окружающей среды, обычно они составляют 3–4 мкм в диаметре , хотя некоторые дрожжи могут вырасти до 40 мкм. [7] Большинство дрожжей размножаются бесполым путем путем митоза , а многие делают это с помощью асимметричного процесса деления, известного как почкование . По своей одноклеточной природе дрожжи можно противопоставить плесневым грибам , у которых растут гифы . Виды грибов, способные принимать обе формы (в зависимости от температуры или других условий), называются диморфными грибами .
Слово «дрожжи» происходит от древнеанглийского gist , gyst и от индоевропейского корня yes- , означающего «кипятить», «пениться» или «пузырь». [13] Дрожжевые микробы, вероятно, являются одними из самых ранних одомашненных организмов. Археологи, раскапывающие египетские руины, обнаружили ранние точильные камни и камеры для выпечки дрожжевого хлеба, а также рисунки 4000-летних пекарен и пивоваренных заводов . [14] Было обнаружено, что сосуды, изученные на нескольких археологических памятниках в Израиле (датируемые примерно 5000, 3000 и 2500 лет назад), в которых, как считалось, содержались алкогольные напитки ( пиво и медовуха ), содержали колонии дрожжей, которые выживали на протяжении тысячелетий, обеспечивая первые прямые биологические доказательства использования дрожжей в ранних культурах. [15] В 1680 году голландский натуралист Антон ван Левенгук впервые наблюдал под микроскопом дрожжи, но в то время не считал их живыми организмами , а скорее шаровидными структурами. [16] поскольку исследователи сомневались, являются ли дрожжи водорослями или грибами. [17] Теодор Шванн признал их грибами в 1837 году. [18] [19]
В 1857 году французский микробиолог Луи Пастер показал, что, пропуская кислород в дрожжевой бульон, можно увеличить рост клеток , но замедлить брожение – это наблюдение позже назвали « эффектом Пастера ». В статье « Мемуар о алкогольном брожении » Пастер доказал, что алкогольное брожение осуществляется живыми дрожжами, а не химическим катализатором. [14] [20]
К концу 18 века были идентифицированы два штамма дрожжей, используемых в пивоварении: Saccharomyces cerevisiae (дрожжи верхового брожения) и S. пасторианус (дрожжи низового брожения). S. cerevisiae продается голландцами на коммерческой основе для выпечки хлеба с 1780 года; а около 1800 года немцы начали производить S. cerevisiae в форме крема. В 1825 году был разработан метод удаления жидкости, позволяющий готовить дрожжи в виде твердых блоков. [21] Промышленное производство дрожжевых блоков было расширено с появлением фильтр-пресса в 1867 году. В 1872 году барон Макс де Спрингер разработал производственный процесс создания гранулированных дрожжей из свекольной патоки . [22] [23] [24] метод, который использовался до первой мировой войны. [25] В Соединенных Штатах дрожжи, передающиеся по воздуху естественным путем, использовались почти исключительно до тех пор, пока коммерческие дрожжи не появились на рынке на выставке Centennial Exposition в 1876 году в Филадельфии, где Чарльз Л. Флейшманн продемонстрировал продукт и процесс его использования, а также подал выпеченный хлеб. . [26]
Механический холодильник (впервые запатентованный в 1850-х годах в Европе) освободил пивоваров и виноделов впервые от сезонных ограничений и позволил им покинуть подвалы и другие земляные помещения. Для Джона Молсона зарабатывал на жизнь в Монреале , который до разработки холодильника , сезон пивоварения длился с сентября по май. Те же сезонные ограничения раньше действовали в сфере винокуренного искусства. [27]
Дрожжи различаются в зависимости от температурного диапазона, в котором они лучше всего растут. Например, Leucosporidium frigidum растет при температуре от -2 до 20 °C (от 28 до 68 °F), Saccharomycestelluris при температуре от 5 до 35 °C (от 41 до 95 °F), а Candida slooffi при температуре от 28 до 45 °C (от 82 до 113 °C). °Ф). [29] Клетки могут пережить замораживание при определенных условиях, при этом жизнеспособность со временем снижается.
Появление белых нитевидных дрожжей, широко известных как дрожжи кам , часто является побочным продуктом лактоферментации (или маринования) некоторых овощей. Обычно это результат воздействия воздуха. Хотя он и безвреден, он может придать маринованным овощам неприятный привкус, и его необходимо регулярно удалять во время брожения. [30]
Экология
Дрожжи очень распространены в окружающей среде и часто выделяются из материалов, богатых сахаром. Примеры включают встречающиеся в природе дрожжи на кожуре фруктов и ягод (например, винограда, яблок или персиков ) и экссудаты растений (например, соки растений или кактусов). Некоторые дрожжи встречаются в почве и насекомых. [31] [32] Было показано, что дрожжи из почвы и кожуры фруктов и ягод доминируют в грибковой сукцессии во время гниения фруктов. [33] Экологическая функция и биоразнообразие дрожжей относительно неизвестны по сравнению с таковыми других микроорганизмов . [34] Дрожжевые грибки, в том числе Candida albicans , Rhodotorula Rubra , Torulopsis и Trichosporon Cutaneum , обитают между пальцами ног людей как часть их кожной флоры . [35] Дрожжи также присутствуют во флоре кишечника млекопитающих и некоторых насекомых. [36] и даже в глубоководной среде обитает множество дрожжей. [37] [38]
Индийское исследование семи видов пчел и девяти видов растений показало, что 45 видов из 16 родов колонизируют нектарники цветов и медовые желудки пчел. Большинство из них были представителями рода Candida ; наиболее распространенным видом в медовых желудках была Dekkera intermedia , а в нектарниках цветков - Candida Blankii . [39] Было обнаружено, что дрожжи, колонизирующие нектарники чемерицы вонючей, повышают температуру цветка, что может способствовать привлечению опылителей за счет увеличения испарения летучих органических соединений . [34] [40] были Черные дрожжи зарегистрированы как партнер в сложных отношениях между муравьями , их мутуалистическим грибом , грибковым паразитом гриба и бактерией, убивающей паразита. Дрожжи оказывают негативное воздействие на бактерии, которые обычно производят антибиотики для уничтожения паразита, поэтому могут повлиять на здоровье муравьев, позволяя паразиту распространяться. [41]
Определенные штаммы некоторых видов дрожжей производят белки, называемые токсинами-убийцами дрожжей, которые позволяют им устранять конкурирующие штаммы. (См. основную статью о дрожжах-убийцах .) Это может вызвать проблемы в виноделии, но потенциально также может быть использовано с пользой, используя штаммы, продуцирующие токсин-киллер, для производства вина. Токсины-убийцы дрожжей также могут иметь медицинское применение при лечении дрожжевых инфекций (см. раздел «Патогенные дрожжи» ниже). [42]
Морские дрожжи, определяемые как дрожжи, выделенные из морской среды, способны лучше расти на среде, приготовленной с использованием морской, а не пресной воды. [43] Первые морские дрожжи были выделены Бернхардом Фишером в 1894 году из Атлантического океана и идентифицированы как Torula sp. и Mycoderma sp. [44] После этого открытия по всему миру были выделены различные другие морские дрожжи из разных источников, включая морскую воду, морские водоросли, морскую рыбу и млекопитающих. [45] Среди этих изолятов некоторые морские дрожжи произошли из наземных местообитаний (сгруппированы как факультативные морские дрожжи), которые были завезены в морскую среду и выжили в ней. Остальные морские дрожжи были сгруппированы как облигатные или аборигенные морские дрожжи, обитающие только в морской среде обитания. [44] Однако не было найдено достаточных доказательств, объясняющих незаменимость морской воды для облигатных морских дрожжей. [43] Сообщалось, что морские дрожжи способны продуцировать множество биологически активных веществ, таких как аминокислоты, глюканы, глутатион, токсины, ферменты, фитаза и витамины, которые могут применяться в пищевой, фармацевтической, косметической и химической промышленности, а также для морская культура и охрана окружающей среды. [43] Морские дрожжи успешно использовались для производства биоэтанола с использованием среды на основе морской воды, что потенциально позволит снизить водный след биоэтанола. [46]
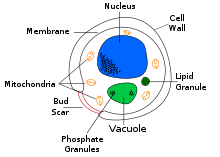
Дрожжи, как и все грибы, могут иметь бесполый и половой репродуктивный цикл. Наиболее распространенным способом вегетативного роста дрожжей является бесполое размножение почкованием . [47] небольшая почка (также известная как пузырь где на родительской клетке формируется или дочерняя клетка). Ядро . материнской клетки распадается на дочернее ядро и мигрирует в дочернюю клетку Затем почка продолжает расти, пока не отделится от родительской клетки, образуя новую клетку. [48] Дочерняя клетка, образующаяся в процессе почкования, обычно меньше материнской клетки. Некоторые дрожжи, в том числе Schizosaccharomyces pombe , размножаются делением, а не почкованием. [47] и тем самым создавая две дочерние клетки одинакового размера.
Гаплоидные делящиеся дрожжи Schizosaccharomyces pombe - это факультативный половой микроорганизм, который может спариваться при ограничении питательных веществ. [3] [50] Воздействие на S. pombe перекиси водорода, агента, вызывающего окислительный стресс, приводящий к окислительному повреждению ДНК, сильно индуцирует спаривание и образование мейотических спор. [51] Почкующиеся дрожжи Saccharomyces cerevisiae размножаются митозом в виде диплоидных клеток при избытке питательных веществ, но при голодании эти дрожжи подвергаются мейозу с образованием гаплоидных спор. [52] Гаплоидные клетки могут затем размножаться бесполым путем путем митоза. Кац Эзов и др. [53] представили данные о том, что в природных популяциях S. cerevisiae преобладают клональное размножение и самоопыление (в форме интратетрадного спаривания). В природе спаривание гаплоидных клеток с образованием диплоидных клеток чаще всего происходит между членами одной и той же клональной популяции, и ауткроссинг встречается редко. [54] Анализ происхождения природных штаммов S. cerevisiae привел к выводу, что ауткроссинг происходит только примерно один раз на каждые 50 000 клеточных делений. [54] Эти наблюдения позволяют предположить, что возможные долгосрочные выгоды от ауткроссинга (например, создание разнообразия), вероятно, будут недостаточными для общего сохранения пола от одного поколения к другому. [ нужна ссылка ] Скорее, краткосрочная выгода, такая как рекомбинационная репарация во время мейоза, [55] может быть ключом к поддержанию пола у S. cerevisiae .
Алкогольные напитки определяются как напитки , содержащие этанол (C 2 H 5 OH). Этот этанол почти всегда производится путем ферментации – метаболизма углеводов . определенными видами дрожжей в анаэробных условиях или условиях с низким содержанием кислорода В таких напитках, как медовуха, вино, пиво или спиртные напитки , на определенном этапе производства используются дрожжи. Дистиллированный напиток — это напиток, содержащий этанол, очищенный перегонкой . Растительный материал, содержащий углеводы, ферментируется дрожжами, при этом образуется разбавленный раствор этанола. Такие спиртные напитки, как виски и ром , получают путем перегонки разбавленных растворов этанола. В конденсате собираются другие компоненты, кроме этанола, включая воду, сложные эфиры и другие спирты, которые (в дополнение к тем, которые обеспечиваются дубом, в котором он может выдерживаться) определяют вкус напитка .
Дрожжевое кольцо, использовавшееся шведскими фермерскими пивоварами в 19 веке для сохранения дрожжей между варками. Пузырьки углекислого газа, образующиеся во время пивоварения [9]
Пивные дрожжи можно разделить на «верхнего брожения» (или «верхового брожения») и «низового брожения» (или «низового брожения»). [59] Верхние дрожжи называются так потому, что во время брожения они образуют пену в верхней части сусла . Примером лучших дрожжей являются Saccharomyces cerevisiae , иногда называемые «элевыми дрожжами». [60] Нижние дрожжи обычно используются для производства пива типа лагер , хотя они также могут производить пиво типа эля . Эти дрожжи хорошо ферментируются при низких температурах. Примером дрожжей с нижней культурой являются Saccharomyces Pastorianus , ранее известные как S. Carlsbergensis .
Десятилетия назад, [ нечеткий ] систематики реклассифицировали S. carlsbergensis (uvarum) как члена S. cerevisiae , отметив, что единственное явное различие между ними - метаболическое. [ сомнительно – обсудить ] Лагерные штаммы S. cerevisiae выделяют фермент мелибиазу, позволяющий им гидролизовать мелибиозу , дисахарид , в более ферментируемые моносахариды . Различия между верхними и нижними культурами, а также холодным и теплым брожением в основном являются обобщениями, используемыми непрофессионалами для общения с широкой публикой. [61]
Наиболее распространенные пивные дрожжи высшего сорта, S. cerevisiae , относятся к тому же виду, что и обычные хлебопекарные дрожжи. [62] Пивные дрожжи также очень богаты необходимыми минералами и витаминами группы B (за исключением B 12 ), что используется в пищевых продуктах, изготовленных из остатков ( побочных ) дрожжей от пивоварения. [63] Однако хлебопекарные и пивные дрожжи обычно принадлежат к разным штаммам, культивируемым с разными характеристиками: штаммы хлебопекарных дрожжей более агрессивны и карбонизуют тесто в кратчайшие сроки; штаммы пивных дрожжей действуют медленнее, но имеют тенденцию производить меньше посторонних привкусов и переносят более высокие концентрации алкоголя (некоторые штаммы до 22%).
Dekkera/Brettanomyces – это род дрожжей, известный своей важной ролью в производстве ламбиков и специальных кислых элей , а также вторичной обработки определенного бельгийского траппистского пива . [64] Таксономия рода Brettanomyces обсуждается с момента его раннего открытия и за прошедшие годы претерпела множество реклассификаций. Ранняя классификация была основана на нескольких видах, которые размножались бесполым путем (анаморфная форма) посредством мультиполярного почкования. [65] Вскоре после этого было обнаружено образование аскоспор и род Dekkera , размножающийся половым путем (телеоморфная форма). в состав систематики был введен [66] Текущая систематика включает пять видов родов Dekkera/Brettanomyces . Это анаморфы Brettanomyces bruxellensis , Brettanomyces anomalus , Brettanomyces custersianus , Brettanomyces naardenensis и Brettanomyces nanus , причем телеоморфы существуют для первых двух видов, Dekkera bruxellensis и Dekkera anomala . [67] Различие между Dekkera и Brettanomyces спорно: Oelofse et al. (2008) цитируют Лоурейро и Малфейто-Феррейру из 2006 года, когда они подтвердили, что современные методы обнаружения молекулярной ДНК не выявили различий между состояниями анаморфа и телеоморфа. За последнее десятилетие Brettanomyces spp. получили все более широкое применение в секторе крафтового пивоварения: несколько пивоварен производили пиво, которое в основном ферментировалось чистыми культурами Brettanomyces spp. Это произошло в результате экспериментов, поскольку существует очень мало информации о ферментативных способностях чистой культуры и ароматических соединениях, продуцируемых различными штаммами. Деккера / Brettanomyces spp. были предметом многочисленных исследований, проведенных за последнее столетие, хотя большинство недавних исследований было сосредоточено на расширении знаний о винодельческой промышленности. Недавние исследования восьми штаммов Brettanomyces , доступных в пивоваренной промышленности, были сосредоточены на ферментации конкретных штаммов и выявили основные соединения, образующиеся во время анаэробной ферментации чистой культуры в сусле. [68]
Дрожжи используются в виноделии ) сахара ( глюкозу и фруктозу , где они преобразуют присутствующие в виноградном соке ( сусле ) в этанол. Дрожжи обычно уже присутствуют на кожуре винограда. Ферментацию можно проводить с помощью этих эндогенных «диких дрожжей». [69] но эта процедура дает непредсказуемые результаты, которые зависят от конкретных типов присутствующих видов дрожжей. По этой причине в сусло обычно добавляют чистую культуру дрожжей; эти дрожжи быстро доминируют в процессе брожения. Дикие дрожжи подавляются, что обеспечивает надежную и предсказуемую ферментацию. [70]
Большинство добавляемых винных дрожжей представляют собой штаммы S. cerevisiae , хотя не все штаммы этого вида подходят. [70] Различные штаммы дрожжей S. cerevisiae обладают разными физиологическими и ферментативными свойствами, поэтому выбранный штамм дрожжей может иметь прямое влияние на готовое вино. [71] Были проведены значительные исследования по разработке новых штаммов винных дрожжей, которые создают нетипичные вкусовые характеристики или повышают сложность вин. [72] [73]
Рост некоторых дрожжей, таких как Zygosaccharomyces и Brettanomyces , в вине может привести к дефектам вина и последующей порче. [74] Brettanomyces при выращивании в вине производит множество метаболитов , некоторые из которых представляют собой летучие фенольные соединения. Вместе эти соединения часто называют « характером Brettanomyces » и часто описывают как « антисептические » или «ароматы скотного двора». Brettanomyces вносят значительный вклад в возникновение дефектов вина в винодельческой промышленности. [75]
Исследователи из Университета Британской Колумбии , Канада, обнаружили новый штамм дрожжей, в котором снижено содержание аминов . Амины в красном вине и Шардоне вызывают неприятный привкус, а у некоторых людей вызывают головные боли и гипертонию. Около 30% людей чувствительны к биогенным аминам, таким как гистамин . [76]
Дрожжи, чаще всего S. cerevisiae , используются в выпечке в качестве разрыхлителя , превращая сбраживаемые сахара, присутствующие в тесте, в углекислый газ . Это приводит к тому, что тесто расширяется или поднимается, поскольку газ образует карманы или пузырьки. Когда тесто выпекается, дрожжи погибают и воздушные карманы «схватываются», придавая выпечке мягкую и рыхлую текстуру. Использование картофеля, воды от варки картофеля, яиц или сахара в хлебном тесте ускоряет рост дрожжей. Большинство дрожжей, используемых в выпечке, относятся к тем же видам, что и при алкогольном брожении. Кроме того, для выпечки иногда используют Saccharomyces exiguus (также известный как S.minor ), дикие дрожжи, обнаруженные в растениях, фруктах и зерновых. При выпечке хлеба дрожжи первоначально дышат аэробно, выделяя углекислый газ и воду. Когда кислород иссякает, начинается брожение , в результате которого в качестве побочного продукта образуется этанол; однако он испаряется во время выпечки. [77]
Блок прессованных свежих дрожжей
Неизвестно, когда дрожжи впервые были использованы для выпечки хлеба. Первые записи, свидетельствующие об этом использовании, пришли из Древнего Египта . [8] Исследователи предполагают, что в теплый день смесь мучной муки и воды оставляли дольше, чем обычно, а дрожжи, которые содержатся в естественных примесях муки, вызвали ее брожение перед выпечкой. Получившийся хлеб был бы легче и вкуснее обычного плоского твердого пирога.
Активные сухие дрожжи, гранулированная форма, в которой дрожжи продаются на коммерческой основе.
Сегодня существует несколько розничных продавцов пекарских дрожжей; Одной из первых разработок в Северной Америке были дрожжи Флейшмана , созданные в 1868 году. Во время Второй мировой войны компания Флейшманн разработала гранулированные активные сухие дрожжи, которые не требовали охлаждения, имели более длительный срок хранения, чем свежие дрожжи, и росли в два раза быстрее. Пекарские дрожжи также продаются в виде свежих дрожжей, спрессованных в квадратный «пирог». Эта форма быстро портится, поэтому ее необходимо использовать вскоре после производства. Слабый раствор воды и сахара можно использовать, чтобы определить, просрочены ли дрожжи. [78] В растворе активные дрожжи будут пениться и пузыриться, ферментируя сахар в этанол и углекислый газ. В некоторых рецептах это называется проверкой дрожжей, поскольку она «доказывает» (проверяет) жизнеспособность дрожжей перед добавлением других ингредиентов. При использовании закваски вместо сахара добавляют муку и воду; это называется расстойкой губки . [ нужна ссылка ]
Когда дрожжи используются для приготовления хлеба, их смешивают с мукой , солью и теплой водой или молоком. Тесто замешивают до тех пор, пока оно не станет гладким, а затем оставляют его подниматься, иногда до тех пор, пока оно не увеличится вдвое. Затем из теста формируют буханки. Некоторое тесто для хлеба отбрасывают после одного подъема и оставляют еще раз подняться (это называется расстойкой теста ), а затем выпекают. Более длительное время подъема дает лучший вкус, но дрожжи могут не поднять хлеб на заключительных стадиях, если изначально оставить его на слишком долгое время. [ нужна ссылка ]
Способность дрожжей превращать сахар в этанол была использована биотехнологической промышленностью для производства этанольного топлива . Процесс начинается с измельчения сырья, такого как сахарный тростник , полевая кукуруза или другие зерновые культуры , а затем добавления разбавленной серной кислоты или грибковых ферментов альфа -амилазы для расщепления крахмалов на сложные сахара. Затем добавляется глюкоамилаза, которая расщепляет сложные сахара на простые. После этого добавляют дрожжи для преобразования простых сахаров в этанол, который затем отгоняют с получением этанола чистотой до 96%. [85]
Дрожжи Saccharomyces были генетически модифицированы для ферментации ксилозы , одного из основных сбраживаемых сахаров, присутствующих в целлюлозной биомассе , такой как сельскохозяйственные отходы, бумажные отходы и древесная щепа. [86] [87] Такая разработка означает, что этанол можно эффективно производить из более дешевого сырья, что делает целлюлозное топливо на основе этанола более конкурентоспособной альтернативой бензиновому топливу. [88]
Ряд сладких газированных напитков можно производить теми же методами, что и пиво, за исключением того, что брожение прекращается раньше, в результате чего образуется углекислый газ, а алкоголя остается лишь следы, оставляя в напитке значительное количество остаточного сахара.
Дрожжевой экстракт , полученный из внутриклеточного содержимого дрожжей и используемый в качестве пищевых добавок или ароматизаторов . Общим методом производства дрожжевого экстракта для пищевых продуктов, таких как Vegemite и Marmite , в промышленных масштабах является тепловой автолиз , то есть добавление соли к суспензии дрожжей, что делает раствор гипертоническим, что приводит к сморщиванию клеток. Это запускает аутолиз дрожжей , при котором пищеварительные ферменты расщепляют их собственные белки на более простые соединения – процесс самоуничтожения. Затем умирающие дрожжевые клетки нагревают для завершения их распада, после чего удаляют шелуху (дрожжи с толстыми клеточными стенками, которые могут дать плохую текстуру). Дрожжевые автолизаты используются в компаниях Vegemite и Promite (Австралия); Мармит (Соединенное Королевство); несвязанный с ним Мармит (Новая Зеландия); Витам-Р (Германия); и Сеновис ( Швейцария ).
Пищевые дрожжевые хлопья желтого цвета. Пищевые дрожжи , представляющие собой цельные высушенные, деактивированные дрожжевые клетки, обычно S. cerevisiae . Обычно он имеет форму желтых хлопьев или порошка, а его ореховый вкус и вкус умами делают его веганским заменителем сырного порошка. [93] Еще одно популярное использование — в качестве начинки для попкорна. Его также можно использовать в пюре и жареном картофеле, а также в яичнице . Он выпускается в виде хлопьев или желтого порошка, по текстуре напоминающего кукурузную муку . В Австралии его иногда продают как «острые дрожжевые хлопья». [94]
Оба типа дрожжевых продуктов, описанных выше, богаты витаминами группы B (кроме витамина B 12, если он не обогащен). [63] что делает их привлекательной пищевой добавкой для веганов. [93] Эти же витамины содержатся и в некоторых упомянутых выше дрожжевых продуктах, например в квасе . [95] Пищевые дрожжи, в частности, содержат мало жира и натрия и являются источником белка и витаминов, а также других минералов и кофакторов, необходимых для роста. Многие марки пищевых дрожжей и спредов с дрожжевыми экстрактами, хотя и не все, обогащены витамином B 12 , который вырабатывается бактериями отдельно . [96]
В 1920 году компания Fleischmann Yeast Company начала продвигать дрожжевые лепешки в рамках кампании «Дрожи для здоровья». Первоначально они подчеркивали важность дрожжей как источника витаминов, полезных для кожи и пищеварения. Их более поздняя реклама заявляла о гораздо более широком спектре преимуществ для здоровья и была осуждена Федеральной торговой комиссией как вводящая в заблуждение . Увлечение дрожжевыми пирожными продолжалось до конца 1930-х годов. [97]
Дрожжи часто используются аквариумистами для выработки углекислого газа (CO 2 ) для питания растений в аквариумах с растениями . [102] Уровни CO 2 от дрожжей регулировать труднее, чем от систем с CO 2 под давлением . Однако низкая стоимость дрожжей делает их широко используемой альтернативой. [102]
24 апреля 1996 года было объявлено, что S. cerevisiae является первым эукариотом, чей геном , состоящий из 12 миллионов пар оснований , был полностью секвенирован в рамках проекта «Геном» . [106] В то время это был самый сложный организм, для которого потребовалось семь лет работы и участие более 100 лабораторий. [107] Вторым видом дрожжей, геном которого был секвенирован, был Schizosaccharomyces pombe , работа над которым была завершена в 2002 году. [108] [109] Это был шестой секвенированный эукариотический геном, состоящий из 13,8 миллионов пар оснований. По состоянию на 2014 год геномы более 50 видов дрожжей были секвенированы и опубликованы. [110]
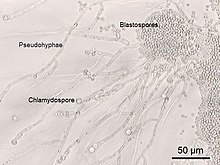
Окраска по Граму Candida albicans из влагалищного мазка. Мелкие овальные хламидоспоры 2–4 мкм . диаметром Микрофотография , Candida albicans показывающая выросты гиф и другие морфологические характеристики.
Дрожжи способны расти в продуктах с низким pH (5,0 или ниже) и в присутствии сахаров, органических кислот и других легко метаболизируемых источников углерода. [123] Во время роста дрожжи метаболизируют некоторые пищевые компоненты и производят конечные продукты метаболизма. Это приводит к изменению физических, химических и сенсорных свойств пищи, и она портится. [124] Рост дрожжей в пищевых продуктах часто наблюдается на их поверхности, например, в сырах или мясе, или в результате ферментации сахара в напитках, таких как соки, и полужидких продуктах, таких как сиропы и джемы . [123] Дрожжи рода Zygosaccharomyces уже давно используются в пищевой промышленности как вызывающие порчу дрожжи . Это происходит главным образом потому, что эти виды могут расти в присутствии высоких концентраций сахарозы, этанола, уксусной кислоты , сорбиновой кислоты , бензойной кислоты и диоксида серы . [74] представляющие некоторые из широко используемых методов консервирования пищевых продуктов . Метиленовый синий используется для проверки наличия живых дрожжевых клеток. [125] В энологии основными дрожжами, вызывающими порчу, являются Brettanomyces bruxellensis.
Индийское исследование семи видов пчел и девяти видов растений показало, что 45 видов дрожжей из 16 родов колонизируют нектарники цветов и медовые желудки пчел. Большинство из них были представителями рода Candida ; Наиболее распространенным видом в желудках медоносных пчел был Dekkera intermedia , а наиболее распространенным видом, колонизирующим нектарники цветков, был Candida Blankii . Хотя механизм не до конца понятен, было обнаружено, что A. indica цветет сильнее, если Candida Blankii . присутствует [39]
В другом примере Spathaspora passalidarum , обнаруженная в пищеварительном тракте жуков-бессов , способствует перевариванию растительных клеток путем ферментации ксилозы . [127]
Многие фрукты производят различные виды сахара, которые привлекают дрожжи, которые ферментируют сахар и превращают его в спирт. Млекопитающие, питающиеся фруктами, находят запах алкоголя привлекательным, поскольку он указывает на спелые, сладкие фрукты, которые обеспечивают больше питательных веществ. В свою очередь, млекопитающие помогают распространять как семена фруктов, так и споры дрожжей. [128] [129]
У дрожжей и маленького ульевого жука мутуалистические отношения. В то время как маленького жука-улья привлекает феромон, выделяемый пчелой-хозяином, дрожжи могут производить аналогичный феромон, который оказывает такое же привлекательное воздействие на маленького жука-улья. Следовательно, дрожжи способствуют заражению SHB, если внутри улья есть дрожжи. [130]
^ Пишкур, Юре; Компаньо, Кончетта (2014). Молекулярные механизмы метаболизма углерода у дрожжей . Спрингер. п. 98. ИСБН 978-3-642-55013-3 . Второй полностью секвенированный дрожжевой геном появился 6 лет спустя от делящихся дрожжей Schizosaccharomyces pombe , которые отделились от S. cerevisiae , вероятно, более 300 миллионов лет назад.
^ Курцман К.П., Пишкур Дж. (2006). «Таксономия и филогенетическое разнообразие дрожжей». В Суннерхаген П., Пискур Дж. (ред.). Сравнительная геномика: использование грибов в качестве моделей . Темы современной генетики. Том. 15. Берлин: Шпрингер. стр. 29–46. дои : 10.1007/b106654 . ISBN 978-3-540-31480-6 .
^ Курцман К.П., Фелл Дж.В. (2005). Габор П., де ла Роза К.Л. (ред.). Биоразнообразие и экофизиология дрожжей . Справочник по дрожжам. Берлин: Шпрингер. стр. 11–30. ISBN 978-3-540-26100-1 .
^ «Что такое дрожжи?» . Виртуальная библиотека дрожжей . 13 сентября 2009 г. Архивировано из оригинала 26 февраля 2009 г. . Проверено 28 ноября 2009 г.
^ Летурно, Ф.; Вилла, П. (1987). «Рост дрожжей Saccharomyces на свекловичной патоке, влияние концентрации субстрата на токсичность алкоголя». Биотехнологические письма . 9 : 53–58. дои : 10.1007/BF01043394 .
^ Миниак (де) М., 1988. Управление цехами спиртового брожения сахарных продуктов (патоки и сточных вод). Пищевая и сельскохозяйственная промышленность 105, 675-688.
^ Бердселл Дж. А., Уиллс С. (2003). Макинтайр Р.Дж., Клегг М.Т. (ред.). Эволюционное происхождение и поддержание половой рекомбинации: обзор современных моделей . Серия «Эволюционная биология» >> Эволюционная биология. Том. 33. Спрингер. стр. 27–137. ISBN 978-0306472619 .
^ Бай Ф.Ю., Чжао Дж.Х., Такашима М., Цзя Дж.Х., Бёкхаут Т., Накасе Т. (2002). «Реклассификация комплексов Sporobolomyces roseus и Sporidiobolus pararoseus с описанием Sporobolomyces phaffii sp. nov». Международный журнал систематической и эволюционной микробиологии . 52 (6): 2309–2314. дои : 10.1099/00207713-52-6-2309 . ПМИД 12508902 .
^ Дополнительную информацию о таксономических различиях см. Доуханик ТМ (1999). «Дрожжи – штаммы и методы обработки». В McCabe JT (ред.). Практичный пивовар . Ассоциация мастеров-пивоваров Америки.
^ MTJ Кастера (1940). Исследование рода дрожжей Brettanomyces (кандидатская диссертация) (на голландском языке). Делфт, Нидерланды: Делфтский университет .
^ Ван дер Вальт, JP (1984). «Деккера 'ван дер Вальт' ». В Крегер-ван Рей, Нью-Джерси (ред.). Дрожжи: таксономическое исследование (3-е изд.). Амстердам: Elsevier Science . стр. 146–150. дои : 10.1016/C2009-0-00558-3 .
^ Макбрайд С., Гарднер Дж. М., де Баррос Лопес М., Йиранек В. (2006). «Поколение новых штаммов винных дрожжей путем адаптивной эволюции». Американский журнал энологии и виноградарства . 57 (4): 423–430. дои : 10.5344/aev.2006.57.4.423 . S2CID 83723719 .
^ Херли Р., де Лувуа Дж., Малхолл А. (1987). «Дрожжи как патогены человека и животных». В Rose AH, Харрисон Дж.С. (ред.). Дрожжи. Том 1: Биология дрожжей (2-е изд.). Нью-Йорк, Нью-Йорк: Академическая пресса . стр. 207–281.
^ Jump up to: а б Курцман КП (2006). «Методы обнаружения, идентификации и подсчета дрожжей, вызывающих порчу». В Блэкберне CDW (ред.). Микроорганизмы, вызывающие порчу пищевых продуктов . Кембридж, Англия: Издательство Woodhead . стр. 28–54. ISBN 978-1-85573-966-6 .
^ Флот Г.Х., Прафаилонг В. (2001). «Дрожжи». В Мойре CJ (ред.). Порча обработанных пищевых продуктов: причины и диагностика . Группа пищевой микробиологии Австралийского института пищевых наук и технологий (AIFST). стр. 383–397. ISBN 978-0-9578907-0-1 .
Arc.Ask3.Ru Номер скриншота №: 239cf69c16466bf2be3da9f26bbe4927__1722676380 URL1:https://arc.ask3.ru/arc/aa/23/27/239cf69c16466bf2be3da9f26bbe4927.html Заголовок, (Title) документа по адресу, URL1: Yeast - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)














 Эта статья содержит цитаты из этого источника, доступного по лицензии Creative Commons Attribution.
Эта статья содержит цитаты из этого источника, доступного по лицензии Creative Commons Attribution.

