Ацетилхолин
Эта статья нуждается в дополнительных цитатах для проверки . ( август 2019 г. ) |
 | |
| Клинические данные | |
|---|---|
| Другие имена | АЧ |
| код АТС | |
| Физиологические данные | |
| Исходные ткани | мотонейроны , парасимпатическая нервная система , головной мозг |
| Целевые ткани | скелетные мышцы , мозг, многие другие органы |
| Рецепторы | никотиновая , мускариновая |
| Агонисты | никотин , мускарин , ингибиторы холинэстеразы |
| Антагонисты | тубокурарин , атропин |
| Предшественник | холин , ацетил-КоА |
| Биосинтез | холин ацетилтрансфераза |
| Метаболизм | ацетилхолинэстераза |
| Идентификаторы | |
| Номер CAS | |
| ПабХим CID | |
| ИЮФАР/БПС | |
| Лекарственный Банк | |
| ХимическийПаук | |
| НЕКОТОРЫЙ | |
| КЕГГ | |
| ЧЭБИ | |
| ЧЕМБЛ | |
| номер Е | E1001(i) (дополнительные химические вещества) |
| Панель управления CompTox ( EPA ) | |
| Информационная карта ECHA | 100.000.118 |
| Химические и физические данные | |
| Формула | С 7 Н 16 Н О 2 |
| Молярная масса | 146.210 g·mol −1 |
Ацетилхолин ( АХ ) — органическое соединение , которое действует в мозге и теле многих видов животных (включая человека) в качестве нейромедиатора . [1] Его название происходит от его химической структуры: это сложный эфир уксусной кислоты и холина . [2] Части тела, которые используют ацетилхолин или подвергаются его воздействию, называются холинергическими .
Ацетилхолин — это нейротрансмиттер, используемый в нервно-мышечных соединениях , другими словами, это химическое вещество, которое выделяют мотонейроны нервной системы для активации мышц. Это свойство означает, что препараты, влияющие на холинергические системы, могут иметь очень опасные эффекты — от паралича до судорог. Ацетилхолин также является нейромедиатором в вегетативной нервной системе , как внутренним передатчиком симпатической нервной системы , так и конечным продуктом, выделяемым парасимпатической нервной системой . [1] Ацетилхолин является основным нейромедиатором парасимпатической нервной системы. [2] [3]
В мозге ацетилхолин действует как нейромедиатор и нейромодулятор . Мозг содержит ряд холинергических областей, каждая из которых выполняет различные функции; например, играет важную роль в возбуждении , внимании , памяти и мотивации . [4]
Ацетилхолин также был обнаружен в клетках неневрального происхождения, а также в микробах. Недавно ферменты, связанные с его синтезом, деградацией и клеточным поглощением, были прослежены до раннего происхождения одноклеточных эукариот. [5] Возбудитель простейших Acanthamoeba spp. продемонстрировали доказательства присутствия ACh, который обеспечивает рост и пролиферативные сигналы через расположенный на мембране гомолог M1-мускаринового рецептора. [6]
Частично из-за его функции активации мышц, а также из-за его функций в вегетативной нервной системе и мозге многие важные лекарства оказывают свое действие, изменяя холинергическую передачу. Многочисленные яды и токсины, вырабатываемые растениями, животными и бактериями, а также химические нервно-паралитические агенты, такие как зарин , причиняют вред, инактивируя или гиперактивируя мышцы, воздействуя на нервно-мышечные соединения. Препараты, действующие на мускариновые рецепторы ацетилхолина , такие как атропин , могут быть ядовитыми в больших количествах, но в меньших дозах они обычно используются для лечения определенных заболеваний сердца и глаз. [ нужна ссылка ] Скополамин или дифенгидрамин , которые также действуют преимущественно на мускариновые рецепторы в головном мозге (особенно на М1 рецептор ), могут вызывать делирий , галлюцинации и амнезию за счет антагонизма рецепторов в этих участках. По состоянию на 2016 год только подтип рецептора М1 был вовлечен в антихолинергический делирий. [7] Привыкание к никотину обусловлено его воздействием на никотиновые рецепторы ацетилхолина в головном мозге.
Химия
[ редактировать ]Ацетилхолин представляет собой молекулу холина по , ацетилированную атому кислорода . Из-за заряженной аммониевой группы ацетилхолин не проникает через липидные мембраны. В связи с этим при наружном введении молекула остается во внеклеточном пространстве и в настоящее время считается, что молекула не проходит через гематоэнцефалический барьер.
Биохимия
[ редактировать ]Ацетилхолин синтезируется в определенных нейронах ферментом холина холин-ацетилтрансферазой из соединений КоА и ацетил- . Холинергические нейроны способны продуцировать АХ. Примером центральной холинергической области является базальное ядро Мейнерта в базальном отделе переднего мозга. [8] [9] Фермент ацетилхолинэстераза превращает ацетилхолин в неактивные метаболиты холин и ацетат . Этот фермент широко распространен в синаптической щели, и его роль в быстром удалении свободного ацетилхолина из синапса важна для правильного функционирования мышц. Некоторые нейротоксины действуют путем ингибирования ацетилхолинэстеразы, что приводит к избытку ацетилхолина в нервно-мышечных соединениях , вызывая паралич мышц, необходимых для дыхания, и останавливая биение сердца.
Функции
[ редактировать ]
Ацетилхолин действует как в центральной нервной системе (ЦНС), так и в периферической нервной системе (ПНС). В ЦНС холинергические проекции от базального отдела переднего мозга к коре головного мозга и гиппокампу поддерживают когнитивные функции этих целевых областей. В ПНС ацетилхолин активирует мышцы и является основным нейромедиатором вегетативной нервной системы. [10] [2]
Клеточные эффекты
[ редактировать ]
Как и многие другие биологически активные вещества, ацетилхолин оказывает свое действие путем связывания и активации рецепторов, расположенных на поверхности клеток. Существует два основных класса рецепторов ацетилхолина: никотиновые и мускариновые . Они названы в честь химических веществ, которые могут избирательно активировать каждый тип рецепторов, не активируя другой: мускарин — соединение, обнаруженное в грибе Amanita muscaria ; никотин содержится в табаке.
Никотиновые ацетилхолиновые рецепторы представляют собой лиганд-управляемые ионные каналы, проницаемые для ионов натрия , калия и кальция . Другими словами, это ионные каналы, встроенные в клеточные мембраны, способные переключаться из закрытого состояния в открытое при связывании с ними ацетилхолина; в открытом состоянии они пропускают ионы. Никотиновые рецепторы бывают двух основных типов: мышечные и нейрональные. Мышечный тип можно избирательно блокировать кураре , нейрональный тип — гексаметонием . Основное расположение рецепторов мышечного типа находится на мышечных клетках, как более подробно описано ниже. Рецепторы нейронального типа расположены в вегетативных ганглиях (как симпатических, так и парасимпатических) и в центральной нервной системе.
Мускариновые рецепторы ацетилхолина имеют более сложный механизм и влияют на клетки-мишени в течение более длительного периода времени. У млекопитающих идентифицировано пять подтипов мускариновых рецепторов, обозначенных от М1 до М5. Все они функционируют как рецепторы, связанные с G-белком , а это означает, что они оказывают свое воздействие через систему вторичных мессенджеров . Подтипы M1, M3 и M5 Gq связаны с ; они повышают внутриклеточные уровни IP 3 и кальция путем активации фосфолипазы C. Их воздействие на клетки-мишени обычно носит возбуждающий характер. Подтипы M2 и M4 Gi ; / Go связаны с они снижают внутриклеточные уровни цАМФ путем ингибирования аденилатциклазы . Их влияние на клетки-мишени обычно носит ингибирующий характер. Мускариновые рецепторы ацетилхолина обнаружены как в центральной нервной системе, так и в периферической нервной системе сердца, легких, верхних отделах желудочно-кишечного тракта и потовых железах.
Нервно-мышечный переход
[ редактировать ]
Ацетилхолин — это вещество, которое нервная система использует для активации скелетных мышц , разновидности поперечно-полосатых мышц. Это мышцы, используемые для всех типов произвольных движений, в отличие от гладкой мышечной ткани , которая участвует в ряде непроизвольных действий, таких как перемещение пищи через желудочно-кишечный тракт и сужение кровеносных сосудов. Скелетные мышцы напрямую контролируются мотонейронами, расположенными в спинном мозге или, в некоторых случаях, в стволе мозга . Эти мотонейроны посылают свои аксоны через двигательные нервы , из которых они выходят для соединения с мышечными волокнами в специальном типе синапсов, называемом нервно-мышечным соединением .
Когда мотонейрон генерирует потенциал действия , он быстро перемещается по нерву, пока не достигнет нервно-мышечного соединения, где инициирует электрохимический процесс, вызывающий высвобождение ацетилхолина в пространство между пресинаптическим окончанием и мышечным волокном. Молекулы ацетилхолина затем связываются с никотиновыми рецепторами ионных каналов на мембране мышечных клеток, вызывая открытие ионных каналов. Затем ионы натрия поступают в мышечную клетку, запуская последовательность шагов, которые в конечном итоге вызывают мышечное сокращение .
Факторы, снижающие высвобождение ацетилхолина (и тем самым влияющие на кальциевые каналы Р-типа ): [11]
- Антибиотики ( клиндамицин , полимиксин )
- Магний: противодействует кальциевым каналам Р-типа.
- Гипокальциемия
- Противосудорожные средства
- Диуретики ( фуросемид )
- Синдром Итона-Ламберта : ингибирует кальциевые каналы P-типа.
- Миастения гравис
- Ботулинический токсин : ингибирует белки SNARE.
Блокаторы кальциевых каналов (нифедипин, дилтиазем) не влияют на Р-каналы. Эти препараты влияют на кальциевые каналы L-типа .
Автономная нервная система
[ редактировать ]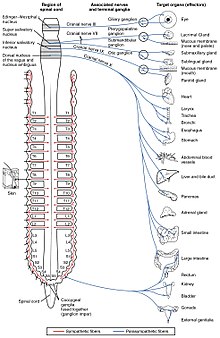
Вегетативная нервная система контролирует широкий спектр непроизвольных и бессознательных функций организма. Ее основными ветвями являются симпатическая нервная система и парасимпатическая нервная система . В широком смысле функция симпатической нервной системы заключается в мобилизации организма к действию; Для описания этого часто используют фразу «бей или беги» . Функция парасимпатической нервной системы заключается в том, чтобы привести организм в состояние, способствующее отдыху, регенерации, пищеварению и воспроизводству; для его описания часто используют фразу «отдыхать и переваривать» или «кормить и размножаться». Обе вышеупомянутые системы используют ацетилхолин, но по-разному.
На схематическом уровне симпатическая и парасимпатическая нервные системы организованы по существу одинаково: преганглионарные нейроны центральной нервной системы посылают проекции нейронам, расположенным в вегетативных ганглиях, которые посылают выходные проекции практически во все ткани тела. В обеих ветвях внутренние связи, проекции центральной нервной системы на вегетативные ганглии, используют ацетилхолин в качестве нейротрансмиттера для иннервации (или возбуждения) нейронов ганглиев. В парасимпатической нервной системе выходные соединения — отростки ганглиозных нейронов к тканям, не принадлежащим к нервной системе, также выделяют ацетилхолин, но действуют на мускариновые рецепторы. В симпатической нервной системе выходные соединения в основном выделяют норадреналин , хотя в некоторых точках, например, при судомоторной иннервации потовых желез, выделяется ацетилхолин.
Прямые сосудистые эффекты
[ редактировать ]Ацетилхолин в сыворотке оказывает прямое влияние на тонус сосудов путем связывания с мускариновыми рецепторами, присутствующими на эндотелии сосудов . Эти клетки реагируют увеличением выработки оксида азота , который сигнализирует окружающим гладким мышцам о необходимости расслабиться, что приводит к расширению сосудов . [12]
Центральная нервная система
[ редактировать ]
В центральной нервной системе АХ оказывает разнообразное влияние на пластичность, возбуждение и вознаграждение . АХ играет важную роль в повышении бдительности, когда мы просыпаемся. [13] в поддержании внимания [14] и в обучении и памяти. [15]
Было показано, что повреждение холинергической (продуцирующей ацетилхолин) системы головного мозга связано с дефицитом памяти, связанным с болезнью Альцгеймера . [16] Также было показано, что ACh способствует быстрому сну. [17]
В стволе мозга ацетилхолин образуется из педункулопонтинного ядра и латеродорсального покрышки, известных под общим названием мезопонтинная покрышка или понтомезэнцефалотегментальный комплекс. [18] [19] В базальном отделе переднего мозга он берет начало из базального ядра Мейнерта и медиального перегородочного ядра :
- Понтомезэнцефалотегментальный комплекс действует преимущественно на М1-рецепторы в стволе мозга , глубоких ядрах мозжечка , мостовых ядрах , голубом пятне , ядре шва , латеральном ретикулярном ядре и нижней оливе . [19] Он также проецируется на таламус , покрышку , базальные ганглии и базальные отделы переднего мозга . [18]
- Базальное ядро Мейнерта действует главным образом на рецепторы М1 в неокортексе .
- Медиальное септальное ядро действует преимущественно на М1-рецепторы в гиппокампе и частях коры головного мозга .
Кроме того, АХ действует как важный внутренний передатчик в полосатом теле , которое является частью базальных ганглиев . Он высвобождается холинергическими интернейронами . У людей, приматов и грызунов эти интернейроны реагируют на выраженные стимулы окружающей среды реакциями, которые по времени совпадают с реакциями дофаминергических нейронов черной субстанции . [20] [21]
Память
[ редактировать ]Ацетилхолин участвует в обучении и памяти несколькими способами. Антихолинергический препарат скополамин ухудшает усвоение новой информации у людей. [22] и животные. [15] У животных нарушение поступления ацетилхолина в неокортекс ухудшает обучение простым задачам распознавания, сравнимым с получением фактической информации. [23] а нарушение поступления ацетилхолина в гиппокамп и прилегающие к нему области коры приводит к забывчивости, сравнимой с антероградной амнезией у людей. [24]
Болезни и расстройства
[ редактировать ]Миастения гравис
[ редактировать ]Заболевание миастения гравис , характеризующееся мышечной слабостью и усталостью, возникает, когда организм ненадлежащим образом вырабатывает антитела против никотиновых рецепторов ацетилхолина и, таким образом, подавляет правильную передачу сигнала ацетилхолина. Со временем концевая пластина двигателя разрушается. Препараты, конкурентно ингибирующие ацетилхолинэстеразу (например, неостигмин, физостигмин или прежде всего пиридостигмин), эффективны при лечении симптомов этого заболевания. Они дают эндогенно высвобождаемому ацетилхолину больше времени для взаимодействия с соответствующим рецептором, прежде чем он будет инактивирован ацетилхолинэстеразой в синаптической щели (пространстве между нервом и мышцей).
Фармакология
[ редактировать ]Блокирование, затруднение или имитация действия ацетилхолина имеет множество применений в медицине. Лекарственные средства, действующие на систему ацетилхолина, являются либо агонистами рецепторов, стимулирующих систему, либо антагонистами, ингибирующими ее. Агонисты и антагонисты ацетилхолиновых рецепторов могут либо оказывать действие непосредственно на рецепторы, либо оказывать свое действие опосредованно, например, воздействуя на фермент ацетилхолинэстеразу , которая разрушает лиганд рецептора. Агонисты повышают уровень активации рецепторов; антагонисты уменьшают его.
Сам по себе ацетилхолин не имеет терапевтической ценности как препарат для внутривенного введения из-за его многогранного действия (неселективности) и быстрой инактивации холинэстеразой. Однако его используют в виде глазных капель, чтобы вызвать сужение зрачка во время операции по удалению катаракты, что способствует быстрому послеоперационному восстановлению.
Никотиновые рецепторы
[ редактировать ]Никотин связывается с никотиновыми рецепторами ацетилхолина и активирует их , имитируя действие ацетилхолина на эти рецепторы. ACh открывает Na + канал при связывании, так что Na + течет в клетку. Это вызывает деполяризацию и приводит к возникновению возбуждающего постсинаптического потенциала. Таким образом, АХ оказывает возбуждающее действие на скелетные мышцы; Электрический отклик быстрый и кратковременный. Кураре — это яд стрел, который действует на никотиновые рецепторы и использовался для разработки клинически полезных методов лечения.
Мускариновые рецепторы
[ редактировать ]Мускариновые рецепторы образуют рецепторные комплексы, связанные с G-белком, и других клеток в клеточных мембранах нейронов . Атропин является неселективным конкурентным антагонистом ацетилхолина мускариновых рецепторов.
Ингибиторы холинэстеразы
[ редактировать ]Многие агонисты рецепторов ACh действуют косвенно, ингибируя фермент ацетилхолинэстеразу . Возникающее в результате накопление ацетилхолина вызывает постоянную стимуляцию мышц, желез и центральной нервной системы, что может привести к фатальным судорогам, если доза высока.
Они являются примерами ингибиторов ферментов и усиливают действие ацетилхолина, замедляя его распад; некоторые из них использовались в качестве нервно-паралитических агентов ( зарин и нервно-паралитический газ VX ) или пестицидов ( органофосфаты и карбаматы ). Многие токсины и яды, вырабатываемые растениями и животными, также содержат ингибиторы холинэстеразы. В клинической практике их применяют в низких дозах. [ почему? ] для обратного действия миорелаксантов , для лечения миастении и лечения симптомов болезни Альцгеймера ( ривастигмин , который увеличивает холинергическую активность в головном мозге).
Ингибиторы синтеза
[ редактировать ]Органические соединения ртути , такие как метилртуть , обладают высоким сродством к сульфгидрильным группам , что вызывает дисфункцию фермента холин-ацетилтрансферазы. Это ингибирование может привести к дефициту ацетилхолина и иметь последствия для двигательной функции.
Ингибиторы высвобождения
[ редактировать ]Ботулинический токсин (ботокс) действует путем подавления высвобождения ацетилхолина, тогда как яд паука черной вдовы ( альфа-латротоксин ) имеет обратный эффект. Ингибирование АХ вызывает паралич . При укусе паука черной вдовы человек теряет запасы АХ, и мышцы начинают сокращаться. Если и когда запасы исчерпаны, наступает паралич .
Сравнительная биология и эволюция
[ редактировать ]Ацетилхолин используется организмами во всех сферах жизни для самых разных целей. Считается, что холин , предшественник ацетилхолина, использовался одноклеточными организмами миллиарды лет назад. [ нужна ссылка ] для синтеза фосфолипидов клеточных мембран. [25] После эволюции переносчиков холина обилие внутриклеточного холина открыло путь для включения холина в другие пути синтеза, включая производство ацетилхолина. Ацетилхолин используется бактериями, грибами и множеством других животных. Многие применения ацетилхолина основаны на его действии на ионные каналы через GPCR, такие как мембранные белки.
Два основных типа рецепторов ацетилхолина, мускариновые и никотиновые рецепторы, эволюционировали, чтобы реагировать на ацетилхолин. Это означает, что эти рецепторы произошли не от общего гомолога, а из отдельных семейств рецепторов. Подсчитано, что семейство никотиновых рецепторов насчитывает более 2,5 миллиардов лет. [25] Аналогичным образом считается, что мускариновые рецепторы отделились от других GPCR по крайней мере 0,5 миллиарда лет назад. Обе эти группы рецепторов развили множество подтипов с уникальным сродством к лигандам и механизмами передачи сигналов. Разнообразие типов рецепторов позволяет ацетилхолину вызывать различные реакции в зависимости от того, какие типы рецепторов активированы, и позволяет ацетилхолину динамически регулировать физиологические процессы. Рецепторы АХ родственны рецепторам 5-НТ3 ( серотонина ), ГАМК и глицина как по последовательности, так и по структуре, что убедительно свидетельствует о том, что они имеют общее эволюционное происхождение. [26]
История
[ редактировать ]В 1867 году Адольф фон Байер расшифровал структуры холина и ацетилхолина и синтезировал их оба, назвав последний « ацетилнейрином ». в своем исследовании [27] [28] Холин является предшественником ацетилхолина. Вот почему Фредерик Уокер Мотт и Уильям Добинсон Халлибертон в 1899 году отметили, что инъекции холина снижают кровяное давление животных. [29] [28] Биологическая активность ацетилхолина впервые была отмечена в 1906 году, когда Рид Хант (1870–1948) и Рене де М. Таво обнаружили, что он снижает кровяное давление в исключительно малых дозах. [30] [28] [31]
В 1914 году Артур Дж. Юинс первым извлек ацетилхолин из природы. он идентифицировал его как снижающий кровяное давление примесь из некоторых Claviceps purpurea экстрактов спорыньи По просьбе Генри Халлетта Дейла . [28] Позже, в 1914 году, Дейл описал действие ацетилхолина на различные типы периферических синапсов, а также отметил, что он снижает кровяное давление кошек при подкожных инъекциях даже в дозах в один нанограмм . [32] [28]
Концепция нейротрансмиттеров была неизвестна до 1921 года, когда Отто Леви заметил, что блуждающий нерв секретирует вещество, которое ингибирует сердечную мышцу во время работы профессором в Университете Граца . Он назвал его vagusstoff («блуждающее вещество»), отметил, что это структурный аналог холина, и предположил, что это ацетилхолин. [33] [34] В 1926 году Лоуи и Э. Навратил пришли к выводу, что это соединение, вероятно, представляет собой ацетилхолин, поскольку вагустофф и синтетический ацетилхолин теряли свою активность аналогичным образом при контакте с лизатами тканей , которые содержали ферменты, разлагающие ацетилхолин (теперь известные как холинэстеразы ). [35] [36] Этот вывод был широко принят. Более поздние исследования подтвердили функцию ацетилхолина как нейромедиатора . [34]
В 1936 году Х. Х. Дейл и О. Лоуи получили Нобелевскую премию по физиологии и медицине за исследования ацетилхолина и нервных импульсов. [28]
См. также
[ редактировать ]- Энн Сильвер
- Ацетилхолинэстераза
- Нервно-мышечный переход
- Никотиновый рецептор ацетилхолина
- Мускариновый рецептор ацетилхолина
Конкретные ссылки
[ редактировать ]- ^ Jump up to: а б Тивари П., Двиведи С., Сингх М.П., Мишра Р., Чанди А. (октябрь 2012 г.). «Основные и современные представления о холинергических рецепторах: обзор» . Азиатско-Тихоокеанский журнал тропических болезней . 3 (5): 413–420. дои : 10.1016/S2222-1808(13)60094-8 . ПМК 4027320 .
- ^ Jump up to: а б с Сэм С., Бордони Б. (2023). «Физиология ацетилхолина» . СтатПерлс . Остров сокровищ (Флорида): StatPearls Publishing. ПМИД 32491757 . Проверено 06 апреля 2023 г.
- ^ Лотт Э.Л., Джонс Э.Б. (июнь 2019 г.). «Холинергическая токсичность» . СтатПерлс . Остров сокровищ (Флорида): StatPearls Publishing. ПМИД 30969605 .
- ^ Капалка ГМ (2010). «Вещества, участвующие в нейротрансмиссии». Диетическая и травяная терапия для детей и подростков . Эльзевир. стр. 71–99 . дои : 10.1016/b978-0-12-374927-7.00004-2 . ISBN 978-0-12-374927-7 .
- ^ Байг А.М., Рана З., Тарик С., Лалани С., Ахмад Х.Р. (март 2018 г.). «Прослежено на временной шкале: открытие ацетилхолина и компонентов холинергической системы человека у примитивных одноклеточных эукариот Acanthamoeba spp». ACS Chem Neurosci . 9 (3): 494–504. дои : 10.1021/acschemneuro.7b00254 . ПМИД 29058403 .
- ^ Байг А.М., Ахмад Х.Р. (июнь 2017 г.). «Доказательства наличия M1-мускаринового гомолога GPCR у одноклеточных эукариот: биоинформатическое 3D-моделирование и эксперименты Acanthamoeba spp» . Дж. Рецепт. Трансдукт сигнала. Рез . 37 (3): 267–275. дои : 10.1080/10799893.2016.1217884 . ПМИД 27601178 . S2CID 5234123 .
- ^ Доусон А.Х., Бакли Н.А. (март 2016 г.). «Фармакологическое лечение антихолинергического делирия – теория, доказательства и практика» . Британский журнал клинической фармакологии . 81 (3): 516–524. дои : 10.1111/bcp.12839 . ПМЦ 4767198 . ПМИД 26589572 .
Делирий связан только с антагонизмом постсинаптических рецепторов М1, и на сегодняшний день другие подтипы рецепторов не задействованы.
- ^ Смитис Дж (2009). «Философия, восприятие и нейробиология». Восприятие . 38 (5): 638–51. дои : 10.1068/p6025 . ПМИД 19662940 . S2CID 45579740 .
- ^ Смитис Дж., Д'Орей де Лантреманж М. (2016). «Природа и функции механизмов сжатия цифровой информации в мозге и в технологиях цифрового телевидения» . Передняя система нейронов . 10 : 40. дои : 10.3389/fnsys.2016.00040 . ПМЦ 4858531 . ПМИД 27199688 .
- ^ Ваксенбаум Дж.А., Редди В., Варакалло М. (2023). «Анатомия вегетативной нервной системы» . СтатПерлс . Остров сокровищ (Флорида): StatPearls Publishing. ПМИД 30969667 . Проверено 06 апреля 2023 г.
- ^ Миллер Р.Д., Эрикссон Л.И., Флейшер Л.А., Винер-Крониш Дж.П., Янг В.Л., ред. (01.01.2009). Анестезия Миллера (7-е изд.). Elsevier Науки о здоровье. стр. 343–47. ISBN 978-0-443-06959-8 .
- ^ Келлог Д.Л., Чжао Дж.Л., Кои Ю., Грин СП (февраль 2005 г.). «Ацетилхолин-индуцированная вазодилатация опосредуется оксидом азота и простагландинами в коже человека». Дж. Прил. Физиол . 98 (2): 629–32. doi : 10.1152/japplphysicalol.00728.2004 . ПМИД 15649880 . S2CID 293055 .
- ^ Джонс Б.Е. (ноябрь 2005 г.). «От бодрствования ко сну: нейронные и химические субстраты». Тренды Фармакол. Наука . 26 (11): 578–86. дои : 10.1016/j.tips.2005.09.009 . ПМИД 16183137 .
- ^ Химмельхебер А.М., Сартер М., Бруно Дж.П. (июнь 2000 г.). «Увеличение высвобождения коркового ацетилхолина во время устойчивого внимания у крыс». Мозговой Res Cogn Мозговой Res . 9 (3): 313–25. дои : 10.1016/S0926-6410(00)00012-4 . ПМИД 10808142 .
- ^ Jump up to: а б Ридли Р.М., Боуз П.М., Бейкер Х.Ф., Кроу Т.Дж. (1984). «Участие ацетилхолина в обучении различению объектов и памяти мартышек». Нейропсихология . 22 (3): 253–63. дои : 10.1016/0028-3932(84)90073-3 . ПМИД 6431311 . S2CID 7110504 .
- ^ Фрэнсис П.Т., Палмер А.М., Снейп М., Уилкок Г.К. (февраль 1999 г.). «Холинергическая гипотеза болезни Альцгеймера: обзор прогресса» . Дж. Нейрол. Нейрохирургия. Психиатрия . 66 (2): 137–47. дои : 10.1136/jnnp.66.2.137 . ПМК 1736202 . ПМИД 10071091 .
- ^ Платт Б., Ридель Г. (август 2011 г.). «Холинергическая система, ЭЭГ и сон». Поведение. Мозговой Рес . 221 (2): 499–504. дои : 10.1016/j.bbr.2011.01.017 . ПМИД 21238497 . S2CID 25323695 .
- ^ Jump up to: а б Вульф, штат Нью-Джерси, Батчер, Л.Л. (май 1986 г.). «Холинергические системы в мозге крысы: III. Проекции от понтомезэнцефалической покрышки на таламус, тектум, базальные ганглии и базальные отделы переднего мозга». Мозговой Рес. Бык . 16 (5): 603–37. дои : 10.1016/0361-9230(86)90134-6 . ПМИД 3742247 . S2CID 39665815 .
- ^ Jump up to: а б Вульф, штат Нью-Джерси, Батчер, Л.Л. (декабрь 1989 г.). «Холенергические системы мозга крысы: IV. Нисходящие проекции понтомезэнцефалической покрышки». Мозговой Рес. Бык . 23 (6): 519–40. дои : 10.1016/0361-9230(89)90197-4 . ПМИД 2611694 . S2CID 4721282 .
- ^ Голдберг Дж. А., Рейнольдс Дж. Н. (декабрь 2011 г.). «Спонтанная активация и вызванные паузы в тонически активных холинергических интернейронах полосатого тела». Нейронаука . 198 : 27–43. doi : 10.1016/j.neuroscience.2011.08.067 . ПМИД 21925242 . S2CID 21908514 .
- ^ Моррис Г., Аркадир Д., Невет А., Ваадия Э., Бергман Х. (июль 2004 г.). «Совпадающие, но разные сообщения дофамина среднего мозга и тонически активных нейронов полосатого тела» . Нейрон . 43 (1): 133–43. дои : 10.1016/j.neuron.2004.06.012 . ПМИД 15233923 .
- ^ Кроу Т.Дж., Гроув-Уайт И.Г. (октябрь 1973 г.). «Анализ дефицита обучения после введения гиосцина человеку» . Бр. Дж. Фармакол . 49 (2): 322–7. дои : 10.1111/j.1476-5381.1973.tb08379.x . ПМЦ 1776392 . ПМИД 4793334 .
- ^ Ридли Р.М., Мюррей Т.К., Джонсон Дж.А., Бейкер Х.Ф. (июнь 1986 г.). «Нарушение обучения после поражения базального ядра Мейнерта у мартышки: модификация холинергическими препаратами». Мозговой Рес . 376 (1): 108–16. дои : 10.1016/0006-8993(86)90904-2 . ПМИД 3087582 . S2CID 29182517 .
- ^ Истон А., Ридли Р.М., Бейкер Х.Ф., Гаффан Д. (июль 2002 г.). «Односторонние поражения холинергических базальных отделов переднего мозга и свода в одном полушарии и нижней височной коры в противоположном полушарии вызывают серьезные нарушения обучения у макак-резус» . Цереб. Кортекс . 12 (7): 729–36. дои : 10.1093/cercor/12.7.729 . ПМИД 12050084 .
- ^ Jump up to: а б Дин Б. (ноябрь 2009 г.). «Эволюция холинергической системы ЦНС человека: привело ли это к возникновению психических заболеваний?». Aust NZJ Психиатрия . 43 (11): 1016–28. дои : 10.3109/00048670903270431 . ПМИД 20001397 . S2CID 31059344 .
- ^ Ортеллс М.О., Лунт Г.Г. (март 1995 г.). «Эволюционная история суперсемейства рецепторов лиганд-управляемых ионных каналов». Тенденции в нейронауках . 18 (3): 121–127. дои : 10.1016/0166-2236(95)93887-4 . ПМИД 7754520 . S2CID 18062185 .
- ^ Байер А (1867). «I. О нейроне» . Ann Chem Юстуса Либиха (на немецком языке). 142 (3): 322–326. дои : 10.1002/jlac.18671420311 .
- ^ Jump up to: а б с д и ж Кавасима К., Фуджи Т., Мориваки Ю., Мисава Х., Хоригучи К. (2015). «Ненейрональная холинергическая система в регуляции иммунной функции с акцентом на α7 нАХР» . Международная иммунофармакология . 29 (1): 127–34. дои : 10.1016/j.intimp.2015.04.015 . ПМИД 25907239 .
- ^ Мотт Ф.В., Халлибертон В.Д. (1899 г.). «VII. Физиологическое действие холина и нейрина» . Философские труды Лондонского королевского общества. Серия Б, содержащая статьи биологического характера . 191 (2001): 211–267. дои : 10.1098/rstb.1899.0007 . ПМЦ 2463419 . ПМИД 20758460 .
- ^ Хант Р., Таво М. (1906). «О физиологическом действии некоторых производных холина и новых методах обнаружения холина». БМЖ . 2 : 1788–1791.
- ^ Доркинс HR (апрель 1982 г.). «Сукаметоний – разработка современного препарата с 1906 года по наши дни» . Медицинская история . 26 (2): 145–68. дои : 10.1017/s0025727300041132 . ПМЦ 1139149 . ПМИД 7047939 .
- ^ Дейл Х.Х. (1914). «Действие некоторых сложных и простых эфиров холина и их отношение к мускарину» . J Pharmacol Exp Ther . 6 (2): 147–190.
- ^ Леви О (1922). «О гуморальной переносимости влияний сердечного нерва». Плуг Arch Ges Phys (на немецком языке). 193 (1): 201–213. дои : 10.1007/BF02331588 . S2CID 34861770 .
- ^ Jump up to: а б Зейзель С.Х. (2012). «Краткая история холина» . Анналы питания и обмена веществ . 61 (3): 254–8. дои : 10.1159/000343120 . ПМЦ 4422379 . ПМИД 23183298 .
- ^ Леви О, Навратил Э (1926). «О гуморальной переносимости влияний сердечного нерва». Плуг Arch Ges Phys (на немецком языке). 214 (1): 678–688. дои : 10.1007/BF01741946 . S2CID 43748121 .
- ^ Циммер Х.Г. (март 2006 г.). «Отто Леви и химическая передача стимуляции блуждающего нерва в сердце» . Клиническая кардиология . 29 (3): 135–6. дои : 10.1002/clc.4960290313 . ПМК 6654523 . ПМИД 16596840 .
Общая библиография
[ редактировать ]- Бреннер GM, Стивенс CW (2006). Фармакология (2-е изд.). Филадельфия, Пенсильвания: У. Б. Сондерс. ISBN 1-4160-2984-2 .
- Канадская ассоциация фармацевтов (2000 г.). Справочник фармацевтических препаратов и специальностей (25-е изд.). Торонто, ON: Webcom. ISBN 0-919115-76-4 .
- Карлсон Н.Р. (2001). Физиология поведения (7-е изд.). Нидхэм-Хайтс, Массачусетс: Аллин и Бэкон. ISBN 0-205-30840-6 .
- Гершон, доктор медицинских наук (1998). Второй мозг . Нью-Йорк, штат Нью-Йорк: HarperCollins. ISBN 0-06-018252-0 .
- Сигал А., Сапру Х.Н. (2006). «Гл. 15» . Основная неврология (пересмотренное 1-е изд.). Филадельфия: Липпинкотт, Уильямс и Уилкинс. стр. 255–267 .
- Хассельмо М.Е. (февраль 1995 г.). «Нейромодуляция и корковая функция: моделирование физиологических основ поведения». Поведение. Мозговой Рес . 67 (1): 1–27. дои : 10.1016/0166-4328(94)00113-T . ПМИД 7748496 . S2CID 17594590 . как PDF
- Ю А.Дж., Даян П. (май 2005 г.). «Неопределенность, нейромодуляция и внимание» . Нейрон . 46 (4): 681–92. дои : 10.1016/j.neuron.2005.04.026 . ПМИД 15944135 . S2CID 15980355 . как PDF